KONUŞ: Bitki hücresinin özellikleri
Toplam kimyasal bileşim neredeyse aynıdır.
Hayvan hücreleriyle benzerlikler: bir membranın, sitozolün, çekirdeğin ve spesifik organellerin varlığı.
Ancak bu ökaryot türleri arasında yapı açısından 2 farklılık vardır: spesifik organellerin varlığı ve metabolizma.
Bir bitki hücresinde var merkezi koful. Bitki hücrelerinin vakuolleri fazla suyu emer, şişer, bu da hücrenin gerilmesine yol açar, vakuol bitki hücresinin hacminin çoğunu kaplar ve bu nedenle bitki hücresinde tüm hücreden geçen yalnızca ince sitoplazma şeritleri kalır. Bu, bir bitki hücresinin boyutunu büyütmenin ekonomik bir yolu olarak hizmet eder. Bu durumda aynı hacmin protein açısından zengin sitoplazma ile doldurulmasına göre daha az enerji tüketilir.
Genç bitki hücrelerinde, hücre büyüdükçe birbirleriyle birleşen ve tüm hücrenin hacminin% 90'ını kaplayan bir veya daha fazla büyük vakuol oluşturan birkaç vakuol bulunabilir. Bu durumda organeller hücrenin çevresine ve çekirdeğe doğru itilir. Vakuolün zarı tonoplasttır. Vakuolün boşluğu hücre özsuyuyla doludur. sulu çözelti inorganik tuzlar, şekerler, organik asitler ve bunların tuzları, proteinleri, düşük molekül ağırlıklı bileşikleri.
Merkezi vakuolün ana işlevi– turgor basıncını korumak. Ozmotik konsantrasyonu koruyan vakuolün hücre özünde çözünmüş moleküller tarafından belirlenir. Konsantrasyon, vakuolün hücrenin iç basıncını koruyan, ona güç ve gerginlik veren bir yapı olarak işlev görmesini sağlar. Tonoplast, şekerlerin taşınmasını sağlayan ATP'ye bağımlı bir proton pompasına sahiptir ve ayrıca çeşitli metabolizmaların vakuollerinden atılımını (çıkarılmasını) sağlayan iyon kanalları da vardır. Hücre özsuyundaki proton konsantrasyonu, 2-5pH'lik asidik bir ortama karşılık gelir. Merkezi vakuol rezervin birikmesini sağlar besinler– şekerler ve proteinler. Bu maddelerin temini, membranların tonoplast ile etkileşime girme yeteneği ile ilişkilidir. Sindirim işlemleri hücre özsu vakuolünün içinde de gerçekleşebilir, çünkü hidrolitik enzimler içerir, tonoplast salgı kesecikleri oluştururken, vakuolden ayrılırlar, ekzosetotik keseciklerle etkileşime girerek içerdikleri bileşenlerin sindirimini sağlarlar. Ayrıca kusurlu dolumun hidrolizine katılarak otofatik döngünün reaksiyonlarını da yürütürler.
Bitki hücre duvarı.
Plazmalemmanın katılımıyla oluşturulmuştur. Esasen dış iskeleti olan hücrenin yüzeyini koruyan hücre dışı çok katmanlı bir oluşumdur. İki yapıdan oluşur: bir çerçeve (selüloz fibril ipliklerinden yapılmış) ve jel benzeri bir matris. Matris polisakkaritler içerir: hemiselülozlar ve pektin maddeleri. Hemiselülozlar, çeşitli heksozlardan (glikoz, manoz, galaktoz) oluşan dallanmış polimer zincirleridir, ayrıca pentozlar (ksiloz, arabinoz) ve üronik asitler (glukuronik ve galakturonik) de olabilir. Hemiselülozun bu bileşenleri birbirleriyle farklı kantitatif oranlarda birleşerek çeşitli kombinasyonlar oluşturur. Ancak hemiselüloz moleküllerinin zincirleri hiçbir zaman kristalleşmez ve fibrilleri sabitlenmez (tespit edilmez). Yüksek miktarda su içerir.
Pektinler. Taşıyan dallanmış, yüksek oranda hidrojenlenmiş polimerleri içeren heterojen bir grup negatif masraflar Birçok galakturonik asit kalıntısının varlığı nedeniyle.
Bu bileşenler sayesinde matris, taban (çerçeve) görevi gören yumuşak plastik bir kütledir.
Bireysel selüloz polimerleri, hidrojen bağları kullanılarak mikrofibriller halinde paketlenir. Sertlik ve güç verir. Aralarında hücre duvarını monolit haline getiren bağlar vardır. Bu tür bağlı selüloz mikrofibrilleri ayrıca pektinlerle çevrilidir. Kalsiyum ve silikon ile etkileşime girebilirler. Ona sertlik veren şey nedir? Jel benzeri yapısı nedeniyle matris, suyun ve küçük moleküllerin diffüz iletimini sağlar.
Hücre duvarları 3 tipe ayrılır: birincil, ikincil ve üçüncül.
Birincil hücrenin %90'ı karbonhidratlardan oluşur. Hücre bölünmesi sırasında, hücrelerin ekvator düzleminde, birbirinden ayrılan kromozomlar (phragmoplast) arasında yer alan bir mikrotübül demeti belirir. Mikrotübüller arasında vakuolar sistemin birçok küçük vezikülleri vardır, CG, orta kısımda fragmoplastlar birbirleriyle birleşmeye başlar. Düzleştirilmiş bir disk veya orta plakanın oluşturulması. Kalloz adı verilen bir polisakkaritten oluşur. Selülozla karşılaştırıldığında daha elastiktir (plazmalemma üzerinde oluşan CG kabarcıklarının içinde bulunmaz). CG vezikülleri, iki yavru hücrenin zarlarının yapımı için gerekli bileşenleri içerir. Küçük vakuollerin füzyon süreci hücrenin merkezinden çevreye doğru meydana gelir ve membran vezikülleri hücrenin yan yüzeyinin yüzeyi ile birleşene kadar devam eder.
Büyüyen birincil hücre duvarı halihazırda 3 katmandan oluşur (amorf bir matristen oluşan orta plaka ve hemiselüloz ve fibriller içeren 2 çevresel katman). Birincil duvar, iki yeni hücresel yapı tarafından hemiselüloz ve selüloz fibrillerinin salgılanmasıyla oluşturulur. Aradaki kalınlığı arttırmak hücre duvarı her biri, giderek daha fazla yeni katman katmanlayarak kalınlaşan hücre zarının maddelerini salgılayacak olan kardeş hücrelerin aktivitesi nedeniyle meydana gelecektir. Plazma zarında selüloz fibrilleri sentezlenir ve polimerize edilir, bunun sonucunda ikincil bir hücre duvarı oluşur (hücreye son şeklini verir). Duvarın kalınlaşması tamamlanır tamamlanmaz, üçe kadar aromatik alkol kalıntısının oksidatif polimerizasyonu yoluyla sentezlenen ve ahşabın ana bileşeni olarak görev yapan hidrofobik bir polimer olan lignin ile modifiye edilir (ahşabın %20-35'i lignindir) ). Lignin, hidrofobik yapısı nedeniyle duvarlarda su yalıtımı oluşturur ve ek bir güçlendirici malzeme görevi görür. Hücre duvarı odunsu hale gelir veya suberize olur (kütin ve suberin nedeniyle). Epidermal hücrelerde, hücre zarlarının yüzeyinde balmumu salgılanır.
Mantarlarda kitin (N-asetilglukozamin) bulunur.
Plastidler.
Tüm plastidlerin ortak özelliği:
Organellerin tüm yüzeyi boyunca yakından kapanan iki zarla çevrilidirler.
İç içerikleri stromadır. İç zarın bölümlerinin istilası ve tomurcuklanması sonucu oluşan zar disklerini içerir. Bu disklere tilakoid adı verilir ve yığınlar (granas) halinde katlanma özelliği vardır. Dördüncü tip membran olan lamel ile etkileşime girerler.
Plastidlerin kendi genomları vardır; her plastid, yaklaşık 100 gen içeren çok sayıda dairesel DNA kopyasına sahiptir. Genom, özel işlevleri yerine getirmek için gereken proteinleri kodlar.
Transkripsiyon ve translasyonda görev alan RNA proteinleri de burada bulunur. Plastidlerden kaynaklanırlar, ancak diğer proteinlerin çoğu nükleer genoma bağlıdır. Bu proteinler sitoplazmada sentezlenir ve plastid içine aktarılmalıdır.
Plastitler diğer organellere veziküler bağlarla bağlanmaz.
Plastidler bir dizi karşılıklı dönüşümü temsil eder.
Proplastid -> lökoplast -> kloroplast -> kromoplast.
Lökoplastlar şunlara dönüşebilir: amiloplastlar, elaioplastlar, aleuron taneleri ve kromoplastlar.
Kloroplast.
Uzatılmış bir şekle sahiptirler, 2-4 mikron boyutundadırlar ve uzunlukları 10 mikron olabilir. Kromatoforlar (büyük kloroplastlar).
Kloroplastın iç boşluğu lümendir. 2 önemli olay: fotosentez, enerji sentezi. Tilakoid membran, klorofilin varlığı ile karakterize edilir; güneş ışığının miktarını emebilir ve güneş enerjisini kimyasal enerjiye dönüştürebilir. Belirli bir dalga boyundaki ışığın emilmesi, klorofilde değişikliklere neden olur. Heyecanlı bir duruma girer. Aktifleştirilmiş klorofilin salınan enerjisi bir dizi ara aşamadan geçerek elektron taşıma zincirine aktarılır, bu da ATP'nin sentezine ve 2 mononükleotid (NAD ve FAD) olan taşıyıcının restorasyonuna yol açar. Enerji, karbondioksit sentezi ve şeker sentezi için harcanır.
Fotosentez: aydınlık ve karanlık faz.
Işık fazı yalnızca ışıkta meydana gelir ve ışığın pigment tarafından emilmesi ve kimyasal bir reaksiyonun (Hill reaksiyonu) yürütülmesi ile ilişkilidir.
Fotosentezin ışık aşamasında güneş ışığının enerjisi klorofilin elektronlarını uyarır. Elektronların mitokondrideki solunum zinciri boyunca taşınmasına benzer şekilde, tilakoid membrandaki oksidatif zincirin bileşenleri boyunca taşınırlar. Kloroplastlarda protonlar stromadan (pH = 8) lümene (pH = 5) pompalanır ve 3 pH birimlik bir gradyan oluşturulur. Bu proton gradyanı, proton zarında 200 mW'lık bir proton hareket kuvveti yaratır, ancak bu neredeyse tamamen pH gradyanından kaynaklanmaktadır. Mitokondriyal solunum zinciriyle karşılaştırıldığında elektronlar farklı yönde hareket eder. Elektronlar fotolizi sırasında sudan alınır ve enerji harcanarak taşıyıcıya aktarılır. O. ışık fazında elimizde: sentezlenmiş ATP molekülleri ve indirgenmiş bir taşıyıcı bulunur. Her iki ürün de karanlık aşamada kullanılır.
Karanlık aşamada, NAD ve ATP'nin azalması nedeniyle karbonhidratlar oluşur. Bu süreç çok aşamalıdır. İçerisinde çok sayıda enzim yer alır. Calvin döngüsü. Öncü yapılardan (proplastidler) kloroplast sayısında bir artış ve diğer plastid formlarının oluşumu meydana gelir.
Proplastidler, ayırt edici özellikleri olmayan küçük çift membranlı keseciklerdir. Daha yoğun içerikleri ve iki zarın varlığı bakımından sitoplazmik vakuollerden farklıdırlar. Proplastidler bitki dokularının bölünmesinde bulunur. Proplastidlerin sayısı bölünme ve tomurcuklanma yoluyla artar. Normal aydınlatma altında proplastidler kloroplastlara dönüşür (boyut artar, uzunlamasına yerleştirilmiş membran kıvrımlarının oluşumu meydana gelir. Bazıları lamel oluşturur, diğerleri - tilakoidler). Karanlıkta gelişirlerse, önce plastidlerin hacminde bir artış olur, ancak bir iç zar sistemi oluşmaz; ayrı bölgelerde biriken, karmaşık kafes yapıları - prolamellar gövdeler oluşturan bir küçük kesecik kütlesi oluşur. (etioplastlar). Membranları protoklorofil (sarı) içerir; ışığın etkisi altında onlardan kloroplastlar oluşur ve protoklorofil gerçek klorofil haline gelebilir (transfer zincirinin bileşenleri olan membranlar sentezlenir).
Lökoplastlar. Kloroplastlardan farklıdır. Depo doku hücrelerinde bulunurlar. Belirsiz morfolojileri onları proplastidlerden ayırmayı zorlaştırır. Lameller olarak görülebilirler ancak normal tilakoid yapılar oluşturma kapasitesine sahiptirler. Karanlıkta çeşitli rezerv maddeleri biriktirirler. Eğer bir karbonhidrat ise, o zaman lökoplastlar amiloplastlara dönüşür (alöron taneleri şeklinde proteinler de olabilir). Ve eğer yağlar elaioplast ise.
Yaprakların gelişimi ve meyvelerin olgunlaşması sırasında kloroplastlardaki renk değişimi ve değişim sürecini gözlemliyoruz. Plastidlerde ek membranlar birikir, klorofil ve nişasta yok edilir. Lameller yok edildiğinde, pigmentlerin (karotenoidlerin) iyice çözündüğü ve renk değişikliğinin ortaya çıktığı lipit damlacıkları salınır. Kromoplastlar, lipoprotein komplekslerinin parçalanmasıyla ilişkili olan dejenere bir plastid formudur.
Plastid genomu.
Kendi genetik sistemi, plastidlerin içinde lokalize olan proteinlerin (tilakoid membran proteinleri) sentezini sağlar. 60 mikrona kadar uzunlukları vardır. 1,3*10 8 Evet. Döngü süresi ve çoğaltma hızı eşleşmiyor. DNA'nın sabitleyici yapısı yoktur. Kloroplastların halka molekülünün tüm özellikleri prokaryotlarınkine yakındır. Kloroplast DNA'sının benzerliği, transkripsiyonun temel düzenleyici işlemlerinin aynı olmasıyla ortaya çıkar. Her türlü RNA'nın sentezi. Kloroplast DNA, ribozomal RNA'yı kodlar. 70S ribozom segmentasyon sabitleri. 17S ve 23S içerirler. Antibiyotiğe (kloramfenikol) duyarlıdır.
KONUŞ: Hücreler arası temaslar
Simbiyotik teori.
Komintsin ve Mereshkovsky.
Bu teori, kloroplastların ve mavi-yeşil alglerin yapısındaki şaşırtıcı benzerlikle desteklenmektedir. Ana işlevleriyle benzerlik, fotosentetik süreçler için neredeyse aynı yetenekler. Çeşitli prokaryotik hücrelerin simbiyozunun bir sonucu olarak nükleer (ökaryotik) hücreler ortaya çıktı. Bu hipoteze göre, ökaryotik bir hücre, gelişiminde diğer hücrelerle çeşitli simbiyoz aşamalarından geçmiştir.
İlk aşamada, anaerobik heterotrofik prokaryotlar gibi hücreler aerobik bakteriler için konakçı haline geldi ve bu da aerobik prokaryotların (bu prokaryotik organizmalar içindeki mitokondri) oluşumuna yol açtı. Bu sürece paralel olarak konakçı hücrede prokaryotik nükleer aparat (genofor), sitoplazmadan izole edilmiş bir çekirdek halinde oluşturulur. Heterotrofik ökaryotik hücreler bu şekilde ortaya çıkabilir. Böyle bir hücre tekrarlanan simbiyoza girer. Örneğin spiroketler veya flagella benzeri bakterilerle. Bu simbiyontların dahil edilmesi, heterotrofik ökaryotik hücrelerde flagella ve siliaların ortaya çıkmasına yol açtı. Ve ek emilimin bir sonucu olarak. Bu, kamçılı ökaryotik hayvan hücrelerinin ortaya çıkmasına yol açtı. Böyle bir organizma aynı zamanda mavi-yeşil algleri de absorbe edebiliyordu ve bu da plastidlerin oluşmasına ve bitki hücresinin oluşmasına yol açıyordu. Zar içi organellerin endosimbiyotik kökenine ilişkin hipotez şüphe götürmez. Mavi-yeşil algler ve hücrelerin gerçek endosembiyozuna ilişkin çok sayıda gerçek bilinmektedir. alt bitkiler ve ortakyaşarların mükemmel bir şekilde çalıştığı ve konakçı hücreye fotosentetik ürünler sağladığı protozoa. Örneğin. Bir terlik siliatı, alg chlorella'yı içerir. Kloroplastlar bazı hücreler tarafından seçilebilir ve endosembiyoz ortakları olarak kullanılabilir. Rotiferler alglerle beslenir. Kloroplastlar sindirim bezlerinin hücrelerinin içinde kendilerine yer bulurlar ve işlevlerini yerine getirmeye devam ederler.
Bitki hücrelerinin fonksiyonlarının özellikleri. Organik maddelerin sentez yöntemi ototrofik beslenmedir.
Bitki hücreleri, hücre duvarının varlığı nedeniyle bitki hücresinin şekil değiştirmesine ve hareket etmesine izin vermez. Bitki hücreleri ise hareket eksikliğini kendi besinlerini sentezleyerek telafi eder. ototroflardır. Bitkiler yaşamları boyunca büyürler, meristemler sürekli bölünerek yeni hücreler oluştururlar; tüm bitkiler modüler organizmalara aittir. Nihai yaşam formları çevrelerine bağlıdır. Bitki hücrelerinin mitoz sırasında da özellikleri vardır:
Yeni hücre duvarının konumu. İş milinin oluşumundan çok önce belirlenirken, iş milinin kendisi bu hücrenin konumuna katılmaz. Mitozdan önce bile hücre döngüsünün G2 fazında bitki hücresinde preprofaz halkası adı verilen spesifik bir yapı oluşur. Hücreyi çevreleyen geniş bir şerit şeklinde korteksin (hücre iskeleti) mikrotübüllerinden oluşur. Çekirdek sağlam kalır (katı). Bu halka bölgesinde, çekirdeğin etrafında iyi oluşturulmuş yoğun bir mikrotübül demetine dönüşene kadar daralmasını sağlayan aktin filamentleri toplanır. Ortaya çıkan yapı, jantın ve jant tellerinin mikrotübüller ve aktin'den oluştuğu ve çekirdeğin göbeği oluşturduğu bir tekerleği andırıyor. Yüzüğün yapısı EPR ve KG elemanları ile zenginleştirilmiştir. Mitozun erken evresinde preprofaz halkasının mikrotübülleri depolimerize olur ve yok edilir. Telofazda ise tam olarak aynı noktada yeni bir hücre duvarı oluşur. o uçak
, ön-faz halkası tarafından belirlenmiştir.
Bitkilerde mitotik milin kutupları sentriol içermez ve doğası gereği daha dağınıktır. Başka bir deyişle bulanık görünüyorlar. Başlangıçta mikrotübüllerden bir profaz mili oluşur. Ondan, nükleer membranın tahrip edilmesinden sonra gerçek bir iğ oluşur ve bu gerçek mitotik milin oluşmasıyla hücrede başka sitoplazmik mikrotübül kalmaz. Bu iş milini kontrol eden nedir, nasıl oluşur? Kromozomlar.
İlk olarak, mikrotübüllerin polimerizasyonu, rastgele yönlenen kromozomların yakınında meydana gelir. Daha sonra motor proteinleri (dynein) mikrotübülleri sıralar, pozitif uçları kromozomlara gönderilir, bu da onların kromozomların kinetokorlarına bağlanmalarını sağlar ve diğerleri mikrotübüllerin negatif uçlarıyla etkileşime girer. Polimerizasyon, sınıflandırma ve bağlanma sonucunda iki kutuplu bir mil oluşur. Nükleer bölünmeden sonra sitoplazmik bölünme meydana gelir. Bu, konumu ön-faz halkası tarafından belirlenen yeni bir hücre septumunun oluşma anına denk gelir. İğ yönelimi ile yeni septum arasındaki ilişki, hayvan hücrelerinde olduğu gibi sıkı değildir.
Bitki hücrelerinin Golgi kompleksi. Fonksiyonları: glikosilasyon (birincil), sıralama (ayırma), hücre duvarı polisakkaritlerinin sentezi (selüloz ve kalloz hariç), enzim komplekslerinin (altıgen rozetler, selüloz sentezi) membrana iletilmesi, veziküllerin vakuollere taşınmasını yönlendirir.
KG, çekirdekten uzakta, plazmaya daha yakın bitki hücrelerinde lokalizedir ve çok sayıda tank yığını şeklinde dağılmıştır. Kompleksin tüm elemanları, hücre içi taşımayı sağlayan bir ağ oluşturan aktin filamentleriyle bağlanır. Aktin yapıları hareketliliği sağlar (miyozin ile birlikte). Bitki hücrelerinde özel bir miyozin - miyozin 8 bulunur. Bu miyozin 8, filamentler oluşturmaz ve yeni oluşan birincil duvarlarda birikir; CG vezikülleri, ikincil hücre duvarını oluşturan polisakkaritler ve yapısal proteinler içerir.
Hücreler arası temaslar.
Bunların birkaç sınıflandırması vardır, farklı yapılandırılmışlardır. Alberts ve Gray (1986) sınıflandırması. Buna göre: kişiler 3 gruba ayrılabilir.
Mekanik (= Yapışkan). Hücre zarında hücre dışı matriks glikoproteinlerinin varlığı nedeniyle hücrelerin birbirine yapışma yeteneği ile ilişkilidir. Plazma zarları arasında yapışma meydana geldiğinde, her zaman glikokaliks ile dolu bir boşluk (20 nm) kalır. Özel CAM proteinleri (cam), hücrelerin birbirine bağlanmasından doğrudan sorumludur: fibronektin, kadherinler, selektinler, integrinler ve diğer proteinler. Bazıları hücreleri moleküller arası etkileşimler yoluyla birbirine bağlar. Diğerleri özel hücreler arası bağlantılar oluşturur. Bu proteinler arasındaki etkileşimler homofilik olabilir, yani. bu durumda komşu hücreler homojen moleküller kullanarak birbirleriyle iletişim kurarlar. Veya yapışmada yer aldıklarında heterofilik çeşitli türler Komşu hücrelerdeki proteinler. Daha fazlası var karmaşık bağlantılar Hücreler arası bağlanma, bağlayıcı ara moleküller tarafından sağlandığında.
Bunlara ek olarak
Kuşak desmozomları. Komşu hücreler arasındaki mesafe 15-20 nm'dir. Orta plaka o kadar parlak değil. Ancak birbirine yapışan ve zarların mekanik bağlantısını sağlayan zar ötesi glikoproteinler vardır. İnce aktin filamentleri (6-7 nm) sitoplazmik tarafta birikir. Plazmalemma boyunca uzanırlar ve hücrenin tüm çevresi boyunca uzanırlar. Fonksiyon: hücrelerin birbirine mekanik olarak yapışması; Aktin filamentleri kasıldığında hücrenin şekli değişebilir; hücre geometrisinde değişikliklere neden olabilir; Mekanik stresi hücreye iletir. Hücre hacmindeki değişiklikten kaynaklanır.
Hemidesmozomlar. Hücrelerin hücre dışı matrisle bağlantısı (Örneğin, bazal membran ile). Fonksiyonel rol, epitel katmanlarının büyük mekanik darbelere dayanmasına izin veren mekaniktir. Dezmozomlar hücrelerin parçalarıdır ancak statik oluşumlar değildir. Dinamiktirler. Kaybolma ve yeniden ortaya çıkma yeteneğine sahiptir. Fagosantral bir reaksiyonla yok edilirler.
Kapanış (sıkı temaslar). Tek katmanlı epitelin karakteristiğidir. İki plazma zarının dış katmanlarının mümkün olduğu kadar yakın olduğu bölge. Bu nedenle elektron mikroskobuyla bakıldığında üç katmanlı bir yapı görülür. Her iki zarın 2 dış katmanı, 2-3 nm kalınlığında tek bir tabaka halinde birleşir. Sıkı temasın tüm alanı boyunca değil, integral membran proteinlerinin bir dizi nokta temasını temsil eder. Sıkı bağlantı bölgesinde 24'ten fazla protein türü bulunmuştur. Bilipid tabakasının zarlarından çıkıntı yapan bu tür proteinler kesişir ve bir ağ (kafes) oluşturur. Sıkı bağlantı tarafında, sitoplazmanın kendisinde, plazmalemmanın yüzeyine paralel olarak yerleştirilmiş çok sayıda fibril (7 nm) vardır. Sıkı bağlantılar glandüler ve bağırsak epitelinin karakteristiğidir. Hücreleri birbirine bağlarlar.Üstelik bu moleküler yapılar hücre içi taşımanın düzenlenmesi, yani Maddelerin hücreler arasındaki boşlukta taşınması.
Sıkı bir temastaki geçirgenlik bariyeri, fibriler iplik benzeri elemanlardan oluşan bir ağ yapısı oluşturan yük taşıyan katmanlar tarafından oluşturulur. İyonlar bu tür gözeneklerden geçebilir; boyutları küçüktür. Ancak çözünebilen bileşenlerin geçişi için bu ipliklerin bütünlüğünün bozulması gerekmektedir. Çözünebilen elementler bu iplikleri koparır ve moleküllerin geçişi sonrasında tekrar bir araya gelirler. Ve böylece molekül temas bariyerinden geçer. Yapısal ve işlevsel olarak sıkı bağlantılar, plazmalemmayı 2 bölüme (apikal ve bazal) ayırır ve sıkı bağlantı, moleküler bileşimin sınırlar içinde korunmasını sağlayan bir septum görevi görür.
İletken kontaklar. Hayvanlarda ve bitkilerde farklılık gösterir.
2 tür hayvan hücresi vardır: boşluk ve kimyasal sentez. Boşluk bağlantıları aracılığıyla moleküller bir hücreden diğerine geçebilir. Ancak kimyasal sentezde hücrelerin doğrudan bir bağlantısı yoktur. Boşluk bağlantıları hücrelerin iletişim bağlantıları olarak kabul edilir. Bu kimyasalların doğrudan transferinde rol oynayan bir yapıdır. Bu tipin karakteristik bir teması, iki komşu hücrenin plazma zarlarının 2-3 nm mesafeye kadar bir araya getirilmesidir. Boşluk temasının kendisi 0,5-5 µm boyutundadır. 2 yarıdan oluşur, her yarının boyutu 7-8 nm'dir, 1-2 nm genişliğinde bir kanalın etrafında bulunurlar. Ve bu 2 yarıya connexons (yarım kanallar) denir. 6 adet bağlayıcı protein alt biriminden oluşurlar. Proteinin moleküler ağırlığı yaklaşık 30 bindir. Proteinler birbirleriyle birleşerek silindirik bir yapı oluşturur ve iki hücrede konneksonların boyutu, sayısı, boyutu ve düzeni tam olarak simetriktir. Boşluk temasları, kütlesi 2000 Dalton'a ulaşan moleküller hariç, 1200 Dalton ağırlığındaki moleküllerin serbest difüzyonuna izin verir. Boşluk bağlantılarının geçirgenliği kanal kapılarının açılıp kapanmasıyla kontrol edilir. Bu süreç denir geçit (kapı mekanizması). Hücre içi pH, kalsiyum iyon akışı veya konneksin protein alt birimlerinin doğrudan fosforilasyonundaki değişikliklerle kontrol edilir. Boşluk kontaklarının işlevleri: organik ve inorganik maddeler. (Örneğin miyokard kası hücrelerinde).
Plasmodesmata bitkilerde bir tür iletken temastır. Bunlar 2 komşu hücreyi birbirine bağlayan ince tübüler sitoplazmik kanallardır. Bu kanalların çapı 20 ila 40 nm arasında değişmektedir. Bu kanalların özellikleri: plazma zarı bir hücreden başka bir hücrenin plazmalemmasına geçer. Bu kanalın ekseni boyunca desmotulün silindirik bir tüpü bir hücreden diğerine uzanır. Lümeni ER hücreleriyle iletişim kurar. Desmotubül ile plazmodesmata membranı arasındaki boşluk sitozol ile doludur. Bu hücreleri çevreleyen plazmalemma, plazmodesmata ve diğer hücrelerin zarına sorunsuz bir şekilde geçer. Sonuç olarak, komşu hücrelerin hiyaloplazması tek bir sisteme (semplast) bağlanır. Ve resmi olarak hiçbir ayrım yoktur (bir hücrenin diğerinden). Plasmodesmata yoluyla serbest taşıma, 800 Dalton'luk bir moleküler ağırlıkla sınırlıdır. Plasmodesmata, ER elementlerinden medyan plakanın birincil oluşumu aşamasında oluşur. Yeni bölünen bir hücrede plazmodezma sayısı hücre başına 1000'e kadar ulaşır. Yaşlanmayla birlikte sayıları azalır ancak kalınlıkları artar. Fonksiyonel rol: çözeltilerin (iyonlar, şekerler, besinler) hücreler arası dolaşımını sağlamak. Olumsuz tarafı ise hastalanmanın daha kolay olmasıdır.
BİTKİ HÜCRELERİNİN KOKULARI
Organoidler genel anlam Tek membranlı yapı prensibine sahip olan ve meydana gelen sadece bitki hücrelerinde.
Boyut ve Adet: Hücrenin yaşına göre belirlenir. Genç hücrelerde, EPS'den kopan küçük keseciklerden kofullar ortaya çıkar. Hücreler büyüyüp farklılaştıkça, küçük vakuoller birbirleriyle birleşerek bir veya daha fazla büyük vakuol oluştururlar ve tüm hücrenin hacminin %80'ini kaplarlar. Sonuç olarak, çekirdek ve organellerin bulunduğu sitoplazma çevreye doğru itilir.
Yapı: merkezi vakuoller sitoplazmadan tek bir zarla ayrılır - tonoplast kalınlığı plazmalemmaya benzer. Vakuol boşluğu doldurulur hücre özü. Hücre özsuyu şunları içerir: inorganik tuzlarşekerler, organik asitler ve bunların tuzları, diğer düşük molekül ağırlıklı maddeler ve ayrıca bazı yüksek molekül ağırlıklı bileşikler (örneğin proteinler).
İşlevler: 1) osmoregülasyon: tonoplast ve plazmalemmanın yarı geçirgenliği nedeniyle hücre özsuyunun karşılık gelen moleküler konsantrasyonu korunur, yani. vakuol bir osmometre görevi görür;
2) boşaltım: suda çözünebilen tüm metabolik ürünler (alkaloidler - nikotin, kafein; polifenoller) tonoplast yoluyla uzaklaştırılabilir;
3) depolama: fosfatlar K + , Na + , Ca 2+ , organik asit tuzları (oksalatlar, sitratlar vb.), şekerler ve proteinler hücre özsuyunda birikir.
RİBozomlar
Zar yapısına sahip olmayan bir organel. Hem prokaryot hem de ökaryot hücrelerinde bulunan genel öneme sahip tek organeldir. Ribozomlar ilk kez tanımlandı 1955 G. J.Palade (Palade granülleri), bunların ribonükleoprotein kompleksleri (RNP) olduğunu kanıtlamıştır.
Kimyasal yapısı: RNP= r-RNA + protein.
Ribozomlar sorumludur %85 RNA'sı, bir hücrede sunulur.
Biçim: Ribozom iki alt birimden oluştuğu için mantar şekline sahiptir: büyük Ve küçük, aralarında yer alır ribozomun fonksiyonel merkezi (FCR) protein biyosentezi sırasında (translasyon süresi), mRNA'nın iki üçlüsünde bulunduğu ve bir enzimatik kompleksin çalıştığı, bir protein molekülünün amino asitlerden birleştirilmesini sağlayan.
Boyutlar: 15 – 35 nanometre. Prokaryotik hücrelerde tam bir ribozomun boyutu 20x17x17 nm, ökaryotik hücrelerde - 25x20x20 nm'dir.
Eğitim yeri: ribozomal alt birimlerin oluşumu çekirdeğin nükleollerinde meydana gelir. Alt birimlerin tam bir ribozom halinde toplanması, magnezyum iyonlarının (Mg2+) konsantrasyonu 0,001 M'ye ulaştığında sitoplazmada meydana gelir; bu konsantrasyon azalırsa, alt birimlerin ayrışması meydana gelir. Mg2+ konsantrasyonu on kat artarak 0,01 M değerine ulaştığında, iki ribozom birbirleriyle etkileşime girerek bir dimer oluşturur.
Kofullar ve hücre özü
Çoğu olgun bitki hücresi, hücre hacminin %70-90'ını kaplayan büyük bir merkezi vakuol ile karakterize edilir. Bu durumda protoplast, tüm organelleriyle birlikte hücre duvarını kaplayan çok ince bir duvar tabakası şeklinde bulunur. Küçük sitoplazmik vakuoller genellikle duvar protoplastında bulunur. Bazen çekirdek hücrenin merkezinde bulunur. nükleer cep sitoplazma, merkezi vakuolden geçen en ince sitoplazmik şeritlerle duvar katmanına bağlanan sitoplazma.
Hücre özsuyu sulu bir çözeltidir çeşitli maddeler Protoplastın hayati aktivitesinin ürünleri olan, esas olarak rezerv maddeleri ve atıklar. Hücre özsuyunun reaksiyonu genellikle hafif asidik veya nötr, daha az sıklıkla alkalidir. Hücre özsuyunu oluşturan maddeler son derece çeşitlidir. Bunlar karbonhidratlar, proteinler, organik asitler ve bunların tuzları, amino asitler, mineral iyonları, alkaloidler, glikozitler, tanenler, pigmentler ve diğer suda çözünen bileşiklerdir. Bunların çoğu, vücutta görünüp kaybolabilen protoplast metabolizmasının ürünleri olan ergastik maddeler grubuna aittir. farklı dönemler hücre yaşamı. Birçok hücre özsuyu maddesi yalnızca bitki hücrelerinde oluşur.
Bitki hücresi özsuyundaki karbonhidratlar, monosakkaritler (glikoz, fruktoz), disakkaritler (sakkaroz) ve polisakkaritler (mukus, inülin) ile temsil edilir.
Glikoz (üzüm şekeri) ve fruktoz (meyve şekeri) vücutta birikir. büyük miktarlar sulu meyvelerde. Sükroz (pancar şekeri), şeker pancarı köklerinde ve şeker kamışı saplarında büyük miktarlarda birikir. Bazı bitki familyaları (cactaceae, Crassulaceae, orkideler), suyu tutan hücre özsuyunda mukus birikmesiyle karakterize edilir. İnülin, Asteraceae'nin yeraltı organlarının hücre özsuyunda nişasta yerine kolloidal bir çözelti olarak biriken bir rezerv polisakkarittir.
Proteinler, olgunlaşan tohumların hücrelerinin boşluklarında kolloidal bir çözelti halinde birikir. Gelişimlerinin ilerleyen aşamalarında tohumlar susuz kaldığında, boşluklardan su uzaklaştırılır, hücre özsuyundaki protein konsantrasyonu artar ve katı jel haline dönüşür. Olgun tohumların kurutulmuş kofullarına denir aleuron taneleri.
Hücre özsuyundaki organik asitlerden en yaygın olanları sitrik, malik, süksinik ve oksaliktir. Bu asitler olgunlaşmamış meyvelerin hücre özsuyunda büyük miktarlarda bulunur ve onlara ekşi bir tat verir. Meyveler olgunlaştığında, organik asitler solunum substratları olarak kullanılabilir, böylece meyvenin ekşi tadı genellikle kaybolur. Organik asitlerin tuzları mineral iyonlarıyla birlikte ozmotik süreçlerde önemli bir rol oynar.
Tanenler (tanenler), buruk bir tada sahip polimerik fenolik bileşiklerdir. Antiseptik özelliklere sahiptirler ve bitki dokularını enfeksiyonlardan ve çürümeden korurlar. Gövde ve kök kabuğunun hücreleri (meşe, söğüt), olgunlaşmamış meyveler (ceviz), yapraklar (çay) ve bazı patolojik büyümeler - safralar özellikle tanenler açısından zengindir. Tanitler tıpta, deri tabaklamada ve kumaşları koyu kahverengiye boyamada kullanılır.
Alkaloidler – çeşitli kimyasal olarak Acı bir tada sahip, nitrojen içeren organik maddeler. Baz özelliklerine sahiptirler ve hücre özsuyunda genellikle tuz formunda bulunurlar. Alkaloit taşıyan bitkilerin çoğu zehirlidir ve otçullar tarafından yenmez. Alkaloid içeren hücrelerde mikroorganizmaların sporları ve mikropları gelişmez ve bitkiler mantar ve bakteri hastalıklarından etkilenmez. Solanaceae, Poppy, Rubiaceae, Ranunculaceae vb. familyalarının temsilcileri özellikle alkaloidler açısından zengindir.
Glikozitler – geniş bir grup doğal maddelerşekerlerin alkoller, aldehitler, fenoller ve diğer maddelerle bileşikleri. Tıpta bir takım bitki glikozitleri kullanılmaktadır. Glikozitler ayrıca hücre özsuyu pigmentlerini de içerir. flavonoidler. Bunlardan bazıları antosiyaninler– hücre özsuyunu kırmızı, mavi veya mor; diğer - flavonlar- sarı. Antosiyaninler birçok bitkinin çiçek rengiyle ilişkilidir. Renk şeması hücre özsuyunun reaksiyonu ile belirlenir: asidikse kırmızı tonlar, nötr ise mor tonlar hakimdir ve hafif alkali bir reaksiyonla mavi tonlar hakimdir. Tonların görünümü ayrıca çeşitli metallerle antosiyanin komplekslerinin oluşumundan da etkilenir. Flavonlar belirler sarı birçok bitkinin yaprakları.
Hücre metabolizmasındaki organik asitlerin, tanenlerin, alkaloitlerin ve hücre özsuyundaki glikozitlerin önemi yeterince açıklanamamıştır. Daha önce bunlar değişimin nihai ürünleri olarak görülüyordu. Birçoğunun metabolik süreçlere yeniden dahil olabileceği ve bu nedenle yedek maddeler olarak değerlendirilebileceği artık gösterilmiştir.
Bitki hücrelerindeki kofullar, rezerv madde ve atık biriktirme görevinin yanı sıra başka bir görevi daha yerine getirir. önemli işlev– turgorun sürdürülmesi. Merkezi vakuolün hücre özsuyundaki iyon ve şeker konsantrasyonu genellikle hücre duvarındakinden daha yüksektir; Tonoplast, bu maddelerin vakuolden difüzyonunu önemli ölçüde yavaşlatır ve aynı zamanda suya kolayca geçirgendir. Bu nedenle vakuole su girecektir. Seçici geçirgen bir zardan suyun bu tek yönlü difüzyon sürecine denir. osmoz. Hücre özsuyuna giren su, duvar protoplastına ve onun aracılığıyla hücre duvarına baskı uygulayarak gergin, elastik hale gelmesine veya turgor hücreler. Turgor, odunsu olmayan bitki organlarının uzaydaki şekil ve konumlarını korumalarının yanı sıra mekanik etkenlere karşı dayanıklılıklarını da sağlar.
Bir hücre, toksik olmayan bir tuz veya şekerin hipertonik bir çözeltisine (yani, hücre özsuyu konsantrasyonundan daha yüksek bir konsantrasyona sahip bir çözelti) yerleştirilirse, o zaman vakuolden ozmotik bir su salınımı meydana gelir. Bunun sonucunda hacmi azalır, elastik duvar protoplastı hücre duvarından uzaklaşır, turgor kaybolur ve plazmoliz hücreler ( Şekil 2.9).
Pirinç. 2.9. Plazmoliz şeması: 1 – turgor durumundaki hücre; 2 – plazmoliz başlangıcı; 3 – tam plazmoliz.
Plazmoliz genellikle geri dönüşümlüdür. Bir hücre suya veya hipotonik bir çözeltiye yerleştirildiğinde, su merkezi vakuol tarafından tekrar kuvvetli bir şekilde emilir, protoplast tekrar hücre duvarına doğru bastırılır ve turgor yenilenir. Plazmoliz, bir hücrenin yaşam durumunun bir göstergesi olarak hizmet edebilir; ölü bir hücre, seçici olarak geçirgen zarlara sahip olmadığı için plazmolize edilmez.
Turgor kaybı bitkinin solmasına neden olur. Yetersiz su koşullarında havada soldurulurken ince hücre duvarları protoplastla aynı anda büzülür ve katlanır.
Turgor basıncı sadece odunsu olmayan bitki parçalarının şeklini korumakla kalmaz, aynı zamanda hücre büyüme faktörlerinden biridir. yükseklik hücreler germe yani suyun emilmesi ve vakuol boyutunun artması nedeniyle. Hayvan hücrelerinin merkezi bir vakuolü yoktur; büyümeleri esas olarak sitoplazma miktarındaki artışa bağlı olarak meydana gelir, bu nedenle hayvan hücrelerinin boyutu genellikle bitki hücrelerinden daha küçüktür.
Merkezi vakuol, meristematik (embriyonik) hücrelerde bulunan çok sayıda küçük vakuolün füzyonundan kaynaklanır. Bu sitoplazmik vakuollerin membranlardan oluştuğu düşünülmektedir. endoplazmik retikulum veya Golgi aygıtı.
Kapsamalar
Kapanımların oluşumu, hücrenin belirli bölgelerinde - vakuolde, hiyaloplazmada, çeşitli organellerde ve daha az sıklıkla hücre duvarında - bazı metabolik ürünlerin aşırı birikmesinden kaynaklanır. Bu maddeler sıklıkla amorf veya kristal formda çökerler. kapanımlar. Kapanımlar var belli bir biçim ve ışık mikroskobu altında açıkça görülebilir. Belirli kapanımların varlığı, bunların şekli ve dağılımı ile bir bitki türü, cinsi ve familyası diğerlerinden ayırt edilebilir, bu nedenle genellikle önemli olarak hizmet ederler. teşhis işareti Tıbbi bitki materyallerini analiz ederken.
Kapsamalar ya yedek maddeler(geçici olarak metabolizmadan uzaklaştırılan bileşikler) veya nihai ürünler değişme. İlk katılım kategorisi şunları içerir: nişasta taneleri,lipit damlaları Ve protein birikintileri; ikinciye - kristaller bazı maddeler.
Nişasta taneleri– bitki hücrelerinin en yaygın kalıntıları. Polisakkarit nişasta– bitkiler için ana rezerv besin türü. O aynı zamanda en önemli bağlantı otçullar tarafından yiyecek olarak kullanılır. Tahıl tanelerinden, patates yumrularından ve muz meyvelerinden elde edilen nişasta insan beslenmesinin en önemli kaynağıdır. Buğday unu neredeyse %75 oranında nişasta tanelerinden oluşur; patates yumrularında ise nişasta %20-30 oranında bulunur. Kimyasal olarak nişasta bir alfa-1,4-D-glukandır, moleküller dallanmış zincirler biçimindedir, nişasta tanesinde yarıçaplar boyunca bulunurlar.
Plastidlerin stromasında nişasta taneleri oluşur. Kloroplastlarda taneler ışık altında depolanır özümleyici (birincil) nişasta, aşırı miktarda şeker olduğunda oluşur - fotosentez ürünleri. Ozmotik olarak aktif olmayan nişastanın oluşumu, kloroplasttaki ozmotik basıncın artmasını önler. Fotosentezin gerçekleşmediği geceleri, asimilatif nişasta enzimler yardımıyla şekerlere hidrolize edilir ve bitkinin diğer kısımlarına taşınır. Yedek (ikincil) nişasta Hücre amiloplastlarında depolanır çeşitli organlar bitkiler (kökler, yeraltı sürgünleri, tohumlar) fotosentetik hücrelerden akan şekerlerden elde edilir. Gerektiğinde depolanan nişasta da şekere dönüştürülür.
Nişasta tanelerinin oluşumu, plastid adı verilen stromanın belirli noktalarında başlar. eğitim merkezleri. Tahıl büyümesi, etrafındaki nişasta katmanlarının art arda birikmesiyle oluşur. eğitim merkezi. Bir tanecikteki bitişik katmanlar, farklı göstergeışığın kırılması ve daha sonra mikroskop altında görülebilmeleri - katmanlı nişasta taneleri. Katmanların düzeni şunlar olabilir: eşmerkezli(buğday) veya eksantrik(patates) ( pirinç. 2.10). Amiloplastta, çevresinde nişasta katmanlarının biriktiği bir eğitim merkezi varsa, o zaman basit iki veya daha fazla ise oluşur karmaşık tahıl birkaç basit olandan oluşur. yarı bileşik tahıl nişasta ilk önce birkaç nokta etrafında biriktirildiğinde oluşur ve daha sonra basit tanelerin temasından sonra etraflarında ortak katmanlar belirir ( pirinç. 2.10).
Nişasta tanelerinin şekli, boyutu, amiloplasttaki sayısı ve yapısı (eğitim merkezinin konumu, katmanlanması, çatlakların varlığı veya yokluğu) genellikle bitki türüne özeldir. pirinç. 2.10). Tipik olarak nişasta taneleri küresel, oval veya mercek şeklindedir, ancak patateslerde düzensizdirler. En büyük taneler (100 mikrona kadar) patates yumru hücrelerinin karakteristiğidir; buğday tanelerinde iki boyuttadırlar - küçük (2-9 mikron) ve daha büyük (30-45 mikron). Mısır tanesi hücreleri küçük taneler (5-30 mikron) ile karakterize edilir. Pirinç, yulaf ve karabuğday karmaşık nişasta tanelerine sahiptir.
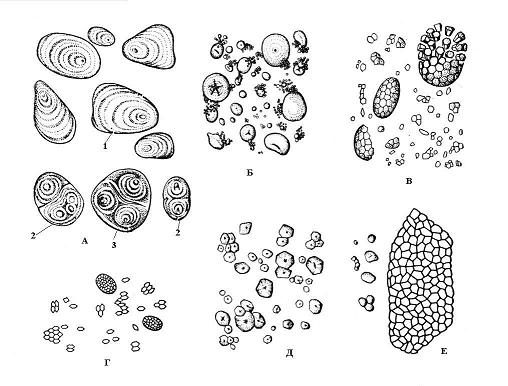
Pirinç. 2.10. Nişasta taneleri çeşitli türler bitkiler: A – patates; B – buğday; B – yulaf; G – pirinç; D – mısır; E – karabuğday; 1 – basit tane; 2 – karmaşık tane; 3 – yarı bileşik tane.
Nişasta reaktifi, bir potasyum iyodür çözeltisi içindeki bir iyot çözeltisidir - Lugol reaktifi. Nişasta tanelerini mavi-mor renklendirir.
Nişasta birikintileri tüm bitki organlarında yaygındır, ancak tohumlar, yeraltı sürgünleri (yumrular, soğanlar, rizomlar) ve odunsu bitkilerin köklerinin ve gövdelerinin iletken dokularının parankimi özellikle zengindir.
Lipid damlaları hemen hemen tüm bitki hücrelerinde bulunur. Yağlı yağlar çok sayıda bitkide birikir ve nişastadan sonra ikinci en önemli yedek besin formudur. Tohumlar ve meyveler özellikle bunlar açısından zengindir. Bazı bitkilerin tohumları (ayçiçeği, pamuk, yer fıstığı) kuru madde ağırlığının %40'ına kadar yağ içerebilir. Bu nedenle bitkisel yağlar esas olarak tohumlardan elde edilir.
Lipid damlacıkları doğrudan hyaloplazmada birikir. Küçük küresel gövdelere benziyorlar, her damla hiyaloplazmadan bir zarla ayrılıyor. Bazen lipid damlalarına denir sferozomlar.
Yağlı yağın reaktifi bir boyadır Sudan III Lipid damlacıkları turuncu-kırmızı renklidir.
Protein kapanımlarıçeşitli hücre organellerinde çeşitli amorf veya kristal birikintiler şeklinde oluşur. Çoğu zaman, protein kristalleri çekirdekte, daha az sıklıkla - hiyaloplazmada, plastid stromada, endoplazmik retikulum sarnıçlarının uzantılarında, peroksizomların ve mitokondri matrisinde bulunabilir. Protein kristallerinin boyutu çoğunlukla ışık mikroskobunun çözünürlüğünün ötesindedir.
Depolama proteinleri basit proteinler kategorisine aittir. proteinler karmaşık proteinlerin aksine - proteinler protoplastın temelini oluşturur. İÇİNDE en büyük sayı kuru tohumların depo dokusunda şu şekilde biriktirilirler: aleuron taneleri, veya protein cisimleri.
Alöron taneleri genellikle küresel şekillidir ve boyutları farklılık gösterir (0,2-20 µm). Bir zarla çevrilidirler ve içine kristalin kalıntıların daldırıldığı amorf bir protein matrisi içerirler - bir (daha az sıklıkla, 2-3) eşkenar dörtgen şekilli ve yuvarlak protein kristali globoidler(birden çoğa) ( pirinç. 2.11). Globoidler fitinden (inositol heksafosforik asit tuzu) oluşur ve rezerv fosfor için bir depolama alanıdır. Kristal içeren aleuron taneciklerine denir karmaşık. Yağlı tohumların (keten, ayçiçeği, kabak, hardal, hint yağı vb.) depo hücrelerinin karakteristik özelliğidirler. Daha az yaygın basit kristal içermeyen, yalnızca amorf protein (baklagiller, pirinç, mısır) içeren aleurone taneleri ( pirinç. 2.12).
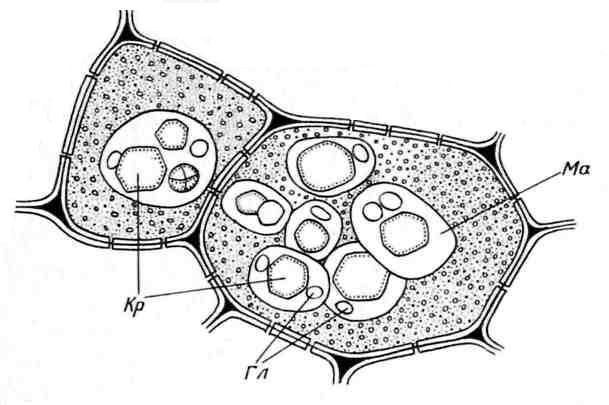
Pirinç. 2.11. Hint fasulyesi tohumlarının endosperm hücrelerindeki aleuron taneleri:Kr.– protein kristalleri; Gl– globoidler; anne– protein matrisi.
Tohum gelişimi sırasında depolama proteinleri vakuollerde biriktirilir. Tohumlar olgunlaştığında, dehidrasyonla birlikte, protein vakuolleri kurur, protein ve fitin çözeltiden çöker ve kristalleşebilir. Tohum çimlenmesi sırasında aleuron taneleri suyu emer, şişer ve yavaş yavaş tipik vakuollere dönüşür. Globoidlerin proteinleri ve maddeleri fidenin büyümesi ve gelişmesi için harcanır.
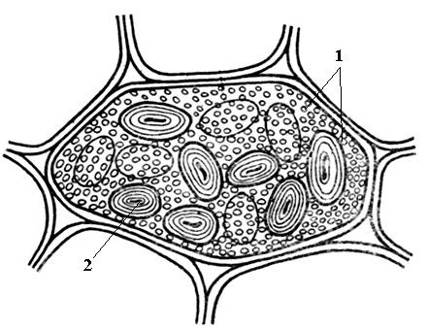
Pirinç. 2.12. Fasulye tohumu kotiledon hücresindeki basit aleurone ve nişasta taneleri: 1 – basit aleuron taneleri; 2 – nişasta tanesi.
Protein kalıntıları Lugol reaktifi ile altın sarısına boyanabilir.
Kalsiyum oksalat kristalleri sıklıkla bitki hücrelerinde bulunur. Sadece vakuollerde biriktirilirler. Kalsiyum oksalat kristallerinin şekli oldukça çeşitlidir ( pirinç. 2.13) ve genellikle tıbbi bitki materyallerinin teşhisinde kullanılan belirli bitkilere özgüdür. Olabilir tek kristaller eşkenar dörtgen, oktahedral veya uzun şekil (banotu yaprakları), Dürzi– küresel kristallerin yıldız şeklinde iç içe büyümeleri (knotweed yaprakları, datura, sinameki, ravent kökleri), Raphidler– demetler halinde toplanan küçük iğne şeklinde kristaller (vadideki zambak yaprakları, kök boya rizomları), stiloidler– daha büyük, çubuk şeklinde kristaller (vadi yapraklarındaki zambak) ve kristal kum– birçok küçük tek kristalden oluşan kümeler (belladona yaprakları). En yaygın olanı drusendir. 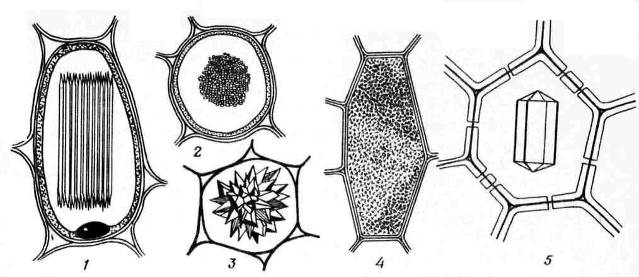
Pirinç. 2.13. Kalsiyum Oksalat Kristal Formları: 1,2 – raphidalar (1 – yandan görünüş, 2 – kesit görünüş); 3 – druzen; 4 – kristal kum; 5 – tek kristal.
Kabuktaki lifler boyunca veya bazı bitkilerin yaprak damarları boyunca (meşe kabuğu, meyan kökü, sinameki yaprakları) kristal taşıyan astar– bulunan paralel satırlar tek kristal kalsiyum oksalat içeren hücreler ( Şekil 2.14).
![]()
Pirinç. 2.14. Sinameki yaprağında kristal astarlı bir damar.
Aşırı iyonları serbest bırakan hayvanlardan farklı olarak dış çevre idrarla birlikte olmayan bitkiler gelişmiş organlar salgılar onları dokularda biriktirmeye zorlanır. Genellikle kalsiyum oksalat kristallerinin hücre yaşamının son ürünü olduğuna ve fazla kalsiyumun uzaklaştırılmasının amaçlandığına inanılır. Nitekim bitkilerin zaman zaman döktüğü organ ve dokularda (yapraklar ve ağaç kabuğu) büyük miktarlarda kristaller oluşur. Ancak vakuollerden kristallerin kaybolabileceğine dair kanıtlar vardır. Bu durumda rezerv kalsiyumun depolandığı yer olarak düşünülebilirler.
Kalsiyum oksalat kristallerinin kimyasal yapısı, konsantre mineral asitler. Hidroklorik asidin etkisi altında kristaller çözülür. Sülfürik asite maruz kaldığında kalsiyum oksalat, çözünmeyen kalsiyum sülfata (alçıtaşı) dönüşerek çok sayıda iğne şeklinde kristal oluşturur.
Kristalin kapanımlara yakın sistolitler. Çoğunlukla kalsiyum karbonat veya silikadan oluşurlar ve hücre duvarının hücre içine çıkıntı yapan çıkıntılarında ortaya çıkan küme şeklindeki oluşumlardır ( pirinç. 2.15). Sistolitler ısırgan otu ve dut ailelerinin bitkilerinin karakteristik özelliğidir. Sistolitlerin önemi henüz açıklığa kavuşturulmamıştır.
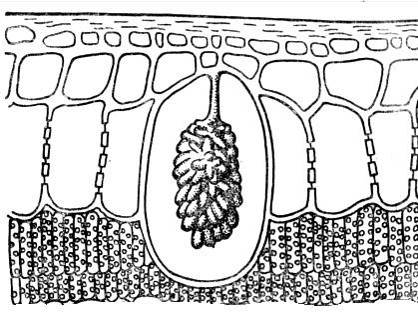
Pirinç. 2.15. Ficus yaprağının epidermal hücresindeki sistolit.
Hücre duvarı
Hücre duvarı ( hücre zarı) – karakteristik özellik Bitki hücresini hayvan hücresinden ayıran özellik. Hücre duvarı hücreye kendine özgü şeklini verir. Duvarın enzimatik olarak çıkarıldığı özel besin ortamlarında kültürlenen bitki hücreleri her zaman küresel bir şekil alır. Hücre duvarı hücreye güç verir ve protoplastı korur; turgor basıncını dengeler ve böylece plazmalemmanın yırtılmasını önler. Hücre duvarlarının toplanması, bitki gövdesini destekleyen ve ona mekanik güç veren iç iskeleti oluşturur.
Hücre duvarı renksiz ve şeffaftır, kolayca geçirgendir güneş ışığı. Genellikle duvarlar suya doyurulur. Hücre duvarı sistemi, suyu ve içinde çözünmüş düşük molekül ağırlıklı bileşikleri taşır (apoplast boyunca taşıma).
Hücre duvarı esas olarak polisakkaritlerden oluşur ve bunlar polisakkaritlerden oluşur. iskelet maddeleri Ve matris maddeleri.
İskelet maddesi bitki hücre duvarı selüloz (lif) bir beta-1,4-D-glukandır. Bu en yaygın olanıdır organik madde biyosfer. Selüloz molekülleri çok uzun, dallanmamış zincirlerdir; birkaç düzinelik gruplar halinde birbirine paralel olarak bulunurlar ve çok sayıda molekül tarafından bir arada tutulurlar. hidrojen bağları. Sonuç olarak, mikrofibriller duvarın yapısal çerçevesini oluşturan ve gücünü belirleyen. Selüloz mikrofibrilleri yalnızca elektron mikroskobu ile görülebilir, çapları 10-30 nm'dir ve uzunlukları birkaç mikrona ulaşır.
Selüloz çözünmez ve suda şişmez. Kimyasal olarak son derece inerttir ve organik solventlerde, konsantre alkalilerde ve seyreltik asitlerde çözünmez. Selüloz mikrofibrilleri elastiktir ve çok gergindir (çeliğe benzer). Bu özellikler selüloz ve ürünlerinin yaygın kullanımını belirlemektedir. Neredeyse tamamı selülozdan oluşan pamuk elyafının dünya üretimi yılda 1,5 10 7 tondur. Selülozdan dumansız barut, asetat ipek ve viskon, selofan ve kağıt elde edilir. Kalitatif reaksiyon bir reaktif ile selüloz üzerinde gerçekleştirildi klor-çinko-iyot selüloz hücre duvarı mavi-mor renge döner.
Mantarlarda hücre duvarının iskelet maddesi kitin– glikozamin kalıntılarından oluşan bir polisakkarit. Kitin selülozdan bile daha dayanıklıdır.
Mikrofibriller amorf bir ortama daldırılmıştır. matris genellikle suya doymuş plastik bir jeldir. Matris, molekülleri birkaç farklı şekerin kalıntılarından oluşan ve selülozdan daha kısa ve dallanmış zincirlerden oluşan karmaşık bir polisakkarit karışımıdır. Matris polisakkaritleri, hücre duvarının güçlü şişme, suya karşı yüksek geçirgenlik ve içinde çözünmüş düşük molekül ağırlıklı bileşikler ve katyon değişim özellikleri gibi özelliklerini belirler. Matris polisakkaritleri iki gruba ayrılır: pektin maddeleri Ve hemiselüloz.
Pektik maddeler suda kuvvetli bir şekilde şişer veya çözülür. Alkaliler ve asitler tarafından kolayca yok edilirler. Pektin maddelerinin en basit temsilcileri suda çözünür pektik asitler– 1,4-bağlarla doğrusal zincirlere (alfa-1,4-D-galakturonan) bağlanan alfa-D-galakturonik asitin (100 birime kadar) polimerizasyon ürünleri. Pektik asitler (pektinler)– bunlar, karboksil gruplarının kısmen metillendiği alfa-D-galakturonik asidin daha yüksek molekül ağırlıklı (100-200 birim) polimerik bileşikleridir. Pektatlar Ve pektinatlar– pektik ve pektik asitlerin kalsiyum ve magnezyum tuzları. Pektik asitler, pektatlar ve pektinatlar, şeker ve organik asitlerin varlığında suda yoğun jeller oluşturacak şekilde çözünürler.
Bitkilerin hücre duvarlarında esas olarak protopektinler- arabinanlar ve galaktanlarla birlikte metoksilenmiş poligalakturonik asidin yüksek molekül ağırlıklı polimerleri; dikotiledonlu bitkilerde galakturonan zincirleri az miktarda ramnoz içerir. Protopektinler suda çözünmez.
Hemiselülozlar nötr şeker kalıntılarından oluşturulan dallı zincirler olup, glikoz, galaktoz, mannoz, ksiloz daha yaygındır; polimerizasyon derecesi 50-300. Hemiselülozlar kimyasal olarak pektin maddelerine göre daha stabildir; hidrolize olmaları daha zordur ve suda daha az şişerler. Hemiselülozlar tohumların hücre duvarlarında yedek madde olarak (hurma, hurma) birikebilir. Pektin maddeleri ve hemiselülozlar karşılıklı geçişlerle birbirine bağlanır. Hücre duvarı matrisinde polisakkaritlerin yanı sıra özel bir yapısal protein de bulunur. Arabinoz şeker kalıntılarına bağlanır ve bu nedenle bir glikoproteindir.
Matris polisakkaritleri, selüloz mikrofibrilleri arasındaki boşlukları doldurmaktan daha fazlasını yapar. Zincirlerinin düzenli bir şekilde dizilmesi ve hem birbirleriyle hem de mikrofibrillerle çok sayıda bağ oluşturması, hücre duvarının gücünü önemli ölçüde artırır.
Bitki hücre duvarları sıklıkla kimyasal modifikasyonlara uğrar. Odunlaşma, veya odunlaşma matriste biriktirildiğinde ortaya çıkar lignin– suda çözünmeyen, fenolik yapıya sahip bir polimer bileşiği. Odunlaşmış hücre duvarı elastikiyetini kaybeder, sertliği ve basınç dayanımı keskin bir şekilde artar ve su geçirgenliği azalır. Lignin için reaktifler şunlardır: 1) floroglüsinol Ve konsantre hidroklorik asit veya sülfürik asit(odunlanmış duvarlar kiraz kırmızısı bir renk alır) ve 2) sülfatanilin, odunlaşmış duvarların etkisi altında limon sarısı olur. Lignifikasyon, ksilemin (ahşap) iletken dokusunun ve sklerenkimanın mekanik dokusunun hücre duvarlarının karakteristiğidir.
Örnekleme, veya suberinizasyon birikmesi sonucu ortaya çıkar içeri hidrofobik polimerlerin hücre duvarı – suberina Ve balmumu. Suberin, polimerik yağ asitlerinin esterlerinin bir karışımıdır. Balmumu monomerleri yağlı alkoller ve balmumu esterleridir. Balmumu organik çözücüler tarafından kolaylıkla ekstrakte edilir ve hızla eriyerek kristaller oluşturur. Suberin, organik çözücülerde erimeyen veya çözünmeyen amorf bir bileşiktir. Alternatif paralel katmanlar oluşturan suberin ve balmumu, tüm hücre boşluğunu içeriden bir film şeklinde kaplar. Suberin filmi pratik olarak suya ve gazlara karşı geçirimsizdir, bu nedenle oluşumundan sonra hücre genellikle ölür. Suberizasyon hücre duvarlarının karakteristiğidir örtü dokusu trafik sıkışıklığı Suberize hücre duvarları için reaktif Sudan III, turuncu-kırmızı renk.
Kütinizasyon Epidermal doku hücrelerinin dış duvarları açığa çıkar. Kutin Ve balmumu hücre duvarının dış yüzeyinde bir film şeklinde alternatif katmanlar halinde biriktirilir - tırnak etleri. Kütin, benzer şekilde yağ benzeri bir polimer bileşiğidir. kimyasal doğa ve suberinin özellikleri. Kütikül, bitkiyi bitkinin yüzeyinden suyun aşırı buharlaşmasından korur. Bir reaktifle lekeleyebilirsiniz Sudan III turuncu-kırmızı renktedir.
Mineralizasyon hücre duvarı matriste birikme nedeniyle oluşur büyük miktar mineral maddeler, çoğunlukla silika (silikon oksit), daha az sıklıkla oksalat ve kalsiyum karbonat. Mineraller duvara sertlik ve kırılganlık kazandırır. Silika birikimi at kuyruğu, saz ve otların epidermal hücrelerinin karakteristik özelliğidir. Silisleşme sonucu elde edilen sap ve yaprakların sertliği, salyangozlara karşı koruyucu bir madde görevi görürken, aynı zamanda bitkilerin lezzetini ve besin değerini de önemli ölçüde azaltır.
Bazı özelleşmiş hücrelerin mukus hücre duvarı. Bu durumda, selüloz ikincil duvarı yerine amorf, yüksek oranda hidratlanmış asidik polisakkaritler şu şekilde biriktirilir: balçık Ve diş etleri kimyasal yapısı pektin maddelerine yakındır. Mukus, mukus çözeltileri oluşturmak için suda iyi çözünür. Diş etleri yapışkandır ve iplikler halinde gerilir. Kuruduğunda azgın bir kıvama sahiptirler. Mukus biriktiğinde protoplast yavaş yavaş hücrenin merkezine doğru itilir, hacmi ve vakuolün hacmi giderek azalır. Sonunda hücre boşluğu tamamen mukusla dolabilir ve hücre ölür. Bazı durumlarda mukus birincil hücre duvarından yüzeye geçebilir. Golgi aygıtı, mukus sentezinde ve salgılanmasında önemli bir rol oynar.
Bitki hücrelerinin salgıladığı mukus, çeşitli işlevler. Böylece kök başlığındaki mukus, yağlayıcı görevi görerek kök ucunun toprakta büyümesini kolaylaştırır. Böcek öldürücü bitkilerin (sundews) mukus bezleri, böceklerin yapıştığı tuzaklayıcı mukus salgılar. Tohum kabuğunun (keten, ayva, muz) dış hücrelerinin salgıladığı müsilaj, tohumun toprak yüzeyine sabitlenmesini sağlar ve fidenin kurumasını önler. Mukus bir reaktifle boyandı metilen mavisi mavi renkte.
Diş etlerinin salınması genellikle bitkiler yaralandığında meydana gelir. Örneğin kiraz ve eriklerde gövde ve dalların yaralı bölgelerinden diş eti akıntısı sıklıkla görülmektedir. Kiraz tutkalı sertleştirilmiş bir sakızdır. Sakız gerçekleştirir koruyucu fonksiyon, yaranın yüzeyden kapatılması. Sakızlar esas olarak baklagil familyalarından (akasya, tragacanth astragalus) ve erik alt familyasından Rosaceae familyasından (kiraz, erik, kayısı) odunsu bitkilerden oluşur. Diş etleri ve müsilajlar tıpta kullanılmaktadır.
Hücre duvarı protoplastın hayati aktivitesinin bir ürünüdür. Golgi aygıtında matris polisakkaritleri, duvar glikoproteini, lignin ve mukus oluşur. Selüloz sentezi, mikrofibrillerin oluşumu ve yönlendirilmesi plazmalemma tarafından gerçekleştirilir. Büyük rol mikrofibrillerin oryantasyonunda, plazmalemmanın yakınında biriken mikrofibrillere paralel olarak yerleştirilmiş mikrotübüllere aittir. Mikrotübüller yok edilirse yalnızca izodiametrik hücreler oluşur.
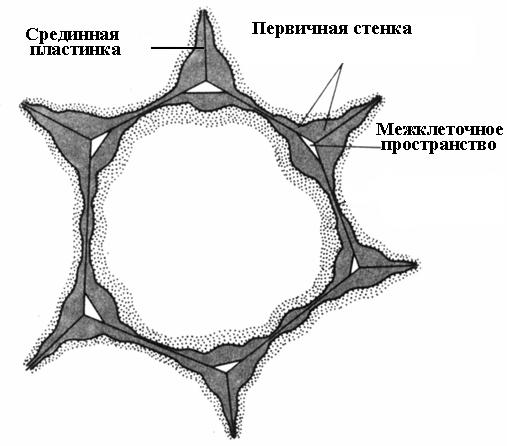
Hücre duvarı oluşumu hücre bölünmesi sırasında başlar. Bölünme düzleminde, iki yavru hücre için ortak olan tek bir katman olan bir hücre plakası oluşur. Yarı sıvı kıvamında pektin maddelerinden oluşur; selüloz yoktur. Yetişkin bir hücrede hücre plakası korunur, ancak değişikliklere uğrar, bu yüzden buna denir. medyan, veya hücreler arası plaka (hücreler arası madde)(pirinç. 2.16). Medyan plaka genellikle çok incedir ve neredeyse ayırt edilemez.
Hücre plakasının oluşmasından hemen sonra yavru hücrelerin protoplastları kendi hücre duvarlarını oluşturmaya başlar. Hem hücre plakasının yüzeyinde hem de daha önce ana hücreye ait olan diğer hücre duvarlarının yüzeyinde içeriden biriktirilir. Bölünmeden sonra hücre, merkezi vakuolün oluşumu ve büyümesiyle ilişkili olarak suyun hücre tarafından yoğun ozmotik emiliminin neden olduğu uzama büyüme fazına girer. Turgor basıncı duvarı germeye başlar, ancak sürekli olarak yeni mikrofibril ve matris maddeleri bölümlerinin birikmesi nedeniyle yırtılmaz. Yeni malzeme kısımlarının birikmesi protoplastın tüm yüzeyi üzerinde eşit olarak meydana gelir, böylece hücre duvarının kalınlığı azalmaz.
Bölünen ve büyüyen hücrelerin duvarlarına denir öncelik. Çok fazla (%60-90) su içerirler. Kuru maddede matris polisakkaritleri (%60-70) hakimdir, selüloz içeriği %30'u geçmez ve lignin yoktur. Birincil duvarın kalınlığı çok küçüktür (0,1-0,5 mikron).
Birçok hücre için, hücre büyümesinin durmasıyla eş zamanlı olarak hücre duvarı birikimi de durur. Bu tür hücreler yaşamın sonuna kadar ince bir birincil duvarla çevrilidir ( pirinç. 2.16).
Pirinç. 2.16. Birincil duvarı olan parankima hücresi.
Diğer hücrelerde hücre son boyutuna ulaştıktan sonra bile duvar birikmesi devam eder. Bu durumda duvar kalınlığı artar ve hücre boşluğunun kapladığı hacim azalır. Bu süreç denir ikincil kalınlaşma duvarlar ve duvarın kendisi denir ikincil(pirinç. 2.17).
İkincil duvar, esas olarak mekanik bir destek işlevi gören ek bir duvar olarak düşünülebilir. Kesinlikle ikincil duvar ahşabın, tekstil elyafının, kağıdın özelliklerinden sorumludur. İkincil duvar, birincil duvara göre önemli ölçüde daha az su içerir; birbirine paralel konumlanmış selüloz mikrofibrilleri (kuru maddenin ağırlığının %40-50'si) hakimdir. Matris polisakkaritlerinden hemiselülozlar (%20-30) tipiktir ve çok az pektin maddesi vardır. İkincil hücre duvarları genellikle lignifikasyona uğrar. Odunlaşmamış ikincil duvarlarda (keten lifleri, pamuk kılları) selüloz içeriği %95'e ulaşabilir. Harika içerik ve mikrofibrillerin kesin olarak sıralanmış yönelimi, yüksek mekanik özellikler ikincil duvarlar. Çoğunlukla, ikincil odunlaşmış hücre duvarına sahip hücreler, ikincil kalınlaşma tamamlandıktan sonra ölürler.
Medyan lamina komşu hücreleri birbirine yapıştırır. Çözünmesi durumunda hücre duvarları birbiriyle temasını kaybeder ve ayrılır. Bu süreç denir maserasyon. Orta plakadaki pektin maddelerinin pektinaz enzimi kullanılarak çözünür bir duruma dönüştürüldüğü ve daha sonra suyla (armut, kavun, şeftali, muz gibi olgunlaşmış meyveler) yıkandığı doğal maserasyon oldukça yaygındır. Orta plakanın tüm yüzey üzerinde değil, yalnızca hücrelerin köşelerinde çözündüğü kısmi maserasyon sıklıkla gözlenir. Turgor basıncı nedeniyle bu yerlerdeki komşu hücreler yuvarlanır ve bunun sonucunda formasyon oluşur. hücreler arası boşluklar(pirinç. 2.16). Hücreler arası boşluklar, su buharı ve gazlarla dolu tek dallı bir ağ oluşturur. Böylece hücreler arası boşluklar hücrelerin gaz alışverişini artırır.
Özellik ikincil duvar - ikincil duvarda kalınlaşmamış alanların kalması sonucunda birincil duvarın üstünde düzensiz birikmesi - gözenekler. İkincil duvar büyük bir kalınlığa ulaşmazsa gözenekler küçük çöküntüler gibi görünür. İkincil duvarı güçlü olan hücrelerde kesitteki gözenekler, hücre boşluğundan birincil duvara kadar uzanan radyal kanallar formundadır. Gözenek kanalının şekline bağlı olarak iki tür gözenek vardır: basit ve hakkında kenarlı(Şekil 2.17).
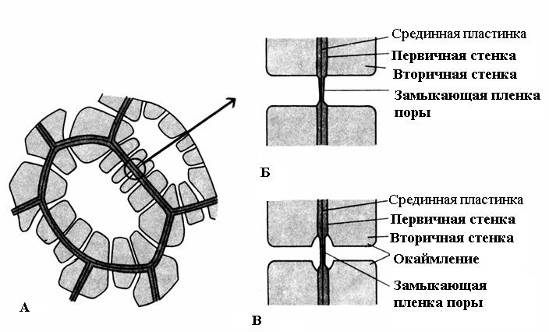
Pirinç. 2.17. Gözenek türleri: A – ikincil duvarlara ve çok sayıda basit gözeneklere sahip hücreler; B – bir çift basit gözenek; B – bir çift kenarlı gözenek.
sen basit gözenekler Gözenek kanalının çapı tüm uzunluğu boyunca aynıdır ve dar bir silindir şeklindedir. Basit gözenekler parankima hücrelerinin, sak ve ağaç liflerinin karakteristiğidir.
İki bitişik hücredeki gözenekler birbirinin karşısında görünme eğilimindedir. Bu ortak gözenekler, orta plaka ve birincil duvardan oluşan ince bir bölmeyle ayrılan tek bir kanala benzer. Komşu hücrelerin bitişik duvarlarının iki gözeneklerinin bu kombinasyonuna denir. gözenek çiftleri ve bir bütün olarak çalışır. Bunları ayıran duvar kısmına denir. gözeneklerin kapanış filmi, veya gözenek zarı. Canlı hücrelerde, gözeneklerin kapanış filmi çok sayıda maddeyle doludur. plazmodezmata(pirinç. 2.18).
Plazmodesmata yalnızca doğuştan gelen bitki hücreleri. Bitişik hücrelerin duvarını geçen sitoplazma şeritleridir. Bir hücredeki plazmodezma sayısı çok fazladır - birkaç yüz ila on binlerce arasında; genellikle plazmodezmata gruplar halinde toplanır. Plazmodesmal kanalın çapı 30-60 nm'dir. Duvarları, komşu hücrelerin plazmalemması ile devam eden plazmalemma ile kaplıdır. Plazmodesmata'nın merkezinde bir membran silindiri vardır - plazmadesmata'nın merkezi çubuğu, her iki hücrenin endoplazmik retikulumunun elemanlarının zarları ile süreklidir. Kanaldaki merkezi çubuk ile plazmalemma arasında, komşu hücrelerin hyaloplazması ile devam eden hyaloplazma vardır.
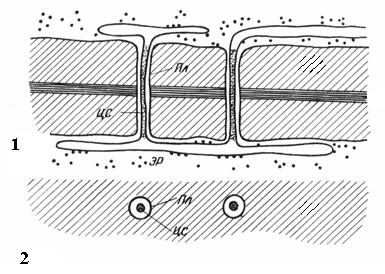
Pirinç. 2.18. Elektron mikroskobu altında Plasmodesmata (şema): 1 – uzunlamasına kesitte; 2 – kesitte; Pl– plazmalemma; CA– plazmodesmata'nın merkezi çubuğu; Acil servis– endoplazmik retikulumun elemanı.
Böylece hücre protoplastları birbirlerinden tamamen izole edilmez, ancak plazmodesmata kanalları aracılığıyla iletişim kurarlar. İyonların ve küçük moleküllerin hücreler arası taşınmasını taşırlar ve ayrıca hormonal uyarıları iletirler. Plasmodesmata yoluyla hücrelerin protoplastları bitki organizması adı verilen tek bir bütün oluşturur simplast ve maddelerin plazmodezmata yoluyla taşınmasına denir simplastik farklı apoplastik Hücre duvarları ve hücreler arası boşluklar boyunca taşıma.
sen kenarlı gözenekler(pirinç. 2.17) kanal, hücre duvarının birikmesi sırasında keskin bir şekilde daralır, bu nedenle hücre boşluğuna açılan gözeneğin iç açıklığı, birincil duvara bitişik olan dıştan çok daha dardır. Kenarlı gözenekler, ahşabın su ileten elemanlarının erken ölen hücrelerinin karakteristiğidir. Bunlarda gözenek kanalı, huni şeklinde kapatma filmine doğru genişler ve ikincil duvar, kanalın genişleyen kısmı üzerinde bir silindir şeklinde asılı kalarak bir gözenek odası oluşturur. Sınırlı gözenek adı, yüzeyden bakıldığında iç açıklığın küçük bir daire veya dar boşluk, dış delik ise daha büyük çaplı bir daire veya daha geniş bir yarık şeklinde iç delik ile sınırlanır.
Gözenekler içeriye taşınmayı kolaylaştırır