Actinomycetes (Actinomycetales, fra det greske aktis - ray, mykes - sopp) er forgrenende bakterier som tilhører fylum Actinobacteria. De er en del av den normale mikrofloraen Fordøyelsessystemet terrestriske virveldyr og virvelløse dyr, og er også tilstede i overflod i jorda og spiller en viktig rolle i økologien og syklusen av stoffer i jorda.
Disse mikroorganismene er årsakene til mange opportunistiske patologier - de som oppstår som et resultat av nedsatt funksjon immunforsvar kropp. Actinomycetes er mye brukt i bioteknologi, da de er en kilde til en rekke antibakterielle og antitumorstoffer.
Ris. 1. Streptomycetes syntetiserer en enorm mengde antibakterielle og antitumormedisiner.
Strukturen til actinomycetes: hvorfor er de bakterier og ikke sopp?
1. Organisering av genetisk materiale
Det arvelige materialet til Actinomycetes er inneholdt i ett molekyl av deoksyribonukleinsyre, som har en ringform og er fritt plassert i cytoplasmaet - den samme formen for organisering av genetisk materiale, kalt en nukleoid, er karakteristisk for andre bakterier. Hos sopp er arvestoffet organisert og inkludert i cellekjernen.
Actinomycete DNA inneholder et stort nummer av GC-par (65-75 % av det totale antallet nukleotider). Denne egenskapen er konstant, er ikke avhengig av mutasjoner, og brukes derfor i taksonomien til mikroorganismer. Dette innholdet av GC-par gjør DNA-et til actinomycetes svært ildfast, derfor tar DNA-analyse av actinomycetes mer tid sammenlignet med andre bakterier.

Ris. 2. Skjematisk struktur cellevegg av Gr+ bakterier.

Ris. 3. Actinomycetes, Gram-farget.
Actinomycetes har en tett bakteriecellevegg, som er plassert utenfor cytoplasmatisk membran og forårsaker deres positive Gram-farging. Som andre gram-positive bakterier består den av flere dusin lag med mureinpolymer (peptidoglykan), som er gjennomsyret av teichoic og lipoteichoic syrer. Lipoteichoic syrer er forankret i den cytoplasmatiske membranen til bakterien og kobler den til celleveggen. Teichoic syrer overføre til celleveggen negativ ladning. Celleveggen til sopp består av andre polymerer - kitin og glukan.
Ris. 4. Soppcellevegg.
3. Cellulære organeller
Actinomycetes, som andre bakterier, har ikke membranorganeller. Actinomycetes har 70S ribosomer, mens sopp har 80S ribosomer, som andre eukaryote organismer.
4. Vekst av kolonier
Dannelsen av mycel under vekst er det actinomycetes ligner mest på sopp. Mycelet i tilfelle av actinomycetes er en forgrenende samling av hyfer. Hyfene er delt av septa i lange bakterieceller som inneholder flere nukleoider. Skilleveggene i en rekke arter kan løpe i en gjensidig vinkelrett retning. Hyfer forgrener seg ved knoppskyting.
Mycel som vokser inn i underlaget (jord, silt eller næringsmedium), kalt substrat. Det gir næringsstoffer til kolonien. Et luftmycel stiger over substratet, og gir kolonien en "fluffiness" - det danner sporer, så vel som såkalte "sekundære metabolitter" (i motsetning til de "primære metabolittene" av substratmyceliet), blant dem er det mange antibakterielle stoffer.
Livssyklus og fysiologi til actinomycetes
I løpet av livssyklusen mest av actinomycetes danner sporer. Noen actinomycetes formerer seg ved fragmentering av myceliet.
1. Sporulering
Actinomycete-sporer stammer fra luftmycel. Dette er eksosporer - de utvikler seg utenfor modercellen. Hyfene til luftmyceliet som sporer utvikler seg fra kalles sporebærere. Sporene kan være inneholdt i en fortykning i enden av sporangium - et sporangium (for eksempel i streptomycetes, actinoplanes og plymelia), eller kan være lokalisert i en kjede langs sporangium (for eksempel i nocardia og actinomadura).
I henhold til antall dannede sporer er actinomycetes delt inn i:
- Monosporøs (for eksempel Saccaromonospora, Micromonospora, Thermomonospora) - danner enkeltsporer, ofte ved knoppskyting og påfølgende separasjon fra morhyfen med en septum;
- Oligosporøs (for eksempel Actinomadura) - danner korte kjeder av sporer langs sporebæreren;
- Polysporøse (de fleste andre actinomycetes, for eksempel Streptomyces, Frankia, Geodermatophilus) - danner mange sporer innelukket i sporangia.

Ris. 5. Sporangium av actinomycetes av slekten Frankia.
Actinomycete-sporer kan være bevegelige - i dette tilfellet har sporen et flagellum og kan bevege seg (sporer av actinoplanes, geodermatophiles og dermatophiles). I de fleste tilfeller er sporene ikke-bevegelige og spres med vind, vann eller dyr.

Ris. 6. Dermatofiler, lysmikroskopi.
Sporulering i actinomycetes er spesielt aktiv under ugunstige forhold. Sporenes motstand mot varme er lav sammenlignet med sporer fra andre bakterier, men de tåler uttørking ikke dårligere enn andre, og har derfor enorm adaptiv betydning. Actinomycetes dominerer andre mikroorganismer i tørr ørkenjord.
Spiring av sverm krever en viss fuktighet eksternt miljø. I nærvær av vann svulmer sporen, enzymer aktiveres og metabolske prosesser settes i gang, ledsaget av frigjøring av vekstrør (fremtidige bakterielegemer) og syntese av nukleinsyrer.
2. Type pust
De fleste actinomycetes er aerobe (de krever oksygen for å opprettholde livet). Fakultative anaerober (bakterier som kan leve både i nærvær og fravær av oksygen) finnes blant arter med et kort mycelstadium, som formerer seg ved fragmentering av mycelet.
3. Syrebestandighet
Actinomycetes har acidotoleranse - motstand i et surt miljø, som gjør at de kan leve i syremettet skogsjord. Syrebestandighet i laboratoriet kan bestemmes ved å farge et preparat som inneholder actinomycetes i henhold til Ziehl-Neelsen (fuksin etterfulgt av etsing med svovelsyre og farging med metylenblått). De fleste aktinomyceter med denne fargen misfarges ikke etter etsning med syre og beholder en rød magentafarge. Alkalisk miljø ugunstig for disse bakteriene: ved forhøyet pH er de utsatt for spordannelse.
4. Funksjoner av metabolisme

Ris. 7. Aerobe aktinomyceter danner pigment på agarskråninger. Fra venstre til høyre: Actinomadura madurae, Nocardia asteroides, Micromonospora.
Dannelsen av "sekundære metabolitter" av luftmycel ble nevnt ovenfor. Blant dem:
- pigmenter som forårsaker forskjellige farger av luftmycel når de vokser på medier;
- flyktige luktstoffer som gir en karakteristisk lukt til jorda etter regn, stillestående vann og huden til noen dyr;
- antibiotika:
en. soppdrepende - polyener;
b. antibakteriell - for eksempel streptomycin, erytromycin, tetracyklin, vankomycin;
c. antitumor - antracykliner, bleomycin.
Hvor bor actinomycetes?
Actinomycetes i det største antallet finnes i jord, og det er betydelig færre mycelformer enn sporer. De spiller en betydelig rolle i dannelsen av humus, som bryter ned organiske stoffer som er vanskelige for andre bakterier å utnytte. I denne forbindelse brukes aktinomyceter som sanitære indikatormikroorganismer i sanitære og epidemiologiske forhold: deres påvisning i store mengder i jord eller vann indikerer tilstedeværelsen av kompost i det tilsvarende substratet.

Ris. 8. Actinomycetes i kompost.
Actinomycetes er symbionter av mange planter, og hjelper dem å fikse nitrogen. Samtidig er mange mikroorganismer av denne klassen patogener av plantesykdommer.

Ris. 9. Potetstreptomycosis.
De finnes også i den normale mikrofloraen i fordøyelsessystemet til en rekke dyr, alt fra jord annelids(for eksempel regnvann) og slutter med store husdyr.
Disse mikroorganismene hjelper til med å bryte ned cellulose, som er rikelig tilstede i plantemat. Hos mennesker finnes aktinomyceter i munnhulen (gummi og plakk), tarm (distal kolon), hud (ansikt, nese, bak ørene, mellom fingrene) og i organer luftveiene(hovedsakelig i de øvre luftveiene).
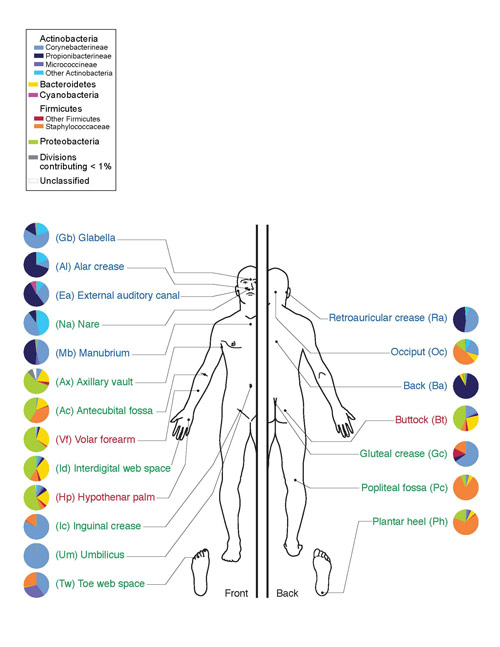
Ris. 10. Mikroflora av menneskelig hud. Filum Actinobacteria er indikert i nyanser av blått, klassen Actinomycetes er indikert i knallblått.
Actinomycetes, hvis kroppens immunreaktivitet er redusert, kan forårsake actinomycosis - opportunistiske sykdommer som består i dannelsen av actinomycosis granulomer - ansamlinger av bakterielegemer som ligner korn av gult svovel ("drusen"), omgitt av immunkompetente celler. Den inflammatoriske reaksjonen fører til smelting av granulomer, dannelse av fistler, noe som fører til perforering av organer og spredning av bakterier med blod.

Ris. 11. Actinomycosis drusen, Gram flekk.

Ris. 12. Aktinomykose i overkjeven hos en ku.

Ris. 13. Human maxillær actinomycosis.
Actinomycetes – fantastiske organismer, fortsatt villedende mange forskere med deres likhet med sopp. Sammen med den potensielle faren i form av opportunistisk actinomycosis, gir disse organismene mennesker fruktbar jord og våpen for å bekjempe smittsomme og onkologiske sykdommer - antibiotika og cytostatika.
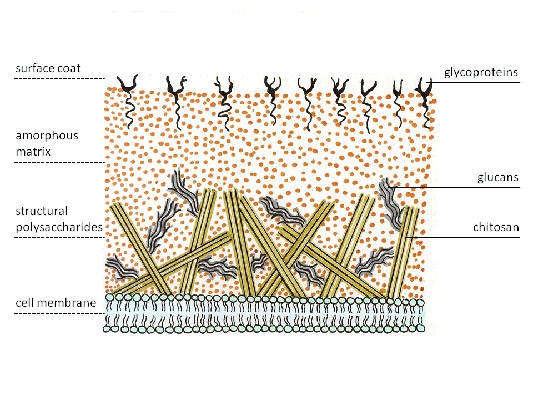
Kjemisk sammensetning og struktur av celleveggen til sopp
Celleveggen til sopp er flerlags, med forskjellige lag dannet av strukturelle karbohydrater som er forskjellige i deres kjemiske sammensetning, som kan deles inn i 3 grupper basert på deres kjemiske sammensetning:
glukosepolymerer(glukan, kitin, cellulose). Glukaner utgjør det ytre laget av celleveggen til de fleste sopp. Det indre laget av soppcelleveggen er dannet av kjeder av kitin, noe som gir den stivhet. Kitin erstatter cellulose, som er fraværende i de fleste sopp, men er en del av celleveggen til oomycetes, som foreløpig ikke tilhører typiske sopp. Deacetylert kitin kalles kitosan, som i kombinasjon med kitin danner celleveggen til zygomyceter.
polymerer av andre monosakkarider (mannose, galaktose, etc.) i motsetning til høyere planter, hvor de danner grunnlaget for matrisen under vanlig navn hemicellulose, mindre typisk for sopp. Unntaket er gjær, hvis cellevegger er spesielt rike på mannosepolymerer kalt mannaner. Det antas at denne veggsammensetningen bedre sikrer spiring.
polymerer av karbohydrater kovalent knyttet til peptider (glykoproteiner) danne det midterste laget av en flerlags cellevegg og lek viktig rolle, både for å opprettholde den strukturelle integriteten til celler og i dens metabolske prosesser med miljøet.
Andre spesifikke trekk ved soppcellen inkluderer: fravær av plastider, som bringer den nærmere en dyrecelle;
Ingen stivelse, som i eumyceter er erstattet av et polysakkarid nær animalsk stivelse glykogen, i oomyceter med et polysakkarid nær laminarin brunalger. Det produseres også en rekke reservekarbohydrater som er spesifikke for sopp.
Utvikling av spesifikke sekundære metabolitter, av hvilke stor rolle antibiotika, fyto- og mykotoksiner, og fytohormoner spiller en rolle.
De spesifikke egenskapene til sopp inkluderer også heterokaryose og paraseksuell prosess.
Hos sopp er fenomenet heterokaryose eller heteronuklearitet svært utbredt, der i en celle lang tid kjerner som er heteroalliske for noen gener er bevart. Heterokaryose erstatter heterozygositet i haploide sopp og fremmer rask tilpasning av sopp til endrede forhold. Tilstedeværelsen av heteronuklearitet skyldes en rekke spesifikke funksjoner sopp:
1. tilstedeværelsen av mer enn én kjerne i en celle
2. den spesifikke strukturen til celleskilleveggen, der det er ett eller flere gjennomgående hull, kalt porer, gjennom hvilke kjerner kan migrere fra en celle til en annen
3. Hyfer innenfor en koloni og til og med forskjellige tettliggende kolonier dyrket fra forskjellige sporer av samme type sopp vokser ofte sammen, som et resultat av at utveksling av kjerner av forskjellige stammer er mulig.
Paraseksuell (pseudofil) prosess. Hvis i heterokaryote celler kjerner som er heteroalleliske på et hvilket som helst sted smelter sammen, vises en heterozygot diploid kjerne. Det kan komme inn i en spore og gi opphav til en diploid heterozygot klon. Under mitoseprosessen kan diploide kjerner gå tilbake til haploid tilstand på grunn av tap av ett sett med kromosomer, eller en utveksling av kromosomseksjoner kan forekomme i dem (mitotisk kryssing). Begge prosessene er ledsaget av rekombinasjon av foreldregener og, følgelig, fenotyper. Paraseksuell (aseksuell) rekombinasjon er et svært sjeldent fenomen, som ikke overstiger en kjerne per million, men på grunn av det store antallet kjerner i myceliet, observeres det konstant i sopppopulasjoner.
Reproduksjon– vegetativ, aseksuell, seksuell.
Vegetativ– fragmentering av thallus, dannelse av klamydosporer, som etter en periode med dvale spirer til mycelium, spirende i gjær.
Aseksuell Reproduksjon i forskjellige sopp kan utføres av mobile og immobile sporer. Zoosporer danner en relativt liten gruppe sopp og sopplignende organismer - akvatiske og noen terrestriske, der genetiske forbindelser med vannlevende sopp og alger. Strukturen til flagella er viktig diagnostisk tegn når de er tildelt et bestemt rike. De aller fleste sopp aseksuell reproduksjon immobile sporer dannes, noe som indikerer deres svært gamle fremvekst på land. I henhold til stedet for dannelse og lokalisering skilles endogene sporangiosporer, dannet i sporangier og eksogene (konidier), og utvikler seg på spesielle hyfer - konidioforer. Konidier dannes i de fleste sopp (ascomycetes, basidiomycetes, noen zygomycetes) som utgjør konidiesporulering, som er ekstremt mangfoldig og mye brukt for å diagnostisere sopp.
Seksuell reproduksjon av sopp har sin egen spesifisitet både i morfologien til den seksuelle prosessen og i mekanismene til genetiske og fysiologisk regulering kjønn og overføring av arvelig informasjon.
Somatogami- den vanligste og enkleste typen seksuell prosess, består av sammensmelting av to udifferensierte kjønnsceller somatiske celler. Noen ganger skjer det selv uten cellefusjon - kjernene inne i cellen smelter sammen. Finnes i de fleste basidiomyceter, pungdyrgjær og noen andre taxaer.
Gametangiogami– gametangia er isolert på det haploide mycelet, og innholdet deres tappes under den seksuelle prosessen. Denne seksuelle prosessen er karakteristisk for de fleste pungdyrsopper. En variant av gametangiogami er zygogami hos zygomycetes.
Gametogami i form av iso-hetero og oogamy, forekommer det mye sjeldnere hos sopp enn hos andre eukaryoter. Iso- og heterogami forekommer bare hos chytridiomycetes. Klassisk oogami med dannelse av sæd og egg kommer ikke til uttrykk i sopp, men svært modifiserte varianter finnes.
I henhold til egenskapene til kjønnsregulering hos sopp, skilles flere typer seksuell prosess ut.
Gynandromixis kan vurderes ved å bruke eksemplet med toboende oomyceter, der oogonier og antheridia utvikler seg på forskjellige thalli, for eksempel sent blight eller potetsopp. Hvis en genetisk homogen stamme dyrkes i en monokultur, reproduserer den kun på aseksuell måte. Hvis mycelene til to stammer er i nærheten, kan morfogenetiske endringer spores i dem under påvirkning av kjemiske sekresjoner av steroidnatur - sexferomoner. Antheridiol induserer dannelsen av antheridia hos partneren, og oogoniol induserer dannelsen av oogonium. I dette tilfellet er reguleringen av sex relativ: om en stamme vil danne antheridia eller oogonia avhenger av det kvantitative forholdet mellom de tilsvarende feromonene i den og dens partner. Derav navnet på den seksuelle prosessen - gynandromixis.
Dimixis eller heterohallisme. Det har lenge vært bemerket at sopp kan være homo- eller heterotaliske. Hos homotalliske arter, under den seksuelle prosessen, smelter genetisk identiske kjerner i myceliet. Hos heterotalliske arter er det nødvendig på et tidspunkt for å gjennomgå den seksuelle syklusen (forskjellig i forskjellige typer sopp) fusjon av etterkommere av sporer (mer presist deres kjerner). For at to stammer skal være seksuelt kompatible, må det være genetisk forskjell (heteroallelisitet) på visse loci kalt paringsloki. De fleste sopp (zygomycetes, ascomycetes og noen basidiomycetes) har ett parringssted med to alleler. Parringsstedet består av flere gener som styrer syntesen av sexferomoner. Slik heterohallisme kalles unifaktoriell eller bipolar. Avkommet av slike sopp etter meiose er delt inn i to selvinkompatible, men gjensidig kompatible grupper i forholdet 1:1, dvs. sannsynligheten for relatert (innavl) og urelatert (utavl) krysning er, som i bifile høyere eukaryoter, 50 %.
I genomet til høyere basidiomyceter er det to parrende loci - A og B, og bare stammer som er heteroalleliske på begge lociene er kompatible med hverandre (Ax Bx er kompatibel med Ay By, men ikke med Ax By og Ay Bx). Slik heterohallisme kalles tofaktor eller tetrapolar. Det reduserer sjansen for innavl med opptil 25 %.
Diaphoromixis– høyere basidiomyceter har ikke to, men mange alleler av parringsstedet, som finnes tilfeldig i forskjellige stammer som utgjør populasjonen. Denne paringskontrollen sikrer 100 % sjanse for utavl, siden stammer med forskjellige alleler er gjensidig kompatible, og det er mange alleler. Som et resultat dannes panmix-hybridpopulasjoner.
Livssykluser til sopp like forskjellige som selve soppene. De viktigste syklusene, deres tilhørighet til avdelingene for sopp
1. Den aseksuelle syklusen er karakteristisk for en enorm gruppe ufullkomne eller mitofungi som har mistet seksuell reproduksjon. Delingen av kjernene deres er utelukkende mitotisk. De aller fleste mitofunger tilhører pungdyrsopper, men på grunn av tapet av den seksuelle prosessen utgjør de formell gruppe ufullkomne sopp eller deuteromycetes.
2. Haploid syklus. Den vegetative thallus bærer haploide kjerner. Etter den seksuelle prosessen (syngami) deler den diploide zygoten (vanligvis etter en hvileperiode) seg meiotisk - zygotisk meiose. Karakteristisk for zygomyceter og mange chytridiomycetes.
3. Den haploid-dikaryote syklusen er preget av det faktum at etter sammensmeltingen av innholdet av gametangia (gametangiogami) eller somatiske celler av haploid mycel (somatogami), danner kjernene dikaryoner (par av genetisk forskjellige kjerner). De deler seg synkront og danner dikaryot mycel. Den seksuelle prosessen ender med fusjonen av dikaryonkjerner, den resulterende zygoten deler seg ved meiose uten hvileperiode. Meiosporer utgjør den seksuelle sporuleringen av pungdyr og basidiomyceter i form av ascosporer og basidiosporer. Når de spirer, dannes haploid mycel. I de aller fleste pungdyrsopper (unntatt gjær og taffrinsopp) Livssyklus den haploide fasen dominerer i form av vegetativt mycel (anamorf), den dikaryote fasen er kortvarig og er representert av askogene hyfer som poser dannes på (teleomorf). Hos basidiomyceter dominerer den dikaryote fasen i livssyklusen den haploide fasen er kortvarig.
4. Den haploid-diploide syklusen i form av en isomorf generasjonsendring er sjelden hos sopp (noen gjærsopp og vannlevende chytridiomyceter).
5. Den diploide syklusen er karakteristisk for oomyceter og noen pungdyrgjær. Den vegetative thallus er diploid, gametisk meiose observeres under dannelsen av gametangia eller gameter.
3. Miljøgrupper sopp
Sopp og sopplignende organismer finnes i alle terrestriske og akvatiske økosystemer, Hvordan den viktigste delen heterotrofisk blokk, okkuperer sammen med bakterier trofisk nivå nedbrytere. Bred bruk sopp i biosfæren bestemmes av en rekke de viktigste funksjonene:
1. De fleste har en mycelstruktur av thallus. (lar deg raskt mestre underlaget og ha en stor kontaktflate med miljøet).
2. Høy hastighet på vekst og reproduksjon, tillater kort tid befolke store masser substrat, danner et stort antall sporer og distribuerer dem over lange avstander.
3. Høy metabolsk aktivitet, manifestert i et bredt spekter av miljøfaktorer.
4. Høy grad av genetisk rekombinasjon, betydelig biokjemisk variasjon, miljømessig plastisitet.
5. Evnen til å raskt gå over til en tilstand av suspendert animasjon, å oppleve ugunstige forhold I løpet av lang tid.
Hoved miljøfaktor for sopp er et matsubstrat. I forhold til denne faktoren skilles hovedgruppene av sopp, som kalles trofiske grupper.
4. en kort beskrivelse av avdelinger av sopp.
Prokaryot celle
Prokaryoter- organismer som, i motsetning til eukaryoter, ikke har en dannet cellekjerne og andre indre membranorganeller(med unntak av flate cisterner hos fotosyntetiske arter, som cyanobakterier). Det eneste store sirkulære (i noen arter - lineære) dobbelttrådet DNA-molekyl, som inneholder hoveddelen av cellens genetiske materiale (den såkalte nukleoiden), danner ikke et kompleks med histonproteiner (det såkalte kromatinet). ). Prokaryoter inkluderer bakterier, inkludert cyanobakterier (blågrønne alger) og arkea. Etterkommerne av prokaryote celler er organellene til eukaryote celler - mitokondrier og plastider. Hovedinnholdet i cellen, som fyller hele volumet, er viskøs granulær cytoplasma.
Eukaryot celle
Eukaryoter- organismer som, i motsetning til prokaryoter, har en dannet cellekjerne, avgrenset fra cytoplasma av en kjernekappe. Det genetiske materialet er inneholdt i flere lineære dobbelttrådete DNA-molekyler (avhengig av typen organisme, kan antallet per kjerne variere fra to til flere hundre), festet fra innsiden til cellekjernens membran og dannes i den enorme flertall (unntatt dinoflagellater) et kompleks med histonproteiner kalt kromatin. Eukaryote celler har et system av indre membraner som i tillegg til kjernen danner en rekke andre organeller (endoplasmatisk retikulum, Golgi-apparat, etc.). I tillegg har de aller fleste permanente intracellulære prokaryote symbionter - mitokondrier, og alger og planter har også plastider.
2) cytoplasma
Cytoplasma - Internt miljø levende eller døde celler, unntatt kjernen og vakuolen, begrenset plasmamembran. Det inkluderer hyaloplasma - det viktigste gjennomsiktige stoffet i cytoplasmaet, de essensielle cellulære komponentene som finnes i det - organeller, samt forskjellige ikke-permanente strukturer - inneslutninger.
Sammensetningen av cytoplasma inkluderer alle typer organiske og uorganiske stoffer. Den inneholder også uløselig avfall av metabolske prosesser og reservedeler næringsstoffer. Hovedstoffet i cytoplasmaet er vann.
Cytoplasma beveger seg konstant, flyter inn i en levende celle, beveger seg med den ulike stoffer, inneslutninger og organeller. Denne bevegelsen kalles syklose. Alle metabolske prosesser finner sted i den.
Cytoplasmaet er i stand til vekst og reproduksjon og kan, hvis det fjernes delvis, gjenopprettes. Imidlertid fungerer cytoplasmaet normalt bare i nærvær av kjernen. Uten den kan ikke cytoplasma eksistere lenge, akkurat som kjernen uten cytoplasma.
Den viktigste rollen til cytoplasma er å forene alle cellulære strukturer (komponenter) og forsyne dem med kjemisk interaksjon. Cytoplasmaet opprettholder også turgoren (volumet) til cellen og opprettholder temperaturen.
3) Cellevegg
Celleveggen- en stiv cellemembran plassert utenfor den cytoplasmatiske membranen og utfører strukturelle, beskyttende og transporterende funksjoner. Finnes i de fleste bakterier, arkea, sopp og planter. Dyr og mange protozoer har ikke cellevegg.
Cellevegger prokaryot
Bakterielle cellevegger er sammensatt av peptidoglykan (murein) og er av to typer: gram-positive og gram-negative. Celleveggen av den gram-positive typen består utelukkende av et tykt lag av peptidoglykan, tett tilstøtende cellemembranen og gjennomsyret av teichoic eller poteichoic syrer. I den gram-negative typen er peptidoglykanlaget mye tynnere, mellom det og plasmamembranen er det et periplasmatisk rom, og på utsiden er cellen omgitt av en annen membran, representert av den såkalte. lipopolysakkarid og er et pyrogent endotoksin fra gramnegative bakterier.
Soppcellevegger
Celleveggene til sopp er sammensatt av kitin og glukaner.
Kjennetegn på sopp
Til soppen i vid forstand inkluderer heterotrofe eukaryoter med en osmotrof type ernæring. Assosiert med kostholdsvaner karaktertrekk strukturen og livsstilen til disse organismene. Mest karakteristiske trekk sopp:
– Den vanligste formen for den vegetative kroppen er mycelium, som består av gjentatte forgrenede tråder som trenger inn i underlaget og absorberer næringsstoffer oppløst i vann fra det.
– Organisk materiale i underlaget, som er en næringskilde for sopp, er det i de fleste tilfeller i form av biopolymerer som ikke er i stand til å trenge gjennom cellemembran. Soppceller produserer depolymeraseenzymer, som frigjøres til underlaget og bryter ned polymerer til enklere. organiske forbindelser, i stand til å bli transportert inn i soppcellen. Slike enzymer kalles vanligvis eksoenzymer.
– Aktiv absorpsjon av stoffer fra substratet skjer på grunn av det enorme turgortrykket, som i soppceller er mye høyere sammenlignet med andre eukaryote organismer.
– Sopp er veldig preget av dannelsen av sporer, som oppstår under både aseksuell og seksuell reproduksjon. Gjenkjennelse av sopp er ikke mulig av den vegetative kroppen, men ved sporulering. På grunn av plasseringen av den vegetative kroppen inne i underlaget og den immobile livsstilen, på reproduksjonstidspunktet, dannes det forskjellige sporebærende strukturer på soppmyceliet, som stiger over underlaget og sporene som dannes på dem fordeles av luftstrømmer over veldig lange avstander.
Moderne systemer sopp
Av moderne ideer, basert på studiet av genomene til ekomorfen "sopp" er en kollektiv gruppe der ekte sopp skilles ut, kongeriket Mycota eller Fungi, hvorav flertallet og mindre antall arter taxa og mindre mangfoldig gruppe sopplignende organismer (pseudomycetes) er en del av kongeriket Straminopila, som også inkluderer fargede alger (Ochrophyta). Kongeriket av ekte sopp forener 4 divisjoner, forskjellig i strukturen til den vegetative kroppen og egenskapene til reproduksjon: Chytridiomycota, Zygomycota, Ascomycota, Basidiomycota.
Taksonomisk betydelige tegn, brukt til å bygge soppsystemer
– Strukturen til den vegetative kroppen
– Kjemisk sammensetning og struktur av celleveggen
– Type seksuell prosess
– Typer sporulering
Typer thalli av sopp og sopplignende organismer
Amøboid eller plasmodial Thallus er en mononukleær eller multinukleær celle blottet for en cellevegg, omgitt av en plasmamembran og et komprimert perifert proteinlag - periplasten. For å forankre i underlaget og øke absorpsjonsflaten kan det dannes rotlignende utvekster som til sammen danner rhizomyceliet. Under reproduksjonen blir enten hele thallus til et enkelt sporangium (holokarpisk thallus), eller det dannes flere sporangier (eukarpisk thallus).
Mycelium thallus og dens modifikasjoner. Grunnleggende strukturelt element– hyfer. Segmentert og ikke-segmentert mycel. Septa og deres dannelse.
Modifikasjoner av mycel.
Hvilestrukturer: klamydosporer og sklerotia.
Ledende og støttende strukturer: tråder og rhizomorfer.
Smittsomme strukturer: appressoria og haustoria
Gjærlignende spirende thallus representerer individuelle, membrandekkede celler eller ustabile, lett desintegrerende cellekjeder som dannes under spirende på grunn av ikke-divergens av mor- og datterceller. Permanent og midlertidig gjærlignende tilstand av thalli.
Kjemisk sammensetning og struktur av celleveggen til sopp
Celleveggen til sopp er flerlags, med forskjellige lag dannet av strukturelle karbohydrater som er forskjellige i deres kjemiske sammensetning, som kan deles inn i 3 grupper basert på deres kjemiske sammensetning:
glukosepolymerer(glukan, kitin, cellulose). Glukaner utgjør det ytre laget av celleveggen til de fleste sopp. Det indre laget av soppcelleveggen er dannet av kjeder av kitin, noe som gir den stivhet. Kitin erstatter cellulose, som er fraværende i de fleste sopp, men er en del av celleveggen til oomycetes, som foreløpig ikke tilhører typiske sopp. Deacetylert kitin kalles kitosan, som i kombinasjon med kitin danner celleveggen til zygomyceter.
polymerer av andre monosakkarider (mannose, galaktose, etc.) I motsetning til høyere planter, hvor de danner grunnlaget for matrisen under det generelle navnet hemicellulose, er de mindre karakteristiske for sopp. Unntaket er gjær, hvis cellevegger er spesielt rike på mannosepolymerer kalt mannaner. Det antas at denne veggsammensetningen bedre sikrer knoppskyting.
polymerer av karbohydrater kovalent knyttet til peptider (glykoproteiner) danner det midterste laget av en flerlags cellevegg og spiller en viktig rolle både for å opprettholde cellenes strukturelle integritet og i dens metabolske prosesser med miljøet.
Andre spesifikke trekk ved soppcellen inkluderer: fravær av plastider, som bringer den nærmere en dyrecelle;
Ingen stivelse, som i eumyceter er erstattet av et polysakkarid nær animalsk stivelsesglykogen, i oomyceter med et polysakkarid nær laminarin av brunalger. Det produseres også en rekke reservekarbohydrater som er spesifikke for sopp.
Utvikling av spesifikke sekundære metabolitter, hvorav antibiotika, fyto- og mykotoksiner og fytohormoner spiller en stor rolle.
De spesifikke egenskapene til sopp inkluderer også heterokaryose og paraseksuell prosess.
Hos sopp er fenomenet heterokaryose eller heteronuklearitet svært utbredt, der kjerner som er heteroalleliske for noen gener forblir i én celle i lang tid. Heterokaryose erstatter heterozygositet i haploide sopp og fremmer rask tilpasning av sopp til endrede forhold. Tilstedeværelsen av forskjellige kjerner skyldes en rekke spesifikke trekk ved sopp:
1. tilstedeværelsen av mer enn én kjerne i en celle
2. den spesifikke strukturen til celleskilleveggen, der det er ett eller flere gjennomgående hull, kalt porer, gjennom hvilke kjerner kan migrere fra en celle til en annen
3. Hyfer innenfor en koloni og til og med forskjellige tettliggende kolonier dyrket fra forskjellige sporer av samme type sopp vokser ofte sammen, som et resultat av at utveksling av kjerner av forskjellige stammer er mulig.
Paraseksuell (pseudofil) prosess. Hvis i heterokaryote celler kjerner som er heteroalleliske på et hvilket som helst sted smelter sammen, vises en heterozygot diploid kjerne. Det kan komme inn i en spore og gi opphav til en diploid heterozygot klon. Under mitoseprosessen kan diploide kjerner gå tilbake til haploid tilstand på grunn av tap av ett sett med kromosomer, eller en utveksling av kromosomseksjoner kan forekomme i dem (mitotisk kryssing). Begge prosessene er ledsaget av rekombinasjon av foreldregener og, følgelig, fenotyper. Paraseksuell (aseksuell) rekombinasjon er et svært sjeldent fenomen, som ikke overstiger en kjerne per million, men på grunn av det store antallet kjerner i myceliet, observeres det konstant i sopppopulasjoner.
Reproduksjon– vegetativ, aseksuell, seksuell.
Vegetativ– fragmentering av thallus, dannelse av klamydosporer, som etter en periode med dvale spirer til mycelium, spirende i gjær.
Aseksuell Reproduksjon i forskjellige sopp kan utføres av mobile og immobile sporer. Zoosporer danner en relativt liten gruppe sopp og sopplignende organismer - akvatiske og noen terrestriske, der genetiske forbindelser med akvatiske sopp og alger er godt synlige. Strukturen til flagellene er et viktig diagnostisk trekk når det tildeles et spesifikt rike. I det store flertallet av sopp, under aseksuell reproduksjon, dannes immobile sporer, noe som indikerer deres svært gamle fremvekst på land. I henhold til stedet for dannelse og lokalisering skilles endogene sporangiosporer, dannet i sporangier og eksogene (konidier), og utvikler seg på spesielle hyfer - konidioforer. Konidier dannes i de fleste sopp (ascomycetes, basidiomycetes, noen zygomycetes) som utgjør konidiesporulering, som er ekstremt variert og mye brukt for å diagnostisere sopp.
Seksuell reproduksjon av sopp har sine egne spesifikasjoner både i morfologien til den seksuelle prosessen og i mekanismene for genetisk og fysiologisk regulering av sex og overføring av arvelig informasjon.
Somatogami- den vanligste og enkleste typen seksuell prosess, består av fusjon av to somatiske celler som ikke er differensiert til kjønnsceller. Noen ganger skjer det selv uten cellefusjon - kjernene inne i cellen smelter sammen. Finnes i de fleste basidiomyceter, pungdyrgjær og noen andre taxaer.
Gametangiogami– gametangia er isolert på det haploide mycelet, og innholdet deres tappes under den seksuelle prosessen. Denne seksuelle prosessen er karakteristisk for de fleste pungdyrsopper. En variant av gametangiogami er zygogami hos zygomycetes.
Gametogami i form av iso-hetero og oogamy, forekommer det mye sjeldnere hos sopp enn hos andre eukaryoter. Iso- og heterogami forekommer bare hos chytridiomycetes. Klassisk oogami med dannelse av sæd og egg kommer ikke til uttrykk i sopp, men svært modifiserte varianter finnes.
I henhold til egenskapene til kjønnsregulering hos sopp, skilles flere typer seksuell prosess ut.
Gynandromixis kan vurderes ved å bruke eksemplet med toboende oomyceter, der oogonier og antheridia utvikler seg på forskjellige thalli, for eksempel sent blight eller potetsopp. Hvis en genetisk homogen stamme dyrkes i en monokultur, reproduserer den seg kun ukjønnet. Hvis mycelene til to stammer er i nærheten, kan morfogenetiske endringer spores i dem under påvirkning av kjemiske sekresjoner av steroidnatur - sexferomoner. Antheridiol induserer dannelsen av antheridia hos partneren, og oogoniol induserer dannelsen av oogonium. I dette tilfellet er reguleringen av sex relativ: om en stamme vil danne antheridia eller oogonia avhenger av det kvantitative forholdet mellom de tilsvarende feromonene i den og dens partner. Derav navnet på den seksuelle prosessen - gynandromixis.
Dimixis eller heterohallisme. Det har lenge vært bemerket at sopp kan være homo- eller heterotaliske. Hos homotalliske arter, under den seksuelle prosessen, smelter genetisk identiske kjerner i myceliet. Hos heterotalliske arter, for å gjennomgå den seksuelle syklusen, på et tidspunkt (forskjellig for forskjellige typer sopp), er fusjonen av sporenes etterkommere (mer presist deres kjerner) nødvendig. For at to stammer skal være seksuelt kompatible, må det være genetisk forskjell (heteroallelisitet) ved visse loci kalt paringsloki. De fleste sopp (zygomycetes, ascomycetes og noen basidiomycetes) har ett parringssted med to alleler. Parringsstedet består av flere gener som styrer syntesen av sexferomoner. Slik heterohallisme kalles unifaktoriell eller bipolar. Avkommet til slike sopp etter meiose er delt inn i to selvinkompatible, men gjensidig kompatible grupper i forholdet 1:1, dvs. sannsynligheten for relatert (innavl) og urelatert (utavl) krysning er, som i bifile høyere eukaryoter, 50 %.
I genomet til høyere basidiomyceter er det to parrende loci - A og B, og bare stammer som er heteroalleliske på begge lociene er kompatible med hverandre (Ax Bx er kompatibel med Ay By, men ikke med Ax By og Ay Bx). Slik heterohallisme kalles tofaktor eller tetrapolar. Det reduserer sjansen for innavl med opptil 25 %.
Diaphoromixis– høyere basidiomyceter har ikke to, men mange alleler av parringsstedet, som finnes tilfeldig i forskjellige stammer som utgjør populasjonen. Denne paringskontrollen sikrer 100 % sjanse for utavl, siden stammer med forskjellige alleler er gjensidig kompatible, og det er mange alleler. Som et resultat dannes panmix-hybridpopulasjoner.
Livssykluser til sopp like forskjellige som selve soppene. De viktigste syklusene, deres tilhørighet til avdelingene for sopp
1. Den aseksuelle syklusen er karakteristisk for en enorm gruppe ufullkomne eller mitofungi som har mistet seksuell reproduksjon. Delingen av kjernene deres er utelukkende mitotisk. De aller fleste mitofunger tilhører pungdyrsoppene, men på grunn av tapet av den seksuelle prosessen utgjør de en formell gruppe ufullkomne sopp eller deuteromycetes.
2. Haploid syklus. Den vegetative thallus bærer haploide kjerner. Etter den seksuelle prosessen (syngami) deler den diploide zygoten (vanligvis etter en hvileperiode) seg meiotisk - zygotisk meiose. Karakteristisk for zygomyceter og mange chytridiomycetes.
3. Den haploid-dikaryote syklusen er preget av det faktum at etter sammensmeltingen av innholdet av gametangia (gametangiogami) eller somatiske celler av haploid mycel (somatogami), danner kjernene dikaryoner (par av genetisk forskjellige kjerner). De deler seg synkront, og danner dikaryot mycel. Den seksuelle prosessen ender med fusjonen av dikaryonkjerner, den resulterende zygoten deler seg ved meiose uten hvileperiode. Meiosporer utgjør den seksuelle sporuleringen av pungdyr og basidiomyceter i form av ascosporer og basidiosporer. Når de spirer, dannes haploid mycel. Hos de aller fleste pungdyrsopper (unntatt gjær- og taphrinsopper) er livssyklusen dominert av den haploide fasen i form av vegetativt mycel (anamorf), den dikaryote fasen er kortvarig og representeres av askogene hyfer som poser på. dannes (teleomorf). Hos basidiomyceter dominerer den dikaryote fasen i livssyklusen den haploide fasen er kortvarig.
4. Den haploid-diploide syklusen i form av en isomorf generasjonsendring er sjelden hos sopp (noen gjærsopp og vannlevende chytridiomyceter).
5. Den diploide syklusen er karakteristisk for oomyceter og noen pungdyrgjær. Den vegetative thallus er diploid, gametisk meiose observeres under dannelsen av gametangia eller gameter.
3. Økologiske grupper av sopp.
Sopp og sopplignende organismer er inkludert i alle terrestriske og akvatiske økosystemer som den viktigste delen av den heterotrofe blokken, sammen med bakterier som okkuperer det trofiske nivået av nedbrytere. Den brede distribusjonen av sopp i biosfæren bestemmes av en rekke viktige funksjoner:
1. De fleste har en mycelstruktur av thallus. (lar deg raskt mestre underlaget og ha en stor kontaktflate med miljøet).
2. Høy hastighet på vekst og reproduksjon, som tillater på kort tid å kolonisere store masser av substrat, danne et stort antall sporer og distribuere dem over lange avstander.
3. Høy metabolsk aktivitet, manifestert i et bredt spekter av miljøfaktorer.
4. Høy grad av genetisk rekombinasjon, betydelig biokjemisk variasjon, miljømessig plastisitet.
5. Evnen til raskt å gå over til en tilstand av suspendert animasjon og overleve ugunstige forhold i lang tid.
Den viktigste miljøfaktoren for sopp er matsubstratet. I forhold til denne faktoren skilles hovedgruppene av sopp, som kalles trofiske grupper.
4. Kort beskrivelse av soppsnitt.
MIKROBIOLOGI, 2010, bind 79, nr. 6, s. 723-733
UDC 582.281(047)
SOPP-CELLEVEGG: MODERNE KONSEPT OM KOMPOSISJON OG BIOLOGISK FUNKSJON
© 2010 E. P. Feofilova1
Etablering Det russiske akademiet Sciences Institute of Microbiology oppkalt etter. S.N. Vinogradsky RAS, Moskva
Mottatt av redaktøren 05.11.2009
Gjennomgangen er viet den lite studerte overflatestrukturen til cellen av filamentøse sopp - celleveggen (CW). Data er gitt om isolasjonsmetoder og renhetstesting for fravær av cytoplasmatisk innhold i CS-fraksjonen og om dens kjemiske sammensetning. De strukturelle (rammene) og intrastrukturelle komponentene til CS - aminopolysakkarider, a- og b-glukaner, proteiner, lipider, uronsyrer, hydrofobiner, sporopollenin og melaniner - vurderes i detalj. Spesiell oppmerksomhet er gitt til kitin, dets ny funksjon i anti-stress beskyttelse av celler, så vel som forskjellene mellom dette aminopolysakkaridet av sopp og kitinet fra alger og leddyr. Fenomenet apikal hyfevekst og deltakelsen av spesielle mikrovesikler i morfogenesen til soppcellen diskuteres. Data er gitt om enzymene som er involvert i syntesen og lyseringen av CS. Til slutt diskuteres den funksjonelle betydningen av SC i sopp sammenlignet med overflatestrukturene til høyere eukaryoter.
Stikkord: filamentøse sopp, cellevegg, isolasjonsmetoder, kjemisk sammensetning, apikal vekst, fysiologiske funksjoner, morfogenese.
Et av hovedproblemene moderne biologi Spørsmålet er hvordan det under utviklingsprosessen skjer dannelsen av morfologisk identiske organismer, hvilke biokjemiske mekanismer og hvilke cellulære strukturer som er involvert i denne prosessen, som har funnet sted på planeten vår i milliarder av år. Ikke mindre interessant er dataene om hvordan stress påvirker morfogenesen og hvilke biopolymerer som kontrollerer ytre form celler. Som forskningsresultater viser senere år, er den voksende hyfen av filamentøse sopp en unik modell som lar oss forstå prosessen med celledannelse og hvordan cellemorfologiens konstanthet opprettholdes under ontogenese. Et veldig stort bidrag til studiet av dette fenomenet ble gitt av data om studiet av sammensetningen og biologisk funksjon CS av filamentøse sopp.
Plantenes CS ble oppdaget i 1665, og CS av sopp først på begynnelsen av 1700-tallet. Dette er imidlertid overfladisk cellestruktur I lang tid ble det praktisk talt ikke studert, siden det ble antatt at CS bare utførte en "ramme"-funksjon, og den ble sammenlignet med veggene i et hus som bærer en bærende belastning. Men på begynnelsen av 1900-tallet endret holdningen til CS seg dramatisk, det begynte å bli studert intensivt, men hovedsakelig i planter og bakterier. Den intensive utviklingen av soppdyrking og bioteknologisk produksjon, der trådformede sopp var produsenter av biologisk aktive stoffer, intensiverte imidlertid utviklingen
1 adresse for korrespondanse (e-post: [e-postbeskyttet]).
slips Vitenskapelig forskning om studiet av sopp-CS. Data om denne overflatestrukturen for perioden frem til 1980-tallet ble oppsummert i verdens første bok om sopp-CS. I de påfølgende årene skjedde hoveddelen av arbeidet på slutten av det 20. og begynnelsen av XXIårhundrer. Fundamentale studier har blitt utført på systematikken, kjemisk sammensetning av CS, apikal hyfevekst, kitinmetabolisme, soppdrepende legemidler, hydrofobiner, kovalente bundne proteiner, enzymer involvert i dannelsen av CS, hyphal forgrening og lysis av CS. Disse dataene er ikke generalisert, men det er diskusjonen av dem sammen som gjør det mulig å vise at denne overflatestrukturen til cellen bærer en multifunksjonell belastning og utfører slik essensielle funksjoner, for eksempel å beskytte cellen mot handlingen ugunstige faktorer, kontroll av morfogenese, deltakelse i reproduksjonsprosesser, bestemmelse av antigene og adhesive egenskaper, kontroll av prosessene for dimorfisme og dannelsen av hvilende soppceller, oppfatning av et eksternt signal og dets overføring til membranen og intracellulære budbringere. Tatt i betraktning det ovennevnte, anså vi det som hensiktsmessig å diskutere følgende data: den kjemiske sammensetningen av CS, den biologiske funksjonen til dens viktigste biopolymerer, strukturen til CS og intercellulære interaksjoner i sopphyfer, fenomenet apikal hyfevekst, og hovedenzymer involvert i dannelsen og lyseringen av CS.
KJEMISK SAMMENSETNING AV CELLULAR
VEGGER AV SVAMP OG BIOLOGISK FUNKSJON AV DERES HOVEDKOMPONENTER
Studerer kjemisk oppbygning CS av sopp begynner med å oppnå en ren fraksjon av denne strukturen, dvs. rensing av CS fra cytoplasmatisk forurensning. Den innledende prosedyren - ødeleggelse av soppceller og vasking av cellulært innhold - utføres ved lav temperatur (ca. 4-5 ° C) for å forhindre den destruktive aktiviteten til nedbrytende enzymer. Før cellene blir ødelagt, fryses de ved en temperatur flytende nitrogen. Celledestruksjon utføres enten i spesielle homogenisatorer eller i presser med " fast trykk Videre bruk av en ultralydsdesintegrator gir gode resultater. Celleinnholdet vaskes kaldt vann og gjentas 4-5 ganger. For mer grundig fjerning av cytoplasmatisk innhold, vask med NaCl, 8 M urea, 1 M ammonium eller 0,5 M eddiksyre. For tiden er det utviklet metoder for isolering av sopp-CSer under deres ontogenese, og disse metodene er betydelig forskjellige for mycel og hvileceller. Avhengig av forskningsretningen, spesielt ved bestemmelse av polysakkaridsammensetningen, ekstraheres lipider fra CS ved å bruke organiske løsningsmidler, for eksempel kloroform og metanol i forholdet 2: 1, noen ganger gode resultater gir ytterligere behandling av CS med svovelsyreeter. Dette løsemidlet tørker CS godt og kan noen ganger erstatte frysetørking. Neste nivå- bestemmelse av renheten til den resulterende CS-fraksjonen. For dette formålet, lys og elektronmikroskopi og spesialiserte maleteknikker. Den mest brukte reaksjonen er basert på interaksjonen mellom I3- og kitosan. Isolert ren CS med Lugols løsning er farget rosa eller lilla, mens intakte CS er røde i fargen. Farging for tilstedeværelse av kjerner med DAPI (4,6-ekt1to-2-rkepyTsn-yo1) brukes også. Det bør spesielt understrekes at isolering av en ren fraksjon av CS er en svært ansvarlig prosedyre, som påfølgende resultater av analysen av dens kjemiske sammensetning avhenger av. I denne forbindelse er et interessant arbeid der en glukan, eller mer presist et kitosan-glukankompleks, ble funnet i myceliet til en representant for slimsopp, selv om alle tidligere studier indikerte fravær av glukan i myceliet til Mucorales.
For tiden er komponentene i sopp-CS delt inn i strukturelle komponenter (kitin, p-(1-3)-p(1-6)-glukaner, p-(1-4)-glukan (cellulose) og intrastrukturelle (de er kalt matrise), som begynte å inkludere mannoproteiner, galakto-mannoproteiner, xylo-mannoproteiner, glukurono-
mannoproteiner og a-(1-3)-glukan. De minst studerte er a-glukaner, som er en lineær polymer av glukose (i Schizosaccharomyces pombe inneholder denne glukanen omtrent 260 glukoserester). Denne polymeren består av to koblede lineære kjeder som inneholder ca. 120 (1-3)-a-D-glukoserester og (1-4)-a-D-glukoserester ved endene av polymermolekylet. Det antas at denne glukanen er nødvendig for prosessen med morfogenese. En vannuløselig glukan ble isolert fra CS av Penicillium roqueforti mycelium, som etter tørking mister sin evne til å løse seg i alkali, men den var i stand til å løses opp i en 10 % løsning av litiumklorid i dimetylsulfoksid. Når man studerer glukan ved metylering, ble det oppnådd to tri-O-metyl-derivater av glukose, tilsvarende 1 - 3 og 1 - "- 4-bindingene mellom monosakkarider i et forhold på ca. 5: 2, og spor av en tetra-O -metylderivat som tilsvarer de terminale ikke-reduserende mono-nosakkarid-restene. Dette resultatet indikerer lineær struktur molekyler som inneholder bare 1 -»- 3 og 1 -»- 4 bindinger mellom glukoserester. Disse dataene ble bekreftet ved analyse av 13C-NMR-spekteret til polysakkaridet, hvorfra a-konfigurasjonen av alle glukoserester også følger. Slike polysakkarider, spesielt etter kjemisk modifikasjon (sulfatering eller karbosimetylering), som kan gjøre dem oppløselige i vann, kan brukes som biologisk aktive polymerer og er av interesse for å studere deres biologiske funksjon i celleveggen til sopp og planter.
Vannuløselige (1 -»- 3)-α-glukaner ble tidligere isolert fra flere arter av høyere og lavere sopp, inkludert Penicillium chrysogenum, og beslektede α-glukaner med to typer bindinger (1-3) og (1 -» - 4), forskjellig i forholdet, ble funnet i høyere planter(Aconitum kusnezoffii Reichb).
Strukturell komponent- cellulose - karakteristisk for oomycete-sopp, som ifølge moderne taksonomi er klassifisert som pseudofungi. Monosakkaridsammensetningen til CS inkluderer glukose, mannose, xylose, og det dominerende sukkeret er glukose, som utgjør opptil 68%. Sammensetningen av sopp-CS inkluderer også aminosyrer, lipider (ikke mer enn 3%) og N-acetyl-glukosamin. Glukaner, som 1,3-glukan, danner et sterkt kompleks med kitin, kalt kitin-glukan-komplekset (CGC), som utgjør skjelettet til soppcellen. Dette komplekset er tilstede i CS av nesten alle sopp, med unntak av zygomycetes, som fungerer som en pålitelig systematisk funksjon. Hos noen sopp ble det funnet et slimete materiale bestående av polysakkarider representert av ß-1,3-glukaner som kun inneholder glukoseenheter forbundet med ß-1,6-bindinger til hver tredje glukoseenhet på overflaten av CS.