De første organismene på jorden var encellede. Hele kroppen til organismen besto av bare én celle. Senere dukket opp flercellede organismer deres kropper besto imidlertid av identiske celler. Og først da begynte organismer ikke bare å bestå av identiske, men også av forskjellige celler. Identiske celler i samme organisme danner vev. I komplekse organismer det kan være en rekke forskjellige vev, så det er en rekke forskjellige celler.
Ved sammensetningen av plantevev kan du bestemme hvilken gruppe de tilhører - alger, moser, bregner eller frøplanter.
Vev inneholder celler som er like i struktur og funksjoner. Vev kan variere i tettheten av celler; i noen kan de være plassert svært nær hverandre og danne rader med celler, i andre kan de ligge som ønsket, ikke tett til hverandre, løst. Mellomrommene mellom celler kalles intercellulære rom, eller intercellulære rom. Vevet inkluderer også intercellulære rom.
Celler pedagogisk stoff dele seg gjennom plantens levetid. Cellene i utdanningsvevet ligger tett til hverandre; de deler seg danner de nye celler og sikrer dermed plantens vekst ikke bare i lengde, men også i tykkelse. I tillegg er celler av planteopplæringsvev i stand til å transformere seg til celler fra andre vev.
Ansvarlig for oppretting og akkumulering av stoffer hovedstoff. Det er i dette vevet klorofyll finnes, takket være det uorganiske stoffer organisk syntetiseres. Hovedvevet finnes hovedsakelig i bladene til planter.
Imidlertid er de viktigste vevene der reserven oppstår næringsstoffer, finnes i frø, modifiserte røtter (potetknoll), stengler (løk) etc.
Utfører en beskyttende funksjon dekkvev . Den beskytter alle planteorganer fra utsiden mot uttørking, skade og overoppheting. I huden på blader og skudd er cellene i integumentært vev tett lukket sammen; de har en gjennomsiktig cellevegg som lar lys passere gjennom. I røtter og stilker kan dekkvevet bli suberisert, og bli til en kork.
Takk til ledende vev stoffer kan bevege seg gjennom hele planten. Stoffer beveger seg i vandige løsninger som strømmer gjennom cellene i ledende vev. U høyere planter ledende vev består av kar, trakeider og silrør. Ledende vev har porer og åpninger som tillater bevegelse av stoffer mellom celler.
Ledende vev er et forgrenet nettverk som forbinder alle planteorganer. Dermed er alle deler av anlegget kombinert til et enkelt system.
Mekanisk stoff lar planter tåle ulike belastninger, for eksempel vind. Mekaniske vevsceller har veldig sterke cellevegger.
Eksistensen av forskjellig vev skyldes det faktum at planteceller på land må prestere ulike funksjoner. Roten er i jorda og absorberer vannløsning, holder også planten i jorden. Bladene utsettes for lys og er ansvarlige for syntesen av organiske stoffer. Stengelen forbinder forskjellige deler av planten med hverandre.
Plantevev er ganske forskjellige. jeg lurer på hva morfologiske trekk av hver slik struktur direkte avhenge av funksjonen den utfører. Det er vanlig å skille mellom flere typer:
- pedagogisk;
- integumentær;
- mekanisk;
- ledende;
- grunnleggende.
Hver struktur har visse funksjoner, som vil bli diskutert nedenfor.
Pedagogisk plantevev
Utdanningsvev kalles også meristemer. Denne strukturen består av små, mangefasetterte celler med tynne vegger. De er tett lukket sammen. Under et mikroskop kan du se at de har en stor kjerne og mange små vakuoler. Et trekk ved dette vevet er cellenes evne til å dele seg konstant. Det er dette som sikrer konstant vekst planter. Det er vanlig å skille mellom følgende typer:
- Primær meristem- hos en voksen plante er dette vevet bevart i spissen av skuddene og spissen av røttene. Det er takket være det at den primære veksten av planten i lengde oppstår.
- Sekundært meristem - representert av kambium og fellogen. Disse vevene gir sekundær vekst av stilken og roten i diameter. Basert på deres plassering skilles apikale, laterale og interkalære sekundære meristemer.
Plante integumentært vev
Dekkvevet er plassert på overflaten av plantekroppen. Hovedfunksjonen er beskyttelse. Slike strukturer er ansvarlige for plantens motstand mot mekanisk påvirkning, beskytte mot plutselige temperatursvingninger og overdreven fordampning av fuktighet, beskytte mot penetrasjon av patogene mikroorganismer. Integumentære elementer er vanligvis delt inn i tre hovedgrupper:
- Epidermis (huden) er et primært vev som består av små, gjennomsiktige og tett sammenkoblede celler. Vanligvis dekker denne typen vev overflaten av blader og unge skudd. Det epidermale laget av blader inkluderer også stomata - formasjoner som er ansvarlige for prosessene med gassutveksling og transpirasjon.
- Periderm er et sekundært integumentært vev som ligger på overflaten av stilken og roten. Den består av pheloggen og er et dødt lag med celler, hvis vegger er impregnert med det vanntette stoffet suberin.
- Bark er et vev som er karakteristisk for trær og noen busker. Dette laget av integumentært vev er den ytre delen av korken.
Ledende plantevev
Hovedfunksjonen til denne gruppen av vev er transport av vann og mineraler gjennom hele plantekroppen. Det er vanlig å skille mellom følgende typer ledende elementer:
- Xylem - sikrer bevegelse av vann med oppløst mineraler fra rotsystemet til den overjordiske delen av planten. Den består av spesielle kar, den såkalte luftrøret og trakeider.
- Floem er vevet som tillater nedadgående strømning. Gjennom silrør føres alle organiske næringsstoffer som syntetiseres av bladene til resten av planteorganene, inkludert rotsystemet.
planter: parenkym
Dette vevet består av små levende celler med tynne vegger. Det er dette som danner grunnlaget for alle organer. Disse inkluderer:
- Assimileringsvev - cellene deres inneholder et stort antall kloroplaster og er ansvarlige for dannelsen av organiske stoffer. Mest av Disse vevene finnes i bladene.
- Lagringsvev - avsatt i celler nyttig materiale. Dette vevet er konsentrert i frukt, røtter og frø.
- Akviferøst vev tjener til å akkumulere og bevare vann. Disse vevene er karakteristiske for planter som lever i varmt og tørt klima, for eksempel kaktus.
- Luftbærende vev - slike vev har enorme intercellulære hulrom som er fylt med luft. Aerenchyma er karakteristisk for myr og
Mekanisk plantevev
Ansvarlig for å skape en sterk ramme. De opprettholder plantens form, noe som gjør den mer motstandsdyktig mot mekanisk påvirkning. Dette vevet består av celler med tykke membraner. De er sterkest utviklet i stammen til planten.
Tekstil - en gruppe celler og ikke-cellulære strukturer som ligner i opprinnelse og struktur, som danner et strukturelt-funksjonelt kompleks og utfører de samme funksjonene.
Klassifisering tar vanligvis hensyn til funksjon, struktur, opprinnelse og plassering av vev. Det er seks hovedgrupper (systemer) av vev:
· System av meristematisk (pedagogisk) vev:
Ø apikale meristem;
Ø lateral meristem;
Ø intercalary meristem;
Ø sårmeristem.
· System av integumentært (kant)vev:
Ø epidermis;
Ø periderm (plugg);
Ø skorpe (rhytidome);
Ø epiblema.
· Hovedstoffsystem:
Ø assimilering (klorofyllbærende) parenkym (klorenkym);
Ø lagringsparenkym.
· Mekanisk (forsterkning) stoffsystem:
Ø collenchyma;
Ø sklerenkym.
· Ledende vevssystem (komplekst vev basert på ledende elementer):
Ø xylem;
Ø Phloem.
· System av utskillende (sekretorisk) vev:
Ø eksterne sekretoriske strukturer;
Ø indre sekretoriske strukturer.
4.1. Utdanningsvev (meristem)
Planter har ubegrenset vekst på grunn av tilstedeværelsen av pedagogisk vev. De er dannet av udifferensierte (parenkymale) runde eller polyedrale celler uten intercellulære mellomrom. Celleveggene er tynne, lett utvidbare, cytoplasmaet er tykt, tyktflytende, kjernen er stor, okkuperer sentral posisjon. Celler av pedagogisk vev er i stand til å dele seg raskt, så de inneholder mange ribosomer og mitokondrier.
Etter opprinnelse skiller de:
· Primære merister - meristem av embryoet. De bestemmer utviklingen av frøplanten og den primære veksten av organer.
· Sekundære merister . De oppstår på grunnlag av de primære. De sikrer vekst av organer hovedsakelig i bredden.
Etter plassering er det:
· Apikale (apikale) meristemer . Plassert i endene av hoved- og sideaksene til stammen og roten, bestemmer de hovedsakelig lengdeveksten til orgelet.
· Laterale (laterale) meristemer . De oppstår på grunn av aktiviteten til primære meristemer. Som regel forårsaker de fortykkelse av aksiale organer.
· Interkalære merister . Områder med intensivt delende celler, vanligvis lokalisert ved skuddnoder eller ved basene bladblader. De er rester av det apikale meristem. Når veksten av internoder eller blader stopper, blir det interkalære meristemet til permanente vev, det vil si at deres aktivitet er kortvarig. Men noen ganger kan disse meristemene fungere i ganske lang tid (for eksempel ved bunnen av internodene til kjerringrokk og frokostblandinger).
· Sår (traumatisk) meristemer . De vises på steder med mekanisk ødeleggelse av vev fra levende celler i forskjellige parenkymale vev. De sikrer tilheling av såret og blokkerer tilgangen til patogener.
4.2. Integumentært vev
Integumentært vev er permanente formasjoner. Etter å ha oppstått, deler cellene i disse vevene seg ikke lenger.
Som oftest, integumentært vev kalt vev som dekker kroppen til en plante og samhandler med eksternt miljø. De beskytter indre stoffer fra handling ugunstige faktorer miljø, regulere gassutveksling og transpirasjon.
Selve integumentvevet inkluderer:
1. Epidermis
Hovedfunksjonene er beskyttelse av unge organer mot uttørking, mekanisk beskyttelse og gassutveksling, noen ganger sekretorisk - cellene fungerer som en beholder for sekret. Dette er det primære integumentære vevet. Oftest i epidermis er det hoved dekkceller, stomatale formasjoner og ulike utvekster ( trichomes). De viktigste integumentære cellene er vanligvis representert av et enkelt lag med tettpakkede celler. Celleveggene er vanligvis kronglete, ytterveggene er tykkere enn de andre.Dette er levende celler med store vakuoler, cytoplasmaet ser ut som et tynt vegglag. ER- og Golgi-apparatet er vanligvis godt utviklet.
Dekkvevet til rotabsorpsjonssonen kalles epibleme.
For gassutveksling og transpirasjon inneholder epidermis spesialopplæring -stomata. De er en gruppe høyt spesialiserte celler. Stomata er en spaltelignende åpning i epidermis, avgrenset av to bønneformede celler. Dette vaktceller. I motsetning til andre epidermale celler inneholder de kloroplaster. Veggene til vaktcellene som vender mot stomatalfissuren er fortykket. De epidermale cellene som omgir vaktcellene kalles sikkerhet eller tilstøtende. Under stomata er gass-luft kammer. Vakt- og sideceller, stomatalfissur og ha-
luftkammerformen stomatal apparat. Stomata er oftest plassert på undersiden av bladet.
2. Cuticle
Den beskyttende funksjonen til epidermis kan forsterkes ved tilstedeværelsen av en kutikula og et voksaktig belegg. Cuticle er en cellefri formasjon. Det er et produkt av protoplastaktivitet og består av et spesielt stoff - kutin og vokslignende stoffer. Vokslignende stoffer kan være en del av skjellaget eller plassert på overflaten. Cuticle og voksaktig belegg finnes på frukt, blader, stilker og blomsterdeler. Kutikulaen og det voksaktige belegget er ugjennomtrengelig for vann og nesten ugjennomtrengelig for gasser.
3. Periderm (kork)
Sekundært integumentært vev bestående av phellems- selve trafikkorkene, fellogen- korkkambium phelloderms- korkparenkym. Den erstatter epidermis, som gradvis dør og avtar. Den dannes hovedsakelig i stengler og røtter. Korken består av vanlige radielle rader med tettsittende celler med suberiserte vegger. Innholdet i cellen dør. Det er ingen intercellulære rom. Kork er ugjennomtrengelig for vann og gasser. Linser dannes i pluggen for gassutveksling og transpirasjon.
4. Ritide, eller skorpe
I de fleste treaktige planter erstattes korken av en skorpe, som noen ganger kalles tertiært integumentært vev. Når en skorpe dannes nytt lag fellogen og periderm legges ned i hovedvevet som ligger dypere enn den første ytre periderm. De nydannede lagene av pluggen skiller seg til periferien av organet, ikke bare peridermen, men også en del av det underliggende parenkymet i cortex. Slik oppstår en tykk flercellet og død formasjon. Siden skorpen ikke kan strekke seg, når stammen tykner, sprekker den og det dannes sprekker.
4.3. Mekaniske (forsterkende) stoffer
Intensivt utviklet i landplanter. Hovedformålet er å forhindre brudd på vev og organer.Stenglene er plassert langsperiferi, i røttene - i sentrum. De består av celler med tykke vegger, ofte lignifiserte.
1. Collenchyma
Utviklet hovedsakelig i stilkene, bladstilkene og bladene til tofrøbladede planter. Som regel finnes det i den perifere delen av organer rett under epidermis eller litt dypere. Dannet av levende, langstrakte celler, som ofte inneholder kloroplaster. Celleveggene er ujevnt fortykket.
2. Sklerenkym
Det viktigste mekaniske vevet til høyere planter. Etter opprinnelse kan den være primær (hvis den ble dannet fra procambium eller fra pericycle) og sekundær (hvis den ble dannet fra kambium). Dannet av celler med jevnt fortykkede, ofte lignifiserte vegger. Protoplasten dør tidlig, og støttefunksjonen utføres av døde celler som kalles fibre. 1. Xylem (tre)
Den består av kar og trakeider som utfører den oppadgående strømmen av vann og mineraler, samt trefibre og treparenkym.
2. Trakeider
Celler forlenget langs organets akse med sterkt skrå endevegger. Penetrering av løsningen fra en tracheide til en annen skjer gjennom porene. Finnes oftere i høyere sporer og gymnospermer.
3. Fartøy
Dannet fra individuelle segmenter som tidligere var celler. Dette er lange mikroskopiske rør. Endeveggene til karsegmentene løses nesten helt opp oggjennomgående hull (perforeringer) vises. Karenes lumen er bredere enn trakeidene. Dette er et mer avansert ledende stoff som strekker seg største utvikling hos angiospermer.
Den består av silelementer, medfølgende følgeceller, bastparenkym og floem (bast) fibre.
4. Silelementer
Den viktigste delen av floem. På grunn av tilstedeværelsen av silelementer gir floem en nedadgående strøm av vann og organisk materiale. Cellene til silelementene har en levende protoplast, gjennom hvilken bevegelsen av vann og organiske stoffer skjer. Protoplaster av naboceller kommuniserer med hverandre gjennom spesielle små hull - perforering. Perforeringer samles i grupper - siktefelt. Det er silceller og silrør.
5. Sil celler
Karakteristisk for høyere spore- og gymnospermplanter. De er svært langstrakte celler med spisse ender. Silfelt ligger spredt langs sideveggene. Modne celler beholder sin kjerne. Silceller mangler medfølgende celler.
6. Silrør
Karakteristisk for angiospermer (fig. 7). Perforeringer samles i grupper og form silplater, som er plassert i enden av cellene. I modne segmenter av silrør er kjernen fraværende, den sentrale vakuolen løses opp, cellesaften kombineres med cytoplasmaet. Cellen forblir imidlertid i live.Protoplasten har form av lange tråder som passerer gjennom perforeringer fra segment til segment.
Ved siden av hvert segment av silrøret er det satellittceller. De deltar i transport av stoffer gjennom silrør.
I planteorganer er xylem og floem hovedsakelig lokalisert i sammensetningen komplekse formasjoner - ledende bunter.
4.5. Hovedstoffer
De danner grunnlaget for organer, fyller mellomrommene mellom andre vev, og gir alle aspekter av intern metabolisme i planter. De kalles parenkymalt eller parenkym. I typiske tilfeller er intercellulære rom godt utviklet.
Mest typisk for blader og grønne assimilerende stengler. Inneholder kloroplaster og utfører funksjonen til fotosyntese. Celler runde eller litt langstrakte oval form. Veggene deres er tynne, aldri lignifisert og noen ganger foldet. Cellene er nesten helt fylt med kloroplaster, bare i midten er det en vakuole. Kjernen og cytoplasmaet inntar en veggposisjon. Delt i søyleformet, eller palisade, Og svampete klorenkym. Kolumnare klorenkymceller er ordnet i ett eller flere lag. Endeveggene vender mot utsiden og innsiden av orgelet. De langsgående veggene er i nær kontakt med hverandre. Cellene til svampete klorenkym er ordnet løst, med store intercellulære rom.
1. Lagringsparenkym
Hovedsakelig utviklet i aksiale organer, reproduktive og vegetative reproduksjonsorganer. Server for å bevare næringsstoffer. Dannet av tynnveggede celler. Det er ingen kloroplaster. Kjernen, cytoplasma og andre organeller inntar først en veggposisjon, og kan deretter forsvinne helt, mens cellene forblir i live. I tørre områder har planter vannlagrende vev. Cellene i slikt vev inneholder mye slim, som hjelper til med å holde på vann.
4.6. Utskillelsesvev
Slipp eller akkumulere ulike stoffer. Cellene i utskillelsesvevet er tynnveggede. Avhengig av arten av de utskilte stoffene, den glatte endoplasmatisk retikulum eller Golgi-apparat. Utskillelsesvev er delt inn i ytre og indre.
Evolusjonært assosiert med integumentært vev. De fremhever annerledes kjemiske substanser spiller spesifikk verdi i livet til planter: noen tiltrekker seg pollinerende insekter, andre er metabolske produkter, etc. Slike stoffer inkluderer:
· nektarier-spesialiserte kjertelutvekster som produserer nektar;
· hydatoder-flercellede formasjoner som skiller ut dråpe-flytende vann og salter oppløst i det;
· osmoforer-spesialiserte celler i epidermis eller spesielle kjertler som skiller ut aromatiske stoffer.
Vev er grupper av celler som er like i opprinnelse, form, struktur og funksjon.
Basert på formen på cellebestanddelene, skilles vev mellom parenkymalt vev, sammensatt av isodiametriske (med omtrent like indre diameter) celler, og prosenkymalt vev, som består av langstrakte celler med forskjellig "lengde" og "bredde" fra hverandre.
Vev er vanligvis klassifisert i henhold til hovedfunksjonen de utfører.
Pedagogiske stoffer, eller merister, sikre kontinuerlig vekst av planter. Meristemceller lang tid beholde evnen til å dele seg uten å bli til permanent vev. Cellene i pedagogisk vev er tynnveggede, fylt med tett cytoplasma, små, og en betydelig del av cellevolumet er okkupert av kjernen. I høyere planter, fra de første stadiene av embryoutvikling, dannes apikale (apikale) meristemer på toppen av stilken og på spissen av roten. Etter hvert som de vokser og forgrener seg, utvikler hvert sideskudd og hver siderot sine egne apikale meristemer. De sørger for at planten vokser i lengde. Laterale meristemer (kambium) kan vises i stengler og røtter. Hos tofrøbladede planter sikrer delingen av kambiumceller veksten av stilker og røtter i tykkelse. Fra permanent parenkymalt vev oppstår noen ganger et sekundært meristem - phellogen (korkkambium), som danner integumentært vev - kork.
Ved bunnen av internodene på stilken og ved bunnen av unge voksende blader er det et interkalært meristem. På slutten av veksten av stilkdelen eller bladet slutter det interkalære meristemet å fungere, og blir til permanent vev.
I assimilasjonsvev fotosyntese skjer. Assimilasjonsvev er hovedsakelig lokalisert i bladet og stilken under epidermis og består av tynnveggede parenkymceller som inneholder kloroplaster.
I lagringsvev reservestoffer deponeres. Disse vevene består av levende parenkymceller. I flerårige planter finnes lagringsvev i stengler, røtter, jordstengler, knoller og løker. Lagringsstoffer kan være karbohydrater, proteiner og fett.
Luftvev (aerenkym) - vev med svært store intercellulære rom, hvis hovedfunksjon er ventilasjon. Intercellulære systemer er koblet til det ytre miljøet gjennom åpninger i integumentære vev (stomata, linser). Gjennom de intercellulære rommene kan oksygen fra bladene, hvor det samler seg som følge av fotosyntesen, trenge inn til røttene, noe som er svært viktig for myr-, vann- og andre planter som lever under forhold med vanskelig gassutveksling.
Integumentært vev beskytte høyere planter fra uttørking, plutselige temperatursvingninger, fra overflødig strålingsenergi, fra mekanisk skade, fra overdreven isolasjon.
I det første leveåret er bladene og de unge stilkene til planten dekket med et lag med tettpakkede celler - epidermis (huden). Epidermale celler er levende, som inneholder en kjerne, tett cytoplasma og små leukoplaster. Når cellene vokser, dannes det vakuoler i dem, ofte oppløst i dem cellesaft antocyaninpigment, som forårsaker den lilla-røde fargen på blader og stilker.
Epidermale celler har enheter som forhindrer planten i å miste vann: deres ytre vegger er kraftig fortykket og dekket med en tynn vanntett film - kutikula (kutikula). Noen planter har et voksaktig belegg over neglebåndet. Noen ganger er veggene til epidermale celler impregnert med silika (hestehaler, sedges, frokostblandinger). Ofte danner epidermale celler utvekster - hår, dekker blader og stilker i et tykt lag. Hårene kan være svært forskjellige i størrelse og form (filamentøse, forgrenede, stjerneformede, etc.). Hvis hårene ender i hevelse og er i stand til å samle seg og skille ut slim, essensielle oljer osv., de kalles kjertel.
Inntreden av luft i planten, frigjøring av oksygen og vanndamp utføres av spesielle formasjoner som er tilstede i epidermis - stomata. Vanligvis, i landplanter, er stomata plassert på undersiden av bladet, nær vannplanter med flytende blader - på toppen. Hver stomi består av et par bønneformede beskyttelsesceller og en stomatalfissur, som er et uttalt intercellulært rom. Beskyttelsescellene i stomata, i motsetning til andre celler i epidermis, inneholder alltid kloroplaster og gjennomgår aktivt fotosyntese. Stomatalfissuren kan ekspandere og trekke seg sammen på grunn av endringer i osmotisk trykk i vaktcellene.
Gjennom åpne stomata, intens diffusjon av vanndamp, oksygen og karbondioksid. Når stomata er lukket, reduseres transpirasjon og gassutveksling kraftig. I planter temperert sone antall stomata varierer avhengig av plantetype og levekår fra 100 til 700 per 1 mm 2 bladoverflate.
I flerårige landplanteorganer blir det primære integumentære vevet (epidermis) etter en tid erstattet av et sekundært, mer pålitelig beskyttende plantevev - en plugg (periderm). Peridermen oppstår som et resultat av aktiviteten til korkkambium (phellogen), som dannes under epidermis. Når fellogenceller deler seg, legger de ut lag med korkceller, som utfører beskyttende funksjoner. Cellerekkene i korken er tett ved siden av hverandre; det er ingen intercellulære mellomrom i korken. Fellogen avleirer lag med phellodermceller inni, som består av levende celler og gir næring til fellogencellene.
Etter hvert som stilken tykner og peridermen dannes, blir overhuden felt og stilken blir fra grønn til brun. Stengler med en dannet periderm er i stand til å overvintre.
For å sikre gassutveksling av indre vev, dannes det såkalte linser i korken. Hos linser er korkceller og levende parenkymceller løst forbundet med hverandre. Gassutveksling skjer gjennom intercellulære rom. Linser på unge stilker ser ut som små tuberkler.
Mest treslag Etter noen år dannes et tertiært dekkende vev - en skorpe. I et epletre forekommer det ved 6-8 år, i et furutre. - kl 8-10, for eik - kl 25-30, for agnbøk - ved 50 år. Skorpen dannes som et resultat av gjentatt avsetning av nye lag av periderm i stadig dypere lag av cortex. Når stammen tykner og fellogenet gjentatte ganger legges ned og aktivt, sprekker det perifere døde vevet i skorpen og overflaten på trestammen blir ujevn. Crusted stammer er beskyttet mot brå endringer temperaturer, fra markskogbranner, fra skade fra dyr.
Utskillelsesvev akkumulerer eller frigjør stoffer som er utelukket fra metabolismen. Internt ekskresjonsvev akkumulerer metabolsk avfall i kroppen. De består av individuelle isolerte celler blant celler i andre vev. Dette er de såkalte idioblastene. I dem samles krystaller av kalsiumoksalat, eteriske oljer, tanniner etc. Beholdere for sekret (for eksempel i sitrusfrukter) og ekskresjonspassasjer (harpikspassasjer av bartrær, tubuli av paraplyer osv.) kan dannes i de intercellulære rommene. Levende celler som samler melkesaft i vakuoler kalles laticifers. Den melkeaktige juicen inneholder harpiks, gummi, essensielle oljer, proteinforbindelser og alkaloider.
Eksternt utskillelsesvev er også mangfoldig. Kjertelhårene som danner fremspringet til noen planter frigjør eteriske oljer, salter og andre stoffer. Hydatoder er grupper av celler assosiert med det ledende vevet i bladet og ender med vannstomata; de skiller ut vann og salter oppløst i den. Eksternt ekskresjonsvev inkluderer nektarier, plassert i blomster og skiller ut en sukkerholdig væske (nektar), som tiltrekker pollinerende insekter; så vel som fordøyelseskjertlene til kjøttetende planter (i soldugg, duggblad, butterwort, etc.), som skiller ut enzymer og syrer som er nødvendige for å fordøye vevet til fangede virvelløse dyr.
Mekanisk (forsterkende) stoffer utføre en støttefunksjon. I unge organer av høyere planter gir cellevegger og turgor tilstrekkelig styrke og en viss form organ. Men i landplanter, når de vokser, er det behov for å danne spesialisert mekanisk vev med fortykkede cellevegger. Avhengig av formen på cellene og metoden for å fortykke veggene deres, skilles tre typer mekaniske vev ut: collenchyma, sclerenchyma (fibre) og sclereids (steinede celler).
Collenchyma er et mekanisk vev som består av levende celler, vanligvis parenkymalt eller litt forlenget. Den finnes i bladstilker og bladblader, i voksende deler av stengler. Avhengig av egenskapene til cellefortykning, skilles kantete og lamellære collenchyma. Det vanligste er vinkelkollenkym, som er preget av delvis fortykkelse av veggene i hjørnene av cellene i kontakt med hverandre. Generelt skapes det en slags sterk forsterkning i stoffet. Celleveggene i slikt vev består av fiber. Ved lamellært collenchyma er delvise fortykkelser av membranene arrangert i parallelle rader.
Sclerenchyma er veldig forskjellig fra collenchyma. Den består vanligvis av prosenkymale (forlengede) celler kalt fibre. Skjellene i dem er jevnt fortykket, ofte lignifisert. Innholdet i cellene dør og mekanisk funksjon utføre fortykkede membraner av vevsceller. Fibrene som utgjør tre kalles trefibre, eller libriform. Fibrene som utgjør basten (floemet) kalles bastfibre. I tekstilindustrien brukes bast ikke-lignifiserende fibre bestående av ren fiber (lin).
Sklereider er celler med sterkt fortykkede lignifiserte membraner som ikke har en fiberform. Slike celler består for eksempel av nøtteskall, frø av saftige druper osv. Steinceller i fruktkjøttet av pære- eller kvedefrukter er parenkymceller med sterkt fortykkede lignifiserte vegger.
Mekanisk vev skaper en sterk ramme av plantekroppen, som er fylt med en elastisk masse av levende celler. Mekanisk vev i stilken ligger langs periferien og øker dermed motstanden mot bøyning og brudd. I roten er de mekaniske elementene konsentrert hovedsakelig i sentrum, noe som hindrer organet i å briste.
Ledende stoffer utføre funksjonen med å lede vann og næringsstoffer inn i plantekropper. Planten har to typer ledende vev: xylem og floem. En oppadgående strøm oppstår gjennom xylem (tre): bevegelse av vann og mineraler oppløst i det fra roten til alle planteorganer. Xylem er et komplekst vev som består av faktiske ledende elementer (kar eller trakeider), som bestemmer vevets natur, samt celler som utfører mekaniske og lagringsfunksjoner.
Trakeider er døde, langstrakte, noen ganger spisse i endene, celler med intakte primærvegger. Gjennom disse veggene (i de avgrensede porene) trenger vann inn fra en trakheid til en annen ved filtrering. Det primære skallet til trakeiden tykner og blir lignifisert, bare mange porer forblir utykke. Trakeider er karakteristiske for tre av bregner og gymnospermer, hvor de utfører både ledende og mekaniske funksjoner.
Det mest perfekte ledende systemet finnes i blomstrende planter. Deres xylem er representert av mer avanserte vannledende elementer - kar. Fartøyet består av en rekke segmenter, i de tverrgående veggene av hvilke hull er dannet. Takket være dem utføres den uhindrede bevegelsen av løsninger gjennom det lange og smale kapillærlignende røret til fartøyet. Membranene i karene, som trakeider, er ujevnt fortykket og impregnert med lignin. Fortykningene gir karene mekanisk styrke. Gjennom ikke-fortykkede områder av kar (porer), kan løsninger også strømme horisontalt inn i nabokar og parenkymceller. I henhold til arten av fortykkelsen av membranene skilles karene i ringformede, spiralformede, skalariforme og punktpore. På grunn av det faktum at i regioner med sesongklima om våren avleirer kambium trakeider eller kar i treet med brede åpninger og en relativt tynn vegg, og om høsten - med smale åpninger og en tykkere vegg, den såkalte veksten ringene er forskjellige. De mekaniske elementene i tre - libriforms - så vel som kar, utviklet seg fra trakeider ved å forbedre den mekaniske snarere enn ledende funksjonen: de libriforme fibrene utviklet et fortykket sekundært skall og antallet og størrelsen på porene ble redusert.
Den nedadgående strømmen av oppløste organiske stoffer som kommer fra bladene utføres av floemet. Floemet består av silrør som assimilater beveger seg gjennom, følgeceller, mekaniske celler (bastfibre) og parenkymceller.
Silrør består av en kjede av levende celler. Mellom tilstøtende celler (segmenter) av røret er det sillignende plater gjennomboret av hull (perforeringer), gjennom hvilke protoplastene til nabosegmentene kommuniserer med cytoplasmatiske ledninger. Silplaten er altså en tverrvegg av to tilstøtende celler som utgjør silrøret. I cellene som danner silrør, er det som regel ingen kjerner, men protoplastene forblir i live og leder aktivt organisk materiale. De medfølgende cellene, eller satellittcellene, er plassert i nærheten av silrørene. De har kjerner og cytoplasma med mange mitokondrier; intensiv metabolisme oppstår i dem. Silrørssegmentene og følgecellene er strukturelt og funksjonelt nært beslektet med hverandre; kjernene til satellittceller koordinerer funksjonene til cytoplasmaet til silrørceller.
Ledende vev, sammen med mekaniske vevsfibre, er organisert i spesielle strukturer - ledende eller vaskulære-fibrøse bunter. Disse buntene trenger gjennom alle planteorganer, og forenes til et enkelt ledende system.
Vaskulære-fibrøse bunter er forskjellige i plasseringen av xylem og floem i forhold til hverandre; Det er konsentriske, sivile, bicollaterale og radiale bunter.
I konsentriske bunter omgir ledende vev av en type ledende vev av en annen type: floem-xylem eller xylem-floem.
I collateral bunter er xylem og floem plassert side om side. I slike bunter vender xylemregionen ofte mot organets akse, og floemregionen vender ofte mot periferien. Slike bunter er karakteristiske for stilkene og bladene til de fleste moderne planter.
I bicollaterale bunter grenser to tråder av floem til xylemet: den ene er nærmere organets akse, den andre er nærmere utsiden.
Komplekse radielle tuer er karakteristiske for planterøtter. I dem er xylemet plassert langs organets radier, mellom dem er det tråder av floem.
Lukkede fibrovaskulære bunter er de der det ikke er kambiale celler og de ikke er i stand til sekundær fortykning på grunn av dannelsen av nye celler.
I tillegg til ledende og mekaniske elementer inneholder åpne vaskulære-fibrøse bunter også kambialvev. Buntkambiumet er et enkelt lag med konstant langsgående delende celler som utvikler seg til nye ledende og mekaniske elementer, og gir dermed sekundær fortykkelse av buntene spesielt og stammen generelt.
Tekstil- en gruppe celler som er strukturelt og funksjonelt forbundet med hverandre, like i opprinnelse, struktur og utfører visse funksjoner i kroppen.
Vev oppsto i høyere planter i forbindelse med deres tilgang til land og nådde den største spesialiseringen på angiospermer, der opptil 80 arter skilles. De viktigste plantevevet:
Pedagogisk,
integumentær,
Ledende,
Mekanisk
Grunnleggende.
Stoffer kan være enkelt og kompleks. Vanlige stofferbestår av én type celle (for eksempel collenchyma, meristem), ogkompleks- fra celler av forskjellige strukturer, utfører, i tillegg til hoved- og tilleggsfunksjoner(epidermis, xylem, floem, etc.).
Pedagogiske stoffer, eller merister, er embryonale vev. Takket være deres langvarige evne til å dele seg (noen celler deler seg gjennom hele livet), deltar meristem i dannelsen av alt permanent vev og danner dermed planten, og bestemmer også dens langsiktige vekst.
Cellene i utdanningsvev er tynnveggede, mangefasetterte, tett lukket, med tett cytoplasma, en stor kjerne og veldig små vakuoler. De er i stand til å dele seg i forskjellige retninger.
I henhold til opprinnelsen til meristemer er det primære og sekundære. Det primære meristemet er embryoet til frøet, og hos en voksen plante forblir det på spissen av røttene og spissen av skuddene, noe som gjør det mulig for dem å vokse i lengde. Videre vekst av roten og stilken i diameter ( sekundær vekst) er gitt sekundære merister- kambium og fullogen. Basert på deres plassering i plantekroppen, skilles apikale (apikale), laterale (laterale), interkalære (interkalære) og sår (traumatiske) meristemer.
Integumentært vev lokalisert på overflaten av alle planteorganer. De opptrer hovedsakelig beskyttende funksjon- beskytte planter mot mekanisk skade, penetrasjon av mikroorganismer, plutselige temperatursvingninger, overdreven fordampning, etc. Avhengig av deres opprinnelse, skilles tre grupper av integumentært vev - epidermis, periderm og skorpe.
Epidermis (epidermis, hud)- primært integumentært vev lokalisert på overflaten av blader og unge grønne skudd (fig. 8.1). Den består av et enkelt lag med levende, tettpakkede celler som ikke har kloroplaster. Cellemembranene er vanligvis kronglete, noe som sikrer deres sterke lukking. Den ytre overflaten av cellene i dette vevet er ofte dekket med et kutikula eller voksaktig belegg, som er en ekstra beskyttelsesanordning. Overhuden til blader og grønne stilker inneholder stomata som regulerer transpirasjon og gassutveksling i planten.
Periderm- sekundært integumentært vev av stengler og røtter, som erstatter epidermis i flerårige (sjeldnere årlige) planter (fig. 8.2.). Dens dannelse er assosiert med aktiviteten til det sekundære meristemet - fellogen (korkkambium), hvis celler deler seg og differensierer i sentrifugalretningen (utover) inn i korken (phellema), og i sentripetalretningen (innover) - til en lag av levende parenkymceller (phelloderm). Kork, phellogen og phelloderm utgjør periderm.
Ris. 8.1. Bladepidermis ulike planter: A-klorofytum; 6 - vanlig eføy: i - duftende geranium; G - hvit morbær; 1- epidermale celler; 2 - stomatale vaktceller; 3 - stomatal fissur.
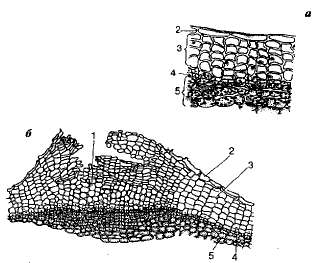
Figur 8.2. Periderm av hyllebærstilk (a - tverrsnitt av et skudd, b - linser): I-utføre stoff; 2 - rester av epidermis; 3 -kork (phellema); 4 - felogen; 5 - phelloderm.
Korkens celler er impregnert med et fettlignende stoff - suberin - og slipper ikke vann og luft gjennom, så innholdet i cellen dør og den fylles med luft. Flerlagskorken danner et slags stengeldeksel som pålitelig beskytter planten mot uønskede påvirkninger. miljø. For gassutveksling og transpirasjon av levende vev som ligger under pluggen, inneholder sistnevnte spesialopplæring -linser; Dette er hull i pluggen fylt med løst anordnede celler.
Skorpe dannet i trær og busker for å erstatte kork. I det dypere vevet i cortex legges nye områder av fellogen ned, og danner nye lag med kork. Som et resultat blir det ytre vevet isolert fra den sentrale delen av stilken, deformert og dør.På overflaten av stilken dannes det gradvis et kompleks av dødt vev, bestående av flere lag med kork og døde deler av bark. En tykk skorpe gir mer pålitelig beskyttelse for planten enn kork.
Ledende stoffer sikre bevegelse av vann og næringsstoffer oppløst i det gjennom hele planten. Det er to typer ledende vev - xylem (tre) og floem (bast).
Xylem er det viktigste vannledende vevet til høyere karplanter, og sikrer bevegelse av vann med mineraler oppløst i det fra røttene til bladene og andre deler av planten (stigende strøm). Den utfører også en støttefunksjon. Xylemet består av trakeider og luftrør (kar) (fig. 8.3), treparenkym og mekanisk vev.
Trakeider De er smale, svært langstrakte døde celler med spisse ender og lignifiserte membraner. Inntrengningen av løsninger fra en tracheide til en annen skjer ved filtrering gjennom porer - fordypninger dekket med en membran. Væske strømmer sakte gjennom trakeidene, siden poremembranen hindrer vannbevegelse. Trakeider finnes i alle høyere planter, og i de fleste kjerringrokk, klubbmoser, bregner og gymnospermer tjener de som det eneste ledende elementet i xylemet. U angiospermer Sammen med trakeider er det kar.
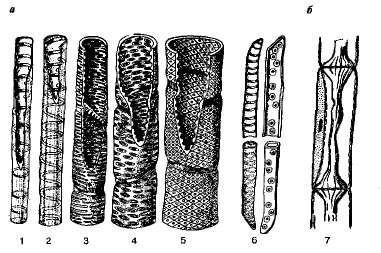
Figur 8.3. Elementer av xylem (a) og floem (6): 1-5 - henholdsvis ringformet, spiralformet, skalariform og porøs (4, 5) luftrør; 6 - ringede og porøse trakeider; 7 - silrør med ledsagercelle.
Luftrør (kar)- dette er hule rør som består av individuelle segmenter plassert over hverandre. I segmentene dannes gjennomgående hull på tverrveggene - perforeringer, eller disse veggene blir fullstendig ødelagt, på grunn av hvilken hastigheten på strømmen av løsninger gjennom karene øker mange ganger. Skallene på karene er impregnert med lignin og gir stilken ekstra styrke. Avhengig av arten av fortykkelsen av membranene, skilles luftrørene ut som ringede, spiralformede, skalariforme, etc. (se fig. 8.3).
Phloem leder organiske stoffer syntetisert i bladene til alle planteorganer (nedadgående strøm). I likhet med xylem er det et komplekst vev og består av silrør med følgeceller (se fig. 8.3), parenkym og mekanisk vev. Silrør er dannet av levende celler plassert over hverandre. Deres tverrvegger er gjennomhullet små hull, danner som en sil. Cellene i silrørene er blottet for kjerner, men inneholder cytoplasma i den sentrale delen, hvis tråder går gjennom hull i tverrskilleveggene inn i naboceller. Silrør, som kar, strekker seg langs hele lengden av planten. Ledsagerceller er koblet til segmentene av siktrørene ved hjelp av tallrike plasmodesmata og utfører tilsynelatende noen av funksjonene som går tapt av siktrørene (enzymsyntese, ATP-dannelse).
Xylem og floem er i nært samspill med hverandre og danner spesielle komplekse grupper- ledende bunter.
Mekaniske stoffer sikre styrken til planteorganene. De danner en ramme som støtter alle planteorganer, og motstår deres brudd, kompresjon og brudd. Hovedkarakteristikkene til strukturen til mekaniske vev, som sikrer deres styrke og elastisitet, er kraftig fortykning og lignifisering av membranene deres, tett lukking mellom celler og fravær av perforeringer i celleveggene.
Mekanisk vev er mest utviklet i stilken, hvor de er representert av bast- og trefibre. I røttene er mekanisk vev konsentrert i midten av organet.
Avhengig av formen på cellene, deres struktur, fysiologisk tilstand og metode for fortykning cellemembraner Det finnes to typer mekanisk vev: collenchyma og sclerenchyma (fig. 8.4).

Ris. 8.4. Mekaniske stoffer: a -vinkelkollenkym; 6- sklerenkym; V -- sclereider fra kirsebær plommefrukter: 1 - cytoplasma, 2-tyknet celleveggen, 3 - poretubuli.
Collenchyma er representert av levende parenkymceller med ujevnt fortykkede membraner, noe som gjør dem spesielt godt tilpasset for å styrke unge voksende organer. Som primære, strekker collenchyma-celler seg lett og forstyrrer praktisk talt ikke forlengelsen av delen av planten der de er lokalisert. Collenchyma er vanligvis lokalisert i separate tråder eller en kontinuerlig sylinder under epidermis til de unge stilkene og bladstilkene, og grenser også til venene i tofrøbladede blader. Noen ganger inneholder collenchyma kloroplaster.
Sklerenkym består av langstrakte celler med jevnt fortykkede, ofte lignifiserte membraner, hvis innhold dør av tidlige stadier. Membranene til sclerenchyma-celler har høy styrke, nær styrken til stål. Dette vevet er bredt representert i de vegetative organene til landplanter og danner deres aksiale støtte.
Det finnes to typer sklerenkymceller: fibre og sklereider. Fibre- disse er lange tynne celler, vanligvis samlet i tråder eller bunter (for eksempel bast- eller trefibre). Sclereids - disse er runde, døde celler med veldig tykke, lignifiserte membraner. De danner frøskallet, nøtteskall, frø av kirsebær, plommer og aprikoser; de gir pærekjøttet sin karakteristiske grove karakter.
Hovedstoff, eller parenkym, består av levende, vanligvis tynnveggede celler som danner grunnlaget for organer (derav navnet vev). Det huser mekanisk, ledende og annet permanent vev. Hovedvevet utfører en rekke funksjoner, og derfor skiller de mellom assimilering (klorenkym), lagring, pneumatisk (aerenkym) og vannførende parenkym (fig. 8.5).

Figur 8.5. Parenkymalt vev: 1-3 - klorofyllbærende (henholdsvis søyleformet, svampete og foldet); 4-lagring (celler med stivelseskorn); 5 - pneumatisk, eller aerenchyma.
Celler assimilering vev inneholder kloroplaster og utfører funksjonen til fotosyntese. Hoveddelen av dette vevet er konsentrert i bladene, en mindre del i unge grønne stengler.
I celler lagring proteiner, karbohydrater og andre stoffer avsettes i parenkymet. Den er godt utviklet i stilkene til treplanter, i røtter, knoller, løker, frukt og frø. Planter av ørkenhabitater (kaktus) og saltmyrer har akvifer parenchyma, som tjener til å akkumulere vann (for eksempel inneholder store eksemplarer av kaktus fra slekten Carnegia opptil 2-3 tusen liter vann i vevet). Vann- og myrplanter utvikles spesiell type hovedstoff - pneumatisk parenkym, eller aerenkym. Aerenchyma-celler danner store luftbærende intercellulære rom, gjennom hvilke luft blir levert til de delene av planten hvis forbindelse med atmosfæren er vanskelig
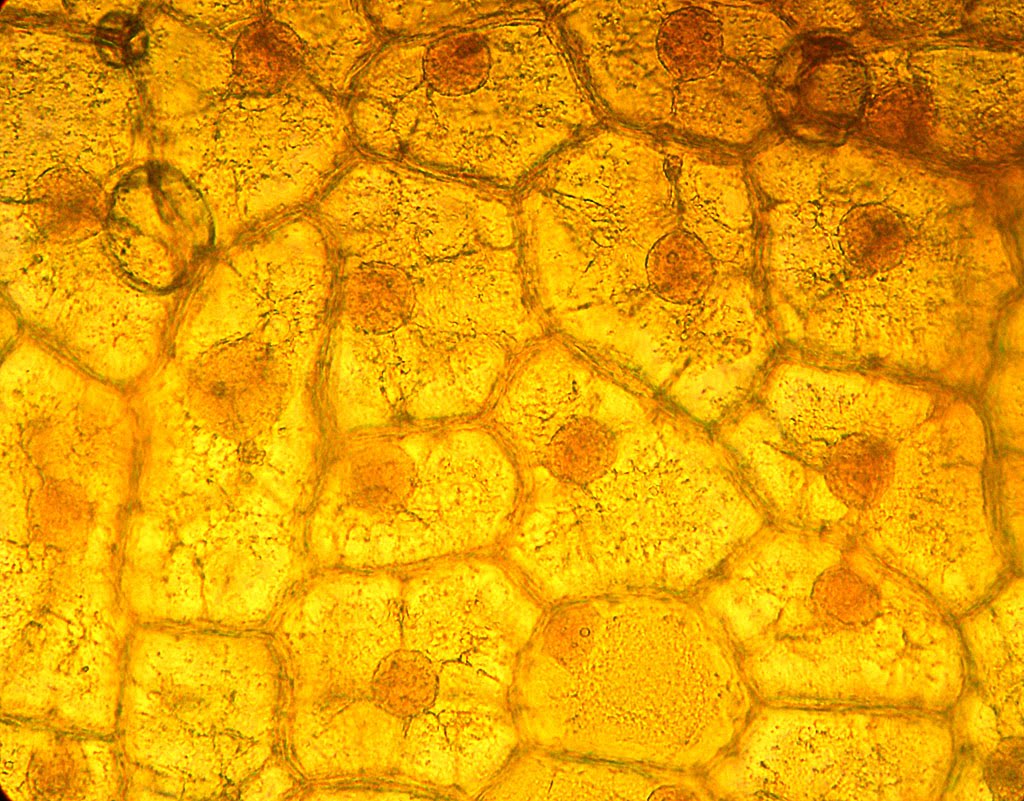
Løk skalerer under et mikroskop