Det blev allerede nævnt ovenfor, at i cellekernen er DNA-molekyler placeret i særlige strukturer kaldet kromosomer. Deres forskning begyndte for over 100 år siden ved hjælp af et konventionelt lysmikroskop. Allerede af slutningen af det 19. århundredeårhundrede blev der opdaget noget om kromosomernes adfærd i celledelingsprocessen, og ideen blev udtrykt om deres deltagelse i overførslen af arvelighed.
Kromosomer bliver synlige i et mikroskop, når en celle deler sig på et bestemt stadie af cellecyklussen kaldet mitose. Kromosomer i denne tilstand er kompakte stavformede strukturer forskellige længder med en nogenlunde konstant tykkelse har de fleste kromosomer en forsnævring, der deler kromosomet i to arme. I området af indsnævringen er der en struktur vigtig for fordobling af kromosomer, kaldet centromere. Når en celle deler sig under mitose, fordobles antallet af kromosomer, hvilket resulterer i, at begge nydannede celler i sidste ende forsynes med det samme standard sæt kromosomer.
Først i 1956 beskrev Y. Tio og A. Levan for første gang det menneskelige kromosomsæt og bestemte kvantitativ sammensætning kromosomer og gav dem deres total morfologiske egenskaber. Faktisk markerede disse værker begyndelsen på studiet af strukturen af det menneskelige genom. Hos mennesker indeholder hver celle i kroppen 46 kromosomer, hvis fysiske længder varierer fra 1,5 til 10 mikron (fig. 7).
Ris. 7. Et mikroskopisk billede af det komplette sæt af kromosomer indeholdt i kernen af hver enkelt menneskelig celle
Lad os minde læseren om, at sættet af kromosomer i alle menneskelige celler (undtagen kønsceller) kaldes diploid (dobbelt), da hvert kromosom er repræsenteret af to kopier (23 par i alt). Hver human somatisk celle (undtagen røde blodlegemer) indeholder 2 komplette sæt kromosomer. Hvert enkelt (haploid) sæt indeholder 23 kromosomer - 22 almindelige kromosomer (autosomer) og et kønskromosom - X eller Y. Således er genomet af hver bestemt person består af 23 par kæmpe DNA-molekyler fordelt på forskellige kromosomer, og hvis vi taler om det menneskelige genom generelt (mænd og kvinder), så samlet antal der er 24 sådanne molekyler. Dette er det første grundlæggende blanding, som blev opnået om det menneskelige genom ved at analysere kromosomer.
En undersøgelse af strukturen (størrelse og form) af menneskelige kromosomer viste, at de fleste af dem udseende ligner kegler, der består af to tykke dele (kromatider) og en tynd indsnævring (centromerer) mellem dem. Ligheden med kegler og ikke med håndvægte er, at centromeren oftest ikke er placeret i midten af kromosomet, men forskydes til en af dens ender. Kromosomstørrelser varierer meget, hvor det korteste kromosom er cirka ti gange mindre end det længste. Dette er det andet princip vigtig information om strukturen af det menneskelige genom - de 24 DNA-molekyler, der udgør det, har forskellige størrelser.
Hvis man sammenligner antallet og størrelsen af kromosomer hos mennesker og hos andre arter af organismer, kan man se enorme forskelle. For eksempel har en ko, hvis genomstørrelse er omtrent lig med det menneskelige genom, 60 par kromosomer. Den kløvede frø indeholder kun 18 kromosomer, men selv de mindste af dem er større end de største menneskelige kromosomer. Hos fugle når antallet af kromosomer tværtimod 40 eller mere, og de er alle meget små i størrelse. Således er mangfoldigheden af kromosomer i naturen meget stor.
Ved hjælp af lysmikroskopi blev størrelsen af alle menneskelige kromosomer bestemt. Derefter blev alle ikke-kønskromosomer nummereret i rækkefølge efter faldende størrelse - fra 1 til 22. Kønskromosomerne blev ikke tildelt et nummer, men blev navngivet X og Y. Som mere præcise efterfølgende undersøgelser viste, viste kromosom 21 sig faktisk at være en smule mindre end 22, men nummereringen af kromosomerne blev ikke ændret (for ikke at forårsage forvirring). Forskellen i kromosomsættene mellem mænd og kvinder er, at kvinder har to køns X-kromosomer (dvs. kromosomerne i alle 23 par er ens), og hos mænd dannes et par med X-kromosomet af manden kønskromosom- Y. Hvert kromosom kan betragtes som et separat bind af en stor 24-binds samling af værker kaldet Encyclopedia of Man.
Menneskelige kønsceller indeholder, i modsætning til cellerne i en voksen organismes krop (somatiske celler), ikke 2 sæt volumener af DNA-tekst, men kun én. Før undfangelsen består hvert enkelt kromosom (et separat bind i Human Encyclopedia) af faderens sæd og mors æg af forskellige kapitler af deres forældres DNA-tekst blandet i forskellige kombinationer. Ethvert af de kromosomer, vi modtog fra vores far, blev dannet i hans testikler, kort før vi blev undfanget. Tidligere, i hele menneskehedens historie, havde et sådant kromosom aldrig eksisteret. Det blev dannet gennem processen med tilfældig blanding, der opstår under deling, og som gradvist dannes af dele af forfædrenes kromosomer på faderens side, der forenes med hinanden. Situationen er den samme med ægs kromosomer, bortset fra at de er dannet i vores mors krop længe før vi bliver født (næsten umiddelbart efter fødslen af moderen selv).
I zygoten, som er dannet som et resultat af sammensmeltningen af en sædcelle og et æg, blandes og blandes de moderlige og faderlige gener i forskellige kombinationer. Dette sker som et resultat af det faktum, at kromosomerne ikke forbliver uændrede gennem generationer - de interagerer med deres tilfældigt stødte par og udveksler materiale med det. Denne igangværende proces kaldes rekombination. Og den næste generation får ofte et hybridkromosom – dels fra bedstefaren og dels fra bedstemoderen. Længere i en række af generationer krydser og divergerer genveje konstant. Som et resultat af sammensmeltningen af et unikt æg med en unik sædcelle opstår et genom, der er unikt i alle henseender. Og i denne forstand er vi alle unikke. Hvert menneskeligt individ gemmer unik genetisk information bestående af en tilfældig kombination forskellige muligheder gener.
Et enkelt gen kan ses som en enhed, der fortsætter med at eksistere over adskillige generationer. Og i denne forstand er genet udødelig! Der er endda et så originalt synspunkt, at det ikke er mennesker selv, men deres gener, der styrer verden, og hver specifik levende organisme tjener kun som et midlertidigt husly for dem. Denne kontroversielle idé kommer fra Richard Dawkins, forfatter til The Selfish Gene. Efter hans mening er gener praktisk talt udødelige, i modsætning til de levende organismer, hvori de findes. Nogle gener er titusinder eller endda hundreder af millioner af år gamle. Gener, for at bruge Dawkins' terminologi, gør hvad de kan for at overleve. De tilpasser sig varme og kulde, vælger et bedre sted for sig selv, migrerer ved hjælp af mennesker og indgår i nye kombinationer. Manden viste sig at være en ret urolig ejer. I tusinder af år har han rejst meget rundt i verden og spredt sin tilstedeværelse, indflydelse og sine ting - gener. (Den nysgerrige læser kan lære mere om R. Dawkins' ideer og argumentation i Bilag 1). Dette synspunkt er langt fra indiskutabelt, og fra den videre fremstilling vil det blive klart for os, at gener for det første ikke er egoistiske, men arbejdsnarkomaner. Der er gener, der er "vogtere" af genomet, gener, der er "pedeller", gener, der er "kokke", og gener, der er "husholdere." Ved at sikre deres eksistens sikrer de vores eksistens.
Umiddelbart efter undfangelsen fremtidige mand repræsenterer kun én celle (zygote), udstyret med et initialt DNA-bibliotek indeholdende 46 volumener. Blandt de 46 bind modtages 23 altid fra faderen og de øvrige 23 fra moderen. Teksterne i de 23 faderlige og 23 moderlige bind, selvom de generelt er meget ens, adskiller sig ikke desto mindre i detaljer. For eksempel er der i faderbind nr. 18 på side 253 en kommandosætning (i form af et gen), der siger, at barnets øjne skal være brune, og i samme moderbind på samme side står der også skrevet. om øjenfarve, men ifølge Denne tekst skal farven være blå. Den første indikation er mere streng (dominerende) end den anden, og som et resultat vil barnets øjne have en brun farve. Det gen, der dikterer dets rettigheder, kaldes dominerende og den, der afstår sine rettigheder - recessiv. Kun de mennesker, der har blå øjenfarve, har følgende i både deres moderlige og faderlige tekster: recessive gener, hvor der er en indikation af blåøjethed. Derefter deler zygoten sig i to celler, hver af dem deler sig igen, og så videre, indtil milliarder af celler dukker op. Processen med celledeling er vist skematisk i fig. 8.
Ved hver celledeling kopieres mængderne af DNA-tekst, der er indeholdt i bibliotekerne, nøjagtigt uden praktisk talt fejl. Den voksne menneskekrop består i gennemsnit af 10 14 celler. For eksempel er der i hjernen og leveren cirka 10 milliarder celler i immunsystem- 300 milliarder celler. I løbet af et menneskes hele liv sker der omkring 10 16 celledelinger i hans krop. Cellulær sammensætning Mange organer fornyes flere gange over 70 års levetid. Og hver af disse celler indeholder de samme 46 bind DNA-tekst.
I slutningen af 60'erne af det 20. århundrede skete der et vigtigt gennembrud i studiet af kromosomer. Det var kun på grund af det faktum, at de begyndte at bruge en speciel maling til deres farvning. kontrastmiddel- akrichine sennep, og så andre forbindelser, der ligner det. Denne farvning gjorde det muligt at identificere inde i kromosomerne stort antal forskellige understrukturer, der ikke kunne påvises under et mikroskop uden farvning. Efter farvning af kromosomerne med et specifikt Giemsa-Romanovsky-farvestof ligner de zebraer: tværgående lyse og mørke striber med varierende farveintensitet er synlige langs hele længden.
Ris. 8. De vigtigste stadier af cellecyklussen fører til celledeling
Disse bånd kaldes kromosomale G-segmenter eller bånd (fig. 9). Mønstret for segmentering varierer meget mellem forskellige kromosomer, men arrangementet af kromosomsegmenter er konstant på hvert kromosom i alle typer af menneskelige celler.
Arten af de striber, der afsløres ved farvning, er endnu ikke helt klarlagt. Det er nu kun blevet fastslået, at regioner af kromosomer svarende til mørke bånd (kaldet R-bånd) replikerer tidligere end lyse områder (kaldet G-bånd). Således har bånddannelsen af kromosomer højst sandsynligt stadig en eller anden betydning, som endnu ikke er fuldt ud forstået.
Farvning af kromosomer lettede i høj grad deres identifikation og bidrog efterfølgende til at bestemme placeringen af gener på dem (genkortlægning).
Ris. 9. Specifikke kromosomale G-segmenter identificeret ved farvning af menneskelige kromosomer og deres betegnelsessystem i henhold til løsningen international konference i Paris i 1971. Tallene under kromosomerne angiver deres tal. X og Y - kønskromosomer, p - kort arm, q - lang arm af kromosomer
Selvom de detaljerede processer, der forekommer under farvning, endnu ikke er helt klare, er det indlysende, at farvemønsteret afhænger af en sådan parameter som det øgede eller reducerede indhold af AT- eller GC-par i individuelle kromosombånd. Og det er en ting mere generel information om genomet - det er ikke homogent; det indeholder regioner beriget med visse nukleotidpar.
Dette kan især skyldes repeterbarheden af visse typer af DNA-nukleotidsekvenser i visse regioner.
Differentiel farvning af kromosomer har fundet bred anvendelse til påvisning og identifikation af små individuelle ændringer i genomet af en bestemt person ( polymorfi), hvilket især fører til forskellige patologier. Et eksempel på dette er opdagelsen af det såkaldte Philadelphia-kromosom, som findes hos patienter med kronisk myeloid leukæmi. Ved hjælp af kromosomfarvning er det blevet fastslået, at hos patienter med denne sygdom forsvinder et bestemt fragment på kromosom 21 og vises for enden af den lange arm af kromosom 9 (fragmentoverførsel eller translokation, forkortet t). Genetikere betegner en sådan begivenhed som t (9; 21). Kromosomanalyse indikerer således, at forskellige DNA-molekyler kan udveksle separate sektioner med hinanden, hvilket resulterer i dannelsen af "hybrider" i genomet, bestående af DNA-molekyler af forskellige kromosomer. Analyse af kromosomernes allerede undersøgte egenskaber gjorde det muligt at danne sig en idé om polymorfi af det menneskelige genom.
For at bestemme lokaliseringen af individuelle gener på kromosomer (det vil sige genkortlægning) bruges et helt arsenal af specielle metoder, ofte meget komplekse i design og udførelse. En af de vigtigste er molekylær hybridisering (dannelse af en hybrid) af et gen eller dets fragment med kromosompræparater fikseret på et fast underlag, isoleret fra celler i ren form(dette kaldes hybridisering in situ). Essensen af hybridiseringsmetoden in situ består i interaktionen (hybridiseringen) mellem denaturerede (uflettede) DNA-strenge i kromosomer og komplementære nukleotidsekvenser af kromosomer tilføjet til præparatet, individuelt enkeltstrenget DNA eller RNA (de kaldes sonder). Hvis der er komplementaritet mellem en af strengene af kromosomalt DNA og proben, dannes der ret stabile molekylære hybrider mellem dem. Proberne er præ-mærket vha forskellige mærker(radioaktiv, fluorescerende osv.). Steder for hybriddannelse på kromosomer identificeres ved placeringen af disse mærker på kromosompræparater. Så selv før fremkomsten af metoder genteknologi og DNA-sekventering har for eksempel afsløret placeringen i det humane genom af gener, der koder for store og små ribosomale RNA'er (rRNA'er). Generne af førstnævnte viste sig at være lokaliseret i fem forskellige humane kromosomer (13, 14, 15, 21 og 22), mens hovedparten af små rRNA-gener ( 5S RNA) er koncentreret ét sted på den lange arm af kromosom 1.
Et eksempel på et billede opnået ved hybridisering af fluorescerende farvestof-mærkede genprober er vist i fig. 10 på den farvede indsats.
Ris. 10. Hybridisering af humane kromosomer med genprober mærket med røde og grønne fluorescerende farvestoffer. Pilene angiver placeringen af de tilsvarende gener i enderne af to forskellige kromosomer (øverst til højre viser en forstørrelse af billedet af hybridiserende kromosomer).
Gener placeret på det samme kromosom defineres som forbundne gener. Hvis gener er placeret på forskellige kromosomer, nedarves de uafhængigt (uafhængig segregation). Når gener er på det samme kromosom (det vil sige forbundet), er de ude af stand til uafhængig adskillelse. Lejlighedsvis kan der forekomme forskellige ændringer i kromosomer i kønsceller som følge af rekombinationsprocesser mellem homologe kromosomer. En af disse processer kaldes krydser over. På grund af krydsning er koblingen mellem gener fra den samme gruppe aldrig fuldstændig. Jo tættere de forbundne gener er på hinanden, jo mindre sandsynligt er det, at placeringen af sådanne gener vil ændre sig hos børn sammenlignet med deres forældre. Måling af hyppigheden af rekombinationer (overkrydsning) bruges til at etablere lineær rækkefølge gener på et kromosom inden for en koblingsgruppe. Ved kortlægning af kromosomer afgøres det således i første omgang, om disse gener er placeret på samme kromosom, uden at specificere hvilket. Efter at mindst et af generne fra en given koblingsgruppe er lokaliseret på et specifikt kromosom (for eksempel ved brug af hybridisering in situ), bliver det klart, at alle andre gener i denne koblingsgruppe er placeret på det samme kromosom.
Det første eksempel på forbindelsen af gener med bestemte kromosomer kan være påvisningen af koblingen af visse arvelige træk med kønskromosomer. For at bevise lokaliseringen af et gen på det mandlige køn Y-kromosom er det nok at vise, at denne egenskab altid kun findes hos mænd og aldrig findes hos kvinder. Bindingsgruppen af det kvindelige X-kromosom er unikt karakteriseret ved fraværet af arvelige egenskaber, der overføres fra far til søn, og arven af moderkarakteristika.
Særlig vigtig for at studere det menneskelige genom i de tidlige stadier af dets forskning var en metode kaldet somatisk cellehybridisering. Når humane somatiske (ikke-reproduktive) celler blandes med celler fra andre dyrearter (oftest blev mus eller kinesiske hamsterceller brugt til dette formål), kan fusion af deres kerner (hybridisering) forekomme i nærvær af visse midler. Når sådanne hybridceller reproducerer, går nogle kromosomer tabt. Ved et lykkeligt uheld for forsøgslederne går de fleste af de menneskelige kromosomer tabt i human-mus hybridceller. Dernæst udvælges hybrider, hvor kun ét menneskeligt kromosom er tilbage. Undersøgelser af sådanne hybrider har gjort det muligt at associere nogle biokemiske egenskaber, der er karakteristiske for humane celler, med visse humane kromosomer. Gradvist, gennem brug af selektive medier, lærte de at opnå bevarelse eller tab af individuelle menneskelige kromosomer, der bærer bestemte gener. Udvælgelsesskemaet, selvom det ved første øjekast ikke var særlig enkelt, viste sig ret godt i forsøget. Således kom de med et særligt selektivt medium, hvor kun de celler, hvori enzymet thymidinkinase er syntetiseret, kan overleve. Hvis vi til hybridisering med humane celler tager mutante museceller, der ikke syntetiserer thymidinkinase, som partner, så vil kun de hybrider, der indeholder humane kromosomer med thymidinkinasegenet, overleve. På denne måde var det for første gang muligt at etablere lokaliseringen af thymidinkinasegenet på humant kromosom 17.
På trods af at studiet af det menneskelige genom på kromosomniveau gav en række vigtige egenskaber, var de de mest generelle og gav relativt lidt til en fuldstændig forståelse af strukturen og funktionen af det genetiske apparat i menneskelige celler.
| |
Vi sagde, at somatiske celler indeholder et dobbelt, diploid sæt kromosomer, og modne kønsceller indeholder et enkelt, haploid sæt. Et diploid sæt kromosomer er også til stede i umodne kønsceller. En halvering af antallet af kromosomer og følgelig DNA, kaldet deres reduktion, sker i processen med gametogenese, det vil sige udviklingen af kønsceller - kønsceller. En halvering af antallet af kromosomer i kønsceller er en forberedelse til fremtidig befrugtning, hvor deres diploide sæt genoprettes på grund af kombinationen af de haploide kromosomsæt af sæd og æg.
Gametogenese forekommer i gonaderne: i testiklerne i den mandlige krop og i æggestokkene hos hunnen. Følgelig kaldes det spermatogenese og oogenese. Fælles for spermato- og oogenese er de første 3 perioder af gametogenesen: reproduktion, vækst og modning. Hankønsceller, der udvikler sig, gennemgår en yderligere fjerde periode - dannelse (fig. 26).
I den første periode af gametogenesen deler kønsceller sig intensivt mitotisk, og deres antal stiger. I denne periode Kønscellerne kaldes henholdsvis spermato- og oogonier. For nemheds skyld viser diagrammet tilfældet, når det haploide sæt kun omfatter 3 kromosomer, hvoraf det ene er seksuelt - gonosom eller heterochromosom (på græsk betyder "heteros" forskelligt) og to ikke-seksuelle - autosomer. I celler er et haploid sæt sortfarvet, det andet er givet konturlinjer. Homologe, utvetydige autosomer af begge sæt tegnes samme form og størrelser (dette er den længste linje og cirkel). Heterokromosomer er afbildet forskelligt - med en lige linje (X-kromosom) og en buet linje af samme længde (Y-kromosom). Kromosomerne ligger separat i cellen.
Kønsceller, der er gået ind i vækstperioden, betegnes som spermato- og oocytter af første orden. De øges i størrelse, især oocytter, og deres nukleare apparat gennemgår omstrukturering. Homologe kromosomer ligger parallelt med hinanden og danner bivalente, hvis antal er lig med antallet af kromosomer i det haploide sæt. Hver af de to kromosomer af den bivalente er til gengæld en parstruktur - en dyade, da den består af to søsterkromatider. Når afstanden mellem disse kromatider bliver veldefineret, ligner de bivalente allerede tetrader. Antallet af tetrader svarer til det haploide antal kromosomer. Det samlede antal påviselige kromatider - fremtidige kromosomer af modne kønsceller - er tetraploide. Mængden af DNA i spermato- og oocytter før modningsperioden er også tetraploid.
Så kommer en modningsperiode, som er karakteriseret ved meiose ("meiose" betyder reduktion) - to hurtigt næste ven efter den anden delinger af kønsceller, hvorunder der sker kromosomreduktion. Mandlige kønsceller, der har fuldført den første modningsdeling, kaldes spermatocytter af anden orden eller prespermatider, og de tilsvarende kvindelige kønsceller kaldes oocytter af anden orden Efter anden modningsdeling bliver prespermatider til operamatider, oocytter af anden orden bliver til modne æg Den første deling af modningen er reduktion Under denne deling fordeles hele kromosomer - dyader - mellem dattercellerne og kromosomsættet bliver haploid.Anden deling af modningen kaldes udligning, ligning, da halvdelene af kromosomerne (dyader), i det væsentlige deres kromatider, divergerer mellem dattercellerne.
Interfasen mellem første og anden modningsdeling kan være meget kort eller endda helt fraværende, da der her på nuværende tidspunkt hverken sker DNA-replikation eller fordobling af antallet af kromosomer i cellerne. Delingen begynder igen, nu af to celler, og hver af barnebarnscellerne modtager et kromosom fra dyaden. Således modtager hver af disse fire celler, som er et resultat af 2 modningsdelinger, et af tetradens elementer. Det haploide antal tetrader svarer til det haploide antal i kromosomsættet af celler, der har gennemgået meiose. På samme måde bliver mængden af DNA, der er tetraploid i begyndelsen af meiosen (i oocytten og spermatocytten af første orden) efter deling i 4 dele haploid i slutningen af den.
Under spermatogenesen opnås fra hver spermatogonia, der er gået ind i vækstperioden, 4 fuldgyldige spermatider som følge af modningsdelinger. Under oogenese bevarer den anden ordens oocyt, der kommer ud fra en førsteordens oocyt, næsten hele størrelsen af modercellen. Den anden dattercelle modtager halvdelen af moderens kromosommateriale og kun en ubetydelig del af sit cytoplasma. Denne lille celle kaldes reduktionslegemet. Et lignende billede gentages under den anden modningsdeling - en andenordens oocyt giver anledning til et æg, der er omtrent lige stort og et andet reduktionslegeme. Samtidig deler det første reduktionslegeme sig også i to. Som et resultat, fra en oogonia, som er gået fra reproduktionsperioden til vækstperioden, og derefter ind i modningsperioden, dannes kun et modent æg. Dette er en fordelagtig tilpasning til forplantning - hele tilførslen af næringsstoffer akkumuleret af den første ordens oocyt i vækstperioden forbliver i ægget, hvilket er nødvendigt for at sikre indledende faser udvikling af fremtidens embryo.
I dannelsesperioden omdannes sædcellen i en typisk celle til en sædcelle, som har en meget kompleks struktur, der sikrer sin rolle som en aktiv, mobil partner i befrugtningshandlingen. Alle komponenter i spermatiden deltager i denne omstrukturering.
Først og fremmest er dens centrioler placeret efter hinanden og bestemmer således den fremtidige sperms lange akse (fig. 27A). Blandt de mange små proakrosomale granuler, der er dannet i spermatocytten - i midten af dens Golgi-apparat, som nu bevæger sig til den forreste ende af cellen (fig. 27B-3), fremkommer et stort akrosomalt (“acron” betyder spids) granulat, som så ligger ved kernen i stedet for sin fremtidige hovedstang (fig. 27B-9). Det lamellære kompleks reduceres, hvilket giver anledning til en akroblastvesikel, der omslutter det akrosomale granula. Spermatidens krop begynder gradvist at forlænge, og cellekernen bliver mere og mere tæt. Det er placeret i den forreste ende af den udviklende sædcelle. Den proksimale centriole ligger bag kernen, og den distale danner en flagel, som et kinetosom. Derefter er det opdelt i to dele, og bagsiden af dem har form af en ring (fig. 27B-8) og bevæger sig væk fra fronten og glider langs den voksende flagelum - den fremtidige aksiale filament af sædhalen. Ringen bliver hængende ved cellens bagkant. På dette tidspunkt er mitokondrier allerede for det meste akkumuleret nær det aksiale gevind. Akroblasten, der vokser, bevæger sig i form af en hætte ind på den forreste del af kernen.
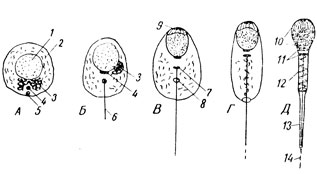
Ved slutningen af dannelsesperioden kommer differentieringen i sektioner godt til udtryk i spermen: hovedet, som i grunden er en fladtrykt og meget kompakt kerne, foran dækket af en kappe, hvorunder akrosomet ligger helt i kanten; en hals dannet af centrioler; mellemliggende, forbindende sektion og hale. Alle mitokondrier er koncentreret i den mellemliggende sektion, som spiraler rundt om den aksiale filament. Den distale grænse af den mellemliggende sektion er den lukkende centriolring. I cytoplasmaet er der relativt mange stoffer (glykogen, lipider), på grund af hvis nedbrydning spermen delvist får energi til bevægelse. I halen, når de undersøges under et optisk mikroskop, skelnes der mellem to sektioner - den vigtigste, dækket af cytoplasma, og den terminale, "bar", bestående kun af halefilamentet.
Ved hjælp af et elektronmikroskop blev det fastslået, at den tynde hætte, der dækker sædkernen, er en fladtrykt cisterne. Dens indre membran støder op til kernen, og dens ydre membran støder op til plasmalemmaet. På bagsiden af sædhovedet dækker plasmalemmaet direkte kernen. Selve kernen er meget tæt fyldt med sammenflettede tråde 40 Å tykke, som er DNP (nukleohiston) molekyler. Kemisk analyse kondenseret kromatin af kernen viser, at den består af cirka halvt DNA og halvt protein. Centriolerne placeret i halsregionen har en struktur, der er typisk for disse organeller. Den mindste af dem, støder op til bagsiden af kernen i dens midterste del, har udseende af en cylinder dannet af 9 par rør. Den distale centriole er mere udviklet. Den er ligesom en kinetochore i forbindelse med 9 par filamentkanter af flagellen, som indeholder 2 mere centrale, ligesom alle flimmerhår og flageller generelt.
Mitokondrierne, der ligger tæt i den mellemliggende sektion omkring den aksiale filament, danner cirka 14 vindinger af spiralen. Tilsyneladende giver de først og fremmest energi til de kontraktile elementer i sæden. I halens hovedsektion er 9 par aksiale filamenter omgivet af ringfibriller, som holdes sammen af to langsgående tykke tråde. I den terminale sektion af halen er der ingen ringfibriller; bundtet af aksiale fibriller, der danner det aksiale filament, er indesluttet i en homogen masse og dækket på overfladen af et plasmalemma. Her støder vi således på en struktur, der er karakteristisk for alle cilia og flageller.
Den samlede længde af sædceller hos mennesker når cirka 60 mk. Den bevæger sig aktivt med en hastighed på 3,5 mm i minuttet. Samtidig roterer den rundt om sin akse med uret og producerer en fuld omgang på 15 minutter. Dens evne til at bevæge sig afhænger af pH og andre egenskaber i miljøet. Levetiden for sædceller i skeden er kun 1 time; i andre kvindelige kønsorganer varer den flere dage. Langs den kvindelige kønsorganer bevæger sæden sig passivt (som følge af muskelsammentrækninger af livmoderens vægge og æggeledere), når den øverste tredjedel af æggelederne, hvor befrugtning sker. Dermed behøver han ikke at spilde energi fra sin generelt ubetydelige reserve for at overvinde en så lang vej for ham.
En moden ægcelle har en sfærisk form; dens diameter hos mennesker er 135 mk. Den er altid dækket af en mikroskopisk synlig membran, og dens struktur adskiller sig fra somatiske celler, hovedsageligt på to måder. For det første er det nuklear-cytoplasmatiske forhold i det mere eller mindre skarpt forskudt til fordel for cytoplasmaet, hvilket forklares af akkumuleringen i oocyttens krop i perioden med vækst af næringsstoffer til det fremtidige embryos behov. For det andet mangler den et cellecenter, som forsvinder i samme vækstperiode. Cellecenter introducerer en sædcelle i ægget under befrugtningen, og herefter begynder den mitotiske deling af zygoten - en organisme placeret på det encellede stadium af ontogenese.
Lad os kort berøre spørgsmålet om kønsbestemmelse, som opstår efter befrugtning af organismen. Som det viste sig, sker dette i tidspunktet for undfangelsen, det vil sige befrugtning, og er forårsaget af kombinationen af heterokromosomer i zygoten. Lad os se på diagrammet (fig. 26) og betragte kromosomsættene i udviklende kvindelige og mandlige kønsceller. Alle oogonier har to X-kromosomer. Det følger, at det haploide sæt af kromosomer af hvert modent æg nødvendigvis vil omfatte et af X-kromosomerne. I spermatogoni er der blandt de to heterokromosomer et X-kromosom og et Y-kromosom. Derfor skal sædcellerne være anderledes – halvdelen med et X-kromosom og halvdelen med et Y-kromosom.
Hos pattedyr og mennesker, når to X-kromosomer kombineres i en zygote, udvikles en kvindelig organisme; i nærværelse af XY-kromosomer udvikles en mandlig organisme. Det ufødte barns køn vil således afhænge af, hvilken af faderens sæd, moderens æg forbinder med (fig. 28). Det humane haploide sæt indeholder et eller andet af heterokromosomer (kønskromosomer - gonosomer) og 22 autosomer (ikke-kønskromosomer). Der skal være 46 kromosomer i et menneskes zygote, embryonale, somatiske og umodne kønsceller - 44+XY i den mandlige krop og 44+XX i den kvindelige. Først efter den første meiotiske deling i de modnende kønsceller reduceres antallet af kromosomer til 23. At implementere normal udvikling nødvendig tilgængelighed diploid sæt af kromosomer uden nogen afvigelser, både kvantitative og strukturelle.
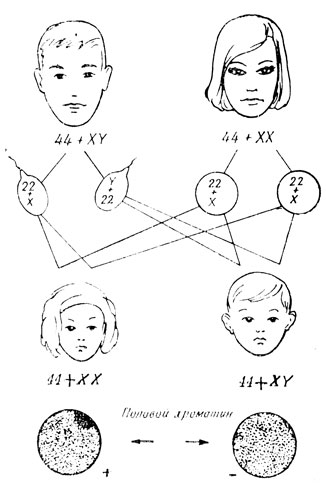
I 1949 fastslog Barr, at hos hunpattedyr og kvinder opfører X-kromosomerne i det diploide sæt sig forskelligt - et af dem, efter celledeling, despiraler, ligesom autosomer, og bliver umuligt at skelne i interfasekernen, det andet forbliver stærkt spiralformet, som f.eks. heterokromatiske kromomerer. Dette heterokromatiske X-kromosom, der er tydeligt synligt i kernen i form af en mørk krop i den sædvanlige farvning af præparater, kaldes kønskromatin. I genetiske eksperimenter for at identificere Barr-legemer (kønskromatin-legemer) er det nemmest at undersøge blodlymfocytter eller afskallede epitelceller. Det viste sig, at et af de to X-kromosomer kunne blive inaktive.
I i sjældne tilfælde Under oogenese adskilles X-kromosomerne ikke under meiosen. Som følge heraf kan der dannes æg, der afviger fra normen: i stedet for et X-kromosom kan der være to, eller der kan slet ikke være noget X-kromosom. I sidstnævnte tilfælde Under befrugtningen kan der opstå to typer zygoter, der i det diploide sæt indeholder enten et Y-kromosom eller et X-kromosom, der indføres i ægget under befrugtning af en sædcelle. En zygote med et Y-kromosom er generelt ikke levedygtig og dør. En zygote med ét X-kromosom hos mennesker har 45 kromosomer: 44+XO. Med denne kombination af kromosomer udvikler zygoten sig til en ringere kvinde af lille statur med rudimentære æggestokke og som følge heraf fraværet af sekundære seksuelle karakteristika. Denne patologi er kendt som Turners syndrom. Det eneste X-kromosom i dette tilfælde er despiraliseret, og derfor påvises kønskromatin ikke i somatiske celler hos sådanne piger.
Resultatet af ikke-disjunktion af X-kromosomer under meiose vil også være afvigelser i sættet af den modsatte natur, nemlig tilstedeværelsen af tre X-kromosomer eller to X-kromosomer og et Y-kromosom i zygoten. En kvindelig organisme med tre X-kromosomer omtales som en "superkvinde" eller "superkvinde" (for mennesker). Et sådant individ kaldes dog kun en "superkvinde" kun betinget, baseret på det overtallige antal X-kromosomer. I virkeligheden, med et sæt af kromosomer 44 + XXX, er der underudvikling af æggestokkene og derfor ofte et tab af fertilitet. Interessant nok er "superkvinder" med fire X-kromosomer (kromosomalt sæt 44 + XXXX) fertile, men har en reduceret mental udvikling. Ved analyse af kønskromatin i somatiske celler hos kvinder med XXX- eller XXXX-kromosomer findes henholdsvis 2 eller 3 Barr-legemer i kernerne. I disse tilfælde er kun ét af alle X-kromosomer således despiraliseret og aktivt i interfasekerner.
Zygoter med et kromosomsæt af type 44+XXY udvikles hos mænd, der lider af Klinefelters syndrom - mental retardering og underudvikling af testiklen, hvilket fører til infertilitet. Deres somatiske celler har kønschromatin og indeholder 1 Barr krop. Andet X, det samme som y normale mænd, fortvivlet i interfase. Lignende udviklingsdefekter observeres i mere X-kromosomer kombineret med et Y-kromosom, nemlig i typerne XXXY, XXXXY og XXXXXY. Tilstedeværelsen af Y-kromosomet i sættet bestemmer udviklingen af et mandligt individ, men et ringere. Antal Barr-legemer i interfasekerner af somatiske celler pr. enhed mindre antal X-kromosomer i et sæt.
Relativt for nylig blev en anden kromosomal anomali opdaget - type XYY. Dyr er "super hanner" med ekstra Y-kromosom afvige stor styrke og aggressivitet. Mænd med to Y-kromosomer er forskellige høj(over 180 cm), stor fysisk styrke, men reduceret mentale evner. Som hos normale mænd indeholder interfasekernerne i deres somatiske celler ikke kønskromatin.
Forskere har fundet ud af, at arven af nogle sygdomme er forbundet med gener placeret på kønskromosomerne. For eksempel overføres medfødte farvesynsforstyrrelser (en sygdom tidligere kaldet farveblindhed), generelt blindhed forårsaget af atrofi af synsnerven, hæmofili (patologiske problemer med at stoppe blødning) gennem X-kromosomet.
Det fænomen, hvor der er ekstra kromosomer i et sæt, eller nogle af kromosomerne mangler, kaldes aneuploidi. Tilstedeværelsen af et ekstra kromosom kaldes trisomi; hvis der er to sådanne ekstra kromosomer, er det dobbelt trisomi. Hvis et kromosom mangler, kaldes det monosomi.
Et eksempel på lidelser forbundet med en stigning i antallet af autosomer er den mest berømte trisomi - en af de mindste kromosomer - den 21. Tilstedeværelsen af 21 kromosomer i kromosomsættet i stedet for to forekommer ved Downs syndrom - en af formerne mental retardering, kombineret med forsinkelser og forstyrrelser fysisk udvikling og nogle gange med tilstedeværelsen af visse deformiteter ( udseende patienter ligner meget og er karakteriseret ved et lille kranium, fladt baghoved, skrå øjenform, bred nedsunket næserygge, halvåben mund, deforme ører). Kønsorganerne er underudviklede, og sekundære seksuelle karakteristika er dårligt udtrykt. Halvdelen af børn med Downs syndrom lever ikke længere end 2 år.
Mange sygdomme er blevet opdaget, der er forårsaget af gener placeret på forskellige autosomer. Blandt dem kan nævnes arvelige psykiske sygdomme, såsom skizofreni og epilepsi.
Alle former for arvelige menneskelige sygdomme forårsaget af forskellige lidelser genotype er genstand for undersøgelse af den i øjeblikket intensivt udviklede gren af cytogenetik - medicinsk genetik.
Husk fra lærebogen “Planter. Bakterie. Svampe og lav,” hvilke processer karakteriserer en celles liv. Hvad er opbygningen af cellekernen? Hvad er kromosomer? Hvad er strukturen af et DNA-molekyle? Hvad er DNA-reduplikation?
Perioden med celleaktivitet fra dets oprindelse til døden kaldes cellelivscyklus eller cellecyklus. I denne periode sker cellevækst, udvikling og reproduktion. Cellecyklus varighed i forskellige celler selv i den samme organisme er forskellig. For eksempel er varigheden af denne cyklus i humane epitelvævsceller omkring 10-15 timer og i leverceller helt år. Cellecyklussen består af to intervaller af forskellig varighed: interfase og celledeling (fig. 66).
Ris. 66. Cellelivscyklus (cellecyklus): 1 - interfase; 2 - mitose
Interfase. En del livscyklus celler mellem to på hinanden følgende delinger kaldes interfase (fra latin inter - mellem og den græske fase - udseende). Det er karakteriseret aktive processer metabolisme, proteinbiosyntese, nukleinsyrer, kulhydrater og lipider. I interfase opstår processer forbundet med cellens liv - dissimilering og assimilering. Energiforsyningen i cellen øges på grund af syntesen af ATP. Alle typer RNA syntetiseres aktivt i kernen, og ribosomer dannes og samles i nukleolen. Cellen gennemgår intensiv vækst, og antallet af alle dens organeller stiger.
Hovedbegivenheden i interfase er DNA-reduplikation - dens selvduplikation. Sådan forbereder cellen sig på at dele sig.
Varigheden af interfasen afhænger af celletypen og er i gennemsnit mindst 90 % af den samlede tid af cellecyklussen. Efter afslutningen af interfasen går cellen ind næste del cyklus - division.
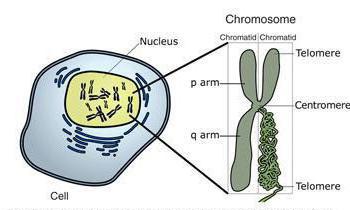
Kromosomernes struktur. Vigtig rolle i cellecyklussen hører til kromosomer. Kromosom er et kompleks af spiraliserede DNA-molekyler og proteiner (fra det græske chromo - farve og somo - krop). De regulerer ikke kun alle metaboliske processer i cellen, men sikrer også transmissionen arvelige oplysninger fra en generation af celler og organismer til en anden. En prokaryot celle indeholder kun ét cirkulært DNA-molekyle, der ikke er bundet til proteiner. Derfor kan det ikke kaldes et kromosom.
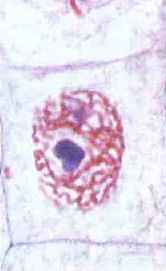
Ris. 67. Kromatintråde i cellelivscyklussens interfase
De fleste kromosomer i interfase er i form af kromatintråde, hvilket gør dem praktisk talt usynlige (fig. 67). Efter reduplicering består hvert kromosom af to DNA-molekyler, der spiraler, forbinder med proteiner og antager forskellige former. De to datter-DNA-molekyler pakkes separat og danner søsterkromatider (fra det græske chrom - farve og eidos - udseende). Søsterkromatider holdes sammen og danner ét kromosom (fig. 68). Stedet for samhørighed mellem to søsterkromatider kaldes en centromere (fra det latinske centrum - midt og meros - del).
Ris. 68. Kromosomstruktur efter DNA-reduplikation: 1 - centromer: 2 - kromosomarme; 3 - søsterkromatider; 4 - DNA-molekyle: 5 - protein
Det er kun muligt at studere formen og størrelsen af kromosomer og bestemme deres antal i en celle under deling, når de er maksimalt spiraliseret, tætpakket, godt farvet og synligt med et lysmikroskop.
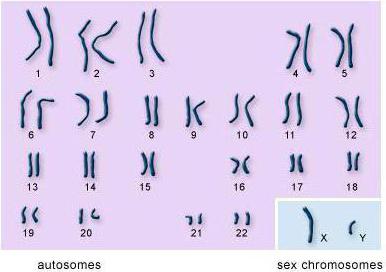
Kromosomsæt af celler. Hver organismes celler indeholder specifikt sæt kromosomer, som kaldes en karyotype (fra græsk karyon - kerne og slåfejl - prøve, form). Hver type organisme har sin egen karyotype. Kromosomer af karyotyper adskiller sig i form, størrelse og sæt genetisk information. Kromosomsættet er strengt individuelt for hver type organisme. Den menneskelige karyotype består således af 23 par kromosomer (fig. 69), frugtfluen Drosophila har 4 par kromosomer, og en af hvedearterne har 14 par.
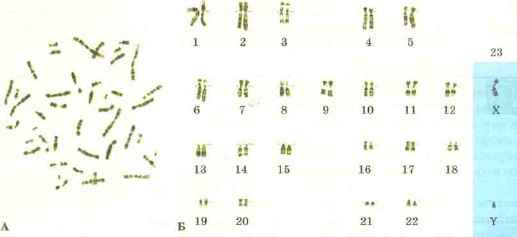
Ris. 69. Kromosomsæt af humane celler: A - generelt fotografi; B - 23 par kromosomer
Karyotype undersøgelser forskellige organismer viste, at deres celler kan indeholde dobbelte og enkelte sæt kromosomer.
Et dobbelt sæt kromosomer består altid af parrede kromosomer, der er identiske i størrelse, form og art af arvelig information. Parrede kromosomer kaldes homologe (fra det græske homos - identiske). Alle ikke-reproduktive humane celler indeholder således 23 par kromosomer, dvs. 46 kromosomer præsenteres i form af 23 par. I Drosophila danner 8 kromosomer 4 par. Parrede homologe kromosomer ligner meget hinanden i udseende. Deres centromerer er de samme steder, og deres gener er placeret i samme sekvens.
Nogle celler kan have et enkelt sæt kromosomer. For eksempel i celler lavere planter- encellede grønne alger har et enkelt sæt kromosomer, hvorimod højere planter og dyr er det dobbelt. Dyres kønsceller har også et enkelt sæt kromosomer. I dette tilfælde er der ingen parrede kromosomer, der er ingen homologe kromosomer, men der er ikke-homologe. Således indeholder menneskelige kønsceller 23 kromosomer. Desuden adskiller det kromosomale sæt af mandlige og kvindelige kønsceller sig i det 23. kromosom. Det ligner i formen de latinske bogstaver X eller Y. Spermatozoer kan have et X- eller Y-kromosom. Æg bærer altid X-kromosomet.
Kromosomsættet betegnes normalt latinsk bogstav s. Det dobbelte sæt betegnes derfor 2p, og det enkelte sæt betegnes p.
Øvelser baseret på det gennemgåede materiale
- Definer en celles livscyklus (cellecyklus).
- Hvad er inferfase? Hvilken begivenhed er den vigtigste i interfase? Begrund dit svar.
- Hvor mange DNA-molekyler består kromosomet af i begyndelsen af interfase og før celledeling?
- Hvordan bestemmes antallet og formen af kromosomer? forskellige typer organismer?
- Hvordan adskiller et dobbelt sæt kromosomer sig fra et enkelt sæt?
- Der er 44 kromosomer i karyotypen af en kanin. Hvor mange kromosomer er der i en kanins ikke-reproduktive celler, og hvor mange er der i kønscellerne?
Vi vil finde svaret på dette spørgsmål og også bestemme den betydning, de har for levende organismer. Hvad er mekanismen for deres placering og konstruktion?
Et lille tilbagetog
Kromosomer er vigtig del genetisk mekanisme. De fungerer som et DNA-lager. Nogle vira har enkeltstrengede molekyler, men i de fleste tilfælde er de dobbeltstrengede og er lineære eller lukkede i en ring. Men DNA er placeret i kromosomer udelukkende i cellulære organismer. Det vil sige, at denne opbevaring i vira ikke bruges i sædvanlig forstand, da mikroorganismen selv spiller en sådan rolle. Når de er oprullet, placeres molekylerne mere kompakt. Kromosomer består af kromatin. Dette er en speciel fiber, der dannes, når eukaryotisk DNA vikler sig om specielle proteinpartikler kaldet histoner. De er placeret med et bestemt interval, så strukturen er stabil.
Om kromosomer
 De er de vigtigste strukturelle elementer cellekerne. På grund af evnen til at reproducere sig selv, kan kromosomer give genetisk forbindelse mellem generationer. Det skal bemærkes forskellen i deres længde hos forskellige dyr og mennesker: deres størrelse kan variere fra fraktioner til snesevis af mikron. Som kemisk grundlag konstruktion bruger nukleoproteiner, der er dannet af proteiner som protaminer og histoner. Kromosomer er løbende placeret i Og det gælder alle evt højere former liv. Så ovenstående udsagn om, hvor kromosomerne er placeret i dyrecelle, med nøjagtig samme tillid kan tilskrives planter. Kig ud af vinduet. Hvilke træer kan du se bagved? Lind, eg, birk, valnød? Eller måske ribs- og hindbærbuske? Når vi besvarer spørgsmålet om, hvor kromosomerne er placeret i de planter, der blev listet, kan vi sige, at de er på samme sted som i dyreorganismer - i
De er de vigtigste strukturelle elementer cellekerne. På grund af evnen til at reproducere sig selv, kan kromosomer give genetisk forbindelse mellem generationer. Det skal bemærkes forskellen i deres længde hos forskellige dyr og mennesker: deres størrelse kan variere fra fraktioner til snesevis af mikron. Som kemisk grundlag konstruktion bruger nukleoproteiner, der er dannet af proteiner som protaminer og histoner. Kromosomer er løbende placeret i Og det gælder alle evt højere former liv. Så ovenstående udsagn om, hvor kromosomerne er placeret i dyrecelle, med nøjagtig samme tillid kan tilskrives planter. Kig ud af vinduet. Hvilke træer kan du se bagved? Lind, eg, birk, valnød? Eller måske ribs- og hindbærbuske? Når vi besvarer spørgsmålet om, hvor kromosomerne er placeret i de planter, der blev listet, kan vi sige, at de er på samme sted som i dyreorganismer - i
Kromosomernes placering i en celle: hvordan valget træffes
 En flercellet eukaryot er ejer af Den er sammensat af genomet af faderen og moderen. Takket være meioseprocessen konjugerer de med hinanden. Dette sikrer processen med at udveksle områder - krydse over. Parring er mulig i disse tilfælde, hvilket er nødvendigt for at sikre funktionen af gener i celler, der ikke deler sig, men er i dvaletilstand. Konsekvensen af dette er, at kromosomerne er i kernen, og for at fortsætte funktionerne i delinger, må de ikke forlade dens grænser. Det er selvfølgelig ikke svært at finde nukleotidrester i selve cellen. Men i de fleste tilfælde er dette enten genomet i mitokondrier eller enkelte dele af helheden, der er brudt af og nu er "frit svævende." Det er meget svært at finde et komplet kromosom uden for kernen. Og sker det, skyldes det udelukkende fysiske skader.
En flercellet eukaryot er ejer af Den er sammensat af genomet af faderen og moderen. Takket være meioseprocessen konjugerer de med hinanden. Dette sikrer processen med at udveksle områder - krydse over. Parring er mulig i disse tilfælde, hvilket er nødvendigt for at sikre funktionen af gener i celler, der ikke deler sig, men er i dvaletilstand. Konsekvensen af dette er, at kromosomerne er i kernen, og for at fortsætte funktionerne i delinger, må de ikke forlade dens grænser. Det er selvfølgelig ikke svært at finde nukleotidrester i selve cellen. Men i de fleste tilfælde er dette enten genomet i mitokondrier eller enkelte dele af helheden, der er brudt af og nu er "frit svævende." Det er meget svært at finde et komplet kromosom uden for kernen. Og sker det, skyldes det udelukkende fysiske skader.
Kromosom sæt
 Dette er navnet på hele sættet af kromosomer, der er til stede i cellekernen. Alle har det biologiske arter har sit eget konstante og karakteristiske sæt, som blev konsolideret under evolutionen. Det kan være af to typer: enkelt (eller haploid, fundet hos dyr) og dobbelt (eller diploid). Sættene adskiller sig i antallet af kromosomer, de indeholder. Så hos heste er deres nummer to. Men i protozoer og nogle spore planter deres antal kan nå tusindvis. Forresten, hvis vi taler om, hvor kromosomerne er placeret i bakterier, skal det bemærkes, at de som regel også er placeret i kernen, men det er også muligt, at de "frit" flyder i cytoplasmaet. Men det gælder udelukkende for encellede organismer. Desuden adskiller de sig ikke kun i mængde, men også i størrelse. En person har 46 kromosomer i sit sæt.
Dette er navnet på hele sættet af kromosomer, der er til stede i cellekernen. Alle har det biologiske arter har sit eget konstante og karakteristiske sæt, som blev konsolideret under evolutionen. Det kan være af to typer: enkelt (eller haploid, fundet hos dyr) og dobbelt (eller diploid). Sættene adskiller sig i antallet af kromosomer, de indeholder. Så hos heste er deres nummer to. Men i protozoer og nogle spore planter deres antal kan nå tusindvis. Forresten, hvis vi taler om, hvor kromosomerne er placeret i bakterier, skal det bemærkes, at de som regel også er placeret i kernen, men det er også muligt, at de "frit" flyder i cytoplasmaet. Men det gælder udelukkende for encellede organismer. Desuden adskiller de sig ikke kun i mængde, men også i størrelse. En person har 46 kromosomer i sit sæt.
Kromosommorfologi
 Det er direkte relateret til deres spiralisering. Så når de er i interfase-stadiet, er de mest udviklede. Men i begyndelsen af delingsprocessen begynder kromosomerne intensivt at forkortes gennem deres spiralisering. Største grad Denne tilstand opstår på metafasestadiet. Relativt korte og tætte strukturer dannes på den. Metafasekromosomet er dannet af to kromatider. De består til gengæld af såkaldte elementære filamenter (chromonemata).
Det er direkte relateret til deres spiralisering. Så når de er i interfase-stadiet, er de mest udviklede. Men i begyndelsen af delingsprocessen begynder kromosomerne intensivt at forkortes gennem deres spiralisering. Største grad Denne tilstand opstår på metafasestadiet. Relativt korte og tætte strukturer dannes på den. Metafasekromosomet er dannet af to kromatider. De består til gengæld af såkaldte elementære filamenter (chromonemata).
Individuelle kromosomer
De skelnes afhængigt af placeringen af centromeren (primær indsnævring). Hvis denne komponent går tabt, så mister kromosomerne evnen til at dele sig. Så den primære forsnævring deler kromosomet i to arme. Sekundære kan også dannes (i dette tilfælde kaldes det resulterende resultat en satellit). Hver type organisme har sine egne specifikke (i antal, størrelse eller form) sæt af kromosomer. Hvis den er dobbelt, er den betegnet som en karyotype.
Kromosomal teori om arvelighed
Disse bærere blev først beskrevet af I.D. Chistyakov i 1874. I 1901 henledte Wilson opmærksomheden på tilstedeværelsen af parallelisme i deres adfærd. Han fokuserede derefter på Mendeleske arvelighedsfaktorer i meiose og befrugtning og konkluderede, at gener er placeret på kromosomer. Gennem 1915-1920 beviste Morgan og hans medarbejdere denne position. De lokaliserede flere hundrede gener til Drosophila-kromosomer, hvilket skabte de første genetisk kort. De data, der blev opnået på dette tidspunkt, dannede grundlag for al efterfølgende udvikling af videnskaben i i denne retning. Også baseret på disse oplysninger, en kromosomteori arvelighed, ifølge hvilken kontinuiteten af celler og hele organismer sikres takket være disse bærere. 
Kemisk sammensætning
Forskningen fortsatte, og under biokemiske og cytokemiske forsøg i 30-50'erne af forrige århundrede blev det fastslået, hvad de var sammensat af. Deres sammensætning er som følger:
- Grundlæggende proteiner (protaminer og histoner).
- Ikke-histonproteiner.
- Variable komponenter. De kan være RNA og surt protein.
Kromosomer dannes af deoxyribonukleoproteinstrenge. De kan forbindes i bundter. I 1953 blev strukturen opdaget, og mekanismen for dens auto-reproduktion blev demonteret. Opnået viden om nukleinkode, tjente som grundlag for fremkomsten ny videnskab- genetik. Nu ved vi ikke kun, hvor kromosomerne er placeret i en celle, men vi har også en idé om, hvad de er lavet af. Når de i almindelige hverdagssamtaler taler om det arvelige apparat, så mener de som regel ét DNA, men man ved nu, at det kun er dets komponent.
Kønskromosomer
De gener, der er ansvarlige for et pattedyrs køn (inklusive mennesker), findes i et særligt par. Der kan være andre tilfælde af organisering, hvor alt er bestemt af forholdet mellem hver type kønskromosom. Dyr, der har denne type definition, kaldes autosomer. Hos mennesker (og også andre pattedyr) bestemmes hunkønnet af de samme kromosomer, som betegnes som X. For mænd bruges X og Y. Men hvordan vælger man, hvilket køn barnet skal have? Til at begynde med modnes den kvindelige bærer (ægget), som X er placeret i. Og køn bestemmes altid af indholdet af spermatocytter. De indeholder både X- og Y-kromosomer i lige store forhold (plus/minus). Kønnet på det ufødte barn afhænger af bæreren, som er den første til at udføre befrugtning. Og som følge heraf kan enten en kvinde (XX) eller en mand (XY) opstå. Så vi fandt ikke kun ud af, hvor kromosomerne er placeret hos mennesker, men fandt også ud af funktionerne i deres placering og kombination, når vi skabte en ny organisme. Det er værd at bemærke, at denne proces er noget lettere for flere simple former livet, derfor, når du stifter bekendtskab med, hvad de har, og hvordan det forløber, kan du måske bemærke små forskelle fra modellen beskrevet her.
Operation
 Kromosomalt DNA kan opfattes som en skabelon, der virker til at syntetisere specifikke messenger-RNA-molekyler. Men denne proces kan kun forekomme under betingelse af despiralisering af et bestemt område. Når vi taler om muligheden for, at et gen eller et helt kromosom virker, skal det bemærkes, at visse betingelser kan være nødvendige for deres funktion. Du har sikkert hørt om insulin? Genet, der er ansvarligt for dets produktion, er til stede i alt menneskelige legeme. Men det kan aktiveres og virker kun, når det er i de nødvendige celler, der skaber bugspytkirtlen. Og der er rigtig mange af sådanne sager. Hvis vi taler om udelukkelse af et helt kromosom fra stofskiftet, så kan vi huske dannelsen af kønskromatinlegemet.
Kromosomalt DNA kan opfattes som en skabelon, der virker til at syntetisere specifikke messenger-RNA-molekyler. Men denne proces kan kun forekomme under betingelse af despiralisering af et bestemt område. Når vi taler om muligheden for, at et gen eller et helt kromosom virker, skal det bemærkes, at visse betingelser kan være nødvendige for deres funktion. Du har sikkert hørt om insulin? Genet, der er ansvarligt for dets produktion, er til stede i alt menneskelige legeme. Men det kan aktiveres og virker kun, når det er i de nødvendige celler, der skaber bugspytkirtlen. Og der er rigtig mange af sådanne sager. Hvis vi taler om udelukkelse af et helt kromosom fra stofskiftet, så kan vi huske dannelsen af kønskromatinlegemet.
Menneskelige kromosomer
I 1922 antog Peitner, at mennesker har 48 kromosomer. Dette blev selvfølgelig ikke sagt i tomt rum, men baseret på visse data. Men i 1956, forskerne Tire og Levan, når de brugte de nyeste metoder En undersøgelse af det menneskelige genom afslørede, at mennesker faktisk kun har 46 kromosomer. De gav også en beskrivelse af vores karyotype. Parrene er nummereret fra et til treogtyve. Selvom det sidste par ofte ikke er tildelt et nummer, men hver for sig kaldes det, det består af.
Konklusion
 Så gennem artiklen har vi bestemt, hvilken rolle kromosomer har, hvor de er placeret, og hvordan de er bygget. Naturligvis fik det menneskelige genom den største opmærksomhed, men dyr og planter blev også overvejet. Vi ved, hvor kromosomerne er placeret i cellen, funktionerne i deres placering samt mulige transformationer, der kan forekomme med dem. Hvis vi taler om genomet, så husk, at det kan være i andre dele, ikke kun kernen. Men hvordan barnet protesterer, er påvirket af præcis, hvad der er i kromosomerne. Desuden afhænger organismens egenskaber ikke meget af mængden af disse. Så efter at have talt om, hvor kromosomerne er placeret plantecelle og dyreorganismer, mener vi, at vores opgave er løst.
Så gennem artiklen har vi bestemt, hvilken rolle kromosomer har, hvor de er placeret, og hvordan de er bygget. Naturligvis fik det menneskelige genom den største opmærksomhed, men dyr og planter blev også overvejet. Vi ved, hvor kromosomerne er placeret i cellen, funktionerne i deres placering samt mulige transformationer, der kan forekomme med dem. Hvis vi taler om genomet, så husk, at det kan være i andre dele, ikke kun kernen. Men hvordan barnet protesterer, er påvirket af præcis, hvad der er i kromosomerne. Desuden afhænger organismens egenskaber ikke meget af mængden af disse. Så efter at have talt om, hvor kromosomerne er placeret plantecelle og dyreorganismer, mener vi, at vores opgave er løst.
Nyttige links:
Celler, kromosomer, celledeling.
Celler, kromosomer, celledeling. Kroppen af hver voksen indeholder mere end hundrede mio celler, mikroskopiske strukturer, der kun når en hundrededel af en millimeter i diameter. Ingen celle ude af stand til at overleve uden for kroppen, medmindre den specifikt dyrkes i en kunstig opløsning.
Kroppens celler varierer i form, størrelse og struktur alt efter den funktion, de udfører. Muskelceller, som for eksempel er lange og tynde, kan trække sig sammen og slappe af og derved tillade kroppen at bevæge sig. Mange nerveceller også lange og tynde, men de er designet til at transmittere impulser, der udgør budskaberne nervesystem, mens leverens sekskantede celler er udstyret med alt nødvendigt for at udføre vitale kemiske processer. Doughnut-formede røde blodlegemer bærer ilt og kuldioxid, mens sfæriske pancreasceller producerer og reparerer hormonet insulin.
På trods af disse variationer er alle celler i kroppen konstrueret efter det samme grundmønster. Langs overfladen af hver celle er der en slags grænsevæg, eller celle membran, indeholdende et gelélignende stof - cytoplasma. Inde i den er cellens kerne, hvor kromosomerne er indeholdt. Cytoplasmaet, selvom det indeholder fra 70 til 80 procent vand, spiller en langt fra passiv rolle. Der sker forskellige processer mellem stoffer opløst i vand kemiske reaktioner; Derudover indeholder cytoplasmaet mange de mindste strukturer, kaldet organeller, som spiller en vigtig rolle.
Celledele
Nogle celler har hårlignende processer kaldet cilia på deres membraner. I næsen, for eksempel, opfanger cilia støvpartikler. Disse cilia kan bevæge sig i bølger i én retning og dirigere ethvert stof.
Cytoplasmaet i alle celler indeholder mikroskopiske, pølseformede organer kaldet mitokondrier, som omdanner ilt og næringsstoffer til den energi, der er nødvendig for alle cellulære aktiviteter.
Disse "energihuse" arbejder ved hjælp af enzymer - komplekse proteiner, der fremskynder kemiske reaktioner i celler og er de mest talrige grundstoffer i muskelceller.
Lysosomer, en anden type mikroskopisk organ i cytoplasmaet, er bittesmå sække fyldt med enzymer, der gør det muligt for cellen at behandle næringsstoffer. De fleste af dem findes i leverceller.
Stoffer produceret af cellen, som er nødvendige for andre dele af kroppen, såsom hormoner, akkumuleres først og lagres derefter i andre bittesmå organer kaldet Golgi-apparatet (intracellulært retikulært apparat).
Mange celler har et helt system af små rør, der betragtes som en slags indre "skelet" i cellen, men alle celler indeholder et system af kanaler - et endoplasmatisk retikulum.
Langs hele maskedannelsen er små sfæriske strukturer kaldet ribosomer, som er ansvarlige for at regulere dannelsen af grundlæggende proteiner, der er nødvendige for alle celler. Proteiner er nødvendige til reparation af strukturer og (i form af enzymer) for kemiske processer i bur og produktion komplekse molekyler såsom hormoner.
Kromosomer
Ud over modne røde blodlegemer, som mister deres kromosomer i de sidste stadier af dannelsen, og æg og sædceller (kønsceller), som indeholder halvdelen af det normale antal kromosomer, indeholder hver celle i kroppen 46 kromosomer, organiseret i 23 par . Det ene kromosom kommer fra moderen, det andet fra faderen. Æg og sæd har kun halvdelen af denne mængde, så under processen med befrugtning af ægget kan det nye væsen garanteres at have det nødvendige antal kromosomer.I befrugtningsøjeblikket begynder generne at give instruktioner til modellering! ny menneske. Faderens kromosomer er ansvarlige for at bestemme køn. Kromosomerne kaldes X og Y, afhængig af deres form. Hos kvinder er begge kromosomer i et par X, men hos mænd er det ene kromosom X, og det andet er Y. Hvis en sæd, der indeholder X, befrugter et æg af X, vil barnet være en pige, men hvis en sæd, der indeholder Y, befrugtes ægget, så bliver barnet en dreng.
Celledeling
Sammen med det faktum, at DNA bærer information, har det også evnen til at reproducere; Uden dette kunne celler hverken duplikere eller transmittere information fra en generation til den næste.
Processen med celledeling, hvor den fordobles, kaldes mitose; dette er en form for deling, der opstår, når et befrugtet æg først vokser til et barn, derefter til en voksen, og når de brugte celler udskiftes. Når en celle ikke deler sig, er kromosomerne ikke synlige i kernen, men når cellen begynder at dele sig, bliver kromosomerne kortere og tykkere, og de ses så at dele sig i to på langs. Disse dobbeltkromosomer adskilles derefter fra hinanden og bevæger sig til modsatte ender af cellen. På det sidste stadie opdeles cytoplasmaet i køn, og der dannes nye vægge omkring to nye celler, som hver har normalt antal kromosomer - 46.
Hver dag dør et enormt antal celler og erstattes af mitose; nogle celler er mere aktive end andre. Når først de er dannet, kan hjerne- og nerveceller ikke udskiftes, men lever, hud og blodceller udskiftes fuldstændigt flere gange om året.
At skabe celler med halvdelen af antallet af kromosomer for at bestemme arvelige egenskaber, kræver en anden delingsmetode, den kaldes meiose. Med denne celledelingsmetode bliver kromosomerne først, som ved mitose, kortere og tykkere og deles i to, men derefter deles kromosomerne i par, så en fra moderen og en fra faderen ligger ved siden af hinanden.
Så hænger kromosomerne meget tæt sammen, og når de fra tid til anden er adskilt fra hinanden, indeholder hvert nyt kromosom allerede flere gener fra moderen og flere gener fra faderen. Herefter deler de to nye celler sig igen, så hvert æg eller sæd indeholder de 23 kromosomer, de skal bruge. Denne udveksling af genetisk materiale gennem meioseprocessen forklarer, hvorfor børn ikke er nøjagtigt som deres forældre, og hvorfor hver person, undtagen enæggede tvillinger, har en unik genetisk sammensætning.