ТКАНИНИ. КЛАСИФІКАЦІЯ ТКАНИН.
В основі організації вищих рослинлежить принцип спеціалізації клітин, який у тому, кожна клітина організму виконує в повному обсязі властиві їй функції, лише деякі, зате повніше і досконало.
Тканини- стійкі, закономірно повторювані комплекси клітин, подібні за походженням, будовою та пристосовані до виконання однієї або кількох функцій.
Існують різні класифікації тканин, але вони досить умовні.
Залежно від основної функції розрізняють кілька груп рослинних тканин.
1. Освітні тканиниабо меристеми,- мають здатність до поділу та формування всіх інших тканин.
2. Покривні тканини:
Первинні;
Вторинні;
Третинні.
3. Основні тканини- складають більшу частину тіла рослини. Розрізняють такі основні тканини:
асиміляційні (хлорофілоносні);
Запасні;
Повітроносні (аеренхіма);
Водоносні.
4. Механічні тканини(опорні, скелетні):
Колленхіма;
Склеренхіма.
5. Провідні тканини:
Ксилема (деревина) - тканина висхідного струму;
Флоема (луб) – тканина низхідного струму.
6. Видільні тканини:
Зовнішні:
Залізисті волоски;
Гідатоди - водяні продихи;
Нектарники;
Внутрішні:
Видільні клітини з ефірними оліями, смолами, дубильними речовинами;
Багатоклітинні вмістилища виділень, млечники.
За здатністю клітин до поділу розрізняють два типи тканин: освітні,або меристеми,і постійні- покривні, видільні, основні, механічні, провідні.
Тканину називають простий,якщо всі її клітини однакові за формою та функціями (паренхіма, склеренхіма, коленхіма). Складнітканини складаються з клітин, неоднакових за формою, будовою та функціями, але пов'язаних загальним походженням (наприклад, ксилема, флоема).
Існує також класифікація тканин, заснована на їхньому походженні (онтогенетична). Відповідно до цієї класифікації розрізняють первинні та вторинні тканини. З первинної меристеми, що знаходиться на верхівці втечі та кінчику кореня, а також із зародка насіння формуються первинні постійнітканини (епідерма, коленхіма, склеренхіма, асиміляційна тканина, епіблема). Клітини постійних тканин нездатні до подальшого поділу. З клітин спеціалізованої меристеми - прокамбія - формуються первинні провіднітканини (первинна ксилема, первинна флоема).
З вторинної меристеми - камбія - формуються вториннітканини: вторинна ксилема, вторинна флоема; з фелогену утворюються пробка, феллодерма, чечевички, що виникають при потовщенні стебла та кореня. Вторинні тканини, як правило, зустрічаються у голонасінних і дводольних покритонасінних рослин. Потужний розвиток вторинних тканин - деревини та лубу характерний для деревних рослин.
ОСВІТНІ ТКАНИНИ
Освітні тканинизавдяки постійному мітотичному поділу їх клітин забезпечують утворення всіх тканин рослини, тобто. фактично формують його тіло. Будь-яка клітина у своєму розвитку проходить три стадії: ембріональну, зростання та стадію диференціації (тобто придбання клітиною певної функції). У міру диференціації зародка первинна меристема зберігається тільки на верхівці майбутньої втечі (у конусі наростання) та на кінчику кореня – апікальні (верхівкові) меристеми.Зародок будь-якої рослини складається з клітин меристеми.
Цитологічна характеристика мерістему.Типові ознаки найвиразніше виражені у верхівкових меристемах. Ці меристеми складені ізодіаметричними багатогранними клітинами, не розділеними міжклітинниками. Їхні оболонки тонкі, містять мало целюлози і здатні розтягуватися.
Порожнина кожної клітини заповнена густою цитоплазмою з відносно великим ядром, що займає центральне положення, і мітозом, що інтенсивно ділиться. У гіалоплазмі багато дифузно розкиданих рибосом, пропластид, мітохондрій та диктіосом. Вакуолей небагато, і вони дрібні. Провідні тканини утворюються з меристеми, що має прозенхімну форму і великі вакуолі - прокамбію і камбію. Клітини прокамбію в поперечному перерізі багатокутні, клітини камбію - прямокутні.
Клітини, що зберігають свої меристематичні властивості, продовжують ділитися, утворюючи нові і нові клітини, звані ініціалями.Частина дочірніх клітин диференціюється, перетворюючись на клітини різних тканин, їх називають похідними ініціалів.Клітини ініціалів можуть ділитися невизначено багато разів, а похідні ініціалів діляться один або кілька разів і перетворюються на постійні тканини.
За походженням розрізняють первинні та вторинні меристеми.
Первинні меристеми
Первинні меристеми походять безпосередньо з меристеми зародка і мають здатність до поділу. По положенню в рослині первинні меристеми можуть бути верхівковими (апікальними), вставковими (інтеркалярними) та бічними (латеральними).
Верхівкові (апікальні) меристеми- такі меристеми, які розташовуються у дорослих рослин на верхівках стебел та кінчиках коріння та забезпечують зростання тіла у довжину. У стебел у конусі наростання виділяють два меристематичні шари: туніку, з якої утворюються покривна тканина і периферична частинапервинної кори, і корпус, з якого утворюються внутрішня частина первинної кори та центральний осьовий циліндр(Рис. 2.3).
Мал. 2.3.Апікальні меристеми стебла: а- поздовжній зріз: 1 – конус наростання; 2 – зачаток листа; 3 - горбок пазушної нирки;
У кінчику кореня розрізняють три шари:
1) дерматоген, з якого утворюється первинна покривно-всмоктуюча тканина – ризодерма;
2) периблему, з якої розвиваються тканини первинної кори;
3) плером, що утворює тканини центрального осьового циліндра.
Бічні (латеральні) меристемиза походженням можуть бути первинними та вторинними, на поперечному зрізі осьових органів мають вигляд кілець. Прикладом первинної бічної меристеми є прокамбій і перицикл. З прокамбіяформуються камбій та первинні елементи судинно-волокнистих пучків (первинна флоема та первинна ксилема), при цьому клітини прокамбію безпосередньо диференціюються в клітини первинних провідних тканин.
Бічні меристеми розташовуються паралельно поверхні органу та забезпечують зростання осьових органів у товщину.
Вставкові (інтеркалярні) меристемичастіше первинні і зберігаються у вигляді окремих ділянок у зонах активного зростання різних частинахрослини (наприклад, в основі черешків листя, біля основ міжвузля). В основі міжвузлів у злаків діяльність цієї меристеми веде до подовження міжвузлів, що забезпечує зростання стебла в довжину.
Вторинні меристеми
До вторинних відносять бічні та ранові меристеми.
Бічні (латеральні) меристемипредставлені камбіємі феллогеном.Вони формуються з промеристем (прокамбію) або постійних тканин шляхом їх дедиференціювання. Клітини камбію діляться перегородками, паралельними поверхні органу (периклінально). З клітин, відкладених камбієм назовні, розвиваються елементи вторинної флоеми, та якщо з клітин, відкладених всередину, - вторинної ксилемы. Камбій, що виник з постійних тканин шляхом дедиференціювання, називають додатковим.За будовою та функцією він не відрізняється від камбію, що виник з промеристем. Феллоген формується із постійних тканин, розташованих у субепідермальних шарах (під епідермою). Ділячись периклінально, феллоген відокремлює назовні майбутні клітини пробки (фелеми), а всередину - клітини фелодерми. Таким чином, феллоген формує вторинну покривну тканину – перидерму. Бічні меристеми розташовуються паралельно поверхні органу та забезпечують зростання осьових органів у товщину.
Раневі меристемиутворюються при пошкодженні тканин та органів. Навколо пошкодження живі клітини дедиференціюються, починають ділитися і цим перетворюються на вторинну меристему. Їхнє завдання - утворити щільну захисну тканину, що складається з паренхімних клітин, - каллюс.Ця тканина білуватого або жовтуватого кольору, її клітини мають великі ядра і досить товсті клітинні стінки. Каллюс виникає при щепленнях, забезпечуючи зрощення щепи з підщепою, і в основі черешків. У ньому можуть закладатися придаткові коріння та нирки, тому його використовують для одержання культури ізольованих тканин.
ПОКРОВНІ ТКАНИНИ
Первинна покривна тканина
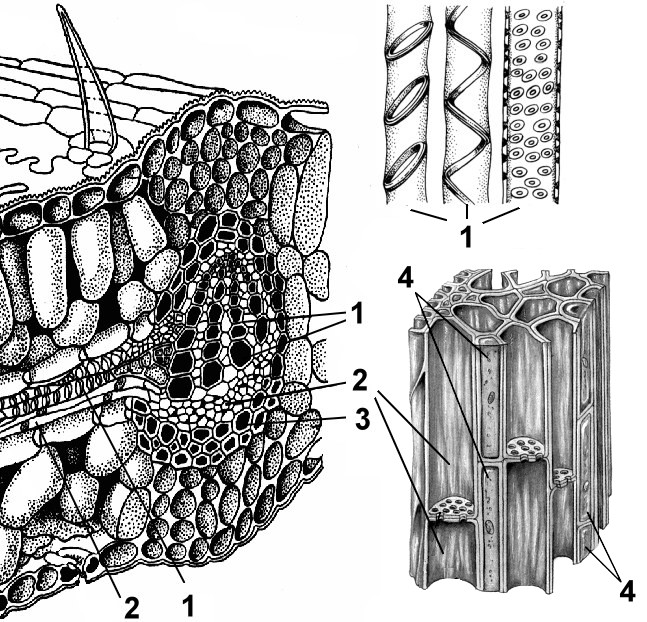
До первинної покривноїтканини відносять епідерму власне епідермальних, навколоустьичних, замикаючих клітин продихом і трихом.
Пектинові речовини та целюлоза, що входять до клітинну стінку, можуть піддаватися ослизненню з освітою слизіві камедей.Вони являють собою полімерні вуглеводи, споріднені з пектиновими речовинами, і відрізняються здатністю до сильного набухання при зіткненні з водою. Камеді в набряклому стані клейкі і можуть витягуватися в нитки, тоді як слизу сильно розпливаються і нитки не витягуються. Пектинові слизу зустрічаються у представників сімейств лілейних, хрестоцвітих, мальвових, липових, розоцвітих, на відміну від целюлозних слизів, що зустрічаються набагато рідше (наприклад, у орхідних).
Устячка
являють собою високоспеціалізовані утворення епідерми, що складаються з двох замикаючих клітин бобоподібної форми та устьичної щілини (своєрідного міжклітинника між ними). Є головним чином листі, але трапляються й у стеблі (рис. 2.6). 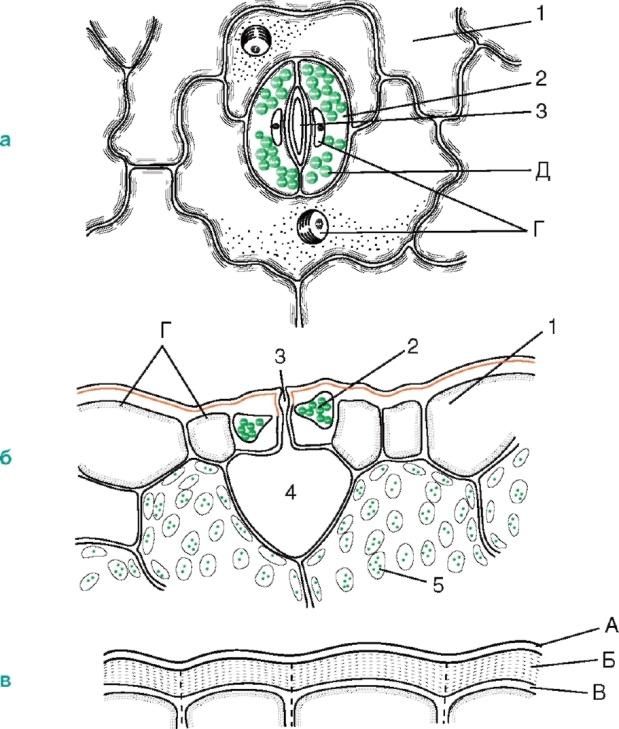
Мал. 2.6.Будова продихи: а, б- Шкірка листа чебрецю (вид зверху та на поперечному зрізі); в- Шкірка зі стебла цереуса (родина кактусових); 1 – власне епідермальні клітини; 2 - замикаючі клітини продихіва; 3 – устьична щілина; 4 - повітроносна порожнина; 5 – клітини хлорофілоносної паренхіми; А – кутикула; Б - кутикулярний шар - оболонка із суберином та воском; В – целюлозний шар стінки; Г - ядро з ядерцем; Д - хлоропласти
Стіни замикаючих клітин потовщені нерівномірно: стінки, спрямовані до щілини (черевні), значно потовщені порівняно зі стінками, спрямованими від щілини (спинними). Щілина може розширюватися і звужуватися, регулюючи транспірацію та газообмін. Під щілиною розташована велика дихальна порожнина (міжклітинник), оточена клітинами мезофілу листка.
Замикаючі клітини оточені навколоустьовими клітинами, які спільно утворюють устьичний комплекс(Рис. 2.7). Розрізняють такі основні типи устьичних комплексів: 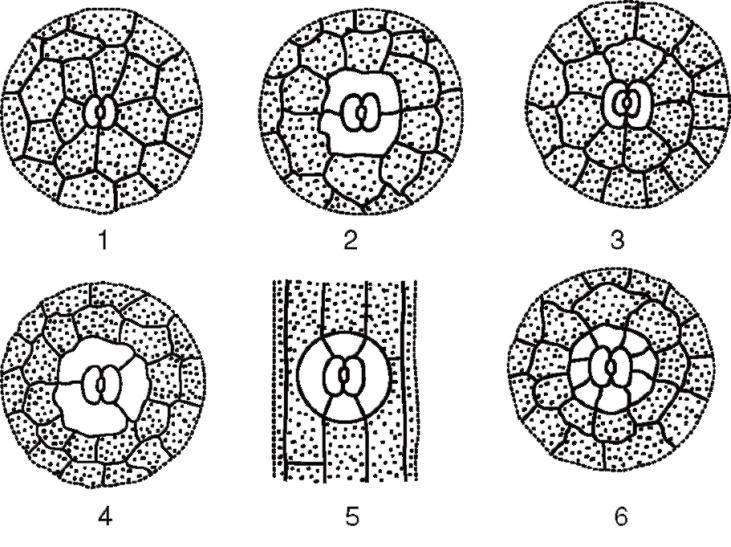
Мал. 2.7.Основні типи устьичного апарату: 1 – аномоцитний (у всіх вищих рослин, крім хвощів); 2 - діацитний (у папоротей та квіткових); 3 - парацитний (у папоротей, хвощів, квіткових та гнетових); 4 – анізоцитний (тільки у квіткових); 5 - тетрацитний (переважно у однодольних); 6 - анциклоцитний (у папоротей, голонасінних та квіткових)
1) аномоцитний(безладний) - замикаючі клітини немає яскраво виражених околоустьичных клітин; характерний всім вищих рослин, крім хвойні;
2) анізоцитний(Нерівноклітинний) - замикаючі клітини продихи оточені трьома навколоустьичними клітинами, одна з яких набагато більша (або менше) інших;
3) парацитний(паралельно-клітинний) - одна навколоустьева клітина (або більше) розташована паралельно замикаючим;
4) діацитний(перехресно-клітинний) – дві навколоустьичні клітини розташовані перпендикулярно замикаючим;
5) тетрацитний(Від грец. tetra- чотири) - головним чином у однодольних;
Продихання розташовані на нижній стороні листа, але у водних рослинз плаваючим листям вони знаходяться тільки на верхній стороні листа. За формою клітин епідерми листа і розташування продихів можна відрізнити однодольне рослина від дводольного (рис. 2.8). Власне епідермальні клітини листка дводольних рослин в контурах хвилясті (рис. 2.9), а у однодольних вони витягнуті, ромбічної форми. 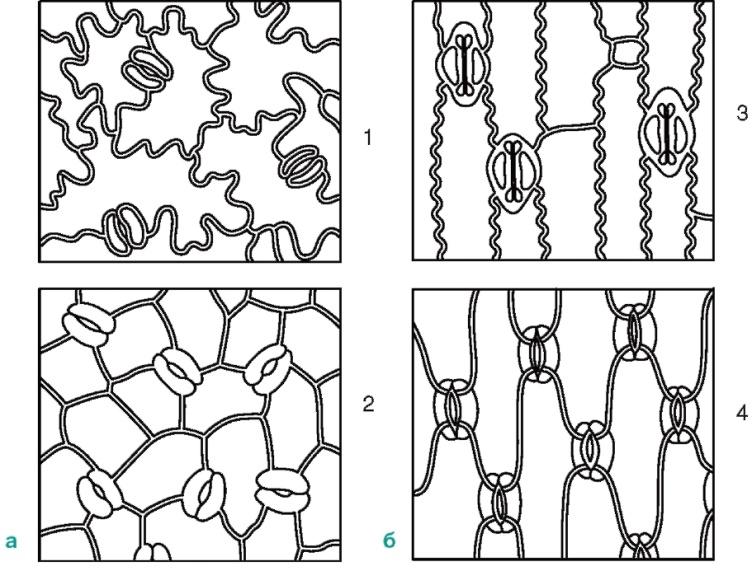
Мал. 2.8.Розташування продихів на епідермі (вид з поверхні): а-дводольні рослини: 1 - Букви; 2 – кавун; б-однодольні рослини: 3 – кукурудза; 4 - ірис
Типи продихів за рівнем розташування щодо поверхні епідермісу можна підрозділити наступним чином.
1.7.1. Продихання, розташовані в одній площині з епідермісом. Найбільш поширений тип і зазвичай у описі мікроскопії лікарської рослинної сировини не вказується, тобто. цей пункт опускається. Діагностичними ознаками будуть або виступаючі, або занурені продихи.
1.7.2. Простирадла устячка - продихи, розташовані над епідермісом. Зазвичай при обертанні мікрогвинта мікроскопа (при опусканні об'єктиву) спочатку виявляються такі продихи, а вже потім з'являються клітини епідермісу, тому на фотографії відобразити їх з поверхні листа практично неможливо, так само як і зобразити на малюнку. В одній площині з епідермісом такі продихи можна побачити на поперечних зрізах, але для цього зріз повинен пройти через продихи, що важко отримати при їх рідкісному розташуванні на листі. Такі продихи характерні, наприклад, для листя мучниці.
1.7.3. Занурені продихи - продихи, занурені в епідерміс. При спостереженні під мікроскопом при обертанні мікрогвинта (при опусканні об'єктиву) спочатку чітко виявляються клітини епідермісу, потім стає можливим чіткіше побачити контури продихів. На фотографіях та малюнках препаратів із поверхні їх також важко відобразити. Зустрічаються в листі конвалії, листі вахти, листках евкаліпта. Іноді поглиблення, в яких розташовуються продихи, вистелені або прикриті волосками і називаються устьичними криптами.
1.8. Типи устьичних клітин
У літературі описується 19 типів, нами обрані лише ті, що використовуються в аналізі лікарської рослинної сировини.
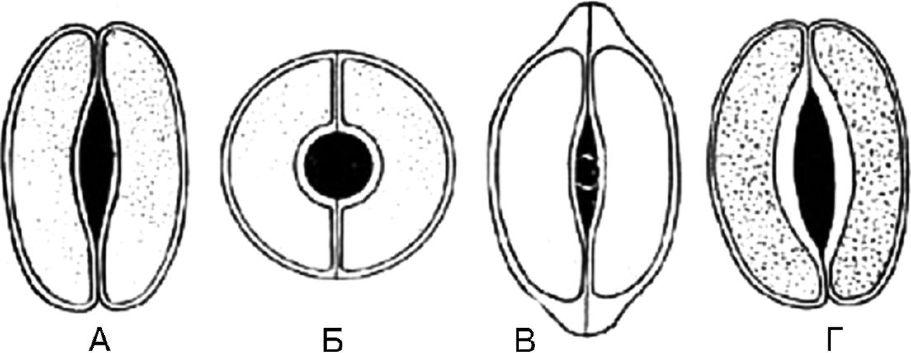
Мал. 63.Типи устьичних клітин. А - сочевицеподібні; Б - сфероподібні; В - ковпачкоподібні; Г – човноподібні
1.8.1. Сочевицеподібні - 2 однакові клітини напівмісячної форми розташовані симетрично. На передній поверхні потовщення оболонки майже рівномірне. Щілина веретеноподібна (рис. 63, А). Тип устьичних клітин характерний більшості рослин.
1.8.2. Сфероподібні - дві однакові, сильно кругоподібно зігнуті клітини розташовані симетрично. На передній поверхні потовщення оболонки майже рівномірне. Щілина кругла (рис. 63, Б).
1.8.3. Ковпачкоподібні - дві однакові клітини напівмісячної форми у полярних частинах мають потовщення у вигляді ковпачка. Щілина веретеноподібна (рис. 63, В). Зустрічаються у наперстянок.
1.8.4. Човноподібні - Внутрішні стінки устьичних клітин потовщені. Щілина веретеноподібна (рис. 63, Г). Спостерігається у траві золототисячника, у листі вахти.
Механізм роботи продихів обумовлений осмотичними властивостями клітин. При освітленні поверхні листа сонцем у хлоропластах замикаючих клітин відбувається активний процес фотосинтезу. Насичення клітин продуктами фотосинтезу, цукрами тягне за собою активне надходження до клітин іонів калію, внаслідок чого концентрація клітинного соку в клітинах, що замикають, зростає. Виникає різниця концентрації клітинного соку навколоустьових та замикаючих клітин. З огляду на осмотичних властивостей клітин вода з околоустьичных клітин надходить у замикаючі, що веде до збільшення обсягу останніх і різкого зростання тургора. Потовщення «черевних» стінок клітин, що замикають, звернених до устьичної щілини, забезпечує нерівномірне розтягнення клітинної стінки; замикаючі клітини набувають вираженої бобоподібної форми, і устьична щілина відкривається. При зниженні інтенсивності фотосинтезу (наприклад, увечері) зменшується утворення цукрів у клітинах, що замикають. Приплив іонів калію припиняється. Концентрація клітинного соку в клітинах, що замикають, знижується в порівнянні з околоустьічними. Вода шляхом осмосу йде з клітин, що замикають, знижуючи їх тургор, в результаті вночі устьична щілина закривається.
Клітини епідерми щільно зімкнуті між собою, завдяки цьому епідерма виконує цілу низку функцій:
Перешкоджає проникненню хвороботворних організмів усередину рослини;
Захищає внутрішні тканини від механічних ушкоджень;
Регулює газообмін та транспірацію;
Через неї виділяються вода, солі;
Може функціонувати як всмоктувальна тканина;
бере участь у синтезі різних речовин, сприйнятті подразнень та у русі листя.
Трихоми - різні за формою, будовою та функціями вирости клітин епідерми: волоски, лусочки, щетинки тощо. Їх поділяють на криючі та залізисті. Залізисті трихоми,на відміну від криючих, мають клітини, що виділяють секрет. Кріючі волоски,утворюючи на рослині шерстистий, повстяний або інший покрив, відбивають частину сонячних променіві цим зменшують транспірацію. Іноді волоски знаходяться тільки там, де розташовані продихи, наприклад, на нижній стороні листа мати-й-мачухи. У деяких рослин живі волоски збільшують загальну поверхню, що випаровує, що сприяє прискоренню транспірації.
Розміри трихом значно варіюють. Найбільш довгі трихоми (до 5-6 см) покривають насіння бавовнику. Кроючі трихоми мають форму простих одно або багатоклітинних, розгалужених або зірчастих волосків. Кріючі трихоми можуть тривалий часзалишатися живими чи швидко відмирати, заповнюючись повітрям.
Від тріхом, що виникають лише за участю епідермальних клітин, відрізняються емергенці, в освіті яких беруть участь і глибше розташовані тканини субепідермальних шарів.
Анатомо-діагностичні ознаки, що мають найбільше значенняі високу варіабельність щодо лікарської сировини. Волоски можуть бути прості та головчасті, які у свою чергу можуть бути одноклітинними та багатоклітинними. Багатоклітинні волоски можуть бути однорядними, дворядними та гіллястими.
ПОКРОВНІ ТКАНИНИ.
- Загальна характеристиката св-ва.
- Будова та ф-ції первинної покривної тканини – епідерми.
Функції епідерми
основ. клітини епідерми
- Будова вторинної покривної тканини – перидерми
Освіта та закладення
Будова сочевичок
- Будова кірки – ретидому.
Загальна характеристика та св-ва.
Покривні тканини покривають тіло рослини згори, тобто. розташовуються на поверхні і безпосередньо контактують з зовнішнім середовищем. З одного боку, вони захищають рослини від будь-яких несприятливих впливів (від проникнення мікроорганізмів, від випаровування вологи), тому клітини покривних тканин щільно зімкнуті, без міжклітинників. З іншого боку, вони повинні забезпечувати зв'язок рослин із довкіллям (газообмін, транспірацію), тому в них є особливі структури.
Таким чином, будова та властивості покривних тканин визначаються виконуваними ними функціями.
Властивості: 1. Багатофункціональність, тобто. це складні тканини, складаються з кількох типів клітин, що виконують різні функції.
2. Забезпечують виборча проникність (для газів, Н 2 Про та ін-в), для чого є спеціальні структури.
3. Постійна змінатканин із віком, або зміна їх функцій.
Розрізняють: первинну, вторинну та третинну покривні тканини, які змінюють одна одну з віком.
2. Первинна покривна тканина – епідерма (шкірка).
Виникла як наслідок еволюційного пристосування рослин до наземних умов життя.
(Епі - грец. "Зверху", "над", дерма - "шкіра").
а) Виникає із зовнішнього шару апікальної (верхівкової) меристеми втечі – протодерми, отже – це первинна покривна тканина; захищає та покриває листя, молоді стебла, квіти та плоди вищих рослин.
Епідерма – первинна покривна тканина рослини.
Зазвичай одношарова, складається з трьох типів клітин, виконують різні ф-ции, тобто. це складна тканина.
Включає:
1) щільно зімкнуті основні клітини епідерми,
2) замикаючих та побічні клітини продихів,
3) трихоми - похідні епідерми у вигляді виростів та волосків.
Функції та властивості епідерми визначаються особливостями будови цих клітин, які ідеально пристосовані до наземного існування рослин.
(При переході до повторного водного способу життя квіткові рослини, наприклад, можуть втратити продихи і захисні структури основних клітин епідерми).
1. Основні клітини епідерми.
1) Утворюють 1 шар живих, прозорих клітин (т.к. розташовуються на зростаючих органах), отже добре пропускають світло до розташованих нижче асим. клітин, де йде фотосинтез.
Захисну та покривну функціюзабезпечують інші особливості:
2) щільно зімкнуті, без міжклітинників
3) товсті зовнішні стінки клітин, тонкі бічні
4) бічні стінки клітин часто мають хвилясту форму, що покращує їх змикання та прилягання один до одного.
5) Оболонка клітини влаштована складно: нижня частинаскладається з целюлози, а верхня кутинізована.
6) Дуже характерне утворення на поверхні оболонки шару кутикули, що регулює зменшення випаровування та газообмін.
Кутикула– тонкий, прозорий шар кутину та рослинних восків на зовнішніх стінках основних клітин. Віск – жироподібне по-во, зменшує проникність для води та газів. Отже, потужність кутикули, розподіл восків і кутину, число та характер пір визначають проникність для розчинів та газів, хімічну стійкість, бактерицидну стійкість.
У вологомустан кутикула більш проникний для газів і рідин, ніж у сухому, тому в дощ через кутикулу йде поглинання водних розчинів(Позакореневе підживлення рослин).
7) Добре розвинені ап. Гольджі та ЕПС, отже в осн. клітинах епідерми йде синтез ряду в-в(вуглеводів, білків тощо) (біосинтетична функція епідерми)
Замикаючі та побічні клітини продихів.
Продихання, або устьичний апарат– це дві замикаючі клітини бобовидної форми з різнопотовщеними стінками та з розташованим між ними міжклітинником – устьичною щілиною (внутрішні стінки устьичних клітин, звернені до щілини товсті, а зовнішні тонкі).
Рідше поряд з ними знаходяться побічні клітини епідерми, що відрізняються від основних клітин.
Під продихом розташована подустьева повітряна порожнина, оточена клітинами паренхіми та пов'язана із системою міжклітинників органу.
Характерно:
1) Є хлоропласти (фотосинтез вуглеводів) та мітохондрії (синтез АТФ). Потрібні, т.к. розкриття устьичної щілини відбувається активно з витратою енергії АТФ, а змикання пасивне.
2) За рахунок різнопотовщеності стінок замикаючих клітин утворюється щілина.
Отже, осн. функції:
1 – транспірація(регульоване випаровування води)
Газообмін.
(Самостійно, вивчити механізм роботи продихів).
Механізм відкривання та закривання устьичної щілини заснований на осмотичних явищах, а також беруть участь усі органоїди протопласту замикаючих клітин.
- - K-Na – насос(механізм швидкого реагування). При високому вмісті в рослині води в плазмалемі замикаючих клітин працюють білки-переносники К + , активно, з витратою енергії АТФ іони К + з навколишніх клітин в цитоплазму, і далі вони надходять у вакуолі замикаючих клітин продихів. При подальшому підвищенні концентрації іонів К + , слідом за ними, пасивно, за градієнтом концентрації, у замикаючі клітини продихів з навколишніх клітин надходить вода. Об'єм вакуолей збільшується, осмотичний тиск у клітинах, що замикають, підвищується, оболонки клітин розтягуються. Через те, що оболонки замикаючих клітин потовщені нерівномірно, зовнішня оболонки розтягуються сильніше, а внутрішні (навернені один до одного) не можуть розтягнутися і розходяться, між ними утворюється щілина – продихи відкриваються. При нестачі води в рослині білки переносники іонів К+ припиняють свою роботу. К-Nа – насос зупиняється. При цьому іони К + за градієнтом концентрацій пасивно залишають устьичні клітини і йдуть в навколишні клітини, слідом за ними йде вода. Клітки, що замикають, втрачають тургор і пасивно стуляються - продихи закриваються.
Т.ч., відкриття устьичної щілини відбувається активно, з витратою енергії, джерелом якої є вуглеводи, що накопичуються хлоропластами. Закриття щілини відбувається пасивно, без витрат енергії.
- – Добова зміна концентрації цукрів(вдень багато, до кінця ночі – мало) (бо є хлоропласти). Протягом дня в хлоропластах клітин, що замикають, йде фотосинтез. Отже, на світлі в них підвищується концентрація глюкози і в міру її накопичення, до вечора, за градієнтом концентрації всередину клітин надходить вода. Замикаючі клітини набувають пружного тургорного стану, продихи розкриваються. Вночі фотосинтез припиняється, глюкоза витрачається на дихання клітин, що замикають, її концентрація знижується. Через це за градієнтом концентрації вода залишає замикаючі клітини, вони втрачають тургор і спадають. Устьична щілина закривається. Вранці процес повторюється. Наразі вважається, що цей механізм обмежений, основним є робота К-Nа – насоса.
Різні рослини виробили певний ритм роботи устькового апарату. У більшості рослин продихи відкриті і вдень і вночі, закриваючись лише при зменшенні вмісту води. У жаркі години продихи зазвичай закриті, але у рослин пустель вони відкриті (за допомогою випаровування вони охолоджують поверхню листя і активніше поглинають воду). У деяких рослин, наприклад у каланхое, продихи відкриваються вночі і закриваються вдень.
Типи продихів:
Важливі під час встановлення спорідненості таксонів рослин.
1.- анамоцитні продихи- мають побічні клітини, що не відрізняються від осн. клітин епідерми (хар-ни всім груп рослин, крім хвощів).
2.- діацитні продихи– мають дві побічні клітини, загальна стінка яких перпендикулярна до устьичної щілини (є у ряду квіткових, у сем. губоцвітих і гвоздичних).
3. – парацитні продихи- побічні клітини розташовуються паралельно замикаючим клітинам і устьичної щілини (є папоротей, хвощів і ряду квіткових рослин).
4. – анізоцитні продихи- Замикаючі клітини оточені трьома побічними, одна з яких або більша, або менше інших (тільки у квіткових рослин).
5. – тетрацитні продихи- мають 4 побічні клітини (є в однодольних).
6. – енциклоцитні продихи- Побічні клітини утворюють вузьке кільце навколо замикаючих клітин (є у папороті, голонасінних і ряду квіткових).
Якщо продихи відкриті, випаровування йде так, ніби епідерми немає зовсім.
Число і розташування продихів варіює у різних рослин. У середньому, у рослини 100-700 продихів на 1 мм 2 . Зазвичай, у наземних рослин більша частинапродихів розташована на нижній стороні листя, зверху їх дуже мало чи ні зовсім. Це пояснюється такими причинами: 1) зверху листя сильніше нагрівається, отже, випаровування йтиме інтенсивніше і втрати вологи будуть більшими; 2) вуглекислий газутворюється в грунті і піднімається вгору, потрапляючи відразу в продихи, до того ж, він важчий за повітря і накопичується зазвичай в нижніх повітряних шарах.
У рослин з вертикально стоїть листям (ребром), наприклад, у евкаліпта, продихи розподілені по обидва боки листа. У водних рослин з плаваючим листям устячка розташовані на верхній стороні.
- Трихоми– волоски, вирости та похідні епідерми.
Форма, будова – систематична ознака визначення видів і пологів рослин.
Бувають: залізисті та криючі.
Залізисті трихомиутворюють та накопичують екскрети та секрети. Це ефірні олії(герань, м'ята та ін) або захисні в-ва (кропива). Крапля олії виділяється під кутикулу, накопичується там, потім при розриві кутикули виходить назовні. Кутикула відновлюється і під нею накопичується нова крапля ефірної олії.
Функції:
1-захисна
2-терморегуляція
3- бактерицидна (ефірні олії – фітонциди)
Кріючі трихомискладаються:
1- часто з мертвих клітин заповнених повітрям (одно-або багатоклітинні мертві трихоми)
Функції: відбиття сонячних променів, сприятливі мікрокліматичні умови для роботи продихів, зменшення нагріву. Отже, зниження транспірації та хутро. захист (рослини пустель та степів).
2- рідко з живих клітин.
Функції: збільшують загальну випаровування. поверхню. Отже, збільшення випаровування (рослини вологих тропіків та тропічних боліт).
Т.ч., епідерма як багатофункціональна складна тканина виконує функції лише у живому вигляді.
1. Регуляція газообміну та транспірації (між рослиною та окр. середовищем)
2. Захисна (хутряна, бактерицидна, від зайвого випаровування води).
3. Видільна (виділяє солі, воду, ефірні олії).
4. Поглинальна (всмоктувальна) (вода з хв. в-вами----позакореневі підживлення).
5. Біосинтетична (синтез орг. в-в: кутинів, восків тощо)
Практич. вик.
- епідермальні волоски (бавовник --- тканина).
- ефірні олії --- парфумеріята медицина.
При подальшому зростанні стебла на 1 місце висувається забезпечення хутра. міцності, захист від втрати води та різких перепадів температур, тому епідерма замінюється вторинною покривною тканиною – перидермою.
3. Вторинна покривна тканина – перидерма.
П. – багатошарова, вторинна покривна тканина.
Виникає після епідерми. Її освіта починається із закладки вторинної меристеми – фелогену (коркового камбію). Він закладається в однорічних пагонах дерев та чагарників до середини літа.
Його освіта може йти 3 способами:
1) найчастіше закладається у клітинах субепідермального шару (під епідермою)
2) іноді феллоген утворюється в глибших шарах кори (малина, смородина),
3) рідко феллоген обр-ся в епідермі при тангенціальному розподілі її клітин.
(верхня клітина залишається епідермальною, а нижня стає клітиною феллогену).
Крім феллогену до складу перидерми входять ще 2 типи клітин, отже це теж складна тканина.
Типи клітин перидерми:
1) фелема (пробка) --- захисна функція
2) феллоген (корковий камбій) --- освітняфункція
3) фелодерма --- Виконує поживну функцію по відношенню до феллогену.
Будова перідерми.
1. Основа – фелоген, одношарова меристема, що виникає з живих паренхімних клітин постійних тканин, отже - вторинна тк.
Його клітини діляться тангентально і відкладають назовні багато шарів клітин пробки – фелеми(головні клітини), а всередину клітини фелодерми.
Спочатку клітини тонкостінні пробки, потім вони опробковевают (на внутрішньої поверхніклітинних оболонок відкладається ізолятор - суберин) і живий вміст їх відмирає і заповнюється повітрям (отже, такі мертві клітини набувають хороших ізолюючих властивостей).
У такому стані вони виконують основні функції:
1) захисна(механічний, захист від втрати вологи, бактерицидний захист, тому що стійкий до руйнування);
2) терморегуляційна(пробка терморегулятор, тому що добре захищає рослину від перепадів t o , отже, відкладається у деревних рослин до кінця літа - початку осені (завдяки пробці рослина поступово промерзає і поступово відтає, що не дає утворитися в цитоплазмі клітин кристалів льоду, які ушкоджують і руйнують клітину).
Феллоген заліковує рани при пошкодженні рослини та при сприятливих умовпостійно формує нові верстви пробки. Тому перидерма – багатошарова тканина. Після розвитку перидерми, епідерма, відрізана від притоку поживних речовині Н 2 О, відмирає і злущується (зелене забарвлення стебла замінюється на коричневу).
Живі тканини, що лежать під пробкою-фелемою, потребують газообміну, тому в перидермі формуються. чечевички - прохідні розриви в пробці,через які відбувається газообмін. Зазвичай чечевички закладаються під продихами. Замість пробки фелогена тут відкладає живі клітини паренхіми.
Поперечний зріз чечевички показує, що шари пробки чергуються з паренхімними клітинами з добре розвиненими міжклітинниками. По міжклітинниках і йде газообмін. Феллоген підстилає паренхімну тканину, т.к. у ньому є міжклітини, він не перешкоджає газообміну.
До зими феллоген відкладає під паренхімним шаром клітин замикаючий шар мертвих клітин пробки-фелеми –чечевичка закривається (т.к. взимку рослина різко скорочує газообмін та обмінні процеси майже до 0).
Навесні під натиском нових паренхімних клітин, що відкладаються феллогеном, цей шар розривається і відновлюється газообмін.
Чечевички у різних видівдерев та чагарників мають свою форму та розмір та змінюються з віком.
Часто у дерев на зміну перидермі приходить кірка – ретидом– третинна покривна тканина (наприклад, у сосни, яблуні, ясена, клена; у платана та евкаліпта – кірки немає).
Шкірка дуже сильна, товста, тріщинувата.
Освіта.
Корка утворюється при багаторазовому закладанні шарів феллогену, та був і перидерми у глибоких тканинах кори.
Живі клітини, опинившись між цими шарами, гинуть і стають частиною кірки.
Т.ч., кірка складається з шарів пробки, що чергуються, і клітин інших, відмерлих тканин кори, тобто. - Складна за складом.
Через те, що мертві тканини пробки не можуть розтягуватися при зростанні стебла в товщину на ретидом утворюються тріщини на мертвих ділянках.
1) захисна (збільшують механічну міцність, стійкість до пожеж тощо);
2) терморегуляційна (запобігає рослині від різких перепадів t o).
МЕХАНІЧНІ ТКАНИНИ
1. Визначення
2. Осн. типи хутра. тканин та їх ф-ції.
3. Значення для людини.
4. Розподіл хутра. тканин у тілі рослин.
Механічні тканини
а) коленхіма - жива (кутова, пластинчаста, пухка)
б) склеренхіма – мертва (волокна, склереїди)
1. Будь-яка рослина потребує опори для збереження своєї цілісності. У рослин чітко простежується еволюція механічних пристроїв при адаптації до життя у різних середовищах. У первинноводних водоростей, що мешкали в щільній та інертній воді хутро. міцність досягається завдяки клітинним целюлозним оболонкам (тобто тільки на клітинному рівні). При виході на сушу у розріджене середовище цього стає недостатньо і у дрібних наземних рослин стійкість досягається завдяки клітинним оболонкам та тургору клітин ( клітинний рівень) . При збільшенні лінійних розмірів міцності клітинних оболонок та тургору стає недостатньо, і виникають особливі механічні тканини, що забезпечують опору рослині (тканинний рівень). Надалі при адаптивній еволюції рослини все краще пристосовуються до повітряно-грунтового середовища та хутра. тканини по-різному і більш оптимально розташовуються в різних органах(Тобто міцність забезпечується на органному та організмовому рівні організації).
Т.о., механічні тканини- Відіграють роль опорної системи і надають міцність рослинам.
Основні функції:
1) опорна(утворюють внутрішній скелет органів рослини)
2) захисна(забезпечують хутр. міцність та стійкість до хутр. впливів)
Виконують функції у поєднанні з іншими тканинами, отже відіграють роль арматуриі часто називаються арматурними.
Механічні (арматурні) тканини- Спеціалізовані тканини, у поєднанні з іншими утворюють арматуру органів і всього тіла рослини.
Виділяють 2 основних типи хутра. тканин: коленхіму та склеренхіму.
1. Колленхіма( "colla" - клей) - складається з живих прозенхімних клітин із тупими та скошеними кінцями.
Характерно:
1. Нерівномірно потовщені оболонки
2. Немає межі між первинною та вторинною оболонками, і вони не здерев'янюють.
3. Це живі клітини, що мають хлоропласти.
Отже,
4. Можуть вести фотосинтез (тому коленхіма з хлоропластами часто називається
хлоренхімуі знаходиться безпосередньо відразу під епідермою).
5. Виконують свої функції лише у стані тургору. Якщо її клітини втрачають воду, то рослини в'януть.
Утворюються в молодих пагонах, у рості листя безпосередньо з апікальних меристем, отже, якби клітини колленхимы були мертвими, з вторинними рівномірними потовщеннями і одревенням, всі вони могли б розтягуватися і зростати шляхом розтягування за іншими тканинами, отже не виконували свої функції. Тому коленхіма – тканина молодих рослин.
Види коленхіми.
Куткова к. –потовщені по кутах частини оболонок 3-5 клітин зливаються і утворюють 3-5-кутники;
Пухка до.- з міжклітинниками між потовщеними ділянками оболонок, що злилися (у водних рослин);
Пластинчаста до.- Потовщені частини оболонок розташовані паралельними шарами.
Колленхіма розвинена в основному у дводольних рослин.
Склеренхіма(грец. "skleros" - твердий) - різновид хутро. тканини, що забезпечує міцність органів та всього тіла рослини. Зустрічається у всіх квіткових (і в однодольних, і дводольних).
Характерно:
1) Клітини часто з рівномірно потовщеними вторинними оболонками, які найчастіше здерев'янюють (мають велику міцність до роздавлювання, але вище крихкість).
2) Мертві клітини. Протопласт, зазвичай, відмирає після формування оболонок.
Тобто. основні функції виконують мертві клітини.
Склеренхімапредставлена волокнамиі склереїдами.
Волокна- Довгі прозенхімні, найчастіше одревеснілі клітини, загострені на кінцях з товстими стінками і вузькою порожниною. Часу мало.
(Винятки, неодревеснелі луб'яні волокна ряду рослин, наприклад, льону).
Волокнабувають: 1) луб'яні(входять до складу лубу-флоеми) (довші і тонкі) і 2) дерев'яні (волокна лібриформа)(Входять до складу деревини).
Можуть бути й інші волокна, які входять до складу інших тканин та розташовуватись групами або поодиноко.
Склереїди– клітини склеренхіми, які не мають форми волокон.
1. – округлі – кам'янисті клітини – брахісклереїди.
2. - гіллясті - астросклереїди і т.д.
Як і волокна утворюють або суцільні групи, шари (у шкаралупі горіхів, кісточці вишні, сливи, персика, абрикоса), або розташовуються поодинці або групами з кількох клітин ідіобластів(Напр., Ідіобласти в м'якоті плодів груші, айви).
Ідіобласти(грец. idios – особливий, blastos – зародок) – клітини тканин, розташовані поодиноко серед масивів інших тканин (характерні для механічних і видільних тканин).
Значення в людини.
Насамперед, волокна
1) Дерев'яні волокна, що утворюють деревину --- будматеріали, папір і т.п.
2) Луб'яні волокна - луб (стопті, дранка і т.п.)
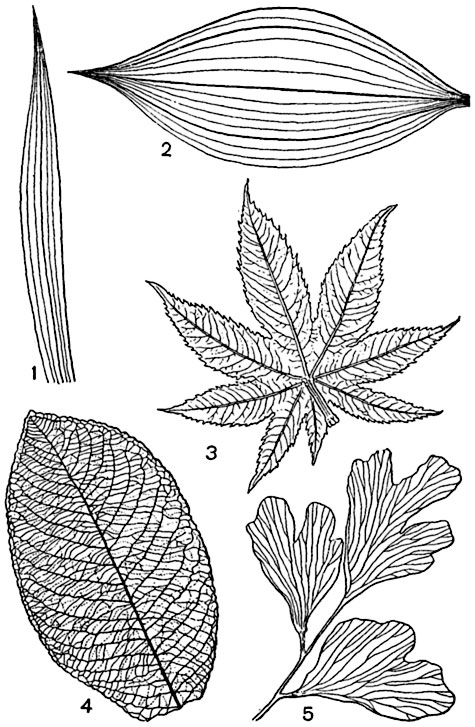






Устячка- це отвори в епідермісі, якими відбувається газообмін. Вони в основному на листі, але є також і на стеблах. Кожне продихання оточене двома замикаючими клітинами, які на відміну від звичайних епідермальних клітин містять хлоропласти. Замикаючі клітини контролюють величину отвору продихи за рахунок зміни своєї тургесцентності. Зовнішній виглядпродихів і замикаючих клітин добре видно на мікрофотографіях, отриманих за допомогою скануючого електронного мікроскопа.
У статті ми вже говорили, як виглядають клітини епідермісу, замикаючі клітини та продихиякщо їх розглядати зверху у світловому мікроскопі. На малюнку наведено схематичне зображення продихів у розрізі. Видно, що стінки замикаючих клітин нерівномірно потовщені: стінка, яка ближче до отвору продиху, звана вентральної, товщі, ніж протилежна, звана дорсальною. Крім того, целюлозні мікрофібрили у стінці орієнтовані таким чином, що вентральна стінка менш еластична, ніж дорсальна. Деякі мікрофібрили утворюють як би обручі навколо клітин, що замикають, схожих на сардельки.
Ці обручі не еластичні, і у міру заповнення клітиниводою,т. е. зростання її тургору, вони не дають збільшуватися її діаметру, дозволяючи розтягуватися тільки в довжину. Але оскільки замикаючі клітини з'єднані своїми кінцями, а тонкі дорсальні стінки розтягуються легше, ніж товсті вентральні, клітини набувають напівкруглої форми. В результаті між двома сусідніми замикаючими клітинами з'являється проміжок, званий устьичною щілиною. Такий же ефект спостерігається, якщо надувати два скріплених кінцями довгастих повітряних кульки, наклеївши вздовж їхніх сторін липку стрічку (імітація нерозтяжної вентральної стінки). Для повноти картини можна нещільно обмотати їх такою ж стрічкою спіраллю, імітувавши целюлозні обручі.
Коли замикаючі клітинивтрачають воду та тургор, устьична щілина закривається. Як відбувається зміна тургесцентності клітин, поки що не ясно.
Відповідно до класичної, так званої « цукро-крохмальной» гіпотезі, у світлий час доби в клітинах, що замикають, підвищується концентрація водорозчинних Сахарів, а отже, осмотичний потенціал їх стає більш негативним, що стимулює надходження в них води шляхом осмосу. Однак нікому ще не вдалося показати, що в замикаючих клітинах накопичується достатня кількість цукру, щоб викликати зміни осмотичного потенціалу, що спостерігаються.
Нещодавно було встановлено, що вдень на світлі в клітинах, що замикають, інтенсивно накопичуються катіони каліюі супутні їм аніони: вони й відіграють роль, що відводилася раніше цукру. Досі неясно, чи врівноважуються при цьому їхні заряди. У деяких вивчених рослин відзначалося накопичення світла великої кількостіаніонів органічних кислот, зокрема малата Разом з тим розміри крохмальних зерен, що з'являються в темряві в хлоропластах клітин, що замикають, зменшуються. Справа в тому, що крохмаль на світлі (необхідні сині промені спектра) перетворюється на малат, можливо, за наступною схемою:
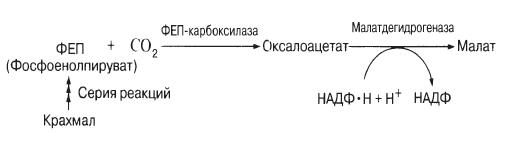
У деяких видів, наприклад у цибулі, крохмалю в клітинах, що замикають, немає. Тому при розкритих продихахмалат не накопичується, а катіони, мабуть, поглинаються разом із неорганічними іонами типу хлорид-іонів.
У темряві калій (К+) виходить із замикаючих клітин навколишні епідермальні клітини. В результаті водний потенціал клітин, що замикають, зростає, і вода з них спрямовується туди, де він нижче. Тургор замикаючих клітин падає, вони змінюють форму, і устьична щілина закривається.
Деякі питання поки щозалишаються без відповіді. Наприклад, чому на ceeiy до замикаючих клітин надходить калій? Яка роль хлоропластів, окрім накопичення крохмалю? Можливо, калій надходить усередину завдяки «включенню» АТФ-ази, локалізованої у плазмалемі. Судячи з деяких даних, цей фермент активується синім світлом. Можливо, АТФаза потрібна для відкачування з клітини протонів (Н+), а катіони калію рухаються в клітину для врівноваження заряду (аналогічний насос, що обговорюється в розд. 13.8.4 працює у флоемі). Справді, як і передбачає ця гіпотеза, всередині клітин рН, що замикають, на світлі падає. У 1979 р. було показано, що у хлоропластах замикаючих клітин кінських бобів (Vtcia faba) немає ферментів циклу Кальвіна, а тилакоїдна система розвинена слабко, хоча хлорофіл там є. Отже, звичайний Сз-фотосинтез не працює, крохмалю в такий спосіб не утворюється. Це, ймовірно, пояснює, чому крохмаль утворюється не вдень, як у звичайних клітинах, що фотосинтезують, а вночі.
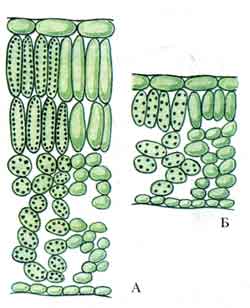
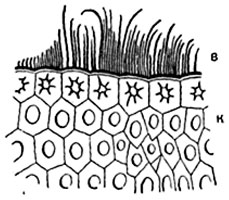
Шкірка, або епідерміс. Шкірка, або епідерміс, є первинну тканинулистя та стебел. Шкірка складається з щільно зімкнутих клітин, що мають у плані у більшості рослин більш-менш звивисті контури (рис. 86). На поперечних розрізах органу клітини шкірки зазвичай мають чотирикутні або п'ятикутні контури. Іноді зовнішня та внутрішня стінки злегка опуклі. Клітини шкірки органів, витягнутих у довжину (листових черешків, листових пластинок лінійного та ланцетного обрису, стебел), зазвичай сильно подовжені в напрямку, паралельному до поздовжньої осі органу. У деяких рослин, наприклад, у багатьох злаків, шкірка складається з клітин декількох типів (рис. 87).
Оболонка епідермальних клітин потовщується зазвичай нерівномірно: у кожній клітині найбільш товста зовнішня стінка, бічні стінки дещо тонші, внутрішня - порівняно тонка. Зустрічаються винятки із цього правила. Бічні та внутрішні стінки клітин зазвичай мають пори (рис. 87), зовнішня ж стінка буває забезпечена ними в вельми поодиноких випадках. Оболонка клітин складається переважно з целюлози. У багатьох рослин (особливо у злаків, осокових, хвощів) зовнішня стінка шкірки просочена кремнеземом; у деяких рослин у зовнішній стінці, рідше у бічних (у драцен) і навіть у внутрішніх відкладаються дрібні кристали або зернятка щавлевокислого кальцію.
Поверхня шкірки покрита плівкою - кутикулою (надшкіркою), що складається з кутина. Кутикула дуже тонка чи зовсім відсутня на поверхні конусів наростання. Часто кутикула має гладку поверхню, в інших випадках утворює виступи у вигляді горбків, перекладин, прямих або хвилястих ребер і т. д. Ці виступи складаються націло з
Мал. 86. Ділянки шкірки листка жовтця повзучого ( Ranunculus repens):
1 - З верхнього боку листа; 2 - З нижньої його сторони.
кутикули або є виступами целюлозної стінки, покритими кутикулою.
На поперечних зрізах шкірки іноді видно, що кутикула пронизує частину товщі стінок. Кутикула сильно заломлює світло і тому добре помітна на зрізах. Ізольована кутикула зазвичай представляє безбарвну суцільну безструктурну плівку. У деяких рослин (наприклад, у золотого дерева Aucuba) кутикула складається з платівок, кожна з яких належить одній епідермальній клітині; над прикордонними ділянками клітин шкірки кутикули немає або дуже тонка.
Потужність кутикули залежить від виду та віку рослини та умов проживання (рис.88); на поверхні стебел з багаторічною шкіркою (у деяких видів калини, клена) кутикула розривається і підновлюється зсередини шляхом утворення нових її нашарувань. Кутикула листя деяких осокових піддається окремінню.
У багатьох рослин у зовнішніх стінках клітин шкірки, іноді в бічних, ще рідше у внутрішніх, утворюються кутикулярні шари- прошарки в целюлозній клітинної оболонки, Що містять кутин (рис. 76) Кутикула та кутикулярні шари нерідко просочені воском; при підігріванні препаратів у воді під покривним склом він виплавляється у вигляді крапель. У деяких рослин віск покриває кутикулу у вигляді дрібних зерен, розташованих рівномірним шаром (на листі та стеблах багатьох злаків, лілейних, наприклад тюльпанів), тонких паличок, часто вигнутих і на кінці закручених (на стеблах деяких злаків, цукрової тростини, рис. 89) , суцільних скорин, тонких - 1μ завтовшки (на листі туй, молодил).
Восковий наліт, як і кутикула, знижує транспірацію
![]()
1 - замикаючі клітини продихів; 2 - побічні клітини; 3 - Клітини епідермісу з численними порами; у порожнині клітин - постінна протоплазма та ядро, у деяких із клітин не замальовані.
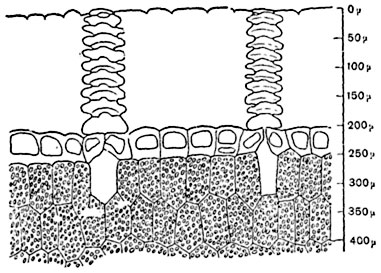
Мал. 88. Частина поперечного розрізу через листоподібний горбок кактуса Ariocarpus retusus; надзвичайно товста кутикула пронизана фігурними ходами над продихами. Під шкіркою – зелена паренхіма.
органів. Віск робить поверхню органів незмочується: з них швидко стікає вода, чим попереджається капілярне закупорювання водою продихів і заселення поверхні рослин дрібними епіфітами.
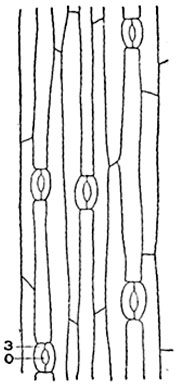
до- Шкірка: в- воскові палички.
Тому зрозуміло, чому стебла та листя багатьох підводних рослин позбавлені кутикули.
Клітини шкірки містять тонкий постінний шар протоплазми з ядром та пластидами та велику центральну вакуолю. Зазвичай із пластид тут є лейкопласти.
У багатьох рослин у вакуолях клітин шкірки – нерідко лише на нижній стороні листка – міститься антоціан.
У вакуолях епідермальних клітин нерідко зустрічаються дубильні речовини, солі щавлевої кислоти у вигляді розчинів (щавлів, кислиць) або кристалів; рідше тут є алкалоїди, отруйні глюкозиди, гіпс (у каперсів - у вигляді кристалів), кальцієві солі яблучної та винно-кам'яної кислот (у винограду).
Хлоропласти та хромопласти в епідермісі квіткових рослин зустрічаються дуже рідко, тільки у деяких тіньових та водних рослин.
Устячка. Для здійснення газообміну між внутрішніми тканинамирослин і довкіллям і для транспірації (випаровування води) в шкірці є продихи.
Устьиці складається з двох спеціалізованих замикаючих клітині щілинного отвору між ними - устьичної щілини. При розгляданні шматочка шкірки, покладеного плазом на предметне скло, продихання зазвичай видно як пара бобовидних або напівмісячних клітин, звернених один до одного увігнутими сторонами (рис. 90-92). Оболонка кожної замикаючої клітини на внутрішній сторонісильно і нерівномірно потовщена, на протилежному, зовнішньому, боці - тонка.
На поперечному розрізі, проведеному через середину устьичної щілини перпендикулярно до неї (рис. 91), видно вирости стінок замикаючих клітин продихів, звані відрогами або дзьобиками.
Щілина продихи має складне обрис: проміжок між зовнішніми відрогами веде напередодні, або передній дворик продиха. Далі йде найвужча частина устьичної щілини. центральна щілина устячка, що знаходиться між тонкими ділянками внутрішніх стін. Центральна щілина розширюється далі у задній дворик. Отвір між відрогами нижніх стінок клітин, що замикають.
Мал. 90. Шкірка листа гіацинту ( Hyacinthus orientalis) з продихами, у плані:
з- замикаючі клітини, о- щілина устячка.
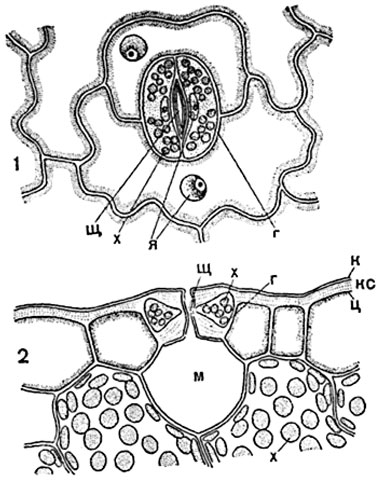
Мал. 91. Устячки з навколишніми клітинами епідермісу (листя чебрецю Thymus):
1 - у плані; 2 - у розрізі; щ- центральна щілина устячка; а- тонка стінка замикаючої клітини продихи, звернена до сусідніх клітин шкірки; у замикаючих клітинах видно ядро ( я) з ядерцем та хлорофілові зерна ( x); до- кутикула; м- міжклітина повітроносна порожнина ("дихальна порожнина").
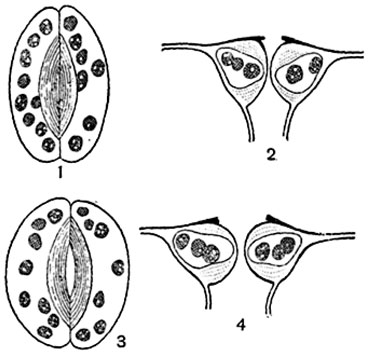
Мал. 92. Устьичний апарат листа буряків ( Beta vulgaris):
1, 3 - продихання в плані; 2, 4 - у розрізі; 1, 2 - продихи закрито; 3, 4 - продихання відкрито.
повідомляє задній дворик з дихальною порожниною (рис. 91, 2 ; 93).
Повертаючись до продиху в плані (рис. 91, 1 ), відзначимо, що щілина устячка в оптичних перерізах, паралельних поверхні епідермісу, має форму вузької двоопуклої лінзи; у плані зазвичай видно обрис щілини лише на рівні зовнішнього отвори і лише на рівні середини центральної щілини. Зовнішні стінки, а в деяких рослин також і внутрішні покриті кутикулою; облямівки двориків іноді складаються суцільно з кутикули. Функціонуючі замикаючі клітки живі; вони містять постінний шар протоплазми з ядром та з хлорофіловими зернами.
Зазвичай продихи розподіляються на поверхні листа або стебла поодинці і більш менш рівномірно (рис. 90). У деяких рослин (наприклад, у злаків) вони розташовуються поздовжніми рядами, а в інших (у олеандра, селезінки) - групами.
Утворення замикаючих клітин продихів відбувається при нерівномірному поділі меристематичних клітин епідермісу, в результаті чого виникають ініціальні клітини продихів. У найпростіших випадках ініціальна клітина стає клітиною продиху. Клітка, що виробляє, утворює шляхом поділу пару замикаючих клітин продихи; розщеплення перегородки з-поміж них веде до утворення устьичной щілини. У деяких рослин ініціальна клітина утворює шляхом поділу цілі групи клітин - пару замикаючих і дві або кілька побічних клітин
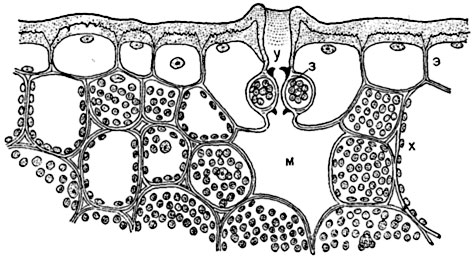
Мал. 93. Частина поперечного розрізу листа Aloe distichaз продихом, зануреним у тканину листа:
е- Шкірка з товстими зовнішніми стінками, покритими потужною кутикулою; у- Порожнину над продихом; з- замикаючі клітини з кутикулярними виступами (позначеними чорним кольором); м- повітроносний міжклітинник ("дихальна порожнина"); х- Клітини хлорофілоносної паренхіми.
продихи; останні відрізняються від інших клітин шкірки меншою величиною та своєрідними обрисами (рис. 94).
Устьичний апарат регулює газообмін рослини. Замикаючі клітини можуть зміщуватися, змінювати об'єм і форму, внаслідок чого змінюється і обрис устьичної щілини: вона може бути більш-менш широко відкрита або зовсім закрита (рис. 92). При відкриванні та закриванні продихів відбуваються зміни живого вмісту замикаючих клітин. При відкритій щілині продихів протоплазма в них більш в'язка, ніж при закритій. Ядро змінює форму: у кормових бобів при тривало відкритому продиху воно стає веретеновидним, при закритому - округло-лопатевим, у жоржини - навпаки (рис. 95).
Відкривання та закривання продихів обумовлюється тургорними явищами. Зміни тургора клітин, що замикають, можуть бути безпосереднім наслідком підведення листа або підвищення в ньому вмісту води.
При підвищенні тургора клітин, що замикають, об'єм їх клітинної порожнини збільшується, в результаті чого стінки віддаляються одна від одної і щілина відкривається. При зниженні тургора замикаючих клітин відбуваються зміни зворотного роду: стінки зближуються до зіткнення і щілину продиху замикається.
Зміни тургору в замикаючих клітинах регулюються перетворенням в них крохмалю хлоропластів на цукор, і назад - цукру на крохмаль: при оцукрювання крохмалю в замикаючих клітинах підвищується концентрація клітинного соку і збільшується їхня сила, що смокче. В результаті клітини, що замикають, насмоктують воду (частково за рахунок сусідніх клітин), що викликає зміну їх обсягу і розкриття устьичної щілини (рис. 96). Перетворення цукру замикаючих клітин на крохмаль тягне аналогічним чином закривання щілини продиху.
Непряма участь у закриванні і відкриванні продихів можуть брати не тільки клітини, сусідні з замикаючими, але і більш далекі побічні клітини продихів і рядові клітини шкірки. При тривалому дощі всі клітини шкірки насичуються водою, тургор їх сильно підвищується і замикаючі клітини продихів здавлюються; в результаті устьичні щілини пасивно замикаються. Через деякий час після припинення дощу нормальна робота продихів відновлюється.
Тургорна теорія дії продихів, висунута Молем в 1856 р., піддалася доповненням з боку ряду вчених, що вказують на можливе значеннязмін проникності протоплазми клітин, що замикають.
У пристрої продихів зустрічається велика різноманітність. Форма щілини може бути значно спрощена або ускладнена порівняно з вищеописаною типовою. Найбільш спрощена вона у рослин з листям, що плаває на воді (біле латаття): щілина має вигляд воронки, широким кінцем що з'єднується з дихальної порожниною, а вузьким переходить у зовнішній отвір; ступінь розкриття щілини продихи залежить від стану цього отвору.
У рослин з листям, розташованим більш-менш горизонтально, особливо ж у дерев і у трав тінистих місцепроживання, продихи розташовані переважно або виключно на їх нижній стороні.
У багатьох рослин, переважно у приурочених до сухих, сонячних і вітряних місцеперебування, продихи занурені в глиб органу за рахунок сильно кутинізованих виступів сусідніх клітин (рис. 93). Іноді продихи розташовуються цілими групами в глибині ямочок (як у олеандра). У деяких видів австралійських бенксій ( Banksia) на дні і навколо ямок з продихами є довгі білі волоски; утворюються, таким чином, притулки із затишшям від вітру та затіненням від сонця.

А- три виробляючі клітини продихів ( 1, 1, 1 ) вже відокремилися перегородками від ініціальних клітин; у сусідніх клітинах відбувається розподіл ядра ( 2, 2 ), що передує утворенню побічної клітини; Б- при кожній з виробляючих клітин трьох продихів утворилися дві побічні клітини ( 3 ); У- у трьох виробляючих клітинах продихів відбувається розподіл ядра ( 4, 4 ), що передує освіті замикаючих клітин; Г- Клітка, що виробляє, поділилася на дві клітини, що диференціюються в пару замикаючих клітин продихи ( 5, 5 ), з устьичною щілиною між ними; 6, 6 - побічні клітини; Д- сформований устьичний апарат, що складається з двох замикаючих клітин ( 5, 5 ) та двох побічних клітин ( 6, 6 ).
Волоски (трихоми). У більшості вищих рослин деякі або багато клітин шкірки утворюють вирости, так звані волоски, що мають різну форму, сосочків, горбків, щетинок, кущиків, зірчастих лусочок і т. д. У найпростішому випадку волосинка представляє сосочок - короткий округло-конічний виріст клітини шкірки. Сосочками ж

а- продихи відкрито, у замикаючих клітинах дуже дрібні крохмальні зерна, ядро округло-амебоїдного обрису, з вакуолей у центрі; б- продихи закрито, у замикаючих клітинах - порівняно великі зерна крохмалю, клітинне ядроверетеноподібно, з великими ядерцями в центрі.
є волоски майже всіх типів у ранній стадіїїх утворення. Зустрічаються волоски, що не відокремлюються перегородкою від клітини шкірки, що виробила їх, як наприклад щетинки багатьох бурачникових. У більшості випадків волосок відокремлюється перегородкою від епідермальної клітини, що його зробила, - "материнської клітини волоска". Волок, що відчленовується, рідко залишається одноклітинним; частіше він стає багатоклітинним. Багатоклітинні волоски дуже різноманітні. Серед них розрізняють 1) лінійні, що складаються з клітин, розташованих у ряд (листя картоплі); 2) кущисто-гіллясті (коров'яки, рис. 97, 6 ); 3) лускаті і зірчасто-лускаті ( Elaeagnus, Мал. 97, 7, 8 ) і 4) масивні, що представляють як би пучок ниткоподібних волосків, що спаялися (волоски багатьох губоцвітих). Вміст волосків деяких категорій у дорослій стадії руйнується, порожнини їх зазвичай заповнюються повітрям. Такі волоски, що мають зазвичай білий, або сірий, або жовтуватий колір, можна вважати одним із пристосувань, що поміряють дію на лист або стебло гарячих променів сонця і послаблюють висушує дію вітрів.
Деякі волоски можуть бути рослиною захистом від поїдання тваринами; так, тісно розташовані жорсткі, гострі щетинки і повстяний покрив оберігають листя і стебла від об'їдання слимаками і частково від поїдання травоїдними ссавцями.
Пекучі волоски. Гаряча волосина кропиви - велика жива клітказ міхуровою основою, що сидить у чашоподібній багатоклітинній "підставці". Волосок представляє в мініатюрі ніби порожню голку шприца і закінчується косо поставленою голівкою. Так як оболонка волоска, багата вапном і кремнеземом, дуже ламка і до того ж у місці переходу "голки" в "головку" дуже тонка, то при найменшому дотику тварини або людини до головки остання відламується, гострий край "голки" встромляється в шкіру, і клітинний сікволосинка, що містить пекучі речовини, впорскується в неї (рис. 97, 9-12 ).
Дотик руки до листя деяких кропив тропічної флори (наприклад, Urtica urentissimaІндонезії) викликає хворобливе чи навіть небезпечне життя запалення.
До покривних тканин відносяться ще перидерма та кірка (опис їх див. на стор. 173-175).
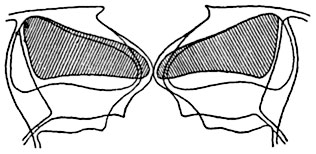
Мал. 96. Схематизований оптичний розріз продихи зимівника ( Heiieborus sp.) у закритому та відкритому стані.
Зовнішні обриси та клітинна порожнина замикаючих клітин показані при закритій щілині продиху і при відкритій щілині. Клітинна порожнина обох замикаючих клітин при закритому продиху заштрихована.