Актиноміцети (Actinomycetales, від грец. Aktis - промінь, mykes - гриб) - це бактерії, що гілкуються, належать типу Актинобактерії (Actinobacteria). Вони є частиною нормальної мікрофлори. травної системиназемних хребетних і безхребетних тварин, а також у достатку присутні в ґрунті і відіграють найважливішу роль в екології та кругообігу речовин у ґрунті.
Ці мікроорганізми є збудниками багатьох опортуністичних патологій – таких, що виникають внаслідок зниження функції імунної системиорганізму. Актиноміцети широко використовуються в біотехнології, оскільки є джерелом цілого ряду антибактеріальних та протипухлинних речовин.
Мал. 1. Стрептоміцети синтезують величезну кількість антибактеріальних та протипухлинних препаратів.
Будова актиноміцетів: чому все ж таки бактерії, а не гриби?
1. Організація генетичного матеріалу
Спадковий матеріал Актиноміцетів укладено в одній молекулі дезоксирибонуклеїнової кислоти, що має кільцеву форму і вільно розташовується в цитоплазмі - така ж форма організації генетичного матеріалу, звана нуклеоїдом, характерна і для інших бактерій. У грибів генетичний матеріал організований і входить до складу клітинного ядра.
ДНК актиноміцетів містить велика кількістьГЦ-пар (65-75% загальної кількості нуклеотидів). Ця ознака постійна, не залежить від мутацій і тому використовується в систематиці мікроорганізмів. Такий вміст ГЦ-пар робить ДНК актиноміцетів дуже тугоплавкою, тому на аналіз ДНК актиноміцетів витрачається більше часу в порівнянні з іншими бактеріями.

Мал. 2. Схематична будоваклітинної стінки Гр+бактерій.

Мал. 3. Актиноміцети, забарвлені за Грамом.
Актиноміцети мають щільну бактеріальну клітинну стінку, яка розташована зовні цитоплазматичної мембраниі зумовлює їхнє позитивне фарбування за Грамом. Як і в інших Грам-позитивних бактерій, вона складається з кількох десятків шарів полімеру муреїну (пептидоглікану), який пронизаний тейхоєвими та ліпотейхоєвими кислотами. Ліпотейхові кислоти заякорені в цитоплазматичній мембрані бактерії і з'єднують її з клітинною стінкою. Тейхові кислоти надають клітинній стінці негативний заряд. Клітинна стінка грибів складається з інших полімерів - хітину і глюкану.
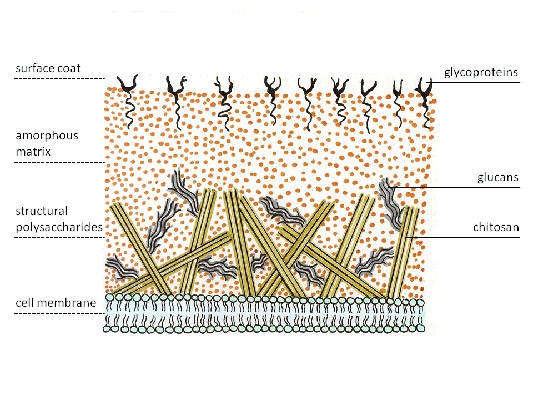
Мал. 4. Клітинна стінка грибів.
3. Клітинні органели
Актиноміцети, подібно до інших бактерій, не мають мембранних органел. У актиноміцетів 70S рибосоми, тоді як гриби мають 80S рибосоми, як і інші еукаріотичні організми.
4. Зростання колоній
Утворення міцелію при зростанні - це те, що найбільше ріднить актиноміцети з грибами. Міцелій у разі актиноміцетів являє собою гіф, що гілкується сукупність. Гіфи розділені перегородками на довгі бактеріальні клітини, які містять кілька нуклеоїдів. Перегородки у ряду видів можуть відбуватися у взаємно перпендикулярному напрямі. Гілкуються гіфи шляхом брунькування.
Міцелій, що вростає в субстрат (землю, мул або живильне середовище), називається субстратним. Він забезпечує колонію поживними речовинами. Над субстратом височить повітряний міцелій, що надає колонії «пухнастість» — він утворює суперечки, а також так звані «вторинні метаболіти» (на відміну від «первинних метаболітів» субстратного міцелію), серед яких багато антибактеріальних речовин.
Життєвий цикл та фізіологія актиноміцетів
У процесі життєвого циклу більша частинаактиноміцети утворюють суперечки. Деякі актиноміцети розмножуються шляхом фрагментації міцелію.
1. Спороутворення
Суперечки актиноміцетів походять із повітряного міцелію. Це екзоспори - вони розвиваються зовні материнської клітини. Гіфи повітряного міцелію, у тому числі розвиваються суперечки, називаються спороносцами. Суперечки можуть утримуватися в потовщенні на кінці спороносця – спорангії (наприклад, у стрептоміцетів, актинопланів та племілій), а можуть розташовуватися ланцюжком вздовж спорангієносця (наприклад, у нокардій та актиномадур).
За кількістю суперечок, що утворюються, актиноміцети діляться на:
- Моноспорові (наприклад, Saccaromonospora, Micromonospora, Thermomonospora) – утворюють поодинокі суперечки, частіше шляхом відбрунькування та подальшого відділення перегородкою від материнського гіфа;
- Олігоспорові (наприклад, Actinomadura) – утворюють короткі ланцюжки спор уздовж спороносця;
- Поліспорові (більшість інших актиноміцетів, наприклад, Streptomyces, Frankia, Geodermatophilus) – утворюють безліч суперечок, укладених у спорангії.

Мал. 5. Спорангій актиноміцетів роду Frankia.
Суперечки актиноміцетів можуть бути рухливими – у такому разі, суперечка має джгутик і може пересуватися (суперечки актинопланів, геодерматофілів та дерматофілів). Найчастіше суперечки нерухомі і поширюються вітром, водою чи тваринами.

Мал. 6. Дерматофіли, світлова мікроскопія.
Споруляція у актиноміцетів особливо активно протікає у несприятливих умовах. Стійкість спор до нагрівання невелика порівняно зі спорами інших бактерій, проте висушування вони витримують не гірше за інших, а тому мають колосальне пристосувальне значення. Актиноміцети домінують над іншими мікроорганізмами в сухих пустельних ґрунтах.
Проростання зграї потребує певної вологості зовнішнього середовища. У присутності води спору набухає, у ній активуються ферменти та запускаються метаболічні процеси, що супроводжуються виходом ростових трубок (майбутніх бактеріальних тіл) та синтезом нуклеїнових кислот.
2. Тип дихання
Більшість актиноміцетів - аероби (потребують кисню для підтримки життєдіяльності). Факультативні анаероби (бактерії, здатні жити як за наявності, так і за відсутності кисню) зустрічаються серед видів з нетривалою міцеліальною стадією, що розмножуються фрагментацією міцелію.
3.Кислотостійкість
Актиноміцети мають ацидотолерантність - стійкість у кислому середовищі, яка дозволяє їм жити в насичених кислотами лісових грунтах. Кислотостійкість у лабораторії може бути визначена забарвленням препарату, що містить актиноміцети, по Цилю-Нільсену (фуксином з подальшим протравленням сірчаною кислотою та забарвленням метиленової синьки). Більшість актиноміцетів при такому забарвленні не знебарвлюються після протруювання кислотою і зберігають червоний фуксиновий колір. Лужне середовищенесприятлива для цих бактерій: при підвищеному рН вони схильні до спороутворення.
4. Особливості метаболізму

Мал. 7. Аеробні актиноміцети утворюють пігмент на скошеному агарі. Зліва направо: Actinomadura madurae, Nocardia asteroides, Micromonospora.
Вище згадувалося утворення повітряним міцелієм «вторинних метаболітів». Серед них:
- пігменти, що зумовлюють різне забарвлення повітряного міцелію при зростанні на середовищах;
- леткі пахучі речовини, що надають характерного запаху ґрунту після дощу, застояній воді, шкірним покривам деяких тварин;
- антибіотики:
a. протигрибкові – полієни;
b. протибактеріальні – наприклад, стрептоміцин, еритроміцин, тетрациклін, ванкоміцин;
c. протипухлинні – антрацикліни, блеоміцин.
Де мешкають актиноміцети?
Актиноміцети в найбільшій кількостівиявляються у ґрунтах, причому міцеліальних форм значно менше, ніж суперечка. Вони відіграють значну роль освіти гумусу, розкладаючи органічні речовини, важкодоступні для утилізації іншими бактеріями. Актиноміцети у зв'язку з цим використовують як санітарно-показові мікроорганізми в санітарно-епідеміологічній справі: виявлення їх у велику кількістьу ґрунті або воді вказує на наявність компосту у відповідному субстраті.

Мал. 8. Актиноміцети у компості.
Актиноміцети є симбіонтами багатьох рослин, допомагаючи їм фіксувати азот. У той же час багато мікроорганізмів цього класу є збудниками захворювань рослин.

Мал. 9. Стрептомікоз картоплі.
Вони також виявляються у складі нормальної мікрофлори травної системи цілого ряду тварин, починаючи від ґрунтових кільчастих хробаків(наприклад, дощових) і закінчуючи великою худобою.
Ці мікроорганізми допомагають розщеплювати целюлозу, яка в достатку присутня в рослинній їжі. У людини актиноміцети виявляються в порожнині рота (ясна та зубний наліт), кишечнику (дистальні відділи товстого кишечника), на шкірі (обличчя, крила носа, за вухами, між пальцями) та в органах дихальної системи(переважно у верхніх дихальних шляхах).
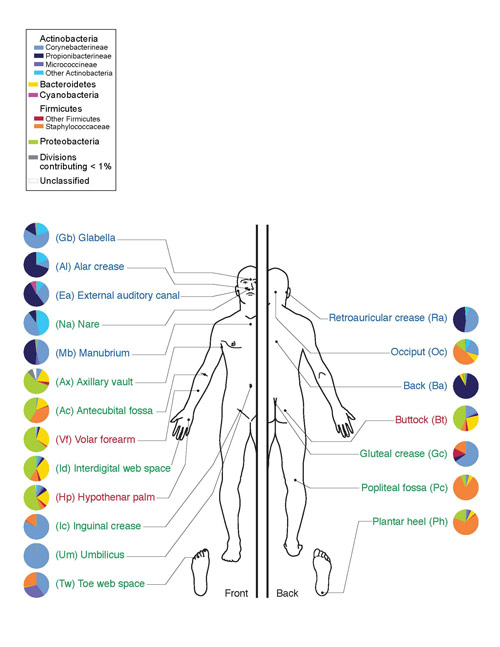
Мал. 10. Мікрофлора шкіри людини. Тип Актинобактерії позначені відтінками синього, клас Актиноміцети – яскраво-блакитним.
Актиноміцети за умови зниження імунної реактивності організму можуть стати причиною актиномікозів – опортуністичних захворювань, що полягають у формуванні актиномікозних гранульом – скупчень бактеріальних тіл, що нагадують зерна жовтої сірки («друз»), оточених імунокомпетентними клітинами. Запальна реакція веде до розплавлення гранульом, утворення свищів, що веде до перфорацій органів та рознесення бактерій кров'ю.

Мал. 11. Актиномікозна друза, забарвлення за Грамом.

Мал. 12. Актиномікоз верхньої щелепи у корови.

Мал. 13. Максілярний актиномікоз людини.
Актиноміцети – дивовижні організми, що до цих пір вводять в оману безліч вчених своєю схожістю з грибами. Поряд з потенційною небезпекою у вигляді опортуністичних актиномікозів, ці організми дарують людині плодоносний ґрунт та зброю для боротьби з інфекційними та онкологічними захворюваннями – антибіотики та цитостатики.
Хімічний склад та структура клітинної стінки грибів
Клітинна стінка грибів багатошарова, при цьому різні шари утворені структурними вуглеводами, що розрізняються за хімічним складом, які за хімічним складом можна розділити на 3 групи:
полімери глюкози(Глюкан, хітин, целюлоза). Глюкани складають зовнішній шар клітинної стінки більшості грибів. Внутрішній шар грибної клітинної стінки утворений ланцюжками хітину, надаючи їй жорсткості. Хітін заміщає целюлозу, яка у більшості грибів відсутня, але входить до складу клітинної стінки ооміцетів, які нині до типових грибів не належать. Деацетильований хітин отримав назву хітозан, який у комплексі з хітином утворює клітинну стінку зигоміцетів.
полімери інших моносахаридів (маннози, галактози та ін.)на відміну від вищих рослин, де вони складають основу матриксу під загальною назвоюгеміцелюлози, менш характерні для грибів. Виняток становлять дріжджі, у клітинних стінках яких особливо багато полімерів маннози, званих маннани. Припускають, що такий склад стінки краще забезпечує брунькування.
полімери вуглеводів, ковалентно пов'язані з пептидами (глікопротеїни)формують серединний шар багатошарової клітинної стінки та грають важливу рольяк у підтримці структурної цілісності клітин, так і в її обмінних процесах з навколишнім середовищем.
До інших специфічних особливостей грибної клітини відносяться: відсутність пластидщо зближує її з клітиною тварин;
Відсутність крохмалю, який у еуміцетів заміщається полісахаридом, близьким до тваринного крохмалю глікогену, у ооміцетів – полісахаридом, близьким до ламінарину бурих водоростей. Виробляється і низку специфічних для грибів запасних вуглеводів.
Вироблення специфічних вторинних метаболітів, з яких велику рольграють антибіотики, фіто- та мікотоксини, фітогормони.
До специфічних особливостей грибів належать також гетерокаріоз та парасексуальний процес.
У грибів дуже поширене явище гетерокаріозу чи різноядерності, у якому у одній клітині тривалий часзберігаються ядра, гетероалельні за деякими генами. Гетерокаріоз замінює гаплоїдним грибам гетерозиготність та сприяє швидкій адаптації грибів до мінливих умов. Наявність різноядерності обумовлено поруч специфічних особливостейгрибів:
1. наявність більше одного ядра у клітці
2. специфічне будова клітинної перегородки, у якій є одне чи кілька наскрізних отворів, званих порами, якими ядра можуть мігрувати з однієї клітини до іншої
3. гіфи всередині однієї колонії і навіть різних близько розташованих колоній, що виросли з різних суперечок одного виду гриба, часто зростаються, внаслідок чого можливий обмін ядрами різних штамів.
Парасексуальний (псевдостатевий)процес. Якщо в гетерокаріотичних клітинах зливаються ядра, гетероалельні за якимось локусом, виникає гетерозиготне диплоїдне ядро. Воно може потрапити у суперечку та дати початок диплоїдному гетерозиготному клону. У процесі мітозів диплоїдні ядра можуть повертатися в гаплоїдний стан внаслідок втрати одного набору хромосом або може відбуватися обмін ділянками хромосом (мітотичний кросинговер). Обидва процеси супроводжуються рекомбінацією батьківських генів і, отже, фенотипів. Парасексуальна (безстатева) рекомбінація дуже рідкісне явище, що не перевищує одного ядра на мільйон, але внаслідок величезної кількості ядер у міцелії постійно спостерігається у популяціях грибів.
Розмноження- Вегетативне, безстатеве, статеве.
Вегетативне- Фрагментація талому, утворення хламідоспор, які після періоду спокою проростають у міцелій, брунькування у дріжджів.
Безстатеверозмноження у різних грибів може здійснюватися рухомими та нерухомими спорами. Зооспори утворюють відносно невелику групу грибів і грибоподібних організмів – водні та деякі наземні, у яких чітко простежуються генетичні зв'язкиз водними грибами та водоростями. Структура джгутиків є важливим діагностичною ознакоюпри віднесенні до певного царства. У переважної більшості грибів при безстатевому розмноженніутворюються нерухомі суперечки, що свідчить про їх дуже давній вихід сушу. За місцем формування і локалізації розрізняють ендогенні спорангіоспори, що утворюються в спорангіях, і екзогенні (конідії), що розвиваються на спеціальних гіфах – конідієносцях. Конідії утворюються у більшості грибів (аскоміцети, базидіоміцети, деякі зигоміцети) складаючи конідіальне спороношення, яке відрізняється величезним різноманіттям і широко використовується для діагностики грибів.
Статеве розмноження грибівмає свою специфіку як у морфології статевого процесу, так і в механізмах генетичної та фізіологічного регулюваннястаті та передачі спадкової інформації.
Соматогамія– найпоширеніший і найпростіший тип статевого процесу, полягає у злитті двох недиференційованих на гамети соматичних клітин. Іноді він протікає навіть без злиття клітин – зливаються ядра усередині клітини. Зустрічається у більшості базидіоміцетів, сумчастих дріжджів та деяких ін. таксонів.
Гаметангіогамія- На гаплоїдному міцелії відокремлюються гаметангії, при статевому процесі зливається їх вміст. Такий статевий процес характерний більшості сумчастих грибів. Варіантом гаметангіогамії є зигогамія у зигоміцетів.
Гаметогаміяу вигляді ізогетеро та оогамії у грибів зустрічається значно рідше, ніж у інших еукаріотів. Ізо-і гетерогамія зустрічається лише у хітридіоміцетів. Класична оогамія з утворенням сперматозоїдів та яйцеклітин у грибів не виражена, а трапляються сильно змінені варіанти.
За особливостями регуляції статі гриби виділяють кілька типів статевого процесу.
Гінандроміксисможна розглянути на прикладі дводомних ооміцетів, у яких оогонії та антеридії розвиваються на різних таломах, наприклад, фітофтора або картопляний гриб. Якщо генетично однорідний штам вирощувати в монокультурі, він розмножується тільки безстатевим способом. Якщо поруч виявляються міцелії двох штамів, то вони простежуються морфогенетичні зміни під впливом хімічних виділень стероїдної природи – статевих феромонов. Антеридіол індукує утворення антеридіїв у партнера, а оогоніол-оогоніїв. При цьому регуляція статі має відносний характер: чи штам утворюватиме антеридії або ж оогонії, залежить від кількісного співвідношення відповідних феромонів у нього та його партнера. Звідси назва статевого процесу – гінандроміксис.
Діміксис чи гетероталізм.Вже давно було помічено, що гриби можуть бути гомо-або гетероталічними. У гомоталличних видів при статевому процесі зливаються генетично ідентичні ядра всередині міцелію. У гетероталлічних видів для проходження статевого циклу необхідно на якомусь етапі (різному у різних видівгрибів) злиття нащадків спор (точніше, їх ядер). Для статевої сумісності двох штамів необхідна генетична відмінність (гетероаллельність) у певних локусах, званих локусами спарювання. У більшості грибів (зигоміцети, аскоміцети, частина базидіоміцетів) є один локус спарювання з двома алелями. Локус спарювання складається з кількох генів, які управляють синтезом статевих феромонів. Такий гетероталізм називається однофакторним або біполярним. Нащадок таких грибів після мейозу поділяється на дві самонесумісні, але взаємно сумісні групи у відсотковому співвідношенні 1:1, тобто. ймовірність спорідненого (інбридингу) та неспорідненого (аутбридингу) схрещування становить як і у двостатевих вищих еукаріотів 50%.
У геномі вищих базидіальних грибів є два локуси спарювання - A і B, причому сумісні один з одним тільки штами гетероалельні по обидва локуси (Ax Bx сумісний з Ay By, але не з Ax By і Ay Bx). Такий гетероталізм називають двофакторним або тетраполярним. Він знижує ймовірність інбридингу до 25%.
Діафороміксис- У вищих базидіоміцетів є не два, а багато алелів локусу спарювання, що випадково зустрічаються у різних штамів, що складають популяцію. Такий контроль спарювання забезпечує 100% ймовірність аутбридингу, так як штами, що мають різні алелі, взаємосумісні, а алелей дуже багато. Внаслідок цього утворюються панміксні гібридні популяції.
Життєві цикли грибівтакож різноманітні як і гриби. Основні цикли, їхня приналежність до відділів грибів
1.Безстатевий цикл характерний для величезної групи недосконалих або мітогрибів, що втратили статеве розмноження. Поділ їх ядер виключно мітотичний. Переважна більшість мітогрибів відноситься до сумчастих грибів, але у зв'язку зі втратою статевого процесу складають формальну групунедосконалих грибів або дейтероміцетів.
2. Гаплоїдний цикл. Вегетативний тал несе гаплоїдні ядра. Після статевого процесу (сингамії) диплоїдна зигота (зазвичай після періоду спокою) ділиться мейотично – зиготичний мейоз. Характерний для зигоміцетів та багатьох хитридіоміцетів.
3. Гаплоїдно-дикаріотичний цикл характеризується тим, що після злиття вмісту гаметангіїв (гаметангіогамія) або соматичних клітин гаплоїдного міцелію (соматогамія) ядра утворюють дикаріони (пари генетично різних ядер). Вони одночасно діляться, утворюючи дикаріотичний міцелій. Статевий процес завершується злиттям ядер дикаріону, що утворюється зигота ділиться мейозом без періоду спокою. Мейоспори становлять статеве спороношення сумчастих та базидіальних грибів у вигляді аскоспор та базидіоспор. При їхньому проростанні утворюється гаплоїдний міцелій. У переважної більшості сумчастих грибів (крім дріжджів та тафринових грибів) у життєвому цикліпереважає гаплоїдна фаза у вигляді вегетативного міцелію (анаморфу), дикаріотична фаза короткочасна та представлена аскогенними гіфами, на яких утворюються сумки (телеоморфи). У базидіальних грибів у життєвому циклі переважає дикаріотична фаза, гаплоїдна фаза короткочасна.
4. Гаплоїдно-диплоїдний цикл у вигляді ізоморфної зміни поколінь у грибів зустрічається рідко (деякі дріжджі та водні хитридіоміцети).
5. Диплоїдний цикл характерний для ооміцетів та деяких сумчастих дріжджів. Вегетативний талом диплоїдний, гаметичний мейоз, спостерігається при формуванні гаметангіїв або гамет.
3. Екологічні групигрибів.
Гриби та грибоподібні організми входять у всі наземні та водні екосистеми, як найважливіша частинагетеротрофного блоку, разом із бактеріями займаючи трофічний рівеньредуцентів. Широке поширеннягрибів у біосфері визначається поруч найважливіших особливостей:
1. Наявність у більшості міцеліальної структури талому. (Дозволяє швидко освоювати субстрат, мати велику поверхню контакту з середовищем).
2. Велика швидкість росту та розмноження, що дозволяють у короткі термінизаселяти великі масисубстрату, утворюючи величезну кількість суперечок і поширювати їх у великі відстані.
3. Висока метаболічна активність, що виявляється у широкому діапазоні значень екологічних факторів.
4. Висока швидкість генетичної рекомбінації, значна біохімічна мінливість, екологічна пластичність.
5. Здатність швидко переходити до стану анабіозу, переживати несприятливі умовипротягом тривалого часу.
Головним екологічним факторомдля грибів є харчовий субстрат. Стосовно цього чинника виділяють основні групи грибів, які називаються трофічними групами.
4. Коротка характеристикавідділів грибів.
Прокаріотична клітина
Прокаріоти- організми, що не володіють, на відміну від еукаріотів, оформленим клітинним ядром та іншими внутрішніми мембранними органоїдами(за винятком плоских цистерн у фотосинтезуючих видів, наприклад, у ціанобактерій). Єдина велика кільцева (у деяких видів - лінійна) дволанцюжкова молекула ДНК, в якій міститься основна частина генетичного матеріалу клітини (так званий нуклеоїд), не утворює комплексу з білками-гістонами (так званого хроматину). До прокаріотів належать бактерії, у тому числі ціанобактерії (синьо-зелені водорості), та археї. Нащадками прокаріотичних клітин є органели еукаріотичних клітин – мітохондрії та пластиди. Основний вміст клітини, що заповнює її об'єм, - в'язка зерниста цитоплазма.
Еукаріотична клітина
Еукаріоти- організми, що мають, на відміну від прокаріотів, оформленим клітинним ядром, відмежованим від цитоплазми ядерною оболонкою. Генетичний матеріал укладений у кількох лінійних дволанцюгових молекулах ДНК (залежно від виду організмів їх число на ядро може коливатися від двох до декількох сотень), прикріплених зсередини до мембрани клітинного ядра і які утворюють у переважній більшості (крім динофлагелят) комплекс з білками-гістонами. хроматином. У клітинах еукаріотів є система внутрішніх мембран, що утворюють, крім ядра, ряд інших органоїдів (ендоплазматична мережа, апарат Гольджі та ін.). Крім того, у переважній більшості є постійні внутрішньоклітинні симбіонти-прокаріоти - мітохондрії, а у водоростей та рослин - також і пластиди.
2) цитоплазма
Цитоплазма - внутрішнє середовищеживої або померлої клітини, крім ядра та вакуолі, обмежена плазматичною мембраною. Включає гіалоплазму - основна прозора речовина цитоплазми, що знаходяться в ній обов'язкові клітинні компоненти - органели, а також різні непостійні структури - включення.
До складу цитоплазми входять усі види органічних та неорганічних речовин. У ній присутні також нерозчинні відходи обмінних процесів та запасні поживні речовини. Основна речовина цитоплазми – вода.
Цитоплазма постійно рухається, перетікає всередині живої клітини, переміщуючи разом із собою різні речовини, включення та органоїди. Цей рух називається циклозом. У ній протікають усі процеси обміну речовин.
Цитоплазма здатна до зростання та відтворення та при частковому видаленні може відновитися. Проте нормально функціонує цитоплазма лише у присутності ядра. Без нього довго існувати цитоплазма неспроможна, як і і ядро без цитоплазми.
Найважливіша роль цитоплазми полягає в поєднанні всіх клітинних структур (компонентів) та забезпеченні їх хімічної взаємодії. Також цитоплазма підтримує тургор(об'єм) клітини, підтримання температури.
3)Клітинна стінка
Клітинна стінка- Жорстка оболонка клітини, розташована зовні від цитоплазматичної мембрани і виконує структурні, захисні та транспортні функції. Виявляється у більшості бактерій, архей, грибів та рослин. Тварини і багато найпростіших мають клітинної стінки.
Клітинні стінкипрокаріот
Клітинні стінки бактерій складаються з пептидоглікану (муреїну) і бувають двох типів: грампозитивного та грамнегативного. Клітинна стінка грампозитивного типу складається виключно з товстого шару пептидоглікану, що щільно прилягає до клітинної мембрани і пронизаного тейхоєвими або потейхоєвими кислотами. При грамнегативному типі шар пептидоглікану істотно тонше, між ним і плазматичною мембраною знаходиться периплазматичний простір, а зовні клітина оточена ще однією мембраною, представленою т.з. ліпополісахаридом і є пірогенним ендотоксином грамнегативних бактерій.
Клітинні стінки грибів
Клітинні стінки грибів складаються з хітину та глюканів.
Характерні особливості грибів
До грибів у широкому значеннівідносять гетеротрофні еукаріоти з обзорного типу харчування. З особливостями харчування пов'язані характерні рисибудови та способу життя цих організмів. Найбільш характерні ознакигрибів:
- Найбільш поширеною формою вегетативного тіла є міцелій, що складається з ниток, що багаторазово гілкуються, які пронизують субстрат і всмоктують з нього розчинені у воді поживні речовини.
– Органічні речовиниу субстраті, які є джерелом харчування для грибів, у більшості випадків знаходяться у формі біополімерів, нездатних проникати через клітинну мембрану. У клітинах грибів виробляються ферменти деполімерази, які виділяються в субстрат і руйнують полімери до більш простих органічних сполук, здатних транспортуватися до грибної клітини. Такі ферменти прийнято називати екзоферментами.
– Активне всмоктування речовин із субстрату здійснюється завдяки величезному тургорному тиску, який у грибних клітинах набагато вищий у порівнянні з іншими еукаріотними організмами.
– Для грибів дуже характерне утворення спор, які з'являються як при безстатевому, так і статевому розмноженні. Впізнавання грибів можливе не по вегетативному тілу, а по спороношенню. У зв'язку з розташуванням вегетативного тіла всередині субстрату і нерухомим способом життя, в момент розмноження на грибному міцелії утворюються різноманітні спороносні структури, які піднімаються над субстратом і спори, що утворюються на них, поширюються повітряними потоками на дуже далекі відстані.
Сучасні системигрибів
за сучасним уявленням, заснованих на вивченні геномів екоморфу «гриби» є збірною групою, в якій виділяють справжні гриби, царство Mycota або Fungi, яких більшість і менша за кількістю видових таксонів і менше різноманітна групагрибоподібних організмів (псевдоміцети) входить до складу царства Straminopila, що поєднує також пофарбовані водорості (Ochrophyta). Царство справжні гриби поєднує 4 відділи, що відрізняються будовою вегетативного тіла та особливостями розмноження: Chytridiomycota, Zygomycota, Ascomycota, Basidiomycota.
Таксономічно значущі ознаки, що використовуються при побудові систем грибів
-Будова вегетативного тіла
– Хімічний склад та структура клітинної стінки
– Тип статевого процесу
- Типи спороношень
Типи таломів грибів та грибоподібних організмів
Амебоїдний або плазмодіальнийТалом є позбавленою клітинної стінки одноядерною або багатоядерною клітиною, оточеною плазматичною мембраною і ущільненим периферичним білковим шаром - перипластом. Для закріплення в субстраті та збільшення всмоктуючої поверхні можуть утворюватися коренеподібні вирости, що у сукупності утворюють ризоміцелій. При розмноженні або весь талом перетворюється на єдиний спорангій (холокарпічний талом), або утворюється кілька спорангіїв (еукарпічний талом).
Міцеліальний таломта його видозміни. Основний структурний елемент- Гіфа. Членистий та нечленистий міцелій. Септи, їхня освіта.
Видозміни міцелію.
Спокою структури: хламідоспори і склероції.
Провідні та підтримуючі структури: тяжі та ризоморфи.
Інфекційні структури: апресорії та гаусторії
Дріжджоподібний брухт талломявляє собою окремі, покриті оболонкою клітини або нестійкі, ланцюжки клітин, що легко розпадаються, які утворюються при брунькаванні внаслідок нерозбіжності материнських і дочірніх клітин. Постійний та тимчасовий дріжджоподібний стан таломів.
Хімічний склад та структура клітинної стінки грибів
Клітинна стінка грибів багатошарова, при цьому різні шари утворені структурними вуглеводами, що розрізняються за хімічним складом, які за хімічним складом можна розділити на 3 групи:
полімери глюкози(Глюкан, хітин, целюлоза). Глюкани складають зовнішній шар клітинної стінки більшості грибів. Внутрішній шар грибної клітинної стінки утворений ланцюжками хітину, надаючи їй жорсткості. Хітін заміщає целюлозу, яка у більшості грибів відсутня, але входить до складу клітинної стінки ооміцетів, які нині до типових грибів не належать. Деацетильований хітин отримав назву хітозан, який у комплексі з хітином утворює клітинну стінку зигоміцетів.
полімери інших моносахаридів (маннози, галактози та ін.)на відміну від вищих рослин, де вони становлять основу матриксу під загальною назвою геміцелюлоза, менш характерні для грибів. Виняток становлять дріжджі, у клітинних стінках яких особливо багато полімерів маннози, званих маннани. Припускають, що такий склад стінки краще забезпечує брунькування.
полімери вуглеводів, ковалентно пов'язані з пептидами (глікопротеїни)формують серединний шар багатошарової клітинної стінки і відіграють важливу роль, як у підтримці структурної цілісності клітин, так і в обмінних процесах з навколишнім середовищем.
До інших специфічних особливостей грибної клітини відносяться: відсутність пластидщо зближує її з клітиною тварин;
Відсутність крохмалю, що у еуміцетів заміщається полісахаридом, близьким до тваринного крохмалю глікогену, у ооміцетів – полісахаридом, близькому до ламінарину бурих водоростей. Виробляється і низку специфічних для грибів запасних вуглеводів.
Вироблення специфічних вторинних метаболітів, з яких велику роль відіграють антибіотики, фіто- та мікотоксини, фітогормони.
До специфічних особливостей грибів належать також гетерокаріоз та парасексуальний процес.
У грибів дуже широко поширене явище гетерокаріозу або різноядерності, при якому в одній клітині тривалий час зберігаються ядра, гетероалельні за деякими генами. Гетерокаріоз замінює гаплоїдним грибам гетерозиготність та сприяє швидкій адаптації грибів до мінливих умов. Наявність різноядерності обумовлено низкою специфічних особливостей грибів:
1. наявність більше одного ядра у клітці
2. специфічне будова клітинної перегородки, у якій є одне чи кілька наскрізних отворів, званих порами, якими ядра можуть мігрувати з однієї клітини до іншої
3. гіфи всередині однієї колонії і навіть різних близько розташованих колоній, що виросли з різних суперечок одного виду гриба, часто зростаються, внаслідок чого можливий обмін ядрами різних штамів.
Парасексуальний (псевдостатевий)процес. Якщо в гетерокаріотичних клітинах зливаються ядра, гетероалельні за якимось локусом, виникає гетерозиготне диплоїдне ядро. Воно може потрапити у суперечку та дати початок диплоїдному гетерозиготному клону. У процесі мітозів диплоїдні ядра можуть повертатися в гаплоїдний стан внаслідок втрати одного набору хромосом або може відбуватися обмін ділянками хромосом (мітотичний кросинговер). Обидва процеси супроводжуються рекомбінацією батьківських генів і, отже, фенотипів. Парасексуальна (безстатева) рекомбінація дуже рідкісне явище, що не перевищує одного ядра на мільйон, але внаслідок величезної кількості ядер у міцелії постійно спостерігається у популяціях грибів.
Розмноження- Вегетативне, безстатеве, статеве.
Вегетативне- Фрагментація талому, утворення хламідоспор, які після періоду спокою проростають у міцелій, брунькування у дріжджів.
Безстатеверозмноження у різних грибів може здійснюватися рухомими та нерухомими спорами. Зооспори утворюють відносно невелику групу грибів і грибоподібних організмів – водні та деякі наземні, у яких чітко простежуються генетичні зв'язки з водними грибами та водоростями. Структура джгутиків є важливою діагностичною ознакою при віднесенні до певного царства. У переважної більшості грибів при безстатевому розмноженні утворюються нерухомі суперечки, що свідчить про їх дуже давній вихід сушу. За місцем формування і локалізації розрізняють ендогенні спорангіоспори, що утворюються в спорангіях, і екзогенні (конідії), що розвиваються на спеціальних гіфах – конідієносцях. Конідії утворюються у більшості грибів (аскоміцети, базидіоміцети, деякі зигоміцети) складаючи конідіальне спороношення, яке відрізняється величезним різноманіттям і широко використовується для діагностики грибів.
Статеве розмноження грибівмає свою специфіку як у морфології статевого процесу, так і в механізмах генетичної та фізіологічної регуляції статі та передачі спадкової інформації.
Соматогамія– найпоширеніший і найпростіший тип статевого процесу, полягає у злитті двох недиференційованих на гамети соматичних клітин. Іноді він протікає навіть без злиття клітин – зливаються ядра усередині клітини. Зустрічається у більшості базидіоміцетів, сумчастих дріжджів та деяких ін. таксонів.
Гаметангіогамія- На гаплоїдному міцелії відокремлюються гаметангії, при статевому процесі зливається їх вміст. Такий статевий процес характерний більшості сумчастих грибів. Варіантом гаметангіогамії є зигогамія у зигоміцетів.
Гаметогаміяу вигляді ізогетеро та оогамії у грибів зустрічається значно рідше, ніж у інших еукаріотів. Ізо-і гетерогамія зустрічається лише у хітридіоміцетів. Класична оогамія з утворенням сперматозоїдів та яйцеклітин у грибів не виражена, а трапляються сильно змінені варіанти.
За особливостями регуляції статі гриби виділяють кілька типів статевого процесу.
Гінандроміксисможна розглянути на прикладі дводомних ооміцетів, у яких оогонії та антеридії розвиваються на різних таломах, наприклад, фітофтора або картопляний гриб. Якщо генетично однорідний штам вирощувати у монокультурі, він розмножується лише безстатевим способом. Якщо поруч виявляються міцелії двох штамів, то вони простежуються морфогенетичні зміни під впливом хімічних виділень стероїдної природи – статевих феромонов. Антеридіол індукує утворення антеридіїв у партнера, а оогоніол-оогоніїв. При цьому регуляція статі має відносний характер: чи штам утворюватиме антеридії або ж оогонії, залежить від кількісного співвідношення відповідних феромонів у нього та його партнера. Звідси назва статевого процесу – гінандроміксис.
Діміксис чи гетероталізм.Вже давно було помічено, що гриби можуть бути гомо-або гетероталічними. У гомоталличних видів при статевому процесі зливаються генетично ідентичні ядра всередині міцелію. У гетероталлічних видів для проходження статевого циклу необхідно на якомусь етапі (різному у різних видів грибів) злиття нащадків спор (точніше, їх ядер). Для статевої сумісності двох штамів необхідна генетична відмінність (гетероаллельність) у певних локусах, званих локусами спарювання. У більшості грибів (зигоміцети, аскоміцети, частина базидіоміцетів) є один локус спарювання з двома алелями. Локус спарювання складається з кількох генів, які управляють синтезом статевих феромонів. Такий гетероталізм називається однофакторним або біполярним. Нащадок таких грибів після мейозу поділяється на дві самонесумісні, але взаємно сумісні групи у відсотковому співвідношенні 1:1, тобто. ймовірність спорідненого (інбридингу) та неспорідненого (аутбридингу) схрещування становить як і у двостатевих вищих еукаріотів 50%.
У геномі вищих базидіальних грибів є два локуси спарювання - A і B, причому сумісні один з одним тільки штами гетероалельні по обидва локуси (Ax Bx сумісний з Ay By, але не з Ax By і Ay Bx). Такий гетероталізм називають двофакторним або тетраполярним. Він знижує ймовірність інбридингу до 25%.
Діафороміксис- У вищих базидіоміцетів є не два, а багато алелів локусу спарювання, що випадково зустрічаються у різних штамів, що складають популяцію. Такий контроль спарювання забезпечує 100% ймовірність аутбридингу, так як штами, що мають різні алелі, взаємосумісні, а алелей дуже багато. Внаслідок цього утворюються панміксні гібридні популяції.
Життєві цикли грибівтакож різноманітні як і гриби. Основні цикли, їхня приналежність до відділів грибів
1.Безстатевий цикл характерний для величезної групи недосконалих або мітогрибів, що втратили статеве розмноження. Поділ їх ядер виключно мітотичний. Переважна більшість мітогрибів відноситься до сумчастих грибів, але у зв'язку зі втратою статевого процесу складають формальну групу недосконалих грибів або дейтероміцетів.
2. Гаплоїдний цикл. Вегетативний тал несе гаплоїдні ядра. Після статевого процесу (сингамії) диплоїдна зигота (зазвичай після періоду спокою) ділиться мейотично – зиготичний мейоз. Характерний для зигоміцетів та багатьох хитридіоміцетів.
3. Гаплоїдно-дикаріотичний цикл характеризується тим, що після злиття вмісту гаметангіїв (гаметангіогамія) або соматичних клітин гаплоїдного міцелію (соматогамія) ядра утворюють дикаріони (пари генетично різних ядер). Вони одночасно діляться, утворюючи дикаріотичний міцелій. Статевий процес завершується злиттям ядер дикаріону, що утворюється зигота ділиться мейозом без періоду спокою. Мейоспори становлять статеве спороношення сумчастих та базидіальних грибів у вигляді аскоспор та базидіоспор. При їхньому проростанні утворюється гаплоїдний міцелій. У переважної більшості сумчастих грибів (крім дріжджів та тафринових грибів) у життєвому циклі переважає гаплоїдна фаза у вигляді вегетативного міцелію (анаморфа), дикаріотична фаза короткочасна та представлена аскогенними гіфами, на яких утворюються сумки (телеморф). У базидіальних грибів у життєвому циклі переважає дикаріотична фаза, гаплоїдна фаза короткочасна.
4. Гаплоїдно-диплоїдний цикл у вигляді ізоморфної зміни поколінь у грибів зустрічається рідко (деякі дріжджі та водні хитридіоміцети).
5. Диплоїдний цикл характерний для ооміцетів та деяких сумчастих дріжджів. Вегетативний талом диплоїдний, гаметичний мейоз, спостерігається при формуванні гаметангіїв або гамет.
3. Екологічні групи грибів.
Гриби та грибоподібні організми входять у всі наземні та водні екосистеми, як найважливіша частина гетеротрофного блоку, разом із бактеріями займаючи трофічний рівень редуцентів. Широке поширення грибів у біосфері визначається низкою найважливіших особливостей:
1. Наявність у більшості міцеліальної структури талому. (Дозволяє швидко освоювати субстрат, мати велику поверхню контакту з середовищем).
2. Велика швидкість зростання та розмноження, що дозволяють у короткі терміни заселяти великі маси субстрату, утворюючи величезну кількість суперечок та поширювати їх на великі відстані.
3. Висока метаболічна активність, що виявляється у широкому діапазоні значень екологічних факторів.
4. Висока швидкість генетичної рекомбінації, значна біохімічна мінливість, екологічна пластичність.
5. Здатність швидко переходити до стану анабіозу, переживати несприятливі умови протягом багато часу.
Головним екологічним фактором для грибів є харчовий субстрат. Стосовно цього чинника виділяють основні групи грибів, які називаються трофічними групами.
4. Коротка характеристика відділів грибів.
МІКРОБІОЛОГІЯ, 2010, том 79, № 6, с. 723-733
УДК 582.281(047)
КЛІТИННА СТІНКА ГРИБІВ: СУЧАСНІ ПОДАННЯ ПРО СКЛАД І БІОЛОГІЧНУ ФУНКЦІЮ
© 2010 р. Є. П. Феофілова1
Установа Російської академіїнаук Інститут мікробіології ім. С.М. Виноградського РАН, Москва
Надійшла до редакції 05.11.2009 р.
Огляд присвячений мало вивченій поверхневій структурі клітини міцеліальних грибів – клітинній стінці (КС). Наводяться дані про методи виділення та перевірки чистоти на відсутність цитоплазматичного вмісту у фракції КС та про її хімічний склад. Детально розглядаються структурні (каркасні) та внутрішньоструктурні компоненти КС - амінополісахариди, а- та в-глюкани, білки, ліпіди, уронові кислоти, гідрофобіни, спорополленін та меланіни. Особлива увагаприділяється хітину, його нової функціїв антистресовому захисті клітин, а також відмінностях цього амінополісахариду грибів від хітину водоростей та Arthropoda. Обговорюється феномен апікального зростання гіфи та участь спеціальних мікровезикул у морфогенезі грибної клітини. Наводяться дані про ферменти, що беруть участь у синтезі та лізисі КС. На закінчення обговорюється функціональна значимість КС у грибів проти поверхневими структурами вищих еукаріотів.
Ключові слова: міцеліальні гриби, клітинна стінка, методи виділення, хімічний склад, апікальне зростання, фізіологічні функціїморфогенез.
Однією з ключових проблем сучасної біологіїє питання - яким чином у процесі розвитку відбувається утворення морфологічно однакових організмів, які біохімічні механізми та які клітинні структури беруть участь у цьому процесі, що вже мільярди років протікає на нашій планеті. Не менш цікавими є дані про те, як впливає стрес на морфогенез і які біополімери контролюють зовнішню формуклітин. Як показують результати досліджень останніх років, зростаюча гіфа мі-целіальних грибів є унікальною моделлю, що дозволяє зрозуміти процес формоутворення клітин і те, як зберігається в онтогенезі сталість клітинної морфології. Дуже великий внесок у дослідження цього феномена зробили дані щодо вивчення складу та біологічної функціїКС міцеліальних грибів.
КС рослин було відкрито 1665 року, а КС грибів лише на початку 18 століття. Однак ця поверхова клітинна структуратривалий час практично не вивчалася, оскільки вважалося, що КС виконує лише "каркасну" функцію, і її порівнювали зі стінами будинку, що несуть опорне навантаження. Але на початку XX століття ставлення до КС різко змінилося, її почали інтенсивно досліджувати, але переважно у рослин та бактерій. Однак інтенсивний розвиток грибівництва та біотехнологічних виробництв, у яких продуцентами БАВ були міцеліальні гриби, інтенсифікували розвиток.
1 Адреса для кореспонденції (e-mail: [email protected]).
тиє наукових дослідженьвивчення КС грибів. Дані за цією поверхневою структурою за період до 80-х років минулого століття були узагальнені в першій у світі книзі з КС грибів. У наступні роки основний масив робіт припав на кінець XX і початок XXIстоліть. Були проведені основні дослідження, присвячені систематиці, хімічному складу КС, апікальному зростанню гіфи, метаболізму хітину, антигрибним препаратам, гідрофобінам, ковалентно. пов'язаним білкам, ферментам, що беруть участь в утворенні КС, розгалуженні гіф та лізисі КС. Ці дані не були узагальнені, але саме обговорення їх у сукупності дозволяє показати, що ця поверхнева структура клітини несе поліфункціональне навантаження та виконує такі найважливіші функції, як, наприклад, захист клітин від дії несприятливих факторів, контроль морфогенезу, участь у процесах репродукції, визначення антигенних і адгезивних властивостей, контроль процесів диморфізму і утворення клітин грибів, що сприймаються, сприйняття зовнішнього сигналу і передача його на мембрану і внутрішньоклітинні месенджери. Враховуючи сказане, ми вважали доцільним зупинитися на обговоренні наступних даних: про хімічний склад КС, біологічну функцію її основних біополімерів, будову КС та міжклітинні взаємодії в гіфах грибів, феномен апікального зростання гіфи, основні ферменти, що беруть участь в утворенні та лізисі КС.
ХІМІЧНИЙ СКЛАД КЛІТИННИЙ
СТІНОКИ ГРИБІВ І БІОЛОГІЧНА ФУНКЦІЯ ЇЇ ОСНОВНИХ КОМПОНЕНТІВ
Вивчення хімічного складуКС грибів починається із отримання чистої фракції цієї структури, тобто. очищення КС від цитоплазматичного забруднення Початкова процедура - руйнування клітин грибів і відмивання клітинного вмісту - здійснюється при низькій температурі (близько 4- 5°С) для того, щоб запобігти руйнівній діяльності деградуючих ферментів. Перед руйнуванням клітин їх заморожують за температури рідкого азоту. Руйнування клітин здійснюється або у спеціальних гомогенізаторах, або на пресах методом " твердого тискуХороші результати дає подальше використання ультразвукового дезінтегратора. Відмивання клітинного вмісту здійснюється холодною водоюі повторюється 4-5 разів. Для більш ретельного видалення цитоплазматичного вмісту використовують відмивання з використанням №С1, 8 М сечовини, 1 М амонію або 0.5 М оцтової кислоти. В даний час розроблені методи виділення КС грибів у процесі їх онтогенезу, причому ці методи істотно різняться для міцелію і клітин, що покояться. Залежно від напряму досліджень, зокрема, при визначенні полісахаридного складу, з КС витягують ліпіди, використовуючи органічні розчинники, наприклад, хлороформ та метанол у співвідношенні 2: 1, іноді хороші результатидає додаткову обробку КС сірчаним ефіром. Цей розчинник добре підсушує КС і іноді може замінити ліофільні сушіння. Наступний етап- Визначення чистоти отриманої фракції КС. З цією метою використовують світлову та електронну мікроскопіюта спеціалізовані методи фарбування. Найчастіше застосовується реакція, заснована на взаємодії I3-з хітозаном. Ізольовані чисті КС із розчином Люголя забарвлюються в рожевий або фіолетовий колір, тоді як інтактні КС мають червоне забарвлення . Використовується також забарвлення на наявність ядер з DAPI (4,6-йек1ето-2-ркепуЦп-йо1). Слід особливо наголосити, що виділення чистої фракції КС є дуже відповідальною процедурою, від якої залежать наступні результати аналізу її хімічного складу. У цьому відношенні цікава робота, в якій у представника мукорових грибів виявлено в міцелії глюкан, точніше, хітозан-глюкановий комплекс, хоча всі попередні дослідженнясвідчили про відсутність у міцелії Mucorales глюкану.
В даний час компоненти КС грибів поділяють на структурні компоненти (хітин, р-(1-3)-р(1-6)-глюкани, р-(1-4)-глюкан (целюлоза), та внутрішньоструктурні (їх називають матрикс) , до яких стали відносити маннопротеїни, галакто-ман-нопротеїни, ксило-маннопротеїни, глюкуроно-
маннопротеїни та а-(1-3)-глюкан . Найменш вивчені а-глюкани, що є лінійним полімером глюкози (у Schizosaccharomyces pombe такий глюкан містить приблизно близько 260 залишків глюкози). Цей полімер складається з двох зв'язаних лінійних ланцюгів, що містять близько 120 (1-3)-а-глюкозних залишків і (1-4)-a-D-глюкозних залишків на кінцях молекули полімеру. Припускають, що цей глюкан необхідний протікання процесу морфогенезу. З КС міцелію Penicillium roqueforti виділено нерозчинний у воді глюкан, який після висушування втрачає здатність розчинятися в лугу, проте його вдалося розчинити у 10% розчині хлориду літію в диме-тилсульфоксиді. При вивченні глюкану методом метилювання були отримані два, три-О-метил-похідних глюкози, що відповідають зв'язкам 1 - 3 і 1 -»- 4 між моносахаридами у співвідношенні близько 5: 2, і сліди тетра-О-метил-похідного, що відповідає кінцевим невосстанавливающим моносахаридним залишкам. Такий результат свідчить про лінійної структуримолекул, що містять тільки 1 -»- 3 і 1 -» - 4 зв'язки між залишками глюкози. Ці дані були підтверджені аналізом спектра 13С-ЯМР полісахариду, з якого також випливає а-конфігурація всіх залишків глюкози. Такі полісахариди, особливо після хімічної модифікації (сульфатування або карбосимети-лювання), здатної надати їм розчинність у воді, можуть бути використані як біологічно активні полімери і становлять інтерес для вивчення їхньої біологічної функції в клітинній стінці грибів і рослин.
Нерозчинні у воді (1-»-3)-а-глюкани були виділені раніше з декількох видів вищих і нижчих грибів, у тому числі з Penicillium chrysoge-num, а споріднені а-глюкани з двома типами зв'язків (1-3) і (1 -»- 4), що відрізняються їх співвідношенням, знаходили в вищих рослинах(Aconi-tum kusnezoffii Reichb).
Структурний компонент– целюлоза – характерний для ооміцетних грибів, яких за сучасною систематикою відносять до псевдогрибів. Моносахаридний склад КС включає глюкозу, маннозу, ксилозу, причому, переважним цукром є глюкоза, що становить до 68%. До складу КС грибів входять також амінокислоти, ліпіди (не більше 3%) та ацетил-глюкозамін. Глюкани, наприклад, 1,3-глюкан, утворюють міцний комплекс з хітином, званий хітин-глюкановим комплексом (ХГК), який становить скелет грибної клітини. Цей комплекс присутній у КС практично у всіх грибів, за винятком зигоміцетів, що є надійною систематичною ознакою. У деяких грибів на поверхні КС виявлений слизовий матеріал, що складається з полісахаридів, представлених ß-1,3-глюканами, що містять тільки глюкозні одиниці, з'єднані ß-1,6-зв'язками з кожною третьою одиницею глюкози