Pedagogiske stoffer pedagogisk vev
(meristemer), plantevev som består av celler som beholder evnen til å dele seg i lang tid. Takket være den gjentatte delingen av disse cellene vokser planter gjennom hele livet (for noen trær er dette hundrevis eller til og med tusenvis av år). Den andre viktige egenskapen til meristemceller er at de gir opphav til spesialiserte celler som danner permanente vev - integumentære, basale, ledende, mekaniske, ekskresjonelle. Avhengig av fordelingen på kroppen til den utviklende planten, skilles fire typer meristemer. Apikale, eller apikale, meristemer sørger for vekst av skudd og røtter i lengden. Laterale, eller laterale, meristemer bestemmer veksten av stengler og røtter i tykkelse og kalles kambium. Intercalary, eller intercalary, meristemer lagres midlertidig i internodene på stilken og ved bunnen av unge blader, noe som sikrer veksten av disse områdene, men blir deretter til permanent vev. Sår, eller traumatiske, meristemer oppstår på steder hvor planten er skadet, hvor de danner en beskyttende ring oss. Spørsmålet er hvordan forskjellige vev dannes fra identiske meristematiske celler, dvs. hvordan celledifferensiering skjer forblir uløst.
.(Kilde: "Biology. Modern illustrated encyclopedia." Sjefredaktør A. P. Gorkin; M.: Rosman, 2006.)
Se hva "pedagogiske stoffer" er i andre ordbøker:
UTDANNINGSTOFF- se meristematiske vev... Ordbok over botaniske termer
videregående opplæringsvev- Synonymer: sekundære meristemer er utdanningsvev som oppstår enten fra primære meristemer eller fra permanente vev. Representert av kambium, fellogen og sårmeristemer...
primære utdanningsvev- Synonymer: primære meristemer, promeristemer, pedagogisk vev avledet direkte fra meristemene til embryoet; Cellene deres har i utgangspunktet evnen til å dele seg. Til P. o. t. inkludere apikale merister av skudd og rot, prokambium, ... ... Anatomi og morfologi av planter
PLANTEVEV- grupper eller komplekser av celler relatert etter felles struktur, opprinnelse, funksjoner og plassering. I samsvar med dette skilles pedagogisk, integumentær, grunnleggende, mekanisk, ledende og ekskretorisk T. r. Pedagogiske stoffer, eller... ... Agricultural Encyclopedic Dictionary
plantevev- plantevev, grupper eller komplekser av celler knyttet til felles struktur, opprinnelse, funksjoner og plassering. I samsvar med dette skilles pedagogisk, integumentær, grunnleggende, mekanisk, ledende og ekskretorisk T. r. Jordbruk. Stor encyklopedisk ordbok
Cellesystemer, som er like i struktur, opprinnelse og funksjon, er forskjellige i størrelse, form og plassering. Sammensetningen av vevet inkluderer for eksempel vevsvæske (fyller de intercellulære rommene) og stoffer som befinner seg mellom cellene. kalsiumsalter ... ... Biologisk leksikon ordbok
Modell av det apikale meristemet "tunica corpus". Meristemer, eller pedagogisk vev, eller meristematisk vev (gammelgresk ... Wikipedia
Tunica body Model Meristems, eller Educational tissues, eller Meristematic tissues (gresk meristos divisible) er et generelt navn for plantevev som består av intensiv deling og opprettholdelse av fysiologisk aktivitet gjennom hele ... ... Wikipedia
Tunica body Model Meristems, eller Educational tissues, eller Meristematic tissues (gresk meristos divisible) er et generelt navn for plantevev som består av intensiv deling og opprettholdelse av fysiologisk aktivitet gjennom hele ... ... Wikipedia
Planteorganer består av vev. Plantevev er en samling av celler som har en lignende struktur, homogen opprinnelse og utfører samme funksjon. Vevet til høyere landplanter har nådd spesiell utvikling. I planter skilles pedagogisk vev ut: integumentært, mekanisk, ledende, grunnleggende og ekskretorisk.
Pedagogisk vev, eller meristem, tjener i planten for vekst de er av primær opprinnelse og sekundær. Det primære utdanningsvevet er lokalisert i vekstkjeglene til røtter og stilker. La oss først se på rotvekstkjegle(Fig. 1, I).
Figur 1. Primært utdanningsvev i rotvekstkjeglen:
JEG - rotvekstkjegle: 1 - celler av pedagogisk vev; 2 - celler av rothetten. II - påfølgende faser av kompleks kjerne- og celledeling(karyokinesis eller mitose): 1 - interfase; 2 - profase; 3 - metafase; 4-5 - anafase; 6-7 - telofase; 8 - cytokinese.
Spissen av roten har en kjegleformet form, dekket med en rothette, under hvilken det primære utdanningsvevet er plassert. Cellene i utdanningsvev er tetraedriske eller mangefasetterte, har tett protoplasma, store kjerner og tynne cellulosemembraner. Vakuolene deres er ikke utviklet eller dårlig utviklet. Disse cellene har evnen til å dele seg kraftig. Celledeling innledes av kompleks deling av kjernen, der det oppstår særegne endringer. På ferdiglagde, godt tilberedte og fargede preparater er kjerner synlige som er i forskjellige faser av kompleks deling, ellers kalt karyokinesis ("karyon" - kjerne, "kinesis" - bevegelse) (fig. 1, II). Tallrike observasjoner har slått fast at i kjernen oppstår først tynne kromatinfilamenter fra kromatinstoffet, som forkortes, tykner og blir til individuelle partikler - kromosomer ("kromo" - farge, "soma" - kropp), ofte buet i form av en hestesko (divisjonsprophase) . Hvert kromosom i profase ser ut til å være delt på langs i to halvdeler, men i virkeligheten forekommer kromosomdobling (reduplikasjon). Nukleolus og kjernefysisk konvolutt forsvinner gradvis. Kromosomene er plassert i den midtre delen av kjernen i ett plan i form av en plate. Halvdelene av hvert kromosom ("datter" kromosomer) passer tett til hverandre, og i denne fasen (metafasen) på tverrgående deler av roten kan du enkelt telle antall kromosomer og undersøke formen deres. Ved polene til kjernen (selv før forsvinningen av kjernemembranen) vises et system av tynne akromatiske (ikke-fargende) filamenter, arrangert i form av to hetter. Trådene vokser og tar form av en spindel. Etter dette begynner divergensen av halvdelene av kromosomene (datterkromosomene) mot de to motsatte polene til cellen (anafase). Divergensen av kromosomer ender med at datterkromosomene, samlet ved cellens poler, flettes sammen med hverandre og blir til to kuler (telofase). Samtidig vises pektinfortykninger på akromatintrådene i den midtre delen av cellen - det dannes en fragmoplast, det vil si rudimentet til den tverrgående membranen. Til slutt er kjernene omgitt av en membran, nukleoler (en eller flere) dannes i dem, og protoplasmaet deles av en tverrgående membran (cytokinesis), noe som resulterer i utseendet til to nye celler.
Prosessen med kjernefysisk og celledeling skjer oftere om natten, hastigheten avhenger av temperatur og fuktighet. Delingsprosessen varer i gjennomsnitt 75-120 minutter, men varer noen ganger flere timer, med den første fasen (profasen) som bruker mest tid. Under celledeling oppstår cellemembranen som et produkt av den vitale aktiviteten til protoplasma og kjerne. Først vises medianplaten. Den består av et intercellulært stoff som limer sammen de primære cellulosemembranene som tilhører to celler.
For første gang ble divergensen av kromosomer mot cellens poler, så vel som andre detaljer om delingen av kjernen og cellen, etablert i 1875 av professor I. D. Chistyakov ved Moskva-universitetet.
Essensen av den komplekse delingen av kjernen og cellen, også kalt mitose, ligger tilsynelatende i det faktum at ved jevn deling av kromosomer i to halvdeler (eller mer presist, dobling av kromosomer), mottar datterceller fra moren mer eller mindre like mengder kromatinsubstans (deoksyribonukleinsyre - DNA), som overfører arv fra modercellen til dattercellene. ("Mitos" er en tråd. Det er vanligvis akseptert at mitose = karyokinesis + cytokinesis). Formen og antallet kromosomer er spesifikke egenskaper for hver planteart (myk hvete 2n = 42 kromosomer, durumhvete 2n = 28, bygg 2n = 14 osv.). Noen ganger er det imidlertid variasjon i antall kromosomer, og celler med forskjellige kromosomer kan observeres i en organisme.
I tillegg til mitose (eller karyokinese), skjer direkte deling av cellekjernen (amitose) i planter - en enkel trekking av kjernen, og noen ganger hele cellen, i to halvdeler.
Sammen med mitose er reduksjon nukleær deling vanlig i planter - meiose, hvor antall kromosomer i nye celler er halvert sammenlignet med modercellen. Denne delingen skjer i planter under sporulering, hvoretter mannlige og kvinnelige kjønnsceller (gameter) oppstår og utvikler seg. Sistnevnte har halvparten av antallet kromosomer i kjernene sine, og slike kjerner kalles haploide i motsetning til de diploide kjernene til vegetative celler som utgjør plantenes vegetative organer. Planters seksuelle prosess består av sammensmelting av to gameter (hann og hunn). Hvis det ikke var noen reduksjon av kromatinstoffet, det vil si en halvering av dets masse, ville i hver ny seksuell generasjon denne substansmassen, og derfor antallet kromosomer, uunngåelig øke. I kjønnsceller (gameter) er kjernene haploide, og først etter befruktningsprosessen blir cellekjernene diploide igjen.
Reduksjonsdivisjon ble oppdaget av forskeren V.I. Belyaev, og forskeren S.G. Navashin oppdaget dobbel befruktning i blomstrende planter og er grunnleggeren av en hel skole med forskere som studerer alle detaljene i kjernefysisk og celledeling.
Primært utdanningsvev gir opphav til andre vev, siden cellene endrer seg, differensierer. Cellenes evne til å dele seg raskt i stilkene og røttene til tofrøbladede planter er bevart for livet i meristematiske celler kalt kambium, på grunn av hvilke stilken og roten tykner. Sekundært utdanningsvev inkluderer de som oppstår fra annet vev. Et eksempel er fellogen, som ofte oppstår fra grunnvevet.
STOFFER. KLASSIFISERING AV STOFFER.
Organiseringen av høyere planter er basert på prinsippet om cellespesialisering, som ligger i det faktum at hver celle i kroppen ikke utfører alle sine iboende funksjoner, men bare noen, men mer fullstendig og perfekt.
Stoffer- stabile, naturlig repeterende komplekser av celler, like i opprinnelse, struktur og tilpasset til å utføre en eller flere funksjoner.
Det finnes ulike klassifiseringer av stoffer, men de er alle ganske vilkårlige.
Avhengig av hovedfunksjonen skilles flere grupper av plantevev ut.
1. pedagogiske stoffer, eller merister,- ha evnen til å dele og danne alt annet vev.
2. Dekker vev:
Hoved;
Sekundær;
Tertiær.
3. Hovedstoffer- utgjør det meste av plantekroppen. Følgende hovedstoffer skilles ut:
Assimilering (klorofyllbærende);
Stockers;
Luftbåren (aerenchyma);
Akviferer.
4. Mekaniske stoffer(støttende, skjelett):
Collenchyma;
Sklerenkym.
5. Ledende stoffer:
Xylem (tre) er et stigende vev;
Floem (floem) er et vev med synkende strømning.
6. Utskillelsesvev:
Utvendig:
Kjertelhår;
Hydatoder er vannstomata;
Sunbirds;
Innvendig:
Utskillelsesceller med essensielle oljer, harpiks, tanniner;
Flercellede beholdere for sekreter, melkemidler.
Basert på cellenes evne til å dele seg, skilles to typer vev: pedagogisk, eller merister, Og fast- integumentær, ekskretorisk, grunnleggende, mekanisk, ledende.
Stoffet heter enkel, hvis alle cellene er identiske i form og funksjon (parenchyma, sclerenchyma, collenchyma). Kompleks vev består av celler som er forskjellige i form, struktur og funksjon, men som er beslektet med en felles opprinnelse (for eksempel xylem, floem).
Det er også en klassifisering av vev basert på deres opprinnelse (ontogenetisk). I henhold til denne klassifiseringen skilles primært og sekundært vev. Fra det primære meristemet som ligger på toppen av skuddet og spissen av roten, samt fra frøembryoet, primære konstanter vev (epidermis, collenchyma, sclerenchyma, assimilasjonsvev, epiblema). Celler av permanent vev er ikke i stand til ytterligere deling. Fra celler av et spesialisert meristem - prokambia - dannes primære ledere vev (primært xylem, primært floem).
Fra det sekundære meristem - kambium - dannes sekundær vev: sekundær xylem, sekundær floem; fra felogen det dannes plugg, phelloderm og linser, som oppstår når stilken og roten tykner. Sekundært vev finnes vanligvis i gymnospermer og tofrøbladede angiospermer. Den kraftige utviklingen av sekundærvev - tre og bast - er karakteristisk for treaktige planter.
UTDANNINGSTOFF
Pedagogiske stoffer Takket være den konstante mitotiske delingen av cellene deres, sikrer de dannelsen av alt plantevev, dvs. faktisk forme kroppen hans. Enhver celle i sin utvikling går gjennom tre stadier: embryonal, vekst og differensieringsstadium (det vil si at cellen får en viss funksjon). Når embryoet differensierer, blir det primære meristemet kun bevart på spissen av det fremtidige skuddet (i vekstkjeglen) og på spissen av roten - apikal (apikal) merister. Embryoet til enhver plante består av meristemceller.
Cytologiske egenskaper ved meristemer. Typiske trekk kommer tydeligst til uttrykk i de apikale meristemene. Disse meristemene er sammensatt av isodiametriske polyedriske celler som ikke er atskilt av intercellulære rom. Skallene deres er tynne, inneholder lite cellulose og er strekkbare.
Hulrommet til hver celle er fylt med tett cytoplasma med en relativt stor kjerne som opptar en sentral posisjon og intensivt deler seg ved mitose. Hyaloplasmaet inneholder mange diffust spredte ribosomer, proplastider, mitokondrier og diktyosomer. Det er få vakuoler og de er små. Ledende vev er dannet av et meristem som har en prosenkymal form og store vakuoler - prokambium og kambium. Prokambiumceller er polygonale i tverrsnitt, kambiumceller er rektangulære.
Celler som beholder sine meristematiske egenskaper fortsetter å dele seg, og danner flere og flere nye celler kalt initialer. Noen av dattercellene differensierer og blir til celler i forskjellige vev, kalles de derivater av initialer. Initialceller kan dele seg uendelig mange ganger, og derivater av initialer deler seg en eller flere ganger og utvikler seg til permanent vev.
Basert på deres opprinnelse skilles primære og sekundære meristemer.
Primære merister
Primære meristemer oppstår direkte fra meristem til embryoet og har evnen til å dele seg. I henhold til deres plassering i planten kan primære meristemer være apikale (apikale), interkalære (interkalære) og laterale (laterale).
Apikale (apikale) meristemer- slike merister som er plassert i voksne planter på toppen av stengler og røtter og sikrer kroppsvekst i lengden. I stilkene, i vekstkjeglen, skilles to meristematiske lag: tunikaen, hvorfra integumentært vev og den perifere delen av primærbarken dannes, og corpus, hvorfra den indre delen av primærbarken og den sentrale aksial sylinder er dannet (fig. 2.3).
Ris. 2.3. Apikale meristem av stammen: EN- lengdesnitt: 1 - vekstkjegle; 2 - blad primordium; 3 - tuberkel av aksillær knopp;
Det er tre lag ved rotspissen:
1) dermatogen, hvorfra det primære integumentære-absorberende vevet - rhizoderm - dannes;
2) periblema, hvorfra vevet i den primære cortex utvikler seg;
3) plerome, danner vevet til den sentrale aksiale sylinderen.
de kan ha primær eller sekundær opprinnelse på et tverrsnitt av aksialorganene de ser ut som ringer. Et eksempel på et primært lateralt meristem er procambium og pericycle. Fra prokambia Kambiet og de primære elementene i de vaskulære-fibrøse buntene (primært floem og primært xylem) dannes, mens prokambiumcellene differensierer direkte til cellene i det primære ledende vevet.
De laterale meristemene er plassert parallelt med overflaten av organet og sikrer vekst av aksialorganene i tykkelse.
Interkalære merister oftere er de primære og er bevart i form av separate områder i soner med aktiv vekst i forskjellige deler av planten (for eksempel ved bunnen av bladstilker, ved bunnen av internoder). Ved bunnen av internodene i korn fører aktiviteten til dette meristemet til forlengelse av internodene, noe som sikrer veksten av stilken i lengden.
Sekundære merister
Sekundære meristemer inkluderer laterale og sårmeristemer.
Laterale (laterale) meristemer presentert kambium Og fellogen. De er dannet fra promerister (prokambium) eller permanente vev ved deres dedifferensiering. Kambiumceller er delt med skillevegger parallelt med overflaten av organet (periklinalt). Elementer av sekundært floem utvikles fra celler avsatt utover av kambium, og elementer av sekundært xylem utvikles fra celler avsatt innover. Kambiet, som oppsto fra permanent vev gjennom dedifferensiering, kalles ytterligere I struktur og funksjon skiller den seg ikke fra kambium, som oppsto fra promerister. Fellogen dannes fra permanent vev som ligger i de subepidermale lagene (under epidermis). Ved å dele periklinalt skiller phellogen fremtidige pluggceller (phelleme) utover, og phellodermceller innover. Dermed danner fellogen et sekundært integumentært vev - periderm. De laterale meristemene er plassert parallelt med overflaten av organet og sikrer vekst av aksialorganene i tykkelse.
Sårmerister dannes når vev og organer er skadet. Rundt skaden dedifferensierer levende celler, begynner å dele seg og transformeres derved til et sekundært meristem. Deres oppgave er å danne et tett beskyttende vev som består av parenkymceller - ring oss. Dette vevet er hvitaktig eller gulaktig i fargen, cellene har store kjerner og ganske tykke cellevegger. Callus oppstår under poding, noe som sikrer sammensmelting av scion med rotstokken, og ved bunnen av stiklingene. Den kan danne tilfeldige røtter og knopper, så den brukes til å oppnå isolerte vevskulturer.
DEKKVEV
Primært integumentært vev
TIL primær integumentær vev inkluderer epidermis, epidermis selv, parastomatale celler, vaktcellene i stomata og trikomene.
Pektinstoffer og cellulose som inngår i celleveggen kan bli utsatt for slimdannelse med dannelsen slim Og tannkjøttet. De er polymere karbohydrater relatert til pektinstoffer og kjennetegnes ved deres evne til å svelle kraftig når de kommer i kontakt med vann. Tannkjøtt i hoven tilstand er klissete og kan trekkes ut i tråder, mens slim er veldig uskarpt og ikke kan trekkes ut i tråder. Pektinslim finnes i representanter for familiene Liliaceae, Cruciferae, Malvaceae, Linden og Rosaceae, i motsetning til celluloseslim, som er mye mindre vanlige (for eksempel hos orkideer).
 Stomata
De er høyt spesialiserte formasjoner av epidermis, bestående av to bønneformede beskyttelsesceller og en stomatal fissur (et slags intercellulært rom mellom dem). De finnes hovedsakelig i bladene, men finnes også i stilken (fig. 2.6).
Stomata
De er høyt spesialiserte formasjoner av epidermis, bestående av to bønneformede beskyttelsesceller og en stomatal fissur (et slags intercellulært rom mellom dem). De finnes hovedsakelig i bladene, men finnes også i stilken (fig. 2.6).
Ris. 2.6. Stomatal struktur: a, b- huden til et timianblad (ovenfra og tverrsnitt); V- skrell fra stammen til Cereus (kaktusfamilien); 1 - faktiske epidermale celler; 2 - vaktceller i stomata; 3 - stomatal fissur; 4 - lufthule; 5 - celler av klorofyllbærende parenkym; A – neglebånd; B - kutikulært lag - skall med suberin og voks; B - celluloselag av veggen; G - kjerne med kjerne; D – kloroplaster
Veggene til vaktcellene er ujevnt fortykket: veggene rettet mot gapet (abdominal) er betydelig tykkere sammenlignet med veggene rettet bort fra gapet (dorsal). Gapet kan utvide seg og trekke seg sammen, og regulere transpirasjon og gassutveksling. Under gapet er det et stort åndedrettshule (intercellulært rom), omgitt av mesofyllceller i bladet.
 Vaktcellene er omgitt av parastomatale celler, som sammen danner stomatal kompleks(Fig. 2.7). Følgende hovedtyper av stomatale komplekser skilles ut:
Vaktcellene er omgitt av parastomatale celler, som sammen danner stomatal kompleks(Fig. 2.7). Følgende hovedtyper av stomatale komplekser skilles ut:
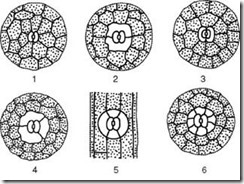
Ris. 2.7. Hovedtyper av stomatale apparater: 1 - anomocytisk (i alle høyere planter, unntatt kjerringrokk); 2 - diacytisk (i bregner og blomstrende planter); 3 - paracytisk (i bregner, kjerringrokk, blomstrende og undertrykkende); 4 - anisocytisk (bare i blomstrende planter); 5 - tetracytisk (hovedsakelig i monocots); 6 - ancyklocytisk (i bregner, gymnospermer og blomstrende planter)
1) anomocytisk(uordnet) – vaktceller har ikke klart definerte parastomatale celler; karakteristisk for alle høyere planter, unntatt bartrær;
2) anisocytisk(ulik celle) - vaktcellene i stomata er omgitt av tre parastomatale celler, hvorav den ene er mye større (eller mindre) enn de andre;
3) paracytisk(parallell celle) - en peristomatal celle (eller flere) er plassert parallelt med vaktcellene;
4) diacytisk(krysscelle) - to parastomatale celler er plassert vinkelrett på vaktcellene;
5) tetrasitt(fra gresk tetra- fire) - hovedsakelig i monocots;
 Stomata er plassert på undersiden av bladet, men hos vannplanter med flytende blader finnes de kun på oversiden av bladet. Basert på formen på bladepidermiscellene og plassering av stomata kan en enfrøbladet plante skilles fra en tofrøbladet plante (fig. 2.8). Selve epidermalcellene i bladene til tofrøbladede planter er bølgete i omriss (fig. 2.9), mens de hos enfrøbladede planter er langstrakte, rombiske i form.
Stomata er plassert på undersiden av bladet, men hos vannplanter med flytende blader finnes de kun på oversiden av bladet. Basert på formen på bladepidermiscellene og plassering av stomata kan en enfrøbladet plante skilles fra en tofrøbladet plante (fig. 2.8). Selve epidermalcellene i bladene til tofrøbladede planter er bølgete i omriss (fig. 2.9), mens de hos enfrøbladede planter er langstrakte, rombiske i form.
Ris. 2.8. Plassering av stomata på epidermis (sett fra overflaten): EN- tofrøbladede planter: 1 - forbokstav; 2 - vannmelon; b- enkimblad: 3 - mais; 4 – iris
Stomatatypene kan deles inn i henhold til deres plasseringsnivå i forhold til overflaten av epidermis som følger.
1.7.1. Stomata ligger i samme plan som epidermis. Den vanligste typen og er vanligvis ikke angitt i beskrivelsen av mikroskopi av medisinske plantematerialer, dvs. denne paragrafen er utelatt. Diagnostiske tegn vil være enten utstående eller nedsenkede stomata.
1.7.2. Utstående stomata - stomata plassert over epidermis. Vanligvis, når mikroskopmikroskopet roteres (når linsen senkes), oppdages slike stomata først, og først da vises epidermale celler, så det er nesten umulig å fange dem på et fotografi fra overflaten av et blad, samt å skildre dem i en tegning. I samme plan som epidermis kan slike stomata sees i tverrsnitt, men for dette må snittet passere gjennom stomata, noe som er vanskelig å få tak i gitt deres sjeldne plassering på bladet. Slike stomata er for eksempel karakteristiske for bjørnebærblader.
1.7.3. Nedsenket stomata - stomata nedsenket i epidermis. Når de observeres under et mikroskop ved å rotere mikroskruen (mens linsen senkes), oppdages epidermiscellene først tydelig, deretter blir det mulig å tydeligere se konturene av stomata. Det er også vanskelig å vise dem i fotografier og tegninger av preparater fra overflaten. Finnes i liljekonvallblader, urblader, eukalyptusblader. Noen ganger er fordypningene der stomata er plassert foret eller dekket med hår og kalles stomatale krypter.
1.8. Typer stomatale celler
Det er 19 typer beskrevet i litteraturen vi har kun valgt ut de som brukes i analyse av medisinske planteråvarer**.
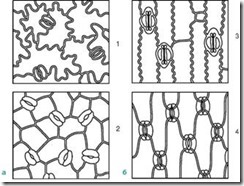
Ris. 63. Typer stomatale celler. A – lentiform; B - sfærisk; B - hetteformet; G – skafoid
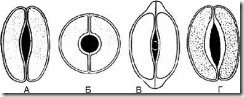
1.8.1. Linseformet - 2 identiske halvmåneformede celler arrangert symmetrisk. På frontplanet er fortykkelsen av skallet nesten jevn. Fissuren er fusiform (Fig. 63, A). Typen stomatale celler er karakteristisk for de fleste planter.
1.8.2. Sfæroidal - to identiske, sterkt sirkulært buede celler er plassert symmetrisk. På frontplanet er fortykkelsen av skallet nesten jevn. Sporet er rundt (fig. 63, B).
1.8.3. Cap-formet - to identiske halvmåneformede celler i de polare delene har fortykkelser i form av en hette. Fissuren er fusiform (fig. 63, B). Finnes i revehansker.
1.8.4. Skafoid - de indre veggene til stomatale celler er fortykket. Fissuren er fusiform (fig. 63, D). Observert i århundregress og urblader.
Mekanismen for stomataoperasjon bestemmes av de osmotiske egenskapene til cellene. Når bladoverflaten er opplyst av solen, skjer en aktiv fotosynteseprosess i vaktcellenes kloroplaster. Metning av celler med fotosyntetiske produkter og sukker innebærer aktiv inntreden av kaliumioner i cellene, som et resultat av at konsentrasjonen av cellesaft i vaktcellene øker. Det er en forskjell i konsentrasjonen av cellesaft i parastomatale og beskyttende celler. På grunn av de osmotiske egenskapene til cellene kommer vann fra de parastomatale cellene inn i vaktcellene, noe som fører til en økning i volumet til sistnevnte og en kraftig økning i turgor. Fortykning av de "abdominale" veggene til beskyttelsescellene som vender mot stomatalfissuren sikrer ujevn strekking av celleveggen; vaktcellene får en distinkt bønneformet form, og stomatalfissuren åpner seg. Når intensiteten av fotosyntesen avtar (for eksempel om kvelden), reduseres dannelsen av sukker i vaktceller. Tilstrømningen av kaliumioner stopper. Konsentrasjonen av cellesaft i vaktceller er redusert sammenlignet med parastomatale celler. Vann forlater beskyttelsescellene ved osmose, og senker deres turgor som et resultat, lukkes stomatalfissuren om natten.
Cellene i epidermis er tett lukket sammen, takket være at epidermis utfører en rekke funksjoner:
Forhindrer penetrasjon av patogene organismer inn i planten;
Beskytter indre vev mot mekanisk skade;
Regulerer gassutveksling og transpirasjon;
Vann og salter frigjøres gjennom det;
Kan fungere som sugevev;
tar del i syntesen av ulike stoffer, oppfatningen av irritasjoner og bevegelse av blader.
Trichomes - utvekster av epidermale celler med forskjellig form, struktur og funksjon: hår, skjell, bust, etc. De er delt inn i dekkende og kjertelformede. kjertel trikomer, i motsetning til coverts har de celler som skiller ut sekreter. Dekker hår danner et ull-, filt- eller annet dekke på planten, de reflekterer en del av solens stråler og reduserer dermed transpirasjonen. Noen ganger finner man hårene bare der stomata er plassert, for eksempel på undersiden av et følblad. Hos noen planter øker levende hår den totale fordampningsoverflaten, noe som bidrar til å akselerere transpirasjonen.
Trikomstørrelser varierer betydelig. De lengste trikomene (opptil 5-6 cm) dekker bomullsfrø. Dekkende trikomer har form av enkle enkelt- eller flercellede, forgrenede eller stjernehår. Dekker trikomer kan forbli i live i lang tid eller raskt dø og fylles med luft.
De skiller seg fra trikomer, som bare oppstår med deltakelse av epidermale celler. emergenter, i dannelsen av hvilke dypere lokaliserte vev i de subepidermale lagene også deltar.
Anatomiske og diagnostiske egenskaper som er av størst betydning og høy variabilitet ved bestemmelse av medisinske råvarer. Hår kan være enkle eller kapitate, som igjen kan være encellede eller flercellede. Flercellede hår kan være enkeltrad, dobbeltrad eller forgrenet.
1.9.1. Enkle hår.
A. Enkle encellede hår
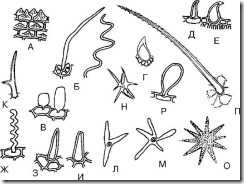
Ris. 68. Enkle encellede hår. A - papillær; B - filamentøs; B - vesikulær; G - spinous; D - krokformet; E – retortformet; F – stump trådlignende korrugert; Z - skarp kjegleformet; I – butt-kjegleformet; K - to-endede; L - trespiss; M, N, O – flerspiss; P - klumpete; P – kølleformet
1. Papillær (Fig. 68, A; Fig. 69-74) - lave, men brede utvekster av epidermale celler. Oftest dannet på kronbladene. De kan være stump-apikale eller akutt-apikale. De finnes på timianblader, liljekonvallblomster, tricolor fiolette blomster, langs kantene av århundreblader, i skjeggete ensiangress, etc.
2. Kjegleformet (Fig. 68, H, I; Fig. 75-79) - rette hår skråstilt i vinkel mot overflaten, som ender i en butt (stump kjegleformet) eller skarp (skarp kjegleformet) ende. De vanligste hårene. Vanligvis er de rette. Hår som skråner mot overflaten er observert på timianblader. Spisse kjegleformede hår finnes på sennablader, tyttebærblader, i trifarget fiolettgress (på blader), i timiangress osv. Stumpe kjegleformede hår finnes på epidermis til anisfrukter og på kronbladene til tricolor fiolette blomster .
3. Filiform (Fig. 68, B, G; Fig. 80, 81) – tynne og lange hår. Kan være rett eller korrugert. Rette trådlignende hår finnes på epidermis til hagtorn og bringebærfrukter. Bølgetrådaktig hår er observert på epidermis av fiolette kronblad (fig. 82).
4. Retortformet (Fig. 68, E; Fig. 83) - hår med en utvidet base og en smal buet eller rett ende. Finnes i humlefrukter og gress.
5. Blæreformet (Fig. 68,B; Fig. 84) – hår i form av små bobler. De finnes for eksempel i immortelle-blomster.
6. Krokformet (Fig. 68, E; Fig. 85-87) - hår, spisse på toppen og buet i form av en krok. Disse hårene kan bli funnet ved bunnen av bjørnebærblader, på overflaten av tyttebærblader, sennablader og anisfrukter.
7. Frynsete (Fig. 88, 89) - lange utvekster av epidermale celler plassert langs kanten av et blad, kronblad eller begerblad. Slike hår finnes for eksempel langs kantene av kronbladene til liljekonvall og fiolett.
8. Ryggformet (Fig. 68, D; Fig. 90) – hårene er nesten runde med en spiss ende. Tricolor fioler er observert i gresset.
9. Kølleformet (Fig. 68, P; Fig. 91) - hår med utvidet endeparti, som minner om en kølle. Finnes på timianblader og tricolor fiolette kronblader.
10. Dobbeltsidig (Fig. 68, K) - hår forgrenet i to ender. Observert i gresset av hyrdevesken.
11. Trespiss (flerspiss) (Fig. 68, L-O) - hår forgrenet i tre (eller flere) ender. Observert i gresset av hyrdevesken.
12. Klumpete (Fig. 68, P) - kjegleformede hår med fremspring (humper) på overflaten.
B. Enkle flercellede enrads hår
Konturen til et flercellet hår kan falle sammen med konturen til et encellet hår, men inneholde to eller flere celler og kalles følgelig et lignende encellet hår som indikerer antall celler, for eksempel et 2-cellet retortformet hår, en 11-cellers filamentøst hår, et kjegleformet hår med 10-15 celler.

Ris. 92. Enkle flercellede hår. A - kjegleformet; B - bobleformet; B - artikulær; G - piskformet; D - fjæraktig; E – T-formet; F - dobbel rad; Z - skjellete; Jeg stråler; L - parallell; M - gaffel; K – bustete
1. Kjegleformet (Fig. 92, A; 93-96). De mest forekommende hårene. Det er rett og tilbøyelig til overflaten, sistnevnte er sjeldne (for eksempel i timian urt). Kjegleformede hår kan være skarpe eller stumpe kjegleformede. Skarpe kjegleformede hår er mer vanlig (i urten av oregano, i bladene av mynte, salvie, etc.). Stumpe kjegler finnes i ringblomstblomster.
2. Retortformet - hår med en utvidet base og en smal buet eller rett ende.
3. Blæreformet (Fig. 92, B; Fig. 97) – hår i form av små bobler. Det er immortelle eggstokker på overflaten.
4. Filiform - tynne og lange flercellede hår.
5. Krokformet - hår spisse på toppen og buet i form av en krok.
6. Larver (Fig. 98, 99) - hår av nesten samme tykkelse i hele lengden, bestående av nesten identiske korte celler, med en butt ende og minner om en larve. De blir observert i gresset til tricolor fioler og i suksesjonsgresset.
7. Frynsete (Fig. 100) - lange flercellede utvekster av epidermale celler plassert langs kanten av et blad, kronblad eller begerblad. Sekvenser finnes i gresset.
8. Uhyggelig (Fig. 92, D; Fig. 101-104) - hår som har en flercellet base, bestående av en kjede av korte celler, og en lang trådformet kronglete terminalcelle. I litteraturen beskrives slike hår som pisklignende, snorlignende, trådaktige eller filtaktige. Hårtypen er karakteristisk for Asteraceae-familien. Finnes i ryllikgress, immortelle-blomster, coltsfoot blader, reinfann blomster, etc.
9. Artikulær (Fig. 92, B; Fig. 105, 106) - hår som har utvidede cellebaser ved artikulasjonsstedene (som ligner et ledd). Lignende hår finnes i morurtgress, på oreganoblader, og sjelden i timiangress.
10. Klumpete - hår som har fremspring (humper) på overflaten. Tocellede tuberøse fibre observeres for eksempel i løvet av thermopsis.
B. Enkle flercellede forgrenede hår
1. Parallell (Fig. 92, L; Fig. 107, 108) - hår, bestående av to lange rette celler (hår), smeltet sammen ved basene. Finnes i lindeblomster, sjelden i nyper og hagtorn.
2. Forked (Fig. 92, M; Fig. 109, 110) - hår, bestående av to lange viklingsceller (hår), smeltet sammen ved basene. Observert i lindblomster, svært sjelden i hagtornblomster og frukter.
3. Stjerneformet (Fig. 111) - hår, bestående av 3 eller flere lange kronglete celler (hår), smeltet sammen ved bunnene. Finnes i lindeblomster.
4. Skjellet (Fig. 92, H) - hår som består av en flercellet plate (i form av en rosett) og en kort stilk (stilken kan være fraværende). Slike hår finnes i havtorn. (Dette navnet på hårene er gitt i samsvar med moderne botanisk terminologi; ifølge andre kilder kalles disse hårstråene stjernehår eller skjellete).
5. T-formet (Fig. 92, E) er et tospisset hår med en- eller flercellet stilk. I litteraturen beskrives de også som hornformede, vippeformede.
6. Cirrus (Fig. 92, D) - flercellede hår, som minner om et forgrenet tre. Finnes for eksempel i mullein.
D. Enkle flercellede flerradshår
1. Dobbel rad (Fig. 92, G; Fig. 112, 113) – hår, hvis celler er ordnet i to rader. De kan observeres i calendula-blomster og tørket gress.
2. Bjelker (Fig. 92, I) - hår som består av to eller flere celler tett sammensmeltet med hverandre og danner en bunt.
3. bustete (Fig. 92 K, Fig. 114) - flercellede hår, bestående av hår av forskjellige lengder sammensmeltet med hverandre og med frie spisse ender. Slike hår er vanligvis hår fra flue (familie Asteraceae). I litteraturen kalles de også taggete komplekse, sylformede.
1.9.2. Kapitere hår. Hovedhår kalles også kjertelhår. De kan være encellede eller flercellede.

Ris. 115. Kapitere hår. A – med encellet stilk og encellet hode; B - med et to-rads ben og et enkeltcellehode; B - med et dobbeltradsben og et dobbeltradshode; G - med en encellet stilk og et flercellet hode; D - med en flercellet stilk og et encellet hode; F - med en flercellet stilk og et flercellet hode; Z – encellet (brennende); I – skjoldbruskkjertelen
A. Encellede hovedhår
Representanter for denne typen hår kan være kjegleformede eller retortformede, men har alltid et hode i enden. Slike hår finnes for eksempel på brennesleblader. B. Flercellede hovedhår (kjertelhår).
1. Hår med flercellet hode og encellet stilk (Fig. 115, D; Fig. 116-120). Finnes i morurtgress og bjørnebærblader.
2. Hår med encellet hode og encellet stilk (Fig. 115, A; Fig. 121-123). Finnes i morurtgress, salvieblader, hyllebærblomster, mynteblader.
3. Hår med encellet hode og flercellet stilk (Fig. 115, D; Fig. 124). Finnes i morurtgress, salvieblader, calendula blomster og kamille pedicels.
4. Hår med flercellet hode og flercellet stilk (Fig. 115, G; Fig. 125). Finnes i morurtgress, bjørnebærblader, lindblomster, calendulablomster, fiolettgress.
5. Hår med encellet hode og torads stilk (Fig. 115, B; Fig. 126). Finnes i blomster av calendula og immortelle.
6. Hår med torads hode og torads stilk (Fig. 115, B; Fig. 127-130). De finnes i blomstene av calendula og immortelle, og i gresset av myrgress.
7. Kølleformet flercellede kjertelhår (fig. 131-133; se fig. 155, E) - hår som har en kontur med en utvidet endedel, som minner om en kølle. De kan observeres for eksempel i tyttebærblader og fiolettgress.
8. Skjoldbruskkjertelen flercellede kjertelhår (Fig. 115, I; Fig. 134) er et skjold av polygonale tynnveggede celler som sitter på en en- eller tocellet kort stilk. Skjelaget på scutellum beveger seg bort fra cellene med den essensielle oljen utskilt under den. De finnes i humlefrukter.
1.10. Arten av fortykkelse av cellevegger og hår som dekker skjellaget
A. Karakter av fortykkelse av cellevegger
1.10.1. Tynnvegget (Fig. 135-137). De fleste hårene er tynnveggede. Celleveggene til flercellede lange tynnveggede hår kollapser noen ganger, og forstyrrer hårets jevne kontur. Slike hår, for eksempel, finnes på den nedre epidermis av kakali-bladet.
1.10.2. Tykkvegget (Fig. 138, 139). Finnes i nyper, strenggress, Dahurian lollipop gress, etc.
1.10.3. Ujevnt fortykket. Du kan se morurt i gresset (Fig. 140). Nypehår med skrå tverrporer tilhører også denne kategorien (fig. 141, 142).
B. Naturen til hårene som dekker neglebåndet
1.10.4. Glatt overflate (Fig. 143). Hår med en slik overflate observeres for eksempel i plantainblader, celandine gress, etc.
1.10.5. vorteaktig overflate (se fig. 1, G; fig. 144, 145) - epidermis danner fremspring i form av tuberkler (vorter). Det er hår for eksempel i fiolettgress, i timiangress, i morurtgress, i mynteblader, i sennablader osv. I dette tilfellet kan overflaten av håret være litt vorteaktig når det dannes små fremspring av skjellaget , og grovt vorteaktig når det dannes betydelige fremspring. I det første tilfellet kan hår i fiolett gress nevnes som et eksempel, i det andre - hår i sennablader og timiangress.
1.10.6. Grov overflate (se fig. 68, P) - neglebåndet danner svært store fremspring, for eksempel hårene på Thermopsis lanceolata-gresset.
10.7. Rynket overflate - folder eller bølger av hårkutikula fra bunnen til toppen (fig. 146). Det observeres oftere i papiller, for eksempel i centaury-gress og hyllebærblomster. Vanlige hår kan imidlertid også ha en rynket overflate, for eksempel i strenggress. I bunnen har hårene på myntebladene en rynket overflate (fig. 147).
1.10.8. Klekket overflate - epidermis danner korte fremspring i form av striper (tuberkler, vorter). Det er sjeldent og er et mellomalternativ mellom en vorteaktig og rynket overflate. Det observeres for eksempel på hårene til hylleblomster (fig. 148).
1.11. Funksjoner av hårfestesteder
1.11.1. Vanlige vedleggssteder (Fig. 149, 150) - hår festes til cellen eller mellom cellene i epidermis. Den vanligste typen finnes for eksempel i følblad, mynteblad, salvieblad, morurt, fiolett urt, etc.
1.11.2. Utvekster fra epidermale celler (se fig. 68, A; 69-74). Observert i hyllebærblomster, centaury gress, motherwort kronblader, skjeggete gentian gress, epidermis av anis frukter, etc.
1.11.3. Ved bunnen av håret dannes en rosett av epidermale celler (Fig. 151, 152). Finnes i sennablader, plantainblader, oreganourter, etc.
1.11.4. Forlenget hårbase (Fig. 153). Finnes i groblad.
1.11.5. Den utvidede delen av bunnen av håret er nedsenket i bunnen av subepidermalt vev - emergenter (se fig. 115, 3). Det kan observeres for eksempel i nesleblader.
1.11.6. Flercellet hårbase (Fig. 154). Du kan for eksempel observere strenger i gresset.
Ofte bryter hårene av og etterlater festepunktene på epidermis, som også må noteres som et anatomisk og diagnostisk tegn på medisinske plantematerialer.
SEKUNDÆR DEKKVEV
Det sekundære integumentære vevet kalles periderm. Dette er et komplekst dekkende vev av stilker, røtter og jordstengler til flerårige planter. Den erstatter epidermis i aksiale organer, som gradvis dør og slynges av. Peridermen er dannet av fellogen (sekundær meristem). Fellogen dannes i epidermis, subepidermalt lag, og til og med i de dype lagene av aksiale organer. Fellogenceller deler seg som følger: pluggceller legges utover, og levende parenkymceller av phelloderm legges innover. Stenglenes phellodermceller inneholder kloroplaster.
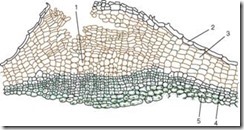 Pluggen består av døde celler hvis cellevegg er impregnert med et fettlignende stoff som kalles suberin. Cellene er arrangert i jevne rader, har en rektangulær form (på et tverrsnitt), og passer tett til hverandre, og danner et flerlagshus. Kork beskytter indre levende vev mot fuktighetstap, plutselige temperatursvingninger og penetrering av mikroorganismer. Levende vev som ligger under pluggen trenger gassutveksling og fjerning av overflødig fuktighet. Det er derfor, under stomata, på grunn av delingen av de subepidermale lagene (selv før utseendet av peridermen), og deretter de fenogene, levende, løst lokaliserte parenkymcellene med mange intercellulære rom, kalt utfører vevingny, som bryter epidermis og skaper mulighet for gassutveksling og transpirasjon med det ytre miljøet. Denne strukturelle formasjonen kalles linser(Fig. 2.12).
Pluggen består av døde celler hvis cellevegg er impregnert med et fettlignende stoff som kalles suberin. Cellene er arrangert i jevne rader, har en rektangulær form (på et tverrsnitt), og passer tett til hverandre, og danner et flerlagshus. Kork beskytter indre levende vev mot fuktighetstap, plutselige temperatursvingninger og penetrering av mikroorganismer. Levende vev som ligger under pluggen trenger gassutveksling og fjerning av overflødig fuktighet. Det er derfor, under stomata, på grunn av delingen av de subepidermale lagene (selv før utseendet av peridermen), og deretter de fenogene, levende, løst lokaliserte parenkymcellene med mange intercellulære rom, kalt utfører vevingny, som bryter epidermis og skaper mulighet for gassutveksling og transpirasjon med det ytre miljøet. Denne strukturelle formasjonen kalles linser(Fig. 2.12).
Ris. 2.12. Strukturen til peridermen med linsen: 1 – støttevevet til linsen; 2 - rester av epidermis; 3 - kork (phellema); 4 - fellogen; 5 - phelloderm
Linser, som ser ut som små tuberkler, er godt synlige på overflaten av skudd av trær og busker (fig. 2.13).
På bjørkestammer er restene deres observert i form av karakteristiske svarte tverrstriper på osp, de har form av diamanter.
Tertiær integumentært vev
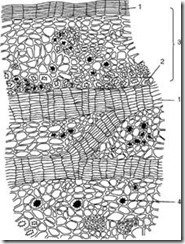 Skorpe (rhytide) er et tertiært integumentært vev som dannes i flerårige planter i roten, stengelen og rhizomet. Hvert år, i de dypere lagene, legges et nytt lag med fellogen og peridermen dannes. Det ytre laget av peridermen - korken - isolerer alt overliggende vev, som et resultat av at de dør. Dermed er samlingen av tallrike peridermer med dødt vev mellom seg skorpen (fig. 2.14).
Skorpe (rhytide) er et tertiært integumentært vev som dannes i flerårige planter i roten, stengelen og rhizomet. Hvert år, i de dypere lagene, legges et nytt lag med fellogen og peridermen dannes. Det ytre laget av peridermen - korken - isolerer alt overliggende vev, som et resultat av at de dør. Dermed er samlingen av tallrike peridermer med dødt vev mellom seg skorpen (fig. 2.14).
Ris. 2.14. Eikeskall: 1 – lag kork; 2 - fibre; 3 - rester av den primære skorpen; 4 – drusen av kalsiumoksalat
(meristematiske celler), og finnes i de delene av planter hvor vekst skjer.
1. Cytologiske trekk ved meristemer
Differensierte planteceller kan ikke dele seg i det hele tatt eller transformeres til andre typer celler. Derfor er celledeling i meristemer nødvendig for å gi nye celler for vekst av annet vev, dannelse av nye organer og strukturen til plantekroppen. Funksjonen til meristematiske celler ligner på dyrestamceller, som ikke differensierer eller differensierer litt, og er i stand til kontinuerlig celledeling. Meristematiske celler er små, og cytoplasma og kjerne fyller cellen fullstendig. Vakuolene er ekstremt små og cytoplasmaet inneholder ikke differensierte plastider (kloroplaster eller kromoplaster), selv om de er tilstede i vestigial form (protoplastider). Meristematiske celler er tettpakket, med nesten ingen intercellulær plass. Celleveggen er en veldig tynn primærcellevegg.
Den fysiologiske aktiviteten til meristemer krever en balanse mellom to antagonistiske prosesser: dannelse av nytt vev og støtte for fornyelse av populasjonen av meristematiske celler.
Meristem- danner plantevev, hvorfra alt annet vev kan dannes, meristematiske celler beholder i lang tid evnen til å dele seg med dannelsen av nye uspesialiserte celler.
Meristemets celler er flate, små, tett plassert ved siden av hverandre, uten intercliniforms, kjernen er plassert i midten av cellene, vakuolene er små, tallrike, cellemembranen er primær, det er bare plastidforløpere.
De cytologiske trekkene til kryssmeristemene kommer typisk til uttrykk i de apikale meristemene. Cellene er isodiametriske polygoner, ikke atskilt av intercellulære rom. Celleveggene er tynne, med lite celluloseinnhold. Cytoplasmaet er tett, kjernen er stor, plassert i sentrum. I cytoplasmaet er det et stort antall ribosomer og mitokondrier (intensiv syntese av proteiner og andre stoffer forekommer). Tallrike vakuoler er veldig små.
Cellene i de laterale meristemene er ulik i størrelse og form. Dette skyldes forskjellen i cellene i permanente vev som de er dannet fra. For eksempel, i kambium er det parenkymale og prosenkymale celler. Parenkymet til de ledende kompleksene er dannet fra de parenkymale initialene, og de ledende elementene selv er dannet fra de prosenkymale initialene.
Meristemer er klassifisert i henhold til deres posisjon i anlegget:
Og etter opprinnelse:
- hoved
- sekundær.
Primære merister er i nær forbindelse med det apikale meristem og direkte fra dem dannes spesielt protoderm, hovedmeristem og procambium. Protoderm er et enkeltlagslag av dårlig differensierte celler som dekker toppen fra utsiden, og epidermis dannes deretter fra den. Cortex er dannet fra hovedmeristem, og den sentrale sylinderen er dannet av procambium, hvis celler er forlenget langs stammens akse.
Sekundære merister penetrere fra spesialiserte celler av modne planteorganer og sikre deres sidevekst (phelogen, pericycle, cambium). Vev dannet fra det primære meristemet kalles primært, og de som dannes fra det sekundære meristemet kalles sekundært. Sekundære meristem bør også omfatte de såkalte traumatiske meristemene, som dannes på steder hvor plantekroppen er skadet og gir regenerering.
Blant de listede typene meristemer er den første som dukker opp i ontogenese det apikale meristemet, hvorfra differensiering av alt annet vev skjer. Den er plassert på toppen (APEX) av stilken og roten og deres sidegrener. Et trekk ved livsaktiviteten til en planteorganisme er spesielt bevaringen av formativ aktivitet i de apikale meristemene gjennom hele ontogenesen, derfor sier de at planter er i stand til ubegrenset vekst.
Struktur av det apikale meristemet (vekstpunkt)
1 - Sentral del
2 - Perifer del
2. Apikalt meristem
4. Interkalært meristem
Intercalary meristem (synonym - intercalary meristem) - celler er lokalisert mellom områder med differensiert vev, og gir intercalary vekst.
Den interkalære meristem, for eksempel, er lokalisert i nodene til korn, ved bunnen av petioles og stamen filamenter. Dette er gjenværende primærmeristemer. De stammer fra taumeristem, men deres transformasjon til permanent vev er forsinket sammenlignet med andre stammevev. Disse delikate meristemene er spesielt merkbare i frokostblandinger. Når det gjelder brød, sørger de for heving av stilkene på grunn av ujevn deling av celler fra under- og oversiden av halmen.
5. Sårmeristem
Dannes når vev og organer blir skadet. Levende celler som omgir de berørte områdene dedifferensierer og begynner å dele seg, det vil si at de blir til et sekundært meristem. Sårmeristemer dannes Kalus- tett vev av en blåaktig eller gulaktig farge, bestående av parenkymceller av forskjellige størrelser, arrangert på en uordnet måte. Callusceller har store kjerner og relativt tykke cellevegger. Callus kan gi opphav til ethvert plantevev eller organ. En plugg dannes i periferien; differensiering av callusceller til andre vev er mulig. Ytterligere røtter og knopper kan dannes i Kalus. Callus oppstår når planter podes og sikrer vekst av scion og rotstokk; ved bunnen av stiklingene. Calus brukes også for å oppnå isolerte vevskulturer.