Ribosomer– ribonukleoproteingranulat 25 nm i størrelse. De består av to underenheter: liten (10 nm) og stor (15 nm), mellom hvilke en tråd av messenger-RNA er lokalisert under proteinbiosyntese (translasjon). I dette tilfellet binder den lille underenheten seg til RNA, og den store katalyserer dannelsen av polypeptidkjeder. Ribosomale underenheter dannes i nukleolene og går deretter ut av kjernen inn i cytoplasmaet gjennom kjerneporer. Sammenstillingen av ribosomer fra deres underenheter skjer før starten av proteinsyntesen, og etter fullføring av syntesen av polypeptidkjeden desintegrerer de igjen.
En syntetisk aktiv celle inneholder flere millioner ribosomer, som utgjør omtrent 5 % av dens tørre masse. Skille gratis ribosomer (ikke assosiert med membraner og lokalisert i hyaloplasma i suspensjon) og ufri ribosomer (assosiert med membraner i det cytoplasmatiske retikulum). Ribosomer kan lokaliseres enkeltvis (i dette tilfellet er de funksjonelt inaktive), men oftere er de koblet i kjeder som er trukket, som perler, på trådlignende messenger RNA-molekyler ( polyribosomer, polysomer). Frie ribosomer syntetiserer proteiner for cellens egne behov, mens ikke-frie ribosomer syntetiserer proteiner for eksport.
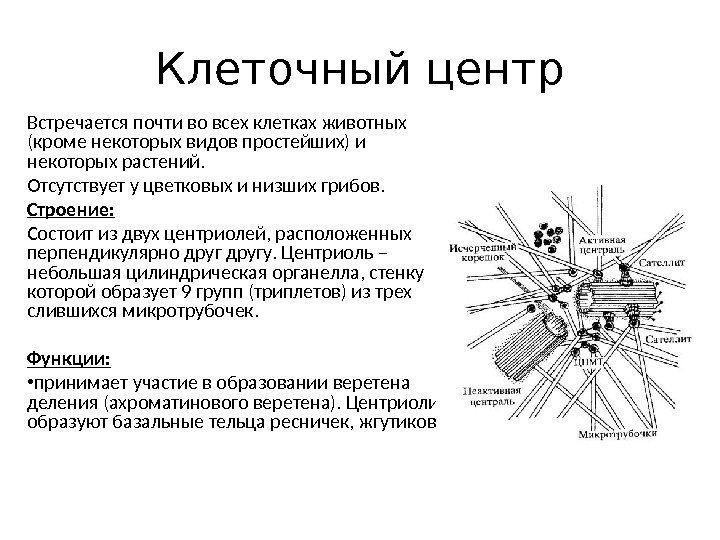
Cellesenter (cytosenter)– består av to plassert vinkelrett på hverandre sentrioler.
Figur 2-8. Cellesenter (A) og sentriolstruktur (B).
1. Centriol.
2. Satellitter.
3. Triplett av mikrotubuli.
4. Mikrotubuli.
(Ifølge V.L. Bykov).
Centriole er en hul sylinder 200 nm tykk og 300-500 nm lang. Sentriolveggen er dannet av 9 tripletter av mikrotubuli, 24 nm tykke, bygget av kuleprotein tubulin. Nabotripletter av mikrotubuli er forbundet i form av broer av proteinmolekyler dynein. Hver triplett av mikrotubuli er også assosiert med sfæriske strukturer - satellitter. Mikrotubuli divergerer fra satellittene til sidene og danner seg centosfære(Figur 2-8).
Cellesenteret tar del i dannelsen av delingsspindelen; Under mitose beveger sentrioler seg mot polene til modercellen. I tillegg deltar sentrioler i dannelsen av flimmerhår og flageller.
Organeller i cytoskjelettet
Cytoskjelettet er et komplekst dynamisk tredimensjonalt nettverk av mikrotubuli, mikrofibriller og mikrofilamenter, som gir: 1) vedlikehold og endring i celleform, 2) distribusjon og bevegelse av cellekomponenter, 3) transport av stoffer inn og ut av cellen, 4) cellemotilitet, 5) deltar i intercellulære forbindelser.
Mikrotubuli har en tykkelse på 24 nm og en lengde på flere mikron. Mikrotubulus veggtykkelse er 5 nm, og lumendiameteren er tilsvarende 14 nm. De består av 13 tubulin-protofibriller som går i en spiral. Mikrotubuli er en del av spindelen og sørger for divergens av kromosomer under mitose, opprettholder cellens form og sikrer mobilitet, og deltar i transporten av makromolekyler i cellen. Assosiert med mikrotubuli er proteinet kinesin, som er et enzym som bryter ned ATP og omdanner energien fra nedbrytningen til mekanisk energi. I den ene enden er kinesinmolekylet koblet til en spesifikk organell, og i den andre, ved hjelp av ATP-energi, glir det langs mikrotubuli og beveger organellen i cytoplasmaet.
Mikrotubuli er et labilt system der dissosiasjonen (ødeleggelsen) av noen mikrotubuli og sammenstillingen (dannelsen) av andre skjer kontinuerlig. Stedet for dannelse av mikrotubuli ( organiseringssentre for mikrotubuli) er små sfæriske legemer satellitter. De er lokalisert i basallegemene til flimmerhårene og cellesenteret.
Mellomfilamenter (mikrofibriller) – proteinfilamenter 8-11 nm tykke. De danner rammen av cellen, opprettholder dens form og elastisitet, og sikrer også den ryddige ordningen av organeller i cellen.
Mikrofilamenter – proteinfilamenter 5-7 nm tykke. De er tilstede i alle celler og er lokalisert i dets kortikale lag (under cytolemma). Sammensetningen av proteinene som danner dem i forskjellige celler forskjellige (aktin, myosin, tropomyosin). De danner skjelettet, rammen til cellen, dens intracellulære kontraktile apparatur, sikrer endringer i form og bevegelse av celler, cytoplasmatisk flyt, endocytose og eksocytose.
Funksjonelle systemer (enheter) i cellen- komplekser av organeller som, under kontroll av kjernen, sikrer utførelsen av viktige cellefunksjoner. Det er: 1) syntetiske apparater (inkluderer endoplasmatisk retikulum, ribosomer, Golgi-kompleks; 2 ) energiapparat (mitokondrier); 3) intracellulært fordøyelsesapparat vania(endosomer, lysosomer, peroksisomer); 4) cytoskjelett (mikrotubuli, mikrofibriller, mikrofilamenter, cellesenter).

 Organeller er permanente intracellulære strukturer som har en spesifikk struktur og utfører tilsvarende funksjoner. Organeller er delt inn i to grupper: membran og ikke-membran. Membranorganeller presenteres i to varianter: dobbelmembran og enkelmembran.
Organeller er permanente intracellulære strukturer som har en spesifikk struktur og utfører tilsvarende funksjoner. Organeller er delt inn i to grupper: membran og ikke-membran. Membranorganeller presenteres i to varianter: dobbelmembran og enkelmembran.
 De doble membrankomponentene er plastider, mitokondrier og cellekjernen. Enkeltmembranorganeller inkluderer organellene i det vakuolære systemet - det endoplasmatiske retikulum, Golgi-komplekset, lysosomer, vakuoler av plante- og soppceller osv. Ikke-membranorganeller inkluderer ribosomer og cellesenteret
De doble membrankomponentene er plastider, mitokondrier og cellekjernen. Enkeltmembranorganeller inkluderer organellene i det vakuolære systemet - det endoplasmatiske retikulum, Golgi-komplekset, lysosomer, vakuoler av plante- og soppceller osv. Ikke-membranorganeller inkluderer ribosomer og cellesenteret
 Enkeltmembranorganeller Endoplasmatisk retikulum (ER) eller retikulum - et komplekst system av kanaler og hulrom ulike former(tubuli, sisterne), penetrerer hele cytoplasmaet. a) Grovt eller granulært endoplasmatisk retikulum: membranene er dekket med små granuler - ribosomer. Funksjoner: syntese av polypeptider, deres delvise modifikasjon og transport b) Glatt eller agranulært, endoplasmatisk retikulum: membraner er blottet for ribosomer, men enzymer av lipid- og karbohydratmetabolisme akkumuleres her. Funksjoner: syntese av lipider, steroider, karbohydrater, deres transport.
Enkeltmembranorganeller Endoplasmatisk retikulum (ER) eller retikulum - et komplekst system av kanaler og hulrom ulike former(tubuli, sisterne), penetrerer hele cytoplasmaet. a) Grovt eller granulært endoplasmatisk retikulum: membranene er dekket med små granuler - ribosomer. Funksjoner: syntese av polypeptider, deres delvise modifikasjon og transport b) Glatt eller agranulært, endoplasmatisk retikulum: membraner er blottet for ribosomer, men enzymer av lipid- og karbohydratmetabolisme akkumuleres her. Funksjoner: syntese av lipider, steroider, karbohydrater, deres transport.

 Funksjoner: Kobler sammen alle cellemembranstrukturer i enhetlig system. Det er overflaten som alle intracellulære prosesser skjer (syntese av membranproteiner, lipider og karbohydrater). Romlig skiller cellen. Transport av stoffer skjer gjennom et system av kanaler.
Funksjoner: Kobler sammen alle cellemembranstrukturer i enhetlig system. Det er overflaten som alle intracellulære prosesser skjer (syntese av membranproteiner, lipider og karbohydrater). Romlig skiller cellen. Transport av stoffer skjer gjennom et system av kanaler.
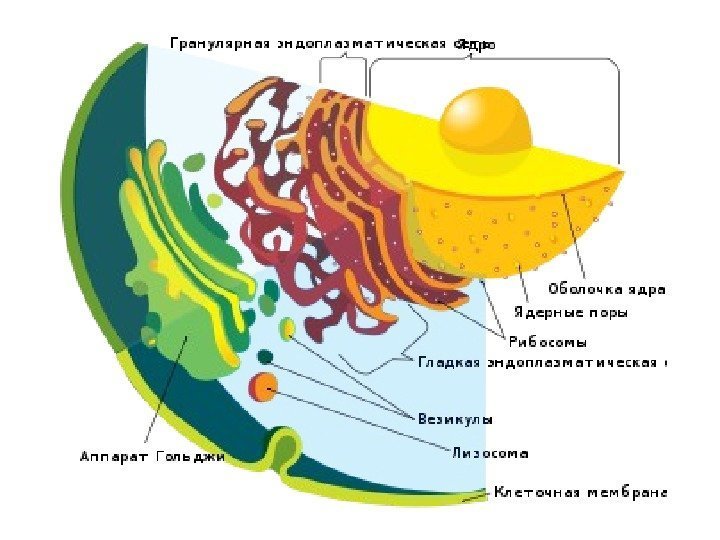
 Golgi-komplekset er tilstede i nesten alle celler (med unntak av erytrocytter og sædceller). Struktur: Et system av stablede, flate membransekker - sisterner, rør og tilhørende vesikler. Funksjoner: Transport av stoffer, hovedsakelig proteiner og lipider som kommer fra endoplasmatisk retikulum, deres foreløpige kjemiske omorganisering, akkumulering, pakking til vesikler, dannelse av lysosomer.
Golgi-komplekset er tilstede i nesten alle celler (med unntak av erytrocytter og sædceller). Struktur: Et system av stablede, flate membransekker - sisterner, rør og tilhørende vesikler. Funksjoner: Transport av stoffer, hovedsakelig proteiner og lipider som kommer fra endoplasmatisk retikulum, deres foreløpige kjemiske omorganisering, akkumulering, pakking til vesikler, dannelse av lysosomer.
 Golgi-apparatet ble oppkalt etter den italienske forskeren Camillo Golgi, som først oppdaget det i 1897
Golgi-apparatet ble oppkalt etter den italienske forskeren Camillo Golgi, som først oppdaget det i 1897

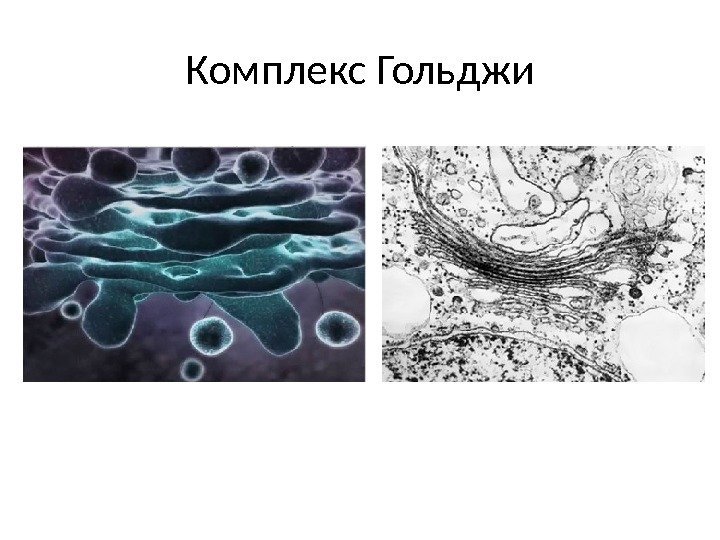
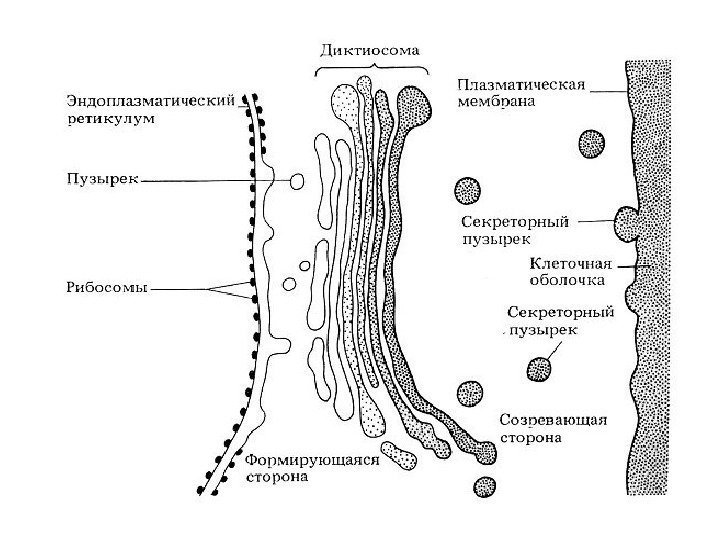
 Funksjoner av Golgi-komplekset 1) sortering, akkumulering og utskillelse av sekretoriske produkter 2) fullføring av post-translasjonell modifikasjon av proteiner 3) akkumulering av lipidmolekyler og dannelse av lipoproteiner 4) dannelse av lysosomer 5) syntese av polysakkarider for dannelse av glykoproteiner , voks, slim, matriksstoffer cellevegger planter (hemicellulose, pektiner) 6) dannelse av celleplaten etter nukleær deling i planteceller 7) deltakelse i dannelsen av akrosomet; dannelse av kontraktile vakuoler av protozoer.
Funksjoner av Golgi-komplekset 1) sortering, akkumulering og utskillelse av sekretoriske produkter 2) fullføring av post-translasjonell modifikasjon av proteiner 3) akkumulering av lipidmolekyler og dannelse av lipoproteiner 4) dannelse av lysosomer 5) syntese av polysakkarider for dannelse av glykoproteiner , voks, slim, matriksstoffer cellevegger planter (hemicellulose, pektiner) 6) dannelse av celleplaten etter nukleær deling i planteceller 7) deltakelse i dannelsen av akrosomet; dannelse av kontraktile vakuoler av protozoer.
 Funksjoner av Golgi-apparatet: I sisternene til Golgi-apparatet modnes proteiner beregnet for sekresjon, transmembranproteiner i plasmamembranen, lysosomproteiner osv. De modnede proteinene beveger seg sekvensielt gjennom organellesisternene, der deres endelige folding skjer. samt modifikasjoner - glykosylering og fosforylering.
Funksjoner av Golgi-apparatet: I sisternene til Golgi-apparatet modnes proteiner beregnet for sekresjon, transmembranproteiner i plasmamembranen, lysosomproteiner osv. De modnede proteinene beveger seg sekvensielt gjennom organellesisternene, der deres endelige folding skjer. samt modifikasjoner - glykosylering og fosforylering.
 Separasjon av proteiner i 3 strømmer: 1. lysosomale - glykosylerte proteiner (med mannose) kommer inn i cis-seksjonen av Golgi-komplekset, noen av dem er fosforylerte, og det dannes en markør for lysosomale enzymer - mannose-6-fosfat. I fremtiden vil disse fosforylerte proteinene ikke gjennomgå modifikasjon, men vil gå inn i lysosomene. 2. konstitutiv eksocytose (konstitutiv sekresjon). Denne strømmen inkluderer proteiner og lipider, som blir komponenter av celleoverflateapparatet, inkludert glykokalyxen, eller de kan være en del av den ekstracellulære matrisen. 3. Induserbar sekresjon - proteiner som fungerer utenfor cellen, celleoverflateapparatet, under Internt miljø kropp. Karakteristisk for sekretoriske celler.
Separasjon av proteiner i 3 strømmer: 1. lysosomale - glykosylerte proteiner (med mannose) kommer inn i cis-seksjonen av Golgi-komplekset, noen av dem er fosforylerte, og det dannes en markør for lysosomale enzymer - mannose-6-fosfat. I fremtiden vil disse fosforylerte proteinene ikke gjennomgå modifikasjon, men vil gå inn i lysosomene. 2. konstitutiv eksocytose (konstitutiv sekresjon). Denne strømmen inkluderer proteiner og lipider, som blir komponenter av celleoverflateapparatet, inkludert glykokalyxen, eller de kan være en del av den ekstracellulære matrisen. 3. Induserbar sekresjon - proteiner som fungerer utenfor cellen, celleoverflateapparatet, under Internt miljø kropp. Karakteristisk for sekretoriske celler.
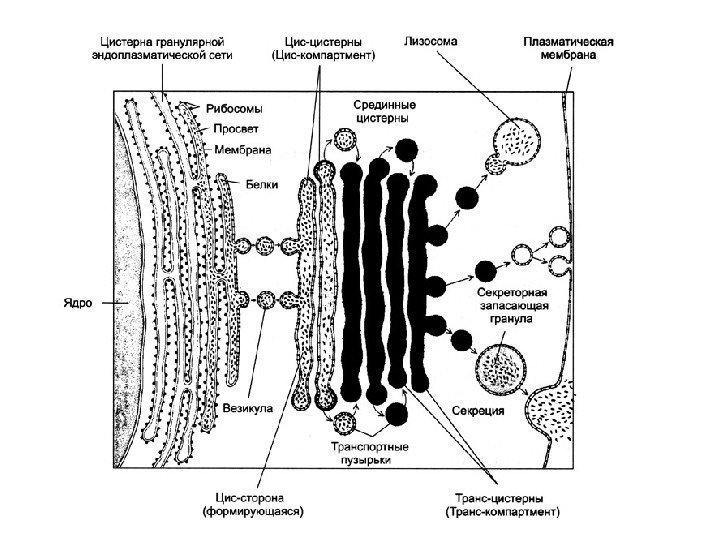
![]() For å avslutte vurderingen av strukturen og driften av en så kompleks membranorganell som Golgi-apparatet, er det nødvendig å understreke at til tross for den tilsynelatende morfologiske homogeniteten til komponentene, vakuolen og sisternen, er det faktisk ikke bare en samling av vesikler, men et slankt, dynamisk, komplekst organisert, polarisert system.
For å avslutte vurderingen av strukturen og driften av en så kompleks membranorganell som Golgi-apparatet, er det nødvendig å understreke at til tross for den tilsynelatende morfologiske homogeniteten til komponentene, vakuolen og sisternen, er det faktisk ikke bare en samling av vesikler, men et slankt, dynamisk, komplekst organisert, polarisert system.

 Lysosomer Finnes i alle celler, spredt over hele cytoplasmaet. Struktur: Enkeltmembranvesikler av forskjellige former og størrelser; inneholder ulike proteolytiske enzymer (ca. 40). Funksjoner: Delta i intracellulær fordøyelse, dvs. nedbryting av store molekyler. De kan også ødelegge strukturene til selve cellen, forårsaker dens død - autolyse.
Lysosomer Finnes i alle celler, spredt over hele cytoplasmaet. Struktur: Enkeltmembranvesikler av forskjellige former og størrelser; inneholder ulike proteolytiske enzymer (ca. 40). Funksjoner: Delta i intracellulær fordøyelse, dvs. nedbryting av store molekyler. De kan også ødelegge strukturene til selve cellen, forårsaker dens død - autolyse.
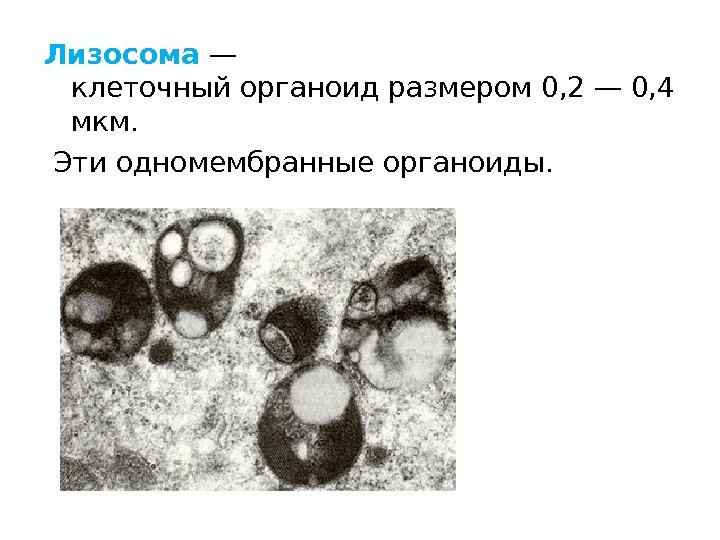
 Lysosomer ble først beskrevet i 1955 av Christian de Duve i dyreceller, og ble senere oppdaget i planteceller.
Lysosomer ble først beskrevet i 1955 av Christian de Duve i dyreceller, og ble senere oppdaget i planteceller.
 Lysosomer Hos planter ligner lysosomer delvis på lysosomer i deres dannelsesmetode og i funksjon. Tilstedeværelsen av lysosomer er karakteristisk for celler av alle eukaryoter. Det er ingen uprokaryote lysosomer, siden de mangler fagocytose og ikke har intracellulær fordøyelse.
Lysosomer Hos planter ligner lysosomer delvis på lysosomer i deres dannelsesmetode og i funksjon. Tilstedeværelsen av lysosomer er karakteristisk for celler av alle eukaryoter. Det er ingen uprokaryote lysosomer, siden de mangler fagocytose og ikke har intracellulær fordøyelse.
 Tegn på lysosomer Et av tegnene på lysosomer er tilstedeværelsen i dem av en rekke enzymer (syrehydrolaser) som er i stand til å bryte ned proteiner, karbohydrater, lipider og nukleinsyrer.
Tegn på lysosomer Et av tegnene på lysosomer er tilstedeværelsen i dem av en rekke enzymer (syrehydrolaser) som er i stand til å bryte ned proteiner, karbohydrater, lipider og nukleinsyrer.
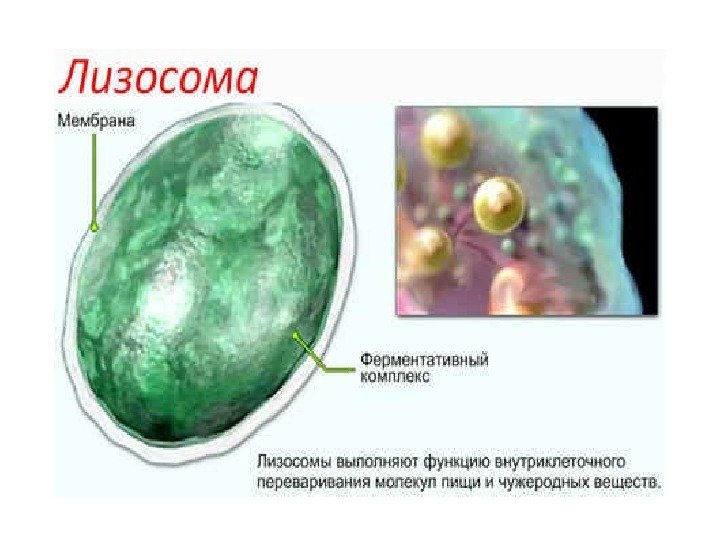
 Enzymene til lysosomer inkluderer katepsiner (vevsproteaser), sur ribonuklease, fosfolipaser og andre lysosomer inneholder enzymer som er i stand til å splitte seg organiske molekyler sulfat (sulfataser) eller fosfat (syre fosfatase) grupper.
Enzymene til lysosomer inkluderer katepsiner (vevsproteaser), sur ribonuklease, fosfolipaser og andre lysosomer inneholder enzymer som er i stand til å splitte seg organiske molekyler sulfat (sulfataser) eller fosfat (syre fosfatase) grupper.
 Dannelse av lysosomer og deres typer Lysosomer dannes fra vesikler (vesikler), separert fra Golgi-apparatet, og vesikler (endosomer), som stoffer kommer inn i under endocytose. Membranene i det endoplasmatiske retikulumet deltar i dannelsen av autolysosomer (autofagosomer). Alle lysosomproteiner syntetiseres på fastsittende ribosomer. utenfor membraner i det endoplasmatiske retikulum og passerer deretter gjennom hulrommet og gjennom Golgi-apparatet.
Dannelse av lysosomer og deres typer Lysosomer dannes fra vesikler (vesikler), separert fra Golgi-apparatet, og vesikler (endosomer), som stoffer kommer inn i under endocytose. Membranene i det endoplasmatiske retikulumet deltar i dannelsen av autolysosomer (autofagosomer). Alle lysosomproteiner syntetiseres på fastsittende ribosomer. utenfor membraner i det endoplasmatiske retikulum og passerer deretter gjennom hulrommet og gjennom Golgi-apparatet.
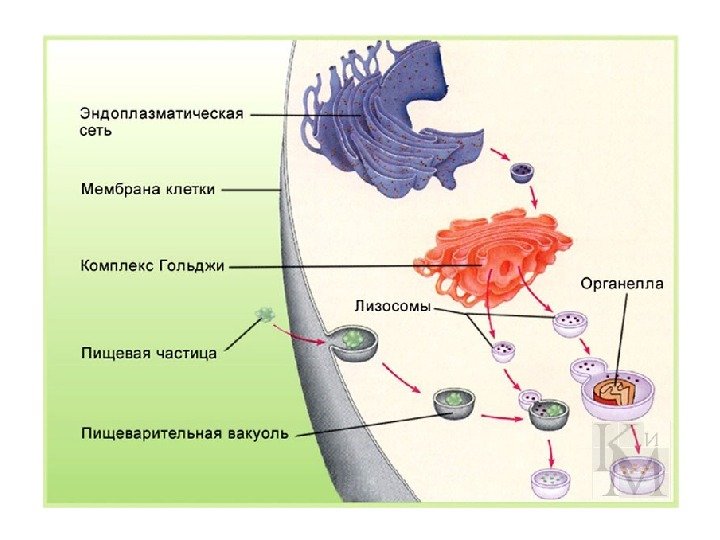
 Lysosomer er heterogene organeller som har forskjellige former, størrelser, ultrastrukturelle og cytokjemiske egenskaper. "Typiske" lysosomer i dyreceller er vanligvis sfæriske eller oval form. Antall lysosomer varierer fra én (en stor vakuole i mange plante- og soppceller) til flere hundre eller tusen (i dyreceller).
Lysosomer er heterogene organeller som har forskjellige former, størrelser, ultrastrukturelle og cytokjemiske egenskaper. "Typiske" lysosomer i dyreceller er vanligvis sfæriske eller oval form. Antall lysosomer varierer fra én (en stor vakuole i mange plante- og soppceller) til flere hundre eller tusen (i dyreceller).
![]() Det er primære og sekundære lysosomer. De førstnevnte er dannet i området av Golgi-apparatet, de inneholder enzymer i inaktiv tilstand, mens sistnevnte inneholder aktive enzymer.
Det er primære og sekundære lysosomer. De førstnevnte er dannet i området av Golgi-apparatet, de inneholder enzymer i inaktiv tilstand, mens sistnevnte inneholder aktive enzymer.
 Blant lysosomer kan man også skille mellom heterolysosomer (fordøyer materiale som kommer inn i cellen fra utsiden gjennom fago- eller pinocytose) og autolysosomer (ødelegger cellens egne proteiner eller organeller).
Blant lysosomer kan man også skille mellom heterolysosomer (fordøyer materiale som kommer inn i cellen fra utsiden gjennom fago- eller pinocytose) og autolysosomer (ødelegger cellens egne proteiner eller organeller).
 Den mest brukte klassifiseringen av lysosomer og tilhørende kompartmenter er: Tidlig endosom - den mottar endocytiske (pinocytotiske) vesikler. Sen endosom - vesikler med materiale absorbert under pinocytose og vesikler fra Golgi-apparatet med hydrolaser kommer inn i det fra det tidlige endosomet.
Den mest brukte klassifiseringen av lysosomer og tilhørende kompartmenter er: Tidlig endosom - den mottar endocytiske (pinocytotiske) vesikler. Sen endosom - vesikler med materiale absorbert under pinocytose og vesikler fra Golgi-apparatet med hydrolaser kommer inn i det fra det tidlige endosomet.
 klassifisering Lysosom - vesikler med en blanding av hydrolaser og fordøyelig materiale kommer inn i det fra det sene endosomet.
klassifisering Lysosom - vesikler med en blanding av hydrolaser og fordøyelig materiale kommer inn i det fra det sene endosomet.
 Fagosomklassifisering - den inkluderer større partikler (bakterier, etc.) absorbert av fagocytose. Fagosomer smelter vanligvis sammen med et lysosom.
Fagosomklassifisering - den inkluderer større partikler (bakterier, etc.) absorbert av fagocytose. Fagosomer smelter vanligvis sammen med et lysosom.
 klassifisering Autophagosome er en region av cytoplasma omgitt av to membraner, vanligvis inkludert noen organeller og dannet under makroautofagi. Fusjonerer med lysosomet.
klassifisering Autophagosome er en region av cytoplasma omgitt av to membraner, vanligvis inkludert noen organeller og dannet under makroautofagi. Fusjonerer med lysosomet.
 klassifisering Multivesikulære legemer - vanligvis omgitt av en enkelt membran, inneholder inni mindre vesikler omgitt av en enkelt membran. De er dannet ved en prosess som minner om mikroautofagi, men inneholder materiale hentet utenfra. Dannelsesstadiet tilsvarer tidlige endosomer.
klassifisering Multivesikulære legemer - vanligvis omgitt av en enkelt membran, inneholder inni mindre vesikler omgitt av en enkelt membran. De er dannet ved en prosess som minner om mikroautofagi, men inneholder materiale hentet utenfra. Dannelsesstadiet tilsvarer tidlige endosomer.
 klassifisering Restlegemer (telolysosomer) er vesikler som inneholder ufordøyd materiale (lipofuscin). I normale celler smelter de sammen med den ytre membranen og forlater cellen ved eksocytose. De akkumuleres med aldring eller patologi.
klassifisering Restlegemer (telolysosomer) er vesikler som inneholder ufordøyd materiale (lipofuscin). I normale celler smelter de sammen med den ytre membranen og forlater cellen ved eksocytose. De akkumuleres med aldring eller patologi.
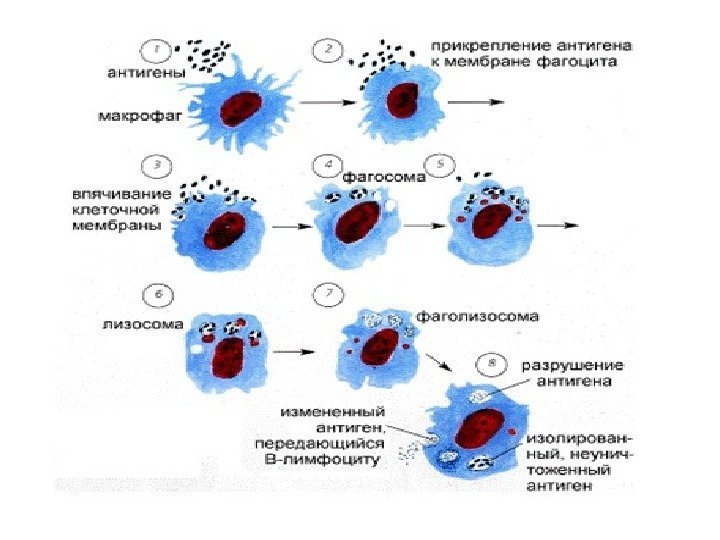
 Funksjoner av lysosomer: fordøyelse av stoffer eller partikler fanget av cellen under endocytose (bakterier, andre celler) - ødeleggelse av strukturer som er unødvendige for cellen, for eksempel under utskifting av gamle organeller med nye, eller fordøyelse av proteiner og andre stoffer som produseres inne i selve cellen
Funksjoner av lysosomer: fordøyelse av stoffer eller partikler fanget av cellen under endocytose (bakterier, andre celler) - ødeleggelse av strukturer som er unødvendige for cellen, for eksempel under utskifting av gamle organeller med nye, eller fordøyelse av proteiner og andre stoffer som produseres inne i selve cellen
 autolyse er selvfordøyelsen av en celle, som fører til dens død (noen ganger er denne prosessen ikke patologisk, men følger med utviklingen av kroppen eller differensieringen av noen spesialiserte celler). Eksempel: Når en rumpetroll forvandles til en frosk, fordøyer lysosomer i halens celler den: halen forsvinner, og stoffene som dannes under denne prosessen absorberes og brukes av andre celler i kroppen. Funksjoner av lysosomer
autolyse er selvfordøyelsen av en celle, som fører til dens død (noen ganger er denne prosessen ikke patologisk, men følger med utviklingen av kroppen eller differensieringen av noen spesialiserte celler). Eksempel: Når en rumpetroll forvandles til en frosk, fordøyer lysosomer i halens celler den: halen forsvinner, og stoffene som dannes under denne prosessen absorberes og brukes av andre celler i kroppen. Funksjoner av lysosomer
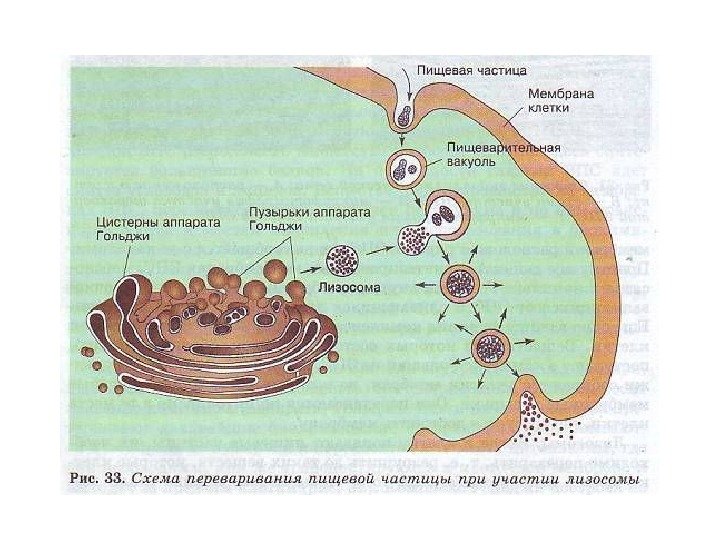
 Klinisk signifikans. Sykdommer assosiert med funksjonsfeil i lysosomer Noen ganger, på grunn av feil funksjon av lysosomer, utvikles lagringssykdommer, der enzymer ikke virker eller fungerer dårlig på grunn av mutasjoner. Et eksempel på lagringssykdommer er amaurotisk idioti på grunn av glykogenlagring. Bruddet av lysosomet og frigjøringen av hyaloplasma-splittende enzymer er ledsaget av en kraftig økning i deres aktivitet. Denne typen økning i enzymaktivitet observeres for eksempel i foci av nekrose under hjerteinfarkt og under påvirkning av stråling.
Klinisk signifikans. Sykdommer assosiert med funksjonsfeil i lysosomer Noen ganger, på grunn av feil funksjon av lysosomer, utvikles lagringssykdommer, der enzymer ikke virker eller fungerer dårlig på grunn av mutasjoner. Et eksempel på lagringssykdommer er amaurotisk idioti på grunn av glykogenlagring. Bruddet av lysosomet og frigjøringen av hyaloplasma-splittende enzymer er ledsaget av en kraftig økning i deres aktivitet. Denne typen økning i enzymaktivitet observeres for eksempel i foci av nekrose under hjerteinfarkt og under påvirkning av stråling.
 Vakuoler Vakuoler er enkeltmembranorganeller som er "beholdere" fylt med vandige løsningerøkologisk og uorganiske stoffer. ER- og Golgi-komplekset deltar i dannelsen av vakuoler. Unge planteceller inneholder mange små vakuoler, som deretter, ettersom cellene vokser og differensierer, smelter sammen og danner én stor sentral vakuole. Den sentrale vakuolen kan oppta opptil 95 % av volumet til en moden celle kjernen og organellene presses mot cellemembran. Membranbegrensende plantevakuol, kalt tonoplast. Væsken som fyller plantevakuolen kalles cellesaft. Del cellesaft inkluderer vannløselige organiske og uorganiske salter, monosakkarider, disakkarider, aminosyrer, endelige eller giftige metabolske produkter (glykosider, alkaloider), noen pigmenter (antocyaniner).
Vakuoler Vakuoler er enkeltmembranorganeller som er "beholdere" fylt med vandige løsningerøkologisk og uorganiske stoffer. ER- og Golgi-komplekset deltar i dannelsen av vakuoler. Unge planteceller inneholder mange små vakuoler, som deretter, ettersom cellene vokser og differensierer, smelter sammen og danner én stor sentral vakuole. Den sentrale vakuolen kan oppta opptil 95 % av volumet til en moden celle kjernen og organellene presses mot cellemembran. Membranbegrensende plantevakuol, kalt tonoplast. Væsken som fyller plantevakuolen kalles cellesaft. Del cellesaft inkluderer vannløselige organiske og uorganiske salter, monosakkarider, disakkarider, aminosyrer, endelige eller giftige metabolske produkter (glykosider, alkaloider), noen pigmenter (antocyaniner).
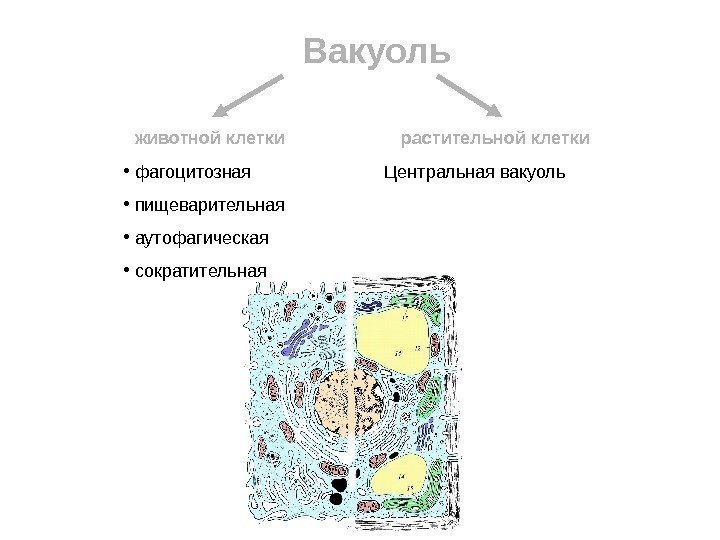 Vakuol dyrecelle Plante-celle fagocytotisk fordøyelseskanal autofagisk kontraktil Sentralvakuole
Vakuol dyrecelle Plante-celle fagocytotisk fordøyelseskanal autofagisk kontraktil Sentralvakuole
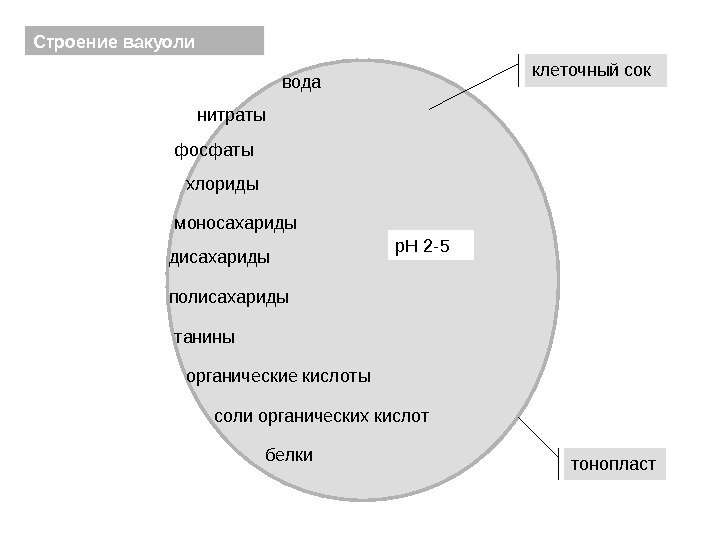 Vakuolens struktur tonoplastcelle saft vann nitrater fosfater klorider monosakkarider disakkarider tanniner organiske syrer salter organiske syrer s. H 2 -5 polysakkarider proteiner
Vakuolens struktur tonoplastcelle saft vann nitrater fosfater klorider monosakkarider disakkarider tanniner organiske syrer salter organiske syrer s. H 2 -5 polysakkarider proteiner
 Funksjon nr. 1 Opprettholde turgortrykk. Vakuolen fungerer som et osmometer og gir cellen nødvendig styrke og tetthet. Funksjon nr. 2 Noen ganger inneholder vakuoler løselige pigmenter. Denne gruppen inkluderer antocyaniner, som er røde, blå eller lilla i fargen, og noen relaterte forbindelser som er gule eller kremfargede. Det er disse pigmentene som hovedsakelig bestemmer fargen på blomster. Akkumulering av reservestoffer og "begravelse" av avfall, dvs. sluttproduktene av cellemetabolismen. Noen ganger ødelegger vakuoler stoffer som er giftige eller unødvendige for cellen. Funksjonsnr.
Funksjon nr. 1 Opprettholde turgortrykk. Vakuolen fungerer som et osmometer og gir cellen nødvendig styrke og tetthet. Funksjon nr. 2 Noen ganger inneholder vakuoler løselige pigmenter. Denne gruppen inkluderer antocyaniner, som er røde, blå eller lilla i fargen, og noen relaterte forbindelser som er gule eller kremfargede. Det er disse pigmentene som hovedsakelig bestemmer fargen på blomster. Akkumulering av reservestoffer og "begravelse" av avfall, dvs. sluttproduktene av cellemetabolismen. Noen ganger ødelegger vakuoler stoffer som er giftige eller unødvendige for cellen. Funksjonsnr.
 Peroksisomer er små vesikler som inneholder et sett med enzymer. Funksjoner: 1) Peroksisomer inneholder proteiner på overflaten av membranen, som fungerer som en reseptor som gjenkjenner signaler på det introduserte proteinet. Av alle peroksimproteinene er det mest kjente enzymet fra gruppen hydroperoksidaser - katalase 2) Delta i metabolske reaksjoner: i metabolismen av lipider, kolesterol, etc.
Peroksisomer er små vesikler som inneholder et sett med enzymer. Funksjoner: 1) Peroksisomer inneholder proteiner på overflaten av membranen, som fungerer som en reseptor som gjenkjenner signaler på det introduserte proteinet. Av alle peroksimproteinene er det mest kjente enzymet fra gruppen hydroperoksidaser - katalase 2) Delta i metabolske reaksjoner: i metabolismen av lipider, kolesterol, etc.
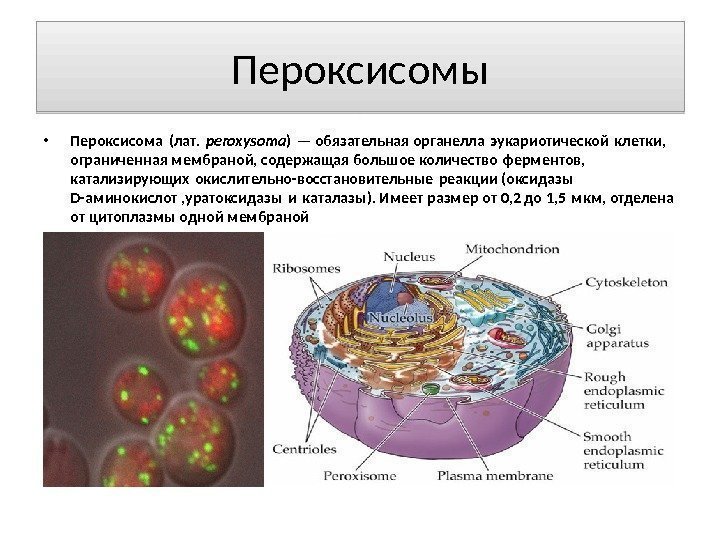 Peroksisomer Peroksisom (lat. peroksysoma) er en obligatorisk organell av en eukaryot celle, avgrenset av en membran, som inneholder et stort nummer av enzymer som katalyserer redoksreaksjoner (D-aminosyreoksidaser, uratoksidase og katalase). Har en størrelse fra 0,2 til 1,5 mikron, atskilt fra cytoplasmaet med en membran
Peroksisomer Peroksisom (lat. peroksysoma) er en obligatorisk organell av en eukaryot celle, avgrenset av en membran, som inneholder et stort nummer av enzymer som katalyserer redoksreaksjoner (D-aminosyreoksidaser, uratoksidase og katalase). Har en størrelse fra 0,2 til 1,5 mikron, atskilt fra cytoplasmaet med en membran
 Funksjoner av peroksisomer Settet med funksjoner til peroksisomer varierer i cellene forskjellige typer. Blant dem: oksidasjon av fettsyrer, fotorespirasjon, ødeleggelse av giftige forbindelser, syntese av gallesyrer, kolesterol og esterholdige lipider, konstruksjon av myelinskjeden nervefibre, fytansyremetabolisme osv. Sammen med mitokondrier er peroksisomer hovedforbrukerne av O 2 i cellen. Peroksisomet inneholder vanligvis enzymer som bruker molekylært oksygen til å abstrahere hydrogenatomer fra visse organiske substrater for å danne hydrogenperoksid: Catalase bruker det resulterende til å oksidere en rekke substrater - for eksempel fenoler, maursyre, formaldehyd og etanol: Denne typen oksidative reaksjoner er spesielt viktig i lever- og nyreceller, hvis peroksisomer nøytraliserer mange giftige stoffer kommer inn i blodet. Nesten halvparten av etanolen som kommer inn i menneskekroppen oksideres til acetaldehyd på denne måten. I tillegg har reaksjonen implikasjoner for avgiftning av cellen fra selve hydrogenperoksid.
Funksjoner av peroksisomer Settet med funksjoner til peroksisomer varierer i cellene forskjellige typer. Blant dem: oksidasjon av fettsyrer, fotorespirasjon, ødeleggelse av giftige forbindelser, syntese av gallesyrer, kolesterol og esterholdige lipider, konstruksjon av myelinskjeden nervefibre, fytansyremetabolisme osv. Sammen med mitokondrier er peroksisomer hovedforbrukerne av O 2 i cellen. Peroksisomet inneholder vanligvis enzymer som bruker molekylært oksygen til å abstrahere hydrogenatomer fra visse organiske substrater for å danne hydrogenperoksid: Catalase bruker det resulterende til å oksidere en rekke substrater - for eksempel fenoler, maursyre, formaldehyd og etanol: Denne typen oksidative reaksjoner er spesielt viktig i lever- og nyreceller, hvis peroksisomer nøytraliserer mange giftige stoffer kommer inn i blodet. Nesten halvparten av etanolen som kommer inn i menneskekroppen oksideres til acetaldehyd på denne måten. I tillegg har reaksjonen implikasjoner for avgiftning av cellen fra selve hydrogenperoksid.
 Ikke-membranorganeller Ribosomer finnes i alle typer celler (inkludert prokaryote). De kan ligge fritt i cytoplasmaet eller koble seg til membranene i ER. Finnes i mitokondrier og plastider. Struktur: Små sfæriske legemer dannet av to ulike underenheter - store og små, som består av 3-4 molekyler ribosomalt RNA og mer enn 50 proteinmolekyler. Ribosomer inneholder alltid magnesiumioner som støtter strukturen deres. Funksjoner: syntese av polypeptidkjeder (det andre stadiet av proteinsyntese er translasjon).
Ikke-membranorganeller Ribosomer finnes i alle typer celler (inkludert prokaryote). De kan ligge fritt i cytoplasmaet eller koble seg til membranene i ER. Finnes i mitokondrier og plastider. Struktur: Små sfæriske legemer dannet av to ulike underenheter - store og små, som består av 3-4 molekyler ribosomalt RNA og mer enn 50 proteinmolekyler. Ribosomer inneholder alltid magnesiumioner som støtter strukturen deres. Funksjoner: syntese av polypeptidkjeder (det andre stadiet av proteinsyntese er translasjon).
 Cellesenter Finnes i nesten alle dyreceller (unntatt noen typer protozoer) og noen planter. Fraværende i blomstrende og nedre sopp. Struktur: Består av to sentrioler plassert vinkelrett på hverandre. Sentriolen er en liten sylindrisk organell, hvis vegg er dannet av 9 grupper (tripletter) av tre smeltede mikrotubuli. Funksjoner: tar del i dannelsen av fisjonsspindelen (achromatinspindelen). Centrioler danner basallegemene til flimmerhår og flageller.
Cellesenter Finnes i nesten alle dyreceller (unntatt noen typer protozoer) og noen planter. Fraværende i blomstrende og nedre sopp. Struktur: Består av to sentrioler plassert vinkelrett på hverandre. Sentriolen er en liten sylindrisk organell, hvis vegg er dannet av 9 grupper (tripletter) av tre smeltede mikrotubuli. Funksjoner: tar del i dannelsen av fisjonsspindelen (achromatinspindelen). Centrioler danner basallegemene til flimmerhår og flageller.
 Mikrotubuli og mikrofilamentstruktur: Et komplekst system filamenter som trenger gjennom hele cytoplasmaet. Trådene er dannet av molekyler av ulike kontraktile proteiner (myosin, tubulin, etc.). Funksjoner: sammen med noen andre elementer danner de cytoskjelettet til cellene, sikrer intracellulær bevegelse av organeller, samt cellebevegelse, sammentrekning av muskelfibre danner trådene til den mitotiske spindelen
Mikrotubuli og mikrofilamentstruktur: Et komplekst system filamenter som trenger gjennom hele cytoplasmaet. Trådene er dannet av molekyler av ulike kontraktile proteiner (myosin, tubulin, etc.). Funksjoner: sammen med noen andre elementer danner de cytoskjelettet til cellene, sikrer intracellulær bevegelse av organeller, samt cellebevegelse, sammentrekning av muskelfibre danner trådene til den mitotiske spindelen
 Rød - kjerne Grønn - mikrotubuli Gul - Golgi-apparat
Rød - kjerne Grønn - mikrotubuli Gul - Golgi-apparat

Atlas: menneskelig anatomi og fysiologi. Fullstendig praktisk veiledning Elena Yurievna Zigalova
Membranorganeller. Transport over membraner
Menneskelige celler er preget av tilstedeværelsen av et stort antall intracellulære membraner, som danner flere rom (fra det engelske rommet - "compartment, compartment"), som skiller seg fra hverandre i struktur og funksjon: cytosol, kjerne, endoplasmatisk retikulum, Golgi-kompleks, mitokondrier, lysosomer, peroksisomer. På grunn av tilstedeværelsen av disse elementene skjer et stort antall forskjellige biokjemiske reaksjoner samtidig i cellen.
Alle membranorganeller er bygget av elementære membraner, hvis strukturelle prinsipp ligner strukturen til plasmalemmaet beskrevet ovenfor. Absorpsjonen av makromolekyler og partikler av celler skjer ved endocytose (fra det greske endon - "inne", kytos - "celle"), frigjøring - ved eksocytose (fra gresk exo - "utenfor", kytos - "celle").
En av essensielle funksjoner Plasmalemmaet er transport. La oss huske at de hydrofobe "halene" av lipider som vender mot hverandre forhindrer penetrasjon av polare vannløselige molekyler. Det er to typer transport: passiv og aktiv. Den første krever ikke energi, den andre er energiavhengig. Som regel bærer den indre (cytoplasmatiske) overflaten av membranen negativ ladning, som letter penetrasjonen av positivt ladede ioner inn i cellen. Vann kommer inn i cellen ved osmose(fra gresk osmos - "trykk, trykk"), som er den langsomme penetrasjonen av vann gjennom en semipermeabel membran som skiller to løsninger med forskjellige konsentrasjoner. Som et resultat utjevnes konsentrasjonene av disse to løsningene.
Diffusjon(fra latin diffusion - "spredning, spredning") er overgangen av ioner eller molekyler forårsaket av deres Brownske bevegelse gjennom membraner fra en sone hvor disse stoffene er i en høyere konsentrasjon til en sone med lavere konsentrasjon til konsentrasjonen er på begge sider av membranen er justert. Spesifikke transportproteiner innebygd i membranen transporterer små polare molekyler over den, med hvert protein som transporterer en klasse med molekyler eller bare en forbindelse. Noen transmembrane proteiner danner kanaler. Aktiv transport utføres av bærerproteiner, mens energi forbrukes som følge av hydrolyse av ATP (adenosintrifosfatsyre) eller protonpotensial. Aktiv transport skjer mot en konsentrasjonsgradient. For å utføre biokjemiske reaksjoner må stoffer komme inn i cellen gjennom endocytose og utskillelse av eksocytose metabolske produkter.
Endocytose. Det finnes flere metoder for endocytose. Inntreden av flytende kolloidale partikler kalles pinocytose, og store svevestøv- fagocytose. For at eksterne molekyler skal komme inn i cellen, må de først bindes av glykokalyx-reseptorer. Cytolemmaet begynner å invaginere, så kommer kantene nærmere og tettere, og splitter av vesikkelen som inneholder de fangede molekylene. Det dannes et endreom som er nedsenket i cytoplasmaet og møtes med lysosomer. Membranene deres smelter sammen. I det resulterende sekundære lysosomet gjennomgår stoffer som kommer inn i cellen ned.
Eksocytose sikrer fjerning av store molekylære forbindelser. Først segregerer de i Golgi-komplekset i form av transportvesikler og blir rettet mot celleoverflaten. Membranen til vesikelen er innebygd i cytolemma, og innholdet i vesikelen vises utenfor cellen.
To varianter er kjent endocytose: fagocytose - absorpsjon av partikler (fra den greske phagos - "sluker" og kytos - "celle") og pinocytose – absorpsjon av oppløste stoffer (fra gresk Pino - "drikke"). Den fagocyterte partikkelen som er innelukket i en membran kalles et fagosom. Under prosessen med endo- og eksocytose er transporterte stoffer innelukket i membranvesikler.
Endoplasmatisk retikulum, eller endoplasmatisk retikulum(ER), er et enkelt sammenhengende hulrom avgrenset av en membran som danner mange invaginasjoner og folder ( se fig. 1). Derfor, på elektrondiffraksjonsmønstre, vises det endoplasmatiske retikulum i form av mange rør, flate eller runde sisterner og membranvesikler. Det er to typer ER: granulære og agranulære. Siden som vender mot cytosolen til den første er dekket med ribosomer, mens siden av den andre er blottet for dem. Funksjon av granulær ER: syntese av proteiner ved ribosomer og transport av proteiner, jevn syntese og utveksling av karbohydrater og lipider (steroidhormoner, glykogen, kolesterol) og nøytralisering (hepatocytter), syntese av klorider, hvorfra det dannes i magen saltsyre. Å være et depot av kalsiumioner, er glatt ER involvert i muskelsammentrekning; avgrenser fremtidige blodplater i megakaryocytter. En av de viktigste funksjonene til ER er syntesen av membranproteiner og lipider for alle cellulære organeller.
Kompleks eller Golgi-apparat(CG), er et sett med tanker, bobler, plater, rør, sekker, avgrenset av en membran, der syntetiserte produkter akkumuleres og pakkes ( se fig. 1). Disse produktene fjernes fra cellen ved hjelp av de komplekse elementene i tillegg syntetiseres polysakkarider, protein-karbohydratkomplekser dannes og transporterte molekyler modifiseres. I et lysmikroskop vises CG i form av et nett eller et system av tubuli og vakuoler. CG er tilstede i alle menneskelige celler, unntatt erytrocytter og kåte skjell i epidermis. I de fleste celler er CG plassert rundt eller i nærheten av kjernen. I CG identifiseres tre membranelementer: flatede sekker (sisterner), vesikler og vakuoler. CG er en tredimensjonal koppformet struktur som består av flere (fra ett til flere hundre) diktyosomer (fra den greske dyktionen - "nettverk"). Hvert diktyosom inneholder 4–8 (i gjennomsnitt 6) parallelle flate cisterner, penetrert av porer med utvidede ender, hvorfra vakuoler som inneholder syntetiserte stoffer splittes av. Cisternene er assosiert med mange membranøse vesikler så vel som større sekretoriske granuler. Elementer av Golgi-komplekset er forbundet med kanaler.
Membranene til Golgi-komplekset dannes og vedlikeholdes av granulære endoplasmatisk retikulum, der membrankomponenter syntetiseres. De transporteres av transportvesikler som spirer fra ER og smelter sammen med CG, hvorfra sekretoriske vesikler stadig spirer, og cisternae-membranene fornyes stadig. De leverer glykokalyxen og syntetiserte stoffer til plasmalemmaet, og sikrer dermed fornyelsen av plasmalemmaet. En av de viktigste funksjonene til CG er proteinsortering.
Lysosomer- membranorganeller som inneholder ca. 50 typer forskjellige hydrolytiske enzymer, som syntetiseres på ribosomer i det granulære endoplasmatiske retikulum, hvorfra de overføres med transportvesikler til CG, hvor de modifiseres. Primære lysosomer knopper fra overflaten av CG. Alle lysosomer i cellen danner et enkelt lysosomalt rom, der et surt pH-miljø konstant opprettholdes, fra 3,5–5,0. Membranene til lysosomer er motstandsdyktige mot enzymene som finnes i dem og beskytter cytoplasmaet fra deres virkning.
Det er fire funksjonelle former lysosomer Primære lysosomer, spiret fra Golgi-komplekset, smelter sammen med fagosomet, form sekundært lysosom(phagolysosom), der fordøyelsen av absorberte stoffer til monomerer skjer. Sistnevnte transporteres gjennom lysosommembranen inn i cytosolen. Ufordøyde stoffer forblir i lysosomet, noe som resulterer i dannelsen gjenværende kropp. I tillegg fordøyer lysosomer skadede strukturer i sin egen celle ( autolysosom).
Peroksisomer er vesikler med en diameter på 0,2 til 0,5 mikron, omgitt av membraner, som inneholder oksidative enzymer (omtrent 40 % av alle proteiner er katalase) som produserer og ødelegger hydrogenperoksid. De bruker molekylært oksygen.
Mitokondrier, som er "cellens energistasjoner", er involvert i prosessene med cellulær respirasjon og konvertering av energi til en form som er tilgjengelig for bruk av cellen. I et lysmikroskop vises mitokondrier som runde, langstrakte eller stavformede strukturer 0,3–5,0 µm lange og 0,2–1,0 µm brede. Antall, størrelse og plassering av mitokondrier avhenger av cellens funksjon og dens energibehov. Dermed når antallet i hver levercelle 2500. Med hjelpen elektronmikroskopi Det er fastslått at mitokondrier er organeller med doble membraner ( ris. 5). Mellom den ytre og indre mitokondriemembranen er det et intermembranrom. Den indre membranen danner mange folder, eller cristae, på grunn av hvilke den indre membranen øker dramatisk. På indre overflate cristae ligger mange elektrontette submitokondrier elementærpartikler(opptil 4000 per 1 mikron 2 membraner), med form som en sopp. I rommet begrenset av den indre mitokondriemembranen er det en finkornet matrise.
Ris. 5. Mitokondrier (ifølge B. Alberts et al.; ifølge C. de Duve, med endringer). JEG - generell ordning bygninger: 1 – ytre membran; 2 - indre membran; 3 - cristae; 4 - matrise; II - diagram av strukturen til crista: 5 - fold av den indre membranen; 6 - soppformede kropper
Mitokondrier inneholder sitt eget DNA, RNA og ribosomer, som er lokalisert i matrisen. Dermed er mitokondrier utstyrt med sine egne genetisk system, nødvendig for deres selvreproduksjon og proteinsyntese. Det bør understrekes at mitokondrielt DNA, RNA og ribosomer skiller seg fra cellens egne og ligner veldig på prokaryote.
MERK FØLGENDE
Hos pattedyr, inkludert mennesker, arves mitokondriegenomet fra moren.
Mitokondrier formerer seg ved å dele eksisterende, uavhengig av delingen av andre mitokondrier og selve cellen.
Skjer konstant i cellene metabolisme(fra det greske metabolske - "endring, transformasjon"), eller metabolisme, som er et sett med prosesser assimilering(reaksjoner av biosyntese av komplekse biologiske molekyler fra enklere) og dissimilering(spaltningsreaksjoner). Som et resultat av dissimilering vil energien som finnes i kjemiske bindinger stoffer. Denne energien brukes av cellen til å utføre ulike jobber, inkludert assimilering. La oss huske at energi verken blir skapt eller ødelagt, den går bare fra en form til en annen, egnet for arbeid. Cellen bruker energien som finnes i de kjemiske bindingene til aminosyrer, monosakkarider og fettsyrer. De dannes som et resultat av fordøyelsen fra proteiner, karbohydrater og fett og kommer inn i cellen.
La oss vurdere energimetabolisme ved å bruke eksemplet på nedbrytning av glukose. Glukose transporteres gjennom plasmamembran, og dens oksygenfrie nedbrytning, eller glykolyse, skjer i cytoplasmaet. Glykolyse er en flertrinns enzymatisk prosess, som et resultat av at to molekyler pyrodruesyre og to ATP-molekyler dannes fra ett glukosemolekyl (som tar hensyn til de to ATP-molekylene som brukes til å utføre reaksjonene). Pyrodruesyre gjennomgår ytterligere oksidasjon (aerob med deltagelse av oksygen) i mitokondrier, som inneholder enzymkjeder som katalyserer reaksjonene til syntesen av ATP (adenosintrifosfat). ATP er en universell bærer og hovedenergiakkumulatoren i cellen. Energien er inneholdt i høyenergibindinger mellom fosforsyrerester.
Når én fosfatgruppe fjernes fra ATP, dannes ADP (adenosin difosfatsyre) og fosfat og fri energi frigjøres som brukes av cellen til å utføre arbeid. I mitokondrier kombineres ADR med en fosforsyrerest for å danne ATP. Som et resultat av glykolyse frigjøres bare ca. 5 % av energien i mitokondrier i prosessen med aerob oksidasjon og lagres i ATP. Per ett glukosemolekyl dannes 36 ATP-molekyler.
Kjerne - hoved- cellestruktur, er tilstede i alle menneskelige celler unntatt erytrocytter og blodplater. I de fleste celler er formen sfærisk eller eggformet, men andre former for kjernen finnes også (ringformet, stavformet, spindelformet, perleformet, bønneformet, segmentert, polymorf, etc.). Størrelsen på kjernene varierer mye fra 3 til 25 mikron. Eggcellen har den største kjernen. De fleste menneskelige celler er mononukleære, men det er binukleære (for eksempel noen nevroner, hepatocytter, kardiomyocytter), og noen strukturer er multinukleære (myosymplast muskelfibre).
I kjernen er det følgende strukturer: nukleær konvolutt, kromatin, nukleolus og nukleoplasma. Kjernen er omgitt av en kjernefysisk konvolutt, bestående av en indre og ytre kjernemembran, hver 8 nm tykk, atskilt av et perinukleært rom (eller atomhylse cisterna) 20–50 nm bredt. Begge er elementære cellemembraner. Ribosomer er festet til den ytre, som går inn i det granulære endoplasmatiske retikulumet. Det perinukleære rommet danner et enkelt hulrom med det endoplasmatiske retikulum ( se fig. 1).
Kjernefysiske konvolutten penetreres av mange ordnede runde atomporer med en diameter på 50–70 nm, som Total okkuperer opptil 25% av overflaten av kjernen. Gjennom kjerneporer skjer selektiv transport av store partikler, samt utveksling av stoffer mellom kjernen og cytosol.
I levende celler er karyoplasma (nukleoplasma) homogen (bortsett fra nukleolen). Etter fiksering og prosessering av vev for lys- eller elektronmikroskopi, blir to typer kromatin synlige (fra gresk kroma - "maling"); godt farget heterokromatin (inaktivt) og lett eukromatin (aktivt).
I den delende kjernen spiraler kromatinet for å danne kromosomer. Kromatinet til den ikke-delte kjernen og kromosomene til den delende kjernen dannes av deoksyribonukleinsyre (DNA) assosiert med RNA og proteiner (histoner og ikke-histoner). Den kjemiske identiteten til kromatin og kromosomer bør understrekes.
I somatiske celler Det er to kopier av hvert kromosom, de kalles homologe. De er identiske i lengde, form, struktur, stripearrangement og bærer de samme genene, som er lokalisert på samme måte. Normal karyotype menneske (fra gresk karyon - "kjernen av en nøtt", skrivefeil - "prøve") inkluderer 22 par autosomer og ett par kjønnskromosomer (XX hunn eller XV hann) ( ris. 6).

Ris. 6. Menneskelig karyotype (frisk mann). jeg - karyotype, generell form; II - metafase kromosomer
Gene en del av DNA karakterisert ved en spesifikk nukleotidsekvens som er ansvarlig for syntesen av et spesifikt protein. Genet er den elementære enheten for arv.
Nukleolen (en eller flere) påvises i alle ikke-delte kjerner i form av en tett, intenst farging avrundet homogen basofil kropp, hvis størrelse er proporsjonal med intensiteten protein syntese. Ribosomer dannes i kjernen. Kjernejuice er den ikke-fargede elektronlyse delen av kjernen - en kolloid løsning av proteiner som omgir kromatinet og nukleolen.
Fra boken Morsomme øvelser for tungen forfatter Olga NovikovskayaHVA ER TRANSPORTENE Se, flyet flyr, nynner, en modig pilot sitter i den brede tuppen av tungen din og uttal lyden [L]. mens du løfter armene til sidene og svinger dem som vinger Vel, åh dette
Fra boken Morsomme øvelser for tungen forfatter Olga NovikovskayaHVA ER TRANSPORTEN Se, flyet flyr, nynner, en modig pilot sitter i det. heve armene til sidene og svinge dem som vinger Vel, denne fallskjermen. Ned oss
Fra boken Medisinsk fysikk forfatter Vera Aleksandrovna Podkolzina Fra boken Histologi forfatter V. Yu Barsukov1. Introduksjon til histologisk forløp. Celleorganeller Histologi er vitenskapen om strukturen, utviklingen og vitale funksjonene til vev i levende organismer. Følgelig studerer histologi et av nivåene for organisering av levende materie - vev Hovedobjektet for histologi i systemet
Fra boken Histologi forfatter V. Yu Barsukov4. Morfologi og funksjoner til celleorganeller (forts.) Lysosomer er de minste organellene i cytoplasmaet, de er legemer avgrenset av en bilipidmembran. Lysosomes funksjon er å sikre intracellulær fordøyelse, dvs. nedbryting av både eksogene og.
Fra bok Normal fysiologi forfatter Nikolay Alexandrovich AgadzhanyanGassutveksling og gasstransport Gassutveksling av O2 og CO2 gjennom alveolær-kapillærmembranen skjer gjennom diffusjon, som skjer i to trinn. I det første trinnet skjer diffusjonsoverføringen av gasser gjennom den lufthematiske barrieren, i det andre trinnet skjer det
Fra bok Levende vann. Hemmelighetene til cellulær foryngelse og vekttap forfatter Lyudmila RudnitskayaVI STYRKER CELLEMEMBRANENE MED ØVELSER Om fordelene fysisk trening mye har blitt sagt. Regelmessig trening forbedrer funksjonen til karsystemet, normaliserer blodtrykk og sukkernivåer, bygger muskelmasse og forbrenner fett. Det er under trening at vår
Fra bok Hemmelig visdom Menneskekroppen forfatter Alexander Solomonovich ZalmanovMembraner Livet er den evige bevegelsen av væsker mellom celler og inne i cellene. Å stoppe denne bevegelsen resulterer i døden. En delvis nedgang i bevegelsen av væsker i et organ forårsaker delvis forstyrrelse. Generell nedbremsing av ekstra- og intracellulære væsker i
Etter hundre år er jeg en ung 20-åring jeg er Guds Ånd, i henhold til Guds hellige vilje blir jeg stadig sunnere og sterkere, nervene mine blir stadig sunnere og sterkere, jeg utvikler meg hele tiden, akselererer gudgitt konstant helbredelse og foryngelse, jeg blir alltid 20 år gammel, selv etter fem år 20 år. Meg og gjennom
Fra boken Alt pusteøvelser. For helsen til de som bryr seg... forfatter Mikhail Borisovich IngerleibKapittel 3. Transport av gasser med blod «Bærer» av oksygen fra lungene til vev og organer og karbondioksid fra vev og organer til lungene er blod. I en fri (oppløst) tilstand overføres en så liten mengde gasser at den trygt kan neglisjeres når
Fra bok Godt syn- Klart sinn lange år! Østens eldste praksis forfatter Andrey Alekseevich LevshinovPust inn gjennom nesen, pust ut gjennom munnen Lær å puste med magepusting slik at du puster inn gjennom nesen og puster ut gjennom munnen (munnen er bare litt åpen, leppene er foldet som et lite rør) - mykt, jevnt. uten
Fra boken The Science of Breathing of Indian Yogis forfatter William Walker AtkinsonKapittel VI PUSTE GJENNOM NESENE OG PUSTE GJENNOM MUNNEN En av de første leksjonene i vitenskapen om pust til yogier er viet til å lære å puste gjennom nesen og vinne vanlig vane- pust gjennom munnen Den menneskelige respirasjonsmekanismen lar ham puste gjennom både nesen og munnen, men for ham er saken sann
Fra boken Conspiracies of a Siberian healer. Utgave 33 forfatter Natalya Ivanovna Stepanova Fra Giris bok. Sport av sterk og sunn forfatter Alexey Ivanovich VorotyntsevKastblokk over hodet (kast over hodet) Kast over hodet - kaste en vekt over hodet og deretter motta den med samme hånd med en rotasjon av overkroppen 180°. Alle kast over hodet utføres med en torsovending på 180°, så når du syner et kast, blir turn