МЕРИСТЕМИ АБО ОСВІТИЛЬНІ ТКАНИНИ
цитоплазма. Хімічний склад, фізичний стан, будова та властивості. Надходження речовин у клітину. Типи руху цитоплазми.
Цитоплазма безбарвна, має слизову консистенцію і містить різні речовини, у тому числі високомолекулярні сполуки, наприклад білки, присутність яких обумовлює колоїдні властивості цитоплазми. Цитоплазма – частина протопласту, укладена між плазмалемою та ядром. Основу цитоплазми становить її матрикс, або гіалоплазма - складна безбарвна, оптично прозора колоїдна система, здатна до оборотних переходів із золю в гель.
У цитоплазмі рослинних клітинє органоїди: невеликі тільця, що виконують спеціальні функції, - пластиди, комплекс Гольджі, ендоплазматична мережа, мітохондрії і т.д. У цитоплазмі здійснюється більша частина процесів клітинного метаболізму, крім синтезу нуклеїнових кислот, що відбуваються в ядрі. Цитоплазма пронизана мембранами – найтоншими (4-10нм) плівками, побудованими переважно з фосфоліпідів та ліпопротеїнів. Мембрани обмежують цитоплазму від клітинної оболонки та вакуолі і всередині цитоплазми утворюють ендоплазматичну мережу (ретикулум) – систему дрібних вакуолей та канальців, з'єднаних один з одним.
Найважливішою властивістю цитоплазми, пов'язаною насамперед із фізико-хімічними особливостями гіалоплазми, є її здатність до руху. У клітинах з однією великою вакуоллю рух здійснюється зазвичай у одному напрямі (циклоз) з допомогою спеціальних органоїдів – микрофиламентов, що становлять нитки особливого білка - актина. Рухаюча гіалоплазма захоплює пластиди та мітохондрії. Клітинний сік, що знаходиться у вакуолях, є водний розчин різних речовин: білків, вуглеводів, пігментів, органічних кислот, солей, алкалоїдів і т.п. Концентрація речовин, що знаходяться в клітинному соку, зазвичай вища за концентрацію речовин у зовнішньому середовищі (ґрунт, водойми). Відмінність концентрацій певною мірою обумовлює можливість надходження в клітину води та ґрунтових розчинів, що до певної міри пояснюється явищем осмосу. У клітині роль напівпроникної мембраниграє цитоплазма. Прикордонні шари цитоплазми, що вистилають оболонку та клітинну вакуоль, проникні тільки для води та деяких розчинів, але непроникні для багатьох розчинених у воді речовин. Ця властивість цитоплазми одержала назву напівпроникності або вибіркову проникність. На відміну від цитоплазми клітинна оболонка проникна всім розчинів, непроникна вона лише твердих частинок. Надходження речовин у клітину не можна зводити лише до осмотичних явищ, що виражені у дорослих клітинах із добре розвиненими вакуолями. Насправді це дуже складний процес, обумовлений багатьма факторами Активну участь у поглинанні речовин бере вся система колоїдів цитоплазми. Інтенсивність руху залежить від температури, ступеня освітлення, постачання киснем тощо.
У дуже молодих клітинах цитоплазма заповнює майже всю їхню порожнину. У міру зростання клітини в цитоплазмі з'являються дрібні вакуолі, заповнені клітинним соком, що є водним розчином різних органічних речовин. Згодом, при подальшому зростанні клітини вакуолі збільшуються в розмірах і, зливаючись, часто утворюють одну велику центральну вакуоль, що відтісняє цитоплазму до оболонки клітини. У таких клітинах усі органоїди розташовуються у тонкому постінній шарі цитоплазми. Іноді ядро залишається у центрі клітини. У цьому випадку цитоплазма, що утворює навколо нього ядерну кишеньку, з'єднується з постінним шаром тонкими цитоплазматичними тяжами.
У шарі цитоплазми розташовані хлоропласти, що вистилають верхню стінку. Вони є майже округлі або злегка овальні тільця. Зрідка можна зустріти пластиди, перетягнуті посередині.
Поняття про тканини. Класифікація тканин. Відмінність постійних тканин від освітніх.
Більшість наземних рослинклітини тіла неоднакові за своїми функціями, будовою та походженням. Ця різнорідність виникла і закріпилася, у процесі пристосування рослин до відмінностей повітряного та ґрунтового середовищ. Системи клітин, структурно і функціонально подібні один до одного і зазвичай мають загальне походження, отримали назву ткачів. Тканини є майже у всіх вищих рослин. Немає їх тільки у частини мохоподібних (печіночники). Водорості та багрянки ( нижчі рослини), як правило, також не мають розвинених тканин.
Комплекси клітин, подібних до функцій, а здебільшогоі за будовою, що мають однакове походження та певну локалізацію в тілі рослини, називають тканинами.
Розподіл тканин у органах рослин та його структура тісно пов'язані з виконанням ними певних фізіологічних функцій.
Тканини, що складаються з одного типу клітин, отримали назву простих, а що складаються з різних типівклітин - складних, чи комплексних. Існують різні класифікаціїтканин, але вони досить умовні. Рослинні тканини ділять на кілька груп залежно від основної функції:
1) меристеми, або освітні тканини (тканини, що складаються з живих тонкостінних клітин, що інтенсивно діляться);
а) верхівкові (апікальні) меристими (розташовані на верхівках стебел і в закінченнях коренів) зумовлюють зростання цих органів у довжину;
б) бічні мерестими - камбій і феллоген (камбій забезпечує потовщення стебла і кореня. Феллоген утворює пробку)
2) покривні (захищають внутрішні тканинирослин від прямого впливу зовнішнього середовища, регулюють випаровування та газообмін)
а) епідерміс;
б) пробка;
3) провідні (забезпечують проведення води, ґрунтових розчинів та продуктів асиміляції, що виробляються листям. Провідні тканини за походженням можуть бути первинними та вторинними.);
а) кселіму або дерев'яна тканина(тканина, що проводить воду)
б) флоема або луб (тканина, яка проводить органічні речовини, утворені рослиною в процесі фотосинтезу);
4) механічні (обумовлюють міцність рослини);
а) коленхіму (складається з паренхіми або кілька подовжених клітин з нерівномірно потовщеними целюлозними стінками);
б) склеренхіма (клітини мають рівномірно потовщені дерев'яні стінки);
1) волокна;
2) склериди;
5) основні (що складаються з однорідних паренхімних клітин, які заповнюють простір між іншими тканинами);
6) секреторні, або видільні (що містять продукти відкидання).
Лише клітини меристематичних тканин здатні поділяти. Клітини інших тканин, зазвичай, до поділу нездатні, та його число збільшується з допомогою діяльності відповідних меристем. Такі тканини називають постійними. Постійні тканини виникають із меристем внаслідок клітинного диференціювання. Диференціювання полягає в тому, що в ході індивідуального розвиткуорганізму (онтогенезу) виникають якісні відмінності між спочатку однорідними клітинами, при цьому змінюються будова та функціональні властивостіклітин. Зазвичай диференціювання необоротне. На хід її впливають речовини, що виконують роль гормонів.
МЕРИСТЕМИ АБО ОСВІТИЛЬНІ ТКАНИНИ
Мерістеми (від грец. «меристос» - поділений), або освітні тканини, мають здатність до поділу та утворення нових клітин. За рахунок меристему формуються всі інші тканини і здійснюється тривалий (протягом усього життя) зростання рослини. У тварин меристеми відсутні, чим пояснюється обмежений період їхнього зростання. Клітини меристем відрізняються високою метаболічною активністю. Одні клітини меристем, що отримали назву ініціальних, затримуються на ембріональній стадії розвитку протягом усього життя рослини, інші поступово диференціюються і перетворюються на клітини різних постійних тканин. Ініціальна клітина меристеми може дати початок будь-якій клітині організму. Тіло наземних рослин – похідне щодо небагатьох ініціальних клітин.
Первинні меристеми мають меристематичну активність, тобто здатні до поділу спочатку. У ряді випадків здатність до активного поділу може знову виникнути і у клітин, що вже майже втратили цю властивість. Такі «знов» меристеми, що виникли, називають вторинними.
У тілі рослини меристеми займають різне становище, що дозволяє їх класифікувати. За становищем у рослині виділяють верхівкові, або апікальні (від латів. «апекс» - верхівка), бічні, або латеральні (від латів. «латус» - бік), та інтеркалярні меристеми.
Апікальні меристеми розташовуються на верхівках осьових органів рослини та забезпечують зростання тіла у довжину, а латеральні – переважно зростання у товщину. Кожна втеча та корінь, а також зародковий корінець, нирка зародка мають апікальну меристему. Апікальні меристеми первинні та утворюють конуси наростання кореня та втечі (рис. 1).
Латеральні меристеми розташовуються паралельно бічним поверхням осьових органів, утворюючи свого роду циліндри, що на поперечних зрізах мають вигляд кілець. Частина належить до первинним. Первинними меристемами є прокамбій та перицикл, вторинними – камбій та феллоген.
Інтеркалярні, або вставкові, меристеми частіше первинні і зберігаються у вигляді окремих ділянок у зонах активного зростання (наприклад, у основ міжвузля, в основах черешків листя).
Існують також ранові меристеми. Вони утворюються в місцях ушкодження тканин і органів і дають початок калюсу - особливої тканини, що складається з однорідних паренхімних клітин, що прикривають місце поразки. Чим інтенсивніше каллюсообразование, тим більше гарантія зрощення підщепи з прищепом і вкорінення живців. Освіта каллюса- необхідна умовакультури тканин рослини на штучних середовищах
Клітини апікальних меристем більш-менш ізодіаметричні за розмірами та багатогранні за формою. Міжклітинників з-поміж них немає, оболонки тонкі, містять мало целюлози. Порожнина клітини заповнена густою цитоплазмою з відносно великим ядром, яке займає центральне становище. Вакуолі численні, дрібні, але під світловим мікроскопом зазвичай непомітні. Ергастичні речовини, як правило, відсутні. Пластид та мітохондрій мало і вони дрібні.
Клітини бічних меристем різні за величиною та формою. Вони приблизно відповідають клітинам тих постійних тканин, які з них надалі виникають. Так, у камбії зустрічаються як паренхімні, і прозенхімні ініціалі. З паренхімних ініціалей утворюється паренхіма провідних тканин, а з прозенхімних - провідні елементи.
Мал. 1. Верхівкова меристема втечі елодеї. А - поздовжній зріз; 5 - конус наростання ( зовнішній вигляді поздовжній зріз); В – клітини первинної меристеми;
Г - паренхімна клітина листа, що сформувався:
1 ~ конус наростання, 2 - зачаток листка, 3 - горбок пазушної нирки
Зони кореня. Особливості морфологічної будови кожної зони у зв'язку з функціями, що виконуються. Зростання кореня в довжину та втовщину.
Поряд із втечею корінь - основний орган вищої рослини, що в типовому випадку виконує функцію мінерального та водного харчування. Інша важлива функціякореня, що тісно пов'язана з основною, - закріплення, рослини в грунті.
Різні частини кореня виконують неоднакові функції та характеризуються певними морфологічними особливостями. Ці частини дістали назву зон (рис. 2). Кінчик кореня зовні завжди прикритий кореневим чохликом, що захищає апікальну меристему. Клітини кореневого чохлика продукують слиз, що покриває поверхню кореня. Завдяки слизу знижується тертя про ґрунт, і його частинки легко прилипають до кореневих закінчень та кореневих волосків.
Кореневий чохлик складається з живих паренхімних клітин, що виникають у більшості однодольних з особливої меристеми, що отримала назву калітрогену, а у дводольних і голонасінних - верхівкової меристеми кінчика кореня. Водні рослини кореневого чохлика зазвичай не мають.
Під чохлом розташовується зона поділу, представлена меристематичною верхівкою кореня, його апексом. Внаслідок активності апікальної меристеми формуються всі інші зони та тканини кореня.
Клітини, що діляться, зосереджені в зоні поділу, що має розміри близько 1 мм. Ця частина кореня помітно відрізняється від інших зон своїм жовтуватим забарвленням. Після зоною розподілу розташовується зона розтягування (зростання). Вона також невелика по протяжності (кілька міліметрів), виділяється світлим забарвленням і прозора. Клітини зони зростання практично не діляться, але здатні розтягуватися в поздовжньому напрямку, проштовхуючи кореневе закінчення в глиб грунту. Вони характеризуються високим тургором, що сприяє активному розсуванню частинок ґрунту. У межах зони зростання відбувається диференціація первинних провідних тканин.
Закінчення зони зростання помітно з появою на епіблемі численних кореневих волосків. Кореневі волоски розташовуються в зоні-всмоктування, функція якої зрозуміла з її назви. На корені вона займає ділянку від кількох міліметрів до кількох сантиметрів. На відміну від зони зростання ділянки цієї зони не зміщуються щодо частинок грунту. Основну масу води та розчинів солей молоде коріння засвоює в зоні всмоктування за допомогою кореневих волосків.
Кореневі волоски з'являються у вигляді невеликих сосочків – виростів клітин епіблеми. Зростання волоска здійснюється біля його верхівки. Оболонка кореневого волоскарозтягується швидко. Після певного часу кореневий волосок відмирає. Тривалість його життя вбирається у 10-20 днів.
(меристематичних клітин) і знаходяться в частинах рослин, де відбувається зростання.
1. Цитологічні особливості меристем
Диференційовані клітини рослин взагалі можуть ділитися чи перетворюватися на клітини інших типів. Тому поділ клітин у меристемах необхідний для забезпечення нових клітин для росту інших тканин, утворення нових органів та забезпечення структури тіла рослини. За функцією меристематичних клітин аналогічні стовбуровим клітинам тварин, які не диференціюються або диференціюються незначно, і здатні до безперервного клітинного поділу. Меристематичні клітини маленькі, а цитоплазма і ядро повністю заповнюють клітину. Вакуолі надзвичайно маленькі, а цитоплазма не містить диференційованих пластид (хлоропластів або хромопластів), хоча вони присутні у рудиментарній формі (протопластиди). Меристематичні клітини щільно упаковані, майже без міжклітинного простору. Клітинна стінка – дуже тонка первинна клітинна стінка.
Фізіологічна діяльність меристем потребує балансу між двома антагоністичними процесами: утворенням нових тканин та підтримкою оновлення популяції меристематичних клітин.
Мерістема- утворююча рослинна тканина, з якої можуть утворюватися всі інші тканини, меристематичних клітин тривалий часзберігають здатність до поділу з утворенням нових неспеціалізованих клітин.
Клітини меристеми сплощені, дрібні, щільно розташовані один біля одного, без міжклітинників, в центрі клітин розташоване ядро, вакуолі дрібні, численні, первинна клітинна оболонка, є лише попередники пластид.
Цитологічні особливості меристем круп типово виражені в апікальних меристемах. Клітини ізодіаметричні багатокутники, не розділені міжклітинниками. Клітинна стінка тонка, з невеликим вмістом целюлози. Цитоплазма густа, ядро велике, розташоване в центрі. У цитоплазмі велика кількість рибосом і мітохнодрій (відбувається інтенсивний синтез білків та інших речовин). Численні вакуолі дуже дрібні.
Клітини латеральних меристем неоднакові за розміром та формою. Це з відмінністю клітин постійних тканин, їх утворюються. Так, наприклад, у камбії є паренхімні та прозенхімні клітини. З паренхімних ініціалів утворюється паренхіма провідних комплексів, з прозенхімних – власне провідні елементи.
Меристеми класифікують за становищем їх у рослині:
І за походженням:
- первинні
- вторинні.
Первинні меристеми знаходяться у тісному зв'язку з апікальною меристемою і безпосередньо з них утворюються, зокрема, протодерма, основна меристема, прокамбій. Протодерма – одношаровий пласт малодиференційованих клітин, що покриває апекс ззовні, надалі з неї формується епідерма. З основної меристеми формується кора, і з прокамбия, клітини якого витягнуті вздовж осі стебла, формується центральний циліндр.
Вторинні меристеми вникають зі спеціалізованих клітин зрілих органів рослин та забезпечують їх латеральне зростання (фелоген, перицикл, камбій). Тканини, що утворюються з первинної меристеми, називаються первинними, а з вторинною – вторинними. До вторинних меристем слід також віднести так звані травматичні меристеми, які формуються в місцях ушкодження тіла рослин та забезпечують регенерацію.
Серед перерахованих типів меристем першої в онтогенезі утворюється апікальна меристема, з якої відбувається диференціація всіх інших тканин. Розташована вона на верхівках (АПЕКС) стебла та кореня та їх бічних відгалужень. Особливістю життєдіяльності рослинного організмує, зокрема, збереження утворюючої діяльності в апікальних меристемах протягом усього онтогенезу, тому кажуть, що рослини здатні до необмеженого зростання.
Будова апікальної меристеми (точки зростання)
1 - Центральна частина
2 - Периферична частина
2. Апікальної меристеми
4. Інтеркалярна меристема
Інтеркалярна меристема (синонім – вставна меристема) – клітини розташовуються між ділянками диференційованих тканин, забезпечуючи вставне зростання.
Інтеркалярна меристема, наприклад, розташовується у вузлах злаків, біля основи черешків, тичинкових ниток. Це залишкові первинні меристеми. Вони виходять з верівкових меристем, але їх перетворення на постійні тканини затримано в порівнянні з іншими тканинами стебла. Це ніжні меристеми особливо добре помітні у злаків. У разі стану хлібів вони забезпечують підняття стебел за рахунок нерівномірного поділу клітин із нижньої та верхньої сторони соломинки.
5. Ранева меристема
Ті, що утворюються при пораненні тканин і органів. Живі клітини, що оточують уражені ділянки, дедиференціюються та починають ділитися, тобто перетворюються на вторинну меристему. Рана меристеми утворюють Калюс- щільну тканину білого або жовтого кольору, що складається з паренхімних клітин різного розміру, розміщених не впорядковано. Клітини калюсу мають великі ядра і відносно товсті клітинні стінки. Калюс може дати початок будь-якій тканині чи органу рослини. На перефірії формується пробка, можлива диференціація клітин калюсу інші тканини. У Калюсі можуть закладатися додаткове коріннята нирки. Калюс виникає при щепленні рослин, забезпечує зростання щепи та підщепи; в основі живців. Також Калюс використовують для одержання культури ізольованих тканин.
Органи рослин складаються із тканин. Рослинною тканиноюназивається сукупність клітин, що мають подібну будову, однорідне походження і виконують одну й ту саму функцію.Особливого розвитку досягли тканини найвищих наземних рослин. У рослинах розрізняють освітні тканини, покривні, механічні, провідні, основні та видільні.
Освітні тканини, або меристеми, служать у рослині для зростання, вони бувають первинного походження та вторинного. Первинна освітня тканина знаходиться в конусах наростання коренів та стебел. Зупинимося спершу на конусі наростання кореня(Рис.1, I).
Рис.1. Первинна освітня тканина в конусі наростання кореня:
I - конус наростання кореня: 1 – клітини освітньої тканини; 2 – клітини кореневого чохлика. II - послідовні фази складного поділу ядра та клітини(Каріокінез, або мітоз): 1 - інтерфаза; 2 - профаза; 3 – метафаза; 4-5 – анафаза; 6-7 – телофаза; 8 – цитокінез.
Кінчик кореня має конусоподібну форму, прикритий кореневим чохликом, під яким і знаходиться первинна освітня тканина. Клітини освітньої тканини чотиригранні або багатогранні, мають густу протоплазму, великі ядра та тонкі целюлозні оболонки. Вакуолі їх не розвинені чи слабо розвинені. Ці клітини мають здатність енергійного поділу. Поділу клітини передує складне розподіл ядра, у якому відбуваються своєрідні зміни. На готових добре приготовлених і пофарбованих препаратах видно ядра, що знаходяться в різних фазах складного поділу, що називається інакше каріокінезом («каріон» - ядро, «кінезес» - рух) (рис.1, II). Численними спостереженнями встановлено, що в ядрі спершу з хроматинової речовини виникають тонкі хроматинові нитки, які коротшають, товщають і перетворюються на окремі частинки - хромосоми ("хромо" - кpacкa, "сома" - тіло), часто вигнуті у вигляді підкови (профа . Кожна хромосома в профазі хіба що розщеплюється вздовж дві половини, а насправді відбувається подвоєння хромосом (редуплікація). Ядро і ядерна оболонка поступово зникають. Хромосоми розташовуються у середній частині ядра в одній площині у вигляді платівки. Половинки кожної хромосоми ("дочірні" хромосоми) щільно прилягають одна до іншої, і в цій фазі (метафазі) на поперечних зрізах кореня легко можна порахувати число хромосом і розглянути їх форму. На полюсах ядра (ще до зникнення ядерної оболонки) з'являється система тонких ахроматинових ниток, що не фарбуються, що розташовуються у вигляді двох ковпачків. Нитки розростаються і набувають вигляду веретена. Після цього починається розбіжність половинок хромосом (дочірніх хромосом) до двох протилежних полюсів клітини (анафаза). Розбіжність хромосом закінчується тим, що дочірні хромосоми, що зібралися біля полюсів клітини, переплітаються між собою, перетворюючись на два клубки (телофаза). Одночасно у середній частині клітини на ахроматинових нитках з'являються пектинові потовщення - утворюється фрагмопласт, тобто зачаток поперечної оболонки. Нарешті, ядра оточуються оболонкою, у яких утворюються ядерця (одне чи кілька), а протоплазма розділяється поперечної оболонкою (цитокинез ), у результаті виникають дві нові клітини.
Процес поділу ядра і клітини відбувається частіше вночі, швидкість його залежить від температури і вологості. Процес поділу в середньому триває 75-120 хвилин, але іноді триває кілька годин, причому першу фазу (профазу) витрачається найбільше часу. Клітинна оболонка при розподілі клітини виникає як продукт життєдіяльності протоплазми та ядра. Спочатку з'являється серединна платівка. Вона складається з міжклітинної речовини, що склеює первинні целюлозні оболонки, що належать двом клітинам.
Вперше розбіжність хромосом до полюсів клітини, як і інші подробиці поділу ядра і клітини, було встановлено 1875 р. професором Московського університету І. Д. Чистяковим.
Сутність складного поділу ядра і клітини, званого також мітозом, полягає, мабуть, у тому, що шляхом рівномірного розщеплення хромосом на дві половинки (або, точніше, подвоєння хромосом) дочірні клітини отримують від материнської більш-менш рівну кількість хроматинової речовини (дезоксирибону кислоти - ДНК), що передає спадковість від материнської клітини дочірнім клітинам. («Мітос» - нитка. Зазвичай прийнято, що мітоз = каріокінез + цитокінез). Форма та кількість хромосом є специфічними ознакамидля кожного виду рослин (пшениця м'яка 2n = 42 хромосоми, тверда пшениця 2n = 28, ячмінь 2n = 14 і т. д.). Проте іноді спостерігається мінливість числа хромосом, й у одному організмі можна спостерігати різнохромосомні клітини.
Крім мітозу (або каріокінезу), в рослинах зустрічається прямий поділ клітинного ядра (амітоз) - просте перетягування ядра, а іноді й усієї клітини на дві половинки.
Поряд з мітозом у рослинах поширений редукційний поділ ядра - мейоз, при якому кількість хромосом у нових клітинах зменшується вдвічі порівняно з материнською клітиною. Такий поділ відбувається у рослин при спороутворенні, за яким виникають і розвиваються чоловічі та жіночі статеві клітини (гамети). Останні мають у своїх ядрах половину кількість хромосом, і такі ядра називаються гаплоїдними на відміну диплоїдних ядер вегетативних клітин, з яких складаються вегетативні органи рослин. Підлоговий процес рослин полягає у злитті двох гамет (чоловічої та жіночої). Якби не відбувалося редукції хроматинової речовини, тобто зменшення його маси вдвічі, то в кожному новому статевому поколінні ця маса речовини, а отже, і кількість хромосом неминуче збільшувалася б. У статевих клітинах (гаметах) ядра бувають гаплоїдними, і лише після процесу запліднення клітинні ядразнову стають диплоїдними.
Редукційний поділ був відкритий вченим В. І. Бєляєвим, а вчений С. Г. Навашин відкрив подвійне запліднення у квіткових рослин і є засновником цілої школивчених, які займаються вивченням всіх деталей поділу ядра та клітини.
Первинна освітня тканина дає початок іншим тканинам, оскільки клітини її, змінюючись, диференціюються. Здатність клітин посилено ділитися в стеблах і коренях дводольних рослин зберігається на все життя в меристематичних клітинах, що мають назву камбію, завдяки якому товщає стебло і корінь. До вторинних освітніх тканин відносяться такі, що виникають з інших тканин. Прикладом може бути феллоген, що часто виникає з основної тканини.
1) Освітні тканини, або меристеми
Тіло рослини формується внаслідок діяльності освітніхтканин, званих меристемами. Основне властивість меристем – здатність клітин до поділу мітозом, з допомогою наявності ініціальних клітин.
У складі меристем розрізняють: 1) ініціалі, 2) похідні від ініціалів. Ініціали зберігають здатність до поділу протягом усього життя рослини і залишаються у складі меристем. Похідні від ініціалів діляться кілька разів і потім перетворюються на постійні тканини.
Особливості ініціалі: густа цитоплазма, велике ядро, багато мітохондрій, дрібні вакуолі або їх немає, тонка клітинна стінка.
За походженням розрізняють: 1) первинні меристеми, які беруть початок безпосередньо від меристема зародка. З них розвиваються первинні постійні тканини. 2) вторинні меристемиз первинної або інших тканин. З них розвиваються постійні вторинні тканини.
Залежно від розташування виділяють чотири типи меристем:
Верхівкові ( апікальні) меристеми- на верхівках стебел та кінцях коренів, забезпечує зростання у довжину.
Бічні (латеральні) меристеми –по колу осьових органів, забезпечуються зростання завтовшки. Первинні бічні меристеми – це прокамбій та перицикл. . Вторинні латеральні меристеми: камбій- виникає з прокамбію - і пробковий камбій (феллоген)- утворюється із клітин постійних тканин. З прокамбію та камбію утворюються провідні тканини, з пробкового камбію – перидерма.
Вставкові (інтеркалярні) меристемизнаходяться в основах міжвузлів пагонів та молодого листя. Вони первинні, оскільки є залишками верхівкових меристем. Вставні меристеми не мають у своєму складі ініціалів і згодом повністю перетворюються на постійні тканини.
Раневі (травматичні) меристемизазвичай утворюються при пошкодженні тканин та органів. Раневі меристеми утворюють каллус- особливу тканину, що складається з однорідних паренхімних клітин, що прикривають місце поранення. З калюсу може виникнути будь-яка тканина або орган рослини.
Практичне використання: квітки застосовують у парфумерії, медицині, в декорі.
2) Мікроскопічна будова однодольних рослин:
а) Виражено первинну кору(лілейні, ірисові). У цих випадках можна бачити в стеблі виразне диференціювання на шкірку, первинну кору та осьовий циліндрз багатошаровим перициклом у вигляді склеренхімного кільця. Первинна кора зазвичай складається з кількох шарів паренхімних хлорофілоносних клітин. Колленхіма, як правило, відсутня. У підземних пагонах первинна кора та центральний циліндр розмежовані особливо різко; первинна кора буває значно ширшою, ніж у наземних стеблах цих рослин, в ендодермі з'являються кільця Каспарі або навіть чашеподібні потовщення клітинних оболонока перицикл представлений переважно паренхімними клітинами.
Б) В інших представників класу однодольних, у родинах пальм, злаків та інших, первинна кора так чітко не виражена. У стеблі кукурудзи під епідермісом іноді знаходяться ділянки хлорофілоносної паренхіми, склеренхіма оточує ці ділянки і між ними прилягає безпосередньо до епідермісу. Периферичні судинні пучки мають потужні склеренхімні обкладки, і поблизу епідермісу утворюється іноді суцільний шар склеренхіми, який занурені судинні пучки.
3) Загальна характеристикавищих рослин
За своїм виглядом, за будовою та біологічним особливостямвищі рослини дуже різноманітні. До них відносяться не тільки квіткові рослини, але і голонасінні, а також папороті, хвощі, плауни та мохи.
Вищі рослини характеризуються рядом ознак. високої організації. Їхнє тіло розчленоване на стебла і листя (на відміну від нижчих).У зв'язку з цим їх називають листостебловими рослинами.
Поява складного розчленування, утворення органів (корінь, стебло, лист) було викликано переходом рослин у нову наземне середовищепроживання. Повітряне середовищехарактеризується набагато більшим вмістом кисню, ніж водна. Ґрунтова – іншими умовами мінерального харчування та особливо водопостачання. Для асиміляції в нових умовах була потрібна перебудова фотосинтезуючого апарату. Так у рослини з'явилося стебло.
Далі для кращого всмоктування ризоїди замінилися корінням. Коріння почало забезпечувати мінеральне харчування, а листя – повітряне харчування вищих рослин.
Для нормального функціонування органів було необхідне швидке пересування води до листя та органічних речовин – з листа до інших частин рослини. Це призвело до розвитку провідної системи у вигляді спеціальних клітин – трахеїд, судин та ситоподібних трубок. Для захисту утворилася покривна тканина, механічна.
Статевий процес став менше завісити від води.
Вищі: рініофіти, псилофіти, мохи, плауноподібні, хвощеподібні, папороті, голонасінні, покритонасінні
ТКАНИНИ. КЛАСИФІКАЦІЯ ТКАНИН.
В основі організації вищих рослин лежить принцип спеціалізації клітин, який полягає в тому, що кожна клітина організму виконує не всі властиві їй функції, а лише деякі, зате повніше і досконаліше.
Тканини- стійкі, закономірно повторювані комплекси клітин, подібні за походженням, будовою та пристосовані до виконання однієї або кількох функцій.
Існують різні класифікації тканин, але вони досить умовні.
Залежно від основної функції розрізняють кілька груп рослинних тканин.
1. Освітні тканиниабо меристеми,- мають здатність до поділу та формування всіх інших тканин.
2. Покривні тканини:
Первинні;
Вторинні;
Третинні.
3. Основні тканини- складають більшу частинутіла рослини. Розрізняють такі основні тканини:
асиміляційні (хлорофілоносні);
Запасні;
Повітроносні (аеренхіма);
Водоносні.
4. Механічні тканини(опорні, скелетні):
Колленхіма;
Склеренхіма.
5. Провідні тканини:
Ксилема (деревина) - тканина висхідного струму;
Флоема (луб) - тканина спадного струму.
6. Видільні тканини:
Зовнішні:
Залізисті волоски;
Гідатоди – водяні продихи;
Нектарники;
Внутрішні:
Видільні клітини з ефірними оліями, смолами, дубильними речовинами;
Багатоклітинні вмістилища виділень, млечники.
За здатністю клітин до поділу розрізняють два типи тканин: освітні,або меристеми,і постійні- покривні, видільні, основні, механічні, провідні.
Тканину називають простий,якщо всі її клітини однакові за формою та функціями (паренхіма, склеренхіма, коленхіма). Складнітканини складаються з клітин, неоднакових за формою, будовою та функціями, але пов'язаних загальним походженням(наприклад, ксилема, флоема).
Існує також класифікація тканин, заснована на їхньому походженні (онтогенетична). Відповідно до цієї класифікації розрізняють первинні та вторинні тканини. З первинної меристеми, що знаходиться на верхівці втечі та кінчику кореня, а також із зародка насіння формуються первинні постійнітканини (епідерма, коленхіма, склеренхіма, асиміляційна тканина, епіблема). Клітини постійних тканин нездатні до подальшого поділу. З клітин спеціалізованої меристеми – прокамбія - формуються первинні провіднітканини (первинна ксилема, первинна флоема).
З вторинної меристеми – камбія - формуються вториннітканини: вторинна ксилема, вторинна флоема; з феллогену утворюються пробка, феллодерма, чечевички, що виникають при потовщенні стебла та кореня. Вторинні тканини, як правило, зустрічаються у голонасінних та дводольних покритонасінних рослин. Потужний розвиток вторинних тканин – деревини та лубу характерний для деревних рослин.
ОСВІТНІ ТКАНИНИ
Освітні тканинизавдяки постійному мітотичному поділу їх клітин забезпечують утворення всіх тканин рослини, тобто. фактично формують його тіло. Будь-яка клітина у своєму розвитку проходить три стадії: ембріональну, зростання та стадію диференціації (тобто придбання клітиною певної функції). У міру диференціації зародка первинна меристема зберігається тільки на верхівці майбутньої втечі (у конусі наростання) і на кінчику кореня – апікальні (верхівкові) меристеми.Зародок будь-якої рослини складається з клітин меристеми.
Цитологічна характеристика меристему.Типові ознаки найвиразніше виражені у верхівкових меристемах. Ці меристеми складені ізодіаметричними багатогранними клітинами, не розділеними міжклітинниками. Їхні оболонки тонкі, містять мало целюлози і здатні розтягуватися.
Порожнина кожної клітини заповнена густою цитоплазмою з відносно великим ядром, що займає центральне положення, і мітозом, що інтенсивно ділиться. У гіалоплазмі багато дифузно розкиданих рибосом, пропластид, мітохондрій та диктіосом. Вакуолей небагато, і вони дрібні. Провідні тканини утворюються з меристеми, що має прозенхімну форму і великі вакуолі - прокамбію і камбію. Клітини прокамбію в поперечному перерізібагатокутні, клітини камбію – прямокутні.
Клітини, що зберігають свої меристематичні властивості, продовжують ділитися, утворюючи нові і нові клітини, звані ініціалями.Частина дочірніх клітин диференціюється, перетворюючись на клітини різних тканин, їх називають похідними ініціалів.Клітини ініціалів можуть ділитися невизначено багато разів, а похідні ініціалів діляться один або кілька разів і перетворюються на постійні тканини.
За походженням розрізняють первинні та вторинні меристеми.
Первинні меристеми
Первинні меристеми походять безпосередньо з меристеми зародка і мають здатність до поділу. По положенню в рослині первинні меристеми можуть бути верхівковими (апікальними), вставковими (інтеркалярними) та бічними (латеральними).
Верхівкові (апікальні) меристеми- такі меристеми, які розташовуються у дорослих рослин на верхівках стебел та кінчиках коріння та забезпечують зростання тіла у довжину. У стебел у конусі наростання виділяють два меристематичні шари: туніку, з якої утворюються покривна тканина і периферична частинапервинної кори і корпус, з якого утворюються внутрішня частина первинної кори і центральний осьовий циліндр (рис. 2.3).
Мал. 2.3.Апікальні меристеми стебла: а- поздовжній зріз: 1 – конус наростання; 2 – зачаток аркуша; 3 – горбок пазушної нирки;
У кінчику кореня розрізняють три шари:
1) дерматоген, з якого утворюється первинна покривно-всмоктуюча тканина – ризодерма;
2) периблему, з якої розвиваються тканини первинної кори;
3) плером, що утворює тканини центрального осьового циліндра.
за походженням можуть бути первинними та вторинними, на поперечному зрізі осьових органів мають вигляд кілець. Прикладом первинної бічної меристеми є прокамбій і перицикл. З прокамбіяформуються камбій та первинні елементи судинно-волокнистих пучків (первинна флоема та первинна ксилема), при цьому клітини прокамбію безпосередньо диференціюються в клітини первинних провідних тканин.
Бічні меристеми розташовуються паралельно поверхні органу та забезпечують зростання осьових органів у товщину.
Вставкові (інтеркалярні) меристемичастіше первинні і зберігаються у вигляді окремих ділянок у зонах активного зростання різних частинахрослини (наприклад, в основі черешків листя, біля основ міжвузля). В основі міжвузлів у злаків діяльність цієї меристеми веде до подовження міжвузлів, що забезпечує зростання стебла в довжину.
Вторинні меристеми
До вторинних відносять бічні та ранові меристеми.
Бічні (латеральні) меристемипредставлені камбіємі феллогеном.Вони формуються з промеристем (прокамбію) або постійних тканин шляхом їх дедиференціювання. Клітини камбію діляться перегородками, паралельними поверхні органу (периклінально). З клітин, відкладених камбієм назовні, розвиваються елементи вторинної флоеми, та якщо з клітин, відкладених всередину, – вторинної ксилемы. Камбій, що виник із постійних тканин шляхом дедиференціювання, називають додатковим.За будовою та функцією він не відрізняється від камбію, що виник з промеристем. Феллоген формується із постійних тканин, розташованих у субепідермальних шарах (під епідермою). Ділячись периклінально, феллоген відокремлює назовні майбутні клітини пробки (фелеми), а всередину – клітини фелодерми. Таким чином, фелоген формує вторинну покривну тканину- Перідерму. Бічні меристеми розташовуються паралельно поверхні органу та забезпечують зростання осьових органів у товщину.
Раневі меристемиутворюються при пошкодженні тканин та органів. Навколо пошкодження живі клітини дедиференціюються, починають ділитися і цим перетворюються на вторинну меристему. Їхнє завдання – утворити щільну захисну тканину, що складається з паренхімних клітин, – каллюс.Ця тканина білуватого або жовтуватого кольору, її клітини мають великі ядра і досить товсті клітинні стінки. Каллюс виникає при щепленнях, забезпечуючи зрощення щепи з підщепою, і в основі черешків. У ньому можуть закладатися придаткові коріння та нирки, тому його використовують для одержання культури ізольованих тканин.
ПОКРОВНІ ТКАНИНИ
Первинна покривна тканина
До первинної покривноїтканини відносять епідерму власне епідермальних, околоустьичных, замикаючих клітин продихом і трихом.
Пектинові речовини та целюлоза, що входять у клітинну стінку, можуть піддаватися ослизненню з утворенням слизіві камедей.Вони являють собою полімерні вуглеводи, споріднені з пектиновими речовинами, і відрізняються здатністю до сильного набухання при зіткненні з водою. Камеді в набряклому стані клейкі і можуть витягуватися в нитки, тоді як слизи сильно розпливаються і нитки не витягуються. Пектинові слизу зустрічаються у представників сімейств лілейних, хрестоцвітих, мальвових, липових, розоцвітих, на відміну від целюлозних слизів, що зустрічаються набагато рідше (наприклад, у орхідних).
 Устячка
являють собою високоспеціалізовані утворення епідерми, що складаються з двох замикаючих клітин бобоподібної форми та устьичної щілини (своєрідного міжклітинника між ними). Є головним чином листі, але трапляються й у стеблі (рис. 2.6).
Устячка
являють собою високоспеціалізовані утворення епідерми, що складаються з двох замикаючих клітин бобоподібної форми та устьичної щілини (своєрідного міжклітинника між ними). Є головним чином листі, але трапляються й у стеблі (рис. 2.6).
Мал. 2.6.Будова продихи: а, б- Шкірка листа чебрецю (вид зверху і на поперечному зрізі); в- Шкірка зі стебла цереуса (родина кактусових); 1 – власне епідермальні клітини; 2 – замикаючі клітини продихи; 3 – устьична щілина; 4 – повітроносна порожнина; 5 – клітини хлорофілоносної паренхіми; А – кутикула; Б – кутикулярний шар – оболонка із суберином та воском; В – целюлозний шар стінки; Г - ядро з ядерцем; Д – хлоропласти
Стіни замикаючих клітин потовщені нерівномірно: стінки, спрямовані до щілини (черевні), значно потовщені порівняно зі стінками, спрямованими від щілини (спинними). Щілина може розширюватися і звужуватися, регулюючи транспірацію та газообмін. Під щілиною розташована велика дихальна порожнина (міжклітинник), оточена клітинами мезофілу листка.
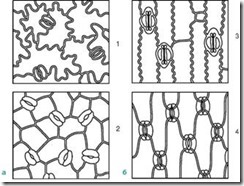
 Замикаючі клітини оточені навколоустьовими клітинами, які спільно утворюють устьичний комплекс(Рис. 2.7). Розрізняють такі основні типи устьичних комплексів:
Замикаючі клітини оточені навколоустьовими клітинами, які спільно утворюють устьичний комплекс(Рис. 2.7). Розрізняють такі основні типи устьичних комплексів:
Мал. 2.7.Основні типи устьичного апарату: 1 – аномоцитний (у всіх вищих рослин, крім хвощів); 2 – діацитний (у папороті та квіткових); 3 – парацитний (у папоротей, хвощів, квіткових та гнетових); 4 – анізоцитний (тільки у квіткових); 5 – тетрацитний (переважно у однодольних); 6 – анциклоцитний (у папоротей, голонасінних та квіткових)
1) аномоцитний(безладний) – замикаючі клітини немає яскраво виражених околоустьичных клітин; характерний всім вищих рослин, крім хвойні;
2) анізоцитний(Нерівноклітинний) – замикаючі клітини продихи оточені трьома навколоустьичними клітинами, одна з яких набагато більша (або менше) інших;
3) парацитний(паралельно-клітинний) – одна навколоустьева клітина (або більше) розташована паралельно замикаючим;
4) діацитний(перехресно-клітинний) – дві навколоустичні клітини розташовані перпендикулярно замикаючим;
5) тетрацитний(Від грец. tetra- чотири) - головним чином у однодольних;
 Продихання розташовані на нижній стороні листа, але у водних рослинз плаваючим листям вони знаходяться тільки на верхній стороні листа. За формою клітин епідерми листа і розташування продихів можна відрізнити однодольне рослина від дводольного (рис. 2.8). Власне епідермальні клітини листка дводольних рослин в контурах хвилясті (рис. 2.9), а у однодольних вони витягнуті, ромбічної форми.
Продихання розташовані на нижній стороні листа, але у водних рослинз плаваючим листям вони знаходяться тільки на верхній стороні листа. За формою клітин епідерми листа і розташування продихів можна відрізнити однодольне рослина від дводольного (рис. 2.8). Власне епідермальні клітини листка дводольних рослин в контурах хвилясті (рис. 2.9), а у однодольних вони витягнуті, ромбічної форми.
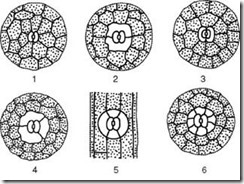
Мал. 2.8.Розташування продихів на епідермі (вид з поверхні): а- дводольні рослини: 1 - Букви; 2 – кавун; б- однодольні рослини: 3 – кукурудза; 4 – ірис
Типи продихів за рівнем розташування щодо поверхні епідермісу можна підрозділити в такий спосіб.
1.7.1. Продихання, розташовані в одній площині з епідермісом. Найбільш поширений тип і зазвичай у описі мікроскопії лікарської рослинної сировини не вказується, тобто. цей пункт опускається. Діагностичними ознакамибудуть або виступаючі, або занурені продихи.
1.7.2. Виступаючі продихи - продихи, розташовані над епідермісом. Зазвичай при обертанні мікрогвинта мікроскопа (при опусканні об'єктиву) спочатку виявляються такі продихи, а вже потім з'являються клітини епідермісу, тому на фотографії відобразити їх з поверхні листа практично неможливо, так само як і зобразити на малюнку. В одній площині з епідермісом такі продихи можна побачити на поперечних зрізах, але для цього зріз повинен пройти через продихи, що важко отримати при їх рідкісному розташуванні на листі. Такі продихи характерні, наприклад, для листя мучниці.
1.7.3. Занурені продихи - продихи, занурені в епідерміс. При спостереженні під мікроскопом при обертанні мікрогвинта (при опусканні об'єктиву) спочатку чітко виявляються клітини епідермісу, потім стає можливим чіткіше побачити контури продихів. На фотографіях та малюнках препаратів із поверхні їх також важко відобразити. Зустрічаються в листі конвалії, листі вахти, листках евкаліпта. Іноді поглиблення, в яких розташовуються продихи, вистелені або прикриті волосками і називаються устьичними криптами.
1.8. Типи устьичних клітин
У літературі описується 19 типів, нами обрані лише ті, що використовуються в аналізі лікарської рослинної сировини.
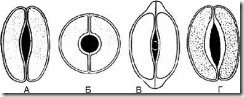
Мал. 63.Типи устьичних клітин. А - сочевицеподібні; Б - сфероподібні; В – ковпачкоподібні; Г – човноподібні
1.8.1. Сочевицеподібні - 2 однакові клітини напівмісячної форми розташовані симетрично. На передній поверхні потовщення оболонки майже рівномірне. Щілина веретеноподібна (рис. 63, А). Тип устьичних клітин характерний більшості рослин.
1.8.2. Сфероподібні - дві однакові, сильно кругоподібно зігнуті клітини розташовані симетрично. На передній поверхні потовщення оболонки майже рівномірне. Щілина кругла (рис. 63, Б).
1.8.3. Ковпачкоподібні - дві однакові клітини напівмісячної форми у полярних частинах мають потовщення у вигляді ковпачка. Щілина веретеноподібна (рис. 63, В). Зустрічаються у наперстянок.
1.8.4. Човноподібні - Внутрішні стінки устьичних клітин потовщені. Щілина веретеноподібна (рис. 63, Г). Спостерігається у траві золототисячника, у листі вахти.
Механізм роботи продихів обумовлений осмотичними властивостями клітин. При освітленні поверхні листа сонцем у хлоропластах замикаючих клітин відбувається активний процесфотосинтезу. Насичення клітин продуктами фотосинтезу, цукрами тягне за собою активне надходження до клітин іонів калію, внаслідок чого концентрація клітинного сокуу замикаючих клітинах зростає. Виникає різниця концентрації клітинного соку навколоустьових та замикаючих клітин. З огляду на осмотичних властивостей клітин вода з околоустьичных клітин надходить у замикаючі, що веде до збільшення обсягу останніх і різкого зростання тургора. Потовщення «черевних» стінок клітин, що замикають, звернених до устьичної щілини, забезпечує нерівномірне розтягнення клітинної стінки; замикаючі клітини набувають вираженої бобоподібної форми, і устьична щілина відкривається. При зниженні інтенсивності фотосинтезу (наприклад, увечері) зменшується утворення цукрів у клітинах, що замикають. Приплив іонів калію припиняється. Концентрація клітинного соку в клітинах, що замикають, знижується в порівнянні з околоустьічними. Вода шляхом осмосу йде з клітин, що замикають, знижуючи їх тургор, в результаті вночі устьична щілина закривається.
Клітини епідерми щільно зімкнуті між собою, завдяки цьому епідерма виконує цілу низку функцій:
Перешкоджає проникненню хвороботворних організмів усередину рослини;
Захищає внутрішні тканини від механічних ушкоджень;
Регулює газообмін та транспірацію;
Через неї виділяються вода, солі;
Може функціонувати як всмоктувальна тканина;
бере участь у синтезі різних речовин, сприйнятті подразнень та у русі листя.
Трихоми - різні за формою, будовою та функціями вирости клітин епідерми: волоски, лусочки, щетинки тощо. Їх поділяють на криючі та залізисті. Залізисті трихоми,на відміну від криючих, мають клітини, що виділяють секрет. Кріючі волоски,утворюючи на рослині шерстистий, повстяний або інший покрив, відбивають частину сонячних променіві цим зменшують транспірацію. Іноді волоски знаходяться тільки там, де розташовані продихи, наприклад, на нижній стороні листа мати-й-мачухи. У деяких рослин живі волоски збільшують загальну поверхню, що випаровує, що сприяє прискоренню транспірації.
Розміри трихом значно варіюють. Найбільш довгі трихоми (до 5-6 см) покривають насіння бавовнику. Кроючі трихоми мають форму простих одно або багатоклітинних, розгалужених або зірчастих волосків. Трихоми, що кроять, можуть тривалий час залишатися живими або швидко відмирати, заповнюючись повітрям.
Від тріхом, що виникають лише за участю епідермальних клітин, відрізняються емергенці, в освіті яких беруть участь і глибше розташовані тканини субепідермальних шарів.
Анатомо-діагностичні ознаки, що мають найбільше значенняі високу варіабельність щодо лікарської сировини. Волоски можуть бути прості та головчасті, які у свою чергу можуть бути одноклітинними та багатоклітинними. Багатоклітинні волоски можуть бути однорядними, дворядними та гіллястими.
1.9.1. Прості волоски.
А. Прості одноклітинні волоски
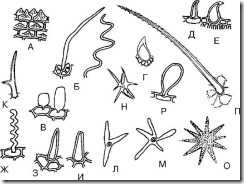
Мал. 68.Прості одноклітинні волоски. А - сосочкоподібний; Б - ниткоподібний; В - міхурові; Г – шипоподібний; Д - гачкоподібний; Е – ретортоподібний; Ж – тупонітеподібний гофрований; З – гостроконусоподібний; І – тупоконусоподібний; К – двокінцевий; Л – трикінцевий; М, Н, Про – багатокінцевий; П - горбистий; Р – булавоподібний
1. Сосочкоподібні (рис. 68, А; рис. 69-74) - невисокі, але широкі вирости епідермальних клітин. Найчастіше утворюються на пелюстках. Можуть бути туповерхівкові та гостроверхівкові. Зустрічаються на листі чебрецю, квітках конвалії, квітках фіалки триколірної, по краю листя золототисячника, в траві тирличу бородатою та ін.
2. Конусоподібні (рис. 68, З, І; рис. 75-79) - прямі і нахилені під кутом до поверхні волоски, що закінчуються тупим (тупоконусоподібні) або гострим (гостроконусоподібні) кінцем. Найбільш поширені волоски. Зазвичай бувають прямі. Нахилені до поверхні волоски спостерігаються на листі чебрецю. Остроконусовидні волоски зустрічаються на листі сенни, листі брусниці, в траві фіалки триколірної (на листі), в траві чебрецю та ін. Тупоконусоподібні волоски є на епідермісі плодів анісу, на пелюстках квіток фіалки триколірної.
3. Ниткоподібні (Рис. 68, Б, Ж; мал. 80, 81) - тонкі і довгі волоски. Можуть бути прямі та гофровані. Прямі ниткоподібні волоски виявляються на епідермісі плодів глоду та малини. Гофровані тупонітеподібні волоски спостерігаються на епідермісі пелюсток фіалки (рис. 82).
4. Ретортоподібні (рис. 68, Е; рис. 83) - волоски з розширеною основою і вузьким вигнутим або прямим закінченням. Зустрічаються у суплодіях хмелю, траві зубчатки.
5. Бульбашкові (рис. 68, В; рис. 84) - волоски у вигляді невеликих пухирів. Їх можна виявити, наприклад, у квітках безсмертника.
6. Гачкоподібні (Рис. 68, Д; мал. 85-87) - волоски, загострені на верхівці і вигнуті у вигляді гачка. Ці волоски можна зустріти біля основи листя мучниці, на поверхні листя брусниці, сенни, плодів анісу.
7. Бахромчасті (Рис. 88, 89) - довгі вирости епідермальних клітин, розташовані по краю листка, пелюстка або чашолистка. Такі волоски є, наприклад, по краю пелюсток конвалії та фіалки.
8. Шипоподібні (рис. 68, Г; рис. 90) – волоски майже округлої форми із загостреним кінцем. Спостерігаються у траві фіалки триколірної.
9. Булавоподібні (рис. 68, Р; рис. 91) - волоски з розширеною кінцевою частиною, що нагадують булаву. Є на пелюстках чебрецю, пелюсток фіалки триколірної.
10. Двоконечні (рис. 68, К) - волоски, розгалужені на два кінці. Спостерігаються в траві грициків.
11. Трикінцеві (багатокінцеві) (Рис. 68, Л-О) - волоски, розгалужені на три (і більше) кінця. Спостерігаються в траві грициків.
12. Бугристі (рис. 68, П) – конусоподібні волоски, що мають виступи (пагорби) на своїй поверхні.
Б. Прості багатоклітинні однорядні волоски
Контур багатоклітинного волоска може збігатися з таким одноклітинного волоска, але містити дві і більше клітин і відповідно називатися як аналогічний одноклітинний волосок із зазначенням кількості клітин, наприклад 2-клітинний ретортовидний волосок, 11-клітинний ниткоподібний волосок, 10-15-клітинний конусовидний волосок.

Мал. 92.Прості багатоклітинні волоски. А – конусоподібний; Б - міхурові; В – суглобовий; Г – бичевидний; Д - перистий; Е - Т-подібний; Ж – дворядні; З - лускатий; І – пучковий; Л – паралельний; М – вильчастий; К – щетинисті
1. Конусоподібні (Мал. 92, А; 93-96). Найбільш широко зустрічаються волоски. Бувають прямі та нахилені до поверхні, останні зустрічаються рідко (наприклад, у траві чебрецю). Конусоподібні волоски можуть бути гостро-і тупоконусоподібні. Найбільш поширені гостроконусоподібні волоски (у траві материнки, у листі м'яти, шавлії та ін.). Тупоконусоподібні зустрічаються у квітках календули.
2. Ретортоподібні - волоски з розширеною основою та вузьким вигнутим або прямим закінченням.
3. Бульбашкові (рис. 92, Б; рис. 97) - волоски у вигляді невеликих пухирів. Є на поверхні зав'язі безсмертника.
4. Ниткоподібні - тонкі та довгі багатоклітинні волоски.
5. Гачкоподібні - волоски, загострені на верхівці та вигнуті у вигляді гачка.
6. Гусеницеподібні (Мал. 98, 99) – волоски практично однакової товщини на всьому своєму протязі, що складаються з майже однакових коротких клітин, з тупим кінцем і нагадують гусеницю. Спостерігаються у траві фіалки триколірної, у траві череди.
7. Бахромчасті (рис. 100) - довгі багатоклітинні вирости епідермальних клітин, розташовані по краю листка, пелюстки або чашолистки. Зустрічаються у траві череди.
8. Бічкоподібні (рис. 92, Г; рис. 101-104) – волоски, що мають багатоклітинну основу, що складається з ланцюга коротких клітин, і довгу ниткоподібну звивисту кінцеву клітину. У літературі такі волоски описують як батога, шнуроподібні, ниткоподібні, повстяні. Тип волосків характерний для сімейства Asteraceae. Зустрічається в траві деревію, квітках безсмертника, листі мати-й-мачухи, квітках пижма та ін.
9. Суглобові (рис. 92, В; рис. 105, 106) - волоски, що мають розширені основи клітин у місцях їх зчленування (що нагадують суглоб). Подібні волоски виявляються в траві собачої кропиви, на пелюстках материнки, рідко в траві чебрецю.
10. Бугристі - волоски, що мають виступи (горби) на своїй поверхні. Двоклітинні горбисті волоки спостерігаються, наприклад, у листі термопсису.
В. Прості багатоклітинні гіллясті волоски
1. Паралельні (рис. 92, Л; рис. 107, 108) - волоски, що складаються з двох довгих прямих клітин (волосків), зрощених основами. Зустрічаються в квітках липи, рідко в плодах шипшини та глоду.
2. Вільчасті (рис. 92, М; рис. 109, 110) - волоски, що складаються з двох довгих звивистих клітин (волосків), зрощених основами. Спостерігаються в квітках липи, дуже рідко в квітках та плодах глоду.
3. Зірчасті (рис. 111) - волоски, що складаються з 3 і більш довгих звивистих клітин (волосків), зрощених основами. Виявляються у квітках липи.
4. Лускаті (рис. 92, З) - волоски, що складаються з багатоклітинної пластинки (у вигляді розетки) і короткої ніжки (ніжка може бути відсутнім). Такі волоски є в обліпиху. (Ця назва волосків наведена відповідно до сучасної ботанічної термінології; за іншими даними ці волоски називають зірчастими або щиткоподібними лускатими).
5. Т-подібні (рис. 92, Е) – двокінцева волосинка, що має одно-, багатоклітинну ніжку. У літературі описуються також як рогоподібні, коромислоподібні.
6. Перисті (Рис. 92, Д) - багатоклітинні волоски, що нагадують дерево, що гілкується. Виявляються, наприклад, у коров'яку.
Г. Прості багатоклітинні багаторядні волоски
1. Дворядні (рис. 92, Ж; рис. 112, 113) - волоски, клітини яких розташовані в два ряди. Їх можна спостерігати у квітках календули, у траві сушениці.
2. Пучкові (рис. 92, І) – волоски, що з двох і більше клітин, щільно зрощених друг з одним, утворюючи пучок.
3. Щетинисті (рис. 92 К, рис. 114) - багатоклітинні волоски, що складаються з волосків, що зрослися один з одним. різної довжинита мають вільні загострені кінці. Такі волоски зазвичай є волосками летучки (родина Asteraceae).У літературі їх ще називають зазубреними складними, шилоподібними.
1.9.2. Головчасті волоски. Головчасті волоски називають також залізистими. Вони можуть бути одноклітинними та багатоклітинними.

Мал. 115.Головчасті волоски. А – з одноклітинною ніжкою та одноклітинною головкою; Б – з дворядною ніжкою та одноклітинною головкою; В – з дворядною ніжкою та дворядною головкою; Г – з одноклітинною ніжкою та багатоклітинною головкою; Д – з багатоклітинною ніжкою та одноклітинною головкою; Ж – з багатоклітинною ніжкою та багатоклітинною головкою; З – одноклітинний (пекючий); І – щиткоподібний
А. Одноклітинні головчасті волоски
Представники цього типу волосків можуть бути конусоподібними або ретортоподібними, але мають на кінці головку. Такі волоски є, наприклад, на листі кропиви. Б. Багатоклітинні головчасті (залізисті) волоски
1. Волоски, що мають багатоклітинну головку та одноклітинну ніжку (Рис. 115, Г; мал. 116-120). Зустрічаються в траві собачої кропиви, листі мучниці.
2. Волоски, що мають одноклітинну головку та одноклітинну ніжку (рис. 115, А; рис. 121-123). Зустрічаються в траві собачої кропиви, листі шавлії, квітках бузини, листі м'яти.
3. Волоски, що мають одноклітинну головку та багатоклітинну ніжку (Рис. 115, Д; мал. 124). Зустрічаються в траві собачої кропиви, листі шавлії, квітках календули і квітконіжках ромашки.
4. Волоски, що мають багатоклітинну головку та багатоклітинну ніжку (Рис. 115, Ж; мал. 125). Зустрічаються в траві собачої кропиви, листі мучниці, квітках липи, квітках календули, траві фіалки.
5. Волоски, що мають одноклітинну головку та дворядну ніжку (Рис. 115, Б; мал. 126). Зустрічаються в квітках календули та безсмертника.
6. Волоски, що мають дворядну головку та дворядну ніжку (рис. 115, У; рис. 127- 130). Зустрічаються в квітках календули і безсмертника, в траві сухоцвіту болотної.
7. Булавоподібні багатоклітинні залізисті волоски (рис. 131-133; див. рис. 155, Д) - волоски, що мають контур з розширеною кінцевою частиною, що нагадують булаву. Їх можна спостерігати, наприклад, у листі брусниці, траві фіалки.
8. Щиткоподібні багатоклітинні залізисті волоски (рис. 115, І; рис. 134) являють собою щиток із багатокутних тонкостінних клітин, що сидить на одно-, двоклітинній короткій ніжці. Кутикула щитка відсувається від клітин, що виділяються під неї. ефірною олією. Вони виявлені у супліддях хмелю.
1.10. Характер потовщеності клітинних стінок і кутикули волосків, що покриває.
А. Характер потовщеності клітинних стінок
1.10.1. Тонкостінні (Рис. 135-137). Більшість волосків тонкостінні. Стінки клітин багатоклітинних довгих тонкостінних волосків іноді спадаються, порушуючи рівний контур волоска. Такі волоски, наприклад, зустрічаються на нижньому епідермісі листа какалії.
1.10.2. Товстостінні (Рис. 138, 139). Зустрічаються у плодах шипшини, траві череди, траві лепедеці даурської та ін.
1.10.3. Нерівномірно потовщені. Можна спостерігати в траві собачої кропиви (рис. 140). До цієї категорії належать волоски шипшини з косопоперечными порами (рис. 141, 142).
Б. Характер покриває кутикули волосків
1.10.4. Гладка поверхня (Рис. 143). Волоски з такою поверхнею спостерігаються, наприклад, у листі подорожника, траві чистотілу та ін.
1.10.5. Бородавчаста поверхня (див. рис. 1, Ж; рис. 144, 145) – епідерміс утворює виступи у вигляді горбків (бородавок). Є волоски, наприклад, у траві фіалки, у траві чебрецю, у траві собачої кропиви, у листі м'яти, у листі сенни та ін. При цьому поверхня волоска може бути слабобородавчастою, коли утворюються невеликі виступи кутикули, і грубобородавчастої, коли виступи утворюються значні. У першому випадку як приклад можна навести волоски у траві фіалки, у другому – волоски листя сенни та трави чебрецю.
1.10.6. Великогорбиста поверхня (див. рис. 68, П) – кутикула утворює дуже великі виступи, наприклад волоски трави термопсису ланцетного.
10.7. Зморшкувата поверхня - Складки або хвилі кутикули волоска від основи до його вершини (рис. 146). Спостерігається частіше у сосочків, наприклад, у траві золототисячника, квітках бузини. Однак зморшкувата поверхня може бути і у звичайних волосків, наприклад, у траві низки. В основі зморшкувату поверхню мають волоски листя м'яти (рис. 147).
1.10.8. Штрихувата поверхня - епідерміс утворює короткі виступи у вигляді штрихів (горбків, бородавок). Рідко зустрічається і є проміжним варіантом між бородавчастою та зморшкуватою поверхнею. Спостерігається, наприклад, на волосинках квіток бузини (рис. 148).
1.11. Особливості місць приєднання волосків
1.11.1. Звичайні місцяприкріплення (рис.149, 150) – волоски приєднуються до клітини чи між клітин епідермісу. Найбільш поширений тип зустрічається, наприклад, у листі мати-й-мачухи, листі м'яти, листі шавлії, траві собачої кропиви, траві фіалки та ін.
1.11.2. Вирости з клітин епідермісу (Див. рис. 68, А; 69-74). Спостерігаються в квітках бузини, в траві золототисячника, пелюстках собачої кропиви, в траві тирличу бородатою, епідермісі плодів анісу та ін.
1.11.3. В основі волоска утворюється розетка з клітин епідермісу (Рис. 151, 152). Є у листі сенни, листі подорожника, траві материнки та інших.
1.11.4. Розширена основа волоска (Рис. 153). Зустрічається у листі подорожника.
1.11.5. Розширена частина основи волоска занурена в цоколь із субепідермальних тканин – емергенці (Див. рис. 115, 3). Можна спостерігати, наприклад, у листі кропиви.
1.11.6. Багатоклітинна основа волосків (Рис. 154). Можна спостерігати, наприклад, у траві низки.
Нерідко волоски обламуються, залишаючи на епідермісі місця свого прикріплення, що також необхідно відзначати як анатомо-діагностичну ознаку лікарської рослинної сировини.
ВТОРИНА ПОКРОВНА ТКАНИНА
Вторинну покривну тканину називають перидермою.Це складна покривна тканина стебел, коренів та кореневищ багаторічних рослин. Вона змінює епідерму осьових органів, яка поступово відмирає та злущується. Перидерма утворюється із феллогену (вторинної меристеми). Феллоген закладається в епідермі, субепідермальному шарі та навіть у глибоких шарах осьових органів. Клітини феллогену поділяються в такий спосіб назовні відкладають клітини пробки, а всередину – живі паренхімні клітини фелодерми. У клітинах фелодерми стебел містяться хлоропласти.
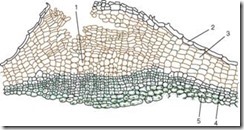 Корок складається з мертвих клітин, у яких клітинна стінка просочена жироподібною речовиною суберином Клітини розташовані рівними рядами, мають прямокутну форму(На поперечному зрізі), щільно прилягають один до одного, формуючи багатошаровий футляр. Корок охороняє внутрішні живі тканини від втрати вологи, різких температурних коливань та проникнення мікроорганізмів. Живі тканини, що лежать під пробкою, потребують газообміну та видалення надлишку вологи. Саме тому під продихом внаслідок поділу субепідермальних шарів (ще до появи перидерми), а надалі і феллогену відкладаються живі, рихло розташовані, з безліччю міжклітин паренхімні клітини, звані виконує тканинанью, яка розриває епідерму і створює можливість газообміну та транспірації з зовнішнім середовищем. Це структурна освітаназивають сочевічкою(Рис. 2.12).
Корок складається з мертвих клітин, у яких клітинна стінка просочена жироподібною речовиною суберином Клітини розташовані рівними рядами, мають прямокутну форму(На поперечному зрізі), щільно прилягають один до одного, формуючи багатошаровий футляр. Корок охороняє внутрішні живі тканини від втрати вологи, різких температурних коливань та проникнення мікроорганізмів. Живі тканини, що лежать під пробкою, потребують газообміну та видалення надлишку вологи. Саме тому під продихом внаслідок поділу субепідермальних шарів (ще до появи перидерми), а надалі і феллогену відкладаються живі, рихло розташовані, з безліччю міжклітин паренхімні клітини, звані виконує тканинанью, яка розриває епідерму і створює можливість газообміну та транспірації з зовнішнім середовищем. Це структурна освітаназивають сочевічкою(Рис. 2.12).
Мал. 2.12.Будова перидерми з чечевичкою: 1 - виконує тканину чечевички; 2 – залишки епідерми; 3 – пробка (фелема); 4 – фелоген; 5 – фелодерма
Чечевички, що мають вигляд невеликих горбків, чітко виділяються на поверхні пагонів дерев та чагарників (рис. 2.13).
На стовбурах берези їх залишки спостерігаються у вигляді характерних поперечних чорних смужок, біля осики вони набувають форми ромбів.
Третинна покривна тканина
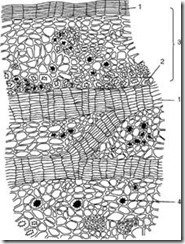 Корка (ритідом)є третинною покривною тканиною, яка утворюється у багаторічних рослин в корені, стеблі, кореневищі. Щороку у глибших шарах закладається новий шарфеллогену і утворюється перидерма. Зовнішній шар перидерми – пробка – ізолює всі тканини, що лежать вище, в результаті чого вони відмирають. Таким чином, сукупність численних перидерм з тканинами, що відмерли між ними, і є кіркою (рис. 2.14).
Корка (ритідом)є третинною покривною тканиною, яка утворюється у багаторічних рослин в корені, стеблі, кореневищі. Щороку у глибших шарах закладається новий шарфеллогену і утворюється перидерма. Зовнішній шар перидерми – пробка – ізолює всі тканини, що лежать вище, в результаті чого вони відмирають. Таким чином, сукупність численних перидерм з тканинами, що відмерли між ними, і є кіркою (рис. 2.14).
Мал. 2.14.Шкіра дуба: 1 – шари пробки; 2 – волокна; 3 – залишки первинної кори; 4 – друзі оксалату кальцію