VAKUOL SEL TUMBUHAN
Organoid maksud umum, mempunyai prinsip struktur membran tunggal dan berlaku sahaja dalam sel tumbuhan.
Saiz dan Kuantiti: ditentukan oleh umur sel. Dalam sel muda, vakuol timbul daripada vesikel kecil yang terputus daripada EPS. Apabila sel membesar dan membezakan, vakuol kecil bergabung antara satu sama lain dan membentuk satu atau lebih vakuol besar, menduduki sehingga 80% daripada isipadu keseluruhan sel. Akibatnya, sitoplasma dengan nukleus dan organel ditolak ke pinggir.
Struktur: vakuol pusat dipisahkan daripada sitoplasma oleh satu membran - tonoplast, yang serupa dengan ketebalan plasmalemma. Rongga vakuol terisi getah sel. Sap sel mengandungi: garam tak organik, gula, asid organik dan garamnya, bahan berat molekul rendah lain, serta beberapa sebatian berat molekul tinggi (contohnya, protein).
Fungsi: 1) osmoregulasi: disebabkan oleh separa kebolehtelapan tonoplast dan plasmalemma, kepekatan molekul sap sel yang sepadan dikekalkan, i.e. vakuol berfungsi sebagai osmometer;
2) perkumuhan: semua produk metabolik larut air (alkaloid - nikotin, kafein; polifenol) boleh dikeluarkan melalui tonoplast;
3) menyimpan: fosfat K + , Na + , Ca 2+ , garam terkumpul dalam sap sel asid organik(oksalat, sitrat, dsb.), gula dan protein.
RIBOSOM
Organel yang tidak mempunyai struktur membran. Ia adalah satu-satunya organel yang mempunyai kepentingan umum yang terdapat dalam sel kedua-dua prokariot dan eukariota. Ribosom pertama kali diterangkan dalam 1955 G. J. Palade (butiran palade), yang membuktikan bahawa ia adalah kompleks ribonukleoprotein (RNP).
Sifat kimia: RNP= r-RNA + protein.
Ribosom menyumbang 85% RNA, dibentangkan dalam sel.
Borang: Ribosom mempunyai bentuk cendawan kerana ia terdiri daripada dua subunit: besar Dan kecil, di antara mereka terletak pusat fungsi ribosom (FCR) , di mana, semasa biosintesis protein (tempoh terjemahan), mRNA terletak dalam dua tripletnya dan kompleks enzimatik beroperasi, memastikan pemasangan molekul protein daripada asid amino.
Dimensi: 15 – 35 nm. Saiz ribosom lengkap dalam sel prokariotik ialah 20x17x17 nm, dalam sel eukariotik - 25x20x20 nm.
Tempat pendidikan: pembentukan subunit ribosom berlaku dalam nukleolus nukleus. Penghimpunan subunit menjadi ribosom lengkap berlaku dalam sitoplasma apabila kepekatan ion magnesium (Mg 2+) mencapai 0.001 M jika kepekatan ini berkurangan, pemisahan subunit berlaku; Apabila kepekatan Mg 2+ meningkat sepuluh kali ganda, mencapai nilai 0.01 M, kedua-dua ribosom berinteraksi antara satu sama lain, membentuk dimer.
KULIAH: Ciri-ciri sel tumbuhan
Jumlah komposisi kimia adalah hampir sama.
Persamaan dengan sel haiwan: kehadiran membran, sitosol, nukleus, dan organel tertentu.
Walau bagaimanapun, antara jenis eukariota ini terdapat 2 perbezaan dalam struktur: kehadiran organel dan metabolisme tertentu.
Dalam sel tumbuhan ada vakuol pusat. Vakuol sel tumbuhan menyerap air yang berlebihan, membengkak, yang membawa kepada regangan sel, vakuol menduduki sebahagian besar isipadu sel tumbuhan, dan oleh itu hanya helai nipis sitoplasma kekal di dalam sel tumbuhan, melalui seluruh sel. Ini berfungsi sebagai cara ekonomi untuk sel tumbuhan untuk meningkatkan saiznya. Dalam kes ini, kurang tenaga digunakan daripada dalam kes mengisi volum yang sama dengan sitoplasma yang kaya dengan protein.
Sel tumbuhan muda mungkin mempunyai beberapa vakuol, yang, apabila sel membesar, bergabung antara satu sama lain dan membentuk satu atau beberapa vakuol besar, yang menduduki sehingga 90% daripada isipadu keseluruhan sel. Dalam kes ini, organel ditolak ke pinggir sel, dan nukleus juga. Membran vakuol ialah tonoplast. Rongga vakuol dipenuhi dengan sap sel, iaitu larutan akueus garam tak organik, gula, asid organik dan garamnya, protein, sebatian berat molekul rendah.
Fungsi utama vakuol pusat– mengekalkan tekanan turgor. Ia ditentukan oleh molekul yang terlarut dalam sap sel vakuol yang mengekalkan kepekatan osmotik. Kepekatan memastikan bahawa vakuol berfungsi sebagai struktur yang mengekalkan tekanan dalaman sel, memberikan kekuatan dan ketegangan. Tonoplast mempunyai pam proton yang bergantung kepada ATP yang memastikan pengangkutan gula, dan terdapat juga saluran ion yang menyediakan perkumuhan (penyingkiran) daripada vakuol pelbagai metabolisme. Kepekatan proton dalam sap sel sepadan dengan persekitaran berasid 2-5pH. Vakuol pusat memastikan pengumpulan rizab nutrien- gula dan protein. Bekalan bahan ini dikaitkan dengan keupayaan membran untuk berinteraksi dengan tonoplast. Proses pencernaan juga boleh berlaku di dalam vakuol sap sel, kerana mengandungi enzim hidrolitik, manakala tonoplast membentuk vesikel rembesan, mereka berpecah daripada vakuol, berinteraksi dengan vesikel eksocetotik, memastikan pencernaan komponen yang terkandung di dalamnya. Mereka juga menjalankan tindak balas kitaran autofatik, mengambil bahagian dalam hidrolisis pengisian yang rosak.
Dinding sel tumbuhan.
Dibentuk dengan penyertaan plasmalemma. Ia adalah pembentukan multilayer ekstraselular yang melindungi permukaan sel, pada asasnya adalah rangka luarannya. Ia terdiri daripada dua struktur: rangka kerja (diperbuat daripada benang fibril selulosa) dan matriks seperti gel. Matriks mengandungi polisakarida: hemiselulosa dan bahan pektin. Hemiselulosa ialah rantai polimer bercabang yang terdiri daripada pelbagai heksosa (glukosa, manosa, galaktosa), mungkin juga terdapat pentosa (xylose, arabinosa) dan asid uronik (glukuronik dan galakturonik). Komponen hemiselulosa ini digabungkan antara satu sama lain dalam nisbah kuantitatif yang berbeza dan membentuk pelbagai kombinasi. Tetapi rantai molekul hemiselulosa tidak pernah mengkristal, dan fibril mereka tidak tetap (tidak dikesan). Mengandungi jumlah air yang tinggi.
Pektin. Kumpulan heterogen yang termasuk bercabang, polimer terhidrogenasi tinggi yang membawa caj negatif disebabkan oleh kehadiran banyak sisa asid galakturonik.
Terima kasih kepada komponen ini, matriks adalah jisim plastik lembut yang berfungsi sebagai asas (bingkai).
Polimer selulosa individu dibungkus ke dalam mikrofibril menggunakan ikatan hidrogen. Memberi ketegaran dan kekuatan. Di antara mereka terdapat ligamen yang mengubah dinding sel menjadi monolit. Mikrofibril selulosa terikat sedemikian dikelilingi oleh pektin. Mereka boleh berinteraksi dengan kalsium dan silikon. Apa yang memberikannya ketegaran. Oleh kerana struktur seperti gel, matriks menyediakan penghantaran meresap air dan molekul kecil.
Dinding sel dibahagikan kepada 3 jenis: primer, sekunder, tertier.
Sel primer terdiri daripada 90% karbohidrat. Semasa pembahagian sel, sekumpulan mikrotubul muncul di satah khatulistiwa sel, terletak di antara kromosom yang menyimpang (phragmoplast). Di antara mikrotubulus terdapat banyak vesikel kecil sistem vakuolar, CG, di bahagian tengah phragmoplasts mula bergabung dengan satu sama lain. Membentuk cakera leper atau plat median. Terdiri daripada polisakarida yang dipanggil callose. Lebih elastik berbanding selulosa (tidak terkandung di dalam gelembung CG, terbentuk pada plasmalemma). Vesikel CG mengandungi komponen yang diperlukan untuk pembinaan membran dua sel anak. Proses percantuman vakuol kecil berlaku dari pusat sel ke pinggir dan berterusan sehingga vesikel membran bergabung dengan permukaan permukaan sisi sel.
Dinding sel primer yang semakin meningkat sudah terdiri daripada 3 lapisan (plat tengah, yang terdiri daripada matriks amorf, dan 2 pinggiran, mengandungi hemiselulosa dan fibril). Dinding primer dibentuk oleh rembesan fibril hemiselulosa dan selulosa oleh dua struktur selular baru. Meningkatkan ketebalan antara dinding sel akan berlaku disebabkan oleh aktiviti sel anak, yang masing-masing, untuk bahagiannya, akan merembeskan bahan membran sel, yang menebal dengan melapisi lebih banyak lapisan baru. Pada membran plasma, fibril selulosa disintesis dan dipolimerkan, mengakibatkan pembentukan dinding sel sekunder (memberi sel bentuk terakhirnya). Sebaik sahaja penebalan dinding selesai, ia diubah suai dengan lignin, polimer hidrofobik, disintesis melalui pempolimeran oksidatif sehingga tiga sisa alkohol aromatik dan berfungsi sebagai komponen utama kayu (20-35% kayu adalah lignin. ). Oleh kerana sifat hidrofobiknya, lignin mencipta kalis air di dinding dan berfungsi sebagai bahan pengukuhan tambahan. Dinding sel menjadi lignifikasi, atau suberized (disebabkan oleh kutin dan suberin). Dalam sel epidermis, lilin dirembes pada permukaan membran sel.
Kulat mempunyai kitin (N-acetylglucosamine).
Plastid.
Sifat umum semua plastid:
Mereka dikelilingi oleh dua membran yang rapat rapat di sepanjang seluruh permukaan organel.
Kandungan dalaman adalah stroma. Ia mengandungi cakera membran yang terbentuk akibat invaginasi dan tunas bahagian membran dalam. Cakera ini dipanggil tilakoid dan mempunyai keupayaan untuk melipat menjadi timbunan (granas). Mereka berinteraksi dengan jenis membran keempat - lamella.
Plastid mempunyai genomnya sendiri, setiap plastid mempunyai beberapa salinan DNA bulat yang mengandungi kira-kira 100 gen. Genom mengekod protein yang diperlukan untuk melaksanakan fungsi khusus.
Protein RNA yang terlibat dalam transkripsi dan terjemahan juga terdapat di sini. Mereka berasal dari plastid, tetapi kebanyakan protein lain bergantung pada genom nuklear. Protein ini disintesis dalam sitoplasma dan mesti diimport ke dalam plastid.
Plastid tidak disambungkan kepada organel lain melalui ikatan vesikular.
Plastid mewakili satu siri transformasi bersama.
Proplastid -> leucoplast -> kloroplas -> kromoplast.
Leukoplas boleh berubah menjadi: amiloplas, elaioplast, butiran aleuron dan kromoplast.
Kloroplas.
Mereka mempunyai bentuk yang memanjang, saiz 2-4 mikron, dan panjang boleh menjadi 10 mikron. Kromatofor (kloroplas besar).
Ruang dalaman kloroplas ialah lumen. 2 fenomena penting: fotosintesis, sintesis tenaga. Membran tilakoid dicirikan oleh kehadiran klorofil ia mampu menyerap kuanta cahaya matahari dan menukar tenaga suria kepada tenaga kimia. Penyerapan cahaya dengan panjang gelombang tertentu menyebabkan perubahan dalam klorofil. Dia masuk ke dalam keadaan teruja. Dan tenaga yang dikeluarkan dari klorofil diaktifkan dipindahkan melalui satu siri peringkat perantaraan ke rantai pengangkutan elektron, yang membawa kepada sintesis ATP dan pemulihan pembawa, iaitu 2 mononukleotida (NAD dan FAD). Tenaga dibelanjakan untuk sintesis karbon dioksida dan sintesis gula.
Fotosintesis: fasa terang dan gelap.
Fasa cahaya hanya berlaku dalam cahaya dan dikaitkan dengan penyerapan cahaya oleh pigmen dan kelakuan tindak balas kimia - tindak balas Hill.
Dalam fasa cahaya fotosintesis, tenaga cahaya matahari mengujakan elektron klorofil. Ia diangkut sepanjang komponen rantai oksidatif dalam membran tilakoid, sama seperti bagaimana elektron diangkut sepanjang rantai pernafasan dalam mitokondria. Dalam kloroplas, proton dipam dari stroma (pH = 8) ke lumen (pH = 5) menghasilkan kecerunan 3 unit pH. Kecerunan proton ini menghasilkan daya motif proton sebanyak 200 mW pada membran proton, tetapi ini hampir keseluruhannya disebabkan oleh kecerunan pH. Berbanding dengan rantai pernafasan mitokondria, elektron bergerak ke arah yang berbeza. Elektron diambil dari air semasa fotolisisnya dan dipindahkan ke pembawa dengan perbelanjaan tenaga. Itu. dalam fasa cahaya kita ada: molekul ATP yang disintesis dan pengangkut yang dikurangkan. Kedua-dua produk digunakan dalam peringkat gelap.
Pada peringkat gelap, karbohidrat terbentuk kerana pengurangan NAD dan ATP. Proses ini berbilang peringkat. Sebilangan besar enzim terlibat di dalamnya. Kitaran Calvin. Peningkatan bilangan kloroplas dan pembentukan bentuk plastid lain berlaku daripada struktur prekursor (proplastid).
Proplastid ialah vesikel dua membran kecil tanpa ciri tersendiri. Mereka berbeza daripada vakuol sitoplasma dalam kandungannya yang lebih padat dan kehadiran dua membran. Proplastid terletak dalam membahagikan tisu tumbuhan. Bilangan proplastid bertambah melalui pembahagian dan tunas. Di bawah pencahayaan biasa, proplastid bertukar menjadi kloroplas (peningkatan saiz, pembentukan lipatan membran yang terletak secara longitudinal berlaku. Beberapa membentuk lamellae, yang lain - tilakoid). Sekiranya mereka berkembang dalam gelap, maka pertama sekali terdapat peningkatan dalam jumlah plastid, tetapi sistem membran dalaman tidak terbentuk, jisim vesikel kecil terbentuk, yang terkumpul di zon berasingan, membentuk struktur kekisi kompleks - badan prolamellar; (etioplast). Membran mereka mengandungi protoklorofil (kuning di bawah pengaruh cahaya, kloroplas terbentuk daripadanya dan protoklorofil boleh berubah menjadi klorofil sebenar (membran, komponen rantaian pemindahan, disintesis).
Leukoplas. Berbeza dengan kloroplas. Mereka ditemui dalam sel tisu simpanan. Morfologi mereka yang tidak menentu menjadikan mereka sukar untuk dibezakan daripada proplastid. Mereka boleh dilihat sebagai lamella tetapi mampu membentuk struktur tilakoid yang normal. Dalam gelap, mereka mengumpul pelbagai bahan rizab. Jika ia adalah karbohidrat, maka leukoplas terbentuk menjadi amiloplast (mungkin juga terdapat protein dalam bentuk bijian aleuron). Dan jika lemak adalah elaioplasts.
Kami memerhatikan proses perubahan warna dan perubahan dalam kloroplas semasa perkembangan kelopak dan pematangan buah. Membran tambahan terkumpul dalam plastid, klorofil dan kanji dimusnahkan. Apabila lamellae dimusnahkan, titisan lipid dilepaskan, di mana pigmen (karotenoid) larut dengan baik dan perubahan warna muncul. Kromoplast adalah bentuk plastid yang merosot, yang dikaitkan dengan pemecahan kompleks lipoprotein.
Genom plastid.
Sistem genetiknya sendiri memastikan sintesis protein yang disetempat di dalam plastid itu sendiri (protein membran tilakoid). Mereka mempunyai panjang sehingga 60 mikron. 1.3*10 8 Ya. Tempoh kitaran dan kelajuan replikasi tidak sepadan. DNA tidak mempunyai struktur penetapan. Semua ciri molekul cincin kloroplas adalah hampir dengan ciri prokariot. Persamaan DNA kloroplas ditunjukkan dalam fakta bahawa proses pengawalseliaan asas transkripsi adalah sama. Sintesis semua jenis RNA. Kod DNA kloroplas untuk RNA ribosom. Pemalar segmentasi ribosom 70S. Ia mengandungi 17S dan 23S. Sensitif kepada antibiotik (chloramphenicol).
KULIAH: Hubungan antara sel
Teori simbiotik.
Komintsin dan Mereshkovsky.
Teori ini disokong oleh persamaan yang menakjubkan dalam struktur kloroplas dan alga biru-hijau. Persamaan dengan fungsi utama mereka, kebolehan yang hampir sama untuk proses fotosintesis. Hasil daripada simbiosis pelbagai sel prokariotik, sel nuklear (eukariotik) muncul. Menurut hipotesis ini, sel eukariotik dalam perkembangannya melalui beberapa peringkat simbiosis dengan sel lain.
Pada peringkat pertama, sel-sel seperti prokariot heterotropik anaerobik menjadi perumah bagi bakteria aerobik, yang membawa kepada pembentukan prokariot aerobik (mitokondria, dalam organisma prokariotik ini). Selari dengan proses ini, dalam sel perumah, radas nuklear prokariotik (genofor) terbentuk menjadi nukleus yang diasingkan daripada sitoplasma. Ini adalah bagaimana sel eukariotik heterotropik boleh timbul. Sel sedemikian memasuki simbiosis berulang. Contohnya, dengan spirochetes atau bakteria seperti flagella. Kemasukan simbion ini membawa kepada kemunculan flagella dan silia dalam sel eukariotik heterotropik. Dan sebagai hasil daripada penyerapan tambahan. Ini membawa kepada kemunculan sel haiwan eukariotik dengan flagela. Organisma sedemikian juga boleh menyerap alga biru-hijau dan ini membawa kepada pembentukan plastid dan pembentukan sel tumbuhan. Hipotesis asal endosimbiotik organel intramembran tidak diragukan lagi. Banyak fakta endosimbiosis sebenar alga biru-hijau dan sel diketahui tumbuhan yang lebih rendah dan protozoa, di mana simbion berfungsi dengan sempurna dan membekalkan sel perumah dengan produk fotosintesis. Contohnya. Ciliate selipar, ia mengandungi chlorella alga. Kloroplas boleh dipilih oleh beberapa sel dan digunakan sebagai endosimbion. Rotifer memakan alga. Kloroplas mendapati diri mereka berada di dalam sel-sel kelenjar pencernaan dan terus melaksanakan fungsinya.
Ciri-ciri fungsi sel tumbuhan. Kaedah sintesis bahan organik adalah pemakanan autotrof.
Sel tumbuhan, kerana kehadiran dinding sel, tidak membenarkan sel tumbuhan berubah bentuk dan bergerak. Dan sel tumbuhan mengimbangi kekurangan pergerakan dengan mensintesis makanan mereka sendiri, i.e. adalah autotrof. Tumbuhan tumbuh sepanjang hayat mereka, meristem sentiasa membahagi dan membentuk sel baru; semua tumbuhan tergolong dalam organisma modular. Bentuk kehidupan terakhir mereka bergantung pada persekitaran mereka. Sel tumbuhan juga mempunyai ciri-ciri dalam perjalanan mitosis:
Kedudukan dinding sel baru. Ia ditentukan lama sebelum pembentukan gelendong, manakala gelendong itu sendiri tidak mengambil bahagian dalam lokasi sel ini. Malah sebelum mitosis, dalam fasa G 2 kitaran sel, struktur tertentu terbentuk dalam sel tumbuhan, yang dipanggil cincin preprophase. Ia terbentuk daripada mikrotubulus korteks (sitokeleton) dalam bentuk jalur lebar yang mengelilingi sel. Nukleus kekal utuh (pepejal). Di kawasan gelang ini, filamen aktin berkumpul, yang memastikan penyempitannya sehingga ia bertukar menjadi berkas mikrotubul padat yang terbentuk dengan baik di sekeliling nukleus. Struktur yang terhasil menyerupai roda, di mana rim dan jejari terdiri daripada mikrotubul dan aktin, dan teras membentuk hab. Struktur cincin diperkaya dengan elemen EPR dan KG. Pada peringkat awal mitosis, mikrotubul cincin praprofasa didepolimerisasi dan dimusnahkan. Dan dalam telofase, dinding sel baru terbentuk tepat pada kapal terbang itu
, yang telah ditetapkan oleh cincin praprofasa.
Dalam tumbuhan, kutub gelendong mitosis tidak mengandungi sentriol dan bersifat lebih meresap. Dalam erti kata lain, mereka kelihatan kabur. Pada mulanya, gelendong prophase terbentuk daripada mikrotubul. Daripadanya, selepas pemusnahan membran nuklear, gelendong sebenar terbentuk, dan dengan pembentukan gelendong mitosis sebenar ini, tiada mikrotubul sitoplasma lain kekal di dalam sel. Apa yang mengawal gelendong ini, apa yang terbentuk? Kromosom.
Pertama, pempolimeran mikrotubul berlaku berhampiran kromosom, yang berorientasikan secara rawak. Kemudian protein motor (dynein) menyusun mikrotubul, hujung positifnya dihantar ke kromosom, yang membolehkan mereka melekat pada kinetokor kromosom, dan yang lain berinteraksi dengan hujung negatif mikrotubul. Hasil daripada pempolimeran, pengisihan, dan lampiran, gelendong bipolar terbentuk. Selepas pembahagian nuklear, pembahagian sitoplasma berlaku. Ini bertepatan dengan saat pembentukan septum sel baru, kedudukannya ditentukan oleh cincin preprophase. Hubungan antara orientasi gelendong dan septum baru tidak ketat, seperti dalam sel haiwan.
Kompleks Golgi sel tumbuhan. Fungsi: glikosilasi (utama), pengasingan (pemisahan), sintesis polisakarida dinding sel (kecuali selulosa dan callose), penghantaran kompleks enzim (roset heksagon, sintesis selulosa) ke membran, mengarah pengangkutan vesikel ke dalam vakuol.
KG disetempatkan dalam sel tumbuhan lebih dekat dengan plasma, lebih jauh dari nukleus, dan tersebar dalam bentuk banyak timbunan tangki. Semua elemen kompleks disambungkan oleh filamen aktin, yang membentuk rangkaian yang memastikan pengangkutan intraselular. Struktur aktin menyediakan mobiliti (bersama-sama dengan myosin). Sel tumbuhan mempunyai miosin khas - myosin 8. Miosin 8 ini tidak membentuk filamen dan terkumpul dalam dinding primer yang baru terbentuk, vesikel CG mengandungi polisakarida dan protein struktur yang membentuk dinding sel sekunder.
Hubungan antara sel.
Terdapat beberapa klasifikasi mereka, mereka berstruktur secara berbeza. Klasifikasi Alberts dan Gray (1986). Menurutnya: kenalan boleh dibahagikan kepada 3 kumpulan.
Mekanikal (= Pelekat). Dikaitkan dengan keupayaan sel untuk melekat antara satu sama lain, kerana kehadiran glikoprotein matriks ekstraselular dalam membran sel. Apabila lekatan berlaku di antara membran plasma, sentiasa ada jurang (20 nm) yang dipenuhi dengan glikokaliks. Protein CAM khas (cam) bertanggungjawab secara langsung untuk menghubungkan sel antara satu sama lain: fibronektin, kadherin, selektin, integrin dan protein lain. Sebahagian daripada mereka menyambung sel antara satu sama lain melalui interaksi antara molekul. Yang lain membentuk sambungan antara sel khas. Interaksi antara protein ini mungkin homofilik, i.e. sel jiran dalam kes ini berkomunikasi antara satu sama lain menggunakan molekul homogen. Atau heterofilik, apabila mereka mengambil bahagian dalam lekatan pelbagai jenis protein pada sel jiran. Ada lagi sambungan yang kompleks, apabila pengikatan antara sel dipastikan oleh molekul perantara penghubung.
Di samping itu
Desmosomes ikat pinggang. Jarak antara sel jiran ialah 15-20 nm. Plat tengah tidak begitu terang. Tetapi terdapat glikoprotein trans-membran yang melekat antara satu sama lain dan menyediakan sambungan mekanikal membran. Filamen aktin nipis (6-7 nm) terkumpul di bahagian sitoplasma. Mereka terletak di sepanjang plasmalemma dan berjalan di sepanjang perimeter sel. Fungsi: lekatan mekanikal sel antara satu sama lain; apabila filamen aktin mengecut, bentuk sel boleh berubah; boleh menyebabkan perubahan dalam geometri sel; menghantar tekanan mekanikal ke sel. Disebabkan oleh perubahan isipadu sel.
Hemidesmosom. Sambungan sel dengan matriks ekstraselular (Contohnya, dengan membran bawah tanah). Peranan berfungsi adalah mekanikal, yang membolehkan lapisan epitelium menahan kesan mekanikal yang besar. Desmosom adalah bahagian sel, tetapi ia bukan pembentukan statik. Mereka dinamik. Mampu menghilang dan muncul semula. Mereka dimusnahkan oleh tindak balas phagocentral.
Penutup (sentuhan ketat). Mereka adalah ciri epithelia satu lapisan. Zon di mana lapisan luar dua membran plasma adalah sedekat mungkin. Oleh itu, apabila dilihat melalui mikroskop elektron, struktur tiga lapisan kelihatan. 2 lapisan luar kedua-dua membran bergabung menjadi satu tebal 2-3 nm. Bukan di seluruh kawasan sentuhan ketat, tetapi mewakili satu siri titik hubungan protein membran integral. Lebih daripada 24 jenis protein ditemui di zon persimpangan ketat. Menonjol dari membran lapisan bilipid, protein tersebut bersilang dan membentuk rangkaian (kekisi). Di sisi persimpangan ketat, dalam sitoplasma itu sendiri terdapat banyak fibril (7 nm), yang terletak selari dengan permukaan plasmalemma. Persimpangan ketat adalah ciri epitelium kelenjar dan usus. Mereka menghubungkan sel antara satu sama lain. Lebih-lebih lagi, ini struktur molekul mengawal selia pengangkutan paraselular, i.e. pengangkutan bahan dalam ruang antara sel.
Halangan kebolehtelapan dalam sentuhan ketat dicipta oleh lapisan pembawa cas yang membentuk struktur rangkaian unsur seperti benang fibril. Ion boleh melalui liang sedemikian saiznya kecil. Walau bagaimanapun, untuk laluan komponen larut, integriti benang ini mesti terganggu. Unsur larut memecahkan benang ini, kemudian mereka bersatu semula selepas laluan molekul. Oleh itu, molekul bergerak melalui penghalang sentuhan. Dari segi struktur dan fungsi, persimpangan ketat membahagikan plasmalemma kepada 2 bahagian (apikal dan basal) dan persimpangan ketat bertindak sebagai septum, yang menyebabkan komposisi molekul dikekalkan dalam had.
Hubungan konduktif. Ia berbeza dalam haiwan dan tumbuhan.
Terdapat 2 jenis sel haiwan: gap dan sintesis kimia. Melalui gap junctions, molekul boleh bergerak dari satu sel ke sel yang lain. Tetapi dalam sintesis kimia, sel tidak mempunyai sambungan langsung. Persimpangan jurang dianggap sebagai persimpangan komunikasi sel. Ini adalah struktur yang terlibat dalam pemindahan langsung bahan kimia. Sentuhan ciri jenis ini ialah penyatuan membran plasma dua sel jiran pada jarak 2-3 nm. Sentuhan jurang itu sendiri mempunyai saiz 0.5-5 µm. Ia terdiri daripada 2 bahagian, saiz setiap separuh adalah 7-8 nm, ia terletak di sekitar saluran 1-2 nm lebar. Dan 2 bahagian ini dipanggil connexons (separuh saluran). Mereka terdiri daripada 6 subunit protein connectin. Berat molekul protein adalah kira-kira 30 ribu. Dengan bergabung antara satu sama lain, protein membentuk struktur silinder dan dalam dua sel saiz, bilangan, saiz, dan susunan sambungan adalah simetri yang ketat. Sentuhan jurang membenarkan resapan bebas molekul seberat 1200 Dalton, tidak termasuk molekul yang jisimnya mencapai 2000 Dalton. Kebolehtelapan simpang jurang dikawal oleh pembukaan dan penutupan pintu saluran. Proses ini dipanggil gerbang (mekanisme pintu). Ia dikawal oleh perubahan pH intraselular, fluks ion kalsium, atau fosforilasi langsung subunit protein connexin. Fungsi sesentuh jurang: menjalankan organik dan bahan bukan organik. (Sebagai contoh, dalam sel otot miokardium).
Plasmodesmata ialah sejenis sesentuh pengalir dalam tumbuhan. Ini adalah saluran sitoplasma tiub nipis yang menghubungkan 2 sel jiran. Diameter saluran ini berbeza dari 20 hingga 40 nm. Ciri-ciri saluran ini: membran plasma daripada satu sel masuk ke plasmalemma sel lain. Di sepanjang paksi saluran ini, tiub silinder desmotule terbentang dari satu sel ke sel yang lain. Lumennya berkomunikasi dengan sel ER. Ruang antara desmotubule dan membran plasmodesmata dipenuhi dengan sitosol. Plasmalemma yang bersempadan dengan sel-sel ini dengan lancar masuk ke dalam plasmodesmata dan membran sel lain. Akibatnya, hyaloplasma sel jiran disambungkan ke dalam satu sistem (symplast). Dan secara rasmi tidak ada pemisahan (satu sel dari yang lain). Pengangkutan percuma melalui plasmodesmata terhad kepada berat molekul 800 Dalton. Plasmodesmata terbentuk pada peringkat pembentukan utama plat median daripada unsur-unsur ER. Dalam sel yang baru dibahagikan, bilangan plasmodesmata mencapai sehingga 1000 setiap sel. Dengan penuaan, bilangan mereka berkurangan, tetapi ketebalannya meningkat. Peranan berfungsi: memastikan peredaran antara sel larutan (ion, gula, nutrien). Sisi negatifnya ialah lebih mudah jatuh sakit.
Vakuol dan getah sel
Kebanyakan sel tumbuhan matang dicirikan oleh vakuol pusat yang besar, menduduki sehingga 70-90% daripada jumlah sel. Dalam kes ini, protoplas dengan semua organel terletak dalam bentuk lapisan dinding yang sangat nipis yang melapisi dinding sel. Vakuol sitoplasma kecil biasanya terdapat dalam protoplas dinding. Kadangkala nukleus terletak di tengah-tengah sel dalam poket nuklear sitoplasma, yang disambungkan ke lapisan dinding oleh helai sitoplasma paling nipis yang melintasi vakuol pusat.
Getah sel ialah larutan akueus pelbagai bahan, yang merupakan produk daripada aktiviti penting protoplas, terutamanya bahan simpanan dan sisa. Tindak balas sap sel biasanya sedikit berasid atau neutral, kurang kerap alkali. Bahan-bahan yang membentuk sap sel sangat pelbagai. Ini adalah karbohidrat, protein, asid organik dan garamnya, asid amino, ion mineral, alkaloid, glikosida, tanin, pigmen dan sebatian larut air yang lain. Kebanyakannya tergolong dalam kumpulan bahan ergastik - produk metabolisme protoplas, yang boleh muncul dan hilang tempoh yang berbeza hayat sel. Banyak bahan sap sel hanya terbentuk dalam sel tumbuhan.
Karbohidrat dalam sap sel tumbuhan diwakili oleh monosakarida (glukosa, fruktosa), disakarida (sukrosa) dan polisakarida (lendir, inulin).
Glukosa (gula anggur) dan fruktosa (gula buah) terkumpul dalam kuantiti yang banyak dalam buah-buahan berair. Sukrosa (gula bit) terkumpul dalam kuantiti yang banyak dalam akar bit gula dan batang tebu. Sebilangan keluarga tumbuhan (cactaceae, Crassulaceae, orkid) dicirikan oleh pengumpulan lendir dalam sap sel, yang mengekalkan air. Inulin ialah polisakarida simpanan, disimpan sebagai larutan koloid dalam sap sel organ bawah tanah Asteraceae dan bukannya kanji.
Protein terkumpul dalam bentuk larutan koloid dalam vakuol sel benih yang masak. Apabila benih mengalami dehidrasi pada peringkat akhir perkembangannya, air dikeluarkan dari vakuol, kepekatan protein dalam sap sel meningkat, dan ia berubah menjadi keadaan gel pepejal. Vakuol dehidrasi benih matang dipanggil bijirin aleuron.
Daripada asid organik dalam sap sel, yang paling biasa ialah sitrik, malik, suksinik dan oksalik. Asid ini terdapat dalam kuantiti yang banyak dalam sap sel buah-buahan yang belum masak, memberikan mereka rasa masam. Apabila buah masak, asid organik boleh digunakan sebagai substrat respirasi, jadi rasa masam buah biasanya hilang. Garam asid organik, bersama-sama dengan ion mineral, memainkan peranan penting dalam proses osmotik.
Tannin (tannin) adalah sebatian fenolik polimer dengan rasa astringen. Mereka mempunyai sifat antiseptik dan melindungi tisu tumbuhan daripada jangkitan dan pereputan. Sel-sel kulit batang dan akar (oak, willow), buah-buahan yang belum masak (walnut), daun (teh) dan beberapa pertumbuhan patologi - hempedu terutamanya kaya dengan tanin. Tannid digunakan dalam perubatan, untuk menyamak kulit, dan mewarnakan kain coklat gelap.
Alkaloid – pelbagai dalam secara kimia bahan organik yang mengandungi nitrogen yang mempunyai rasa pahit. Mereka mempunyai sifat asas dan terdapat dalam sap sel, biasanya dalam bentuk garam. Banyak tumbuhan yang mengandungi alkaloid adalah beracun dan tidak dimakan oleh herbivor. Dalam sel yang mengandungi alkaloid, spora dan kuman mikroorganisma tidak berkembang, dan tumbuhan tidak terjejas oleh penyakit kulat dan bakteria. Wakil-wakil keluarga Solanaceae, Poppy, Rubiaceae, Ranunculaceae, dan lain-lain amat kaya dengan alkaloid.
Glikosida - kumpulan yang luas bahan semulajadi, sebatian gula dengan alkohol, aldehid, fenol dan bahan lain. Sebilangan glikosida tumbuhan digunakan dalam perubatan. Glikosida juga termasuk pigmen sap sel - flavonoid. Sebahagian daripada mereka adalah antosianin– beri sap sel merah, biru atau ungu; lain - flavon- kuning. Antosianin dikaitkan dengan warna bunga banyak tumbuhan. Skema warna ditentukan oleh tindak balas sap sel: jika ia berasid, maka ton merah mendominasi, jika neutral, ton ungu mendominasi, dan dengan tindak balas yang sedikit beralkali, ton biru mendominasi. Penampilan warna juga dipengaruhi oleh pembentukan kompleks antosianin dengan pelbagai logam. Flavon menentukan kuning kelopak sebilangan tumbuhan.
Kepentingan asid organik, tanin, alkaloid dan glikosida sap sel dalam metabolisme sel belum cukup dijelaskan. Sebelum ini, mereka dianggap sebagai produk akhir pertukaran. Kini telah ditunjukkan bahawa banyak daripada mereka boleh terlibat semula dalam proses metabolik dan oleh itu boleh dianggap sebagai bahan simpanan.
Sebagai tambahan kepada fungsi mengumpul bahan rizab dan sisa, vakuol dalam sel tumbuhan melakukan yang lain fungsi penting- mengekalkan turgor. Kepekatan ion dan gula dalam sap sel vakuol pusat biasanya lebih tinggi daripada di dinding sel; Tonoplast dengan ketara memperlahankan resapan bahan-bahan ini dari vakuol dan pada masa yang sama mudah telap ke air. Oleh itu, air akan memasuki vakuol. Proses satu arah resapan air melalui membran telap terpilih ini dipanggil osmosis. Air yang memasuki sap sel memberikan tekanan pada protoplas dinding, dan melaluinya pada dinding sel, menyebabkan keadaan tegang, keanjalan, atau turgor sel. Turgor memastikan bahawa organ tumbuhan tidak berlign mengekalkan bentuk dan kedudukannya di angkasa, serta ketahanannya terhadap faktor mekanikal.
Jika sel diletakkan dalam larutan hipertonik beberapa garam atau gula bukan toksik (iaitu, dalam larutan kepekatan yang lebih tinggi daripada kepekatan sap sel), maka pembebasan osmotik air daripada vakuol berlaku. Akibatnya, isipadunya berkurangan, protoplas dinding elastik bergerak menjauhi dinding sel, turgor hilang, dan plasmolisis sel( Rajah 2.9).
nasi. 2.9. Skim plasmolisis: 1 – sel dalam keadaan turgor; 2 - permulaan plasmolisis; 3 – plasmolisis lengkap.
Plasmolisis biasanya boleh diterbalikkan. Apabila sel diletakkan di dalam air atau larutan hipotonik, air sekali lagi diserap dengan kuat oleh vakuol pusat, protoplas sekali lagi ditekan pada dinding sel, dan turgor dipulihkan. Plasmolisis boleh berfungsi sebagai penunjuk keadaan hidup sel; sel mati tidak diplasmolisis, kerana ia tidak mempunyai membran telap terpilih.
Kehilangan turgor menyebabkan tumbuhan menjadi layu. Apabila layu di udara dalam keadaan bekalan air yang tidak mencukupi, dinding sel nipis mengecut serentak dengan protoplas dan menjadi terlipat.
Tekanan turgor bukan sahaja mengekalkan bentuk bahagian tumbuhan yang tidak berlign, ia juga merupakan salah satu faktor pertumbuhan sel, memberikan ketinggian sel regangan, iaitu disebabkan oleh penyerapan air dan peningkatan saiz vakuol. Sel haiwan tidak mempunyai vakuol pusat; pertumbuhannya berlaku terutamanya disebabkan oleh peningkatan jumlah sitoplasma, oleh itu saiz sel haiwan biasanya lebih kecil daripada sel tumbuhan.
Vakuol pusat timbul daripada gabungan banyak vakuol kecil yang terdapat dalam sel meristematik (embrionik). Vakuol sitoplasma ini dianggap terbentuk oleh membran retikulum endoplasma atau radas Golgi.
Kemasukan
Pembentukan kemasukan disebabkan oleh pengumpulan berlebihan produk metabolik tertentu di bahagian tertentu sel - dalam vakuol, hyaloplasma, pelbagai organel, dan kurang kerap di dinding sel. Bahan-bahan ini sering memendakan dalam bentuk amorf atau kristal - kemasukan. Kemasukan mempunyai bentuk tertentu dan boleh dilihat dengan jelas di bawah mikroskop cahaya. Dengan kehadiran kemasukan tertentu, bentuk dan pengedarannya, satu spesies, genera dan keluarga tumbuhan boleh dibezakan daripada yang lain, oleh itu mereka sering berfungsi sebagai penting. tanda diagnostik semasa menganalisis bahan tumbuhan ubatan.
Kemasukan adalah sama ada bahan ganti(sebatian dikeluarkan sementara daripada metabolisme), atau produk akhir pertukaran. Kategori kemasukan pertama termasuk bijirin kanji,penurunan lipid Dan mendapan protein; ke yang kedua - kristal beberapa bahan.
Bijirin kanji– kemasukan sel tumbuhan yang paling biasa. Polisakarida kanji– jenis nutrien simpanan utama untuk tumbuhan. Dia juga yang paling banyak sambungan penting digunakan sebagai makanan oleh herbivor. Pati daripada bijirin bijirin, ubi kentang, dan buah pisang adalah sumber pemakanan manusia yang paling penting. Tepung gandum mengandungi hampir 75% bijirin kanji dalam ubi kentang, kanji membentuk 20-30%. Secara kimia, kanji adalah alpha-1,4-D-glukan, molekulnya mempunyai bentuk rantai bercabang, dalam butiran kanji mereka terletak di sepanjang jejari.
Bijirin kanji terbentuk dalam stroma plastid. Dalam kloroplas, bijirin dimendapkan dalam cahaya kanji asimilasi (utama)., terbentuk apabila terdapat lebihan gula - hasil fotosintesis. Pembentukan kanji tidak aktif secara osmotik menghalang peningkatan tekanan osmotik dalam kloroplas. Pada waktu malam, apabila fotosintesis tidak berlaku, kanji asimilasi dihidrolisiskan kepada gula dengan bantuan enzim dan diangkut ke bahagian lain tumbuhan. Kanji ganti (sekunder). disimpan dalam amiloplas sel pelbagai organ tumbuhan (akar, pucuk bawah tanah, biji) daripada gula yang mengalir daripada sel fotosintesis. Jika perlu, kanji simpanan juga ditukar kepada gula.
Pembentukan butiran kanji bermula pada titik tertentu dalam stroma plastid, dipanggil pusat-pusat pendidikan. Pertumbuhan bijirin berlaku melalui pemendapan berturut-turut lapisan kanji di sekeliling pusat pendidikan. Lapisan bersebelahan dalam satu butir mungkin ada penunjuk berbeza pembiasan cahaya, dan kemudian ia boleh dilihat di bawah mikroskop - berlapis-lapis bijirin kanji. Susunan lapisan boleh sepusat(gandum) atau sipi(kentang) ( nasi. 2.10). Sekiranya terdapat satu pusat pendidikan dalam amiloplast, di sekeliling lapisan kanji didepositkan, maka ringkas, jika dua atau lebih, maka ia terbentuk bijirin kompleks, terdiri daripada beberapa yang mudah. bijirin separa kompleks terbentuk apabila kanji mula-mula didepositkan di sekitar beberapa titik, dan kemudian, selepas sentuhan butiran mudah, lapisan biasa muncul di sekelilingnya ( nasi. 2.10).
Bentuk, saiz, nombor dalam amiloplast dan struktur (kedudukan pusat pendidikan, lapisan, kehadiran atau ketiadaan retak) bijirin kanji selalunya khusus untuk spesies tumbuhan ( nasi. 2.10). Biasanya, bijian kanji adalah sfera, ovoid atau berbentuk kanta, tetapi dalam kentang ia tidak teratur. Bijirin terbesar (sehingga 100 mikron) adalah ciri sel ubi kentang; dalam bijirin gandum ia mempunyai dua saiz - kecil (2-9 mikron) dan lebih besar (30-45 mikron). Sel bijirin jagung dicirikan oleh bijirin kecil (5-30 mikron). Beras, oat, dan soba mempunyai bijirin kanji yang kompleks.
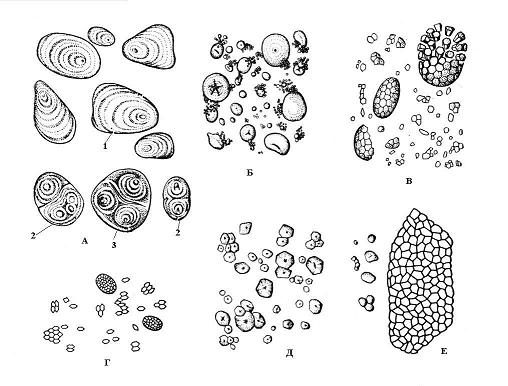
nasi. 2.10. Bijirin kanji pelbagai jenis tumbuhan: A – kentang; B – gandum; B - oat; G – beras; D – jagung; E – soba; 1 – bijirin mudah; 2 - bijirin kompleks; 3 – bijirin separa kompaun.
Reagen untuk kanji ialah larutan iodin dalam larutan kalium iodida - Reagen Lugol. Ia mewarnakan bijirin kanji biru-ungu.
Mendapan kanji tersebar luas di semua organ tumbuhan, tetapi benih, pucuk bawah tanah (ubi, mentol, rizom), dan parenkim tisu konduktor akar dan batang tumbuhan berkayu sangat kaya dengannya.
Lipid jatuh terdapat dalam hampir semua sel tumbuhan. Minyak lemak terkumpul dalam sebilangan besar tumbuhan dan merupakan bentuk rizab nutrien kedua terpenting selepas kanji. Biji dan buah-buahan sangat kaya dengannya. Benih sesetengah tumbuhan (bunga matahari, kapas, kacang tanah) boleh mengandungi sehingga 40% minyak mengikut berat bahan kering. Oleh itu, lemak sayuran diperoleh terutamanya daripada biji benih.
Titisan lipid terkumpul terus dalam hyaloplasma. Mereka kelihatan seperti badan sfera kecil, setiap titisan dipisahkan dari hyaloplasma oleh membran. Kadang-kadang titisan lipid dipanggil sferosom.
Reagen untuk minyak lemak ialah pewarna SudanIII, titisan lipid berwarna jingga-merah.
Kemasukan protein dalam bentuk pelbagai mendapan amorf atau kristal terbentuk dalam pelbagai organel sel. Selalunya, kristal protein boleh didapati dalam nukleus, kurang kerap - dalam hyaloplasma, stroma plastid, dalam lanjutan tangki retikulum endoplasma, matriks peroksisom dan mitokondria. Saiz hablur protein selalunya melebihi resolusi mikroskop cahaya.
Protein simpanan tergolong dalam kategori protein ringkas - protein, tidak seperti protein kompleks - protein, membentuk asas protoplas. Mereka disimpan dalam kuantiti yang paling banyak dalam tisu simpanan benih kering dalam bentuk bijirin aleuron, atau badan protein.
Bijirin Aleurone biasanya berbentuk sfera dan berbeza dalam saiz (0.2-20 µm). Mereka dikelilingi oleh membran dan mengandungi matriks protein amorfus, di mana kemasukan kristal direndam - satu (kurang kerap, 2-3) kristal protein berbentuk rombohedral dan bulat globoid(dari satu kepada banyak) ( nasi. 2.11). Globoid terdiri daripada phytin (garam asid heksafosforik inositol) dan merupakan tapak simpanan untuk rizab fosforus. Bijirin Aleurone yang mengandungi kristal dipanggil kompleks. Mereka adalah ciri sel penyimpanan biji minyak (rami, bunga matahari, labu, mustard, minyak jarak, dll.). Kurang biasa ringkas bijirin aleuron yang tidak mengandungi kristal, tetapi hanya protein amorf (kekacang, beras, jagung) ( nasi. 2.12).
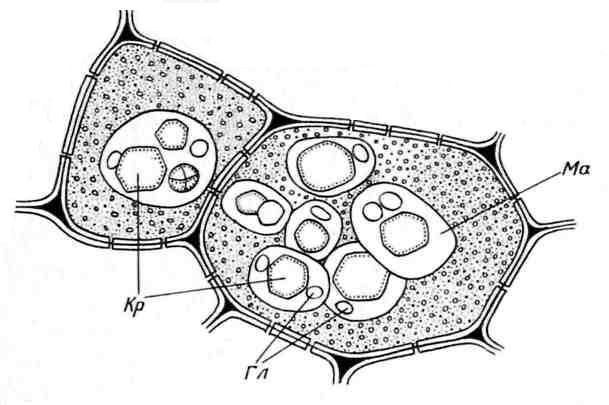
nasi. 2.11. Bijirin Aleurone dalam sel endosperm biji kastor:Kr- kristal protein; Gl– globoid; Mak- matriks protein.
Semasa perkembangan benih, protein simpanan diendapkan dalam vakuol. Apabila benih masak, disertai dengan dehidrasinya, vakuol protein menjadi kering, protein dan phytin termendap keluar dari larutan dan boleh mengkristal. Semasa percambahan biji benih, bijian aleuron menyerap air, membengkak dan beransur-ansur berubah menjadi vakuol biasa. Protein dan bahan globoid dibelanjakan untuk pertumbuhan dan perkembangan anak benih.
nasi. 2.12. Aleuron ringkas dan bijirin kanji dalam sel kotiledon biji kacang: 1 – butir aleuron mudah; 2 - bijirin kanji.
Kemasukan protein boleh diwarnakan kuning keemasan dengan reagen Lugol.
Kristal kalsium oksalat sering dijumpai dalam sel tumbuhan. Mereka disimpan hanya dalam vakuol. Bentuk kristal kalsium oksalat agak pelbagai ( nasi. 2.13) dan selalunya khusus untuk tumbuhan tertentu, yang digunakan dalam diagnosis bahan tumbuhan ubatan. boleh jadi kristal tunggal rombohedral, oktahedral atau bentuk memanjang (daun henbane), Druze– percambahan kristal sfera berbentuk bintang (daun knotweed, datura, senna, akar rhubarb), jeram– hablur kecil berbentuk jarum dikumpulkan dalam tandan (lily daun lembah, rizom madder), styloid– lebih besar, hablur berbentuk batang (lily daun lembah) dan pasir kristal– gugusan banyak kristal tunggal kecil (daun belladonna). Yang paling biasa ialah drusen. 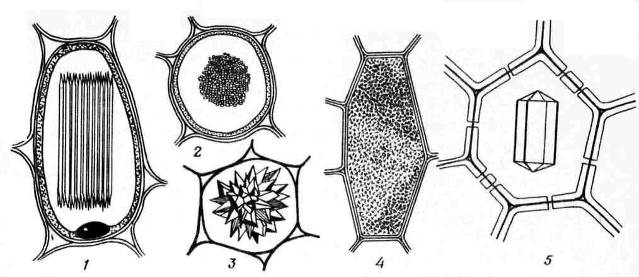
nasi. 2.13. Bentuk Kristal Kalsium Oksalat: 1,2 – raphidas (1 – pandangan sisi, 2 – pandangan keratan rentas); 3 – drusen; 4 – pasir kristal; 5 - kristal tunggal.
Di sepanjang gentian dalam kulit atau di sepanjang urat daun beberapa tumbuhan (kulit kayu oak, akar licorice, daun senna) lapisan galas kristal– terletak baris selari sel dengan kristal tunggal kalsium oksalat ( Rajah.2.14).
![]()
nasi. 2.14. Urat dengan lapisan kristal dalam daun senna.
Tidak seperti haiwan, yang membebaskan ion berlebihan semasa persekitaran luaran bersama-sama dengan air kencing, tumbuhan yang tidak mempunyai organ yang dibangunkan rembesan terpaksa mengumpulnya di dalam tisu. Biasanya dipercayai bahawa kristal kalsium oksalat adalah produk akhir hayat sel, bertujuan untuk membuang kalsium yang berlebihan. Sesungguhnya, kristal terbentuk dalam kuantiti yang banyak dalam organ dan tisu yang ditumpahkan oleh tumbuhan dari semasa ke semasa (daun dan kulit kayu). Walau bagaimanapun, terdapat bukti bahawa kristal boleh hilang dari vakuol. Dalam kes ini, mereka boleh dianggap sebagai tempat pemendapan kalsium rizab.
Sifat kimia kristal kalsium oksalat boleh disahkan oleh tindakan pekat asid mineral. Di bawah pengaruh asid hidroklorik, kristal larut. Apabila terdedah kepada asid sulfurik, kalsium oksalat berubah menjadi kalsium sulfat (gipsum) yang tidak larut, membentuk banyak kristal berbentuk jarum.
Dekat dengan kemasukan kristal cystoliths. Mereka paling kerap terdiri daripada kalsium karbonat atau silika dan merupakan pembentukan berbentuk kelompok yang timbul pada unjuran dinding sel yang menonjol ke dalam sel ( nasi. 2.15). Cystoliths adalah ciri tumbuhan dari keluarga jelatang dan mulberi. Kepentingan cystoliths masih belum dijelaskan.
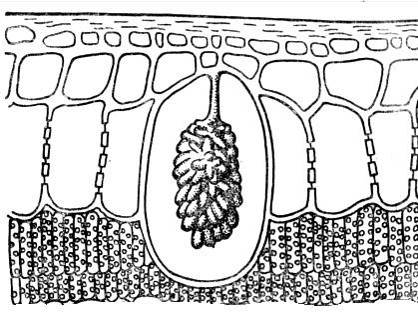
nasi. 2.15. Cystolith dalam sel epidermis daun ficus.
Dinding sel
dinding sel ( membran sel) – ciri ciri sel tumbuhan, membezakannya daripada sel haiwan. Dinding sel memberikan bentuk khusus kepada sel. Sel tumbuhan yang dibiakkan pada media nutrien khas, di mana dindingnya dikeluarkan secara enzimatik, sentiasa mengambil bentuk sfera. Dinding sel memberikan kekuatan sel dan melindungi protoplas; ia mengimbangi tekanan turgor dan dengan itu menghalang pecah plasmalemma. Gabungan dinding sel membentuk rangka dalaman yang menyokong badan tumbuhan dan memberikan kekuatan mekanikal.
Dinding sel tidak berwarna dan telus, mudah telap cahaya matahari. Biasanya dinding tepu dengan air. Sistem dinding sel mengangkut air dan sebatian berat molekul rendah yang terlarut di dalamnya (pengangkutan sepanjang apoplast).
Dinding sel terutamanya terdiri daripada polisakarida, yang boleh dibahagikan kepada bahan rangka Dan bahan matriks.
Bahan rangka dinding sel tumbuhan ialah selulosa (serat), iaitu beta-1,4-D-glucan. Ini adalah yang paling biasa bahan organik biosfera. Molekul selulosa adalah rantai tidak bercabang yang sangat panjang, ia terletak selari antara satu sama lain dalam kumpulan beberapa dozen dan disatukan oleh banyak ikatan hidrogen. Akibatnya, mikrofibril, yang mencipta kerangka struktur dinding dan menentukan kekuatannya. Mikrofibril selulosa hanya boleh dilihat dengan mikroskop elektron, diameternya ialah 10-30 nm, dan panjangnya mencapai beberapa mikron.
Selulosa tidak larut dan tidak membengkak dalam air. Ia sangat lengai secara kimia dan tidak larut dalam pelarut organik, alkali pekat dan asid cair. Mikrofibril selulosa adalah anjal dan sangat tegangan (serupa dengan keluli). Sifat-sifat ini menentukan penggunaan meluas selulosa dan produknya. Pengeluaran gentian kapas dunia, yang hampir keseluruhannya terdiri daripada selulosa, adalah 1.5 10 7 tan setahun. Serbuk mesiu tanpa asap, sutera asetat dan viscose, selofan, dan kertas diperoleh daripada selulosa. Reaksi kualitatif dijalankan pada selulosa dengan reagen klorin-zink-iodin, dinding sel selulosa bertukar menjadi biru-ungu.
Dalam kulat, bahan rangka dinding sel ialah kitin– polisakarida yang dibina daripada sisa glukosamin. Kitin lebih tahan lama daripada selulosa.
Mikrofibril direndam dalam amorfus matriks, biasanya gel plastik tepu air. Matriks adalah campuran kompleks polisakarida, molekulnya terdiri daripada sisa-sisa beberapa gula yang berbeza dan rantaian yang lebih pendek dan bercabang daripada selulosa. Polisakarida matriks menentukan sifat dinding sel seperti pembengkakan yang kuat, kebolehtelapan tinggi kepada air dan sebatian berat molekul rendah yang terlarut di dalamnya, dan sifat pertukaran kation. Polisakarida matriks dibahagikan kepada dua kumpulan - bahan pektin Dan hemiselulosa.
Bahan pektik sangat membengkak atau larut dalam air. Mereka mudah dimusnahkan oleh alkali dan asid. Wakil paling mudah bahan pektin adalah larut air asid pektik– produk pempolimeran asid alfa-D-galakturonik (sehingga 100 unit), dihubungkan oleh 1,4-ikatan ke dalam rantai linear (alfa-1,4-D-galacturonan). Asid pektin (pektin)– ini adalah sebatian polimer dengan berat molekul yang lebih tinggi (100-200 unit) asid alfa-D-galakturonik, di mana kumpulan karboksil sebahagiannya dimetilasi. Pektat Dan pectinates– garam kalsium dan magnesium daripada asid pektik dan pektik. Asid pektik, pektat dan pektinat larut dalam air dengan kehadiran gula dan asid organik untuk membentuk gel padat.
Dinding sel tumbuhan mengandungi terutamanya protopektin– polimer berat molekul tinggi asid poligalakturonik methoxylated dengan arabinan dan galaktan dalam tumbuhan dikotiledon, rantai galacturonan mengandungi sejumlah kecil rhamnose; Protopektin tidak larut dalam air.
Hemiselulosa adalah rantai bercabang yang dibina daripada sisa gula neutral, glukosa, galaktosa, mannose, xilosa adalah lebih biasa; darjah pempolimeran 50-300. Hemiselulosa secara kimia lebih stabil daripada bahan pektin lebih sukar untuk dihidrolisis dan kurang mudah membengkak dalam air. Hemiselulosa boleh dimendapkan dalam dinding sel benih sebagai bahan simpanan (kurma, kesemak). Bahan pektik dan hemiselulosa dihubungkan dengan peralihan bersama. Sebagai tambahan kepada polisakarida, protein struktur khas terdapat dalam matriks dinding sel. Ia terikat kepada sisa gula arabinosa dan oleh itu merupakan glikoprotein.
Polisakarida matriks melakukan lebih daripada sekadar mengisi ruang antara mikrofibril selulosa. Rantaian mereka disusun dengan teratur dan membentuk banyak ikatan antara satu sama lain dan dengan mikrofibril, yang meningkatkan kekuatan dinding sel dengan ketara.
Dinding sel tumbuhan sering mengalami pengubahsuaian kimia. Lignifikasi, atau lignifikasi berlaku apabila ia didepositkan dalam matriks lignin– sebatian polimer yang bersifat fenolik, tidak larut dalam air. Dinding sel berlign kehilangan keanjalannya, kekerasan dan kekuatan mampatannya meningkat dengan mendadak, dan kebolehtelapannya kepada air berkurangan. Reagen untuk lignin ialah: 1) phloroglucinol Dan asid hidroklorik pekat atau asid sulfurik(dinding berlign memperoleh warna merah ceri) dan 2) sulfataniline, di bawah pengaruh dinding lignified menjadi kuning limau. Lignifikasi adalah ciri dinding sel tisu pengalir xilem (kayu) dan tisu mekanikal sklerenkim.
Pensampelan, atau suberinisasi berlaku akibat pemendapan daripada dalam dinding sel polimer hidrofobik - suberina Dan lilin. Suberin ialah campuran ester asid lemak polimer. Monomer lilin ialah alkohol berlemak dan ester lilin. Lilin mudah diekstrak oleh pelarut organik dan cepat cair, membentuk kristal. Suberin adalah sebatian amorf yang tidak cair atau larut dalam pelarut organik. Suberin dan lilin, membentuk lapisan selari berselang-seli, melapisi seluruh rongga sel dari dalam dalam bentuk filem. Filem suberin boleh dikatakan tidak telap air dan gas, jadi selepas pembentukannya sel biasanya mati. Suberisasi adalah ciri dinding sel penutup tisu kesesakan lalu lintas Reagen untuk dinding sel suberisasi ialah Sudan III, warna oren-merah.
Coutinization Dinding luar sel tisu epidermis terdedah. Kutin Dan lilin dimendapkan dalam lapisan berselang-seli pada permukaan luar dinding sel dalam bentuk filem - kutikula. Cutin ialah sebatian polimer seperti lemak yang serupa dengannya sifat kimia dan sifat suberin. Kutikula melindungi tumbuhan daripada penyejatan air yang berlebihan dari permukaan tumbuhan. Anda boleh mengotorkannya dengan reagen Sudan III dalam warna oren-merah.
Mineralisasi dinding sel berlaku kerana pemendapan dalam matriks kuantiti yang banyak bahan mineral, selalunya silika (silikon oksida), kurang kerap oksalat dan kalsium karbonat. galian memberikan kekerasan dan kerapuhan dinding. Pemendapan silika adalah ciri sel epidermis ekor kuda, sedges dan rumput. Ketegaran batang dan daun yang diperoleh akibat silisifikasi berfungsi sebagai agen pelindung terhadap siput, dan juga mengurangkan kelazatan dan nilai pemakanan tumbuhan dengan ketara.
Sesetengah sel khusus mempunyai lendir dinding sel. Dalam kes ini, bukannya dinding sekunder selulosa, polisakarida berasid amorfus dan terhidrat tinggi didepositkan dalam bentuk lendir Dan gusi, dekat sifat kimia dengan bahan pektin. Lendir larut dengan baik dalam air untuk membentuk larutan lendir. Gusi melekit dan meregang menjadi benang. Apabila kering, mereka mempunyai konsistensi horny. Apabila lendir dimendapkan, protoplas secara beransur-ansur ditolak ke arah pusat sel, isipadu dan isipadu vakuol secara beransur-ansur berkurangan. Akhirnya, rongga sel mungkin terisi sepenuhnya dengan lendir, dan sel mati. Dalam sesetengah kes, lendir boleh melalui dinding sel primer ke permukaan. Radas Golgi memainkan peranan utama dalam sintesis dan rembesan lendir.
Lendir yang dirembeskan oleh sel tumbuhan melakukan pelbagai fungsi. Oleh itu, lendir penutup akar berfungsi sebagai pelincir, memudahkan pertumbuhan hujung akar di dalam tanah. Kelenjar lendir tumbuhan insektivor (sundews) merembeskan lendir perangkap yang melekat pada serangga. Lendir yang dirembeskan oleh sel luar kulit benih (rami, quince, pisang raja) mengikat benih ke permukaan tanah dan melindungi anak benih daripada kering. Lendir diwarnai dengan reagen metilena biru dalam warna biru.
Pembebasan gusi biasanya berlaku apabila tumbuhan cedera. Sebagai contoh, pelepasan gusi dari kawasan yang cedera pada batang dan dahan sering diperhatikan dalam ceri dan plum. Gam ceri ialah gusi yang mengeras. Gum melakukan fungsi pelindung, menutup luka dari permukaan. Gusi terbentuk terutamanya dalam tumbuhan berkayu daripada keluarga kekacang (akasia, astragalus tragacanth) dan Rosaceae daripada subfamili plum (ceri, plum, aprikot). Gusi dan lendir digunakan dalam perubatan.
Dinding sel adalah hasil daripada aktiviti penting protoplas. Polisakarida matriks, glikoprotein dinding, lignin dan lendir terbentuk dalam radas Golgi. Sintesis selulosa, pembentukan dan orientasi mikrofibril dijalankan oleh plasmalemma. Peranan besar dalam orientasi mikrofibril, ia tergolong dalam microtubules, yang terletak selari dengan mikrofibril yang didepositkan berhampiran plasmalemma. Jika mikrotubul dimusnahkan, hanya sel isodiametrik yang terbentuk.
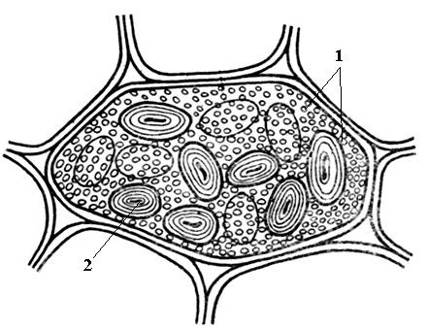
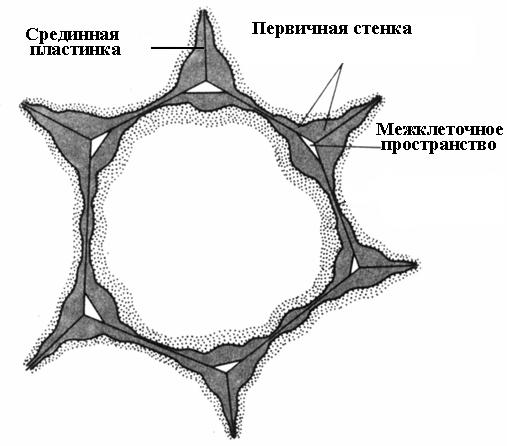
Pembentukan dinding sel bermula semasa pembahagian sel. Dalam satah pembahagian, plat sel terbentuk, satu lapisan biasa kepada dua sel anak. Ia terdiri daripada bahan pektin yang mempunyai konsistensi separa cecair; tiada selulosa. Dalam sel dewasa, plat sel dipelihara, tetapi mengalami perubahan, itulah sebabnya ia dipanggil median, atau plat antara sel (bahan antara sel)(nasi. 2.16). Plat median biasanya sangat nipis dan hampir tidak dapat dibezakan.
Sejurus selepas pembentukan plat sel, protoplas sel anak mula meletakkan dinding sel mereka sendiri. Ia dimendapkan dari dalam kedua-dua permukaan plat sel dan pada permukaan dinding sel lain yang sebelum ini dimiliki oleh sel induk. Selepas pembahagian, sel memasuki fasa pertumbuhan pemanjangan, yang disebabkan oleh penyerapan osmotik air yang kuat oleh sel yang berkaitan dengan pembentukan dan pertumbuhan vakuol pusat. Tekanan turgor mula meregangkan dinding, tetapi ia tidak koyak kerana fakta bahawa bahagian baru mikrofibril dan bahan matriks sentiasa didepositkan ke dalamnya. Pemendapan bahagian baru bahan berlaku sama rata di seluruh permukaan protoplas, jadi ketebalan dinding sel tidak berkurangan.
Dinding sel yang membelah dan membesar dipanggil utama. Mereka mengandungi banyak (60-90%) air. Bahan kering didominasi oleh matriks polisakarida (60-70%), kandungan selulosa tidak melebihi 30%, dan tiada lignin. Ketebalan dinding primer sangat kecil (0.1-0.5 mikron).
Bagi kebanyakan sel, pemendapan dinding sel terhenti serentak dengan pemberhentian pertumbuhan sel. Sel-sel tersebut dikelilingi oleh dinding primer yang nipis sehingga akhir hayat ( nasi. 2.16).
nasi. 2.16. Sel parenkim dengan dinding primer.
Dalam sel lain, pemendapan dinding berterusan walaupun selepas sel mencapai saiz akhir. Dalam kes ini, ketebalan dinding meningkat, dan jumlah yang diduduki oleh rongga sel berkurangan. Proses ini dipanggil penebalan sekunder dinding, dan dinding itu sendiri dipanggil menengah(nasi. 2.17).
Dinding sekunder boleh dianggap sebagai tambahan, melaksanakan terutamanya fungsi sokongan mekanikal. Tepat sekali dinding sekunder bertanggungjawab terhadap sifat kayu, gentian tekstil, kertas. Dinding sekunder mengandungi lebih sedikit air daripada dinding primer; ia dikuasai oleh mikrofibril selulosa (40-50% daripada berat bahan kering), yang terletak selari antara satu sama lain. Daripada polisakarida matriks, hemiselulosa (20-30%) adalah tipikal, dan terdapat sangat sedikit bahan pektin. Dinding sel sekunder biasanya mengalami lignifikasi. Dalam dinding sekunder tidak berlignifikasi (gentian kulit rami, bulu kapas), kandungan selulosa boleh mencapai 95%. Kandungan yang hebat dan orientasi mikrofibril yang diperintahkan dengan ketat menentukan sifat mekanikal yang tinggi pada dinding sekunder. Selalunya, sel-sel yang mempunyai dinding sel lignifikasi sekunder mati selepas penebalan sekunder selesai.
Lamina median melekatkan sel-sel jiran bersama-sama. Jika ia dibubarkan, dinding sel kehilangan sentuhan antara satu sama lain dan terpisah. Proses ini dipanggil maserasi. Maserasi semula jadi agak biasa, di mana bahan pektin plat tengah ditukar menjadi keadaan larut menggunakan enzim pectinase dan kemudian dibasuh dengan air (buah pir, tembikai, pic, pisang yang terlalu masak). Maserasi separa sering diperhatikan, di mana plat tengah tidak larut di seluruh permukaan, tetapi hanya di sudut sel. Oleh kerana tekanan turgor, sel-sel jiran di tempat-tempat ini dibulatkan, mengakibatkan pembentukan ruang antara sel(nasi. 2.16). Ruang antara sel membentuk rangkaian bercabang tunggal, yang diisi dengan wap air dan gas. Oleh itu, ruang antara sel meningkatkan pertukaran gas sel.
Ciri dinding sekunder - pemendapan yang tidak sekata di atas dinding primer, akibatnya kawasan yang tidak menebal kekal di dinding sekunder - liang pori. Jika dinding sekunder tidak mencapai ketebalan yang besar, liang-liang kelihatan seperti lekukan kecil. Dalam sel dengan dinding sekunder yang kuat, liang-liang dalam keratan rentas mempunyai bentuk saluran radial yang memanjang dari rongga sel ke dinding primer. Berdasarkan bentuk saluran liang, terdapat dua jenis liang: ringkas dan tentang bertepi(Gamb. 2.17).
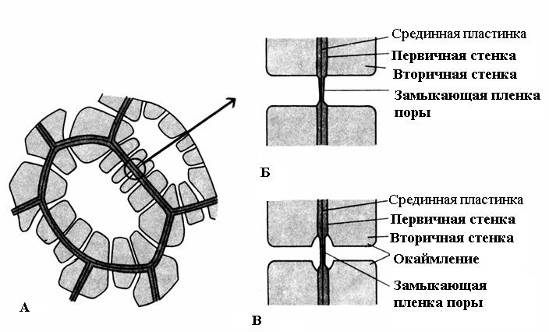
nasi. 2.17. Jenis pori: A – sel dengan dinding sekunder dan banyak liang mudah; B - sepasang liang mudah; B – sepasang pori yang bersempadan.
U pori-pori mudah Diameter saluran liang adalah sama sepanjang keseluruhan panjangnya dan mempunyai bentuk silinder sempit. Liang-liang mudah adalah ciri-ciri sel parenkim, kulit kayu dan gentian kayu.
Liang-liang dalam dua sel bersebelahan cenderung kelihatan bertentangan antara satu sama lain. Liang-liang biasa ini kelihatan seperti satu saluran, dipisahkan oleh sekatan nipis plat tengah dan dinding utama. Gabungan dua liang dinding bersebelahan sel jiran ini dipanggil sepasang pori dan berfungsi sebagai satu keseluruhan. Bahagian dinding yang memisahkan mereka dipanggil filem penutup liang, atau membran pori. Dalam sel hidup, filem penutup liang meresap dengan banyak plasmodesmata(nasi. 2.18).
Plasmodesmata hanya terdapat dalam sel tumbuhan. Ia adalah helai sitoplasma yang melintasi dinding sel bersebelahan. Bilangan plasmodesmata dalam satu sel adalah sangat besar - dari beberapa ratus hingga puluhan ribu biasanya plasmodesmata dikumpulkan dalam kumpulan. Diameter saluran plasmodesmal ialah 30-60 nm. Dindingnya dipenuhi dengan plasmalemma, berterusan dengan plasmalemma sel bersebelahan. Di tengah plasmodesmata terdapat silinder membran - rod pusat plasmadesmata, berterusan dengan membran unsur-unsur retikulum endoplasma kedua-dua sel. Antara batang pusat dan membran plasma di dalam saluran terdapat hyaloplasma, berterusan dengan hyaloplasma sel bersebelahan.
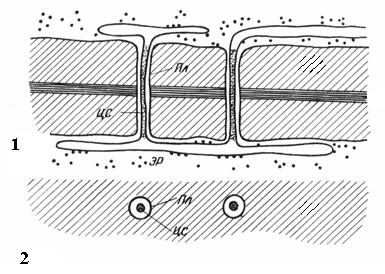
nasi. 2.18. Plasmodesmata di bawah mikroskop elektron (rajah): 1 – pada bahagian membujur; 2 - pada keratan rentas; Pl– plasmalemma; CA– batang tengah plasmodesmata; ER– unsur retikulum endoplasma.
Oleh itu, protoplas sel tidak sepenuhnya diasingkan antara satu sama lain, tetapi berkomunikasi melalui saluran plasmodesmata. Mereka membawa pengangkutan antara sel ion dan molekul kecil, dan juga menghantar rangsangan hormon. Melalui plasmodesmata, protoplas sel dalam organisma tumbuhan membentuk satu keseluruhan yang dipanggil simplast, dan pengangkutan bahan melalui plasmodesmata dipanggil simplastik tidak seperti apoplastik pengangkutan di sepanjang dinding sel dan ruang antara sel.
U pori bersempadan(nasi. 2.17) saluran mengecil secara mendadak semasa pemendapan dinding sel, jadi pembukaan dalaman liang, membuka ke dalam rongga sel, jauh lebih sempit daripada yang luaran, bersempadan dengan dinding utama. Liang-liang bersempadan adalah ciri-ciri sel mati awal unsur-unsur pengalir air kayu. Di dalamnya, saluran liang mengembang berbentuk corong ke arah filem penutup, dan dinding sekunder tergantung dalam bentuk penggelek di atas bahagian saluran yang diperluas, membentuk ruang liang. Nama pori bersempadan berasal dari fakta bahawa apabila dilihat dari permukaan, bukaan dalaman kelihatan seperti bulatan kecil atau jurang sempit, manakala lubang luar, seolah-olah, bersempadan dengan bahagian dalam dalam bentuk bulatan diameter yang lebih besar atau celah yang lebih lebar.
Liang pori memudahkan pengangkutan ke dalam