Клітини рослин оточені щільною полісахаридною оболонкою, вистеленою зсередини плазмалемою.
Освіта клітинної стінкивідбувається в метафазі та телофазі клітинного поділу. У екваторіальній зонірозподілу виникає серединна платівка, що складається з пектату кальцію, яка, наростаючи від центру до периферії, відокремлює одну новостворену клітину від іншої. Середня платівка з того й іншого боку покривається первинною клітинною стінкою. Зростання завтовшки відбувається за рахунок накладання нових шарів з боку вмісту кожної клітини. Зростання клітини в довжину починається з розпушування матриксу. У цьому процесі важливу рольграють фітогормон. У порожнини, що утворилися, надходять нові порції матеріалу, з якого будується клітинна стінка. Синтез та транспорт цих речовин здійснюються головним чином вакуолями апарату Гольджі.
Клітинну стінку клітин, що діляться і ростуть розтягуванням, називають первинної.Після припинення зростання клітини на первинну клітинну стінку зсередини відкладаються нові шари і виникає міцна вториннаКлітинна стінка.
До складу клітинної стінки входять структурні компоненти(целюлоза у рослин, хітин у грибів), компоненти матриксу стінки (геміцелюлози, пектин, білки), інкрустуючі компоненти (лігнін, суберин) та речовини, що відкладаються на поверхні стінки (кутин та воску). Клітинні стінки можуть містити також силікати та карбонати кальцію.
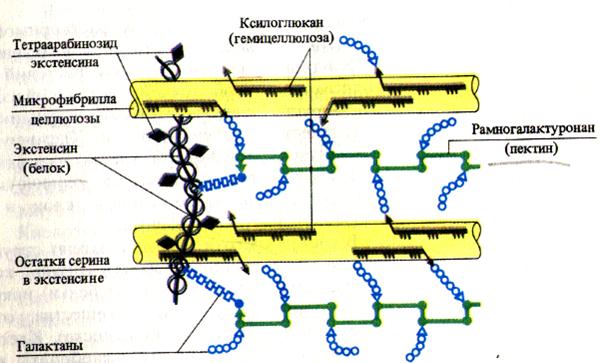
Целюлоза(полімер b-D-глюкози), геміцелюлози(полімери гексоз та пентоз) та пектинові речовини(Похідні уронових кислот) є вуглеводними компонентами клітинних стінок. Целюлоза та пектинові речовини адсорбують воду, забезпечуючи обводненість клітинної стінки. Пектинові речовини, що містять багато карбоксильних груп, зв'язують іони двовалентних металів, які здатні обмінюватися на інші катіони (Н+, К+ тощо). Це зумовлює катіонообмінну здатність клітинних стін рослин. Крім вуглеводних компонентів до складу матриксу клітинної стінки входить також структурний білок, званий екстенсин.Це глікопротеїн, що містить понад 20% L-оксипроліну від суми амінокислот. За цією ознакою білок клітинних стінок рослин подібний до міжклітинного білка тварин - колагеном.
Целюлоза: А – структура молекули целюлози; Б – асоціації молекули целюлози: 1 – міцела, 2 – мікрофібрила, 3 – макрофібрила
Основним інкрустуючим речовиною клітинної стінки є лігнін.Інтенсивна лігніфікація клітинних стін починається після припинення росту клітини. Лігнін є полімером з нерозгалуженою молекулою, що складається з ароматичних спиртів (п-кумарового, коніферилового, синапового). Руйнування та конденсація лігніну в ґрунті - один із факторів утворення гумусу. Інтенсивна лігніфікація (просочування шарів целюлози лігніном) клітинних оболонок починається після припинення росту клітини. Лігнін може відкладатися окремими ділянками - у вигляді кілець, спіралей або сітки, як це спостерігається в оболонках клітин провідної тканини - ксилеми, або суцільним шаром, за винятком тих місць, де здійснюються контакти між сусідніми клітинами у вигляді плазмодесм. Лігнін скріплює целюлозні волокна і діє як дуже твердий і жорсткий каркас, що підсилює міцність клітинних стінок на розтяг і стиск. Він же забезпечує клітинам додатковий захист від фізичних та хімічних впливівзнижує водопроникність. Вміст лігніну в оболонці сягає 30%. Інкрустація їм клітинних оболонок призводить до їх озеленення,яке часто спричиняє відмирання живого вмісту клітини. Лігнін у поєднанні з целюлозою надає особливі властивостідеревини, які роблять її незамінним будівельним матеріалом.
У регуляції водного та теплового режимурослин беруть участь тканини, стінки клітин яких просочені суберином.Відкладення суберину робить стінки важко проникними для води та розчинів (наприклад, в ендодермі, перидермі). Суберінвідкладається на оболонку зсередини і робить її практично непроникною для води та розчинів. В результаті протопласт клітини відмирає та клітина заповнюється повітрям. Такий процес називається випробуванням.Спостерігається пробковування оболонки клітин у покривних тканинахбагаторічних деревних рослин – перидермі, кірці, а також в ендодермі кореня. Суберин - основна речовина коркових утворень, якому ці утворення переважно зобов'язані своїми властивостями: непроникністю для води, для газів і малою теплопровідністю.
Поверхня епідермальних клітин рослин захищена гідрофобними речовинами. кутиномі восками:Попередники цих сполук секретуються з цитоплазми на поверхню, де відбувається їх полімеризація. Шар кутину зазвичай пронизаний полісахаридними компонентами стінки (целюлозою, пектином) і утворює кутикулу. Кутикула бере участь у регуляції водного режиму тканин та захищає клітини від пошкоджень та проникнення інфекції.
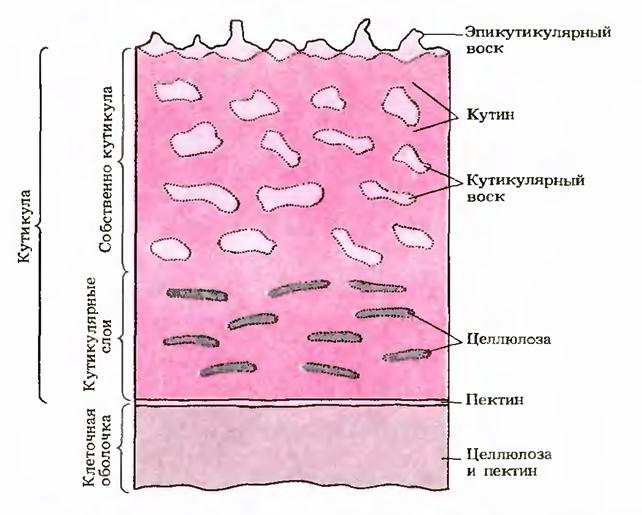
В оболонках епідермальних клітин деяких рослин (злаків, осок та ін.) накопичується велика кількістьмінеральних речовин (Мінералізація),в першу чергу карбонату кальцію та кремнезему. При мінералізації листя і стебла рослин стають жорсткими, твердими і меншою мірою поїдаються тваринами.
У первинних клітинних стінках частку целюлози припадає до 30% сухої маси стінки. Кількість геміцелюлоз та пектинових речовин змінюється залежно від об'єкта. Разом з білками пектинові речовини можуть становити близько 30% сухої маси клітини, причому кількість білка досягає 5-10%. Близько 40% посідає частку геміцелюлоз.
1) Клітинна стінка - структурна освіта. Функція: надає міцності та форми, захищає протопласт від зовнішніх умов, бере участь у проведенні та поглинанні речовин.
Основа клітинної оболонки (склад) - високополімерні вуглеводи (целюлоза, тобто клітковина - не перетравлюється, вказує на низьку продуктивність), молекули целюлози зібрані в складні пучки (міцелії), міцелії об'єднуються в фібрили, їх проміжки заповнені геміцелюкою. з'єднання) та пектином (корисні, набухають у воді, є джерелом енергії).
Розрізняють первинну та вторинну клітинні оболонки. Меристематичні та молоді зростаючі клітини мають первинну клітиннуоболонку, тонку, багату на пектин і геміцелюлозу; фібрили целюлози в матриксі первинної клітинної оболонки розташовані невпорядковано.
Вторинна клітиннаоболонка утворюється зазвичай після досягнення клітиною остаточного розміру і накладається шарами на первинну з боку протопласту. У вторинній клітинній оболонці переважає целюлоза, її фібрили розташовуються впорядкування, паралельно, але напрямок їх у кожному шарі інше, що підвищує міцність клітинної оболонки. У вторинній клітинній оболонці є отвори (пори), де клітини поділяють лише первинна оболонка та плазмодесми (цитоплазматичні містки, що з'єднують сусідні клітини рослин).
Видозміни клітинної стінки:
- Здеревніння клітинної оболонки відбувається в результаті відкладення лігніну (невуглеводний компонент у фібрилах), клітини втрачає еластичність, але можуть пропускати воду. Ці клітини найчастіше мертві, ніж живі. Стінки деяких клітин можуть містити: віск, кутину, суберин. Функції: надає клітині форми; відокремлює одну клітину від іншої, є скелетом для кожної клітини та надає міцності всій рослині, виконує захисну функцію.
- Опробування викликається особливою жироподібною речовиною - суберином. Такі оболонки стають непроникними для води і газів, також вони не пропускають тепло, вміст клітин з пробковілими оболонками відмирає.
- Кутинізація полягає у виділенні жироподібної речовини кутину. Зазвичай кутинізуються зовнішні стінки шкірки листя і "трав'янистих стебел. Це робить їх менш проникними для води, зменшує випаровування води у рослин, охороняє від перегріву та ультрафіолету. Кутин утворює на поверхні органу плівку, яка називається кутикулою.
- Мінералізація клітинних оболонок - це відкладення: кремнезему та солей кальцію. Найбільш сильно інкрустуються оболонки клітин шкірки листя та стебел злаків, осок, хвощів. Листям злаків та осок можна поранити руки.
- Ослизнення оболонок - перетворення целюлози та пектинових речовин у слизу та камеді. Ослизнення добре спостерігається на насінні льону, що знаходилося у воді. Утворення слизів сприяє кращому поглинанню води насінням та прикріпленню їх до ґрунту.
2) Розмноження:здатність окремо взятої особини дати початок цілій серії собі подібних.
Ділять на: статеве та безстатеве (власне безстатеве та вегетативне)
Вегетативне: нові особини розвиваються з окремих вегетативних органів або їх взаємодій Здійснюється завдяки регініраціям (св-во відновлювати із частини тіла організм). Біо значення: новий організм подібний до материнського.
Способи вегетативного розмноження:
- розмноження живцями (частиною рослини, яка не заражена, садять у субстрат, спородину),
- розмноження методом щеплення (шляхом зрощування частин кількох рослин, що застосовується в садівництві),
- розмноження бульбами (м'ясисті бульби з пити в-вами садять у землю, живородна гречка),
- розмноження нащадками (утворюють пагони на коренях, осика),
- розмноження цибулинками (восени відсаджують від самої рослини в землю)
- розмноження вусами (повзучі пагони, укорінюються придат корінням, кістяника, суниця)
- розмноження кореневищами (підземна втеча, запас піт-в, конвалія, фіалка, пирій)
Використання вегетативного розмноження людиною. Решта див у 40.
З давніх пірлюдина, культивуючи рослини, стала використовувати вегетативне розмноження. Наприклад, вирощування картоплі, суниці, бананау всіх країнах світу здійснюється лише вегетативним шляхом – бульбами, вусами та кореневищами.
Використання вегетативного відтворення рослин у сільськогосподарській практиці отримало назву штучного вегетативного розмноження.
Основні прийоми штучного вегетативного розмноження зводяться до повторення тих, що відбуваються у рослин у природних умовах.
Люди часто використовують розмноження живцями - частинами зеленого або здерев'янілого втечі (виноград, смородина, агрус, троянда, гвоздика, фікус), бульбами (картопля, жоржина, батат, топінамбур), листям (сенполія, глоксинія, бегонія), цибулинами (цибуля, часник, тюльпан, нарцис), розподілом куща (смородина, піретрум)та відведеннями (аґрус, жимолість, клематис), вусами (полуниця), кореневищами (цукрова тростина, іриси, флокси), кореневими нащадками (слива, малина, вишня, бузок).
3) Гарбузові. Форма трави. Корінь стрижневий. Стебло: лазить, стеле, кучерявий Лист: простий, черешковий, без прилистків.
Формула: роздільностатевий
1) правильний жіночий Ca (5) Co (5) A 0 G (3) оцвітина під зав'яззю
2) правильний чоловічий Ca (5) З (5) А 2+2+1 G 0
Суцвіття поодиноке. Плід: гарбуза
Представники: огірок, диня, гарбуз, кавун., кабачок
Значення: харчове, кормове
Клітинна оболонказдатна до потовщення та видозміни. В результаті цього утворюється ве вторинна структура. Потовщення оболонки відбувається шляхом накладання нових шарів на нервпчпуго оболонку. Зважаючи на те, що накладення йде вже іа тверду оболонку, Фібрили целюлози в кожному шарі лежать паралельно, а в сусідніх шарах - під кутом один до одного. Цим досягається значна міцність та твердість вторинної оболонки. У міру того, як число шарів фібрил целюлози стає більше і товщина стінки збільшується, вона втрачає еластичність і здатність до зростання. У вторинній клітинній стінці вміст целюлози значно зростає, у деяких випадках до 60% і більше. У міру подальшого старіння клітин матрикс оболонки може заповнюватися різними речовинами- лігніном, суберином (зревніння або пробковування оболонки). Лігнін утворюється з геміцелюлози н пектинових речовин.
Клітинна оболонка деревного волокна має кілька шарів: первинний, який називається зовнішньою оболонкою волокна, та вторинний (стінка, що складається, у свою чергу, із трьох шарів: зовнішнього, середнього та внутрішнього). Між первинними стінками клітин знаходиться шар міжклітинного речовини, з якого волокна з'єднуються друг з одним. Вторинна стінка відносно товста і є головною масою об'єму клітини.
У вторинних шарах клітинних стін деревини сосни накопичувалися в великих кількостяхманнани (22%) та уроновий ангідрид (25%).
[ ...]
Фаза потовщення клітинної стінки. Як відбувається потовщення. У період розростання протопласт оточений лише первинною стінкою. Коли ж деревна клітка досягає свого найбільшого розмірупо поверхні або незабаром після цього стінка клітини потовщується. Це викликано нашаруванням вторинної стінки на первинну, причому цей новий шарвиникає в результаті подальшої діяльностіпротопласту всередині порожнини клітини. Природно, клітини, у яких протопласт зник, що неспроможні продовжувати потовщувати свої стінки. Утворення вторинної стінки є ознакою незворотної зміни в клітині, подальше розростання якої вже виключено, але не обов'язково виключається подальший поділ за умови, що дочірні клітини, що отримуються таким чином, займають такий же обсяг, як і початкова клітина.
М.1іп - килим, покривало). Він складається з таблитчастих тонкостінних клітин із густою цитоплазмою. Зазвичай він однорядний, але іноді буває дворядним або багаторядним. Клітини ноту ма спочатку одноядерні, пізніше вони часто стають двоядерними або навіть багатоядерними. Тапетум є фізіологічно надзвичайно активною тканиною: його клітини містять ферменти, гормони, і поживний матеріал, що використовується в процесі мікроспорогепезу. Є деякі підстави вважати секреторний тип в еволюційному відношенні первинним, а амебоїдний - вторинним.
Необхідно, однак, зазначити, що ці дані слід розглядати як наближені, оскільки вихідні препарати не були ретельно очищені.
Важко визначити розташування в клітинній стінці поліуронідних геміцелюлоз, тому що реагенти, що використовуються для їх виявлення, впливають і на лігнін. Деякі дослідники припускають, що цементуючим речовиною між фібрилами та різними шарами клітинної стінки є геміцелюлози. Коен вважає навіть, що лігнін вторинної стінки має однакову природуз геміцелюлозами. Підставою для такого припущення є, мабуть, той факт, що деякі вуглеводи при обробці сильними кислотамиможуть давати нерозчинні залишки певного малюнка. Слід підкреслити, проте, що ділянки, як ретельно оброблені реагентами, що розчиняють геміцелюлози, так і не оброблені ними, дають при дії 72% сірчаної кислоти залишки дуже схожої структури.
Для з'ясування складу окремих шарів клітинних стінок була зроблена спроба кількісного визначенняксилоуронідів у різних шарах трахеїд та лібриформа. Вимірювання проводилися на волокнах із червоної японської сосни, європейської ялиці, бука та берези. Для цього волокна обережно нітрували в оцтовому середовищі ангідриду і чотирихлористого вуглецю. Потім зовнішній нітрований шар видаляли розчиненням в ацетоні, після чого контролювали вміст пентозанів у залишку фурфуролу. Було встановлено, що пентозани в волокнах по шарах розділені нерівномірно. Найбільша кількістьпентозанов знайдено у зовнішніх шарах волокон і концентрація їх падає від периферії до центру. Так, зовнішні шари волокон хвойної деревини містять 50-80% пентозанів, а у листяних майже 100%. У вторинних шарах клітинних стінок у хвойних вміст пентозанів виявилося трохи більше 2-4%, а й у листяних 8-10%. Таким чином, хімічний методпідтвердив результати, отримані раніше методом сорбції ультрафіолетового світла.
Розрізняють первинний лігнін, що знаходиться в здеревнених клітинних стінках (природний лігнін) і вторинний - ізольований лігнін. Останній є значною мірою речовиною, зміненою в процесі ізолювання та забрудненою домішками сторонніх речовин. Зміна лігніну виявляється у відщепленні метоксильних груп, внутрішньомолекулярної конденсації та інших ознаках.
Багато відмінностей між типами тканин обумовлені будовою клітинної стінки, особливо вторинної. Як ми вже говорили, утворення первинної клітинної стінки відбувається в процесі розтягування клітини, і, отже, вона повинна мати властивість розтяжності, тоді як вторинна стінка формується вже після того, як подовження припинилося.
Престон
Одночасно з цими внутрішніми змінамизовнішня тверда стінка ооспори розщеплюється на її вершині на п'ять зубців, даючи вихід проростку, що виникає з центральної клітини (рис. 269, 3). Перший поділ центральної клітини відбувається поперечною перегородкою, перпендикулярною до її довгої осі, і призводить до утворення двох функціонально різних клітин. З однієї, більшої клітини надалі утворюється стебловий пагін, який на початковій стадіїРозвиток називають передлітком, з іншої, меншої клітини - перший ризоїд. Обидва вони ростуть шляхом поперечних клітинних поділів. Передліток росте вгору і досить швидко зеленіє, заповнюючись хлоропластами, перший ризоїд прямує вниз і залишається безбарвним (рис. 269, 4). Після низки клітинних поділів, які повідомляють їм будова однорядних ниток, відбувається їх диференціювання на вузли і міжвузля, і подальший їх верхівковий ріст протікає так, як було описано вище для стебла. З вузлів передлітка виникають вторинні передлітки, мутовки листя і бічні гілки стебла, з вузлів першого різозу - вторинні різоїди та їх каламутні волоски. Таким шляхом і формується таллом, що складається з кількох стеблових пагонів у верхній частині та кількох складних ризоїдів у нижній частині (рис. 2G9, 5).
Надмолекулярна структура. На рис 6.10 наведено модель структури клітинної стінки. Вона включає 2 основні шари: первинну стінку Р і вторинну Остання підрозділяється на 3 шари: 5], 5 , шар М, серединна пластинка, є міжклітинною речовиною, що з'єднує клітини між собою.
У наступних розділах (єм. частина II) буде вичерпно розглянута хімія клітинних стінок, відносні кількостілігніну в них та інші родинні теми. Однак закінчуючи розгляд четвертої та кінцевої фази онтогенезу деревної клітини, слід згадати деякі явища, які тим чи іншим шляхом пов'язані з лигннфикацией, як її пошшают ботаніки. Подібно до утворення і розростання клітин, а також потовщення клітинних стінок, лігніфікація може відбуватися лише за життя клітинного протопласта, так як відмерлі клітини не можуть лігніфікувати свої стінки. Процес лігніфікації може бути закінчений у шарі міжклітинної речовини і в первинній стінці, але може продовжуватися у вторинній стінці, навіть якщо цей названий останнім шар ще доволі зростає в товщину. У деревині дерев лігніфікація часто дуже швидко закінчується в шарі, що примикає до внутрішньої сторони камбію, зазвичай майже одночасно з тим, коли нові клітини досягли свого найбільшого розміру, а вторинні стінки своєї кінцевої товщини. Це пояснює, чому заболонь при однаковому вмісті вологи так само або майже так само міцна, як ядерна деревина.
Детальне дослідження розподілу лігніну і полісахаридів в дерев'яних ялинках і березах, що здервіли, клітинних стінках, вимірюванням інтенсивності абсорбції тонкого пучка. ультрафіолетових променівпри проходженні їх через прозорий зріз підтвердило переважне розташування лігніну в серединній платівці та первинній стінці, а також частково у зовнішніх шарах вторинної стінки. У серединній платівці ялинової деревини вміст лігніну досягає 73%, а у вторинній стінці – не більше 16%. Звідси випливає, що полісахариди зосереджені переважно у вторинному шарі. Була зроблена спроба виміряти цим методом взаємне розташуванняцелюлози та геміцелюлоз. Для цього полісахариди спочатку були перетворені на пофарбовані сполуки, що абсорбують світло.
У більшості клітин ясно розрізняються зони, що чергуються, більшого або меншого відкладення лігніну, які створюють видимість концентричних кілець. При протилежному процесі, коли клітинна стінка обробляється делигнифицирующими. реагентами, малюнок целюлози залишається тим самим. Це свідчить про те, що існують, мабуть, дві взаємопроникні системи, що складаються з целюлози та інших полісахаридів, а інша з лігніну. Бейлі та Керр показали, що розміри частинок сягають 0,1 і менше. Проміжки чи смуги пояснюють існування щодо великих «фібрил», помічених деякими дослідниками. Крім переважаючих концентричних малюнків, у волокнах деяких видів деревини проявляється розташування радіальних ліній чи комбінація обох типів. Клітини стиснутої деревини часто мають жорсткі, майже тверді смуги лігніну поруч із порожниною клітини та радіально-розташовані пластинки його, відокремлені зонами полісахаридної речовини, у середній частині стінки клітини.
До складу лишайників входять багато елементів та речовини. Усі їх можна поділити на дві великі групи- первинні та вторинні. До первинних належать ті речовини, які безпосередньо беруть участь у клітинному обміні речовин; їх побудовано тіло лишайників. До вторинних відносяться кінцеві продукти обміну речовин, які зазвичай розташовані на стінках гіф. Багато хто з цих вторинних лишайникових речовин (у більш старій літературі їх називали лишайниковими кислотами) специфічні для лишайників і не зустрічаються в організмах інших систематичних груп.
Риттер, Людтке та ін. повідомили, що при обробці деревних волокон різними реагентами, що викликають набухання, вторинна стінка (а також, ймовірно, і первинна) розпадається на ниткоподібні фрагменти або фібрили. Ріттер розділив ці фібрили на веретеноподібні тіла, які у свою чергу, на сферичні одиниці . Значення таких щодо великих структурних одиниць(Довжина веретеноподібних тіл приблизно 4 [х] неясно, зважаючи на описану вище тонкопористої структури вторинної стінки. Ні в залишках лігніну після розчинення целюлози, ні в залишках целюлози після розчинення лігніну не виявляється помітних проміжків, що вказують на межі названих одиниць клітинних стінок. Крім того, нещодавно проведеними дослідженнями за допомогою електронного мікроскопа у структурі клітинних стінок не було встановлено присутності подібних порівняно великих одиниць.
При оцінці дії різних дереворуйнівних грибів на рослинну тканинунеобхідно враховувати, що окремі гіфи їх. рухаються в товщі клітинних стін вибірково. Так, гриби білої гнилі віддають перевагу серединній платівці і первинній оболонці, де зосереджений головним чином лігнін. Гриби червоної або бурої гнилі, навпаки, вважають за краще проходити по вторинній оболонці, найбільш багатій на вуглеводи. Відповідно відрізняється і фарбування пошкодженої ними деревини. Більш детально ці питання будуть розглянуті надалі.
Дослідження трахеїд та лібриформа за допомогою поляризаційного та електронного мікроскопа, а також рентгенографії дозволили встановити існування у клітинних стінках п'яти концентричних шарів: зовнішньої, або первинної, стінки та вторинної стінки. Вторинна стінка у свою чергу поділяється на три шари, які зазвичай позначаються 81, вг і Бз. Крім того, між первинними стінками сусідніх клітин розташовується серединна пластинка, що склеює їх (рис. 35).
Підвищення виходів під час використання водяної пари пояснюється тим, що прискорюється винесення цінних продуктів із реакційного простору та затримується розвиток реакцій вторинного розпаду. Крім того, при дотику водяної пари з капілярною системою деревини на поверхневих шарах її можлива конденсація пари, що створює умови для термічного розкладанняу кислій водному середовищі. При цьому реакції розкладання відбуваються в першу чергу в шарах клітинної стінки, які розташовані внутрішніх сторінклітинних порожнин і складаються переважно з нетермостійких геміцелюлоз, що легко відщеплюють ацетильні групи та частину пов'язаних з ними метоксилів, утворюючи відповідно оцтову кислотуі метиловий спирт.
Навряд чи правильно називати клітинами сегменти, що складають нитки сфероплеї, і не тільки тому, що вони мають безліч ядер і хлоропластів (і, отже, є явно вторинними утвореннями), а й тому, що поперечні перегородки, що їх відокремлюють, не схожі на клітинні стінки інших багатоклітинних. зелених водоростей. Вони сильно варіюють за формою, а також за способом та місцем освіти (рис. 226, 4-6). Часто поперечні перегородки мають вигляд кільцевих внутрішніх потовщень на стінках клітини, які не стуляються в центрі, так що залишається отвір, через який проходить цитоплазматичний тяж (рис. 226, 4). В інших випадках замість перегородок утворюються спеціальні пробки. І, нарешті, в будь-якому місці нитки можуть виникати групи тяжів, що радіально сходяться, нагадують скелетні тяжі каулерпи і грають механічну роль.
Зовні від плазматичної мембраниїх клітин немає додаткової щільної клітинної стінки чи вона складається з хітину, рідко з целюлози. Запасні вуглеводи зазвичай у формі глікогену (тварини крохмалю).
Маркс-Фігіпі та Пепцел вивчали зміну СП бавовняної целюлози на різних стадіяхдозрівання бавовни. Вони показали, що в'язкість розчинів бавовняної целюлози знижується за кілька годин після відкриття коробочки. Целюлоза вторинної клітинної стінки в волокнах коробок бавовни, що не розкрилися. невеликої зрілості(Вихід целюлози-18%) має єдиний максимум на кривій розподілу при СП 14 000. Близько 10% матеріалу має нижчу молекулярну вагу (СП 1500-2500), ця целюлоза міститься в первинній клітинній стінці.
Положення місць утворення мікрофібрил по відношенню до поверхні мембрани цитоплазми може бути різним. Так, у бактерій цей процес протікає в середовищі, значно віддаленому від поверхні клітини і, отже, від мембрани. Очевидно, аналогічним чином синтез протікає і потовщених первинних стінках клітин епідермісу колеоптилей вівса, оскільки синтез целюлози у разі здійснюється рівномірно але товщині клітинної стінки . В оболонках асцидій відкладення целюлози відбувається, мабуть, також у місцях, віддалених від поверхні секреторних клітин, хоча достатньо переконливих доказів цього припущення немає. Навпаки, мікрофібрили вторинних стінок клітин рослин, можливо, утворюються на внутрішній поверхні стінки, безпосередньої близькостівід мембрани цитоплазми. Оскільки целюлози у вторинних стінках значно більші, ніж у первинних, можна зробити висновок, що більшість целюлозних мікрофібрил утворюється поблизу мембрани цитоплазми. Однак це не є обов'язковим.
Одним із методів, заснованих на цьому принципі, є метод визначення реакційної здатностіцелюлози по картині набухання ксантогенатів в ізо-пропіловому спирті. Процес набухання при взаємодії волокна з розчинником схематично можна представити в такий спосіб: рідина проникає всередину волокна, внаслідок чого об'єм волокна збільшується. Потім відбувається розрив слабкого зовнішнього еластичного шару вторинної клітинної стінки волокна і в місцях розриву утворюються здуття («буси»). Залишки цього шару утворюють на набряклому волокні перетяжки та манжети. Потім зовнішній шар відокремлюється і волокно рівномірно набухає, на ньому утворюються поперечні смуги і волокно ділиться на пакети дисків окремі диски, які надалі розчиняються.
Залежність міцності деревини від вмісту вологи. Так як міцність і жорсткість деревини частково визначаються силами зчеплення, що зв'язують молекули, будь-який агент, що зменшує ці сили, змінює її міцність в цілому. Одним з таких агентів є вода, тому міцність деревини збільшується з зменшенням вмісту вологи не тільки в результаті підвищеної щільності, що походить від усушки, але також через присутність вторинних валентних сил зчеплення1. Так як наявність води в кількості, що перевищує точку насичення волокна, не змінює характеру клітинної стінки, то втрата або придбання капілярної (вільної) води практично не впливає на показники міцності деревини.
Структури, що містять багато лігніну, забарвлюються у темно-коричневий колір до чорного, тоді як слабко лігніфіковані зони забарвлюються у світло-жовтий колір до бурштинового. Результати цієї кольорової реакції повністю підтверджують попередні роботи з дослідження хімії клітинної стінки. Вторинні стінки волокнистих елементів у деревини листяних порід, що ростуть у помірному кліматі, світліші, отже, вони менш лігніфіковані, ніж вторинні стінки хвойних порід. Стінки судин у листяних порід пофарбовані в більш темний колір, Чим навколишні волокнисті елементи, отже, вони містять більше лігніну; мембрани пір також сильно лігніфіковані.
Ця операція здійснювалася на зрізах, що одеревіли, попередньо звільнених від лігніну за допомогою хлориту натрію в оцтовокислому середовищі. Потім зрізи були оброблені п-фенілаз; бензоїлхлоридом з метою етерифікації полісахаридів. Яскраво пофарбовані в оранжево-червоний колірзрізи після набухання у піридині фотометрировались. Піддаючи такій обробці зрізи, що складаються з холоцелюлози, до та після видалення геміцелюлоз, вдалося встановити, що основна маса геміцелюлоз у деревині ялини та берези зосереджена у зовнішніх шарах вторинної стінки. Так, при екстракції зрізу ялинової холоцелюлози 16%-ньш їдким натромбуло встановлено, що із зовнішніх шарів клітини витягується до 60-80%, із середини клітинної стінки близько 50% та з шару Бз лише 16% розчинних у лугу геміцелюлоз від загальної кількості полісахаридів. Аналогічна картина спостерігалася й у поперечних зрізів либриформа з деревини березы.[ ...]
Досліди Ріттера, а пізніше Бейлі та ін. показали, що незалежно від можливої присутності пектинових поліуронідів у серединній платівці, вона складається головним чином з лігніну, як його розуміють хіміки (нерозчинний у холодній 72%-ній сірчаній кислоті, розчинний після хлорування та обробки слабкими підставамичи основними солями). Крім того, Ріттер довів, що більша частиналігніну знаходиться саме в цьому шарі. Це твердження суперечило переважав у той час думці про присутність більшої частини лігніну в інших шарах, особливо у вторинній стінці. Пізніше було доведено, що в таких випадках вторинна стінка, що здається широкою і об'ємною, насправді подібна до павутини, яка після висихання зіщулюється і перетворюється на розрізнені шматочки. Якщо первинні стінки включені в складну серединну пластинку, то цілком імовірно, що тут знаходиться і більша частина лігніну.
Кальцієві канали виявлено й у мембранах рослинних клітин. Показано регулювання входу 45Са2+ мікросоми, виділені з колеоп-тилів кукурудзи та гіпокотилів гарбуза, світлом, ПУК та залежність цієї реакції від кальмодуліну. Для функціонування потенціалзалежних Са2+-каналів (харова водорість Ыие11ор,ш) необхідна наявність М§2+. Стан цих потенціалзалежних каналів контролюється системою ферментів, що рейдують рівень цАМФ у клітині. Були також отримані дані, що свідчать про прямій діїекзогенного цАМФ на поглинання 45Са2+ у клітинах СМатуєотопт гетскагсШ (мутант без клітинної стінки). Дані наведені на рис. 4.1 свідчать про регуляторну дію цАМФ на поглинання Са2+ клітинами. Це свідчить про можливість взаєморегуляції двох систем вторинних посередників - цАМФ і Са2+. У дослідах із тваринними клітинами посилення поглинання Са2+ під дією цАМФ пояснюється фосфорилюванням білків потенціалзалежних Са2+-каналів і внаслідок цього збільшенням перебування їх у відкритому стані.
Вивченню дії ультразвуку на целюлозні волокна присвячено багато досліджень. Деякі дослідники зіставляли чи поєднували вплив ультразвуку з різними механічними впливами. Так, Яйме, Кронерт і Нейхауз вивчали дію ультразвуку на целюлозні волокна порівняно з високочастотними механічними коливаннями і показали, що ультразвук із частотою 20-3000 кгц розпушує структуру волокна, збільшує ступінь його набухання та зневоднення. Механічна міцність паперу, виготовленого з таких целюлоз, підвищується, особливо міцність до роздирання. Аналогічно діють і високочастотні механічні коливання. Івасакі, Ліндберг та Мейєр вважають, що загальна картина змін структури волокна під дією ультразвуку у водному середовищі подібна до змін структури волокон при механічному розмелі. При цьому відбуваються глибокі зміни морфологічної структури волокон, що призводять до зсувів у вторинній клітинній стінці, відриву великих шматків від первинної стінки, потім набухання вторинної стінки та її дефібрилювання. У роботі Сафонової і Клен-ковой щодо мікрофотографій волокон, підданих ультразвуковому впливу у питній воді, показано, що є й інші, глибші порушення у структурі волокна, що стає пронизаним цілою мережею численних поперечних каналів. Зазначається, що волокна ранньої деревини та волокна, що не піддавалися висушуванню, більш сприйнятливі до дії ультразвуку.
Рослинні клітини, подібно до клітин прокаріотів і грибів, укладені в порівняно жорстку клітинну стінку. Матеріал для побудови цієї стінки клітини секретує сама укладена в ній жива клітка(Протопласт). За своїм хімічним складом клітинні стінки рослин відрізняються від клітинних стінок прокаріотів та грибів (табл. 2.1), але цим структурам властиві деякі загальні функції, А саме функції опори та захисту; крім того, і ті та інші обмежують рухливість клітин. Клітинна стінка, що відкладається під час поділу клітин рослини, називається первинною клітинною стінкою. Пізніше внаслідок потовщення вона може перетворитися на вторинну клітинну стінку. У цьому розділі ми опишемо процес утворення первинної клітинної стінки. На рис. 7.21 відтворено електронну мікрофотографія, на якій можна бачити одну з ранніх стадійцього процесу.
Будова клітинної стінки
Первинна клітинна стінка складається з целюлозних мікрофібрил, занурених у матрикс, до складу якого входять складні полісахариди. Целюлоза теж є полісахаридом (її хімічна будоваописано у розд. 5.2.3). Особливо важливе значеннядля тієї ролі, яку целюлоза виконує у клітинних стінках, мають її волокнисту будову та високу міцність на розрив, порівнянну з міцністю сталі. Окремі молекули целюлози – це довгі полісахаридні ланцюги. Безліч таких молекул, пошитих один з одним поперечними. водневими зв'язками, зібрані в міцні пучки, звані мікрофібрилами. Занурені в матрикс мікрофібрили утворюють каркас клітинної стінки. Матрікс клітинної стінки складається з полісахаридів, які для зручності опису ділять зазвичай на пектиниі геміцелюлозив залежності від їх розчинності в різних розчинниках, які використовуються для екстракції. Пектини, або пектинові речовинипри екстракції зазвичай виділяються першими, оскільки їх розчинність вище. Це - змішана група кислих полісахаридів (побудованих з моносахаридів арабінози та галактози, галактуронової кислоти, що належить до класу цукрових кислот, та метанолу). Довгі молекули пектинових речовин можуть бути лінійними чи розгалуженими. Середня платівка, що скріплює стінки сусідніх клітин, складається з клейких студнеподібних пектатів магнію та кальцію. У клітинних стінках деяких дозріваючих плодів нерозчинні пектинові речовини перетворюються знову на розчинні пектини. При додаванні цукру ці останні утворюють гелі; тому їх використовують як желіруючі речовини.
Геміцелюлози- це змішана група полісахаридів, розчинних у лугах (до них відносяться полімери ксилози, галактози, маннози, глюкози та глюкоманнози). У геміцелюлоз, як і у целюлози, молекули мають форму ланцюга, проте їх ланцюги коротші, менш упорядковані та сильніше розгалужені.
Клітинні стінки гідратовані: 60-70% їхньої маси зазвичай становить вода. По вільному простору клітинної стінки вода переміщається безперешкодно. Присутність води впливає на хімічні та фізичні властивостіполісахаридів клітинної стінки.
Матеріали з підвищеною механічною міцністю, подібні до матеріалу клітинної стінки, тобто складаються більш ніж з одного компонента, називаються композиційними матеріаламиабо композитами; їхня міцність зазвичай вища, ніж у кожного з компонентів окремо. Системи з волокон і матриці (в техніці основу композиційного матеріалу називають не матриксом, а матрицею. - Прим. перев.) знаходять широке застосування в техніці, так що на вивчення їх властивостей як у техніці, так і в біології витрачається багато зусиль. Матриця, що працює на стискування, передає напругу волокнам, що працюють на розтяг. Вона ж забезпечує абразивну стійкість і, мабуть, стійкість до несприятливих хімічних впливів, можливих у тих чи інших умовах. У будівельній справі здавна застосовується залізобетон, тобто поєднання бетону із сталевою арматурою. Пізніше з'явився легший композиційний матеріал, у якому роль матриці грає пластик, а роль арматури – скляне чи вуглецеве волокно. Деревина є композиційним матеріалом; своєю міцністю вона завдячує клітинним стінкам. Приклад жорстких композиційних матеріалів біологічного походженняможуть також служити кістка, хрящ і покриває екзоскелет членистоногих кутикула. Існують і гнучкі композиційні матеріалинаприклад сполучна тканина.
У деяких клітин, наприклад у клітин мезофілу листа, протягом усього життя є тільки первинна клітинна стінка. Однак у більшості клітин на внутрішню поверхнюпервинної клітинної стінки (назовні від плазматичної мембрани) відкладаються додаткові шари целюлози, тобто виникає вторинна клітинна стінка. Зазвичай це відбувається після того, як клітина досягне свого максимального розміру, і лише деякі клітини, наприклад клітини коленхіми, продовжують зростання під час цієї фази. Вторинне потовщення клітинних стінок рослини не слід плутати з вторинним потовщенням (вторинним зростанням) самої рослини, тобто зі збільшенням товщини стовбура в результаті додавання нових клітин.
У будь-якому шарі вторинного потовщення целюлозні волокна розташовуються під тим самим кутом, але в різних шарах цей кут різний, чим і забезпечується ще більша міцність структури. Таке розташування волокон целюлозних показано на рис. 7.27.
Деякі клітини, такі, як трахеальні елементи ксилеми і клітини склеренхіми, зазнають інтенсивної лігніфікацію(Деревесень); при цьому всі шари целюлози (первинний і три вторинних) просочуються лігніном - складною полімерною речовиною, що не належить до полісахаридів. У клітинах протоксилеми відкладення лігніну мають кільцеву, спіральну або сітчасту форму, як видно на рис. 8.11. В інших випадках лігніфікація буває суцільною, якщо не рахувати так званих порових полів, тобто тих ділянок у первинній клітинній стінці, через які здійснюються контакт між сусідніми клітинами за допомогою групи плазмодесм (розд. 8.1.3 та рис. 8.7). Лігнін скріплює целюлозні волокна та утримує їх на місці. Він діє як дуже жорсткий і жорсткий матрикс, що посилює міцність клітинних стін на розтяг і особливо на стиск (запобігає прогинам). Він забезпечує клітинам додатковий захист від несприятливих фізичних і хімічних впливів. Разом з целюлозою, що залишається в клітинних стінках, лігнін надає деревині тих особливих властивостей, які роблять її незамінним будівельним матеріалом.
Функції клітинної стінки
Нижче перераховуються основні функції клітинних стінок рослин.
1. Клітинні стінки забезпечують окремим клітинам та рослині в цілому механічну міцність та опору. У деяких тканинах міцність посилюється завдяки інтенсивній лігніфікації клітинних стінок (невелика кількість лігніну є у всіх клітинних стінках).
2. Відносна жорсткість клітинних стінок і опір розтягуванню обумовлюють тургесцентність клітин, коли осмотичним шляхом надходить вода. Це посилює опорну функцію переважають у всіх рослинах і є єдиним джерелом опори для трав'янистих рослин й у таких органів, як листя, т. е. там, де відсутня вторинний зростання. Клітинні стінки також захищають клітини від розриву гіпотонічному середовищі.
3. Орієнтація целюлозних мікрофібрил обмежує і певною мірою регулює як ріст, так і форму клітин, оскільки від розташування цих мікрофібрил залежить здатність клітин до розтягування. Якщо, наприклад, мікрофібрили розташовуються впоперек клітини, оточуючи її як би обручами, то клітина, яку шляхом осмосу надходить вода, буде розтягуватися в поздовжньому напрямку.
4. Система пов'язаних один з одним клітинних стінок ( апопласт) служить головним шляхом, яким пересуваються вода і мінеральні речовини. Клітинні стінки скріплені між собою за допомогою серединних платівок. У стінках є невеликі пори, крізь які проходять цитоплазматичні тяжі, які називаються плазмодесмами. Плазмодесми пов'язують живий вміст окремих клітин - об'єднують усі протопласти в єдину систему, в так званий симпласт.
5. Зовнішні клітинні стінки епідермальних клітин покриваються особливою плівкою-кутикулою, що складається з воскоподібної речовини кутину, що знижує втрати води та зменшує ризик проникнення в рослину хвороботворних організмів. У пробковій тканині клітинні стінки після завершення вторинного зростання просочуються суберином, що виконує подібну функцію.
6. Клітинні стінки судин ксилеми, трахеїд та ситоподібних трубок (з ситоподібними пластинками) пристосовані для далекого транспорту речовин по рослині. Це питання у гол. 8 та 14.
7. Стінки клітин ендодерми кореня просякнуті суберином і тому є бар'єром на шляху руху води (розд. 14.1.5).
8. У деяких клітин їх видозмінені стінки зберігають запаси поживних речовин; таким способом, наприклад, запасаються геміцелюлози в деяких насінні.
9. У передаточних клітин площу поверхні клітинних стінок збільшено і відповідно збільшено площу поверхні плазматичної мембрани, що підвищує ефективність перенесення речовин шляхом активного транспорту(Розд. 14.8.6).
Освіта клітинної стінкипочинається під час поділу клітини. У площині поділу утворюється клітинна платівка, єдиний шар, загальний двох дочірніх клітин. Вона складається з пектинових речовин, що мають напіврідку консистенцію; целюлоза відсутня. У дорослій клітині клітинна платівка зберігається, але зазнає змін, тому її називають серединний, або міжклітинною платівкою (міжклітинною речовиною) (рис. 2.16). Середня платівка зазвичай дуже тонка і майже невиразна.
Відразу після утворення клітинної платівки протопласти дочірніх клітин починають відкладати власну клітинну стінку. Вона відкладається зсередини як на поверхні клітинної пластинки, так і на поверхні інших клітинних стінок, що раніше належали материнській клітині. Після поділу клітина вступає у фазу росту розтягуванням, який зумовлений інтенсивним осмотичним поглинанням клітиною води, пов'язаним із утворенням та зростанням центральної вакуолі. Тургорний тиск починає розтягувати стіну, але вона не рветься завдяки тому, що в неї постійно відкладаються нові порції мікрофібрил та речовин матриксу. Відкладення нових порцій матеріалу відбувається рівномірно по всій поверхні протопласту, тому товщина стінки клітин не зменшується.
Стінки клітин, що діляться і ростуть, називають первинними. Вони містять багато (60-90%) води. У сухій речовині переважають матричні полісахариди (60-70%), вміст целюлози не перевищує 30%, відсутня лігнін. Товщина первинної стінки дуже невелика (01-05 мкм).
Для багатьох клітин відкладення клітинної стінки припиняється одночасно із припиненням росту клітини. Такі клітини оточені тонкою первинною стінкою до кінця життя ( рис. 2.16).
Мал. 2.16. Паренхімна клітка з первинною стінкою.
В інших клітин відкладення стінки продовжується і після досягнення клітиною остаточного розміру. При цьому товщина стінки збільшується, а об'єм, який займає порожнина клітини, скорочується. Такий процес має назву вторинного потовщеннястінки, а саму стінку називають вторинної(рис. 2.17).
Вторинна стінка може розглядатися як додаткова, що виконує головним чином механічну опорну функцію. Саме вторинна стінка є відповідальною за властивості деревини, текстильного волокна, паперу. Вторинна стінка містить значно менше води, ніж первинна; в ній переважають мікрофібрили целюлози (40-50% від маси сухої речовини), які розташовуються паралельно один до одного. З полісахаридів матриксу характерні геміцелюлози (20-30%), пектинових речовин дуже мало. Вторинні клітинні стінки, як правило, зазнають здерев'янення. У неодревесневших вторинних стінках (луб'яні волокна льону, волоски бавовнику) вміст целюлози може досягати 95%. Великий змісті строго впорядкована орієнтація мікрофібрил визначають високі механічні властивостівторинних стін. Часто клітини, що мають вторинну одревесневу клітинну стінку, після завершення вторинного потовщення відмирають.
Середня платівка склеює сусідні клітини. Якщо її розчинити, стінки клітин втрачають зв'язок один з одним і роз'єднуються. Цей процес називається мацерація. Досить звичайна природна мацерація, при якій пектинові речовини серединної платівки переводяться в розчинний стан за допомогою ферменту пектинази і потім вимиваються водою (перестиглі плоди груші, дині, персика, банана). Часто спостерігається часткова мацерація, при якій серединна пластинка розчиняється не по всій поверхні, а лише у кутах клітин. Внаслідок тургорного тиску сусідні клітини у цих місцях округляються, внаслідок чого утворюються міжклітинні(рис. 2.16). Міжклітини утворюють єдину розгалужену мережу, яка заповнюється парами води та газами. Таким чином, міжклітини покращують газообмін клітин.
Характерна риса вторинної стінки – її нерівномірне відкладення поверх первинної стінки, внаслідок чого у вторинній стінці залишаються незтовщені ділянки – пори. Якщо вторинна стінка не досягає великої товщини, пори виглядають як дрібні заглиблення. У клітин із потужною вторинною стінкою пори у розрізі мають вигляд радіальних каналів, що йдуть від порожнини клітини до первинної стінки. За формою порового каналу розрізняють пори двох типів – простіі про каймлені(Рис. 2.17).
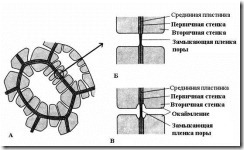
Мал. 2.17. Типи пір: А – клітини з вторинними стінками та численними простими порами; Б - пара простих пір; В – пара облямованих пір.
У простого часудіаметр порового каналу на всій довжині однаковий і має форму вузького циліндра. Прості пори характерні для паренхімних клітин, луб'яних та деревних волокон.
Пори у двох суміжних клітинах, як правило, виникають одна проти одної. Ці загальні пори мають вигляд одного каналу, розділеного тонкою перегородкою із серединної платівки та первинної стінки. Така сукупність двох пор суміжних стін сусідніх клітин носить назву пари пірта функціонує як одне ціле. Розділяючий їхній канал ділянка стінки називається замикаючою плівкою пори, або поровий мембраною. У живих клітинах замикаюча плівка пори пронизана численними плазмодесмами(рис. 2.18).
Плазмодесмивластиві тільки рослинним клітинам. Вони є тяжі цитоплазми, що перетинають стінку суміжних клітин. Число плазмодесм в одній клітці дуже велике – від кількох сотень до десятків тисяч, зазвичай плазмодесми зібрані до груп. Діаметр плазмодесменного каналу становить 30-60 нм. Його стінки вистелені плазмалемою, безперервною з плазмалемою суміжних клітин. У центрі плазмодесми проходить мембранний циліндр. центральний стрижень плазмодесми, безперервний з мембранами елементів ендоплазматичної мережіобох клітин. Між центральним стрижнем та плазмалемою в каналі знаходиться гіалоплазма, безперервна з гіалоплазмою суміжних клітин.
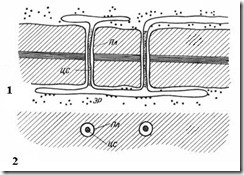
Мал. 2.18. Плазмодесми під електронним мікроскопом (схема): 1 - на поздовжньому зрізі; 2 – на поперечному зрізі; Пл- Плазмалема; ЦС– центральний стрижень плазмодесми; ЕР- Елемент ендоплазматичного ретикулуму.
Таким чином, протопласти клітин не повністю ізольовані один від одного, а повідомляються каналами плазмодесм. Ними відбувається міжклітинний транспорт іонів і дрібних молекул, і навіть передаються гормональні стимули. За допомогою плазмодесм протопласти клітин у рослинному організміутворюють єдине ціле, зване симпластом, а транспорт речовин через плазмодесми отримав назву симпластичногона відміну від апопластичноготранспорту по клітинних стінках та міжклітинниках.
У облямованих пір(рис. 2.17)канал різко звужується в процесі відкладення клітинної стінки, тому внутрішній отвір пори, що виходить у порожнину клітини, набагато вже, ніж зовнішнє, що упирається в первинну стінку. Облямовані пори характерні для клітин водопровідних елементів деревини, що рано відмирають. У них поровий канал у напрямку замикаючої плівки воронковидно розширюється, а вторинна стінка нависає у вигляді валика над розширеною частиною каналу, утворюючи камеру пори. Назва обрамленої пори походить від того, що при розгляді з поверхні внутрішній отвір має вигляд маленького кола або вузької щілини, тоді як зовнішній отвір як би облямовує внутрішній у вигляді кола більшого діаметра або ширшої щілини.
Пори полегшують транспорт води та розчинених речовин від клітини до клітини, водночас не знижуючи міцності клітинної стінки.