Название тканей отражает их положение – они покрывают тело растений и располагаются на границе с внешней средой. Характерной особенностью покровных тканей является то, что они состоят из плотно сомкнутых живых или мертвых клеток. Покровные ткани выполняют прежде всего барьерную функцию и предохраняют внутренние ткани растения от пересыхания и повреждения. К важнейшим функциям покровных тканей относятся также регуляция транспирации и газообмена. В дополнение к основным функциям покровные ткани способны всасывать и выделять вещества, препятствовать проникновению патогенных микроорганизмов. Таким образом, покровные ткани в определенной мере многофункциональны.
В эволюционном плане покровные ткани имеют очень дре внее происхождение, они возникли как следствие перехода растений к жизни на суше. Во всяком случае, тело первых известных наземных растений (риниофитов) было покрыто эпидермой с устьицами.
В онтогенезе современных растений покровные ткани возникают из поверхностных слоев апикальных меристем – из протодермы или дерматогена в результате антиклинальных делений.
В зависимости от происхождения и времени появления различают 3 типа покровных тканей – первичные, вторичные и третичные. К первичным тканям, образующимся в результате дифференцировки клеток первичных меристем, относятся эпидерма (эпидермис, кожица), которая покрывает листья и молодые зеленые стебли, части цветка, плоды, ризодерма (включая эпиблему), покрывающая молодой корешок. Эпидерма (от греч. epi – сверху и derma – кожа) дифференцируется из протодермы. Чаще всего она образована одним слоем тесно сомкнутых клеток, без межклетников (исключая устьичные щели), нередко с извилистыми стенками, что усиливает прочность сцепления. В удлиненных органах растения основные клетки эпидермы вытянуты вдоль продольной оси (на стебле, черешках); на листе – более или менее соответствуют его очертаниям. Наружная поверхность клеток эпидермы обычно утолщена и часто покрыта слоем кутикулы или, восковым налетом различной толщины. Эпикутикулярный воск, откладываясь в виде гладкого слоя, либо палочек и нитей, поднимающихся над поверхностью, создает беловатый или голубоватый налет на листьях и плодах некоторых растений (фикус, слива, яблоня и др.). И кутикула, и воск синтезируются протопластом клеток. Утолщение наружных стенок, наличие кутина и воска не только снижают транспирацию органов, но и защищают растение от проникновения вирусов, бактерий и гиф грибов. У ряда растений (злаков, осок, хвощей) в наружной стенке откладывается кремнезем. У хвойных растений, некоторых злаков, у купены оболочки основных клеток эпидермы одревесневают. Клетки эпидермы многих семян содержат полисахариды, которые при увлажнении набухают и образуют слизи. Семена при этом легко приклеиваются к почве или к движущимся предметам, что способствует их фиксации или распространению.
Как правило, клетки эпидермы живые. Они содержат тонкий постенный слой цитоплазмы с ядром, а иногда и небольшое количество фотосинтетически малоактивных хлоропластов, а также лейкопласты. Эпидермальные клетки лепестков и сочных плодов содержат хромопласты. В крупной центральной вакуоли могут накапливаться вещества вторичного метаболизма – алкалоиды, эфирные масла, таннины, пигменты, оксалат кальция и др.
Нередко клетки эпидермы образуют поверхностные выросты – трихомы (волоски). Они могут быть одноклеточными и многоклеточными, простыми и разветвленными, звездчатыми и головчатыми и др. Трихомы формируют опушение. В функциональном отношении трихомы делят на кроющие и железистые. Одни из них остаются живыми (узамбарские фиалки), другие утрачивают живое содержимое и оказываются мертвыми. Кроющие трихомы выполняют функции защиты растения от перегрева (листья мать-и-мачехи), от избыточной транспирации (многие растения пустынь), от повреждения животными и насекомыми. Эпифитные растения, например, из семейства Бромелиевые используют трихомы для поглощения из воздуха паров воды и минеральных веществ. В самом молодом возрасте кроющие трихомы выполняют функцию транспирации. Железистые трихомы обычно сохраняют живое содержимое клеток. Фактически, их можно рассматривать как выделительные структуры, способные накапливать и выделять в окружающую среду или под кутикулу продукты метаболизма (эфирные масла, смолы, токсичные для растения вещества и др.). У насекомоядных растений выделяются пищеварительные ферменты (росянка, непентес). Нередко железистые волоски заканчиваются одной или несколькими выделительными клетками, или образуют головку (крапива, томат, пеларгония, подорожник, ромашка аптечная и др.). Основная функция железистых волосков связана с выведением токсичных солей из тканей листа (например, у лебеды – Atriplex), защитой органа от перегрева в результате выделения летучих эфирных масел, химической и отчасти механической защитой растений от насекомых. Разнообразное строение трихом иногда используют в качестве диагностического признака в систематике, при анализе лекарственного растительного сырья.
Кроме волосков на эпидерме некоторых растений образуются выросты, называемые эмергенцами (от лат. emergentis – выбивающийся). В их образовании принимают участие не только клетки эпидермы, но и слои клеток, лежащие под ней. Нередко эмергенцы представляют собой довольно крупные образования, похожие на колючки. Однако в отличие от колючек, на побеге растения они расположены без определенного порядка, сдираются вместе с эпидермой. Эмергенцами являются шипы розы, малины, ежевики, шипы на плодах дурмана, эхиноцистиса, многих зонтичных, конского каштана. Эмергенцы выполняют в основном защитную функцию. Таким образом, перечисленные особенности строения эпидермы позволяют защищать молодые органы растений от неблагоприятных факторов внешней среды.
Важнейшей функцией эпидермы является активная регуляция транспирации и газообмена. Эта функция осуществляется через устьичный аппарат, или устьичный комплекс, состоящий из устьица и функционально связанных с ним околоустьичных (побочных) клеток. Устьице представляет собой образование из двух замыкающих клеток и устьичной щели между ними, которая является межклетником. Замыкающие клетки двудольных чаще всего имеют бобовидную форму. Вогнутые, обращенные к щели (вентральные = брюшные = внутренние) стенки клеток утолщены значительно сильнее, чем выпуклые (дорсальные = спинные = внешние) стенки. Кроме того, на них образуются кутикулярные выросты, в результате чего устьичная щель имеет вид небольшого воронковидно-расширенного на концах канала. Наружное расширение перед входом в щель называют передним двориком, внутреннее, за щелью – задним двориком. Под замыкающими клетками расположена подустьичная, или воздушная, камера, где скапливаются пары воды, углекислый газ или кислород. В замыкающих клетках устьиц есть хлоропласты, но отсутствуют плазмодесмы, что, в известной мере, свидетельствует об их сравнительной изолированности от остальных клеток эпидермиса.
Устьица работают в автоматическом режиме, что определяется рядом морфологических и физиологических факторов. Механизм работы устьиц не вполне ясен.
Регулирование ширины устьичной щели, и, следовательно, интенсивности испарения, происходит в результате изменения тургорного давления в замыкающих клетках устьиц. При избытке в растении воды клетки устьица активно (т. е. против градиента концентрации, с затратой энергии) оттягивают от окружающих клеток и органоидов самой замыкающей клетки одновалентные катионы, прежде всего калия. Это ведет к повышению осмотического давления в клетках устьица, их объем и давление на оболочку увеличиваются за счет всасывания воды из окружающих клеток. Микроструктура оболочек замыкающих клеток устьица играет решающую роль в изменении очертаний (движении) клеток и открывании устьичной щели. Целлюлозные микрофибриллы, из которых состоит клеточная оболочка, расположены таким образом, что стенка, обращенная к отверстию, менее эластична, а некоторые волокна образуют своеобразные «обручи» вокруг замыкающих клеток. По мере всасывания воды и повышения тургесцентности, эти «обручи» не дают клетке расширяться, но позволяют удлиняться. Второй фиксатор находится на концах замыкающих клеток, там, где они соединяются друг с другом. Увеличивающееся тургорное давление заставляют внешние (дорсальные) более эластичные стенки отодвигаться. При этом радиальные мицеллы вовлекают в это движение внутренние (вентральные) стенки устьичных клеток. В результате устьичная щель открывается. При закрывании устьиц происходит пассивный отток ионов калия по градиенту концентрации. Осмотическое давление снижается, вода уходит из замыкающих клеток, объем вакуолей уменьшается, растянутые тонкие стенки клеток спадаются, устьичная щель смыкается. Источником энергии для работы устьичного аппарата служит гидролиз запасных углеводов, накапливающихся в процессе фотосинтеза в замыкающих клетках.
Разные растения выработали свой ритм работы устьичного аппарата. У большинства растений устьица открыты и днем, и ночью. В жару устьица обычно закрыты. При значительном насыщении организма водой (например, в результате длительных дождей) устьичные клетки набухают, их сдавливают окружающие клетки эпидермы, в результате устьичная щель также закрывается.
Количество и распределение устьиц варьируют в зависимости от вида растений и условий жизни. Число их колеблется от нескольких десятков до нескольких тысяч на 1 мм 2 поверхности эпидермы. Общая площадь устьичных отверстий составляет не более 1–2 % площади листа. Однако транспирация при открытых устьичных щелях эквивалентна испарению примерно с 70 % соответствующей открытой водной поверхности. При закрытых устьицах транспирация резко снижается, испарение воды происходит только через клеточные стенки и кутикулу. Таким образом, эпидерма весьма эффективно регулирует газообмен и транспирацию.
При развитии устьиц (у покрытосеменных растений) материнская клетка, или клетка-предшественница (меристемоид) замыкающих клеток, образуется в результате неравного деления протодермальной клетки и является меньшей из двух образовавшихся дочерних клеток. Эта меньшая клетка в свою очередь делится и образует две замыкающие клетки. Межклетное вещество между ними набухает, растворяется и образуется устьичная щель. Одновременно формируются и околоустьичные клетки.
По способу образования в онтогенезе растения выделяют три типа устьичных аппаратов: перигенный , мезогенный и мезо-перигенный . При формировании перигенного устьичного аппарата меристемоид делится и образует только пару замыкающих клеток. Такое устьице окружено обычными основными клетками меристемы, как у ириса, пеларгонии. При развитии мезогенного аппарата меристемоид дает начало и замыкающим, и всем околоустьичным клеткам (злаки, гвоздичные, крестоцветные). Если меристемоид дает начало замыкающим клеткам устьица и хотя бы одной побочной клетке, при этом другие околоустьичные клетки образуются из иных меристематических клеток, говорят об образовании мезо-перигенного аппарата. На основании числа побочных клеток и их расположения относительно длинной оси устьичной щели выделяют типы устьичных аппаратов. Они имеют определенную топографию (внешний вид и расположение), достаточно постоянны и поэтому могут использоваться для целей систематики. В фармакогнозии типы устьичных аппаратов используют для диагностики подлинности растений при их микроанализе. Изучением устьичных аппаратов занимается наука стоматография (от лат. stoma – рот). Классификацию устьичных аппаратов предложили английские ботаники К. Меткаф и Л. Чок в 1950 г.
Аномоцитный (от греч. anomos – беспорядочный и kitos – клетка), или беспорядочно-клетный тип, встречается у разных групп высших растений, за исключением хвощей. Побочные клетки в этом случае отсутствуют или не отличаются от остальных клеток эпидермы. Чаще встречается у более примитивных покрытосеменных – у растений из сем. Лютиковые, Гераниевые.
Анизоцитный (от греч аnisos – неравный), или неравноклеточный тип обнаружен только у цветковых растений. Замыкающие клетки окружены тремя побочными, одна из которых либо заметно крупнее, либо заметно мельче остальных. Чаще встречается у растений сем. Крестоцветные (капуста), а также Толстянковые (очиток, молодило).
Парацитный (от греч. para – рядом), или параллельноклеточный тип, характеризуется двумя побочными клетками, расположенными параллельно длинной оси устьица. Характерен для покрытосеменных растений сем. Мареновые (подмаренник, ясменник), Маревые, Мятликовые (Злаки). Встречается у хвощей, папоротников, гнетовых.
Диацитный (от греч. di – две), или перекрестноклеточный тип также, имеет две околоустьичные клетки, расположенные перпендикулярно длинной оси устьица. Характерен для покрытосеменных растений сем. Губоцветные (мята, шалфей и др.), Гвоздичные (гвоздика, звездчатка). Встречается у папоротников (нефролепис).
Тетрацитный (от греч. tetra – четыре) тип встречается главным образом у покрытосеменных однодольных растений. Устьице окружено четырьмя симметрично расположенными околоустьичными клетками: две из них параллельны устьичной щели, а две другие примыкают к полюсам замыкающих клеток (традесканция).
Энциклоцитный тип найден у ряда покрытосеменных, голосеменных и папоротников. Побочные клетки образуют узкое кольцо вокруг замыкающих клеток.
Замыкающие клетки устьиц могут располагаться на одном уровне с основными эпидермальными клетками либо находиться выше их или значительно ниже (например, погруженные устьица в листе сосны).
Если наземные органы растения покрыты эпидермой, то первичной покровной тканью корня является ризодерма (от греч. rhyza – корень и derma – кожа) или эпиблема, образующая корневые волоски. Она возникает из дерматогена – наружных клеток апикальной меристемы корня вблизи корневого чехлика и покрывает молодые окончания корней. Это функционально одна из важнейших тканей, т. к. именно через нее происходит поглощение воды и минеральных веществ из почвы, т. е. осуществляется корневое питание растения. По своему строению эпиблема близка к эпидерме, но она имеет и свои особенности, связанные с местоположением и основной функцией. В эпиблеме отсутствуют устьица. Клетки эпиблемы тонкостенны, имеют первичное строение, содержат большую вакуоль и более вязкую цитоплазму; в связи с активным всасыванием богаты митохондриями. В них отсутствуют хлоропласты, нет и кутикулы. Потенциально каждая клетка эпиблемы способна к образованию выроста – корневого волоска. Однако нередко эту функцию выполняют лишь некоторые специальные клетки, получившие название трихобласты. Корневые волоски обычно одноклеточные, не отделены, в отличие от трихом эпидермы, клеточной перегородкой, существуют короткое время. Достигая в длину 1–2 мм, корневые волоски живут 15–20 дней, а затем отмирают.
Первичная покровная ткань существует разный период времени и только у активно растущих органов. У травянистых растений эпидерма сохраняется до конца жизни. В стеблях многолетних древесных растений (деревьев и кустарников), а также на корнях голосеменных и двудольных растений, для которых характерно вторичное утолщение, на смену первичным покровным тканям формируется вторичная – перидерма (от греч. peri – возле, около). Обычно перидерма образуется на годичных побегах к концу вегетационного периода, но иногда формируется и на второй (вереск, некоторые рододендроны) или третий год жизни растения (толокнянка). Она берет начало от особой вторичной образовательной ткани – феллогена , или пробкового камбия. Феллоген стебля возникает чаще всего из субэпидермальных клеток основной паренхимы, которая сохраняет слабую меристематическую активность. Иногда феллоген возникает в самой эпидерме (у ив, груши, рябины, калины). У корней, а также в стеблях некоторых растений (малины, шиповника, иван-чая), феллоген нередко образуется из перицикла.
Феллоген может закладываться как отдельными участками, которые впоследствии сливаются, так и цельным кольцом по всей окружности осевого органа. Клетки его имеют таблитчатую форму. Они делятся периклинально, параллельно поверхности органа, последовательно отделяя ковнутри клетки феллодермы , или пробковой кожицы, а кнаружи – клетки феллемы , или пробки. Поскольку при каждом делении клеток феллогена одна из дочерних сохраняет меристематические свойства, а вторая дифференцируется в клетку постоянных тканей (феллему или феллодерму), м ежду феллемой и феллодермой сохраняется длительно функционирующий слой феллогена. Клетки феллогена образуют больше клеток феллемы, нежели клеток феллодермы. В итоге феллодерма состоит чаще всего из 1–3 слоев живых паренхимных клеток, содержащих запасные вещества и выполняющих трофическую функцию по отношению к феллогену. Слой феллемы толще. Ее клетки плотно сомкнуты между собой, строго таблитчатой формы, без межклетников, расположены вертикальными рядами. Молодые клетки феллемы живые, однако, впоследствии, часто еще до окончания роста клетки, на первичную оболочку изнутри откладывается суберин, иногда в чередовании с воском. Так как в стенках клеток феллемы пор нет, протопласт опробковевших клеток отмирает. Их полости заполняются воздухом либо смолистыми или дубильными веществами, имеющими темную окраску, поэтому зеленый цвет побегов переходит в бурый. У березы оболочки клеток феллемы пропитаны бетулином, который придает ветвям белый цвет и к тому же обладает антисептическими свойствами. Клетки пробки водо- и газонепроницаемы, хорошо предохраняют внутренние ткани растения от высыхания, колебаний температуры, болезнетворных организмов. Особенно важна перидерма для растений, живущих в условиях сезонных изменений. На стволах некоторых деревьев, например, пробкового дуба, амурского бархата, образуется мощный слой пробки, достигающий нескольких сантиметров толщины. Таким образом, перидерма является сложной комплексной тканью, в состав которой входят как живые (феллоген, феллодерма), так и отмирающие клетки (феллема). При образовании перидермы эпидерма отмирает и слущивается.
Обладая перечисленными свойствами, перидерма хорошо изолирует внутренние ткани от окружающей среды. Вместе с тем живые ткани нуждаются в газообмене и удалении избытка влаги. Поэтому при образовании перидермы первыми начинают формироваться чечевички – специальные образования, которые обеспечивают проветривание внутренних тканей. Феллоген прежде всего закладывается под устьицами (но не под каждым) там, где идут процессы газообмена и транспирации. Образующиеся в результате работы феллогена клетки имеют форму, близкую к изодиаметрической, расположены рыхло, благодаря наличию больших межклетников. Совокупность этих клеток называется выполняющей, или заполняющей тканью. По межклетникам этой выполняющей ткани происходит нерегулируемые (в отличие от устьиц) газообмен и транспирация. В феллогене, который подстилает выполняющую ткань, также есть узкие межклетники, поэтому он не мешает газообмену. С наступлением холодного сезона феллоген откладывает под выполняющей тканью замыкающий слой , состоящий из более мелких, опробковевающих и теснее сомкнутых клеток и чечевички «закрываются». Весной при активизации работы феллогена этот слой под напором новых клеток разрывается и чечевички вновь начинают функционировать. Чечевички обычно выступают над перидермой и имеют вид бугорков различной формы, характерной для разных видов растений. Например, у осины, тополя чечевички ромбовидной формы, у бузины они округлые, у березы линейно вытянуты по окружности ствола и имеют вид черных черточек на белом фоне. Чечевички хорошо заметны на клубнях картофеля, на плодах яблонь и груш. В корнях чечевичек обычно нет.
У большинства древесных растений перидерма осевых органов со временем заменяется ритидомом , или коркой – третичной покровной тканью. Она формируется в результате многократного последовательного заложения новых слоев перидермы в глубже лежащих тканях первичной коры. В результате ткани, заключенные между перидермами, лишаются снабжения водой и питательными веществами, в них откладываются конечные продукты обмена веществ (смолы, танины, оксалаты кальция, алкалоиды и др.), образуются склереиды. Со временем эти ткани отмирают, образуя мощный покровный комплекс. Изнутри корка ежегодно прирастает, а с поверхности растрескивается, разрушается и сбрасывается. Различают ритидом кольчатый и чешуйчатый. Кольчатый ритидом образуется в том случае, если перидермы закладываются более или менее непрерывным слоем. В результате корка снимается со ствола как чулок или расщепляется на длинные полосы (например, у виноградной лозы, клематиса, жимолости, земляничного дерева, кипариса, вишни). Однако чаще встречается чешуйчатый ритидом, при формировании которого, новые перидермы закладываются в виде отдельных перекрывающихся фрагментов. Подобный ритидом образуется у сосны, груши, яблони.
Образование ритидома происходит у разных видов растений в разном возрасте: у виноградной лозы на втором году жизни, у яблони и груши – на седьмой – восьмой год, у сосны в 8–10 лет, у дуба в 25–30 лет, у граба – через полвека. Ритидом обеспечивает защиту деревьев от перепадов температуры, солнечных ожогов, от низовых пожаров, повреждений микроорганизмами, животными и др.
Эпидерма, перидерма и ритидом относятся к сложным тканям, т. к. состоят из разных типов клеток.
Конец работы -
Эта тема принадлежит разделу:
РАСТЕНИЙ, ЗНАЧЕНИЕ МОРФОЛОГИИ
ИСТОРИЯ РАЗВИТИЯ МОРФОЛОГИИ... РАСТЕНИЙ ЗНАЧЕНИЕ МОРФОЛОГИИ КАК НАУЧНОЙ... ТКАНИ РАСТЕНИЙ...
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ:
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
Первичная покровная ткань.
Кожицу листьев и стеблей называют эпидермой
, кожицу корня – эпиблемой
.
Основные функции эпидермы – защита молодых органов от высыхания, механическая защита и газообмен. Эпидерма
, как правило, представлена одним слоем плотно сомкнутых
клеток, на внешней поверхности жироподобное вещество кутин образует защитную пленку – кутикулу. На поверхности кутикулы часто имеется восковой налет. Стенки клеток обычно извилистые, наружные стенки толще остальных.
Зпиблема, ризодерма (от греческого epiblema - покрывало, derma - кожа и rhiza - корень) - первичная покровная ткань корня, выполняющая функции защиты и поглощения питательных веществ. На поверхности клеток образуются боковые выросты – корневые волоски. Устьица и кутикула у эпиблемы отсутствуют.
Экзодерма (от экзо... и греч. derma - кожа) - часть первичной коры, корня, одно- или многоклеточное образование, находящееся непосредственно под первичной покровной тканью - эпиблемой . Особенно характерна для однодольных растений, у которых со временем заменяет эпиблему. Клетки экзодермы остаются живыми, хотя их стенки опробковевают. В экзодерме иногда встречаются пропускные клетки, хотя есть данные, что транспорт веществ может осуществляться и непосредственно через экзодерму.
Для газообмена и транспирации в эпидерме имеются специальные образования – устьица
.
Устьице
представляет собой щелевидное отверстие в эпидерме, ограниченное двумя клетками бобовидной формы. Это замыкающие клетки. В отличие от остальных клеток эпидермы они содержат хлоропласты
. Стенки замыкающих клеток, обращенные в сторону устьичной щели, утолщены. Клетки эпидермы, окружающие замыкающие, называют побочными или прилегающими. Под устьицем находится газовоздушная камера. Замыкающие и побочные клетки, устьичная щель и газовоздушная камера образуют устьичный аппарат. Устьица чаще
располагаются на нижней стороне листа
.
Иногда клетки эпидермы образуют различные придатки, волоски и чешуйки (трихомы). Волоски выполняют защитную функцию, сильное опушение защищает растение от перегрева и потери влаги. Железистые волоски выполняют защитную функцию (например, у крапивы).
Вторичная покровная ткань
, перидерма
.
Перидерма
(от пери... и греч derma - кожа) - вторичная покровная ткань стеблей и корней, корневищ и клубней, редко листьев и плодов. Иногда рассматривается как комплекс тканей. Образуется из вторичной меристемы - феллогена. Состоит из:
феллемы
– собственно пробки
,
феллогена
– пробкового камбия
и
феллодермы
– пробковой паренхимы (внутренняя живая ткань)
…
Из-за изолирующего многослойного пробкового покрова перидерма непроницаема для воды и воздуха. Обмен с внешней средой осуществляется через чечевички - прорывы в перидерме, образующиеся над устьицами. Перидерма сменяет эпидерму, которая постепенно отмирает и слущивается. Закладывается преимущественно в стеблях и корнях.
Третичная покровная ткань
Ткань, образующаяся из вторичной ткани в результате происходящих в ней новообразовательных процессов (напр., заложение прослоек камбия в глубже расположенной вторичной ткани).
К третичной ткани относится ребристая корка
на поверхности толстых стволов древесных растений.
Корка (ритидом)
, третичная покровная ткань. При образовании корки новый слой феллогена и перидермы закладывается в основной ткани, лежащей глубже первой наружной перидермы. Вновь образовавшиеся слои пробки отчленяют к периферии органа не только перидерму, но и часть лежащей под ней паренхимы коры. Так возникает толстое многоклеточное и мертвое образование. Так как корка не может растягиваться, при утолщении ствола она лопается и образуются трещины.

Корка (ритидом)
образуется у многолетних растений в корне, стебле, корневище.
Каждый год в более глубоких слоях закладывается новый слой феллогена и образуется перидерма. Наружный слой перидермы - пробка - изолирует все вышележащие ткани, в результате чего они отмирают. Таким образом, совокупность многочисленных перидерм с отмершими между ними тканями и является коркой.
Ботаника-наука о растенях.
Отлич.черты раст.клетки
Пластиды
3.Тканями
Образовательные ткани.
Ботаника-наука о растенях.
Ботаника охватывает широкий круг проблем: закономерности внешнего и внутреннего строения (морфология и анатомия) растений, их систематику, развитие в течение геологического времени (эволюция) и родственные связи (филогения), особенности прошлого и современного распространения по земной поверхности (география растений), взаимоотношения со средой (экология растений), сложение растительного покрова (фитоценология, или геоботаника), возможности и пути хозяйственного использования растений (ботаническое ресурсоведение, или экономическая ботаника).
По объектам исследования в ботанике выделяют фикологию (альгологию) - науку о водорослях, микология - о грибах, лихенологию - о лишайниках, бриологию - о мхах и др.; изучение микроскопических организмов, преимущественно из мира растений (бактерий, актиномицетов, некоторых грибов и водорослей), выделяют в особую науку - микробиологию. Болезнями растений, вызываемыми вирусами, бактериями и грибами, занимается фитопатология.
Ботаника пользуется как наблюдением, так и сравнительным, историческим и экспериментальным методами, включающими сбор и составление коллекций, наблюдение в природе и на опытных участках, эксперимент в природе и в условиях специализированных лабораторий, математическую обработку полученной информации. Наряду с классическими методами регистрации тех или иных признаков изучаемых растений используется весь арсенал современных химических, физических и кибернетических методов исследования.
Отлич.черты раст.клетки
Природа и основные функции веществ различны. Главнейшие из этих веществ: простые белки; некоторые углеводы, в частности глюкоза, сахароза и крахмал или близкий к нему инулин, а также целлюлоза, запасные жиры и жироподобные вещества - соединения первичного метаболизма; продукты вторичного метаболизма - танниды, полифенольные соединения, алкалоиды, изопренпроизводные и др.относится также обычный во многих растениях оксалат кальция Почти все вещества независимо от их природы в той или иной мере могут вновь вовлекаться в процессы активного метаболизма клетки.Важнейшая группа - запасные вещества. Это белки,углеводы (исключая целлюлозу) и жиры.Целлюлоза и близкие к ней по химической природе гемицеллюлозы и пектиновые вещества - структурные компоненты клеточной стенки. Вторичные метаболиты выполняют различные функции, но главнейшая из них связана с "организацией" защиты от поедания растений животными, внедрения болезнетворных микроорганизмов и т.д. Таким образом, они являются группой защитных веществ. Оксалат кальция чаще всего рассматривается в качестве конечного продукта метаболизма, формы "захоронения" ненужных или вредных клетке веществ. В технике, пищевой промышленности и медицине широко используются крахмал, глюкоза и сахароза. Танниды, или дубильные вещества, соединения полифенольные и изопренпроизводные - основа для получения большинства медицинских препаратов растительного происхождения. Пектиновые вещества широко применяются в кондитерской промышленности, а также в медицине. Некоторые из эргастических веществ крайне ядовиты. Чаще это алкалоиды, некоторые гликозиды, полипептиды (у бледной поганки). Значение запасных веществ для фармации и медицины.
Крахмал, в медицине и фармации для приготовления присыпок, паст (густых мазей), а также при производстве таблеток. Целлюлоза, велико и промышленное значение целлюлозы – из этого вещества изготовляют хлопчатобумажные ткани и бумагу.Клетки выделительных тканей по форме обычно паренхимные и тонкостенные. Они долго остаются живыми, выделяя секрет. Клетки-идиобласты по мере накопления большого количества секрета лишаются протопласта и стенки их нередко опробковевают.Ткани внутренней секреции могут быть представлены отдельными клетками-идиобластами, вместилищами выделений, смоляными ходами, эфирномасляными каналами и млечниками. Пластиды - бесцветные или окрашенные тельца в протоплазме растительных клеток, представляющие собой сложную систему внутренних мембран и выполняющие различные функции. Бесцветные пластиды называют лейкопластами, различно окрашенные - хромопластами, зеленые - хлоропластами.
Только в растительных клетках присутствуют пластиды. Эти мембранные органеллы в зависимости от окраски можно разделить на лейкопласты, хромопласты и хлоропласты. В клетке высших растений содержится около 40 хлоропластов в которых происходит фотосинтез. Как и митохондрии, хлоропласта содержат собственную ДНК и рибосомы. Они способны к автономному размножению, не зависящему от деления клетки. В зависимости от окраски, связанной с наличием или отсутствием тех или иных пигментов, различают три основных типа пластид:
Хлоропласты (зеленого цвета), - хромопласты (желтого, оранжевого или красного цвета) и - лейкопласты (бесцветные).
Самые большие из них - хлоропласты - достигают у высших растений 4-10 мкм длины и хорошо различимы в световой микроскоп. Хлоропласты встречаются во всех зеленых органах растений, лейкопласты весьма обычны в клетках органов, скрытых от солнечного света, - корнях, корневищах, клубнях, а также в ситовидных элементах некоторых покрытосеменных. Хромопласты содержатся в клетках лепестков многих растений, зрелых окрашенных плодах (томаты, шиповник, рябина), иногда - в корнеплодах (морковь).
Строение пластид может быть рассмотрено на примере хлоропластов. Они имеют оболочку, образованную двумя мембранами: наружной и внутренней. Внутренняя мембрана вдается в полость хлоропласта немногочисленными выростами. Мембранная оболочка отграничивает от гиалоплазмы клетки матрикс хлоропласта, так называемую строму. Как строма, так и выросты внутренней мембраны формируют в полости хлоропласта сложную систему мембранных поверхностей, отграничивающих особые плоские мешки, называемые тилакоидами, или ламеллами. Группы дисковидных тилакоидов связаны друг с другом таким образом, что их полости оказываются непрерывными. Эти тилакоиды образуют стопки (наподобие стопки монет), или граны. Тилакоиды стромы объединяют граны между собой. В мембранах тилакоидов сосредоточен главнейший пигмент зеленых растений - хлорофилл и вспомогательные пигменты - каротиноиды. Внутренняя структура хромопластов и лейкопластов проще. Граны в них отсутствуют.
В строме хлоропластов содержатся ферменты и рибосомы, отличающиеся от рибосом цитоплазмы меньшими размерами. Часто имеются один или несколько небольших зерен первичного ассимиляционного крахмала. Генетический аппарат хлоропластов автономен, они содержат свою собственную ДНК.
Основная функция хлоропластов - фотосинтез. Центральная роль в этом процессе принадлежит хлорофиллу, точнее - нескольким его модификациям. Световые реакции фотосинтеза осуществляются преимущественно в гранах, темновые - в строме хлоропласта. И хлоропласты, и митохондрии способны синтезировать собственные белковые молекулы, так как обладают собственной ДНК.
Помимо фотосинтеза, в хлоропластах осуществляется синтез АТФ и АДФ (фосфорилирование), синтез и гидролиз липидов, ассимиляционного крахмала и белков, откладывающихся в строме.
В лейкопластах пигменты отсутствуют, но здесь может осуществляться синтез и накопление запасных питательных веществ, в первую очередь крахмала, иногда белков, редко жиров. Очень часто в лейкопластах формируются зерна вторичного запасного крахмала.
Красноватая или оранжевая окраска хромопластов связана с присутствием в них каротиноидов. Считается, что хромопласты - конечный этап в развитии пластид, иначе говоря, это стареющие хлоропласты и лейкопласты. Наличие хромопластов отчасти определяет яркую окраску многих цветков, плодов и осенних листьев.
3.Тканями
называются группы клеток, которые имеют одинаковое происхождение, сходное строение и выполняют в организме одинаковую функцию.
Каждый тип ткани отличается характерным строением клеток, строго соответствующим той функции, которую данная ткань выполняет.
В соответствии с формой, строением и характером соединения клеток ткани могут быть рыхлые и плотные, живые и мертвые, паренхимные и прозенхимные. Все разнообразие растительных тканей принято подразделять на 6 главных групп: 1) образовательные, или меристемы, 2) основные, 3) покровные, 4) механические, 5) проводящие и 6) выделительные.Образовательные ткани.
Меристемы или образовательные ткани, обладают способностью к делению и образованию новых клеток. За счет меристем формируются все прочие ткани и осуществляется длительный рост растения. У животных меристемы отсутствуют, чем объясняется ограниченный период их роста. Клетки меристем отличаются высокой метаболической активностью. Одни клетки меристем, получившие название инициальных, задерживаются на эмбриональной стадии развития в течение всей жизни растения, другие постепенно дифференцируются и превращаются в клетки различных постоянных тканей.
Первичные меристемы способны к делению изначально. В ряде случаев способность к активному делению может вновь возникнуть и у клеток, уже почти утративших это свойство. Такие "вновь" возникшие меристемы называют вторичными.
В теле растения меристемы занимают различное положение, что позволяет их классифицировать. По положению в растении выделяют верхушечные, или апикальные,боковые, или латеральные, и интеркалярные меристемы.
Апикальные меристемы располагаются на верхушках осевых органов растения и обеспечивают рост тела в длину, а латеральные - преимущественно рост в толщину. Каждый побег и корень, а также зародышевый корешок, почечка зародыша имеют апикальную меристему. Апикальные меристемы первичны и образуют конусы нарастания корня и побега.
Латеральные меристемы располагаются параллельно боковым поверхностям осевых органов, образуя своего рода цилиндры, на поперечных срезах имеющие вид колец. Часть из них относится к первичным. Первичными меристемами являются прокамбий и перицикл, вторичными - камбий и феллоген.
Интеркалярные, или вставочные, меристемы чаще первичны и сохраняются в виде отдельных участков в зонах активного роста (например, у оснований междоузлии, в основаниях черешков листьев).
Покровные ткани, классификация, функции.
Они предохраняют органы растения от высыхания, от температурных воздействий, механических повреждений и других неблагоприятных воздействий окружающей среды. При изменениях возраста органов и их функций, покровные ткани закономерно сменяют одна другую. По происхождению различают первичные (эпидерма, ризодерма, веламен), вторичные (перидерма) и третичные (корка или ритидом) покровные ткани.
Первичные покровные ткани Эпидерма образуется из поверхностного слоя апикальной меристемы - протодермы. Она покрывает листья, плоды, части цветка и молодые стебли. Кроме защитной функции, эпидерма регулирует процессы транспирации и газообмена, принимает участие в синтезе различных веществ и др. В состав эпидермы входит несколько морфологически различных клеток: основные клетки эпидермы, замыкающие и побочные клетки устьиц, трихомы (выросты эпидермы). Клетки эпидермы живые, имеют ядра, лейкопласты, вакуоли, хлоропласты (только в замыкающих клетках устьиц). Эпидерма у большинства растений однослойная, реже многослойная. Клетки первичной покровной ткани плотно примыкают друг к другу, и не имеют межклетников. С наружной стороны вся эпидерма покрыта сплошным слоем кутикулы (прерывается только над устьичными щелями).
Ризодерма (эпиблема) образована апикальной меристемой корня. Она покрывает молодые корневые окончания и именно через ризодерму происходит поглощение воды и минеральных солей из почвы. Кроме того, она взаимодействует с микроорганизмами почвы, из корня в почву выделяются вещества, помогающие почвенному питанию. Клетки ризодермы имеют очень тонкие оболочки. У первичной покровной ткани корня нет кутикулы, вследствие чего эти клетки имеют оболочки легко проницаемые для воды. На небольшом расстоянии от кончика корня образуются корневые волоски - выросты ризодермы.
Веламен, как и ризодерма, происходит из поверхностного слоя апикальной меристемы корня. Эта своеобразная ткань покрывает корни эпифитов и некоторых других растений, приспособленных к жизни на периодически пересыхающих почвах (аспидистра, аспарагус, алоэ, кливия). Веламен от ризодермы отличается многослойностью. Протопласт веламена отмирает и поэтому всасывает воду не осмотическим, а капиллярным путем.
Эмергенцы (нем., единственное число Emergenz, от лат. emerge - появляюсь, выхожу), выросты на поверхности стеблей и листьев растений, образованные не только эпидермисом, но и глубже лежащими тканями (в отличие от волосков ). Это - жгучие волоски крапивы, шипы на стеблях розы и на плодах дурмана, железки на листьях росянки и др.Вторичная покровная ткань - Перидерма. Перидерма сложная, многослойная вторичная покровная ткань стеблей и корней многолетних(реже однолетних) растений 1 . Перидерма сменяет первичные покровные ткани осевых органов, которые постепенно отмирают и слущиваются. У большинства двудольных и голосемянных перидерма обычно появляется в течение первого вегетативного периода в тех частях корня и побега, которые прекратили рост в длину. В основе перидермы стебля лежит вторичная меристема - феллоген. Феллоген стебля чаще возникает из клеток основной паренхимы, лежащей под эпидермой и сохранившей слабую меристематическую активность. У корней основой образования феллогена может быть перицикл. Феллоген может закладываться отдельными участками, позднее сливающимися, либо сразу по всей окружности осевого органа. Прежде всего образуется инициальный ряд уплощенных по форме таблитчатых клеток. Позднее инициальные клетки периклинально делятся, формируя два ряда меристематических клеток. Внутренний ряд дифференцируется как феллодерма (живые паренхим-ные клетки) и далее не делится. Наружный - в ходе серии периклинальных делений формирует различное количество слоев клеток пробки, длительное время, сохраняя меристематическую активность. Феллоген способен делиться также и антиклинально, обеспечивая правильное соотношение тканей в разрастающихся органах. Пробка, феллоген и феллодерма образуют перидерму. По мере того как формируется перидерма, зеленый цвет побегов переходит в бурый. «Вызревшие» к осени побеги первого года способны выдержать зимние морозы. Пробка состоит из отмерших клеток, лишенных межклетников. Их оболочки пропитаны суберином. Клетки пробки воздухо- и водонепроницаемы. Многослойная пробка образует защитный футляр, предохраняющий живые ткани от потери влаги, от резких температурных колебаний и проникновения болезнетворных микроорганизмов. На стволах и сучьях некоторых деревьев (пробковый дуб, бархат амурский) формируется мощный слой пробки, достигающий нескольких сантиметров толщины. Такую пробку используют для промышленного получения укупорочных материалов. Живые ткани, лежащие под пробкой, нуждаются в газообмене и удалении избытка влаги. Поэтому в перидерме довольно рано образуются чечевички - отверстия, «заполненные» рыхлой выполняющей тканью, состоящей из закругленных паренхимных слабоопробковевших клеток с многочисленными межклетниками. Через чечевички осуществляется «проветривание» стебля. Чечевички, Имеющие вид небольших бугорков различной формы, хорошо заметны на поверхности молодых побегов деревьев и кустарников. Чечевички в виде мелких крапин заметны и на некоторых плодах, например яблоках и грушах. По мере утолщения побегов форма чечевичек меняется. На стволах березы их остатки заметны в виде характерных поперечных полосок и черточек. Перидерма образуется, как сказано, у двудольных и хвойных растений. У однодольных (пальмы, драцены, алоэ) иногда путем многократного деления и опробковения периферических клеток первичной коры образуется защитная ткань функционально сходная с перидермой. В многолетних осевых органах двудольных и хвойных растений обычно развивается несколько перидерм. Каждая последующая закладывается глубже предыдущей. Со временем живые клетки наружных перидерм и располагающихся между ними тканей отмирают, образуя мощный покровный комплекс - корку.Она формируется на стволах, многолетних ветвях и корнях деревьев. В отличие от более или менее гладкой перидермы корка нередко трещиноватая и неровная (чешуйчатая пробка). Существует, правда, и относительно гладкая, так называемая кольцевая корка (например, у винограда). Эти различия связаны с особенностями заложения внутренних перидерм. Толстая корка надежно предохраняет стволы деревьев от механических повреждений, лесных низовых пожаров, резкой смены температур, ожогов прямыми солнечными лучами, проникновения патогенных микроорганизмов. Обычно у древесных пород корка начинает закладываться в зрелом возрасте: у сосны в 8-10 лет, у дуба в 25-30 лет. Ежегодно наращиваясь, с годами она может достигать значительной толщины.
Под покровной тканью следует понимать комплекс клеток, расположенных снаружи всех органов растения.
При попытке дать покровной ткани более точное определение, соответствующее характеру каждого отдельного случая, (встречается ряд трудностей. Покровные ткани различных органов весьма разнообразны по строению и по функциям. Покровные ткани надземных органов - листа, стебля - служат для защиты от высыхания и повреждения внутренних, более нежных тканей, выполняют выделительную функцию, при их посредничестве осуществляется газообмен с окружающей средой и пр. Покровы же корней, особенно окончаний их, поглощают воду с растворенными в ней минеральными веществами, содействуют растворению некоторых твердых частиц почвы и защищают внутренние ткани корня от механических повреждений. Покровы осевых органов у многолетних растений к концу первого вегетационного периода в большинстве случаев начинают сильно изменяться: покровы первого года вегетации отмирают, а из них или из других тканей, расположенных в непосредственной близости, образуется покровная ткань другого типа как по структуре, так и по ряду дополнительных функций. Следовательно, необходимо различать первичные покровные ткани и вторичные покровные ткани . Последние или возникают вновь, или представляют собой видоизменение структуры первичной ткани.
Первичная покровная ткань всех растительных органов называется кожицей, или эпидермисом .
Эпидермис образуется из поверхностного слоя первичной меристемы, называемого протодермой . Эпидермис состоит из клеток, как правило, изодиаметрического характера и всегда с живым содержимым. При отмирании протопласта эпидермальных клеток последние перестают служить покровом и защитой находящимся за ними тканям и заменяются другой покровной тканью.
Если рассматривать эпидермальные клетки снаружи, то можно убедиться, что очертания их в разных органах и у разных растений весьма различны. Очертания эпидермальных клеток формируются в большой зависимости от соотношения в скорости роста органа в целом и его поверхности. Например, клетки эпидермиса листа с широкой пластинкой, одинаково разрастающейся как в длину, так и в ширину, имеют извилистые очертания; а клетки эпидермиса стебля молодого растения, сильно вытягивающегося в длину, значительно более вытянуты и имеют менее извилистые стенки, чем клетки эпидермиса листа того же растения. Линейные листья однодольных растений отличаются длинными эпидермальными клетками.
Клетки эпидермиса весьма прочно соединены друг с другом, значительно прочнее, чем с клетками подстилающей эпидермис ткани. Вследствие этого эпидермис часто легко отрывается от прочих тканей, но сам не разрывается и сдирается относительно большими лоскутами. Эпидермис обычно не имеет межклетников, но бывают и исключения. Например, межклетники встречаются в эпидермисе лепестков. Кроме того, в эпидермисе других надземных, преимущественно зеленых, органов имеются щелевидные межклетники, окруженные специальными замыкающими их клетками, - устьица .
В молодых эпидермальных клетках почти вся полость клетки заполнена протопластом с отчетливо видным округлым клеточным ядром и достаточно заметными, хотя и мелкими, лейкопластами. По мере разрастания клетки и увеличения размеров вакуолей цитоплазма занимает постенное положение, и ядро, если оно расположено не в центре клетки, а в цитоплазме, прижатой к оболочке, сплющивается и вытягивается.
Во вполне выросшей эпидермальной клетке вакуоля занимает почти всю клетку, она наполнена клеточным соком, бесцветным или окрашенным растворенными в нем пигментами. Протопласт в виде очень тонкого слоя, нередко с трудом различимого, прижат к оболочке. Часто в такой клетке пластиды разрушаются, совершенно исчезая. Это наблюдается преимущественно у светолюбивых травянистых растений. Впрочем и у таких растений иногда в эпидермальных клетках нижней стороны листа есть хлорофилловые зерна. У растений, произрастающих в тени, хлоропласты часто имеются во всех эпидермальных клетках. У некоторых растений пластиды в эпидермисе листьев сохраняются в виде мелких шаровидных лейкопластов, группирующихся вокруг клеточного ядра.

В эпидермальных клетках образуются различные продукты жизнедеятельности протопласта. Особенно своеобразны цистолиты. , они представляют собой весьма объемистые отложения углекислой извести, возникающие в гипертрофически разросшихся клетках эпидермиса (идиобластах, в данном случае - литоцистах). Если подействовать на эти цистолиты кислотой, например, соляной, то основная масса цистолита растворится и останется лишь целлюлозная основа, прикрепленная к оболочке клетки, на которой откладывается углекислый кальций. Форма и размеры цистолитов у различных растений различны. Они свойственны определенным видам растений. В нашем примере приводится цистолит в листьях конопли. Классическим примером служат цистолиты в листьях фикуса.
Оболочка клеток эпидермиса, особенно наружная стенка их, часто пропитывается или солями кальция, или соединениями кремния (злаки, хвощи). На поверхности эпидермальных клеток очень часто выделяются кутикула и кутикулярные слои. Кутикула затягивает всю поверхность сформировавшегося эпидермиса, прерываясь только устьичными щелями. Кроме кутикулы, снаружи ее у большинства растений выделяется воск в виде тонкого мелкозернистого налета. У некоторых тропических растений восковой покров на листьях достигает значительной толщины (0,5 см). И кутикула и восковой налет не смачиваются водой и предохраняют ткани, расположенные под ними, от потери воды.
Условия произрастания определяют некоторые особенности структуры эпидермальных тканей растения. Эпидермис листьев и стеблей водных растений почти не имеет кутикулы и тем более воскового налета, а у растений засушливых местообитаний кутикула, как правило, образуется. Оболочки эпидермальных клеток надземных органов некоторых растений одревесневают, становятся весьма толстыми, с сильно сократившимися размерами клеточных полостей. При этом часто одревесневают не только клетки самого эпидермиса, но и подстилающие его клетки. Поверхность эпидермиса надземных органов очень редко остается гладкой и однородной в течение всего времени существования этой ткани. Даже на кутикуле почти всегда возникают бугорки, складки, штрихи, придающие поверхности различный, иногда довольно прихотливый рисунок. Кроме того, клетки эпидермиса часто образуют разнообразнейшие выросты - волоски . Волоски бывают одноклеточными и многоклеточными, ветвистыми и неветвистыми. Форма волосков также варьирует: существуют волоски звездчатые, головчатые, в виде чешуек, прицепок и др. Форма, размеры и распределение волосков на поверхности органов характерны для определенных групп растений, и эти признаки часто служат систематическими.
Обычно волоски отделяются перегородкой от той эпидермальной клетки, на которой они образовались. Но есть волоски, представляющие собой даже во вполне развитом состоянии не более как выросты клеток эпидермиса, не отделяющиеся от них специальной перегородкой. Если такие выросты эпидермальных клеток очень короткие, то их называют сосочками . Сосочки особенно распространены на лепестках цветков, придавая им бархатистость.
Многоклеточные волоски нередко чрезвычайно сильно разрастаются, составляющие же их клетки одревесневают и волоски становятся твердыми. Такие волоски называют шипами (малина, старые стебли тыквы, стебли некоторых сортов розы, барбариса и др.).
Все многообразие волосков, руководствуясь физиологическим принципом, можно свести к двум основным типам: кроющие и железистые . Очень часто волоски обоих типов существуют на одном и том же органе. Например, на рисунке изображены простые кроющие и железистые волоски прицветной чешуйки рудбекии.

Кроющие волоски остаются живыми сравнительно недолго. Когда протопласт волоска отмирает, остов его, состоящий из оболочки, сохраняется, наполняясь воздухом. Обычно эти наполненные воздухом блестящие белые остовы волосков и образуют столь характерный волосяной покров тела растений.
Морфологическое разнообразие кроющих волосков представлено на рисунке, на котором изображены волоски частей цветка растений из семейства норичниковых. Все волоски за исключением двух одноклеточные.

Железистые волоски сохраняют живое содержимое своих клеток несколько дольше кроющих. Протопласт клеток железистых волосков, особенно верхушечных, образующих головку волоска клеток, нередко выделяется значительной вакуолизацией и почти всегда содержит крупное клеточное ядро. Если протопласт клеток железистого волоска разрушается, то нежные оболочки их спадаются и волосок, засохнув, отпадает, не оставляя после себя почти никакого следа. Морфология железистых волосков так же, как и кроющих, беспредельно разнообразна. Обычно выделения железистых волосков состоят или из смолистых веществ, или же из эфирных масел, сахаристых веществ, слизей, а иногда просто из воды и растворенных в ней веществ различной природы.
Железистые волоски, покрывающие, например, почечные чешуйки и кроющие листья почек, выделяют смолистое вещество, как бы закупоривающее почку и предохраняющее нежные листочки от высыхания. Возможно, что выделения железистых волосков являются конечными продуктами сложных биохимических процессов, протекающих в растительных организмах.
Железистые волоски, выделяющие воду и имеющие специально приспособленную для этого структуру, называют гидатодами .
Среди железистых волосков особенно своеобразны по своей организации так называемые жгучие волоски, общеизвестным примером которых являются жгучие волоски крапивы. Стенки волоска крапивы пропитаны соединениями кремния, вследствие чего становятся очень ломкими, особенно у верхушки. Такой волосок обламывается от легчайшего соприкосновения, причем кончик волоска превращается в острую иглу, легко вонзающуюся в кожу животного или человека. Такие же волоски с еще более жгучим секретом образуются на различных частях Alonsoa. В большинстве других случаев оболочки железистых волосков нежные, даже если они и выделяют раздражающий кожу секрет, как это имеет место, например, у некоторых видов борщевика.
Значение волосков разнообразно и во многих случаях еще не выяснено. Принято считать, что наиболее распространенной функцией волосков типа кроющих является предохранение поверхности тела растения от излишней потери воды. Блестящие белые волоски, с одной стороны, отражают прямые солнечные лучи, с другой - густым сплетением создают над эпидермисом среду, способствующую уменьшению транспирации. Однако специальными опытами по крайней мере в отношении частей цветка показано, что покрывающие их жизнедеятельные волоски увеличивают интенсивность транспирации.
Обычно ознакомление с различными волосками основано на изучении волосков, покрывающих уже выросшие органы. За исключением железистых волосков, описываются волоски мертвые, лишенные живого содержимого. В живом состоянии такие волоски, несомненно, более связаны с процессами жизнедеятельности растительного организма и играют иную физиологическую роль, чем отмершие сухие волоски. Подтверждением этому положению является развитие волосков на колосках злаков. На молодых цветочных и колосковых чешуях колоса, когда он еще находится в стадии так называемой трубки и окончательно еще не оформился, по краям чешуй, вдоль средней жилки и на молодой ости, если колос будет остистым, расположены волоски с хорошо выраженным крупным клеточным ядром и густой цитоплазмой. Эти волоски пребывают в наиболее активном жизненном состоянии. Из всех частей колоска колосковая и цветковая чешуи развиваются в первую очередь, а волоски на них формируются раньше всех прочих клеток, составляющих ткани этих чешуй. Волоски на чешуях колоска остаются в деятельном состоянии во все время формирования завязи и зародышевого мешка в ней. Как только зародышевый мешок готов к оплодотворению или, в других случаях, когда оплодотворение произойдет, содержимое волосков начинает быстро отмирать. Волоски, расположенные вдоль средней жилки и ости, успевшие значительно разрастись, превращаются в шипики и бугорки, претерпевают одревеснение и придают колосу хорошо известную на ощупь шероховатость.
Таким образом, волоски на начинающем развиваться колоске злака в своем деятельном состоянии связаны с важнейшими процессами в жизни растения - формированием и созреванием полового аппарата, во время которых идет активнейший обмен веществ.
Форма волосков на молодых колосковых чешуях разнообразна.

На молодых завязях различных растений часто образуется много волосков, в содержимом которых имеется алкалоид или какое-либо другое физиологически активное вещество. Все эти волоски в период формирования завязи находятся в жизнедеятельном состоянии, а после оплодотворения начинают отмирать и отпадают. На рисунке изображены железистые волоски завязи девясила; позднее, после оплодотворения, они быстро исчезают. Ясно, что волоски, столь хорошо и в изобилии развивающиеся на поверхности завязи и отпадающие после оплодотворения, в своем развитии связаны с интенсивно идущими в завязи процессами формирования женских половых элементов.

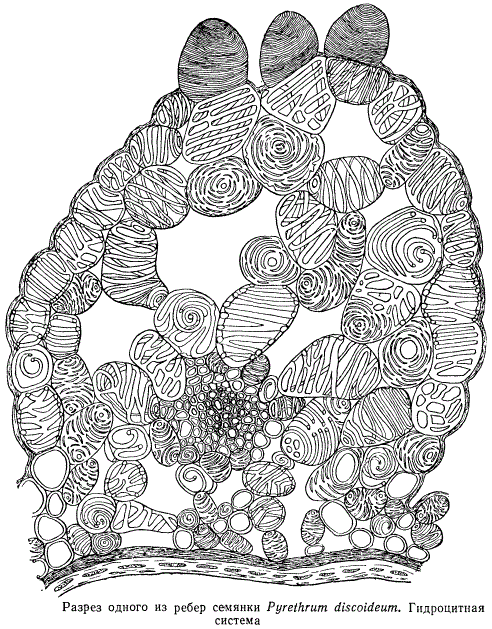
На семянках одной из ромашек, произрастающих в пустыне, можно найти своеобразные волоски, изображенные на рисунке. Каждый волосок состоит из двух морфологически хорошо отличающихся частей: основания, состоящего из нескольких члеников - отдельных клеток, и верхушки, занятой одной ослизняющейся клеткой. Каждый членик основания снабжен целлюлозными тяжами утолщений оболочки, образующими изящный переплет - подобие каркаса. На рисунке все три волоска представляют собой различные стадии ослизнения конечной клетки. Ослизняющиеся клетки и волоски на поверхности плодов некоторых растений, произрастающих в пустынных местностях, обособляются относительно очень рано, они хорошо различимы на далеко еще не зрелых плодах. Часто такие клетки в виде коротких толстых волоскоподобных образований располагаются на верхушках ребер семянок, причем при посредстве особых клеток, называемых гидроцитами , оболочки которых снабжены разнообразными утолщениями, наружные волоскоподобные клетки сообщаются с сосудистыми пучками.

Таким образом, создается своеобразная гидроцитная система, играющая большую роль в водном режиме созревающего плода.
По созревании семянки волоскоподобные гигроскопические образования засыхают и остаются на поверхности в виде небольших бугорков и бородавочек или опадают, или же превращаются в ослизняющиеся клетки.
Эпидермис образуется не только снаружи органа, но и внутри, если в органе формируется специальная полость. Например, ткань, выстилающая полость плодов, по своему происхождению идентична эпидермису, покрывающему плоды снаружи. В связи с нахождением покровной ткани внутри полости, где влажность воздуха и прочие условия отличаются от условий внешней среды, особенности структуры внутреннего эпидермиса плода несколько иные, чем наружного эпидермиса.
Клетки внутреннего эпидермиса плода гороха соединены между собой рыхло, кутикула или очень тонка или отсутствует. Значительная часть клеток вытянута в волоски различной длины с тонкими оболочками. Местами и особенно около семян образуется густой пушок из сосочкоподобных волосков, облекающий семя со всех сторон. То же наблюдается в плодах конских бобов, только покров, снабженный волосками, отличается более густым и равномерным скоплением последних.
Несомненно, образование таких сосочкоподобных волосков, соприкасающихся с развивающимися семенами, имеет какое-то физиологическое значение в процессе развития семян.
У некоторых растений оболочки внутреннего эпидермиса плода довольно значительно утолщаются и иногда одревесневают, особенно ко времени окончательного созревания плода. Хорошим примером такого строения внутреннего эпидермиса плода является эпидермис, покрывающий изнутри стенки коробочек мака. Волоски в таких случаях развиваются не всегда и никогда не достигают значительной длины; кутикула выражена более заметно, поверхность эпидермиса часто бывает гладкой, клетки имеют утолщенные оболочки и плотно сомкнуты между собой, иногда среди них встречаются устьица.
Таким образом, роль волосков и подобных им образований в жизни растительных организмов весьма разнообразна и меняется в зависимости от условий произрастания и возраста растений.
Продолжительность существования эпидермиса на надземных побегах невелика и обычно ограничивается одним вегетационным периодом. Но есть исключения и не столь редкие, особенно у растений более южных районов. Например, листья лавровишни и лавра существуют больше одного года, следовательно, и эпидермис их сохраняется больше одного года. Есть деревья, у которых эпидермис на побегах сохраняется несколько лет, даже на толстых стволах многолетних деревьев. Примерами служат эвкалипт, платан, стеркулия, а на севере - серая ольха, черемуха и др. Многолетен эпидермис также на хвое сосны, ели, туи и др.
На поверхности многолетних эпидермисов развивается относительно толстая кутикула. У хвойных оболочки эпидермальных клеток одревесневают.
В отличие от надземных частей растений ткань, покрывающая молодые части корней, лишена кутикулы вследствие чего клетки этой ткани имеют оболочки, легко проницаемые для воды. Кроме того, у большинства растений при соответствующих условиях клетки, покрывающие конечные участки корней, образуют тонкие выросты, не отделяющиеся от образовавших их клеток перегородками. Эти клетки называются корневыми волосками . Корневые волоски у различных растений бывают различной длины, но в общем длина их колеблется от 0,15 до 8,0 мм. На корнях проростков гороха, выращенных во влажной атмосфере, длина корневых волосков равна 2,5 мм, у конских бобов - 0,8 мм. У одного и того же растения в зависимости от условий среды, в которой развиваются корни, длина корневых волосков может значительно колебаться. В более сухих условиях корневых волосков образуется больше, чем во влажных; у растений, произрастающих в почве, покрытой водой, корневые волоски часто совершенно не образуются.
Оболочка корневых волосков целлюлозная, очень нежная, кутикула на жизнедеятельных волосках не образуется. Наиболее обычна цилиндрическая форма корневых волосков. Ветвистые корневые волоски формируются лишь у редких растений, но при слипании с частицами почвы строгость цилиндрической формы их нарушается. По существу, корневые волоски представляют собой выросты клеток эпидермиса, нередко сильно удлиненные. Однако у немногих растений корневые волоски все же отделяются перегородкой от породившей их эпидермальной клетки.
Начинают развиваться корневые волоски не на самом кончике корня, а на некотором расстоянии от него, примерно на 0,5-3,0 мм от верхушки. Зона, занимаемая корневыми волосками, по длине корня не очень велика, причем длина зоны зависит от многих условий, но не превышает нескольких миллиметров. Корневые волоски недолговечны: достигнув некоторого предела существования, они отмирают, сморщиваются и отпадают. Отмирание корневых волосков происходит в том конце занимаемой ими зоны, где расположены наиболее старые волоски, а на противоположном полюсе зоны происходит новообразование волосков. Корневые волоски постоянно возникают вновь в более молодых участках кончика корня и отмирают в более старых, структура которых становится более постоянной. Таким образом, разветвлениями корня охватываются все новые и новые частицы почвы, с которыми слипаются корневые волоски. Корневые волоски расположены на кончиках корней в занимаемой ими зоне очень густо. Например, у кукурузы на 1 мм 2 поверхности кончика корешка приходится около 425 корневых волосков, а у гороха при тех же самых условиях - 232. За счет образования корневых волосков поверхность корня сильно увеличивается. Так, поверхность покрытой корневыми волосками части кончика корня кукурузы в 5,5 раз больше поверхности такой же части корня без волосков, а у гороха - даже в 12,2 раза. Увеличение поверхности корня содействует выполнению одной из основных его функций - поглощению воды и растворенных в ней веществ.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter .
Покровные ткани являются пограничными. Они располагаются на поверхности органов растений, что обусловливает особенности их строения и многообразие выполняемых функций.
1. Покровные ткани обеспечивают связь растений с окружающей средой. При этом они являются не просто пассивным барьером, но и активными участниками взаимодействий. Например, клетки эпидермы участвуют в определении совместимости пыльцы и рыльца пестика; в развитии процессов, связанных с иммунитетом растений, реакцией на изменение температуры и влажности воздуха и почвы.
2. Покровные ткани осуществляют транспирацию и газообмен и их регулирование.
3. Покровные ткани участвуют в выделении продуктов метаболизма.
4. Покровные ткани защищают растения от поражения болезнетворными вирусами, бактериями, грибами; от поедания животными.
5. Покровные ткани предохраняют растения от суточных и сезонных перепадов температуры.
С учётом происхождения, местоположения, особенностей клеточного строения и выполняемых функций среди покровных тканей выделяют эпидерму, эпиблему, веламен, перидерму и корку. По мнению ряда учёных сюда же относится и семенная кожура.
> Первичные покровные ткани
Эпидермис. Эпиблема. Веламен
Эпидерма (эпидермис) является первичной покровной тканью, характерной для всех высших растений. У покрытосеменных она располагается на поверхности листьев, стеблей и их видоизменений, а также на репродуктивных органах - цветках и плодах. Кроме того, эпидермальные клетки могут выстилать внутреннюю поверхность эндокарпия - внутреннего слоя плодовой оболочки. Эпидерма характерна для травянистых растений. У деревьев и кустарников она развивается на молодых частях побегов, а по мере их старения замещается перидермой.
Эпидерма возникает из протодермы, порождаемой туникой конуса нарастания побега. Это сложная ткань, в состав которой входят основные и побочные клетки, замыкающие клетки устьиц, а также трихомы - выросты эпидермы. Клетки эпидермы располагаются однослойно. Однако у некоторых тропических растений (пальмы, орхидеи и др.) эпидерма бывает многослойной.
Основные клетки эпидермы стеблей, черешков листьев, а у однодольных и листовых пластинок, имеют удлиненную форму, вытянутую вдоль оси органа. Например, в стеблях злаков длина клеток эпидермы достигает 300 мкм. В листовых пластинках большинства двудольных, на цветках и плодах эпидермальные клетки имеют паренхимную и складчатую форму. Основные клетки эпидермы содержат развитые вакуоли, но лишены хлоропластов и не принимают участия в фотосинтезе. Их клеточные оболочки имеют ряд особенностей. Внутренняя оболочка тонкая, гладкая. Антиклинальная, т.е. поперечная, часто бывает извилистой, что увеличивает площадь соприкосновения смежных клеток эпидермы, а следовательно - силу их сцепления. Наружная оболочка наиболее прочная. Она образована более толстым слоем целлюлозы. Химический состав оболочек изменяется в ходе онтогенеза. У хвощей, осок и злаков оболочки часто пропитываются солями кальция, у овса - кремнеземом, у сахарного тростника - суберином. Реже, у бамбуковидных мятликовых, наблюдается одревеснение оболочек. В цитоплазме эпидермальных клеток могут образовываться кристаллы: рафиды (виноград, недотрога), друзы (свекла), цистолиты (фикус), реже - кристаллический песок (картофель).
Протопласт эпидермы активно вырабатывает и выделяет на наружную поверхность высокомолекулярные органические соединения, которые в совокупности образуют кутикулу. В составе катикулы первый слой образуют разветвленные молекулы пектина, затем располагаются глыбки растительного воска, кутин и эпикутикулярный воск. В зависимости от плотности сложения и скульптурированности поверхностного слоя, кутикула может быть плотной и блестящей, как у листьев фикуса, либо рыхлой и матовой, как у стеблей ржи, плодов сливы или яблони. Кутикула снижает интенсивность транспирации, отражает лучи солнца и защищает лист от перегрева, уменьшает возможность проникновения патогенов в растение.
Среди основных клеток эпидермы на верхней стороне листа мятликовых встречаются крупные моторные клетки с большими вакуолями. В жаркую пору они обеспечивают сворачивание листа в трубку (кукуруза) или складывание листовой пластинки (ковыль), что защищает растение от избыточного испарения влаги.
На основных клетках эпидермы часто образуются выросты, или трихомы, которые бывают по выполняемым функциям кроющими и железистыми, а по структуре - волосками и эмергенцами. Кроющие волоски заполнены воздухом и имеют белёсую окраску. Они хорошо отражают прямые солнечные лучи и уменьшают испарение воды. Железистые волоски - это живые клетки, вырабатывающие и выделяющие эфирные масла и другие конечные продукты метаболизма. Волоски образуются из клеток эпидермы. Они бывают одноклеточными и многоклеточными, простыми и ветвистыми. Одноклеточные простые волоски характерны для термопсиса и подмаренника, одноклеточные разветвленные - для пастушьей сумки и маттиолы. Многоклеточные простые встречаются у пеларгонии, многоклеточные ветвистые - у лаванды, а звездчатые - у лоха и облепихи.
Эмергенцы - это поверхностные структуры, образуемые клетками эпидермы и более глубоко расположенных тканей. К ним относятся шипы на плодах у сельдерейных, шипы на стеблях малины и ежевики, жгучие волоски листьев и стеблей крапивы. Они защищают растения от поедания.
Замыкающие клетки устьиц располагаются между основными клетками эпидермы. Они являются активными участниками транспирации и газообмена. Замыкающие клетки имеют небольшие размеры, содержат хлоропласты и попарно располагаются в эпидерме, не срастаясь между собой. Между ними находится устьичная щель, а под ними - подустьичная воздушная полость.
У однодольных замыкающие клетки имеют гантелевидную форму. Они сужены в средней части и расширены по краям. В расширенной части клеточные оболочки тонкие и растяжимые, а в середине - утолщенные и нерастяжимые. У двудольных замыкающие клетки имеют, при просмотре сверху, бобовидную форму. У них утолщены и нерастяжимы участки оболочки, обращенные к устьичной щели. Наружные оболочки этих клеток имеют выросты, которые возвышаются над устьичной щелью и образуют небольшую полость - передний дворик. Если такие же выросты возникают ниже устьичной щели, то они образуют задний, или внутренний, дворик, который непосредственно связан с подустьичной воздушной полостью. В совокупности замыкающие клетки, устьичная щель, передний и задний дворик, а также подустьичная воздушная полость образуют устьице, регулирующее транспирацию и газообмен. Число устьиц достаточно велико. У культурных растений на 1 мм 2 листовой поверхности размещается от 100 до 700 устьиц.
Работа устьиц сводится к регулированию ширины устьичной щели посредством сложных физиологических процессов. Устьица могут открываться в разное время суток. Это зависит от физиологического состояния растений, химического состава и концентрации клеточного сока, обеспеченности клеток водой и углекислым газом, освещенности листа, температуры воздуха и силы ветра.
Утреннее открывание устьиц можно схематично представить следующим образом. Под влиянием света в хлоропластах замыкающих клеток проходит фотосинтез. В результате концентрация их клеточного сока увеличивается, что приводит к поступлению воды из соседних, побочных, клеток и возрастанию тургорного давления. Под действием тургорного давления тонкие участки оболочек замыкающих клеток растягиваются в стороны, противоположные от устьичной щели, и толстые участки оболочки отходят друг от друга. Устьице открывается. При снижении тургорного давления в замыкающих клетках устьица закрываются. Подробно работа устьиц и процесс транспирации изучаются в курсе физиологии растений.
Побочные клетки располагаются рядом с замыкающими клетками устьиц и участвуют в регулировании их водного режима. По размерам они меньше основных и не имеют хлоропластов. Расположение побочных клеток является диагностическим признаком, используемым в систематике растений. Известно более десятка типов устьиц в зависимости от взаимного расположения замыкающих и побочных клеток. Например, при аномоцитном типе, характерном для винограда, красного перца и других цветковых растений, многочисленные побочные клетки не отличаются по размерам от основных клеток эпидермы. При перицитном, известном у папоротников - одна побочная клетка окружает устьице. У двудольных часто встречается парацитный тип устьица, когда две небольшие побочные клетки располагаются параллельно замыкающим. Для однодольных характерен тетрацитный тип, где четыре побочные клетки расположены вокруг устьица так, что плоскости их стенок не совпадают с плоскостью расположения устьичной щели.
Расположение устьиц на поверхности листа также имеет систематическое значение. Например, у однодольных они располагаются ровными рядами, а у двудольных - рассеяно по всей поверхности листовой пластинки. Как правило, устьица располагаются на нижней стороне листа. Это уменьшает испарение воды наземными растениями с плагиотропным, т.е. параллельным поверхности Земли расположением листьев (яблоня, свекла и др.). Если лист тяготеет к вертикальному, ортотропному, положению в пространстве, то устьица на нем могут располагаться с обоих сторон, как у кукурузы или лука. У водных растений, листья которых плавают на поверхности воды, устьица находятся на верхней стороне, как у типичной для белорусских водоёмов кубышки желтой.
Эпиблема. Особым видом первичной покровной ткани является эпиблема, которая образуется меристематическими клетками поверхности кончика корня. Клетки эпиблемы живые, тонкостенные, с развитой вакуолью, располагаются на поверхности корня в зоне всасывания. Одни из них, трихобласты, образуют особые одноклеточные выросты - корневые волоски, которые активно всасывают из почвы воду и растворенные в ней вещества. Другие, атрихобласты, корневых волосков не имеют. Жизнь корневых волосков ограничена двумя - тремя неделями. Затем они разрушаются и слущиваются. Одновременно, по мере роста кончика корня, на нем образуются новые корневые волоски.
Эпиблема на корнях однодольных сохранается на протяжении всей жизни растения. У двудольных в связи с переходом корня к вторичному анатомическому строению эпиблема сохраняется лишь в зоне корневых волосков, а в зоне проведения она замещается перидермой.
Веламен - это первичная покровная ткань воздушных корней тропических эпифитов и лазающих лиан.
Эпифиты произрастают в щелях ветвей, заполненных гумусом, и у оснований листьев крупных деревьев, используя их лишь как опору или субстрат, например, тениофиллум и фаленопсис из семейства Орхидные. Они поселяются также на скалах. Лазающие лианы, к которым относятся культурные виды - ваниль плосколистная (сем. Орхидные) и монстера деликатесная (сем.Ароидные), в качестве субстрата используют почву. У них образуются длинные стебли, прикрепляющиеся к опорным деревьям с помощью воздушных корней, что позволяет выносить листья и цветки в верхний ярус, к свету. У многих из них, как у монстеры, на стебле образуются питающие воздушные корни, растущие вниз. Достигнув почвы, они проникают в неё, ветвятся и всасывают воду и минеральные вещества.
Веламен образуется из протодермы и состоит из нескольких слоев мертвых клеток, оболочки которых имеют сетчатые или спиральные утолщения, придающие им прочность. Благодаря наличию в оболочках многочисленных пор и сквозных отверстий, веламен как губка капиллярным путём всасывает дождевую воду, а также конденсирующуюся атмосферную влагу. Таким образом, веламен выполняет защитную роль и регулирует водный режим растений.