Ribosomer– ribonukleoproteingranulat 25 nm i størrelse. De består af to underenheder: lille (10 nm) og stor (15 nm), mellem hvilke en streng af messenger-RNA er placeret under proteinbiosyntese (translation). I dette tilfælde binder den lille underenhed til RNA, og den store katalyserer dannelsen af polypeptidkæder. Ribosomale underenheder dannes i nukleolerne og forlader derefter kernen ind i cytoplasmaet gennem nukleare porer. Samlingen af ribosomer fra deres underenheder sker før starten af proteinsyntesen, og efter afslutningen af syntesen af polypeptidkæden desintegrerer de igen.
En syntetisk aktiv celle indeholder flere millioner ribosomer, som udgør omkring 5 % af dens tørre masse. Skelne gratis ribosomer (ikke forbundet med membraner og placeret i hyaloplasmaet i suspension) og ufri ribosomer (associeret med membraner i det cytoplasmatiske reticulum). Ribosomer kan lokaliseres enkeltvis (i dette tilfælde er de funktionelt inaktive), men oftere er de forbundet i kæder, der er spændt, som perler, på trådlignende messenger RNA-molekyler ( polyribosomer, polysomer). Frie ribosomer syntetiserer proteiner til cellens egne behov, mens ikke-frie ribosomer syntetiserer proteiner til eksport.
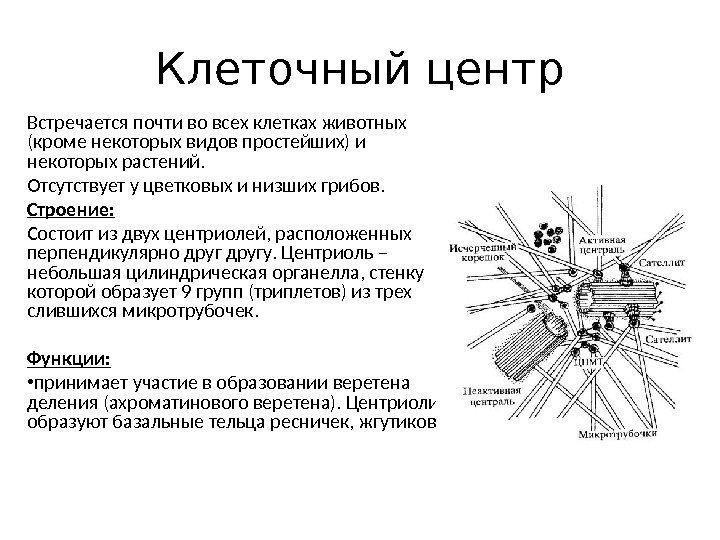
Cellecenter (cytocenter)– består af to placeret vinkelret på hinanden centrioler.
Figur 2-8. Cellecenter (A) og centriolstruktur (B).
1. Centriol.
2. Satellitter.
3. Triplet af mikrotubuli.
4. Mikrotubuli.
(Ifølge V.L. Bykov).
Centriole er en hul cylinder 200 nm tyk og 300-500 nm lang. Centriolvæggen er dannet af 9 tripletter af mikrotubuli, 24 nm tykke, bygget af kugleformet protein tubulin. Nabotripletter af mikrotubuli er forbundet i form af broer af proteinmolekyler dynein. Hver triplet af mikrotubuli er også forbundet med sfæriske strukturer - satellitter. Mikrotubuli divergerer fra satellitterne til siderne og danner centosfæren(Figur 2-8).
Cellecentret deltager i dannelsen af delingsspindlen; Under mitose bevæger centrioler sig mod modercellens poler. Derudover deltager centrioler i dannelsen af cilia og flageller.
Cytoskelettets organeller
Cytoskelettet er et komplekst dynamisk tredimensionelt netværk af mikrotubuli, mikrofibriller og mikrofilamenter, som giver: 1) vedligeholdelse og ændring i celleform, 2) distribution og bevægelse af cellekomponenter, 3) transport af stoffer ind og ud af cellen, 4) cellemotilitet, 5) deltager i intercellulære forbindelser.
Mikrotubuli har en tykkelse på 24 nm og en længde på flere mikrometer. Mikrotubulus vægtykkelse er 5 nm, og lumendiameteren er tilsvarende 14 nm. De består af 13 tubulin protofibriller, der løber i en spiral. Mikrotubuli er en del af spindlen og sørger for divergensen af kromosomer under mitose, opretholder cellens form og sikrer dens mobilitet og deltager i transporten af makromolekyler i cellen. Forbundet med mikrotubuli er proteinet kinesin, som er et enzym, der nedbryder ATP og omdanner energien fra dets nedbrydning til mekanisk energi. I den ene ende er kinesinmolekylet forbundet med en bestemt organel, og i den anden glider det ved hjælp af ATP-energi langs mikrotubuli og bevæger organellen i cytoplasmaet.
Mikrotubuli er et labilt system, hvor dissociationen (destruktion) af nogle mikrotubuli og samlingen (dannelsen) af andre sker kontinuerligt. Stedet for dannelse af mikrotubuli ( organiseringscentre for mikrotubuli) er små sfæriske legemer satellitter. De er placeret i de basale kroppe af cilia og cellecentret.
Mellemfilamenter (mikrofibriller) – proteinfilamenter 8-11 nm tykke. De danner rammen af cellen, bevarer dens form og elasticitet, og sikrer også den ordnede indretning af organeller i cellen.
Mikrofilamenter – proteinfilamenter 5-7 nm tykke. De er til stede i alle celler og er placeret i dets kortikale lag (under cytolemmaet). Sammensætningen af de proteiner, der danner dem i forskellige celler forskellige (aktin, myosin, tropomyosin). De danner skelettet, cellens ramme, dets intracellulære kontraktile apparat, sikrer ændringer i cellernes form og bevægelse, cytoplasmatisk flow, endocytose og exocytose.
Funktionelle systemer (enheder) i cellen- komplekser af organeller, der under kontrol af kernen sikrer udførelsen af vigtige cellefunktioner. Der er: 1) syntetiske apparater (inklusive endoplasmatisk retikulum ribosomer, Golgi-kompleks; 2 ) energi apparat (mitokondrier); 3) intracellulært fordøjelsesapparat vania(endosomer, lysosomer, peroxisomer); 4) cytoskelet (mikrotubuli, mikrofibriller, mikrofilamenter, cellecenter).

 Organeller er permanente intracellulære strukturer, der har en specifik struktur og udfører tilsvarende funktioner. Organeller er opdelt i to grupper: membran og ikke-membran. Membranorganeller præsenteres i to varianter: dobbeltmembran og enkeltmembran.
Organeller er permanente intracellulære strukturer, der har en specifik struktur og udfører tilsvarende funktioner. Organeller er opdelt i to grupper: membran og ikke-membran. Membranorganeller præsenteres i to varianter: dobbeltmembran og enkeltmembran.
 De dobbelte membrankomponenter er plastider, mitokondrier og cellekerne. Enkeltmembranorganeller omfatter organellerne i det vakuolære system - det endoplasmatiske retikulum, Golgi-komplekset, lysosomer, vakuoler af plante- og svampeceller osv. Ikke-membranorganeller omfatter ribosomer og cellecentret
De dobbelte membrankomponenter er plastider, mitokondrier og cellekerne. Enkeltmembranorganeller omfatter organellerne i det vakuolære system - det endoplasmatiske retikulum, Golgi-komplekset, lysosomer, vakuoler af plante- og svampeceller osv. Ikke-membranorganeller omfatter ribosomer og cellecentret
 Enkeltmembranorganeller Endoplasmatisk reticulum (ER) eller reticulum - et komplekst system af kanaler og hulrum forskellige former(tubuli, cisterner), der penetrerer hele cytoplasmaet. a) Ru eller granulært endoplasmatisk retikulum: membranerne er dækket af små granula - ribosomer. Funktioner: syntese af polypeptider, deres delvise modifikation og transport b) Glat eller agranulært, endoplasmatisk retikulum: membraner er blottet for ribosomer, men enzymer af lipid- og kulhydratmetabolisme akkumuleres her. Funktioner: syntese af lipider, steroider, kulhydrater, deres transport.
Enkeltmembranorganeller Endoplasmatisk reticulum (ER) eller reticulum - et komplekst system af kanaler og hulrum forskellige former(tubuli, cisterner), der penetrerer hele cytoplasmaet. a) Ru eller granulært endoplasmatisk retikulum: membranerne er dækket af små granula - ribosomer. Funktioner: syntese af polypeptider, deres delvise modifikation og transport b) Glat eller agranulært, endoplasmatisk retikulum: membraner er blottet for ribosomer, men enzymer af lipid- og kulhydratmetabolisme akkumuleres her. Funktioner: syntese af lipider, steroider, kulhydrater, deres transport.

 Funktioner: Forbinder alle cellemembranstrukturer i samlet system. Det er overfladen, hvorpå alle intracellulære processer forekommer (syntese af membranproteiner, lipider og kulhydrater). Rumligt adskiller cellen. Transporten af stoffer sker gennem et system af kanaler.
Funktioner: Forbinder alle cellemembranstrukturer i samlet system. Det er overfladen, hvorpå alle intracellulære processer forekommer (syntese af membranproteiner, lipider og kulhydrater). Rumligt adskiller cellen. Transporten af stoffer sker gennem et system af kanaler.
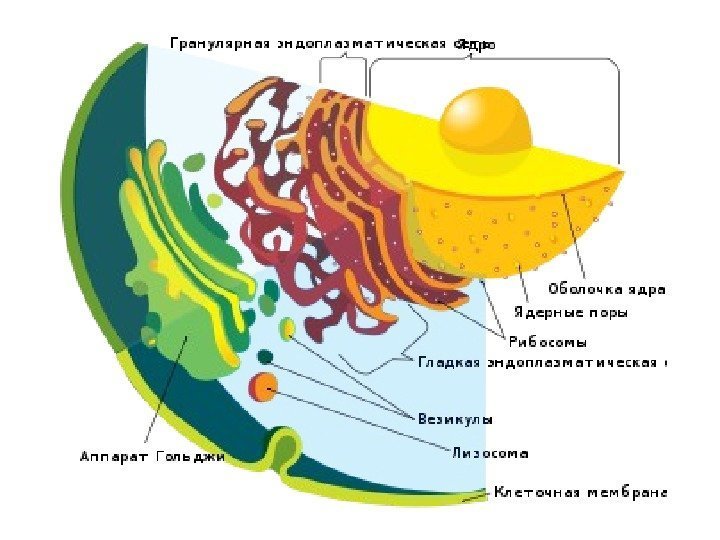
 Golgi-komplekset er til stede i næsten alle celler (med undtagelse af erytrocytter og sædceller). Struktur: Et system af stablede flade membransække - cisterner, rør og tilhørende vesikler. Funktioner: Transport af stoffer, hovedsageligt proteiner og lipider, der kommer fra det endoplasmatiske retikulum, deres foreløbige kemiske omlejring, akkumulering, pakning i vesikler, dannelse af lysosomer.
Golgi-komplekset er til stede i næsten alle celler (med undtagelse af erytrocytter og sædceller). Struktur: Et system af stablede flade membransække - cisterner, rør og tilhørende vesikler. Funktioner: Transport af stoffer, hovedsageligt proteiner og lipider, der kommer fra det endoplasmatiske retikulum, deres foreløbige kemiske omlejring, akkumulering, pakning i vesikler, dannelse af lysosomer.
 Golgi-apparatet blev opkaldt efter den italienske videnskabsmand Camillo Golgi, som først opdagede det i 1897
Golgi-apparatet blev opkaldt efter den italienske videnskabsmand Camillo Golgi, som først opdagede det i 1897

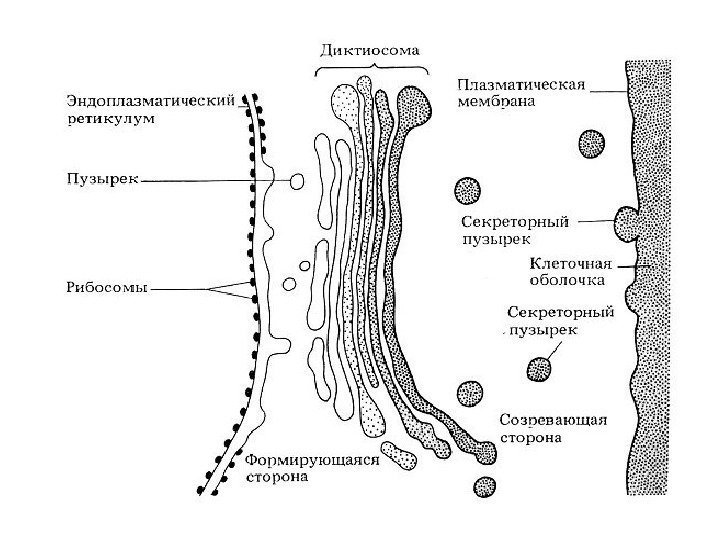
 Funktioner af Golgi Complex 1) sortering, akkumulering og udskillelse af sekretoriske produkter 2) færdiggørelse af post-translationel modifikation af proteiner 3) akkumulering af lipidmolekyler og dannelse af lipoproteiner 4) dannelse af lysosomer 5) syntese af polysaccharider til dannelse af glycoproteiner , voks, slim, matrixstoffer cellevægge planter (hemicellulose, pektiner) 6) dannelse af cellepladen efter kernedeling i planteceller 7) deltagelse i dannelsen af akrosomet; dannelse af kontraktile vakuoler af protozoer.
Funktioner af Golgi Complex 1) sortering, akkumulering og udskillelse af sekretoriske produkter 2) færdiggørelse af post-translationel modifikation af proteiner 3) akkumulering af lipidmolekyler og dannelse af lipoproteiner 4) dannelse af lysosomer 5) syntese af polysaccharider til dannelse af glycoproteiner , voks, slim, matrixstoffer cellevægge planter (hemicellulose, pektiner) 6) dannelse af cellepladen efter kernedeling i planteceller 7) deltagelse i dannelsen af akrosomet; dannelse af kontraktile vakuoler af protozoer.
 Golgi-apparatets funktioner: I Golgi-apparatets cisterner modnes proteiner beregnet til sekretion, transmembranproteiner i plasmamembranen, lysosomproteiner osv. De modnende proteiner bevæger sig sekventielt gennem organelcisternerne, hvori deres endelige foldning sker. samt modifikationer - glykosylering og phosphorylering.
Golgi-apparatets funktioner: I Golgi-apparatets cisterner modnes proteiner beregnet til sekretion, transmembranproteiner i plasmamembranen, lysosomproteiner osv. De modnende proteiner bevæger sig sekventielt gennem organelcisternerne, hvori deres endelige foldning sker. samt modifikationer - glykosylering og phosphorylering.
 Adskillelse af proteiner i 3 strømme: 1. lysosomale - glykosylerede proteiner (med mannose) kommer ind i cis-sektionen af Golgi-komplekset, nogle af dem fosforyleres, og der dannes en markør for lysosomale enzymer - mannose-6-phosphat. I fremtiden vil disse phosphorylerede proteiner ikke undergå modifikation, men vil komme ind i lysosomerne. 2. konstitutiv exocytose (konstitutiv sekretion). Denne strømning inkluderer proteiner og lipider, som bliver komponenter i celleoverfladeapparatet, herunder glycocalyx, eller de kan være en del af den ekstracellulære matrix. 3. Inducerbar sekretion - proteiner, der fungerer uden for cellen, celleoverfladeapparatet, under indre miljø legeme. Karakteristisk for sekretoriske celler.
Adskillelse af proteiner i 3 strømme: 1. lysosomale - glykosylerede proteiner (med mannose) kommer ind i cis-sektionen af Golgi-komplekset, nogle af dem fosforyleres, og der dannes en markør for lysosomale enzymer - mannose-6-phosphat. I fremtiden vil disse phosphorylerede proteiner ikke undergå modifikation, men vil komme ind i lysosomerne. 2. konstitutiv exocytose (konstitutiv sekretion). Denne strømning inkluderer proteiner og lipider, som bliver komponenter i celleoverfladeapparatet, herunder glycocalyx, eller de kan være en del af den ekstracellulære matrix. 3. Inducerbar sekretion - proteiner, der fungerer uden for cellen, celleoverfladeapparatet, under indre miljø legeme. Karakteristisk for sekretoriske celler.
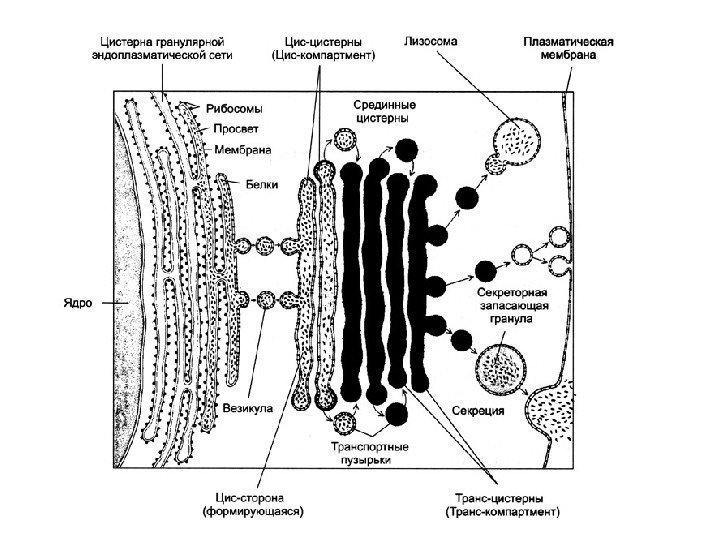
![]() For at afslutte overvejelsen af strukturen og driften af en så kompleks membranorganel som Golgi-apparatet, er det nødvendigt at understrege, at på trods af den tilsyneladende morfologiske homogenitet af dets komponenter, vakuolen og cisternen, er det faktisk ikke kun en samling af vesikler, men et slankt, dynamisk, komplekst organiseret, polariseret system.
For at afslutte overvejelsen af strukturen og driften af en så kompleks membranorganel som Golgi-apparatet, er det nødvendigt at understrege, at på trods af den tilsyneladende morfologiske homogenitet af dets komponenter, vakuolen og cisternen, er det faktisk ikke kun en samling af vesikler, men et slankt, dynamisk, komplekst organiseret, polariseret system.

 Lysosomer Findes i alle celler, spredt over hele cytoplasmaet. Struktur: Enkeltmembran-vesikler af forskellige former og størrelser; indeholder forskellige proteolytiske enzymer (ca. 40). Funktioner: Deltage i intracellulær fordøjelse, dvs. nedbrydning af store molekyler. De kan også ødelægge strukturerne i selve cellen, hvilket forårsager dens død - autolyse.
Lysosomer Findes i alle celler, spredt over hele cytoplasmaet. Struktur: Enkeltmembran-vesikler af forskellige former og størrelser; indeholder forskellige proteolytiske enzymer (ca. 40). Funktioner: Deltage i intracellulær fordøjelse, dvs. nedbrydning af store molekyler. De kan også ødelægge strukturerne i selve cellen, hvilket forårsager dens død - autolyse.
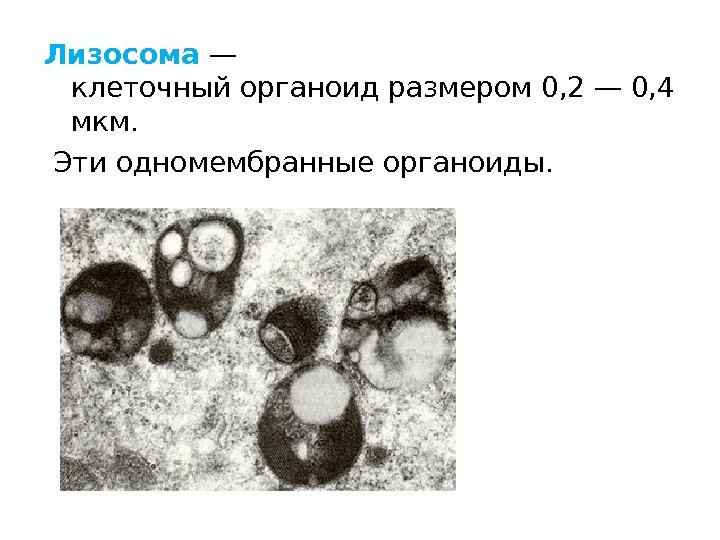
 Lysosomer blev først beskrevet i 1955 af Christian de Duve i dyreceller og blev senere opdaget i planteceller.
Lysosomer blev først beskrevet i 1955 af Christian de Duve i dyreceller og blev senere opdaget i planteceller.
 Lysosomer Hos planter ligner lysosomer til dels lysosomer i deres dannelsesmetode og i funktion. Tilstedeværelsen af lysosomer er karakteristisk for celler af alle eukaryoter. Der er ingen uprokaryote lysosomer, da de mangler fagocytose og ikke har intracellulær fordøjelse.
Lysosomer Hos planter ligner lysosomer til dels lysosomer i deres dannelsesmetode og i funktion. Tilstedeværelsen af lysosomer er karakteristisk for celler af alle eukaryoter. Der er ingen uprokaryote lysosomer, da de mangler fagocytose og ikke har intracellulær fordøjelse.
 Tegn på lysosomer Et af tegnene på lysosomer er tilstedeværelsen i dem af en række enzymer (syrehydrolaser), der er i stand til at nedbryde proteiner, kulhydrater, lipider og nukleinsyrer.
Tegn på lysosomer Et af tegnene på lysosomer er tilstedeværelsen i dem af en række enzymer (syrehydrolaser), der er i stand til at nedbryde proteiner, kulhydrater, lipider og nukleinsyrer.
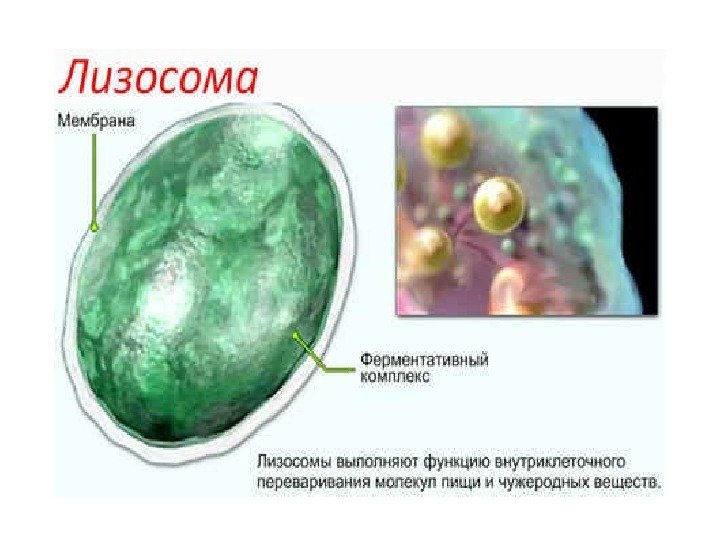
 Lysosomers enzymer omfatter cathepsiner (vævsproteaser), sur ribonuklease, phospholipaser m.fl. Lysosomer indeholder enzymer, der er i stand til at spalte sig fra organiske molekyler sulfat (sulfataser) eller fosfat (syre fosfatase) grupper.
Lysosomers enzymer omfatter cathepsiner (vævsproteaser), sur ribonuklease, phospholipaser m.fl. Lysosomer indeholder enzymer, der er i stand til at spalte sig fra organiske molekyler sulfat (sulfataser) eller fosfat (syre fosfatase) grupper.
 Dannelse af lysosomer og deres typer Lysosomer dannes af vesikler (vesikler), adskilt fra Golgi-apparatet, og vesikler (endosomer), som stoffer trænger ind i under endocytose. Membranerne i det endoplasmatiske reticulum deltager i dannelsen af autolysosomer (autofagosomer). Alle lysosomproteiner syntetiseres på fastsiddende ribosomer. uden for membraner i det endoplasmatiske reticulum og passerer derefter gennem dets hulrum og gennem Golgi-apparatet.
Dannelse af lysosomer og deres typer Lysosomer dannes af vesikler (vesikler), adskilt fra Golgi-apparatet, og vesikler (endosomer), som stoffer trænger ind i under endocytose. Membranerne i det endoplasmatiske reticulum deltager i dannelsen af autolysosomer (autofagosomer). Alle lysosomproteiner syntetiseres på fastsiddende ribosomer. uden for membraner i det endoplasmatiske reticulum og passerer derefter gennem dets hulrum og gennem Golgi-apparatet.
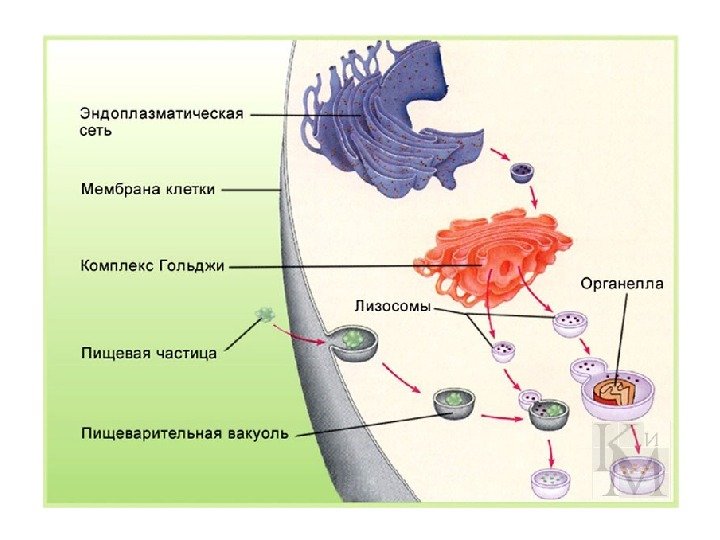
 Lysosomer er heterogene organeller, der har forskellige former, størrelser, ultrastrukturelle og cytokemiske egenskaber. "Typiske" lysosomer i dyreceller er sædvanligvis sfæriske eller oval form. Antallet af lysosomer varierer fra én (en stor vakuole i mange plante- og svampeceller) til flere hundrede eller tusinder (i dyreceller).
Lysosomer er heterogene organeller, der har forskellige former, størrelser, ultrastrukturelle og cytokemiske egenskaber. "Typiske" lysosomer i dyreceller er sædvanligvis sfæriske eller oval form. Antallet af lysosomer varierer fra én (en stor vakuole i mange plante- og svampeceller) til flere hundrede eller tusinder (i dyreceller).
![]() Der er primære og sekundære lysosomer. Førstnævnte er dannet i området af Golgi-apparatet, de indeholder enzymer i en inaktiv tilstand, mens sidstnævnte indeholder aktive enzymer.
Der er primære og sekundære lysosomer. Førstnævnte er dannet i området af Golgi-apparatet, de indeholder enzymer i en inaktiv tilstand, mens sidstnævnte indeholder aktive enzymer.
 Blandt lysosomer kan man også skelne mellem heterolysosomer (fordøjelsesmateriale, der kommer ind i cellen udefra gennem fago- eller pinocytose) og autolysosomer (destruktion af cellens egne proteiner eller organeller).
Blandt lysosomer kan man også skelne mellem heterolysosomer (fordøjelsesmateriale, der kommer ind i cellen udefra gennem fago- eller pinocytose) og autolysosomer (destruktion af cellens egne proteiner eller organeller).
 Den mest udbredte klassifikation af lysosomer og tilhørende kompartmenter er: Tidligt endosom - det modtager endocytiske (pinocytotiske) vesikler. Sen endosom - vesikler med materiale absorberet under pinocytose og vesikler fra Golgi-apparatet med hydrolaser kommer ind i det fra det tidlige endosom.
Den mest udbredte klassifikation af lysosomer og tilhørende kompartmenter er: Tidligt endosom - det modtager endocytiske (pinocytotiske) vesikler. Sen endosom - vesikler med materiale absorberet under pinocytose og vesikler fra Golgi-apparatet med hydrolaser kommer ind i det fra det tidlige endosom.
 klassificering Lysosom - vesikler med en blanding af hydrolaser og fordøjeligt materiale kommer ind i det fra det sene endosom.
klassificering Lysosom - vesikler med en blanding af hydrolaser og fordøjeligt materiale kommer ind i det fra det sene endosom.
 Fagosomklassificering - den omfatter større partikler (bakterier osv.) absorberet af fagocytose. Fagosomer smelter normalt sammen med et lysosom.
Fagosomklassificering - den omfatter større partikler (bakterier osv.) absorberet af fagocytose. Fagosomer smelter normalt sammen med et lysosom.
 klassificering Autophagosom er en region af cytoplasma omgivet af to membraner, som normalt omfatter nogle organeller og dannet under makroautofagi. Sammensmelter med lysosomet.
klassificering Autophagosom er en region af cytoplasma omgivet af to membraner, som normalt omfatter nogle organeller og dannet under makroautofagi. Sammensmelter med lysosomet.
 klassificering Multivesikulære legemer - normalt omgivet af en enkelt membran, indeholder inde i mindre vesikler omgivet af en enkelt membran. De er dannet ved en proces, der minder om mikroautofagi, men indeholder materiale opnået udefra. Dannelsesstadiet svarer til tidlige endosomer.
klassificering Multivesikulære legemer - normalt omgivet af en enkelt membran, indeholder inde i mindre vesikler omgivet af en enkelt membran. De er dannet ved en proces, der minder om mikroautofagi, men indeholder materiale opnået udefra. Dannelsesstadiet svarer til tidlige endosomer.
 klassificering Restlegemer (telolysosomer) er vesikler indeholdende ufordøjet materiale (lipofuscin). I normale celler smelter de sammen med den ydre membran og forlader cellen ved eksocytose. De akkumuleres med aldring eller patologi.
klassificering Restlegemer (telolysosomer) er vesikler indeholdende ufordøjet materiale (lipofuscin). I normale celler smelter de sammen med den ydre membran og forlader cellen ved eksocytose. De akkumuleres med aldring eller patologi.
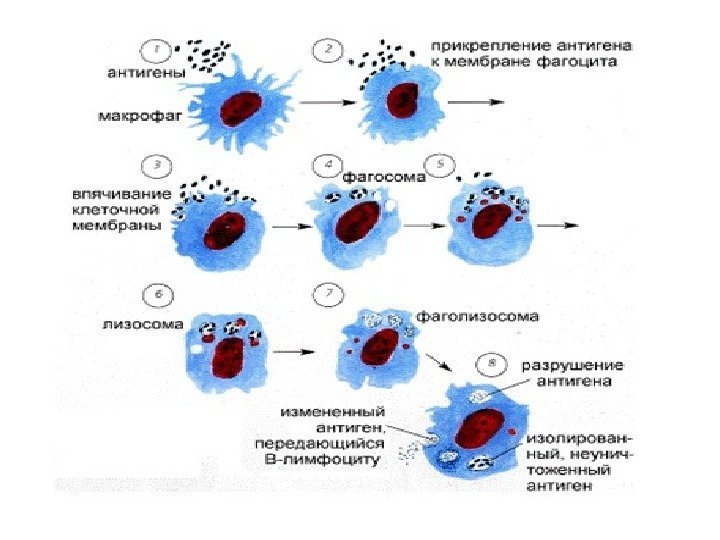
 Lysosomers funktioner: fordøjelse af stoffer eller partikler fanget af cellen under endocytose (bakterier, andre celler); autofagi - ødelæggelse af strukturer, der er unødvendige for cellen, for eksempel under udskiftning af gamle organeller med nye eller fordøjelse af proteiner og andre stoffer produceret inde i selve cellen
Lysosomers funktioner: fordøjelse af stoffer eller partikler fanget af cellen under endocytose (bakterier, andre celler); autofagi - ødelæggelse af strukturer, der er unødvendige for cellen, for eksempel under udskiftning af gamle organeller med nye eller fordøjelse af proteiner og andre stoffer produceret inde i selve cellen
 autolyse er en celles selvfordøjelse, hvilket fører til dens død (nogle gange er denne proces ikke patologisk, men ledsager kroppens udvikling eller differentieringen af nogle specialiserede celler). Eksempel: Når en haletudse forvandles til en frø, fordøjer lysosomer i halens celler den: halen forsvinder, og de stoffer, der dannes under denne proces, absorberes og bruges af andre celler i kroppen. Lysosomers funktioner
autolyse er en celles selvfordøjelse, hvilket fører til dens død (nogle gange er denne proces ikke patologisk, men ledsager kroppens udvikling eller differentieringen af nogle specialiserede celler). Eksempel: Når en haletudse forvandles til en frø, fordøjer lysosomer i halens celler den: halen forsvinder, og de stoffer, der dannes under denne proces, absorberes og bruges af andre celler i kroppen. Lysosomers funktioner
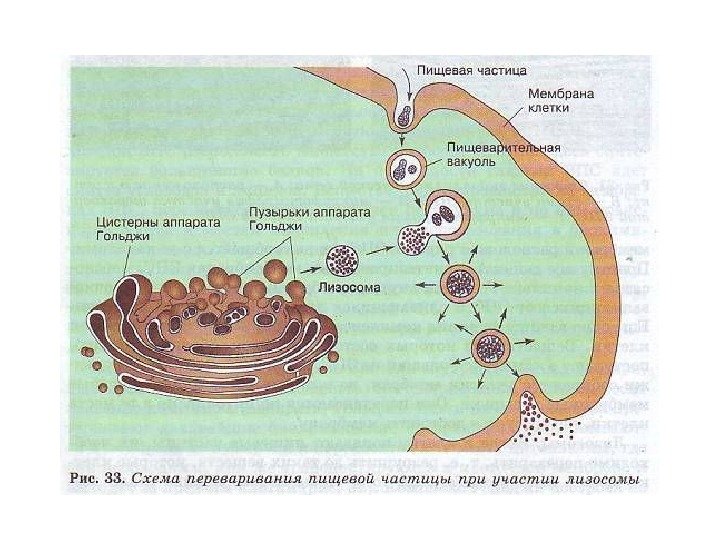
 Klinisk betydning. Sygdomme forbundet med fejlfunktion af lysosomer Nogle gange, på grund af ukorrekt funktion af lysosomer, udvikles der lagersygdomme, hvor enzymer ikke virker eller fungerer dårligt på grund af mutationer. Et eksempel på lagringssygdomme er amaurotisk idioti på grund af glykogenlagring. Bruddet af lysosomet og frigivelsen af hyaloplasmaspaltende enzymer er ledsaget af en kraftig stigning i deres aktivitet. Denne form for stigning i enzymaktivitet observeres for eksempel i foci af nekrose under myokardieinfarkt og under påvirkning af stråling.
Klinisk betydning. Sygdomme forbundet med fejlfunktion af lysosomer Nogle gange, på grund af ukorrekt funktion af lysosomer, udvikles der lagersygdomme, hvor enzymer ikke virker eller fungerer dårligt på grund af mutationer. Et eksempel på lagringssygdomme er amaurotisk idioti på grund af glykogenlagring. Bruddet af lysosomet og frigivelsen af hyaloplasmaspaltende enzymer er ledsaget af en kraftig stigning i deres aktivitet. Denne form for stigning i enzymaktivitet observeres for eksempel i foci af nekrose under myokardieinfarkt og under påvirkning af stråling.
 Vacuole Vacuoles er enkeltmembranorganeller, der er "beholdere" fyldt med vandige opløsningerøkologiske og uorganiske stoffer. ER- og Golgi-komplekset deltager i dannelsen af vakuoler. Unge planteceller indeholder mange små vakuoler, som så, efterhånden som cellerne vokser og differentierer, smelter sammen og danner én stor central vakuole. Den centrale vakuole kan optage op til 95% af volumenet af en moden celle; kernen og organellerne skubbes mod celle membran. Membranbegrænsende plantevakuole, kaldet tonoplast. Væsken, der fylder plantevakuolen, kaldes cellesaft. En del cellesaft omfatter vandopløselige organiske og uorganiske salte, monosaccharider, disaccharider, aminosyrer, endelige eller toksiske metaboliske produkter (glykosider, alkaloider), nogle pigmenter (anthocyaniner).
Vacuole Vacuoles er enkeltmembranorganeller, der er "beholdere" fyldt med vandige opløsningerøkologiske og uorganiske stoffer. ER- og Golgi-komplekset deltager i dannelsen af vakuoler. Unge planteceller indeholder mange små vakuoler, som så, efterhånden som cellerne vokser og differentierer, smelter sammen og danner én stor central vakuole. Den centrale vakuole kan optage op til 95% af volumenet af en moden celle; kernen og organellerne skubbes mod celle membran. Membranbegrænsende plantevakuole, kaldet tonoplast. Væsken, der fylder plantevakuolen, kaldes cellesaft. En del cellesaft omfatter vandopløselige organiske og uorganiske salte, monosaccharider, disaccharider, aminosyrer, endelige eller toksiske metaboliske produkter (glykosider, alkaloider), nogle pigmenter (anthocyaniner).
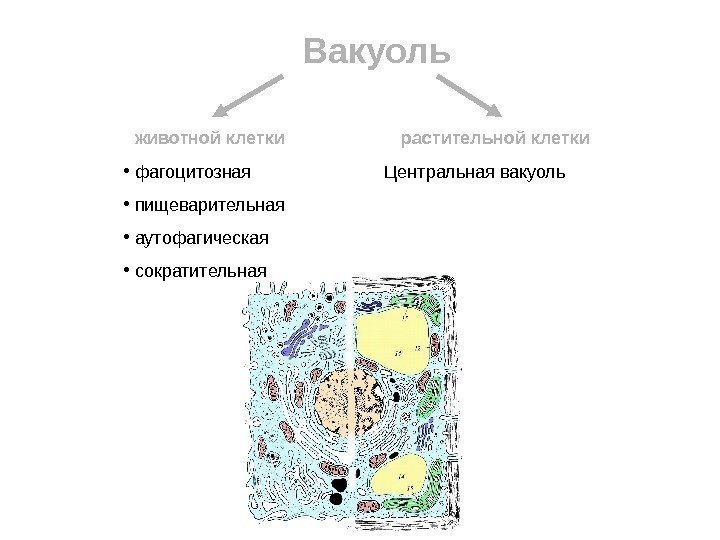 Vakuole dyrecelle plantecelle fagocytotisk fordøjelse autofagisk kontraktil Central vakuole
Vakuole dyrecelle plantecelle fagocytotisk fordøjelse autofagisk kontraktil Central vakuole
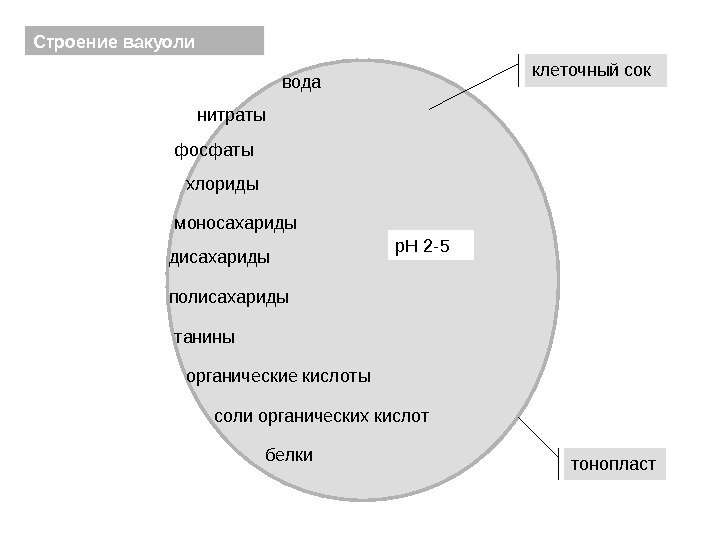 Vakuolens struktur tonoplastcelle saftvand nitrater fosfater chlorider monosaccharider disakkarider tanniner organiske syrer salte organiske syrer s. H2-5 polysaccharider proteiner
Vakuolens struktur tonoplastcelle saftvand nitrater fosfater chlorider monosaccharider disakkarider tanniner organiske syrer salte organiske syrer s. H2-5 polysaccharider proteiner
 Funktion nr. 1 Opretholdelse af turgortryk. Vakuolen fungerer som et osmometer og giver cellen den nødvendige styrke og tørhed. Funktion nr. 2 Nogle gange indeholder vakuoler opløselige pigmenter. Denne gruppe omfatter anthocyaniner, som er røde, blå eller lilla i farven, og nogle relaterede forbindelser, der er gule eller cremefarvede. Det er disse pigmenter, der hovedsageligt bestemmer farven på blomster. Ophobning af reservestoffer og "nedgravning" af affald, dvs. slutprodukterne af cellemetabolisme. Nogle gange ødelægger vakuoler stoffer, der er giftige eller unødvendige for cellen. Funktion nr.
Funktion nr. 1 Opretholdelse af turgortryk. Vakuolen fungerer som et osmometer og giver cellen den nødvendige styrke og tørhed. Funktion nr. 2 Nogle gange indeholder vakuoler opløselige pigmenter. Denne gruppe omfatter anthocyaniner, som er røde, blå eller lilla i farven, og nogle relaterede forbindelser, der er gule eller cremefarvede. Det er disse pigmenter, der hovedsageligt bestemmer farven på blomster. Ophobning af reservestoffer og "nedgravning" af affald, dvs. slutprodukterne af cellemetabolisme. Nogle gange ødelægger vakuoler stoffer, der er giftige eller unødvendige for cellen. Funktion nr.
 Peroxisomer er små vesikler, der indeholder et sæt enzymer. Funktioner: 1) Peroxisomer indeholder proteiner på overfladen af membranen, som fungerer som en receptor, der genkender signaler på det introducerede protein. Af alle peroximproteinerne er det bedst kendte enzym fra gruppen af hydroperoxidaser - katalase 2) Deltager i metaboliske reaktioner: i metabolismen af lipider, kolesterol mv.
Peroxisomer er små vesikler, der indeholder et sæt enzymer. Funktioner: 1) Peroxisomer indeholder proteiner på overfladen af membranen, som fungerer som en receptor, der genkender signaler på det introducerede protein. Af alle peroximproteinerne er det bedst kendte enzym fra gruppen af hydroperoxidaser - katalase 2) Deltager i metaboliske reaktioner: i metabolismen af lipider, kolesterol mv.
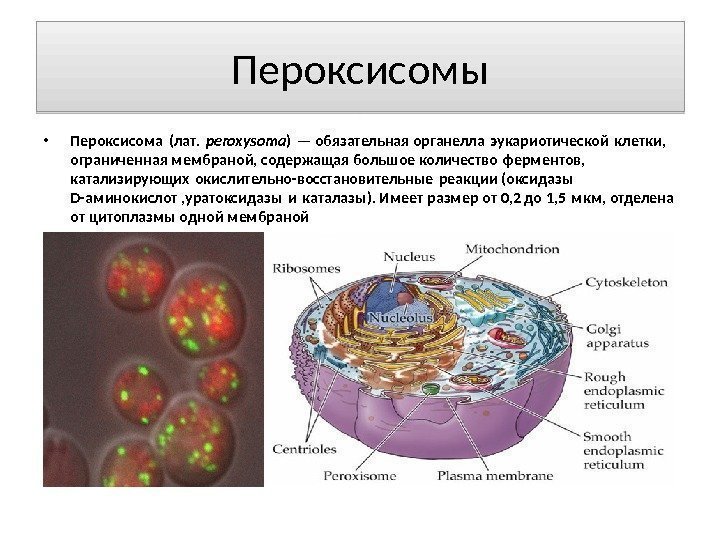 Peroxisomer Peroxisom (lat. peroxysoma) er en obligatorisk organel af en eukaryot celle, afgrænset af en membran, der indeholder et stort antal af enzymer, der katalyserer redoxreaktioner (D-aminosyreoxidaser, uratoxidase og katalase). Har en størrelse fra 0,2 til 1,5 mikron, adskilt fra cytoplasmaet med en membran
Peroxisomer Peroxisom (lat. peroxysoma) er en obligatorisk organel af en eukaryot celle, afgrænset af en membran, der indeholder et stort antal af enzymer, der katalyserer redoxreaktioner (D-aminosyreoxidaser, uratoxidase og katalase). Har en størrelse fra 0,2 til 1,5 mikron, adskilt fra cytoplasmaet med en membran
 Funktioner af peroxisomer Sættet af funktioner af peroxisomer varierer i celler forskellige typer. Blandt dem: oxidation af fedtsyrer, fotorespiration, ødelæggelse af giftige forbindelser, syntese af galdesyrer, kolesterol og esterholdige lipider, konstruktion af myelinskeden nervefibre, fytansyremetabolisme osv. Sammen med mitokondrier er peroxisomer hovedforbrugerne af O 2 i cellen. Peroxisomet indeholder normalt enzymer, der bruger molekylært oxygen til at abstrahere hydrogenatomer fra visse organiske substrater for at danne hydrogenperoxid: Catalase bruger det resulterende til at oxidere en række forskellige substrater - for eksempel phenoler, myresyre, formaldehyd og ethanol: Denne type oxidative reaktioner er især vigtig i lever- og nyreceller, hvis peroxisomer neutraliserer mange giftige stoffer kommer ind i blodbanen. Næsten halvdelen af den ethanol, der kommer ind i menneskekroppen, oxideres på denne måde til acetaldehyd. Derudover har reaktionen betydning for afgiftningen af cellen fra selve hydrogenperoxid.
Funktioner af peroxisomer Sættet af funktioner af peroxisomer varierer i celler forskellige typer. Blandt dem: oxidation af fedtsyrer, fotorespiration, ødelæggelse af giftige forbindelser, syntese af galdesyrer, kolesterol og esterholdige lipider, konstruktion af myelinskeden nervefibre, fytansyremetabolisme osv. Sammen med mitokondrier er peroxisomer hovedforbrugerne af O 2 i cellen. Peroxisomet indeholder normalt enzymer, der bruger molekylært oxygen til at abstrahere hydrogenatomer fra visse organiske substrater for at danne hydrogenperoxid: Catalase bruger det resulterende til at oxidere en række forskellige substrater - for eksempel phenoler, myresyre, formaldehyd og ethanol: Denne type oxidative reaktioner er især vigtig i lever- og nyreceller, hvis peroxisomer neutraliserer mange giftige stoffer kommer ind i blodbanen. Næsten halvdelen af den ethanol, der kommer ind i menneskekroppen, oxideres på denne måde til acetaldehyd. Derudover har reaktionen betydning for afgiftningen af cellen fra selve hydrogenperoxid.
 Ikke-membranorganeller Ribosomer findes i alle typer celler (inklusive prokaryote). De kan ligge frit i cytoplasmaet eller forbindes med membranerne i ER. Findes i mitokondrier og plastider. Struktur: Små sfæriske legemer dannet af to ulige underenheder - store og små, som består af 3-4 molekyler ribosomalt RNA og mere end 50 proteinmolekyler. Ribosomer indeholder altid magnesiumioner, der understøtter deres struktur. Funktioner: syntese af polypeptidkæder (den anden fase af proteinsyntesen er translation).
Ikke-membranorganeller Ribosomer findes i alle typer celler (inklusive prokaryote). De kan ligge frit i cytoplasmaet eller forbindes med membranerne i ER. Findes i mitokondrier og plastider. Struktur: Små sfæriske legemer dannet af to ulige underenheder - store og små, som består af 3-4 molekyler ribosomalt RNA og mere end 50 proteinmolekyler. Ribosomer indeholder altid magnesiumioner, der understøtter deres struktur. Funktioner: syntese af polypeptidkæder (den anden fase af proteinsyntesen er translation).
 Cellecenter Findes i næsten alle dyreceller (undtagen nogle typer protozoer) og nogle planter. Fraværende i blomstrende og nedre svampe. Struktur: Består af to centrioler placeret vinkelret på hinanden. Centriolen er en lille cylindrisk organel, hvis væg er dannet af 9 grupper (tripletter) af tre fusionerede mikrotubuli. Funktioner: deltager i dannelsen af fissionsspindlen (achromatinspindelen). Centrioler danner de basale kroppe af cilia og flageller.
Cellecenter Findes i næsten alle dyreceller (undtagen nogle typer protozoer) og nogle planter. Fraværende i blomstrende og nedre svampe. Struktur: Består af to centrioler placeret vinkelret på hinanden. Centriolen er en lille cylindrisk organel, hvis væg er dannet af 9 grupper (tripletter) af tre fusionerede mikrotubuli. Funktioner: deltager i dannelsen af fissionsspindlen (achromatinspindelen). Centrioler danner de basale kroppe af cilia og flageller.
 Mikrotubuli og mikrofilamentstruktur: Et komplekst system filamenter, der trænger ind i hele cytoplasmaet. Trådene er dannet af molekyler af forskellige kontraktile proteiner (myosin, tubulin osv.). Funktioner: sammen med nogle andre elementer danner de cellernes cytoskelet, sikrer intracellulær bevægelse af organeller, såvel som cellebevægelse, sammentrækning af muskelfibre danner trådene i den mitotiske spindel
Mikrotubuli og mikrofilamentstruktur: Et komplekst system filamenter, der trænger ind i hele cytoplasmaet. Trådene er dannet af molekyler af forskellige kontraktile proteiner (myosin, tubulin osv.). Funktioner: sammen med nogle andre elementer danner de cellernes cytoskelet, sikrer intracellulær bevægelse af organeller, såvel som cellebevægelse, sammentrækning af muskelfibre danner trådene i den mitotiske spindel
 Rød - kerne Grøn - mikrotubuli Gul - Golgi-apparat
Rød - kerne Grøn - mikrotubuli Gul - Golgi-apparat

Atlas: Menneskets anatomi og fysiologi. Komplet praktisk guide Elena Yurievna Zigalova
Membranorganeller. Transport over membraner
Menneskelige celler er karakteriseret ved tilstedeværelsen af et stort antal intracellulære membraner, der danner flere rum (fra det engelske rum - "compartment, compartment"), der adskiller sig fra hinanden i struktur og funktion: cytosol, kerne, endoplasmatisk reticulum, Golgi-kompleks, mitokondrier, lysosomer, peroxisomer. På grund af tilstedeværelsen af disse elementer forekommer et stort antal forskellige biokemiske reaktioner samtidigt i cellen.
Alle membranorganeller er bygget af elementære membraner, hvis strukturelle princip svarer til strukturen af plasmalemmaet beskrevet ovenfor. Absorptionen af makromolekyler og partikler af celler sker ved endocytose (fra den græske endon - "inde", kytos - "celle"), frigivelse - ved exocytose (fra den græske exo - "udenfor", kytos - "celle").
En af væsentlige funktioner Plasmalemmaet er transport. Lad os huske på, at de hydrofobe "haler" af lipider, der vender mod hinanden, forhindrer indtrængning af polære vandopløselige molekyler. Der er to typer transport: passiv og aktiv. Den første kræver ikke energi, den anden er energiafhængig. Som regel bærer den indre (cytoplasmatiske) overflade af membranen negativ ladning, hvilket letter indtrængning af positivt ladede ioner i cellen. Vand kommer ind i cellen ved osmose(fra det græske osmos - "skub, tryk"), som er den langsomme indtrængning af vand gennem en semipermeabel membran, der adskiller to opløsninger med forskellige koncentrationer. Som et resultat udlignes koncentrationerne af disse to opløsninger.
Diffusion(fra latin diffusion - "spredning, spredning") er overgangen af ioner eller molekyler forårsaget af deres Brownske bevægelse gennem membraner fra en zone, hvor disse stoffer er i en højere koncentration til en zone med en lavere koncentration, indtil koncentrationen er på begge sider af membranen er justeret. Specifikke transportproteiner indbygget i membranen transporterer små polære molekyler hen over den, hvor hvert protein transporterer én klasse af molekyler eller kun én forbindelse. Nogle transmembrane proteiner danner kanaler. Aktiv transport udføres af bærerproteiner, mens der forbruges energi som følge af hydrolyse af ATP (adenosintriphosphatsyre) eller protonpotentiale. Aktiv transport sker mod en koncentrationsgradient. For at udføre biokemiske reaktioner skal stoffer ind i cellen igennem endocytose og udskillelse af eksocytose metaboliske produkter.
Endocytose. Der er flere metoder til endocytose. Indgangen af flydende kolloide partikler kaldes pinocytose, og store partikler- fagocytose. For at eksterne molekyler kan komme ind i cellen, skal de først bindes af glycocalyx-receptorer. Cytolemmaet begynder at invaginere, så kommer dets kanter tættere og tættere og spalter vesiklen, der indeholder de fangede molekyler. Der dannes et endreom, som er nedsænket i cytoplasmaet og mødes med lysosomer. Deres membraner smelter sammen. I det resulterende sekundære lysosom bliver stoffer, der kommer ind i cellen, nedbrudt.
Exocytose sikrer fjernelse af store molekylære forbindelser. Først adskiller de sig i Golgi-komplekset i form af transportvesikler og ledes til celleoverfladen. Vesiklens membran er indlejret i cytolemmaet, og vesiklens indhold vises uden for cellen.
Der kendes to varianter endocytose: fagocytose - absorption af partikler (fra den græske phagos - "fortærende" og kytos - "celle") og pinocytose – absorption af opløste stoffer (fra græsk Pino - "drikke"). Den fagocyterede partikel indesluttet i en membran kaldes et fagosom. Under processen med endo- og exocytose er transporterede stoffer indesluttet i membranvesikler.
Endoplasmatisk retikulum eller endoplasmatisk retikulum(ER), er et enkelt kontinuerligt hulrum afgrænset af en membran, der danner mange invaginationer og folder ( se fig. 1). Derfor optræder det endoplasmatiske retikulum på elektrondiffraktionsmønstre i form af mange rør, flade eller runde cisterner og membranvesikler. Der er to typer ER: granulær og agranulær. Den side, der vender mod cytosolen af den første, er dækket af ribosomer, mens siden af den anden er blottet for dem. Funktion af granulær ER: syntese af proteiner ved ribosomer og transport af proteiner, glat syntese og udveksling af kulhydrater og lipider (steroidhormoner, glykogen, kolesterol) og neutralisering (hepatocytter), syntese af klorider, hvorfra det dannes i maven saltsyre. Da det er et depot af calciumioner, er glat ER involveret i muskelsammentrækning; afgrænser fremtidige blodplader i megakaryocytter. En af de vigtigste funktioner i ER er syntesen af membranproteiner og lipider for alle cellulære organeller.
Komplekse eller Golgi-apparater(CG), er et sæt af tanke, bobler, plader, rør, sække, afgrænset af en membran, hvori syntetiserede produkter akkumuleres og emballeres ( se fig. 1). Disse produkter fjernes fra cellen ved hjælp af de komplekse elementer, derudover syntetiseres polysaccharider, dannes protein-kulhydratkomplekser, og transporterede molekyler modificeres. I et lysmikroskop vises CG i form af et net eller et system af tubuli og vakuoler. CG er til stede i alle menneskelige celler, undtagen erytrocytter og hornskæl i epidermis. I de fleste celler er CG placeret omkring eller nær kernen. I CG identificeres tre membranelementer: fladtrykte sække (cisterner), vesikler og vakuoler. CG er en tredimensionel skålformet struktur bestående af flere (fra et til flere hundrede) diktyosomer (fra den græske dyktion - "netværk"). Hvert diktyosom indeholder 4-8 (i gennemsnit 6) parallelle flade cisterner, gennemtrængt af porer med udvidede ender, hvorfra vakuoler indeholdende syntetiserede stoffer spaltes fra. Cisternerne er forbundet med mange membranøse vesikler såvel som større sekretoriske granula. Elementer af Golgi-komplekset er forbundet med kanaler.
Golgi-kompleksets membraner dannes og vedligeholdes af granuleret endoplasmatisk retikulum, hvori membrankomponenter syntetiseres. De transporteres af transportvesikler, der knopper fra ER og smelter sammen med CG, hvorfra sekretoriske vesikler konstant knopper, og cisternernes membraner fornyes konstant. De leverer glycocalyx og syntetiserede stoffer til plasmalemmaet og sikrer dermed fornyelsen af plasmalemmaet. En af CG'ets vigtigste funktioner er proteinsortering.
Lysosomer- membranorganeller indeholdende omkring 50 typer af forskellige hydrolytiske enzymer, som syntetiseres på ribosomer af det granulære endoplasmatiske retikulum, hvorfra de overføres med transportvesikler til CG, hvor de modificeres. Primære lysosomer knopper fra overfladen af CG. Alle lysosomer i cellen danner et enkelt lysosomalt rum, hvor der konstant opretholdes et surt pH-miljø, der spænder fra 3,5-5,0. Lysosomers membraner er resistente over for enzymerne indeholdt i dem og beskytter cytoplasmaet mod deres virkning.
Der er fire funktionelle former lysosomer Primære lysosomer, spiret fra Golgi-komplekset, fusioneret med fagosomet, form sekundært lysosom(phagolysosom), hvor fordøjelsen af absorberede stoffer til monomerer finder sted. Sidstnævnte transporteres gennem den lysosomale membran ind i cytosolen. Ufordøjede stoffer forbliver i lysosomet, hvilket resulterer i dannelsen resterende krop. Derudover fordøjer lysosomer beskadigede strukturer i deres egen celle ( autolysosom).
Peroxisomer er vesikler med en diameter på 0,2 til 0,5 mikron, omgivet af membraner, der indeholder oxidative enzymer (ca. 40 % af alle proteiner er katalase), der producerer og ødelægger hydrogenperoxid. De bruger molekylær oxygen.
Mitokondrier, som er "cellens energistationer", er involveret i processerne med cellulær respiration og omdannelse af energi til en form, der er tilgængelig til brug for cellen. I et lysmikroskop fremstår mitokondrier som runde, aflange eller stavformede strukturer 0,3-5,0 µm lange og 0,2-1,0 µm brede. Antallet, størrelsen og placeringen af mitokondrier afhænger af cellens funktion og dens energibehov. Således når deres antal i hver levercelle 2500. Med hjælpen elektronmikroskopi Det er blevet fastslået, at mitokondrier er organeller med dobbeltmembraner ( ris. 5). Mellem de ydre og indre mitokondrielle membraner er der et intermembranrum. Den indre membran danner talrige folder, eller cristae, på grund af hvilke den indre membran øges dramatisk. På indre overflade cristae ligger mange elektrontætte submitochondriale elementære partikler(op til 4000 pr. 1 mikron 2 membraner), der har form som en svamp. I det rum, der er begrænset af den indre mitokondriemembran, er der en finkornet matrix.
Ris. 5. Mitokondrier (ifølge B. Alberts et al.; ifølge C. de Duve, som ændret). jeg – almindelig ordning bygninger: 1 – ydre membran; 2 - indre membran; 3 - cristae; 4 - matrix; II - diagram over strukturen af crista: 5 - fold af den indre membran; 6 – svampeformede kroppe
Mitokondrier indeholder deres eget DNA, RNA og ribosomer, som er placeret i matrixen. Således er mitokondrier udstyret med deres egne genetiske system, nødvendige for deres selvreproduktion og proteinsyntese. Det skal understreges, at mitokondrielt DNA, RNA og ribosomer adskiller sig fra cellens egne og minder meget om prokaryote.
OPMÆRKSOMHED
Hos pattedyr, herunder mennesker, arves mitokondriegenomet fra moderen.
Mitokondrier formerer sig ved at dele eksisterende, uanset delingen af andre mitokondrier og selve cellen.
Sker konstant i celler stofskifte(fra det græske stofskifte - "ændring, transformation") eller metabolisme, som er et sæt af processer assimilering(reaktioner af biosyntese af komplekse biologiske molekyler fra simplere) og dissimilation(spaltningsreaktioner). Som et resultat af dissimilering er energien indeholdt i kemiske bindinger stoffer. Denne energi bruges af cellen til at udføre forskellige jobs, herunder assimilering. Lad os huske på, at energi hverken skabes eller ødelægges, den går kun fra en form til en anden, egnet til at udføre arbejde. Cellen bruger den energi, der er indeholdt i de kemiske bindinger af aminosyrer, monosaccharider og fedtsyrer. De dannes som et resultat af fordøjelsen fra proteiner, kulhydrater og fedtstoffer og kommer ind i cellen.
Lad os overveje energimetabolisme ved at bruge nedbrydningen af glukose som et eksempel. Glukose transporteres igennem plasma membran, og dets iltfrie nedbrydning, eller glykolyse, sker i cytoplasmaet. Glykolyse er en flertrins enzymatisk proces, som et resultat af hvilken to molekyler pyrodruesyre og to ATP-molekyler dannes ud fra et glukosemolekyle (under hensyntagen til de to ATP-molekyler, der bruges til at udføre reaktionerne). Pyrodruesyre gennemgår yderligere oxidation (aerob med deltagelse af ilt) i mitokondrier, som indeholder enzymkæder, der katalyserer syntesen af ATP (adenosintrifosfat). ATP er en universel bærer og den vigtigste energiakkumulator i cellen. Energien er indeholdt i højenergibindinger mellem fosforsyrerester.
Når én fosfatgruppe fjernes fra ATP, dannes ADP (adenosin diphosphatsyre) og fosfat, og der frigives fri energi, som bruges af cellen til at udføre arbejde. I mitokondrier kombineres ADR med en phosphorsyrerest for at danne ATP. Som et resultat af glykolyse frigives kun omkring 5% af energien; resten frigives i mitokondrier i processen med aerob oxidation og lagres i ATP. Per et glukosemolekyle dannes 36 ATP-molekyler.
Kerne - vigtigste cellestruktur, er til stede i alle humane celler undtagen erytrocytter og blodplader. I de fleste celler er dens form sfærisk eller ægformet, men andre former for kernen findes også (ringformet, stangformet, spindelformet, perleformet, bønneformet, segmenteret, polymorf osv.). Størrelsen af kernerne varierer meget fra 3 til 25 mikron. Ægcellen har den største kerne. De fleste menneskelige celler er mononukleære, men der er binukleære (for eksempel nogle neuroner, hepatocytter, cardiomyocytter), og nogle strukturer er multinukleære (myosymplast muskelfibre).
I kernen er der følgende strukturer: nuklear kappe, kromatin, nukleolus og nukleoplasma. Kernen er omgivet af en nuklear kappe, bestående af en indre og ydre kernemembran, hver 8 nm tyk, adskilt af et perinukleært rum (eller kernehylster cisterne) 20-50 nm bredt. Begge er elementære cellemembraner. Ribosomer er knyttet til den ydre, som passerer ind i det granulære endoplasmatiske retikulum. Det perinukleære rum danner et enkelt hulrum med det endoplasmatiske retikulum ( se fig. 1).
Kernehylden gennemtrænges af mange ordnede runde kerneporer med en diameter på 50–70 nm, som Total optager op til 25% af kernens overflade. Gennem kerneporer sker der selektiv transport af store partikler, samt udveksling af stoffer mellem kerne og cytosol.
I levende celler er karyoplasma (nukleoplasma) homogen (undtagen nukleolus). Efter fiksering og behandling af væv til lys- eller elektronmikroskopi bliver to typer kromatin synlige (fra det græske chroma - "maling"); velfarvet heterochromatin (inaktivt) og let euchromatin (aktivt).
I den delende kerne spiraler kromatin og danner kromosomer. Kromatinet i den ikke-delende kerne og kromosomerne i den delende kerne er dannet af deoxyribonukleinsyre (DNA) forbundet med RNA og proteiner (histoner og ikke-histoner). Den kemiske identitet af kromatin og kromosomer bør understreges.
I somatiske celler Der er to kopier af hvert kromosom, de kaldes homologe. De er identiske i længde, form, struktur, stribearrangement og bærer de samme gener, som er lokaliseret på samme måde. Normal karyotype menneske (fra det græske karyon - "kerne af en nød", stavefejl - "prøve") inkluderer 22 par autosomer og et par kønskromosomer (XX kvinde eller XV mand) ( ris. 6).

Ris. 6. Menneskelig karyotype (sund mand). I – karyotype, generel form; II - metafase kromosomer
Gene en sektion af DNA karakteriseret ved en specifik nukleotidsekvens, der er ansvarlig for syntesen af et specifikt protein. Genet er den elementære enhed for arv.
Nukleolus (en eller flere) detekteres i alle ikke-delende kerner i form af en tæt, intenst farvende afrundet homogen basofil krop, hvis størrelse er proportional med intensiteten proteinsyntese. Ribosomer dannes i nukleolus. Kernejuice er den ikke-farvede elektron-lyse del af kernen - en kolloid opløsning af proteiner, der omgiver kromatin og nukleolus.
Fra bogen Sjove øvelser for tungen forfatter Olga NovikovskayaHVAD ER TRANSPORTERNE? Se, flyet letter. Flyet flyver, nynner, en modig pilot sidder i det. Bid den brede tungespids med tænderne og udtal lyden [L] i lang tid, mens du løfter dine arme til siderne og svinger dem som vinger. Nå, åh det her
Fra bogen Sjove øvelser for tungen forfatter Olga NovikovskayaHVAD ER TRANSPORTEN Se, flyet letter. Flyet flyver, nynner, en modig pilot sidder i det. Bid den brede tungespids med tænderne og udtal lyden [L] i lang tid, mens løfte dine arme til siderne og svinge dem som vinger. Nå, denne faldskærm. Ned os
Fra bogen Medicinsk Fysik forfatter Vera Aleksandrovna Podkolzina Fra bogen Histologi forfatter V. Yu. Barsukov1. Introduktion til histologisk forløb. Celleorganeller Histologi er videnskaben om strukturen, udviklingen og vitale funktioner af levende organismers væv. Følgelig studerer histologi et af niveauerne for organisering af levende stof - væv. Histologiens hovedobjekt i systemet
Fra bogen Histologi forfatter V. Yu. Barsukov4. Celleorganellers morfologi og funktioner (fortsat) Lysosomer er cytoplasmaets mindste organeller, de er legemer afgrænset af en bilipid membran Lysosomers funktion er at sikre intracellulær fordøjelse, dvs. nedbrydning af både eksogene og
Fra bog Normal fysiologi forfatter Nikolay Alexandrovich AgadzhanyanGasudveksling og gastransport Gasudveksling af O2 og CO2 gennem alveolær-kapillærmembranen sker gennem diffusion, som sker i to trin. I det første trin sker diffusionsoverførslen af gasser gennem den lufthæmatiske barriere, i det andet trin sker det
Fra bog Levende vand. Hemmeligheder bag cellulær foryngelse og vægttab forfatter Lyudmila RudnitskayaVI STYRKER CELLEMEMBRANER MED ØVELSER Om fordelene fysisk træning meget er blevet sagt. Regelmæssig træning forbedrer karsystemets funktion, normaliserer blodtryk og sukkerniveauer, opbygger muskelmasse og forbrænder fedt. Det er under træningen, at vores
Fra bog Hemmelig visdom menneskelige legeme forfatter Alexander Solomonovich ZalmanovMembraner Liv er den evige bevægelse af væsker mellem celler og indeni celler. At stoppe denne bevægelse resulterer i døden. En delvis opbremsning i bevægelsen af væsker i et organ forårsager delvis uorden. Generel opbremsning af ekstra- og intracellulære væsker i
Efter hundrede år er jeg en ung 20-årig jeg er Guds Ånd, ifølge Guds hellige vilje bliver jeg hele tiden sundere og stærkere, mine nerver bliver hele tiden sundere og stærkere, jeg udvikler mig konstant, accelererer gudgiven konstant helbredelse og foryngelse, jeg bliver altid 20 år gammel. Jeg er selv efter fem år 20 år gammel. Mig og igennem
Fra bogen Alt åndedrætsøvelser. For sundheden for dem, der bekymrer sig... forfatter Mikhail Borisovich IngerleibKapitel 3. Transport af gasser med blod "Bærer" af ilt fra lungerne til væv og organer og carbondioxid fra væv og organer til lungerne er blod. I en fri (opløst) tilstand overføres en så lille mængde gasser, at den sikkert kan negligeres, når
Fra bog Godt syn- klart sind lange år! Østens ældste praksis forfatter Andrey Alekseevich LevshinovTræk vejret ind gennem næsen, ånd ud gennem munden Lær at trække vejret med abdominal vejrtrækning, så du trækker vejret ind gennem næsen og ånder ud gennem munden (munden er kun lidt åben, læberne er foldet som et lille rør) - blødt, glat, uden
Fra bogen The Science of Breathing of Indian Yogis forfatter William Walker AtkinsonKapitel VI TRÆKNING GENNEM NÆSEVÆRE OG VEJLEDNING GENNEM MUNDEN En af de første lektioner i yogiernes videnskab om vejrtrækning er viet til at lære at trække vejret gennem næsen og vinde sædvanlig vane- trække vejret gennem munden Den menneskelige åndedrætsmekanisme gør det muligt for ham at trække vejret gennem både næse og mund, men for ham er sagen sand
Fra bogen Conspiracies of a Siberian healer. Udgave 33 forfatter Natalya Ivanovna Stepanova Fra Giris bog. Sport af stærk og sund forfatter Alexey Ivanovich VorotyntsevBlok af kast over hovedet (kast over hovedet) Kast over hovedet - kaste en vægt over hovedet og derefter modtage den med samme hånd med en rotation af torsoen 180°. Alle kast over hovedet udføres med en torsodrejning på 180°, så når man kalder et kast, drejes