Planteceller er omgitt av en tett polysakkaridmembran, foret på innsiden med plasmalemma.
utdanning celleveggen oppstår under metafase og telofase av celledeling. I ekvatorial sone deling vises en midtplate, bestående av kalsiumpektat, som vokser fra sentrum til periferien og skiller en nydannet celle fra en annen. Midtplaten er dekket på begge sider av primærcelleveggen. Vekst i tykkelse oppstår på grunn av pålegging av nye lag fra innholdet i hver celle. Cellevekst i lengde begynner med at matrisen løsner. I denne prosessen viktig rolle fytohormoner spiller. Nye deler av materiale som celleveggen er bygget av kommer inn i de dannede hulrommene. Syntesen og transporten av disse stoffene utføres hovedsakelig av vakuoler fra Golgi-apparatet.
Celleveggen til celler som deler seg og vokser i forlengelse kalles hoved. Etter at celleveksten stopper, avsettes nye lag på den primære celleveggen fra innsiden og en sterk sekundær celleveggen.
Celleveggen inneholder strukturelle komponenter(cellulose i planter, kitin i sopp), veggmatrisekomponenter (hemicelluloser, pektin, proteiner), encrusting komponenter (lignin, suberin) og stoffer avsatt på veggoverflaten (kutin og voks). Cellevegger kan også inneholde kalsiumsilikater og karbonater.
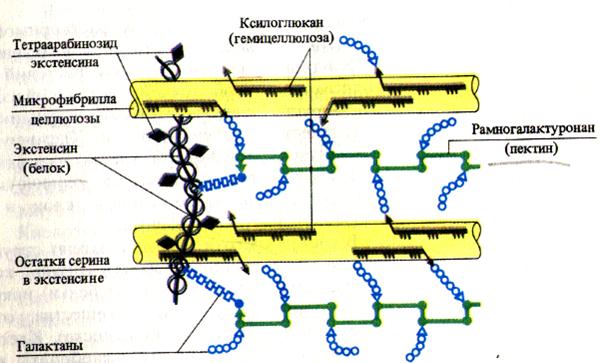
Cellulose(b-D-glukosepolymer), hemicelluloser(polymerer av heksoser og pentoser) og pektinstoffer(uronsyrederivater) er karbohydratkomponenter i celleveggene. Cellulose og pektinstoffer adsorberer vann, og gir hydrering til celleveggen. Pektinstoffer som inneholder mange karboksylgrupper binder toverdige metallioner, som kan byttes ut med andre kationer (H+, K+, etc.). Dette bestemmer kationbytterkapasiteten til plantecelleveggene. I tillegg til karbohydratkomponenter inkluderer celleveggmatrisen også et strukturelt protein kalt extensin. Dette er et glykoprotein som inneholder mer enn 20 % L-hydroksyprolin av de totale aminosyrene. I henhold til denne funksjonen ligner proteinet til plantecellevegger det intercellulære proteinet til dyr - kollagen.
Cellulose: A – strukturen til cellulosemolekylet; B – cellulosemolekylassosiasjoner: 1 – micelle, 2 – mikrofibril, 3 – makrofibril
Celleveggens hovedinnkapsling er lignin Intensiv lignifisering av cellevegger begynner etter at celleveksten opphører. Lignin er en polymer med et uforgrenet molekyl bestående av aromatiske alkoholer (p-kumarsyre, koniferyl, synapisk). Ødeleggelsen og kondenseringen av lignin i jorda er en av faktorene i dannelsen av humus. Intensiv lignifisering (impregnering av celluloselag med lignin) av cellemembraner begynner etter at celleveksten opphører. Lignin kan avsettes i separate seksjoner - i form av ringer, spiraler eller et nettverk, som observert i celleveggene til ledende vev - xylem, eller i et sammenhengende lag, med unntak av de stedene hvor kontakter er laget mellom naboer. celler i form av plasmodesmata. Lignin holder cellulosefibre sammen og fungerer som et veldig hardt og stivt stillas, som øker strekk- og trykkstyrken til celleveggene. Det gir også cellene ekstra beskyttelse mot fysisk og kjemiske påvirkninger, reduserer vannpermeabiliteten. Lignininnholdet i skallet når 30%. Encrustation av cellemembraner med det fører til deres lignifisering, som ofte medfører død av det levende innholdet i cellen. Lignin kombinert med cellulose gir spesielle egenskaper tre, som gjør det til et uunnværlig byggemateriale.
I regulering av vann og termisk regime planter involverer vev hvis cellevegger er impregnert suberin. Avsetningen av suberin gjør veggene vanskelig å permeable for vann og løsninger (for eksempel i endoderm, periderm). Suberin avsettes på skallet fra innsiden og gjør det praktisk talt ugjennomtrengelig for vann og løsninger. Som et resultat dør celleprotoplasten og cellen fylles med luft. Denne prosessen kalles suberisering. Suberisering av cellemembranen observeres i integumentært vev flerårige treaktige planter - periderm, skorpe, og også i endodermis av roten. Suberin er hovedstoffet i korkformasjoner, som disse formasjonene hovedsakelig skylder sine egenskaper: ugjennomtrengelig for vann, gasser og lav varmeledningsevne.
Overflaten til planteepidermale celler er beskyttet av hydrofobe stoffer - cutin Og voks: Forløperne til disse forbindelsene skilles ut fra cytoplasmaet til overflaten, hvor deres polymerisering skjer. Kutinlaget penetreres vanligvis av polysakkaridveggkomponenter (cellulose, pektin) og danner neglebåndet. Kutikula er involvert i reguleringen av vannregimet til vev og beskytter celler mot skade og infeksjon.
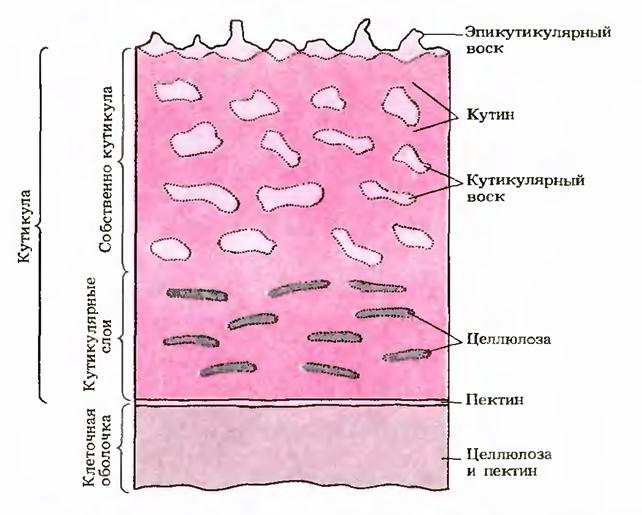
Akkumulerer i membranene til epidermalcellene til noen planter (korn, frø, etc.) et stort nummer av mineraler (mineralisering), primært kalsiumkarbonat og silika. Når de mineraliseres, blir bladene og stilkene til planter tøffe, harde og mindre sannsynlighet for å bli spist av dyr.
I primære cellevegger utgjør cellulose opptil 30 % av veggens tørre masse. Mengden hemicelluloser og pektinstoffer varierer avhengig av objektet. Sammen med proteiner kan pektinstoffer utgjøre omtrent 30 % av cellens tørre masse, med proteinmengden på 5 – 10 %. Omtrent 40 % kommer fra hemicellulose.
1) Celleveggen - strukturell utdanning. Funksjon: gir styrke og form, beskytter protoplasten mot ytre forhold, deltar i ledning og absorpsjon av stoffer.
Grunnlaget for cellemembranen (sammensetningen) er høypolymerkarbohydrater (cellulose, dvs. fiber - fordøyes ikke, indikerer lav produktivitet), cellulosemolekyler samles i komplekse bunter (mycelier), mycelier kombineres til fibriller, deres rom er fylt med hemicellulose (halvfiber - mindre stabil forbindelse) og pektin (nyttig, svelle i vann, er en energikilde).
Det er primære og sekundære cellemembraner. Meristematiske og unge voksende celler har primær celle skall, tynt, rikt på pektin og hemicellulose; Cellulosefibriller i matrisen til den primære celleveggen er ordnet på en uordnet måte.
Sekundær cellulær skallet dannes vanligvis når cellen når sin endelige størrelse og er lagt over lag på den primære fra protoplastsiden. I den sekundære cellemembranen dominerer cellulose dens fibriller på en ordnet, parallell måte, men deres retning i hvert lag er forskjellig, noe som øker styrken til cellemembranen. I den sekundære celleveggen er det åpninger (porer), hvor cellene er separert kun av den primære celleveggen og plasmodesmata (cytoplasmatiske broer som forbinder naboplanteceller).
Celleveggmodifikasjoner:
- Lignifisering av cellemembranen skjer som et resultat av avsetning av lignin (en ikke-karbohydratkomponent i fibrillerne mister sin elastisitet, men kan tillate vann å passere gjennom). Disse cellene er oftere døde enn levende. Noen cellevegger kan inkludere: voks, cutina, suberin. Funksjoner: gir cellen form; skiller en celle fra en annen, er skjelettet for hver celle og gir styrke til hele planten, utfører en beskyttende funksjon.
- Korkdannelse er forårsaket av et spesielt fettlignende stoff - suberin. Slike skjell blir ugjennomtrengelige for vann og gasser, de lar heller ikke varme passere gjennom innholdet i celler med suberiserte skall.
- Cutinisering innebærer frigjøring av det fettlignende stoffet cutin. Vanligvis er de ytre veggene av huden på blader og urteaktige stilker kutiniserte. Dette gjør dem mindre gjennomtrengelige for vann, reduserer fordampning av vann i planter, og beskytter dem mot overoppheting og ultrafiolett stråling kalt kutikula.
- Mineralisering av cellemembraner er avsetning av: silika og kalsiumsalter. Cellemembranene i huden til blader og stilker av frokostblandinger, korn og kjerringrokk er mest belagte. Bladene til frokostblandinger og korn kan skade hendene dine.
- Slimdannelse av skjell er omdannelsen av cellulose og pektinstoffer til slim og tannkjøtt. Slim er tydelig observert på linfrø som var i vann. Dannelsen av slim fremmer bedre absorpsjon av vann av frøene og deres feste til jorda.
2) Reproduksjon: evnen til et enkelt individ til å gi opphav til en hel serie av sitt eget slag.
Delt inn i: seksuell og aseksuell (riktig aseksuell og vegetativ)
Vegetativ: nye individer utvikler seg fra individuelle vegetative organer eller deres interaksjoner. Det utføres takket være regenerering (evnen til å gjenopprette en organisme fra en del av kroppen). Biologisk betydning: den nye organismen ligner den mors.
Metoder for vegetativ formering:
- forplantning ved stiklinger (en del av planten som ikke er infisert er plantet i et substrat, sporodina),
- formering ved poding (ved å spire deler av flere planter, brukt i hagearbeid),
- forplantning med knoller (kjøttfulle knoller med pita plantes i bakken, viviparous bokhvete),
- forplantning av avkom (danner skudd på røtter, osp),
- forplantning med løker (om høsten plantes de fra selve planten ned i bakken)
- formering ved hjelp av ranker (krypende skudd, roting, drupes, jordbær)
- forplantning av jordstengler (underjordiske skudd, pitabrød, liljekonvall, fiolett, hvetegress)
Bruk av vegetativ forplantning av mennesker. Resten er 40 cm.
MED i lang tid mennesker, som dyrket planter, begynte å bruke vegetativ forplantning. For eksempel å vokse poteter, jordbær, banan i alle land i verden utføres det bare med vegetative midler - knoller, ranker og rhizomer.
Bruken av vegetativ reproduksjon av planter i landbrukspraksis kalles kunstig vegetativ forplantning.
De viktigste metodene for kunstig vegetativ forplantning kommer ned til å gjenta de som forekommer i planter under naturlige forhold.
Folk bruker ofte forplantning ved stiklinger - deler av grønne eller treaktige skudd (druer, rips, stikkelsbær, roser, nellik, ficus), knoller (poteter, dahlia, søtpotet, jordskokk), blader (saintpaulia, gloxinia, begonia), pærer (løk, hvitløk, tulipan, påskelilje), dele busken (rips, pyrethrum) og lagdeling (stikkelsbær, kaprifol, klematis), bart (jordbær), jordstengler (sukkerrør, iris, floks), rotskudd (plomme, bringebær, kirsebær, syrin).
3) Gresskar. Form: urter. Trykk på roten. Stengel: klatrende, krypende, klatrende Blad: enkel, petiolate, uten stipler.
Formel: tobo
1) vanlig hunn Ca (5) Co (5) A 0 G (3) perianth under eggstokken
2) korrekt hann Ca (5) Co (5) A 2+2+1 G 0
Blomsterstanden er enslig. Frukt: gresskar
Representanter: agurk, melon, gresskar, vannmelon, zucchini
Betydning: mat, fôr
Cellemembran i stand til å tykne og modifisere. Som et resultat av dette dannes et slør sekundær struktur. Fortykkelse av membranen skjer ved å legge nye lag på nervemembranen. På grunn av at pålegget allerede er i gang hardt skall, cellulosefibriller i hvert lag ligger parallelt, og i tilstøtende lag - i en vinkel til hverandre. Dette oppnår betydelig styrke og hardhet til det sekundære skallet. Ettersom antall cellulosefibrilllag øker og veggtykkelsen øker, mister den sin elastisitet og evne til å vokse. I den sekundære celleveggen øker celluloseinnholdet betydelig, i noen tilfeller opptil 60 % eller mer. Når cellene fortsetter å eldes, kan membranmatrisen fylles opp ulike stoffer- lignin, suberin (lignifisering eller suberisering av skallet). Lignin dannes av hemicellulose og pektinstoffer.[...]
Celleveggen til trefiber har flere lag: primær, som kalles fiberens ytre skall, og sekundær (veggen, som igjen består av tre lag: ytre, midtre og indre). Mellom de primære celleveggene er det et lag av intercellulær substans, gjennom hvilken fibrene er forbundet med hverandre. Sekundærveggen er relativt tykk og representerer hoveddelen av cellevolumet.[...]
I de sekundære lagene av celleveggene av furu akkumulert i store mengder mannaner (22 %) og uronsyreanhydrid (25 %).[...]
[ ...]
Cellevegg fortykningsfase. Hvordan oppstår fortykning? I vekstperioden er protoplasten kun omgitt av primærveggen. Når når en trecelle sin største størrelse langs overflaten eller kort tid etter blir celleveggen tykkere. Dette er forårsaket av lagdeling av sekundærveggen på primæren, og dette nytt lag oppstår som et resultat videre aktiviteter protoplast inne i cellehulen. Celler der protoplasten har forsvunnet kan naturligvis ikke fortsette å tykne veggene sine. Dannelsen av en sekundærvegg er et tegn på en irreversibel endring i cellen, hvis videre vekst allerede er utelukket, men ytterligere deling er ikke nødvendigvis utelukket, forutsatt at dattercellene som oppnås på denne måten opptar samme volum som den opprinnelige cellen. [...]
M.1ip - teppe, sengeteppe). Den består av tabellformede, tynnveggede celler med tett cytoplasma. Det er vanligvis enkeltrad, men noen ganger dobbeltrad eller flerrad. Notoma-celler er i utgangspunktet mononukleære, men senere blir de ofte binukleære eller til og med flerkjernede. Tapetum er et fysiologisk ekstremt aktivt vev: cellene inneholder enzymer, hormoner og næringsmateriale som brukes i prosessen med mikrosporogese. Det er noen grunner til å anse den sekretoriske typen som primær i evolusjonære termer, og amøboidtypen som sekundær.[...]
Det skal imidlertid bemerkes at disse dataene bør betraktes som omtrentlige, siden de opprinnelige preparatene ikke ble grundig renset.[...]
Det er vanskelig å bestemme celleveggplasseringen til polyuronidhemicelluloser fordi reagensene som brukes til å identifisere dem også påvirker lignin. Noen forskere antyder at hemicellulose er det sementerende stoffet mellom fibrillene og de ulike lagene i celleveggen. Cohen mener selv at sekundærveggen lignin har samme natur med hemicelluloser. Grunnlaget for denne antakelsen ser ut til å være det faktum at noen karbohydrater, når de behandles sterke syrer kan produsere uløselige rester av et spesifikt mønster. Det skal imidlertid understrekes at områder, både nøye behandlet med reagenser som løser opp hemicelluloser og ikke behandlet med dem, gir rester med svært lik struktur når de utsettes for 72 % svovelsyre.[...]
For å klargjøre sammensetningen av individuelle lag av cellevegger ble det gjort et forsøk kvantifisering xyloronider i forskjellige lag av trakeider og libriforme. Det ble målt på fibre fra japansk rødfuru, europeisk gran, bøk og bjørk. For dette formål ble fibrene forsiktig nitreret i et medium av eddiksyreanhydrid og karbontetraklorid. Deretter ble det ytre nitrerte laget fjernet ved oppløsning i aceton, hvoretter innholdet av pentosaner i resten ble kontrollert med furfural. Det ble funnet at pentosaner i trefibre er ujevnt delt i lag. Største kvantum pentosaner finnes i de ytre lagene av fibre og deres konsentrasjon avtar fra periferien til sentrum. Dermed inneholder de ytre lagene av bartrefibre 50-80 % pentosaner, mens løvtre inneholder nesten 100 %. I de sekundære lagene av celleveggene til bartrær viste det seg at innholdet av pentosaner ikke var mer enn 2-4%, og i løvtrær var det 8-10%. Dermed, kjemisk metode bekreftet resultatene oppnådd tidligere ved sorpsjonsmetoden for ultrafiolett lys.[...]
Det skilles mellom primær lignin, lokalisert i lignifiserte cellevegger (naturlig lignin) og sekundær lignin – isolert lignin. Sistnevnte er i stor grad et stoff som er modifisert under isolasjonsprosessen og forurenset med urenheter av fremmede stoffer. En endring i lignin kommer til uttrykk i eliminering av metoksylgrupper, intramolekylær kondensasjon og andre funksjoner.[...]
Mange forskjeller mellom vevstyper skyldes strukturen til celleveggen, spesielt den sekundære. Som vi allerede har sagt, skjer dannelsen av den primære celleveggen under prosessen med celleforlengelse, og derfor må den ha egenskapen å utvide seg, mens den sekundære veggen dannes etter at forlengelsen har stoppet.[...]
Preston
Samtidig med disse interne endringer den ytre harde veggen av oosporen deler seg ved toppen i fem tenner, og gir opphav til en frøplante som kommer ut av sentralcellen (fig. 269, 3). Den første delingen av sentralcellen skjer av en tverrskillevegg vinkelrett på dens langakse og fører til dannelsen av to funksjonelle ulike celler. Fra en, større celle dannes det deretter et stengelskudd, som det første stadiet utvikling kalles en pre-voksen, fra en annen, mindre celle - den første rhizoiden. Begge vokser ved tverrgående celledelinger. Den pre-voksne vokser oppover og blir grønn ganske raskt, fylles med kloroplaster går ned og forblir fargeløs (fig. 269, 4). Etter en serie celledelinger, som gir dem strukturen til enkeltradsfilamenter, skjer deres differensiering til noder og internoder, og deres videre apikale vekst fortsetter som beskrevet ovenfor for stammen. Fra nodene til forveksten oppstår sekundære forskudd, hvirvler av blader og laterale grener av stilken, fra nodene til den første rhizoiden - sekundære rhizoider og deres hvirvlende hår. På denne måten dannes en thallus bestående av flere stengelskudd i øvre del og flere komplekse rhizoider i nedre del (fig. 2G9, 5).[...]
Supramolekylær struktur. Figur 6.10 viser en modell av strukturen til celleveggen. Den inkluderer 2 hovedlag: den primære veggen P og den sekundære Sistnevnte er delt inn i 3 lag: 5], 5, Lag M, midtplaten, er en intercellulær substans som forbinder celler med hverandre.[...]
I påfølgende avsnitt (del II) vil kjemien til cellevegger bli grundig diskutert, relative mengder lignin i dem og andre relaterte temaer. Imidlertid avslutter vurderingen av den fjerde og siste fasen av ontogenese vedcelle, er det verdt å nevne noen fenomener som på en eller annen måte er forbundet med lignifisering, som botanikere kaller det. I likhet med dannelsen og spredningen av celler, så vel som fortykkelsen av celleveggene, kan lignifisering bare skje i løpet av den cellulære protoplastens levetid, siden døde celler ikke kan lignifisere veggene deres. Lignifiseringsprosessen kan fullføres i laget av intercellulær substans og i primærveggen, men kan fortsette i sekundærveggen, selv om dette sistnevnte laget fortsatt øker sentripetalt i tykkelse. I treved ender lignifiseringen ofte svært raskt i laget som grenser til innsiden av kambiumet, vanligvis nesten samtidig med det tidspunktet når de nye cellene har nådd sin største størrelse og sekundærveggene har nådd sin endelige tykkelse. Dette forklarer hvorfor spindelved, med samme fuktighetsinnhold, er like sterk eller nesten like sterk som kjerneved.[...]
Detaljert studie av fordelingen av lignin og polysakkarider i de lignifiserte celleveggene til gran- og bjørkeved ved å måle absorpsjonsintensiteten til en tynn stråle ultrafiolette stråler når de passerte dem gjennom en gjennomsiktig seksjon, bekreftet den den dominerende plasseringen av lignin i midtplaten og primærveggen, så vel som delvis i de ytre lagene av sekundærveggen. I midtplaten av granved når lignininnholdet 73%, og i sekundærveggen - ikke mer enn 16%. Det følger at polysakkarider konsentreres hovedsakelig i det sekundære laget. Det ble forsøkt å måle ved denne metoden gjensidig ordning cellulose og hemicelluloser. For å gjøre dette ble polysakkarider først omdannet til fargede forbindelser som absorberer lys.[...]
I de fleste celler er vekslende soner med større eller mindre ligninavsetning tydelig synlige, noe som gir utseendet til konsentriske ringer. I motsatt prosess, når celleveggen behandles med delignifiserende midler. reagenser, forblir cellulosemønsteret det samme. Dette indikerer at det ser ut til å være to interpenetrerende systemer, det ene består av cellulose og andre polysakkarider, og det andre av lignin. Bailey og Kerr viste at partikkelstørrelser når 0,1¡x og mindre. Hullene eller båndene forklarer de relativt store "fibrillerne" som noen forskere har sett. I tillegg til de dominerende konsentriske mønstrene, viser fibrene til noen tresorter et arrangement av radielle linjer eller en kombinasjon av begge typer. Celler av komprimert tre har ofte seige, nesten solide bånd av lignin nær cellehulen og radielt anordnede plater av det, atskilt av soner med polysakkaridstoff, i den midtre delen av celleveggen.[...]
Lav inneholder mange grunnstoffer og stoffer. Alle kan deles i to store grupper- primær og sekundær. Primære stoffer inkluderer de stoffene som direkte deltar i cellulær metabolisme; Kroppen av lav er bygget av dem. Sekundære produkter inkluderer sluttproduktene av metabolisme, vanligvis plassert på veggene til hyfene. Mange av disse sekundære lavstoffene (i eldre litteratur ble de kalt lavsyrer) er spesifikke for lav og finnes ikke i organismer fra andre systematiske grupper.[...]
Ritter, Lüdtke, et al. rapporterte at når trefibre behandles med forskjellige svellemidler, desintegrerer sekundærveggen (og sannsynligvis også primærveggen) til trådlignende fragmenter eller fibriller. Ritter delte disse fibrillene inn i spindelformede legemer, og disse igjen i sfæriske enheter. Betydningen av slike relativt store strukturelle enheter(lengden på de fusiforme legemene er ca. 4[x) er uklar, på grunn av den fint porøse strukturen til sekundærveggen beskrevet ovenfor. Verken ligninrestene etter celluloseoppløsning eller celluloserestene etter ligninoppløsning viser noen merkbare hull som indikerer grensene til disse celleveggenhetene. I tillegg har nyere studier ved bruk av et elektronmikroskop ikke påvist tilstedeværelsen av så relativt store enheter i strukturen til celleveggene.[...]
Ved vurdering av effekten av ulike treråtnende sopp på plantevev det er nødvendig å ta hensyn til at deres individuelle hyfer. bevege seg selektivt gjennom cellevegger. Hvitråtesopper foretrekker således midtplaten og primærskallet, hvor lignin hovedsakelig er konsentrert. Rød eller brun råtesopp, tvert imot, foretrekker å passere gjennom det sekundære skallet, som er rikest på karbohydrater. Følgelig er fargen på treet som er skadet av dem også forskjellig. Disse spørsmålene vil bli diskutert mer detaljert senere.[...]
Studier av trakeider og libriformer ved bruk av et polariserende og elektronmikroskop, samt radiografi, har etablert eksistensen av fem konsentriske lag i celleveggene: den ytre eller primære veggen og sekundærveggen. Sekundærveggen er igjen delt i tre lag, vanligvis betegnet 81, vg og B3. I tillegg er det mellom primærveggene til nabocellene en midtplate som limer dem sammen (fig. 35).[...]
Økningen i utbytte ved bruk av vanndamp forklares av det faktum at fjerningen av verdifulle produkter fra reaksjonsrommet akselereres og utviklingen av sekundære dekomponeringsreaksjoner forsinkes. I tillegg, når vanndamp kommer i kontakt med kapillærsystemet til tre på overflatelagene, er dampkondensering mulig, noe som skaper forhold for termisk dekomponering i surt vannmiljø. I dette tilfellet skjer nedbrytningsreaksjoner først og fremst i lagene av celleveggen, som er lokalisert med indre sider cellehulrom og består overveiende av ikke-varmebestandige hemicelluloser, som lett spalter acetylgrupper og deler av metoksylene knyttet til dem, og danner hhv. eddiksyre og metylalkohol.[...]
Det er neppe riktig å kalle segmentene som utgjør filamentene til kulecellene celler, ikke bare fordi de har mange kjerner og kloroplaster (og derfor tydeligvis er sekundære formasjoner), men også fordi de tverrgående partisjonene som skiller dem er ikke like. til celleveggene til andre flercellede grønnalger. De varierer mye i form, så vel som i metode og sted for dannelse (fig. 226, 4-6). Ofte har tverrgående skillevegger form av ringformede indre fortykkelser på celleveggene som ikke lukker seg i midten, slik at det blir igjen et hull som den cytoplasmatiske tråden går gjennom (fig. 226, 4). I andre tilfeller, i stedet for skillevegger, dannes spesielle plugger. Og til slutt kan grupper av radielt konvergerende snorer dukke opp hvor som helst i tråden, som ligner skjelettstrengene til caulerpa og spiller en mekanisk rolle.
Utenfor fra plasmamembran cellene deres har ikke en ekstra tett cellevegg eller den består av kitin, sjelden cellulose. Lagringskarbohydrater er vanligvis i form av glykogen (animalsk stivelse).[...]
Marx-Figipi og Pepcel studerte endringen i DP av bomullsmasse ved ulike stadier modning av bomull. De viste at viskositeten til bomullscelluloseløsninger sank flere timer etter åpning av esken. Cellulose av den sekundære celleveggen i fibrene til uåpnede bomullsboller liten modenhet(celluloseutbytte - 18%) har et enkelt maksimum på distribusjonskurven ved DP 14.000. Omtrent 10% av materialet har en lavere molekylvekt (DP 1500-2500), denne cellulosen finnes i den primære celleveggen.[.. .]
Plasseringen av stedene for dannelse av mikrofibriller i forhold til overflaten av den cytoplasmatiske membranen kan være forskjellig. I bakterier skjer denne prosessen i et miljø som er betydelig fjernet fra celleoverflaten og derfor fra membranen. Syntesen foregår tilsynelatende på lignende måte i de fortykkede primærveggene til epidermalcellene til havrekoleoptiler, siden cellulosesyntese i dette tilfellet skjer jevnt over hele celleveggens tykkelse. I membranene til ascidians forekommer tilsynelatende celluloseavsetning også på steder fjernt fra overflaten av sekretoriske celler, selv om det ikke er tilstrekkelig overbevisende bevis for denne antakelsen. Tvert imot, mikrofibriller av sekundære plantecellevegger er sannsynligvis dannet på den indre overflaten av veggen, i nærhet fra den cytoplasmatiske membranen. Siden det er mye mer cellulose i sekundærveggene enn i primærveggene, kan det konkluderes med at flertallet av cellulosemikrofibriller dannes nær den cytoplasmatiske membranen. Dette er imidlertid ikke obligatorisk.[...]
En av metodene basert på dette prinsippet er metoden for å bestemme reaktivitet cellulose i henhold til mønsteret for svelling av xanthater i isopropylalkohol. Svelleprosessen under samspillet mellom fiber og løsningsmiddel kan representeres skjematisk på følgende måte: Væske trenger inn i fiberen og får volumet til fiberen til å øke. Da sprekker det svake elastiske ytre laget av den sekundære celleveggen til fiberen og hevelser ("perler") dannes på bruddstedene. Restene av dette laget danner innsnevringer og mansjetter på den hovne fiberen. Deretter separeres det ytre laget og fiberen svulmer jevnt, det dannes tverrstriper på den og fiberen deles i pakker med skiver og separate disker, som deretter oppløses.[...]
Avhengighet av trestyrke på fuktighetsinnhold. Siden styrken og stivheten til tre delvis bestemmes av kohesive krefter som binder molekylene sammen, endrer ethvert middel som reduserer disse kreftene sin totale styrke. Et slikt middel er vann, så styrken til treet øker når fuktighetsinnholdet avtar, ikke bare som følge av den økte tettheten som følge av krymping, men også på grunn av tilstedeværelsen av sekundære valenskohesive krefter1. Siden tilstedeværelsen av vann i en mengde som overstiger metningspunktet til fiberen ikke endrer celleveggens natur, har tap eller oppsamling av kapillært (fritt) vann praktisk talt ingen effekt på treets styrke.
Strukturer som inneholder mye lignin er mørkebrune til svarte, mens svakt lignifiserte områder er lysegule til ravgule. Resultatene av denne fargereaksjonen bekrefter fullt ut tidligere arbeid med celleveggkjemi. Sekundærveggene til de fibrøse elementene i hardtre som vokser i tempererte klimaer er lysere i fargen og derfor mindre lignifisert enn sekundærveggene til bartre. Veggene i blodårene i løvtrær er mer farget mørk farge enn de omkringliggende fibrøse elementene, derfor inneholder de mer lignin; poremembranene er også sterkt lignifiserte.[...]
Denne operasjonen ble utført på lignifiserte seksjoner, tidligere frigjort fra lignin ved bruk av natriumkloritt i et eddiksyremedium. Seksjonene ble deretter behandlet med p-fenylase; benzoylklorid for forestring av polysakkarider. Sterkt farget oransje-rød farge Seksjonene ble fotometeret etter svelling i pyridin. Ved å utsette seksjoner bestående av holocellulose for slik behandling før og etter fjerning av hemicelluloser, kunne man konstatere at hoveddelen av hemicelluloser i gran- og bjørkeved er konsentrert i de ytre lagene av sekundærveggen. Således, ved utvinning av et snitt av gran holocellulose, 16% nsh kaustisk soda Det ble funnet at opptil 60-80 % av de alkaliløselige hemicellulosene av den totale mengden polysakkarider ekstraheres fra de ytre lagene av cellen, ca. 50 % fra midten av celleveggen og fra B3-laget. Et lignende bilde ble observert for tverrsnitt av libriform fra bjørkeved.[...]
Eksperimenter av Ritter, og senere av Bailey et al. viste at, uavhengig av mulig tilstedeværelse av pektiske polyuronider i midtplaten, består den hovedsakelig av lignin, slik kjemikere forstår det (uløselig i kald 72 % svovelsyre, løselig etter klorering og behandling svake grunner eller basiske salter). Dessuten beviste Ritter det mest av lignin ligger i dette laget. Denne uttalelsen var i strid med det rådende synet på den tiden om at det meste av ligninet var tilstede i andre lag, spesielt i sekundærveggen. Det ble senere bevist at i slike tilfeller er den tilsynelatende brede og voluminøse sekundærveggen faktisk som et edderkoppnett, som etter tørking krymper og blir til spredte stykker. Hvis primærveggene er inkludert i en kompleks midtplate, er det svært sannsynlig at det meste av ligninet er lokalisert her.
Kalsiumkanaler finnes også i plantecellemembraner. Reguleringen av inngangen til 45Ca2+ mikrosomer isolert fra maiskoleoptiler og gresskarhypokotyler med lys, PAA, og avhengigheten av denne reaksjonen av kalmodulin ble vist. For funksjonen til spenningsstyrte Ca2+-kanaler (karofytiske alger Lieu11op,m), er tilstedeværelsen av Mg2+ nødvendig. Tilstanden til disse spenningsstyrte kanalene styres av et system av enzymer som overvåker nivået av cAMP i cellen. Det ble også innhentet data som indikerer det direkte handling eksogen cAMP på opptak av 45Ca2+ i cellene i syklonen (mutant uten cellevegg). Dataene vist i fig. 4.1, indikerer den regulatoriske effekten av cAMP på absorpsjonen av Ca2+ av celler. Dette indikerer muligheten for gjensidig regulering av to systemer med andre budbringere - cAMP og Ca2+. I forsøk med dyreceller er økningen i Ca2+-opptak under påvirkning av cAMP forklart av fosforylering av proteiner av spenningsavhengige Ca2+-kanaler og som et resultat en økning i deres tilstedeværelse i åpen tilstand.[...]
Mange studier har blitt viet til å studere effekten av ultralyd på cellulosefibre. Noen forskere har sammenlignet eller kombinert effekten av ultralyd med ulike mekaniske påvirkninger. Således studerte Yaime, Kronert og Neuhaus effekten av ultralyd på cellulosefibre sammenlignet med høyfrekvente mekaniske vibrasjoner og viste at ultralyd med en frekvens på 20-3000 kHz løsner fiberstrukturen, øker graden av dens svelling og dehydrering. Den mekaniske styrken til papir laget av slike celluloser økes, spesielt rivestyrken. Høyfrekvente fungerer på samme måte. mekaniske vibrasjoner. Iwasaki, Lindberg og Meyer mener at det generelle mønsteret av endringer i fiberstruktur under påvirkning av ultralyd i et vannholdig miljø ligner på endringer i fiberstruktur under mekanisk sliping. I dette tilfellet oppstår dyptgripende endringer i den morfologiske strukturen til fibrene, noe som fører til forskyvninger i den sekundære celleveggen, separasjon av store stykker fra primærveggen, deretter til hevelse av sekundærveggen og dens defibrillering. I arbeidet til Safonova og Klenkova, når de studerte mikrofotografier av fibre utsatt for ultralyd i vann, ble det vist at det er andre, dypere forstyrrelser i strukturen til fiberen, som blir penetrert av et helt nettverk av mange tverrgående kanaler. Det bemerkes at tidlige trefibre og fibre som ikke er tørket er mer utsatt for ultralyd.
Planteceller, som cellene til prokaryoter og sopp, er innelukket i en relativt stiv cellevegg. Materialet for konstruksjonen av denne celleveggen skilles ut av cellen selv. levende celle(protoplast). Når det gjelder deres kjemiske sammensetning, skiller plantecelleveggene seg fra celleveggene til prokaryoter og sopp (tabell 2.1), men disse strukturene er preget av noen generelle funksjoner, nemlig funksjonene støtte og beskyttelse; i tillegg begrenser begge cellemotiliteten. Celleveggen avsatt under plantecelledeling kalles den primære celleveggen. Senere, som et resultat av fortykning, kan det bli en sekundær cellevegg. I denne delen beskriver vi prosessen med dannelsen av den primære celleveggen. I fig. 7.21 gjengir et elektronmikrofotografi der du kan se en av tidlige stadier denne prosessen.
Celleveggstruktur
Den primære celleveggen består av cellulosemikrofibriller innebygd i en matrise som inneholder komplekse polysakkarider. Cellulose er også et polysakkarid (dets kjemisk struktur beskrevet i sekt. 5.2.3). Spesielt viktig for rollen som cellulose spiller i cellevegger, har den en fibrøs struktur og høy strekkfasthet som kan sammenlignes med stål. Individuelle cellulosemolekyler er lange polysakkaridkjeder. Mange slike molekyler tverrbundet seg til hverandre hydrogenbindinger, samlet i sterke bunter kalt mikrofibriller. Mikrofibriller nedsenket i matrisen danner rammeverket til celleveggen. Celleveggmatrisen består av polysakkarider, som for enkelhets skyld vanligvis er delt inn i pektiner Og hemicelluloser avhengig av deres løselighet i forskjellige løsemidler som brukes til ekstraksjon. Pektiner, eller pektinstoffer, isoleres vanligvis først under ekstraksjon fordi deres løselighet er høyere. Det er en blandet gruppe sure polysakkarider (bygget av monosakkaridene arabinose og galaktose, galakturonsyre, som tilhører klassen av sukkersyrer, og metanol). Lange molekyler av pektinstoffer kan være lineære eller forgrenede. Median plate, som holder veggene til nabocellene sammen, består av klebrige gelatinøse pectates av magnesium og kalsium. I celleveggene til enkelte modnende frukter omdannes uløselige pektinstoffer tilbake til løselige pektiner. Når sukker tilsettes, danner disse sistnevnte geler; derfor brukes de som geleringsmidler.
Hemicelluloser- dette er en blandet gruppe polysakkarider som er løselige i alkalier (disse inkluderer polymerer av xylose, galaktose, mannose, glukose og glukomanose). Hemicelluloser, som cellulose, har kjedeformede molekyler, men kjedene deres er kortere, mindre ordnet og mer forgrenet.
Celleveggene er hydrert: 60-70 % av massen deres er vanligvis vann. Vann beveger seg fritt gjennom det ledige rommet i celleveggen. Tilstedeværelsen av vann påvirker kjemiske og fysiske egenskaper cellevegg polysakkarider.
Materialer med økt mekanisk styrke, lik celleveggmaterialet, dvs. bestående av mer enn én komponent, kalles komposittmaterialer eller kompositter; deres styrke er vanligvis høyere enn for hver av komponentene separat. Systemer av fibre og en matrise (i prosjektering kalles grunnlaget for et komposittmateriale ikke en matrise, men en matrise. - Oversettelse) er mye brukt i teknologi, så det brukes mye krefter på å studere egenskapene deres både innen teknologi og i biologi. Kompresjonsmatrisen overfører spenning til strekkfibrene. Det gir også slitebestandighet og, tilsynelatende, motstand mot uønskede kjemiske påvirkninger som er mulig under visse forhold. Armert betong har lenge vært brukt i konstruksjon, det vil si en kombinasjon av betong med stålarmering. Senere dukket det opp et lettere komposittmateriale, der rollen til matrisen spilles av plast, og rollen som forsterkning spilles av glass eller karbonfiber. Tre er et komposittmateriale; den skylder sin styrke til celleveggene. Et eksempel på stive komposittmaterialer biologisk opprinnelse Ben, brusk og skjellaget som dekker eksoskjelettet til leddyr kan også tjene. Det er også fleksible komposittmaterialer, slik som bindevev.
Noen celler, som bladmesofyllceller, har bare en primær cellevegg gjennom hele livet. Men i de fleste celler indre overflate Etter dannelsen av den primære celleveggen (utenfor plasmamembranen), avsettes ytterligere lag med cellulose, dvs. en sekundær cellevegg vises. Dette skjer vanligvis etter at cellen har nådd sin maksimale størrelse, og bare noen få celler, for eksempel collenchyma-celler, fortsetter å vokse i denne fasen. Sekundær fortykkelse av plantecelleveggene må ikke forveksles med sekundær fortykning (sekundær vekst) av selve planten, det vil si en økning i tykkelsen på stammen som følge av tilsetning av nye celler.
I ethvert lag med sekundær fortykkelse er cellulosefibre plassert i samme vinkel, men i forskjellige lag er denne vinkelen forskjellig, noe som sikrer enda større styrke på strukturen. Dette arrangementet av cellulosefibre er vist i fig. 7,27.
Noen celler, for eksempel trakeale xylem-elementer og sklerenkymceller, gjennomgår intense lignifisering(lignifisering); i dette tilfellet er alle lag av cellulose (primær og tre sekundære) impregnert med lignin - et komplekst polymert stoff som ikke er relatert til polysakkarider. I protoksylemceller har ligninavsetninger en ring-, spiral- eller nettverksform, som man kan se på fig. 8.11. I andre tilfeller er lignifiseringen kontinuerlig, bortsett fra de såkalte porefeltene, det vil si de områdene i den primære celleveggen som kontakt mellom naboceller skjer gjennom ved bruk av en gruppe plasmodesmata (avsnitt 8.1.3 og fig. 8.7). Lignin binder cellulosefibre sammen og holder dem på plass. Den fungerer som en veldig hard og stiv matrise, som øker strekkstyrken og spesielt trykkfastheten til celleveggene (hindrer henging). Det gir også cellene ekstra beskyttelse mot uønskede fysiske og kjemiske påvirkninger. Sammen med cellulose som er igjen i celleveggene, gir lignin tre de spesielle egenskapene som gjør det til et uunnværlig byggemateriale.
Funksjoner av celleveggen
Hovedfunksjonene til plantecellevegger er listet opp nedenfor.
1. Cellevegger gir individuelle celler og planten som helhet mekanisk styrke og støtte. I noen vev forsterkes styrken ved omfattende lignifisering av celleveggene (små mengder lignin er tilstede i alle cellevegger).
2. Den relative stivheten til celleveggene og motstanden mot strekk bestemmer cellenes turgiditet når vann kommer inn i dem ved osmose. Dette forbedrer støttefunksjonen i alle planter og fungerer som den eneste støttekilden for urteaktige planter og for organer som blader, dvs. der sekundærvekst er fraværende. Cellevegger beskytter også cellene mot brudd i et hypotont miljø.
3. Orienteringen av cellulosemikrofibriller begrenser og regulerer til en viss grad både vekst og form av cellene, siden cellenes evne til å strekke seg avhenger av plasseringen av disse mikrofibrillerne. Hvis for eksempel mikrofibriller er plassert på tvers av cellen og omgir den som med bøyler, vil cellen som vann kommer inn i ved osmose strekke seg i lengderetningen.
4. Et system av cellevegger som er forbundet med hverandre ( apoplast) fungerer som hovedvei langs hvilken vann og mineraler. Cellevegger festes til hverandre ved hjelp av medianplater. Veggene har små porer som cytoplasmatiske tråder passerer, kalt plasmodesmata. Plasmodesmata binder det levende innholdet i individuelle celler - de forener alle protoplaster til enhetlig system, i den såkalte simplast.
5. De ytre celleveggene til epidermale celler er dekket med en spesiell neglebåndsfilm, bestående av et voksaktig stoff kalt cutin, som reduserer vanntap og reduserer risikoen for at patogener kommer inn i planten. I korkvev, etter fullføring av sekundær vekst, er celleveggene impregnert med suberin, som utfører en lignende funksjon.
6. Celleveggene til xylemkar, trakeider og silrør (med silplater) er tilrettelagt for langtransport av stoffer gjennom hele planten. Denne problemstillingen er omtalt i kap. 8 og 14.
7. Veggene til endodermale rotceller er impregnert med suberin og fungerer derfor som en barriere for bevegelse av vann (avsnitt 14.1.5).
8. I noen celler lagrer deres modifiserte vegger reserver næringsstoffer; På denne måten lagres for eksempel hemicelluloser i enkelte frø.
9. I overføringsceller økes overflatearealet av celleveggene og overflatearealet til plasmamembranen økes tilsvarende, noe som øker effektiviteten av overføring av stoffer ved å aktiv transport(Pkt. 14.8.6).
Celleveggdannelse begynner under celledeling. I delingsplanet dannes en celleplate, et enkelt lag felles for de to dattercellene. Den består av pektinstoffer som har en halvflytende konsistens; det er ingen cellulose. I en voksencelle er celleplaten bevart, men gjennomgår endringer, derfor kalles den median, eller intercellulær plate (intercellulær substans) (ris. 2.16). Medianplaten er vanligvis veldig tynn og nesten umulig å skille.
Umiddelbart etter dannelsen av celleplaten begynner protoplastene til dattercellene å legge ned sin egen cellevegg. Det avsettes fra innsiden både på overflaten av celleplaten og på overflaten av andre cellevegger som tidligere tilhørte modercellen. Etter deling går cellen inn i forlengelsesvekstfasen, som er forårsaket av intens osmotisk absorpsjon av vann av cellen assosiert med dannelsen og veksten av sentralvakuolen. Turgortrykk begynner å strekke veggen, men det rives ikke på grunn av det faktum at nye deler av mikrofibriller og matrisestoffer hele tiden avsettes i den. Avsetningen av nye deler av materiale skjer jevnt over hele overflaten av protoplasten, slik at tykkelsen på celleveggen ikke reduseres.
Veggene til celler som deler og vokser kalles hoved. De inneholder mye (60-90%) vann. Tørrstoffet domineres av matrisepolysakkarider (60-70%), celluloseinnholdet overstiger ikke 30%, og det er ingen lignin. Tykkelsen på primærveggen er veldig liten (0,1-0,5 mikron).
For mange celler opphører celleveggavsetning samtidig med opphør av cellevekst. Slike celler er omgitt av en tynn primærvegg til slutten av livet ( ris. 2.16).
Ris. 2.16. Parenkymcelle med en primær vegg.
I andre celler fortsetter veggavsetning selv etter at cellen når sin endelige størrelse. I dette tilfellet øker veggtykkelsen, og volumet som er okkupert av cellehulen reduseres. Denne prosessen kalles sekundær fortykning vegger, og selve veggen kalles sekundær(ris. 2.17).
Sekundærveggen kan betraktes som ekstra, og utfører hovedsakelig en mekanisk støttefunksjon. Det er sekundærveggen som er ansvarlig for egenskapene til tre, tekstilfiber og papir. Sekundærveggen inneholder betydelig mindre vann enn primærveggen; den domineres av cellulosemikrofibriller (40-50 % av tørrstoffvekten), som er plassert parallelt med hverandre. Av matrikspolysakkaridene er hemicelluloser (20-30%) typiske, og det er svært få pektinstoffer. Sekundære cellevegger gjennomgår vanligvis lignifisering. I ikke-lignifiserte sekundærvegger (linbastfibre, bomullshår) kan celluloseinnholdet nå 95 %. Flott innhold og strengt bestilt orientering av mikrofibriller bestemme høy mekaniske egenskaper sekundære vegger. Ofte dør celler som har en sekundær lignifisert cellevegg etter at den sekundære fortykningen er fullført.
Median lamina limer naboceller sammen. Hvis det løses opp, mister celleveggene kontakten med hverandre og skilles. Denne prosessen kalles maserasjon. Naturlig maserasjon er ganske vanlig, der pektinstoffene i midtplaten omdannes til en løselig tilstand ved hjelp av enzymet pektinase og deretter vaskes bort med vann (overmodne frukter av pære, melon, fersken, banan). Ofte observeres delvis maserasjon, hvor midtplaten ikke løses opp over hele overflaten, men kun i hjørnene av cellene. På grunn av turgortrykk blir naboceller på disse stedene avrundet, noe som resulterer i dannelsen intercellulære rom(ris. 2.16). De intercellulære rommene danner et enkelt forgrenet nettverk, som er fylt med vanndamp og gasser. Dermed forbedrer intercellulære rom gassutveksling av celler.
Et karakteristisk trekk ved sekundærveggen er dens ujevne avsetning på toppen av primærveggen, som et resultat av at utykkede områder forblir i sekundærveggen - porene. Hvis sekundærveggen ikke når en stor tykkelse, ser porene ut som små fordypninger. I celler med sterk sekundærvegg ser porene i tverrsnitt ut som radielle kanaler som strekker seg fra cellehulen til primærveggen. Basert på formen på porekanalen er det to typer porer: enkel og om kantet(Fig. 2.17).
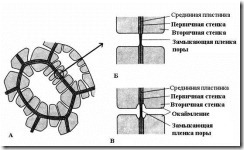
Ris. 2.17. Poretyper: A – celler med sekundærvegger og mange enkle porer; B - et par enkle porer; B – par avkantede porer.
U enkle porer Diameteren på porekanalen er den samme langs hele lengden og har form som en smal sylinder. Enkle porer er karakteristiske for parenkymceller, bast- og trefibre.
Porene i to tilstøtende celler har en tendens til å vises overfor hverandre. Disse vanlige porene ser ut som en kanal, atskilt av en tynn skillevegg mellom midtplaten og primærveggen. Denne kombinasjonen av to porer av tilstøtende vegger av naboceller kalles par porer og fungerer som en helhet. Seksjonen av veggen som skiller dem kalles avslutningsfilm porene, eller poremembran. I levende celler er porens lukkefilm gjennomsyret av mange plasmodesmata(ris. 2.18).
Plasmodesmata kun iboende planteceller. De er tråder av cytoplasma som krysser veggen til tilstøtende celler. Antallet plasmodesmata i en celle er veldig stort - fra flere hundre til titusenvis samles vanligvis plasmodesmata i grupper. Diameteren til den plasmodesmale kanalen er 30-60 nm. Veggene er foret med plasmalemma, kontinuerlig med plasmalemmaet til tilstøtende celler. I midten av plasmodesmata er det en membransylinder - sentral stang plasmodesmata, kontinuerlig med elementmembraner endoplasmatisk retikulum begge cellene. Mellom den sentrale stangen og plasmamembranen i kanalen er det hyaloplasma, kontinuerlig med hyaloplasmaen til tilstøtende celler.
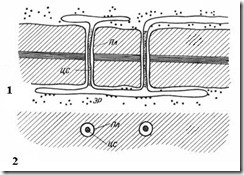
Ris. 2.18. Plasmodesmata under et elektronmikroskop (diagram): 1 – på et lengdesnitt; 2 - på et tverrsnitt; Pl– plasmalemma; CA- sentral stang av plasmodesmata; ER– element i det endoplasmatiske retikulum.
Dermed er ikke celleprotoplaster fullstendig isolert fra hverandre, men kommuniserer gjennom plasmodesmata-kanaler. De bærer intercellulær transport av ioner og små molekyler, og overfører også hormonelle stimuli. Gjennom plasmodesmata, protoplaster av celler i planteorganisme danne en enkelt helhet kalt simplast, og transport av stoffer gjennom plasmodesmata kalles symplastisk I motsetning til apoplastisk transport langs cellevegger og intercellulære rom.
U avgrensede porer(ris. 2.17) kanalen smalner kraftig under avsetningen av celleveggen, så den indre åpningen av poren, som åpner seg inn i cellehulen, er mye smalere enn den ytre, og støter mot primærveggen. Avgrensede porer er karakteristiske for tidlig døende celler av vannledende elementer av tre. I dem utvider porekanalen seg traktformet mot lukkefilmen, og sekundærveggen henger i form av en rulle over den utvidede delen av kanalen og danner et porekammer. Navnet avgrenset pore kommer fra det faktum at sett fra overflaten, ser den indre åpningen ut som en liten sirkel eller smal åpning, mens det ytre hullet så å si grenser til det indre i form av en sirkel med større diameter eller en bredere spalte.
Porene letter transporten av vann og oppløste stoffer fra celle til celle uten å redusere styrken til celleveggen.