МЕРИСТЕМЫ ИЛИ ОБРАЗОВАТЕ ЛЬНЫЕ ТКАНИ
Цитоплазма. Химический состав, физическое состояние, строение и свойства. Поступление веществ в клетку. Типы движения цитоплазмы.
Цитоплазма бесцветная, имеет слизистую консистенцию и содержит различные вещества, в том числе и высокомолекулярные соединения, например белки, присутствие которых обусловливает коллоидные свойства цитоплазмы. Цитоплазма – часть протопласта, заключенная между плазмалеммой и ядром. Основу цитоплазмы составляет ее матрикс, или гиалоплазма, - сложная бесцветная, оптически прозрачная коллоидная система, способная к обратимым переходам из золя в гель.
В цитоплазме растительных клеток имеются органоиды: небольшие тельца, выполняющие специальные функции, - пластиды, комплекс Гольджи, эндоплазматическая сеть, митохондрии и т.д. В цитоплазме осуществляется большая часть процессов клеточного метаболизма, исключая синтез нуклеиновых кислот, происходящих в ядре. Цитоплазма пронизана мембранами – тончайшими (4-10нм) пленками, построенными в основном из фосфолипидов и липопротеинов. Мембраны ограничивают цитоплазму от клеточной оболочки и вакуоли и внутри цитоплазмы образуют эндоплазматическую сеть (ретикулум) – систему мелких вакуолей и канальцев, соединенных друг с другом.
Важнейшим свойством цитоплазмы, связанным прежде всего с физико-химическими особенностями гиалоплазмы, является ее способность к движению. В клетках с одной крупной вакуолью движение осуществляется обычно в одном направлении (циклоз) за счет особых органоидов – микрофиламентов, представляющих собой нити особого белка - актина. Движущая гиалоплазма увлекает пластиды и митохондрии. Клеточный сок, находящийся в вакуолях, представляет собой водный раствор различных веществ: белков, углеводов, пигментов, органических кислот, солей, алкалоидов и т.п. Концентрация веществ, находящихся в клеточном соке, обычно выше концентрации веществ во внешней среде (почва, водоемы). Различие концентраций в известной мере обусловливает возможность поступления в клетку воды и почвенных растворов, что до некоторой степени объясняется явлением осмоса. В клетке роль полупроницаемой мембраны играет цитоплазма. Пограничные слои цитоплазмы, выстилающие оболочку и клеточную вакуоль, проницаемы только для воды и некоторых растворов, но непроницаемы для многих растворенных в воде веществ. Это свойство цитоплазмы получило название полупроницаемости или избирательной проницаемости. В отличие от цитоплазмы клеточная оболочка проницаема для всех растворов, непроницаема она только для твердых частиц. Поступление веществ в клетку нельзя сводить только к осмотическим явлениям, которые выражены во взрослых клетках с хорошо развитыми вакуолями. В действительности это очень сложный процесс, обусловленный многими факторами. Активное участие в поглощении веществ принимает вся система коллоидов цитоплазмы. Интенсивность движения зависит от температуры, степени освещения, снабжения кислородом и т.д.
В очень молодых клетках цитоплазма заполняет почти всю их полость. По мере роста клетки в цитоплазме появляются мелкие вакуоли, заполненные клеточным соком, представляющим собой водный раствор различных органических веществ. Впоследствии, при дальнейшем росте клетки, вакуоли увеличиваются в размерах и, сливаясь, часто образуют одну большую центральную вакуоль, оттесняющую цитоплазму к оболочке клетки. В таких клетках все органоиды располагаются в тонком постенном слое цитоплазмы. Иногда ядро остается в центре клетки. В этом случае цитоплазма, образующая вокруг него ядерный кармашек, соединяется с постенным слоем тонкими цитоплазматическими тяжами.
В слое цитоплазмы расположены хлоропласты, выстилающие верхнюю стенку. Они представляют собой почти округлые или слегка овальные тельца. Изредка можно встетить пластиды, перетянутые посередине.
Понятие о тканях. Классификация тканей. Отличие постоянных тканей от образовательных.
У большинства наземных растений клетки тела неодинаковы по своим функциям, строению и происхождению. Эта разнородность возникла и закрепилась, в процессе приспособления растений к различиям воздушной и почвенной сред. Системы клеток, структурно и функционально сходны друг с другом и обычно имеющие общее происхождение, получили название ткачей. Ткани имеются почти у всех высших растений. Нет их только у части моховидных (печеночники). Водоросли и багрянки (низшие растения), как правило, также не имеют развитых тканей.
Комплексы клеток, сходных по функциям, а большей частью и по строению, имеющих одинаковое происхождение и определенную локализацию в теле растения, называют тканями.
Распределение тканей в органах растений и их структура тесно связаны с выполнением ими определенных физиологических функций.
Ткани, состоящие из одного типа клеток, получили название простых, а состоящие из разных типов клеток - сложных, или комплексных. Существуют различные классификации тканей, но все они достаточно условны. Растительные ткани делят на несколько групп в зависимости от основной функции:
1) меристемы, или образовательные ткани (ткани состоящие из живых тонкостенных, интенсивно делящихся клеток);
а) верхушечные (апикальные) меристимы (расположенны на верхушках стеблей и в окончаниях корней) обусловливают рост этих органов в длину;
б) боковые мерестимы – камбий и феллоген (камбий обеспечивает утолщение стебля и корня. Феллоген образует пробку)
2) покровные (защищают внутренние ткани растений от прямого влияния внешней среды, регулируют испарение и газообмен)
а) эпидермис;
б) пробка;
3) проводящие (обеспечивают проведение воды, почвенных растворов и продуктов ассимиляции, вырабатываемых листьями. Проводящие ткани по происхождению могут быть первичными и вторичными.);
а) кселима или древесная ткань (ткань проводящая воду)
б) флоэма или луб (ткань проводящая органические вещества, образованные растением в процессе фотосинтеза);
4) механические (обусловливают прочность растения);
а) коленхима (состоит из паренхимы или несколько удлиненных клеток с неравномерно утолщенными целлюлозными стенками);
б) склеренхима (клетки имеют равномерно утолщенные одревесневшие стенки);
1) волокна;
2) склериды;
5) основные (состоящие из однородных паренхимных клеток, которые заполняют пространство между другими тканями);
6) секреторные, или выделительные (содержащие продукты отброса).
Лишь клетки меристематических тканей способны к делению. Клетки прочих тканей, как правило, к делению неспособны, и их число увеличивается за счет деятельности соответствующих меристем. Такие ткани называют постоянными. Постоянные ткани возникают из меристем в результате клеточной дифференцировки. Дифференцировка заключается в том, что в ходе индивидуального развития организма (онтогенеза) возникают качественные различия между первоначально однородными клетками, при этом изменяются строение и функциональные свойства клеток. Обычно дифференцировка необратима. На ход ее оказывают влияние вещества, выполняющие роль гормонов.
МЕРИСТЕМЫ ИЛИ ОБРАЗОВАТЕ ЛЬНЫЕ ТКАНИ
Меристемы (от греч. «меристос» - делимый), или образовательные ткани, обладают способностью к делению и образованию новых клеток. За счет меристем формируются все прочие ткани и осуществляется длительный (в течение всей жизни) рост растения. У животных меристемы отсутствуют, чем объясняется ограниченный период их роста. Клетки меристем отличаются высокой метаболической активностью. Одни клетки меристем, получившие название инициальных, задерживаются на эмбриональной стадии развития в течение всей жизни растения, другие постепенно дифференцируются и превращаются в клетки различных постоянных тканей. Инициальная клетка меристемы принципиально может дать начало любой клетке организма. Тело наземных растений - производное относительно немногих инициальных клеток.
Первичные меристемы обладают меристематической активностью, т. е. способны к делению изначально. В ряде случаев способность к активному делению может вновь возникнуть и у клеток, уже почти утративших это свойство. Такие «вновь» возникшие меристемы называют вторичными.
В теле растения меристемы занимают различное положение, что позволяет их классифицировать. По положению в растении выделяют верхушечные, или апикальные (от лат. «апекс» - верхушка), боковые, или латеральные (от лат. «латус» - бок), и интеркалярные меристемы.
Апикальные меристемы располагаются на верхушках осевых органов растения и обеспечивают рост тела в длину, а латеральные - преимущественно рост в толщину. Каждый побег и корень, а также зародышевый корешок, почечка зародыша имеют апикальную меристему. Апикальные меристемы первичны и образуют конусы нарастания корня и побега (рис. 1).
Латеральные меристемы располагаются параллельно боковым поверхностям осевых органов, образуя своего рода цилиндры, на поперечных срезах имеющие вид колец. Часть из них относится к первичным. Первичными меристемами являются прокамбий и перицикл, вторичными - камбий и феллоген.
Интеркалярные, или вставочные, меристемы чаще первичны и сохраняются в виде отдельных участков в зонах активного роста (например, у оснований междоузлии, в основаниях черешков листьев).
Существуют также раневые меристемы. Они образуются в местах повреждения тканей и органов и дают начало каллюсу - особой ткани, состоящей из однородных паренхимных клеток, прикрывающие место поражения Каллюсо-образовательная способность растений используется в практике садоводства при размножении их черенками и прививками. Чем интенсивнее каллюсообразование, тем больше гарантия срастания подвоя с привоем и укоренения черенков. Образование каллюса- необходимое условие культуры тканей растения на искусственных средах.
Клетки апикальных меристем более или менее изодиаметричны по размерам и многогранны по форме. Межклетников между ними нет, оболочки тонкие, содержат мало целлюлозы. Полость клетки заполнена густой цитоплазмой с относительно крупным ядром, занимающим центральное положение. Вакуоли многочисленные, мелкие, но под световым микроскопом обычно не заметны. Эргастические вещества, как правило, отсутствуют. Пластид и митохондрий мало и они мелки.
Клетки боковых меристем различны по величине и форме. Они примерно соответствуют клеткам тех постоянных тканей, которые из них в дальнейшем возникают. Так, в камбии встречаются как паренхимные, так и прозенхимные инициали. Из паренхимных инициалей образуется паренхима проводящих тканей, а из прозенхимных - проводящие элементы.
Рис. 1. Верхушечная меристема побега элодеи. А - продольный срез; 5 - конус нарастания (внешний вид и продольный срез); В - клетки первичной меристемы;
Г - паренхимная клетка сформировавшегося листа:
1 ~ конус нарастания, 2 - зачаток листа, 3 - бугорок пазушной почки
Зоны корня. Особенности морфологического строения каждой зоны в связи с выполняемыми функциями. Рост корня в длину и втолщину.
Наряду с побегом корень - основной орган высшего растения, в типичном случае выполняющий функцию минерального и водного питания. Другая важная функция корня, тесно связана с основной, - закрепление, растения в почве.
Различные части корня выполняют неодинаковые функции и характеризуются определенными морфологическими особенностями. Эти части получили название зон (рис. 2). Кончик корня снаружи всегда прикрыт корневым чехликом, защищающим апикальную меристему. Клетки корневого чехлика продуцируют слизь, покрывающую поверхность корня. Благодаря слизи снижается трение о почву, и ее частицы легко прилипают к корневым окончаниям и корневым волоскам.
Корневой чехлик состоит из живых паренхимных клеток, возникающих у большинства однодольных из особой меристемы, получившей название калиптрогена, а у двудольных и голосеменных - из верхушечной меристемы кончика корня. Водные растения корневого чехлика обычно не имеют.
Под чехликом располагается зона деления, представленная меристематической верхушкой корня, его апексом. В результате активности апикальной меристемы формируются все прочие зоны и ткани корня.
Делящиеся клетки сосредоточены в зоне деления, имеющей размеры около 1 мм. Эта часть корня заметно отличается от прочих зон своей желтоватой окраской. Вслед за зоной деления располагается зона растяжения (роста). Она также невелика по протяженности (несколько миллиметров), выделяется светлой окраской и как бы прозрачна. Клетки зоны роста практически не делятся, но способны растягиваться в продольном направлении, проталкивая корневое окончание в глубь почвы. Они характеризуются высоким тургором, что способствует активному раздвиганию частиц почвы. В пределах зоны роста происходит дифференциация первичных проводящих тканей.
Окончание зоны роста заметно по появлению на эпиблеме многочисленных корневых волосков. Корневые волоски располагаются в зоне-всасывания, функция которой понятна из ее названия. На корне она занимает участок от нескольких миллиметров до нескольких сантиметров. В отличие от зоны роста участки этой зоны уже не смещаются относительно частиц почвы. Основную массу воды и растворов солей молодые корни усваивают в зоне всасывания с помощью корневых волосков.
Корневые волоски появляются в виде небольших сосочков - выростов клеток эпиблемы. Рост волоска осуществляется у его верхушки. Оболочка корневого волоска растягивается быстро. По прошествии определенного времени корневой волосок отмирает. Продолжительность его жизни не превышает 10-20 дней.
(меристематических клеток), и находятся в частях растений, где происходит рост.
1. Цитологические особенности меристем
Дифференцированные клетки растений вообще не могут делиться или превращаться в клетки других типов. Поэтому, деление клеток в меристемах необходим для обеспечения новых клеток для роста других тканей, образование новых органов и обеспечения структуры тела растения. За функцией меристематических клеток аналогичные стволовым клеткам животных , которые не дифференцируются или дифференцируются незначительно, и способны к непрерывному клеточного деления. Меристематических клеток маленькие, а цитоплазма и ядро полностью заполняют клетку. Вакуоли чрезвычайно маленькие, а цитоплазма не содержит дифференцированных пластид (хлоропластов или хромопластов), хотя они присутствуют в рудиментарной форме (протопластиды). Меристематических клеток плотно упакованы, почти без межклеточного пространства. Клеточная стенка - очень тонкая первичная клеточная стенка.
Физиологическая деятельность меристем требует баланса между двумя антагонистическими процессами: образованием новых тканей и поддержкой обновление популяции меристематических клеток.
Меристема - образующая растительная ткань, из которой могут образовываться все остальные ткани, меристематических клеток длительное время сохраняют способность к делению с образованием новых неспециализированных клеток.
Клетки меристемы уплощенные, мелкие, плотно расположены друг возле друга, без межклитинникив, в центре клеток расположено ядро, вакуоли мелкие, многочисленные, клеточная оболочка первичная, имеются лишь предшественники пластид.
Цитологические особенности меристем круп типично выражены в апикальных меристем. Клетки изодиаметрични многоугольники, не разделены межклетников. Клеточные стенки тонкие, с небольшим содержанием целлюлозы. Цитоплазма густая, ядро большое, расположено в центре. В цитоплазме большое калькисть рибосом и митохнодрий (происходит интенсивный синтез белков и других веществ). Многочисленные вакуоли очень мелкие.
Клетки латеральных меристем неодинаковы по размеру и форме. Это связано с отличием клеток постоянных тканей, из них образуются. Так, например, в камбии является паренхимные и прозенхимни клетки. С паренхимных инициалы образуется паренхима ведущих комплексов, с прозенхимних - собственно проводящие элементы.
Меристемы классифицируют по положению их в растении:
И по происхождению:
- первичные
- вторичные.
Первичные меристемы находятся в тесной связи с апикальной меристемы и непосредственно из них образуются, в частности, протодерма, основная меристема, прокамбий. Протодерма - однослойный пласт малодифференцированных клеток, покрывающий апекс извне, в дальнейшем из нее формируется эпидерма. Из основной меристемы формируется кора, а с прокамбия, клетки которого вытянуты вдоль оси стебля, формируется центральный цилиндр.
Вторичные меристемы вникают из специализированных клеток зрелых органов растений и обеспечивают их латеральный рост (фелоген, перицикл, камбий). Ткани, образуются из первичной меристемы, называются первичными, а с вторичной - вторичными. К вторичным меристем следует также отнести так называемые травматические меристемы, которые формируются в местах повреждения тела растений и обеспечивают регенерацию.
Среди перечисленных типов меристем первой в онтогенезе появляется апикальной меристемы, из которой происходит дифференциация всех других тканей. Расположена она на верхушках (АПЕКС) стебля и корня и их боковых ответвлений. Особенностью жизнедеятельности растительного организма является, в частности, сохранение образующей деятельности в апикальных меристем течение всего онтогенеза, поэтому говорят, что растения способны к неограниченному росту.
Строение апикальной меристемы (точки роста)
1 - Центральная часть
2 - Периферическая часть
2. Апикальной меристемы
4. Интеркалярный меристема
Интеркалярный меристема (синоним - вставная меристема) - клетки располагаются между участками дифференцированных тканей, обеспечивая вставной рост.
Интеркалярный меристема, например, располагается в узлах злаков, у основания черешков, тычиночных нитей. Это остаточные первичные меристемы. Они исходят из веривкових меристем, но их преобразование в постоянные ткани задержано по сравнению с другими тканями стебля. Это нежные меристемы особенно хорошо заметны у злаков. В случае состояние хлебов они обеспечивают поднятие стеблей за счет неравномерного деления клеток из нижнего и верхнего стороны соломинки.
5. Раневая меристема
Образующиеся при ранении тканей и органов. Живые клетки, окружающие пораженные участки, дедиференциюються и начинают делиться, то есть превращаются на вторичную меристему. Рана меристемы образуют Калюс - плотную ткань биловатого или желтоватого цвета, состоящая из паренхимных клеток разного размера, расположенных не упорядочено. Клетки каллуса имеют крупные ядра и относительно толстые клеточные стенки. Калюс может дать начало любой ткани или органа растения. На перефирии формируется пробка, возможна дифференциация клеток каллуса на другие ткани. В Калюсе могут закладаться дополнительные корни и почки. Калюс возникает при привиты растений, обеспечивает рост привоя и подвоя; в основе черенков. Также Калюс используют для получения культуры изолированных тканей.
Органы растений состоят из тканей. Растительной тканью называется совокупность клеток, имеющих сходное строение, однородное происхождение и выполняющих одну и ту же функцию. Особенного развития достигли ткани высших наземных растений. В растениях различают образовательные ткани, покровные, механические, проводящие, основные и выделительные.
Образовательные ткани, или меристемы, служат в растении для роста, они бывают первичного происхождения и вторичного. Первичная образовательная ткань находится в конусах нарастания корней и стеблей. Остановимся сперва на конусе нарастания корня (рис.1, I).
Рис.1. Первичная
образовательная ткань в конусе нарастания корня
:
I - конус
нарастания корня
: 1 - клетки образовательной
ткани; 2 - клетки корневого чехлика. II - последовательные фазы сложного
деления ядра и клетки
(кариокинез, или митоз): 1
- интерфаза; 2 - профаза; 3 - метафаза;
4-5 - анафаза; 6-7 - телофаза; 8 -
цитокинез.
Кончик корня имеет конусообразную форму, прикрыт корневым чехликом, под которым и находится первичная образовательная ткань. Клетки образовательной ткани четырехгранные или многогранные, имеют густую протоплазму, крупные ядра и тонкие целлюлозные оболочки. Вакуоли их не развиты или слабо развиты. Эти клетки обладают способностью энергичного деления. Делению клетки предшествует сложное деление ядра, в котором происходят своеобразные изменения. На готовых хорошо приготовленных и окрашенных препаратах видны бывают ядра, находящиеся в разных фазах сложного деления, называемого иначе кариокинезом («карион» - ядро, «кинезес» - движение) (рис.1, II). Многочисленными наблюдениями установлено, что в ядре сперва из хроматинового вещества возникают тонкие хроматиновые нити, которые укорачиваются, утолщаются и превращаются в отдельные частицы - хромосомы («хромо» - кpacкa, «сома» - тело), часто изогнутые в виде подковы (профаза деления). Каждая хромосома в профазе как бы расщепляется вдоль на две половины, а на самом деле происходит удвоение хромосом (редупликация). Ядрышко и ядерная оболочка постепенно исчезают. Хромосомы располагаются в средней части ядра в одной плоскости в виде пластинки. Половинки каждой хромосомы («дочерние» хромосомы) плотно прилегают одна к другой, и в этой фазе (метафазе ) на поперечных срезах корня легко можно сосчитать число хромосом и рассмотреть их форму. На полюсах ядра (еще до исчезновения ядерной оболочки) появляется система тонких ахроматиновых (неокрашивающихся) нитей, располагающихся в виде двух колпачков. Нити разрастаются и принимают вид веретена. Вслед за этим начинается расхождение половинок хромосом (дочерних хромосом) к двум противоположным полюсам клетки (анафаза ). Расхождение хромосом заканчивается тем, что дочерние хромосомы, собравшиеся у полюсов клетки, переплетаются между собой, превращаясь в два клубка (телофаза ). Одновременно в средней части клетки на ахроматиновых нитях появляются пектиновые утолщения - образуется фрагмопласт, то есть зачаток поперечной оболочки. Наконец, ядра окружаются оболочкой, в них образуются ядрышки (одно или несколько), а протоплазма разделяется поперечной оболочкой (цитокинез ), в результате чего возникают две новые клетки.
Процесс деления ядра и клетки происходит чаще в ночные часы, быстрота его зависит от температуры и влажности. Процесс деления в среднем продолжается 75-120 минут, но иногда длится несколько часов, причем на первую фазу (профазу) затрачивается больше всего времени. Клеточная оболочка при делении клетки возникает как продукт жизнедеятельности протоплазмы и ядра. Вначале возникает срединная пластинка. Она состоит из межклеточного вещества, склеивающего первичные целлюлозные оболочки, принадлежащие двум клеткам.
Впервые расхождение хромосом по направлению к полюсам клетки, так же как и другие подробности деления ядра и клетки, было установлено в 1875 г. профессором Московского университета И. Д. Чистяковым.
Сущность сложного деления ядра и клетки, называемого также митозом, заключается, по-видимому, в том, что путем равномерного расщепления хромосом на две половинки (или, точнее, удвоения хромосом) дочерние клетки получают от материнской более или менее равное количество хроматинового вещества (дезоксирибонуклеиновой кислоты - ДНК), которое передает наследственность от материнской клетки дочерним клеткам. («Митос» - нить. Обычно принято, что митоз = кариокинез + цитокинез). Форма и количество хромосом являются специфическими признаками для каждого вида растений (пшеница мягкая 2n = 42 хромосомы, пшеница твердая 2n = 28, ячмень 2n = 14 и т. д.). Однако иногда наблюдается изменчивость числа хромосом, и в одном организме можно наблюдать разнохромосомные клетки.
Кроме митоза (или кариокинеза), в растениях встречается прямое деление клеточного ядра (амитоз) - простое перетягивание ядра, а иногда и всей клетки на две половинки.
Наряду с митозом в растениях распространено редукционное деление ядра - мейоз , при котором количество хромосом в новых клетках уменьшается в два раза по сравнению с материнской клеткой. Такое деление происходит у растений при спорообразовании, вслед за которым возникают и развиваются мужские и женские половые клетки (гаметы). Последние имеют в своих ядрах половинное количество хромосом, и такие ядра называются гаплоидными в отличие от диплоидных ядер вегетативных клеток, из которых состоят вегетативные органы растений. Половой процесс растений заключается в слиянии двух гамет (мужской и женской). Если бы не происходило редукции хроматинового вещества, то есть уменьшения его массы в два раза, то в каждом новом половом поколении эта масса вещества, а следовательно, и количество хромосом неизбежно увеличивались бы. В половых клетках (гаметах) ядра бывают гаплоидными, и только после процесса оплодотворения клеточные ядра вновь становятся диплоидными.
Редукционное деление было открыто ученым В. И. Беляевым, а ученый С. Г. Навашин открыл двойное оплодотворение у цветковых растений и является основателем целой школы ученых, занимающихся изучением всех деталей деления ядра и клетки.
Первичная образовательная ткань дает начало другим тканям, так как клетки ее, изменяясь, дифференцируются. Способность клеток усиленно делиться в стеблях и корнях двудольных растений сохраняется на всю жизнь в меристематических клетках, носящих название камбия , благодаря которому утолщается стебель и корень. Ко вторичным образовательным тканям относятся такие, которые возникают из других тканей. Примером может служить феллоген, часто возникающий из основной ткани.
1) Образовательные ткани, или меристемы
Тело растения формируется в результате деятельности образовательных тканей, называемых меристемами . Основное свойство меристем – способность клеток к делению митозом, за счет наличия инициальных клеток.
В составе меристем различают : 1) инициали , 2) производные от инициалей . Инициали сохраняют способность к делению в течение всей жизни растения и всегда остаются в составе меристем. Производные от инициалей делятся некоторое число раз и затем превращаются в постоянные ткани.
Особенности инициале : густая цитоплазма, крупное ядро, много митохондрий, мелкие вакуоли или их нет, тонкая клеточная стенка.
По происхождению различают : 1) первичные меристемы , которые берут начало непосредственно от меристем зародыша. Из них развиваются первичные постоянные ткани. 2) вторичные меристемы , из первичной или других тканей. Из них развиваются постоянные вторичные ткани.
В зависимости от местоположения выделяют четыре типа меристем:
Верхушечные ( апикальные) меристемы - на верхушках стеблей и концах корней, обеспечивает рост в длину.
Боковые (латеральные) меристемы – по окружности осевых органов, обеспечиваются рост в толщину. Первичные боковые меристемы – это прокамбий и перицикл. Вторичные латеральные меристемы: камбий – возникает из прокамбия - и пробковый камбий (феллоген) - образуется из клеток постоянных тканей. Из прокамбия и камбия образуются проводящие ткани, из пробкового камбия – перидерма.
Вставочные (интеркалярные) меристемы находятся в основаниях междоузлий побегов и молодых листьев. Они первичны, поскольку являются остатками верхушечных меристем. Вставочные меристемы не имеют в своем составе инициалей и со временем полностью превращаются в постоянные ткани.
Раневые (травматические) меристемы обычно образуются при повреждении тканей и органов. Раневые меристемы образуют каллус – особую ткань, состоящую из однородных паренхимных клеток, прикрывающих место поранения. Из каллуса может возникнуть любая ткань или орган растения.
Практическое использование: цветки применяются в парфюмерии, медицине, в декоре.
2) Микроскопическое строение однодольных растений:
А) Выражена первичная кора
(лилейные,ирисовые). В этих случаях можно видеть в стебле отчетливую дифференцировку на кожицу, первичную кору и осевой цилиндр с многослойным перициклом в виде склеренхимного кольца. Первичная кора обычно состоит из нескольких слоев паренхимных хлорофиллоносных клеток. Колленхима, как правило, отсутствует. В подземных побегах первичная кора и центральный цилиндр разграничены особенно резко; первичная кора бывает значительно более широкой, чем в наземных стеблях этих же растений, в эндодерме появляются кольца Каспари или даже чашеобразные утолщения клеточных оболочек, а перицикл представлен преимущественно паренхимными клетками.
Б) У других представителей класса однодольных, в семействах пальм, злаков и других, первичная кора так четко не выражена
. В стебле кукурузы под эпидермисом иногда находятся участки хлорофиллоносной паренхимы, склеренхима окружает эти участки и между ними прилегает непосредственно к эпидермису. Периферические сосудистые пучки имеют мощные склеренхимные обкладки, и вблизи эпидермиса образуется иногда сплошной слой склеренхимы, в который погружены сосудистые пучки.
3) Общая характеристика высших растений
По своему облику, по строению и биологическим особенностям высшие растения очень разнообразны. К ним относятся не только цветковые растения, но и голосеменные, а также папоротники, хвощи, плауны и мхи.
Высшие растения характеризуются рядом признаков более высокой организации. Их тело расчленено на стебли и листья(отличие от низших).
В связи с этим их называют листостебельными растениями
.
Появление сложного расчленения, образование органов (корень, стебель, лист) было вызвано переходом растений в новую наземную среду обитания. Воздушная среда характеризуется гораздо большим содержанием кислорода, чем водная. Почвенная – иными условиями минерального питания и особенно водоснабжения. Для ассимиляции в новых условиях потребовалась перестройка фотосинтезирующего аппарата. Так у растения появился стебель.
Далее для лучшего всасывания ризоиды заменились на корни. Корни стали обеспечивать минеральное питание, а листья – воздушное питание высших растений.
Для нормального функционирования органов было необходимо быстрое передвижение воды к листьям и органических веществ – из листа в другие части растения. Это привело к развитию проводящей системы в виде специальных клеток – трахеид, сосудов и ситовидных трубок. Для защиты образовалась покровная ткань, механическая.
Половой процесс стал меньше завесить от воды.
Высшие: риниофиты, псилофиты,мхи,плауновидные,хвощевидные,папоротники, голосеменные, покрытосеменне
ТКАНИ. КЛАССИФИКАЦИЯ ТКАНЕЙ.
В основе организации высших растений лежит принцип специализации клеток, который заключается в том, что каждая клетка организма выполняет не все присущие ей функции, а только некоторые, но зато более полно и совершенно.
Ткани - устойчивые, закономерно повторяющиеся комплексы клеток, сходные по происхождению, строению и приспособленные к выполнению одной или нескольких функций.
Существуют различные классификации тканей, но все они достаточно условны.
В зависимости от основной функции различают несколько групп растительных тканей.
1. Образовательные ткани, или меристемы, - обладают способностью к делению и формированию всех прочих тканей.
2. Покровные ткани:
Первичные;
Вторичные;
Третичные.
3. Основные ткани - составляют большую часть тела растения. Различают следующие основные ткани:
Ассимиляционные (хлорофиллоносные);
Запасающие;
Воздухоносные (аэренхима);
Водоносные.
4. Механические ткани (опорные, скелетные):
Колленхима;
Склеренхима.
5. Проводящие ткани:
Ксилема (древесина) – ткань восходящего тока;
Флоэма (луб) – ткань нисходящего тока.
6. Выделительные ткани:
Наружные:
Железистые волоски;
Гидатоды – водяные устьица;
Нектарники;
Внутренние:
Выделительные клетки с эфирными маслами, смолами, дубильными веществами;
Многоклеточные вместилища выделений, млечники.
По способности клеток к делению различают два типа тканей: образовательные, или меристемы, и постоянные - покровные, выделительные, основные, механические, проводящие.
Ткань называют простой, если все ее клетки одинаковы по форме и функциям (паренхима, склеренхима, колленхима). Сложные ткани состоят из клеток, неодинаковых по форме, строению и функциям, но связанных общим происхождением (например, ксилема, флоэма).
Существует также классификация тканей, основанная на их происхождении (онтогенетическая). Согласно этой классификации различают первичные и вторичные ткани. Из первичной меристемы, находящейся на верхушке побега и кончике корня, а также из зародыша семени формируются первичные постоянные ткани (эпидерма, колленхима, склеренхима, ассимиляционная ткань, эпиблема). Клетки постоянных тканей неспособны к дальнейшему делению. Из клеток специализированной меристемы – прокамбия - формируются первичные проводящие ткани (первичная ксилема, первичная флоэма).
Из вторичной меристемы – камбия - формируются вторичные ткани: вторичная ксилема, вторичная флоэма; из феллогена образуются пробка, феллодерма, чечевички, возникающие при утолщении стебля и корня. Вторичные ткани, как правило, встречаются у голосеменных и двудольных покрытосеменных растений. Мощное развитие вторичных тканей – древесины и луба характерно для древесных растений.
ОБРАЗОВАТЕЛЬНЫЕ ТКАНИ
Образовательные ткани благодаря постоянному митотическому делению их клеток обеспечивают образование всех тканей растения, т.е. фактически формируют его тело. Любая клетка в своем развитии проходит три стадии: эмбриональную, роста и стадию дифференциации (то есть приобретения клеткой определенной функции). По мере дифференциации зародыша первичная меристема сохраняется только на верхушке будущего побега (в конусе нарастания) и на кончике корня – апикальные (верхушечные) меристемы. Зародыш любого растения состоит из клеток меристемы.
Цитологическая характеристика меристем. Типичные признаки наиболее отчетливо выражены в верхушечных меристемах. Эти меристемы составлены изодиаметрическими многогранными клетками, не разделенными межклетниками. Их оболочки тонки, содержат мало целлюлозы и способны растягиваться.
Полость каждой клетки заполнена густой цитоплазмой с относительно крупным ядром, занимающим центральное положение, и интенсивно делящейся митозом. В гиалоплазме много диффузно разбросанных рибосом, пропластид, митохондрий и диктиосом. Вакуолей немного, и они мелкие. Проводящие ткани образуются из меристемы, имеющей прозенхимную форму и крупные вакуоли, – прокамбия и камбия. Клетки прокамбия в поперечном сечении многоугольные, клетки камбия – прямоугольные.
Клетки, сохраняющие свои меристематические свойства, продолжают делиться, образуя все новые и новые клетки, называемые инициалями. Часть дочерних клеток дифференцируется, превращаясь в клетки различных тканей, их называют производными инициалей. Клетки инициалей могут делиться неопределенно много раз, а производные инициалей делятся один или несколько раз и превращаются в постоянные ткани.
По происхождению различают первичные и вторичные меристемы.
Первичные меристемы
Первичные меристемы происходят непосредственно из меристемы зародыша и обладают способностью к делению. По положению в растении первичные меристемы могут быть верхушечными (апикальными), вставочными (интеркалярными) и боковыми (латеральными).
Верхушечные (апикальные) меристемы - такие меристемы, которые располагаются у взрослых растений на верхушках стеблей и кончиках корней и обеспечивают рост тела в длину. У стеблей в конусе нарастания выделяют два меристематических слоя: тунику, из которой образуются покровная ткань и периферическая часть первичной коры, и корпус, из которого образуются внутренняя часть первичной коры и центральный осевой цилиндр (рис. 2.3).
Рис. 2.3. Апикальные меристемы стебля: а - продольный срез: 1 – конус нарастания; 2 – зачаток листа; 3 – бугорок пазушной почки;
В кончике корня различают три слоя:
1) дерматоген, из которого образуется первичная покровно-всасывающая ткань – ризодерма;
2) периблему, из которой развиваются ткани первичной коры;
3) плером, образующий ткани центрального осевого цилиндра.
по происхождению могут быть первичными и вторичными, на поперечном срезе осевых органов имеют вид колец. Примером первичной боковой меристемы служат прокамбий и перицикл. Из прокамбия формируются камбий и первичные элементы сосудисто-волокнистых пучков (первичная флоэма и первичная ксилема), при этом клетки прокамбия непосредственно дифференцируются в клетки первичных проводящих тканей.
Боковые меристемы располагаются параллельно поверхности органа и обеспечивают рост осевых органов в толщину.
Вставочные (интеркалярные) меристемы чаще первичные и сохраняются в виде отдельных участков в зонах активного роста в различных частях растения (например, в основании черешков листьев, у оснований междоузлий). В основании междоузлий у злаков деятельность этой меристемы ведет к удлинению междоузлий, что обеспечивает рост стебля в длину.
Вторичные меристемы
К вторичным относят боковые и раневые меристемы.
Боковые (латеральные) меристемы представлены камбием и феллогеном. Они формируются из промеристем (прокамбия) или постоянных тканей путем их дедифференцировки. Клетки камбия делятся перегородками, параллельными поверхности органа (периклинально). Из клеток, отложенных камбием наружу, развиваются элементы вторичной флоэмы, а из клеток, отложенных внутрь, – вторичной ксилемы. Камбий, возникший из постоянных тканей путем дедифференцировки, называют добавочньм. По строению и функции он не отличается от камбия, возникшего из промеристем. Феллоген формируется из постоянных тканей, расположенных в субэпидермальных слоях (под эпидермой). Делясь периклинально, феллоген отделяет наружу будущие клетки пробки (феллемы), а внутрь – клетки феллодермы. Таким образом, феллоген формирует вторичную покровную ткань – перидерму. Боковые меристемы располагаются параллельно поверхности органа и обеспечивают рост осевых органов в толщину.
Раневые меристемы образуются при повреждении тканей и органов. Вокруг повреждения живые клетки дедифференцируются, начинают делиться и тем самым превращаются во вторичную меристему. Их задача – образовать плотную защитную ткань, состоящую из паренхимных клеток, – каллюс. Эта ткань беловатого или желтоватого цвета, ее клетки имеют крупные ядра и достаточно толстые клеточные стенки. Каллюс возникает при прививках, обеспечивая срастание привоя с подвоем, и в основании черенков. В нем могут закладываться придаточные корни и почки, поэтому его используют для получения культуры изолированных тканей.
ПОКРОВНЫЕ ТКАНИ
Первичная покровная ткань
К первичной покровной ткани относят эпидерму собственно эпидермальных, околоустьичных, замыкающих клеток устьица и трихом.
Пектиновые вещества и целлюлоза, входящие в клеточную стенку, могут подвергаться ослизнению с образованием слизей и камедей. Они представляют собой полимерные углеводы, родственные пектиновым веществам, и отличаются способностью к сильному набуханию при соприкосновении с водой. Камеди в набухшем состоянии клейкие и могут вытягиваться в нити, тогда как слизи сильно расплываются и в нити не вытягиваются. Пектиновые слизи встречаются у представителей семейств лилейных, крестоцветных, мальвовых, липовых, розоцветных, в отличие от целлюлозных слизей, встречающихся гораздо реже (например, у орхидных).
 Устьица
представляют собой высокоспециализированные образования эпидермы, состоящие из двух замыкающих клеток бобовидной формы и устьичной щели (своеобразного межклетника между ними). Имеются главным образом в листьях, но встречаются и в стебле (рис. 2.6).
Устьица
представляют собой высокоспециализированные образования эпидермы, состоящие из двух замыкающих клеток бобовидной формы и устьичной щели (своеобразного межклетника между ними). Имеются главным образом в листьях, но встречаются и в стебле (рис. 2.6).
Рис. 2.6. Строение устьица: а, б - кожица листа тимьяна (вид сверху и на поперечном срезе); в - кожица со стебля цереуса (семейство кактусовых); 1 – собственно эпидермальные клетки; 2 – замыкающие клетки устьица; 3 – устьичная щель; 4 – воздухоносная полость; 5 – клетки хлорофиллоносной паренхимы; А – кутикула; Б – кутикулярный слой – оболочка с суберином и воском; В – целлюлозный слой стенки; Г – ядро с ядрышком; Д – хлоропласты
Стенки замыкающих клеток утолщены неравномерно: стенки, направленные к щели (брюшные), значительно утолщены по сравнению со стенками, направленными от щели (спинными). Щель может расширяться и сужаться, регулируя транспирацию и газообмен. Под щелью расположена крупная дыхательная полость (межклетник), окруженная клетками мезофилла листа.
 Замыкающие клетки окружены околоустьичными клетками, которые совместно образуют устьичный комплекс
(рис. 2.7). Различают следующие основные типы устьичных комплексов:
Замыкающие клетки окружены околоустьичными клетками, которые совместно образуют устьичный комплекс
(рис. 2.7). Различают следующие основные типы устьичных комплексов:
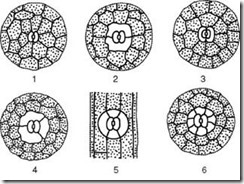
Рис. 2.7. Основные типы устьичного аппарата: 1 – аномоцитный (у всех высших растений, кроме хвощей); 2 – диацитный (у папоротников и цветковых); 3 – парацитный (у папоротников, хвощей, цветковых и гнетовых); 4 – анизоцитный (только у цветковых); 5 – тетрацитный (главным образом у однодольных); 6 – анциклоцитный (у папоротников, голосеменных и цветковых)
1) аномоцитный (беспорядочный) – замыкающие клетки не имеют ярко выраженных околоустьичных клеток; характерен для всех высших растений, исключая хвойные;
2) анизоцитный (неравноклеточный) – замыкающие клетки устьица окружены тремя околоустьичными клетками, одна из которых намного крупнее (или меньше) остальных;
3) парацитный (параллельно-клеточный) – одна околоустьичная клетка (или более) расположена параллельно замыкающим;
4) диацитный (перекрестно-клеточный) – две околоустьичные клетки расположены перпендикулярно замыкающим;
5) тетрацитный (от греч. tetra - четыре) – главным образом у однодольных;
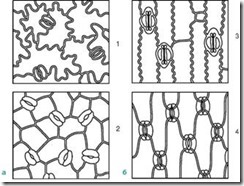 Устьица расположены на нижней стороне листа, но у водных растений с плавающими листьями они находятся только на верхней стороне листа. По форме клеток эпидермы листа и расположению устьиц можно отличить однодольное растение от двудольного (рис. 2.8). Собственно эпидермальные клетки листа двудольных растений в очертаниях волнистые (рис. 2.9), а у однодольных они вытянутые, ромбической формы.
Устьица расположены на нижней стороне листа, но у водных растений с плавающими листьями они находятся только на верхней стороне листа. По форме клеток эпидермы листа и расположению устьиц можно отличить однодольное растение от двудольного (рис. 2.8). Собственно эпидермальные клетки листа двудольных растений в очертаниях волнистые (рис. 2.9), а у однодольных они вытянутые, ромбической формы.
Рис. 2.8. Расположение устьиц на эпидерме (вид с поверхности): а - двудольные растения: 1 - буквица; 2 – арбуз; б - однодольные растения: 3 – кукуруза; 4 – ирис
Типы устьиц по уровню расположения относительно поверхности эпидермиса можно подразделить следующим образом.
1.7.1. Устьица, расположенные в одной плоскости с эпидермисом. Наиболее распространенный тип и обычно в описании микроскопии лекарственного растительного сырья не указывается, т.е. данный пункт опускается. Диагностическими признаками будут либо выступающие, либо погруженные устьица.
1.7.2. Выступающие устьица - устьица, расположенные над эпидермисом. Обычно при вращении микровинта микроскопа (при опускании объектива) сначала обнаруживаются такие устьица, а уже затем появляются клетки эпидермиса, поэтому на фотографии запечатлеть их с поверхности листа практически невозможно, равно как и изобразить на рисунке. В одной плоскости с эпидермисом такие устьица можно увидеть на поперечных срезах, но для этого срез должен пройти через устьице, что трудно получить при их редком расположении на листе. Такие устьица характерны, например, для листьев толокнянки.
1.7.3. Погруженные устьица - устьица, погруженные в эпидермис. При наблюдении под микроскопом при вращении микровинта (при опускании объектива) сначала четко обнаруживаются клетки эпидермиса, затем становится возможным более четко увидеть контуры устьиц. На фотографиях и рисунках препаратов с поверхности их также трудно отобразить. Встречаются в листьях ландыша, листьях вахты, листьях эвкалипта. Иногда углубления, в которых располагаются устьица, выстланы или прикрыты волосками и называются устьичными криптами.
1.8. Типы устьичных клеток
В литературе описывается 19 типов , нами выбраны только те, которые используются в анализе лекарственного растительного сырья**.
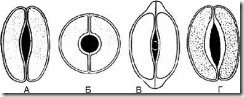
Рис. 63. Типы устьичных клеток. А – чечевицевидные; Б – сферовидные; В – колпачковидные; Г – ладьевидные
1.8.1. Чечевицевидные - 2 одинаковые клетки полулунной формы расположены симметрично. На фронтальной плоскости утолщение оболочки почти равномерное. Щель веретеновидная (рис. 63, А). Тип устьичных клеток характерен для большинства растений.
1.8.2. Сферовидные - две одинаковые, сильно кругообразноизогнутые клетки расположены симметрично. На фронтальной плоскости утолщение оболочки почти равномерное. Щель круглая (рис. 63, Б).
1.8.3. Колпачковидные - две одинаковые клетки полулунной формы в полярных частях имеют утолщения в виде колпачка. Щель веретеновидная (рис. 63, В). Встречаются у наперстянок.
1.8.4. Ладьевидные - внутренние стенки устьичных клеток утолщены. Щель веретеновидная (рис. 63, Г). Наблюдается в траве золототысячника, в листьях вахты.
Механизм работы устьиц обусловлен осмотическими свойствами клеток. При освещении поверхности листа солнцем в хлоропластах замыкающих клеток происходит активный процесс фотосинтеза. Насыщение клеток продуктами фотосинтеза, сахарами влечет за собой активное поступление в клетки ионов калия, вследствие чего концентрация клеточного сока в замыкающих клетках возрастает. Возникает разность концентрации клеточного сока околоустьичных и замыкающих клеток. В силу осмотических свойств клеток вода из околоустьичных клеток поступает в замыкающие, что ведет к увеличению объема последних и резкому возрастанию тургора. Утолщение «брюшных» стенок замыкающих клеток, обращенных к устьичной щели, обеспечивает неравномерное растяжение клеточной стенки; замыкающие клетки приобретают выраженную бобовидную форму, и устьичная щель открывается. При снижении интенсивности фотосинтеза (например, вечером) уменьшается образование сахаров в замыкающих клетках. Приток ионов калия прекращается. Концентрация клеточного сока в замыкающих клетках снижается по сравнению с околоустьичными. Вода путем осмоса уходит из замыкающих клеток, понижая их тургор, в результате ночью устьичная щель закрывается.
Клетки эпидермы плотно сомкнуты между собой, благодаря этому эпидерма выполняет целый ряд функций:
Препятствует проникновению болезнетворных организмов внутрь растения;
Защищает внутренние ткани от механических повреждений;
Регулирует газообмен и транспирацию;
Через нее выделяются вода, соли;
Может функционировать как всасывающая ткань;
принимает участие в синтезе различных веществ, восприятии раздражений и в движении листьев.
Трихомы - различные по форме, строению и функциям выросты клеток эпидермы: волоски, чешуйки, щетинки и т.п. Их подразделяют на кроющие и железистые. Железистые трихомы, в отличие от кроющих, имеют клетки, выделяющие секрет. Кроющие волоски, образуя на растении шерстистый, войлочный или иной покров, отражают часть солнечных лучей и тем самым уменьшают транспирацию. Иногда волоски находятся только там, где расположены устьица, например, на нижней стороне листа мать-и-мачехи. У некоторых растений живые волоски увеличивают общую испаряющую поверхность, что способствует ускорению транспирации.
Размеры трихом значительно варьируют. Наиболее длинные трихомы (до 5-6 см) покрывают семена хлопчатника. Кроющие трихомы имеют форму простых одно или многоклеточных, разветвленных или звездчатых волосков. Кроющие трихомы могут длительное время оставаться живыми или быстро отмирать, заполняясь воздухом.
От трихом, возникающих только при участии эпидермальных клеток, отличаются эмергенцы, в образовании которых участвуют и более глубоко расположенные ткани субэпидермальных слоев.
Анатомо-диагностические признаки, имеющие наибольшее значение и высокую вариабельность при определении лекарственного сырья. Волоски могут быть простые и головчатые, которые в свою очередь могут быть одноклеточными и многоклеточными. Многоклеточные волоски могут быть однорядными, двухрядными и ветвистыми.
1.9.1. Простые волоски.
А. Простые одноклеточные волоски
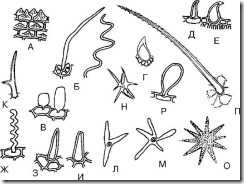
Рис. 68. Простые одноклеточные волоски. А – сосочковидный; Б – нитевидный; В – пузыревидные; Г – шиповидный; Д – крючковидный; Е – ретортовидный; Ж – тупонитевидный гофрированный; З – остроконусовидный; И – тупоконусовидный; К – двухконечный; Л – трехконечный; М, Н, О – многоконечный; П – бугристый; Р – булавовидный
1. Сосочковидные (рис. 68, А; рис. 69-74) – невысокие, но широкие выросты эпидермальных клеток. Чаще образуются на лепестках. Могут быть туповерхушечные и островерхушечные. Встречаются на листьях чабреца, цветках ландыша, цветках фиалки трехцветной, по краю листьев золототысячника, в траве горечавки бородатой и др.
2. Конусовидные (рис. 68, З, И; рис. 75-79) – прямые и наклоненные под углом к поверхности волоски, оканчивающиеся тупым (тупоконусовидные) или острым (остроконусовидные) концом. Наиболее широко распространенные волоски. Обычно бывают прямые. Наклоненные к поверхности волоски наблюдаются на листьях чабреца. Остроконусовидные волоски встречаются на листьях сенны, листьях брусники, в траве фиалки трехцветной (на листьях), в траве чабреца и др. Тупоконусовидные волоски имеются на эпидермисе плодов аниса, на лепестках цветков фиалки трехцветной.
3. Нитевидные (рис. 68, Б,Ж; рис. 80, 81) – тонкие и длинные волоски. Могут быть прямые и гофрированные. Прямые нитевидные волоски обнаруживаются на эпидермисе плодов боярышника и малины. Гофрированные тупонитевидные волоски наблюдаются на эпидермисе лепестков фиалки (рис. 82).
4. Ретортовидные (рис. 68, Е; рис. 83) – волоски с расширенным основанием и узким изогнутым или прямым окончанием. Встречаются в соплодиях хмеля, траве зубчатки.
5. Пузыревидные (рис. 68,В; рис. 84) – волоски в виде небольших пузырей. Их можно обнаружить, например, в цветках бессмертника.
6. Крючковидные (рис. 68,Д; рис. 85-87) – волоски, заостренные на верхушке и изогнутые в виде крючка. Эти волоски можно встретить у основания листьев толокнянки, на поверхности листьев брусники, сенны, плодов аниса.
7. Бахромчатые (рис. 88, 89) – длинные выросты эпидермальных клеток, расположенные по краю листа, лепестка или чашелистика. Такие волоски имеются, например, по краю лепестков ландыша и фиалки.
8. Шиповидные (рис. 68, Г; рис. 90) – волоски почти округлой формы с заостренным концом. Наблюдаются в траве фиалки трехцветной.
9. Булавовидные (рис. 68, Р; рис. 91) – волоски с расширенной конечной частью, напоминающие булаву. Имеются на лепестках чабреца, лепестках фиалки трехцветной.
10. Двухконечные (рис. 68, К) – волоски, разветвленные на два конца. Наблюдаются в траве пастушьей сумки.
11. Трехконечные (многоконечные) (рис. 68, Л-О) – волоски, разветвленные на три (и более) конца. Наблюдаются в траве пастушьей сумки.
12. Бугристые (рис. 68, П) – конусовидные волоски, имеющие выступы (бугры) на своей поверхности.
Б. Простые многоклеточные однорядные волоски
Контур многоклеточного волоска может совпадать с таковым одноклеточного волоска, но содержать две и более клеток и соответственно называться как аналогичный одноклеточный волосок с указанием количества клеток, например 2-клеточный ретортовидный волосок, 11- клеточный нитевидный волосок, 10-15-клеточный конусовидный волосок.

Рис. 92. Простые многоклеточные волоски. А – конусовидный; Б – пузыревидный; В – суставчатый; Г – бичевидный; Д – пе-ристый; Е – Т-образный; Ж – двухрядные; З – чешуйчатый; И – пучковый; Л – параллельный; М – вильчатый; К – щетинистые
1. Конусовидные (рис. 92, А; 93-96). Наиболее широко встречающиеся волоски. Бывают прямые и наклоненные к поверхности, последние встречаются редко (например, в траве тимьяна). Конусовидные волоски могут быть остро- и тупоконусовидные. Более распространены остроконусовидные волоски (в траве душицы, в листьях мяты, шалфея и др.). Тупоконусовидные встречаются в цветках календулы.
2. Ретортовидные - волоски с расширенным основанием и узким изогнутым или прямым окончанием.
3. Пузыревидные (рис. 92, Б; рис. 97) – волоски в виде небольших пузырей. Имеются на поверхности завязи бессмертника.
4. Нитевидные - тонкие и длинные многоклеточные волоски.
5. Крючковидные - волоски, заостренные на верхушке и изогнутые в виде крючка.
6. Гусеницевидные (рис. 98, 99) – волоски практически одинаковой толщины на всем своем протяжении, состоящие из почти одинаковых коротких клеток, с тупым концом и напоминающие гусеницу. Наблюдаются в траве фиалки трехцветной, в траве череды.
7. Бахромчатые (рис. 100) – длинные многоклеточные выросты эпидермальных клеток, расположенные по краю листа, лепестка или чашелистика. Встречаются в траве череды.
8. Бичевидные (рис. 92, Г; рис. 101-104) – волоски, имеющие многоклеточное основание, состоящее из цепи коротких клеток, и длинную нитевидную извилистую конечную клетку. В литературе такие волоски описывают как кнутовидные, шнуровидные, нитевидные, войлочные. Тип волосков характерный для семейства Asteraceae. Встречается в траве тысячелистника, цветках бессмертника, листьях мать-и-мачехи, цветках пижмы и др.
9. Суставчатые (рис. 92, В; рис. 105, 106) – волоски, имеющие расширенные основания клеток в местах их сочленения (напоминающие сустав). Подобные волоски обнаруживаются в траве пустырника, на лепестках душицы, редко в траве чабреца.
10. Бугристые - волоски, имеющие выступы (бугры) на своей поверхности. Двухклеточные бугристые волоки наблюдаются, например, в листве термопсиса.
В. Простые многоклеточные ветвистые волоски
1. Параллельные (рис. 92, Л; рис. 107, 108) – волоски, состоящие из двух длинных прямых клеток (волосков), сросшихся основаниями. Встречаются в цветках липы, редко в плодах шиповника и боярышника.
2. Вильчатые (рис. 92, М; рис. 109, 110) – волоски, состоящие из двух длинных извилистых клеток (волосков), сросшихся основаниями. Наблюдаются в цветках липы, очень редко в цветках и плодах боярышника.
3. Звездчатые (рис. 111) – волоски, состоящие из 3 и более длинных извилистых клеток (волосков), сросшихся основаниями. Обнаруживаются в цветках липы.
4. Чешуйчатые (рис. 92, З) – волоски, состоящие из многоклеточной пластинки (в виде розетки) и короткой ножки (ножка может отсутствовать). Такие волоски имеются в облепихе. (Данное название волосков приведено в соответствии с современной ботанической терминологией ; по другим данным эти волоски называют звездчатыми или щитковидными чешуйчатыми ).
5. Т-образные (рис. 92, Е) – двухконечный волосок, имеющий одно-, многоклеточную ножку. В литературе описываются также как рогообразные, коромыслообразные.
6. Перистые (рис. 92, Д) – многоклеточные волоски, напоминающие ветвящееся дерево. Обнаруживаются, например, в коровяке.
Г. Простые многоклеточные многорядные волоски
1. Двухрядные (рис. 92, Ж; рис. 112, 113) – волоски, клетки которых расположены в два ряда. Их можно наблюдать в цветках календулы, в траве сушеницы.
2. Пучковые (рис. 92, И) – волоски, состоящие из двух и более клеток, плотно сросшихся друг с другом, образуя пучок.
3. Щетинистые (рис. 92 К, рис. 114) – многоклеточные волоски, состоящие из сросшихся друг с другом волосков разной длины и имеющие свободные заостренные концы. Такие волоски обычно являются волосками летучки (семейство Asteraceae). В литературе их еще называют зазубренными сложными, шилообразными.
1.9.2. Головчатые волоски. Головчатые волоски называют также железистыми. Они могут быть одноклеточными и многоклеточными.

Рис. 115. Головчатые волоски. А – с одноклеточной ножкой и одноклеточной головкой; Б – с двухрядной ножкой и одноклеточной головкой; В – с двухрядной ножкой и двурядной головкой; Г – с одноклеточной ножкой и многоклеточной головкой; Д – с многоклеточной ножкой и одноклеточной головкой; Ж – с многоклеточной ножкой и многоклеточной головкой; З – одноклеточный (жгучий); И – щитковидный
А. Одноклеточные головчатые волоски
Представители этого типа волосков могут быть конусовидными или ретортовидными, но всегда имеют на конце головку. Такие волоски есть, например, на листьях крапивы. Б. Многоклеточные головчатые (железистые) волоски
1. Волоски, имеющие многоклеточную головку и одноклеточную ножку (рис. 115, Г; рис. 116-120). Встречаются в траве пустырника, листьях толокнянки.
2. Волоски, имеющие одноклеточную головку и одноклеточную ножку (рис. 115, А; рис. 121-123). Встречаются в траве пустырника, листьях шалфея, цветках бузины, листьях мяты.
3. Волоски, имеющие одноклеточную головку и многоклеточную ножку (рис. 115, Д; рис. 124). Встречаются в траве пустырника, листьях шалфея, цветках календулы и цветоножках ромашки.
4. Волоски, имеющие многоклеточную головку и многоклеточную ножку (рис. 115, Ж; рис. 125). Встречаются в траве пустырника, листьях толокнянки, цветках липы, цветках календулы, траве фиалки.
5. Волоски, имеющие одноклеточную головку и двухрядную ножку (рис. 115, Б; рис. 126). Встречаются в цветках календулы и бессмертника.
6. Волоски, имеющие двухрядную головку и двухрядную ножку (рис. 115, В; рис. 127- 130). Встречаются в цветках календулы и бессмертника, в траве сушеницы топяной.
7. Булавовидные многоклеточные железистые волоски (рис. 131-133; см. рис. 155, Д) – волоски, имеющие контур с расширенной конечной частью, напоминающие булаву. Их можно наблюдать, например, в листьях брусники, траве фиалки.
8. Щитковидные многоклеточные железистые волоски (рис. 115, И; рис. 134) представляют собой щиток из многоугольных тонкостенных клеток, сидящий на одно-, двухклеточной короткой ножке. Кутикула щитка отодвигается от клеток выделяющимся под нее эфирным маслом. Они обнаружены в соплодиях хмеля.
1.10. Характер утолщенности клеточных стенок и покрывающей кутикулы волосков
А. Характер утолщенности клеточных стенок
1.10.1. Тонкостенные (рис. 135-137). Большинство волосков тонкостенные. Стенки клеток многоклеточных длинных тонкостенных волосков иногда спадаются, нарушая ровный контур волоска. Такие волоски, например, встречаются на нижнем эпидермисе листа какалии.
1.10.2. Толстостенные (рис. 138, 139). Встречаются в плодах шиповника, траве череды, траве лепедецы даурской и др.
1.10.3. Неравномерно утолщенные. Можно наблюдать в траве пустырника (рис. 140). К этой же категории относятся волоски шиповника с косопоперечными порами (рис. 141, 142).
Б. Характер покрывающей кутикулы волосков
1.10.4. Гладкая поверхность (рис. 143). Волоски с такой поверхностью наблюдаются, например, в листьях подорожника, траве чистотела и др.
1.10.5. Бородавчатая поверхность (см. рис. 1, Ж; рис. 144, 145) – эпидермис образует выступы в виде бугорков (бородавок). Имеются волоски, например, в траве фиалки, в траве чабреца, в траве пустырника, в листьях мяты, в листьях сенны и др. При этом поверхность волоска может быть слабобородавчатой, когда образуются небольшие выступы кутикулы, и грубобородавчатой, когда выступы образуются значительные. В первом случае в качестве примера можно привести волоски в траве фиалки, во втором – волоски листьев сенны и травы чабреца.
1.10.6. Крупнобугристая поверхность (см. рис. 68, П) – кутикула образует очень большие выступы, например волоски травы термопсиса ланцетного.
10.7. Морщинистая поверхность - складки или волны кутикулы волоска от основания к его вершине (рис. 146). Наблюдается чаще у сосочков, например, в траве золототысячника, цветках бузины. Однако морщинистая поверхность может быть и у обычных волосков, например, в траве череды. У основания морщинистую поверхность имеют волоски листьев мяты (рис. 147).
1.10.8. Штриховатая поверхность - эпидермис образует короткие выступы в виде штрихов (бугорков, бородавок). Редко встречается и является промежуточным вариантом между бородавчатой и морщинистой поверхностью. Наблюдается, например, на волосках цветков бузины (рис. 148).
1.11. Особенности мест присоединения волосков
1.11.1. Обычные места прикрепления (рис.149, 150) – волоски присоединяются к клетке или между клеток эпидермиса. Наиболее распространенный тип встречается, например, в листьях мать-и-мачехи, листьях мяты, листьях шалфея, траве пустырника, траве фиалки и др.
1.11.2. Выросты из клеток эпидермиса (см. рис. 68, А; 69-74). Наблюдаются в цветках бузины, в траве золототысячника, лепестках пустырника, в траве горечавки бородатой, эпидермисе плодов аниса и др.
1.11.3. У основания волоска образуется розетка из клеток эпидермиса (рис. 151, 152). Имеется в листьях сенны, листьях подорожника, в траве душицы и др.
1.11.4. Расширенное основание волоска (рис. 153). Встречается в листьях подорожника.
1.11.5. Расширенная часть основания волоска погружена в цоколь из субэпидермальных тканей – эмергенцы (см. рис. 115, 3). Можно наблюдать, например, в листьях крапивы.
1.11.6. Многоклеточное основание волосков (рис. 154). Можно наблюдать, например, в траве череды.
Нередко волоски обламываются, оставляя на эпидермисе места своего прикрепления, что также необходимо отмечать как анатомо-диагностический признак лекарственного растительного сырья.
ВТОРИЧНАЯ ПОКРОВНАЯ ТКАНЬ
Вторичную покровную ткань называют перидермой. Это сложная покровная ткань стеблей, корней и корневищ многолетних растений. Она сменяет эпидерму осевых органов, которая постепенно отмирает и слущивается. Перидерма образуется из феллогена (вторичной меристемы). Феллоген закладывается в эпидерме, субэпидермальном слое и даже в глубоких слоях осевых органов. Клетки феллогена делятся следующим образом наружу откладывают клетки пробки, а внутрь – живые паренхимные клетки феллодермы. В клетках феллодермы стеблей содержатся хлоропласты.
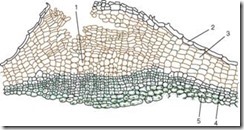 Пробка состоит из мертвых клеток, у которых клеточная стенка пропитана жироподобным веществом суберином. Клетки расположены ровными рядами, имеют прямоугольную форму (на поперечном срезе), плотно прилегают друг к другу, формируя многослойный футляр. Пробка охраняет внутренние живые ткани от потери влаги, резких температурных колебаний и проникновения микроорганизмов. Живые ткани, лежащие под пробкой, нуждаются в газообмене и удалении избытка влаги. Именно поэтому под устьицем вследствие деления субэпидермальных слоев (еще до появления перидермы), а в дальнейшем и феллогена откладываются живые, рыхло расположенные, со множеством межклетников паренхимные клетки, называемыевыполняющей тка
нью
, которая разрывает эпидерму и создает возможность газообмена и транспирации с внешней средой. Это структурное образование называют чечевичкой
(рис. 2.12).
Пробка состоит из мертвых клеток, у которых клеточная стенка пропитана жироподобным веществом суберином. Клетки расположены ровными рядами, имеют прямоугольную форму (на поперечном срезе), плотно прилегают друг к другу, формируя многослойный футляр. Пробка охраняет внутренние живые ткани от потери влаги, резких температурных колебаний и проникновения микроорганизмов. Живые ткани, лежащие под пробкой, нуждаются в газообмене и удалении избытка влаги. Именно поэтому под устьицем вследствие деления субэпидермальных слоев (еще до появления перидермы), а в дальнейшем и феллогена откладываются живые, рыхло расположенные, со множеством межклетников паренхимные клетки, называемыевыполняющей тка
нью
, которая разрывает эпидерму и создает возможность газообмена и транспирации с внешней средой. Это структурное образование называют чечевичкой
(рис. 2.12).
Рис. 2.12. Строение перидермы с чечевичкой: 1 – выполняющая ткань чечевички; 2 – остатки эпидермы; 3 – пробка (феллема); 4 – феллоген; 5 – феллодерма
Чечевички, имеющие вид небольших бугорков, отчетливо выделяются на поверхности побегов деревьев и кустарников (рис. 2.13).
На стволах березы их остатки наблюдаются в виде характерных поперечных черных полосок, у осины они принимают форму ромбов.
Третичная покровная ткань
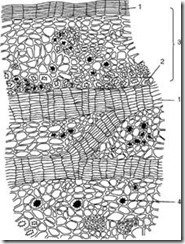 Корка (ритидом)
представляет собой третичную покровную ткань, которая образуется у многолетних растений в корне, стебле, корневище. Каждый год в более глубоких слоях закладывается новый слой феллогена и образуется перидерма. Наружный слой перидермы – пробка – изолирует все вышележащие ткани, в результате чего они отмирают. Таким образом, совокупность многочисленных перидерм с отмершими между ними тканями и есть корка (рис. 2.14).
Корка (ритидом)
представляет собой третичную покровную ткань, которая образуется у многолетних растений в корне, стебле, корневище. Каждый год в более глубоких слоях закладывается новый слой феллогена и образуется перидерма. Наружный слой перидермы – пробка – изолирует все вышележащие ткани, в результате чего они отмирают. Таким образом, совокупность многочисленных перидерм с отмершими между ними тканями и есть корка (рис. 2.14).
Рис. 2.14. Корка дуба: 1 – слои пробки; 2 – волокна; 3 – остатки первичной коры; 4 – друзы оксалата кальция