МЕРИСТЕМИ ИЛИ ОБРАЗОВНИ ТКАНИНИ
Цитоплазма. Хемиски состав, физичка состојба, структура и својства. Влегување на супстанции во клетката. Видови на цитоплазматско движење.
Цитоплазмата е безбојна, има мукозна конзистентност и содржи различни супстанции, вклучувајќи високомолекуларни соединенија, на пример протеини, чие присуство ги одредува колоидните својства на цитоплазмата. Цитоплазмата е дел од протопластот, затворен помеѓу плазмалемата и јадрото. Основата на цитоплазмата е нејзината матрица или хијалоплазма, комплексен безбоен, оптички транспарентен колоиден систем способен за реверзибилни транзиции од сол во гел.
Во цитоплазмата растителни клеткиима органели: мали тела кои вршат посебни функции - пластиди, комплекс Голџи, ендоплазматичен ретикулум, митохондриите итн. Повеќето од процесите на клеточниот метаболизам се одвиваат во цитоплазмата, со исклучок на синтезата нуклеински киселини, кои се јавуваат во јадрото. Во цитоплазмата се пробиваат мембрани - најтенки (4-10 nm) филмови, изградени главно од фосфолипиди и липопротеини. Мембраните ја ограничуваат цитоплазмата од клеточната мембрана и вакуолата и внатре во цитоплазмата формираат ендоплазматичен ретикулум (ретикулум) - систем од мали вакуоли и тубули поврзани едни со други.
Најважното својство на цитоплазмата, поврзано првенствено со физичко-хемиските својства на хијалоплазмата, е неговата способност да се движи. Во клетките со една голема вакуола, движењето обично се врши во една насока (циклоза) поради специјални органели - микрофиламенти, кои се филаменти на посебен протеин - актин. Движечката хијалоплазма ги внесува пластидите и митохондриите. Клеточниот сок кој се наоѓа во вакуоли е воден раствор разни материи: протеини, јаглени хидрати, пигменти, органски киселини, соли, алкалоиди итн. Концентрацијата на супстанции кои се наоѓаат во клеточниот сок обично е поголема од концентрацијата на супстанции во надворешната средина (почва, водни тела). Разликата во концентрациите до одреден степен ја одредува можноста водните и почвените раствори да навлезат во клетката, што донекаде се објаснува со феноменот на осмоза. Улога во ќелијата полупропустлива мембранацитоплазмата игра. Граничните слоеви на цитоплазмата што ја обложуваат клеточната мембрана и клеточната вакуола се пропустливи само за вода и некои раствори, но непропустливи за многу супстанции растворени во вода. Ова својство на цитоплазмата се нарекува полупропустливост или селективна пропустливост. За разлика од цитоплазмата, клеточната мембрана е пропустлива за сите раствори, непробојна е само за цврсти честички. Влегувањето на супстанции во клетката не може да се сведе само на осмотски феномени, кои се изразени во возрасни клетки со добро развиени вакуоли. Всушност тоа е многу тежок процес, поради многу фактори. Целиот систем на цитоплазматски колоиди зема активно учество во апсорпцијата на супстанциите. Интензитетот на движење зависи од температурата, степенот на осветлување, снабдувањето со кислород итн.
Во многу млади клетки, цитоплазмата ја исполнува речиси целата празнина. Како што расте клетката, во цитоплазмата се појавуваат мали вакуоли исполнети со клеточен сок, кој е воден раствор од различни органска материја. Последователно, со понатамошен раст на клетките, вакуолите се зголемуваат во големина и, спојувајќи се, често формираат една голема централна вакуола, туркајќи ја цитоплазмата кон клеточната мембрана. Во таквите клетки, сите органели се наоѓаат во тенок ѕиден слој на цитоплазмата. Понекогаш јадрото останува во центарот на клетката. Во овој случај, цитоплазмата, формирајќи нуклеарен џеб околу неа, е поврзана со ѕидниот слој со тенки цитоплазматски нишки.
Слојот на цитоплазмата содржи хлоропласти што го обложуваат горниот ѕид. Тие се речиси тркалезни или малку овални тела. Повремено можете да видите пластиди повлечени во средината.
Концепт на ткаенини. Класификација на ткаенини. Разлика помеѓу трајните ткива и образовните ткива.
Повеќето копнени растенијаКлетките на телото не се исти по нивните функции, структура и потекло. Оваа хетерогеност се појави и беше консолидирана во процесот на прилагодување на растенијата на разликите во воздухот и почвата. Системите на клетки кои се структурно и функционално слични едни на други и обично имаат заедничко потекло се нарекуваат ткајачи. Скоро секој има ткаенини виши растенија. Само некои бриофити (црн дроб) ги немаат. Алги и црвена трева ( пониски растенија), по правило, исто така, немаат развиени ткива.
Комплекси на клетки кои се слични по функција и во поголемиот дела по структура, кои имаат исто потекло и специфична локализација во телото на растението, се нарекуваат ткива.
Распределбата на ткивата во органите на растенијата и нивната структура се тесно поврзани со извршувањето на одредени физиолошки функции.
Ткивата што се состојат од еден тип на клетки се нарекуваат едноставни, а оние што се состојат од различни типовиклетки - сложени, или сложени. Постои различни класификацииткаенини, но сите тие се сосема конвенционални. Растителните ткива се поделени во неколку групи во зависност од нивната главна функција:
1) меристеми или образовни ткива (ткива што се состојат од живи тенкоѕидни клетки кои интензивно се делат);
а) апикални (апикални) меристими (лоцирани на врвовите на стеблата и на краевите на корените) го одредуваат растот на овие органи во должина;
б) странични мерестими - камбиум и фелоген (камбиумот обезбедува задебелување на стеблото и коренот. Фелоген формира приклучок)
2) интегрален (заштити внатрешни ткаенинирастенијата од директно влијание на надворешната средина, го регулираат испарувањето и размената на гасови)
а) епидермисот;
б) плута;
3) спроводливи (обезбедете спроводливост на вода, почвени раствори и производи за асимилација произведени од листовите. Спроводните ткива можат да бидат примарни и секундарни по потекло.);
а) кселима или дрвена ткаенина(ткаенина спроводливи за вода)
б) флоем или флоем (ткиво што ги спроведува органските материи формирани од растението за време на фотосинтезата);
4) механички (одреди јачината на растението);
а) колонхим (се состои од паренхим или неколку издолжени клетки со нерамномерно задебелени целулозни ѕидови);
б) склеренхим (клетките имаат рамномерно задебелени лигнифицирани ѕидови);
1) влакна;
2) склериди;
5) основна (која се состои од хомогени паренхимски клетки кои го исполнуваат просторот помеѓу другите ткива);
6) секреторен или екскреторен (што содржи отпадни производи).
Само клетките на меристемските ткива се способни за поделба. Клетките на другите ткива, по правило, не се способни за поделба, а нивниот број се зголемува поради активноста на соодветните меристеми. Таквите ткива се нарекуваат трајни. Постојаните ткива произлегуваат од меристемите како резултат на клеточната диференцијација. Диференцијацијата лежи во тоа што за време на индивидуален развојорганизам (онтогенеза), квалитативните разлики се јавуваат помеѓу првично хомогени клетки, додека структурата и функционални својстваклетките. Обично диференцијацијата е неповратна. На нејзиниот напредок влијаат супстанции кои делуваат како хормони.
МЕРИСТЕМИ ИЛИ ОБРАЗОВНИ ТКАНИНИ
Меристемите (од грчкиот „meristos“ - делливи), или образовните ткива, имаат способност да се делат и да формираат нови клетки. Поради меристемите се формираат сите други ткива и настанува долгорочен (доживотен) раст на растението. На животните им недостасуваат меристеми, што го објаснува нивниот ограничен период на раст. Меристемските клетки се карактеризираат со висока метаболичка активност. Некои меристемски клетки, наречени почетни, остануваат во ембрионската фаза на развој во текот на животот на растението, додека други постепено се разликуваат и се претвораат во клетки од различни трајни ткива. Почетната клетка на меристемот, во принцип, може да доведе до која било клетка од телото. Телото на копнените растенија е дериват на релативно малку почетни клетки.
Примарните меристеми имаат меристемска активност, односно првично се способни за делење. Во некои случаи, способноста за активно делење може повторно да се појави во клетките кои речиси го изгубиле ова својство. Ваквите „новопојавени“ меристеми се нарекуваат секундарни.
Во телото на растението, меристемите заземаат различни позиции, што им овозможува да се класифицираат. Според нивната положба во растението, се разликуваат апикални или апикални (од латинскиот „apex“ - врв), странични или странични (од латинскиот „latus“ - страна) и интеркаларни меристеми.
Апикалните меристеми се наоѓаат на врвовите на аксијалните органи на растението и обезбедуваат раст на телото во должина, додека страничните меристеми првенствено обезбедуваат раст во дебелина. Секој изданок и корен, како и ембрионскиот корен и пупка на ембрионот, имаат апикален меристем. Апикалните меристеми се примарни и формираат конуси од растот на коренот и ластарот (сл. 1).
Страничните меристеми се наоѓаат паралелно со страничните површини на аксијалните органи, формирајќи еден вид цилиндри кои изгледаат како прстени во пресеци. Некои од нив се примарни. Примарни меристеми се прокамбиум и перицикл, секундарни се камбиум и фелоген.
Меѓукаларните, или интеркаларни, меристемите се често примарни и се зачувани во форма на посебни делови во зоните на активен раст (на пример, во основата на меѓујазлите, во основата на ливчињата на листовите).
Постојат и рани меристеми. Тие се формираат на места на оштетување на ткивата и органите и даваат калус - специјално ткиво кое се состои од хомогени паренхимски клетки кои го покриваат местото на оштетување.Способноста за формирање на калус на растенијата се користи во хортикултурната практика при нивното размножување со сечи и калемење . Колку е поинтензивно формирањето на калусот, толку е поголема гаранцијата за спојување на подлогата со потомството и искоренувањето на сечињата. Формирање на калус - неопходен условкултура на растителни ткива на вештачки подлоги.
Клетките на апикалните меристеми се повеќе или помалку изодијаметриски по големина и полиедарски во форма. Меѓу нив нема меѓуклеточни простори, лушпите се тенки и содржат малку целулоза. Клеточната празнина е исполнета со густа цитоплазма со релативно големо јадро кое зафаќа централна позиција. Вакуолите се многубројни, мали, но обично не се забележуваат под светлосен микроскоп. Ергастичните супстанции обично се отсутни. Пластидите и митохондриите се малку и мали.
Клетките на страничните меристеми се разликуваат по големина и форма. Тие грубо одговараат на клетките на оние трајни ткива кои подоцна произлегуваат од нив. Така, и паренхимните и прозенхималните иницијали се наоѓаат во камбиумот. Паренхимот на спроводливите ткива се формира од паренхимните иницијали, а спроводните елементи се формираат од прозенхималните иницијали.
Ориз. 1. Апикален меристем на изданокот Елодеа. А - надолжен пресек; 5 - конус за раст ( изгледи надолжен пресек); Б - клетки на примарниот меристем;
G - паренхимна клетка на формираниот лист:
1 ~ конус за раст, 2 - лист примордиум, 3 - туберкула на аксиларни пупки
Корени зони. Карактеристики на морфолошката структура на секоја зона во врска со извршените функции. Раст на коренот во должина и дебелина.
Заедно со ластарот, коренот е главниот орган на вишото растение, кој обично ја извршува функцијата на минерална и водена исхрана. Друго важна функцијакорен, тесно поврзан со главниот, - закотвување на растението во почвата.
Различни делови од коренот вршат различни функции и се карактеризираат со одредени морфолошки карактеристики. Овие делови се нарекуваат зони (сл. 2). Врвот на коренот е секогаш покриен однадвор со капа на коренот што го штити апикалниот меристем. Клетките на капачето на коренот произведуваат слуз што ја обложува површината на коренот. Благодарение на слузта, се намалува триењето со почвата, а нејзините честички лесно се лепат до врвовите на коренот и коренските влакна.
Коренската капа се состои од живи паренхимски клетки, кои произлегуваат кај повеќето еднокомници од посебен меристем наречен калиптроген, а кај двокотиледоните и гимноспермите - од апикалниот меристем на врвот на коренот. Водните растенија обично немаат коренска капа.
Под капачето има зона на поделба, претставена со меристемскиот врв на коренот, неговиот врв. Како резултат на активноста на апикалниот меристем, се формираат сите други зони и ткива на коренот.
Клетките кои се делат се концентрирани во зоната на делење, која е околу 1 мм. Овој дел од коренот е значително различен од другите зони по својата жолтеникава боја. По зоната на поделба е зона на проширување (раст). Мал е и по должина (неколку милиметри), се истакнува со светла боја и навидум е проѕирен. Клетките на зоната на раст практично не се делат, но се способни да се протегаат во надолжната насока, туркајќи го коренскиот крај длабоко во почвата. Тие се карактеризираат со висок тургор, што придонесува за активно ширење на честичките на почвата. Во зоната на раст, се јавува диференцијација на примарните спроводливи ткива.
Крајот на зоната на раст е забележлив со појавата на бројни коренски влакна на епиблемата. Влакната од коренот се наоѓаат во зоната на вшмукување, чија функција е јасна од неговото име. На коренот зафаќа површина од неколку милиметри до неколку сантиметри. За разлика од зоната на раст, деловите од оваа зона повеќе не се поместуваат во однос на честичките на почвата. Младите корени го апсорбираат најголемиот дел од вода и солени раствори во зоната на апсорпција со помош на влакна од коренот.
Влакната од коренот се појавуваат во форма на мали папили - израстоци на клетките на епиблема. Растот на косата се јавува на нејзиниот врв. Школка корен косабрзо се протега. По одредено време, коренот на косата изумира. Неговиот животен век не надминува 10-20 дена.
(меристемски клетки), а се наоѓаат во деловите на растенијата каде што се јавува раст.
1. Цитолошки карактеристики на меристемите
Диференцираните растителни клетки воопшто не можат да се поделат или да се трансформираат во други видови клетки. Затоа, клеточната делба во меристемите е неопходна за да се обезбедат нови клетки за раст на други ткива, формирање на нови органи и структура на телото на растението. Функцијата на меристемските клетки е слична на животинските матични клетки, кои не се разликуваат или малку се разликуваат и се способни за континуирана клеточна делба. Меристемските клетки се мали, а цитоплазмата и јадрото целосно ја исполнуваат клетката. Вакуолите се исклучително мали и цитоплазмата не содржи диференцирани пластиди (хлоропласти или хромопласти), иако тие се присутни во остаток форма (протопластиди). Меристемските клетки се цврсто спакувани, без речиси никаков меѓуклеточен простор. Клеточен ѕид - многу тенок примарен клеточен ѕид.
Физиолошката активност на меристемите бара рамнотежа помеѓу два антагонистички процеси: формирање на нови ткива и поддршка за обновување на популацијата на меристемски клетки.
Меристем- формирање растително ткиво од кое може да се формираат сите други ткива, меристемски клетки долго времеја задржуваат способноста да се делат со формирање на нови неспецијализирани клетки.
Клетките на меристемот се срамнети со земја, мали, густо лоцирани една до друга, без интерклиниформи, јадрото се наоѓа во центарот на клетките, вакуолите се мали, многубројни, клеточната мембрана е примарна, има само пластидни прекурсори.
Цитолошките карактеристики на круп меристемите се типично изразени во апикалните меристеми. Клетките се изодијаметриски полигони, кои не се одделени со меѓуклеточни простори. Клеточните ѕидови се тенки, со мала содржина на целулоза. Цитоплазмата е густа, јадрото е големо, сместено во центарот. Во цитоплазмата има голем број на рибозоми и митохондрии (настанува интензивна синтеза на протеини и други материи). Бројни вакуоли се многу мали.
Клетките на страничните меристеми се нееднакви по големина и форма. Ова се должи на разликата во клетките на трајните ткива од кои се формираат. На пример, во камбиумот има паренхимални и прозенхимални клетки. Паренхимот на водечките комплекси се формира од паренхимните иницијали, а самите спроводливи елементи се формираат од прозенхимните иницијали.
Меристемите се класифицираат според нивната положба во растението:
И по потекло:
- основно
- секундарно.
Примарни меристеми се во тесна врска со апикалниот меристем и директно од нив се формираат, особено, протодермот, главниот меристем и прокамбиумот. Протодермот е еднослоен слој од слабо диференцирани клетки што го покрива врвот однадвор; епидермисот последователно се формира од него. Кората се формира од главниот меристем, а централниот цилиндар е формиран од прокамбиумот, чии клетки се издолжени по оската на стеблото.
Секундарни меристеми продираат од специјализирани клетки на зрели растителни органи и обезбедуваат нивен страничен раст (фелоген, перицикл, камбиум). Ткивата формирани од примарниот меристем се нарекуваат примарни, а оние што се формираат од секундарниот меристем се нарекуваат секундарни. Секундарните меристеми треба да ги вклучуваат и таканаречените трауматски меристеми, кои се формираат на места каде што телото на растението е оштетено и обезбедуваат регенерација.
Помеѓу наведените типови на меристеми, првиот што се појавува во онтогенезата е апикалниот меристем, од кој настанува диференцијација на сите други ткива. Се наоѓа на врвовите (APEX) на стеблото и коренот и нивните странични гранки. Карактеристики на животот растителен организаме, особено, зачувување на формативната активност во апикалните меристеми во текот на целата онтогенеза, затоа велат дека растенијата се способни за неограничен раст.
Структура на апикалниот меристем (точка на раст)
1 - централен дел
2 - Периферен дел
2. Апикален меристем
4. Интеркаларен меристем
Интеркаларен меристем (синоним - интеркаларен меристем) - клетките се наоѓаат помеѓу областите на диференцирани ткива, обезбедувајќи интеркаларен раст.
Интеркаларниот меристем, на пример, се наоѓа во јазлите на житарките, во основата на ливчињата и филаментите на стаменот. Овие се резидуални примарни меристеми. Тие произлегуваат од меристемите на јажето, но нивната трансформација во трајни ткива е одложена во споредба со другите матични ткива. Овие нежни меристеми се особено забележливи кај житариците. Кај лебот тие обезбедуваат подигање на стеблата поради нерамномерната поделба на клетките од долната и горната страна на сламата.
5. Рана меристем
Се формира кога ткивата и органите се повредени. Живите клетки кои ги опкружуваат погодените области се дедиференцираат и почнуваат да се делат, односно се претвораат во секундарен меристем. Се формираат меристеми на рана Калус- густа ткаенина од виолетова или жолтеникава боја, кој се состои од паренхимски клетки различни големини, се наоѓа вон ред. Клетките на калусот имаат големи јадра и релативно дебели клеточни ѕидови. Калусот може да доведе до секое растително ткиво или орган. На периферијата се формира приклучок, можна е диференцијација на клетките на калусот во други ткива. Во Калус тие можат да лежат дополнителни корении бубрезите. Калусот се јавува кога растенијата се накалемени и го обезбедува растот на потомството и подлогата; во основата на сечињата. Калусот се користи и за добивање на изолирани ткивни култури.
Растителните органи се составени од ткива. Растително ткивое збир на клетки кои имаат слична структура, хомогено потекло и вршат иста функција.Ткивата на повисоките копнени растенија достигнаа посебен развој. Кај растенијата се разликуваат образовните ткива: интегрални, механички, проводни, основни и екскреторни.
Едукативни ткаенини, или меристемите, служат во растението за раст; тие се од примарно потекло и од секундарни. Примарното образовно ткиво се наоѓа во конусите за раст на корените и стеблата. Ајде да погледнеме прво конус за раст на коренот(Сл. 1, I).
Сл.1. Примарно образовно ткиво во конусот за раст на коренот:
јас - конус за раст на коренот: 1 - клетки на едукативно ткиво; 2 - клетки на капачето на коренот. II - последователни фази на сложена нуклеарна и клеточна делба(кариокинеза, или митоза): 1 - интерфаза; 2 - профаза; 3 - метафаза; 4-5 - анафаза; 6-7 - телофаза; 8 - цитокинеза.
Врвот на коренот е во форма на конус, покриен со корен капа, под кој се наоѓа примарното воспитно ткиво. Клетките на образовното ткиво се тетраедрални или повеќеслојни, имаат густа протоплазма, големи јадра и тенки целулозни мембрани. Нивните вакуоли не се развиени или слабо развиени. Овие клетки имаат способност енергично да се делат. На клеточната делба и претходи сложена поделба на јадрото, во која се случуваат чудни промени. На готови, добро подготвени и обоени препарати видливи се јадра кои се во различни фази комплексна поделба, инаку наречена кариокинеза („карион“ - јадро, „кинези“ - движење) (сл. 1, II). Бројни набљудувања утврдиле дека во јадрото, тенки хроматински филаменти најпрво произлегуваат од хроматинската супстанца, кои се скратуваат, згуснуваат и се претвораат во поединечни честички - хромозоми („хромо“ - боја, „сома“ - тело), често закривени во форма на потковица (профаза на поделба) . Се чини дека секој хромозом во профазата е поделен по должина на две половини, но во реалноста се случува удвојување на хромозомите (редупликација). Јадрото и нуклеарната обвивка постепено исчезнуваат. Хромозомите се наоѓаат во средишниот дел на јадрото во една рамнина во форма на плоча. Половините од секој хромозом (хромозоми „ќерка“) цврсто се вклопуваат една до друга, и во оваа фаза (метафаза) на попречните делови од коренот, можете лесно да го броите бројот на хромозоми и да ја испитате нивната форма. На половите на јадрото (дури и пред исчезнувањето на нуклеарната мембрана), се појавува систем на тенки ахроматски (не боење) филаменти, распоредени во форма на две капачиња. Конците растат и добиваат форма на вретено. По ова, дивергенцијата на половините од хромозомите (ќерките хромозоми) започнува кон двата спротивни пола на клетката (анафаза). Дивергенцијата на хромозомите завршува со ќерките хромозоми, собрани на половите на клетката, испреплетени едни со други, претворајќи се во две топки (телофаза). Во исто време, на ахроматинските нишки во средишниот дел на клетката се појавуваат задебелувања на пектин - се формира фрагмопласт, односно рудимент на попречната мембрана. Конечно, јадрата се опкружени со мембрана, во нив се формираат јадра (едно или повеќе), а протоплазмата е поделена со попречна мембрана (цитокинеза), што резултира со појава на две нови клетки.
Процесот на нуклеарна и клеточна делба се јавува почесто ноќе, неговата брзина зависи од температурата и влажноста. Процесот на поделба трае во просек 75-120 минути, но понекогаш трае и неколку часа, при што во првата фаза (профаза) се троши најмногу време. За време на клеточната делба, клеточната мембрана се појавува како производ на виталната активност на протоплазмата и јадрото. Прво, се појавува средната плоча. Се состои од меѓуклеточна супстанција која ги лепи примарните целулозни мембрани кои припаѓаат на две клетки.
За прв пат, дивергенцијата на хромозомите кон половите на клетката, како и други детали за поделбата на јадрото и клетката, беше утврдена во 1875 година од професорот на Московскиот универзитет И. Д. Чистјаков.
Суштината на сложената поделба на јадрото и клетката, исто така наречена митоза, очигледно лежи во фактот дека со подеднакво делење на хромозомите на две половини (или, поточно, удвојување на хромозомите), ќерките клетки добиваат од мајката повеќе или помалку еднакви количини хроматинска супстанца (деоксирибонуклеинска киселина).киселини – ДНК), која ја пренесува наследноста од матичната клетка на ќерките ќерки. („Митос“ е нишка. Вообичаено е прифатено дека митоза = карјокинеза + цитокинеза). Обликот и бројот на хромозомите се специфични знациза секој растителен вид (мека пченица 2n = 42 хромозоми, тврда пченица 2n = 28, јачмен 2n = 14 итн.). Сепак, понекогаш постои варијабилност во бројот на хромозоми, а клетките со различни хромозоми може да се забележат во еден организам.
Покрај митозата (или кариокинезата), кај растенијата се јавува директна поделба на клеточното јадро (амитоза) - едноставно повлекување на јадрото, а понекогаш и на целата клетка, на две половини.
Заедно со митозата, редукцијата на нуклеарната поделба е честа кај растенијата - мејозата, во која бројот на хромозоми во новите клетки е намален за половина во споредба со матичната клетка. Оваа поделба се јавува кај растенијата за време на спорулација, по што се појавуваат и се развиваат машки и женски полови клетки (гамети). Последните имаат половина од бројот на хромозоми во нивните јадра, а таквите јадра се нарекуваат хаплоидни за разлика од диплоидните јадра на вегетативните клетки кои ги сочинуваат вегетативните органи на растенијата. Сексуалниот процес на растенијата се состои од спојување на две гамети (машки и женски). Ако немаше намалување на хроматинската супстанција, односно преполовување на нејзината маса, тогаш во секоја нова сексуална генерација оваа маса на супстанција, а со тоа и бројот на хромозоми, неизбежно ќе се зголеми. Во половите клетки (гамети), јадрата се хаплоидни, и тоа само по процесот на оплодување клеточни јадраповторно стануваат диплоидни.
Редукционата поделба беше откриена од научникот В.И.Белјаев, а научникот С.Г.Навашин откри двојно оплодување кај цветните растенија и е основач целото училиштенаучниците вклучени во проучувањето на сите детали за нуклеарната и клеточната делба.
Примарното образовно ткиво предизвикува други ткива, бидејќи неговите клетки, менувајќи се, се разликуваат. Способноста на клетките брзо да се делат во стеблата и корените на двокотиледоните растенија е зачувана доживотно во меристемски клетки наречени камбиум, поради што стеблото и коренот се згуснуваат. Средните образовни ткива ги вклучуваат оние што произлегуваат од други ткива. Пример е фелогенот, кој често произлегува од земното ткиво.
1) Образовни ткива, или меристеми
Како резултат на активноста се формира растителното тело едукативниткива наречени меристемите. Главното својство на меристемите е способноста на клетките да се делат со митоза, поради присуството на почетни клетки.
Составот на меристемите вклучува: 1) иницијали, 2) деривати на иницијали. Иницијалите ја задржуваат способноста да се делат во текот на животот на растението и секогаш остануваат дел од меристемот. Дериватите од иницијалите се делат одреден број пати, а потоа се трансформираат во трајни ткива.
Карактеристики почетна: густа цитоплазма, големо јадро, многу митохондрии, мали или никакви вакуоли, тенок клеточен ѕид.
По потекло се разликуваат: 1) основно меристемите, кои потекнуваат директно од меристемите на ембрионот. Од нив се развиваат примарни трајни ткива. 2) секундарно меристемите, од примарни или други ткива. Од нив се развиваат трајни секундарни ткива.
Во зависност од локацијата, постојат четири типа меристеми:
Апикална ( апикални) меристеми- на врвовите на стеблата и краевите на корените, обезбедува раст во должина.
Странични (странични) меристеми -долж обемот на аксијалните органи, се обезбедува раст во дебелина. Примарни странични меристеми се прокамбиумот и перициклот. . Секундарни странични меристеми: камбиум– произлегува од прокамбиум - и плута камбиум (фелоген)- формиран од клетки на трајни ткива. Проводните ткива се формираат од прокамбиум и камбиум, а перидермот се формира од плута камбиум.
Интеркаларни меристемисе наоѓаат во основите на меѓујазлите на ластарите и младите лисја. Тие се примарни бидејќи се остатоци од апикални меристеми. Меѓукаларните меристеми немаат иницијали во својот состав и со текот на времето целосно се претвораат во трајни ткива.
Рана (трауматски) меристемиобично се формираат кога ткивата и органите се оштетени. Се формираат меристеми на рана калус- специјално ткиво кое се состои од хомогени паренхимски клетки кои го покриваат местото на повредата. Секое ткиво или орган на растение може да произлезе од калус.
Практична употреба: цвеќињата се користат во парфимерија, медицина и декорација.
2) Микроскопска структура на едноколини:
А) Изразен примарен кортекс(крин, ирис). Во овие случаи, во стеблото може да се види јасна диференцијација на кожа, примарен кортекс и аксијален цилиндарсо повеќеслоен перицикл во вид на склеренхимски прстен. Примарниот кортекс обично се состои од неколку слоеви паренхимални клетки кои носат хлорофил. Коленхим обично е отсутен. Во подземните пука, примарниот кортекс и централниот цилиндар се особено остро разграничени; примарниот кортекс е многу поширок отколку во приземните стебла на истите растенија; во ендодермот се појавуваат каспаријански прстени или дури и задебелувања во облик на чаша клеточните мембрани, а перициклот е претставен главно со паренхимски клетки.
Б) Кај другите претставници од класата на монокрите, во семејствата на палми, треви и други, примарната кора е толку јасна не се изразува. Во стебленцето на пченката, под епидермисот, понекогаш има области на паренхим што носи хлорофил; склеренхимот ги опкружува овие области и меѓу нив е во непосредна близина на епидермисот. Периферните васкуларни снопови имаат моќни склеренхимски обвивки, а во близина на епидермисот понекогаш се формира континуиран слој на склеренхим, во кој се потопуваат васкуларните снопови.
3) општи карактеристикивиши растенија
По изглед, структура и биолошки карактеристикивишите растенија се многу разновидни. Тие вклучуваат не само цветни растенија, но и гимносперми, како и папрати, коњско опавче, мов и мов.
Вишите растенија се карактеризираат со голем број карактеристики повеќе висока организација. Нивното тело е поделено на стебла и лисја (разлика од долните).Во овој поглед, тие се нарекуваат лиснати растенија.
Појавата на сложена поделба, формирање на органи (корен, стебло, лист) беше предизвикана од преминот на растенијата во нов копнена срединаживеалиште. Воздушна срединасе карактеризира со многу поголема содржина на кислород од вода. Почва - други услови за минерална исхрана и особено снабдување со вода. За асимилација во нови услови, беше потребно реструктуирање на фотосинтетичкиот апарат. Така растението развило стебло.
Следно, за подобра апсорпција, ризоидите беа заменети со корени. Корените почнаа да обезбедуваат минерална исхрана, а листовите обезбедуваа воздушна исхрана за повисоките растенија.
За нормално функционирање на органите, потребно беше брзо да се премести водата до лисјата и органските материи од листот до другите делови на растението. Ова доведе до развој на спроводен систем во форма на специјални ќелии - трахеиди, садови и цевки со сито. За заштита, формирано е покривно ткиво, механичко.
Сексуалниот процес стана помалку зависен од вода.
Виши растенија: ринофити, псилофити, мовови, ликофити, коњски опавчиња, папрати, гимносперми, ангиосперми
ТКАНИНИ. КЛАСИФИКАЦИЈА НА ТКАФИНИ.
Организацијата на вишите растенија се заснова на принципот на специјализација на клетките, кој лежи во фактот дека секоја клетка на телото не ги извршува сите свои својствени функции, туку само некои, но поцелосно и посовршено.
Ткаенини- стабилни, природно повторувачки комплекси на клетки, слични по потекло, структура и прилагодени за извршување на една или повеќе функции.
Постојат различни класификации на ткаенини, но сите тие се сосема произволни.
Во зависност од главната функција, се разликуваат неколку групи растителни ткива.
1. едукативни ткаенини,или меристеми,- имаат способност да ги делат и формираат сите други ткива.
2. Ткива за покривање:
Примарен;
Секундарна;
терцијарно.
3. Главни ткаенини- Шминка повеќеторастително тело. Се разликуваат следниве главни ткаенини:
Асимилација (носи со хлорофил);
Чорапи;
Воздушно (аеренхим);
Водоносни слоеви.
4. Механички ткаенини(потпорна, скелетна):
Коленхим;
Склеренхим.
5. Проводни ткаенини:
Ксилемот (дрвото) е растечко ткиво;
Флоем (флоем) е ткиво на опаѓачки тек.
6. Екскреторни ткива:
Надворешен:
жлезди влакна;
Хидатодите се водни стомати;
Сончевите птици;
Внатрешна:
Екскреторни клетки со есенцијални масла, смоли, танини;
Повеќеклеточни садови за секрет, лактификатори.
Врз основа на способноста на клетките да се делат, се разликуваат два вида ткива: едукативни,или меристеми,И постојана- интегрален, екскреторен, основен, механички, проводен.
Ткаенината се нарекува едноставно,ако сите негови клетки се идентични по форма и функција (паренхим, склеренхим, колонхим). Комплексенткивата се состојат од клетки кои не се идентични по форма, структура и функција, но се поврзани заедничко потекло(на пр. ксилем, флоем).
Исто така, постои класификација на ткивата врз основа на нивното потекло (онтогенетски). Според оваа класификација, се разликуваат примарни и секундарни ткива. Од примарниот меристем лоциран на врвот на ластарот и врвот на коренот, како и од семенскиот ембрион, примарни константиткива (епидермис, колонхим, склеренхим, асимилирачко ткиво, епиблема). Клетките на трајните ткива не се способни за понатамошна поделба. Од клетки на специјализиран меристем - прокамбија - се формираат примарни проводнициткива (примарен ксилем, примарен флоем).
Од секундарниот меристем - камбиум - се формираат секундарноткива: секундарен ксилем, секундарен флоем; од фелоген Се формираат приклучок, фелодерм и леќа, кои настануваат кога стеблото и коренот се згуснуваат. Секундарните ткива обично се наоѓаат во гимносперми и дикотиледони ангиосперми. Моќниот развој на секундарните ткива - дрво и кори - е карактеристичен за дрвенестите растенија.
ЕДУКАТИВНИ ТКАНИНИ
Едукативни ткаениниБлагодарение на постојаната митотична поделба на нивните клетки, тие обезбедуваат формирање на сите растителни ткива, т.е. всушност го обликува неговото тело. Секоја клетка во својот развој поминува низ три фази: ембрионална, фаза на раст и диференцијација (односно, клетката добива одредена функција). Како што ембрионот се разликува, примарниот меристем е зачуван само на врвот на идното пука (во конусот на раст) и на врвот на коренот - апикален (апикален) меристемите.Ембрионот на кое било растение се состои од меристемски клетки.
Цитолошки карактеристики на меристемите.Типичните карактеристики се најјасно изразени во апикалните меристеми. Овие меристеми се составени од изодијаметриски полиедарски клетки кои не се одделени со меѓуклеточни простори. Нивните лушпи се тенки, содржат малку целулоза и се растегливи.
Шуплината на секоја клетка е исполнета со густа цитоплазма со релативно големо јадро кое зазема централна позиција и интензивно се дели со митоза. Хијалоплазмата содржи многу дифузно расфрлани рибозоми, пропластиди, митохондрии и диктјозоми. Има малку вакуоли и тие се мали. Спроводните ткива се формираат од меристем кој има прозенхимална форма и големи вакуоли - прокамбиум и камбиум. Прокамбиумските клетки во пресекполигонални, камбиумските ќелии се правоаголни.
Клетките кои ги задржуваат своите меристемски својства продолжуваат да се делат, формирајќи се повеќе и повеќе нови клетки наречени иницијали.Некои од ќерките клетки се разликуваат, претворајќи се во клетки од различни ткива, тие се нарекуваат деривати на иницијали.Почетните клетки можат да се делат неодредено многу пати, а дериватите на иницијалите се делат еден или повеќе пати и се развиваат во трајни ткива.
Врз основа на нивното потекло, се разликуваат примарни и секундарни меристеми.
Примарни меристеми
Примарните меристеми произлегуваат директно од меристемот на ембрионот и имаат способност да се делат. Според нивната положба во растението, примарните меристеми можат да бидат апикални (апикални), интеркаларни (меѓукаларни) и странични (странични).
Апикални (апикални) меристеми- такви меристеми кои се наоѓаат во возрасни растенија на врвовите на стеблата и врвовите на корените и обезбедуваат раст на телото во должина. Во конусот на раст на стеблата се разликуваат два меристемски слоја: туниката, од која интегралното ткиво и периферен делпримарниот кортекс и телото од кое се формираат внатрешниот дел на примарната кора и централниот аксијален цилиндар (сл. 2.3).
Ориз. 2.3.Апикални меристеми на стеблото: А- надолжен пресек: 1 – конус на раст; 2 – лист примордиум; 3 – туберкула на аксиларни пупки;
Постојат три слоја на врвот на коренот:
1) дерматоген, од кој се формира примарното ткиво што апсорбира интегрално ткиво - ризодерм;
2) периблма, од која се развиваат ткивата на примарниот кортекс;
3) плером, формирајќи ги ткивата на централниот аксијален цилиндар.
по потекло тие можат да бидат примарни и секундарни, на пресек на аксијалните органи изгледаат како прстени. Пример за примарен страничен меристем е прокамбиумот и перициклот. Од прокамбијаСе формираат камбиумот и примарните елементи на васкуларно-влакнестите снопови (примарен флоем и примарен ксилем), додека прокамбиумските клетки директно се диференцираат во клетките на примарните спроводливи ткива.
Латералните меристеми се наоѓаат паралелно со површината на органот и обезбедуваат раст на аксијалните органи во дебелина.
Интеркаларни меристемипочесто примарни и зачувани во вид на посебни области во зони на активен раст во разни деловирастенија (на пример, во основата на листните ливчиња, во основата на меѓујазлите). Во основата на меѓујазлите кај житарките, активноста на овој меристем доведува до издолжување на меѓујазлите, со што се обезбедува раст на стеблото во должина.
Секундарни меристеми
Секундарните меристеми вклучуваат странични и ранети меристеми.
Странични (странични) меристемипрезентирани камбиумИ фелоген.Тие се формираат од промеристеми (прокамбиум) или трајни ткива со нивната дедиференцијација. Камбиумските клетки се поделени со прегради паралелни со површината на органот (периклинално). Елементите на секундарниот флоем се развиваат од клетките депонирани нанадвор од камбиумот, а елементите на секундарната ксилема се развиваат од клетките депонирани навнатре. Камбиумот, кој настанал од трајните ткива преку дедиференцијација, се нарекува дополнителниВо структурата и функцијата не се разликува од камбиумот, кој настанал од промеристемите. Фелогенот се формира од трајни ткива лоцирани во субепидермалните слоеви (под епидермисот). Поделувајќи се периклински, фелогенот ги одвојува идните приклучни клетки (фелема) нанадвор, а фелодермските клетки навнатре. Така, фелоген формира секундарна покривно ткиво– перидерм. Латералните меристеми се наоѓаат паралелно со површината на органот и обезбедуваат раст на аксијалните органи во дебелина.
Меристеми на ранисе формираат кога ткивата и органите се оштетени. Околу оштетувањето, живите клетки се дедиференцираат, почнуваат да се делат и со тоа се трансформираат во секундарен меристем. Нивната задача е да формираат густо заштитно ткиво составено од паренхимски клетки - калус.Ова ткиво има белузлава или жолтеникава боја, неговите клетки имаат големи јадра и прилично дебели клеточни ѕидови. Калусот се јавува за време на калемењето, обезбедувајќи спојување на потомството со подлогата и во основата на сечињата. Може да формира адвентивни корени и пупки, па затоа се користи за добивање на изолирани ткивни култури.
ПОКРИВАЧНО ТКИВО
Примарно интегрално ткиво
ДО примарен интеграленткивата го вклучуваат епидермисот, самиот епидермал, парастоматалните клетки, чуварите на стоматите и трихомите.
Пектичките супстанции и целулозата вклучени во клеточниот ѕид може да бидат предмет на формирање на слуз со формирањето лигитеИ непцата.Тие се полимерни јаглехидрати поврзани со пектинските материи и се карактеризираат со нивната способност силно да отекуваат кога се во контакт со вода. Непцата во отечена состојба се лепливи и може да се извлечат во конци, додека слузта е многу матна и не може да се извлече во нишки. Пектичките муцилажи се наоѓаат кај претставниците на семејствата Liliaceae, Cruciferae, Malvaceae, Linden и Rosaceae, за разлика од целулозните муцилажи, кои се многу поретки (на пример, кај орхидеи).
 Стомати
Тие се високо специјализирани формации на епидермисот, кои се состојат од две заштитни клетки во облик на грав и стоматална пукнатина (еден вид меѓуклеточен простор меѓу нив). Ги има главно во листовите, но ги има и во стеблото (сл. 2.6).
Стомати
Тие се високо специјализирани формации на епидермисот, кои се состојат од две заштитни клетки во облик на грав и стоматална пукнатина (еден вид меѓуклеточен простор меѓу нив). Ги има главно во листовите, но ги има и во стеблото (сл. 2.6).
Ориз. 2.6.Стоматална структура: а, б- кора од лист од мајчина душица (горен поглед и пресек); В- кора од стеблото на Цереус (фамилија кактуси); 1 – вистински епидермални клетки; 2 – чуварски клетки на стомите; 3 – стомачна пукнатина; 4 – воздушна шуплина; 5 – клетки на паренхим што носи хлорофил; А – кутикула; Б – кутикуларен слој – школка со суберин и восок; Б – целулозниот слој на ѕидот; G – јадро со јадро; Д – хлоропласти
Ѕидовите на заштитните ќелии се нерамномерно задебелени: ѕидовите насочени кон празнината (абдоминална) се значително подебели во споредба со ѕидовите насочени подалеку од јазот (дорзална). Јазот може да се прошири и собира, регулирајќи ја транспирацијата и размената на гасови. Под јазот има голема респираторна празнина (меѓуклеточен простор), опкружена со мезофилни клетки на листот.
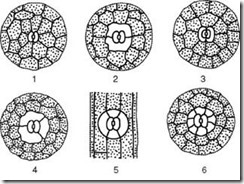 Чуварите клетки се опкружени со парастоматални клетки, кои заедно се формираат стомален комплекс(Сл. 2.7). Се разликуваат следниве главни типови на стоматични комплекси:
Чуварите клетки се опкружени со парастоматални клетки, кои заедно се формираат стомален комплекс(Сл. 2.7). Се разликуваат следниве главни типови на стоматични комплекси:
Ориз. 2.7.Главните типови на стомачни апарати: 1 – аномоцитни (кај сите виши растенија, освен конска опашка); 2 – дијацитни (кај папрати и цветни растенија); 3 – парацитски (кај папрати, конска опашка, цветни и угнетувачки); 4 – анизоцитна (само кај цветни растенија); 5 – тетрацитна (главно кај едноколини); 6 – анциклоцитен (кај папрати, гимносперми и цветни растенија)
1) аномоцитна(неуредно) – чуварите клетки немаат јасно дефинирани парастоматални клетки; карактеристични за сите повисоки растенија, со исклучок на четинари;
2) анизоцитна(нееднаква клетка) – чуварите на стомите се опкружени со три парастоматални клетки, од кои едната е многу поголема (или помала) од другите;
3) парацитичен(паралелна клетка) - една перистоматална клетка (или повеќе) се наоѓа паралелно со заштитните клетки;
4) дијацитичен(вкрстени клетки) - две парастоматални клетки се наоѓаат нормално на заштитните клетки;
5) тетрацит(од грчки тетра- четири) – главно во едноколини;
 Стомите се наоѓаат на долната страна на листот, но водни растенијасо пловечки лисја, тие се само на горната страна на листот. Врз основа на обликот на лисните епидермални клетки и локацијата на стомите, еднокотиледоно растение може да се разликува од двокотиледоно (сл. 2.8). Вистинските епидермални клетки на листовите на двокотиледоните растенија се брановидни по контури (сл. 2.9), додека кај еднокотиледоните растенија тие се издолжени, ромбични по форма.
Стомите се наоѓаат на долната страна на листот, но водни растенијасо пловечки лисја, тие се само на горната страна на листот. Врз основа на обликот на лисните епидермални клетки и локацијата на стомите, еднокотиледоно растение може да се разликува од двокотиледоно (сл. 2.8). Вистинските епидермални клетки на листовите на двокотиледоните растенија се брановидни по контури (сл. 2.9), додека кај еднокотиледоните растенија тие се издолжени, ромбични по форма.
Ориз. 2.8.Локација на стомите на епидермисот (поглед од површината): А- дикотиледони растенија: 1 - почетна буква; 2 – лубеница; б- едноколини: 3 – пченка; 4 - ирис
Видовите на стомати може да се поделат според нивното ниво на локација во однос на површината на епидермисот на следниот начин.
1.7.1. Стомите се наоѓаат во иста рамнина како и епидермисот. Најчестиот тип и обично не е наведен во описот на микроскопијата на лековити растителни материјали, т.е. овој став е изоставен. Дијагностички знациќе има или испакнати или потопени стоми.
1.7.2. Испакнати стомати - стомати лоцирани над епидермисот. Вообичаено, кога се ротира микроскопскиот микроскоп (кога леќата е спуштена), прво се откриваат такви стоми, па дури тогаш се појавуваат епидермални клетки, па речиси е невозможно да се фатат на фотографија од површината на листот, како и да ги прикаже на цртеж. Во истата рамнина како и епидермисот, таквите стоми може да се видат во попречни пресеци, но за ова, делот мора да помине низ стомите, што е тешко да се добие со оглед на нивната ретка локација на листот. Таквите стоми се карактеристични, на пример, за лисјата од мечкино грозје.
1.7.3. Потопени стомати - стомати потопени во епидермисот. Кога се набљудуваат под микроскоп со ротирање на микрошрафот (при спуштање на леќата), епидермалните клетки најпрво јасно се откриваат, а потоа станува возможно појасно да се видат контурите на стоматите. Исто така, тешко е да се прикажат на фотографии и цртежи на препарати од површината. Се наоѓа во лисјата на крин на долината, лисјата на часовникот, лисјата од еукалиптус. Понекогаш вдлабнатините во кои се наоѓаат стомите се обложени или покриени со влакна и се нарекуваат стомачни крипти.
1.8. Видови стоматални клетки
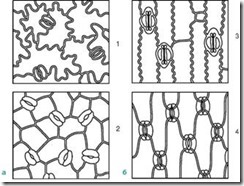
Во литературата се опишани 19 типови; ги избравме само оние што се користат во анализата на медицинските растителни суровини**.
Ориз. 63.Видови стоматални клетки. А – леќа форма; Б – сферично; Б – во облик на капа; Г – скафоид
1.8.1. Леќата - 2 идентични ќелии во форма на полумесечина наредени симетрично. На фронталната рамнина, задебелувањето на школка е речиси униформно. Пукнатината е фузиформна (сл. 63, А). Типот на стомачни клетки е карактеристичен за повеќето растенија.
1.8.2. Сфероиден - две идентични, силно кружно заоблени ќелии се наоѓаат симетрично. На фронталната рамнина, задебелувањето на школка е речиси униформно. Слот е кружен (слика 63, Б).
1.8.3. Во облик на капа - две идентични ќелии во облик на полумесечина во поларните делови имаат задебелувања во форма на капа. Пукнатината е фузиформна (сл. 63, Б). Пронајден во ракавици за лисици.
1.8.4. Скафоид - внатрешните ѕидови на стоматалните клетки се задебелени. Пукнатината е фузиформна (сл. 63, D). Забележано во стоковска трева и лисја од часовници.
Механизмот на работа на стомите се одредува според осмотските својства на клетките. Кога површината на листот е осветлена од сонцето, хлоропластите на заштитните ќелии подлежат на активен процесфотосинтеза. Заситеноста на клетките со фотосинтетички производи и шеќери повлекува активно влегување на јони на калиум во клетките, што резултира со концентрација клеточен сокзголемување на заштитните ќелии. Постои разлика во концентрацијата на клеточниот сок на парастоматалните и заштитните клетки. Поради осмотските својства на клетките, водата од парастоматалните клетки влегува во заштитните клетки, што доведува до зголемување на волуменот на вторите и нагло зголемување на тургорот. Задебелувањето на „абдоминалните“ ѕидови на заштитните клетки свртени кон стоматалната пукнатина обезбедува нерамномерно истегнување на клеточниот ѕид; заштитните клетки добиваат посебна форма во облик на грав, а стоматалната пукнатина се отвора. Кога интензитетот на фотосинтезата се намалува (на пример, во вечерните часови), формирањето на шеќери во заштитните ќелии се намалува. Приливот на јони на калиум престанува. Концентрацијата на клеточниот сок во заштитните клетки е намалена во споредба со парастоматалните клетки. Водата ги напушта заштитните клетки со осмоза, намалувајќи го нивниот тургор; како резултат на тоа, стоматалната пукнатина се затвора ноќе.
Клетките на епидермисот се цврсто затворени заедно, благодарение на што епидермисот врши голем број функции:
Спречува пенетрација на патогени организми во растението;
Ги штити внатрешните ткива од механички оштетувања;
Регулира размена и транспирација на гасови;
Преку него се ослободуваат вода и соли;
Може да функционира како ткиво за вшмукување;
учествува во синтезата на различни супстанции, перцепцијата на иритации и движењето на листовите.
Трихоми - израстоци на епидермални клетки со различна форма, структура и функции: влакна, лушпи, влакна итн. Тие се поделени на покривни и жлезди. вроден трихоми,за разлика од прикриените, тие имаат клетки кои лачат секрет. Покривање на влакнатаформирање на волнена, филц или друга покривка на растението, одразувајќи дел од сончеви зрациа со тоа и намалување на транспирацијата. Понекогаш влакната се наоѓаат само таму каде што се наоѓаат стомите, на пример, на долната страна на листот подбел. Кај некои растенија, живите влакна ја зголемуваат вкупната испарувачка површина, што помага да се забрза транспирацијата.
Големините на трихом значително се разликуваат. Најдолгите трихоми (до 5-6 см) ги покриваат семките од памук. Покривните трихоми имаат форма на едноставни единечни или повеќеклеточни, разгранети или ѕвездени влакна. Покриените трихоми може да останат живи долго време или брзо да умрат, полнејќи се со воздух.
Тие се разликуваат од трихоми, кои произлегуваат само со учество на епидермалните клетки. итни случаи, во чие формирање учествуваат и подлабоко лоцирани ткива на субепидермалните слоеви.
Анатомски и дијагностички знаци кои имаат највисока вредности висока варијабилност во одредувањето на медицинските суровини. Влакната може да бидат едноставни или капитатни, кои пак може да бидат едноклеточни или повеќеклеточни. Повеќеклеточните влакна можат да бидат едноредни, дворедни или разгранети.
1.9.1. Едноставни влакна.
А. Едноставни едноклеточни влакна
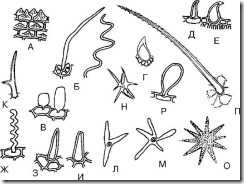
Ориз. 68.Едноставни едноклеточни влакна. А – папиларен; Б – филаментозен; Б – везикуларен; Г – спинозен; D – во облик на кука; Е – во форма на реплика; F – тап брановиден налик на конец; Z – во облик на остар конус; I – во форма на тап конус; К – двокрај; L – трикратна; М, Н, О – повеќекратни; P – грутка; P – во форма на клуб
1. Папиларен (Сл. 68, А; Сл. 69-74) - ниски, но широки израстоци на епидермалните клетки. Најчесто се формираат на ливчиња. Тие можат да бидат тапи-апикални или акутно-апикални. Ги има на лисјата од мајчина душица, цветовите на крин од долината, тробојните виолетови цветови, по должината на рабовите на листовите од стотумот, во брадестата гентинска трева итн.
2. Во облик на конус (Сл. 68, H, I; Сл. 75-79) - прави влакна наклонети под агол на површината, завршувајќи со тап (во облик на тап конус) или остар (остар конусен) крај. Најчести влакна. Обично тие се прави. На листовите од мајчина душица се забележуваат влакна наклонети кон површината. Зашилени влакна во облик на конус се наоѓаат на листовите од сена, листовите од лингон, во тробојната виолетова трева (на лисјата), во тревата мајчина душица итн. .
3. Филиформни (Сл. 68, Б, Г; Сл. 80, 81) – тенки и долги влакна. Може да биде исправен или брановиден. На епидермисот на плодовите од глог и малина се наоѓаат права влакна слични на конец. Брановидни конец-како влакната се забележани на епидермисот на виолетовите ливчиња (сл. 82).
4. Во облик на реплика (Сл. 68, Е; Сл. 83) - влакна со проширена основа и тесен заоблен или прави крај. Се наоѓа во овошјето од хмељ и тревата.
5. Во облик на мочен меур (Сл. 68,Б; Сл. 84) – влакна во вид на мали меурчиња. Тие можат да се најдат, на пример, во цвеќињата на смил.
6. Во облик на кука (Сл. 68, Е; Сл. 85-87) - влакна, зашилени на врвот и закривени во форма на кука. Овие влакна може да се најдат во основата на листовите од мечкино грозје, на површината на листовите од јаболчницата, листовите од сена и плодовите од анасон.
7. Реси (Сл. 88, 89) - долги израстоци на епидермалните клетки лоцирани по должината на работ на лист, ливче или сепал. Таквите влакна се наоѓаат, на пример, по должината на рабовите на ливчињата од крин и виолетова боја.
8. Во облик на 'рбет (Сл. 68, Д; Сл. 90) – влакната се речиси кружни во форма со зашилен крај. Во тревата се забележани тробојни темјанушки.
9. Во форма на клуб (Сл. 68, П; Сл. 91) - влакна со проширен краен дел, кои потсетуваат на палка. Се наоѓа на ливчиња од мајчина душица и виолетови ливчиња во тробојка.
10. Двоен крај (Сл. 68, К) - влакна разгранети на два краја. Забележано во тревата од овчарската чанта.
11. Трикратно (повеќе точки) (Сл. 68, L-O) - влакна разгранети на три (или повеќе) краеви. Забележано во тревата од овчарската чанта.
12. Трнлив (Сл. 68, P) - влакна во облик на конус со испакнатини (испакнатини) на нивната површина.
Б. Едноставни повеќеклеточни влакна со еден ред
Контурата на повеќеклеточното влакно може да се совпадне со онаа на едноклеточното влакно, но содржи две или повеќе клетки и, соодветно, се нарекува слична едноклеточна влакно што го покажува бројот на клетки, на пример, влакно во облик на 2-клетка, 11-клеточна филаментозна коса, влакно во форма на конус од 10-15 клетки.

Ориз. 92.Едноставни повеќеклеточни влакна. А - во форма на конус; Б – во облик на меур; Б – артикуларно; G – во облик на камшик; Д – пердувести; Е – Т-облик; F – двоен ред; Z – лушпест; I – зрак; L – паралелно; М – чаталесто; К – наежвам
1. Во облик на конус (Сл. 92, А; 93-96). Најраспространетите влакна. Постојат прави и наклонети кон површината, вторите се ретки (на пример, во тревата мајчина душица). Влакната во облик на конус може да бидат во форма на остри или тапи конусни. Почести се влакната во облик на остри конуси (во билката од оригано, во листовите од нане, жалфија итн.). Тапи конуси се наоѓаат во цветовите на невен.
2. Во облик на реплика - влакна со проширена основа и тесен заоблен или прави крај.
3. Во облик на мочен меур (Сл. 92, Б; Сл. 97) – влакна во вид на мали меурчиња. На површината има јајници од смил.
4. Филиформни - тенки и долги повеќеклеточни влакна.
5. Во облик на кука - влакна насочени на врвот и закривени во форма на кука.
6. Гасеници (Сл. 98, 99) - влакна со речиси иста дебелина по целата должина, составени од речиси идентични кратки ќелии, со тап крај и кои потсетуваат на гасеница. Тие се забележани во тревата на тробојните темјанушки и во тревата на сукцесијата.
7. Реси (Сл. 100) - долги повеќеклеточни израстоци на епидермалните клетки лоцирани по должината на работ на лист, ливче или сепал. Секвенци се наоѓаат во тревата.
8. Неверојатни (Сл. 92, Д; Сл. 101-104) - влакна кои имаат повеќеклеточна основа, која се состои од синџир на кратки ќелии и долга филаментозна извртена завршна клетка. Во литературата, таквите влакна се опишани како камшички, жици, нишки или филц. Видот на косата е карактеристичен за семејството Asteraceae. Се наоѓа во тревата од ајдучка трева, цвеќињата од смил, лисјата од подбел, цветовите од танзи итн.
9. Зглобна (Сл. 92, Б; Сл. 105, 106) - влакна кои имаат проширени клеточни основи на местата на нивната артикулација (налик на зглоб). Слични влакна се наоѓаат во тревата од мајчина матица, на ливчиња од оригано и ретко во тревата од мајчина душица.
10. Трнлив - влакна кои имаат испакнатини (испакнатини) на нивната површина. Двоклеточни туберозни влакна се забележани, на пример, во зеленилото на термопсис.
Б. Едноставни повеќеклеточни разгранети влакна
1. Паралелно (Сл. 92, L; Сл. 107, 108) - влакна, составени од две долги прави ќелии (влакна), споени на основите. Се среќава во цветовите на липата, поретко кај шипинките и глоговите.
2. чаталестите (Сл. 92, М; Сл. 109, 110) - влакна, составени од две долги намотани ќелии (влакна), споени на основите. Забележано кај цветовите од липа, многу ретко кај цветовите и плодовите на глог.
3. Во облик на ѕвезда (Сл. 111) - влакна, составени од 3 или повеќе долги згрчени ќелии (влакна), споени на основите. Се наоѓа во цветовите од липа.
4. Лушпеста (Сл. 92, H) - влакна кои се состојат од повеќеклеточна плоча (во форма на розета) и кратко стебленце (стебленцето може да отсуствува). Ваквите влакна се наоѓаат во морскиот тревник. (Ова име за влакната е дадено во согласност со модерната ботаничка терминологија; според други извори, овие влакна се нарекуваат ѕвездести или коримбозни лушпести).
5. Т-форма (Сл. 92, Д) е двокрака влакно со едно или повеќеклеточно стебленце. Во литературата тие се опишани и како во облик на рог, во облик на рокер.
6. Цирус (Сл. 92, Д) - повеќеклеточни влакна, кои потсетуваат на разгрането дрво. Најден, на пример, во лопен.
Г. Едноставни повеќеклеточни повеќередни влакна
1. Двоен ред (Сл. 92, Г; Сл. 112, 113) – влакна, чии ќелии се наредени во два реда. Тие можат да се забележат во цвеќињата на невен и исушената трева.
2. Греди (Сл. 92, I) - влакна кои се состојат од две или повеќе клетки цврсто споени една со друга, формирајќи пакет.
3. наежвам (Сл. 92 К, Сл. 114) - повеќеклеточни влакна, составени од влакна споени едни со други различни должинии има слободни зашилени краеви. Таквите влакна се обично влакна на мувата (семејство Asteraceae).Во литературата тие се нарекуваат и назабени сложени, во облик на шило.
1.9.2. Капитирајте влакна. Капитатните влакна се нарекуваат и вродени влакна. Тие можат да бидат едноклеточни или повеќеклеточни.

Ориз. 115.Капитирајте влакна. А – со едноклеточно стебленце и едноклеточна глава; Б – со дворедна нога и едноклеточна глава; Б – со дворедна нога и дворедна глава; G – со едноклеточно стебленце и повеќеклеточна глава; Д – со повеќеклеточно стебленце и едноклеточна глава; F – со повеќеклеточно стебленце и повеќеклеточна глава; Z – едноклеточен (гори); Јас - тироидна жлезда
A. Едноклеточни капитатни влакна
Претставниците на овој тип на влакна можат да бидат во форма на конус или во форма на ретер, но секогаш имаат глава на крајот. Таквите влакна се наоѓаат, на пример, на лисјата од коприва. Б. Повеќеклеточни капитатни (жлезди) влакна
1. Влакна со повеќеклеточна глава и едноклеточно стебленце (Сл. 115, Д; Сл. 116-120). Се наоѓа во тревата и лисјата од мечкино грозје.
2. Влакна кои имаат едноклеточна глава и едноклеточно стебленце (Сл. 115, А; Сл. 121-123). Се наоѓа во тревата од мајчина трева, лисја од жалфија, цветови од бозел, лисја од нане.
3. Влакна со едноклеточна глава и повеќеклеточно стебленце (Сл. 115, Д; Сл. 124). Се наоѓа во тревата од мајчина трева, лисјата на жалфија, цвеќињата на невен и петелките од камилица.
4. Влакна со повеќеклеточна глава и повеќеклеточно стебленце (Сл. 115, Г; Сл. 125). Се наоѓа во тревата од мајчина трева, лисја од мечкино грозје, цвеќиња од липа, цвеќиња од невен, виолетова трева.
5. Влакна со едноклеточна глава и стебленце во два реда (Сл. 115, Б; Сл. 126). Се наоѓа во цветовите на невен и смил.
6. Влакна со дворедна глава и дворедна дршка (Сл. 115, Б; Сл. 127-130). Тие се наоѓаат во цветовите на невен и смил, и во тревата на барска трева.
7. Во форма на клуб повеќеклеточни вродени влакна (сл. 131-133; види Сл. 155, Д) - влакна кои имаат контура со проширен краен дел, што потсетува на палка. Тие можат да се забележат, на пример, во лисјата на лингонбери и виолетова трева.
8. Тироидната жлезда повеќеклеточните вродени влакна (сл. 115, I; Сл. 134) се штит од полигонални тенкоѕидни клетки кои седат на едно или две-клеточно кратко стебленце. Кутикулата на scutellum се оддалечува од клетките секретирани под него етерично масло. Ги има во плодовите од хмељ.
1.10. Природата на задебелување на клеточните ѕидови и влакната што ја покриваат кутикулата
А. Карактер на задебелување на клеточните ѕидови
1.10.1. Тенки ѕидови (Сл. 135-137). Повеќето влакна се со тенки ѕидови. Клеточните ѕидови на повеќеклеточните долги тенкоѕидни влакна понекогаш пропаѓаат, нарушувајќи ја рамномерната контура на косата. Таквите влакна, на пример, се наоѓаат на долниот епидермис на листот какали.
1.10.2. Дебели ѕидови (Сл. 138, 139). Ги има во шипка, жичана трева, трева од лижавче дахуриска итн.
1.10.3. Нерамномерно задебелена. Во тревата може да се види мајчина боја (сл. 140). Во оваа категорија спаѓаат и влакната од шипинка со коси попречни пори (сл. 141, 142).
Б. Природата на влакната што ја покриваат кутикулата
1.10.4. Мазна површина (Сл. 143). Влакната со таква површина се забележани, на пример, во лисјата на хлебните, тревата celandine итн.
1.10.5. брадавица површина (види Сл. 1, Г; Сл. 144, 145) - епидермисот формира испакнатини во форма на туберкули (брадавици). Има влакна, на пример, во виолетова трева, во трева од мајчина душица, во трева од мајчина трева, во лисја од нане, во листови од сена итн. Во овој случај, површината на косата може да биде малку брадавичка кога се формираат мали испакнатини на кутикулата , и грубо брадавици кога се формираат значителни испакнатини. Во првиот случај, влакната во виолетова трева може да се наведат како пример, во вториот - влакна од лисја од сена и трева од мајчина душица.
1.10.6. Груба површина (види Сл. 68, P) - кутикулата формира многу големи испакнатини, на пример, влакната на тревата Thermopsis lanceolata.
10.7. Збрчкана површина - набори или бранови на кутикулата на косата од основата до нејзиниот врв (сл. 146). Почесто е забележан кај папилите, на пример, кај центавирската трева и цветовите на бозелот. Сепак, обичните влакна може да имаат и збрчкана површина, на пример, во жичаната трева. Во основата, влакната од листовите нане имаат збрчкана површина (сл. 147).
1.10.8. Отворена површина - епидермисот формира кратки испакнатини во форма на ленти (туберкули, брадавици). Ретко е и е средна опција помеѓу брадавичка и збрчкана површина. Тоа е забележано, на пример, на влакната на постарите цветови (сл. 148).
1.11. Карактеристики на местата за прицврстување на косата
1.11.1. Редовни местаприлози (Сл. 149, 150) - влакната се прикачени на клетката или помеѓу клетките на епидермисот. Најчестиот вид се среќава, на пример, во лисјата од подбел, лисјата од нане, лисјата од жалфија, тревата од мајчина трева, билката виолетова итн.
1.11.2. Израстоци од епидермалните клетки (види Сл. 68, А; 69-74). Забележано во цветовите на бозелот, стоковата трева, ливчињата од мајчина трева, брадестата трева на генијалецот, епидермисот на плодовите од анасон итн.
1.11.3. Во основата на косата се формира розета од епидермални клетки (Сл. 151, 152). Се наоѓа во листовите сена, лисјата на хлебните, билката од оригано итн.
1.11.4. Продолжена основа за коса (сл. 153). Се наоѓа во лисјата на хлебните.
1.11.5. Проширениот дел од основата на косата е потопен во основата на субепидермалните ткива - избувнувачки (види Сл. 115, 3). Може да се забележи, на пример, во лисјата од коприва.
1.11.6. Повеќеклеточна основа за коса (сл. 154). Можете да набљудувате, на пример, жици во тревата.
Честопати влакната се откинуваат, оставајќи ги нивните точки за прицврстување на епидермисот, што исто така треба да се забележи како анатомски и дијагностички знак на лековити растителни материјали.
СЕКУНДАРНО ПОКРИВНО ТКИВО
Секундарното интегрално ткиво се нарекува перидерма.Ова е сложено покривно ткиво на стеблата, корените и ризомите на повеќегодишните растенија. Го заменува епидермисот на аксијалните органи, кој постепено изумира и излева. Перидермот е формиран од фелоген (секундарен меристем). Фелогенот се формира во епидермисот, субепидермалниот слој, па дури и во длабоките слоеви на аксијалните органи. Фелогените клетки се делат на следниов начин: клетките од плута се поставени нанадвор, а живите паренхимски клетки на фелодермот се поставени навнатре. Фелодермските клетки на стеблата содржат хлоропласти.
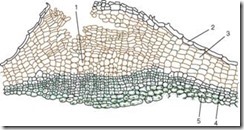 Плута се состои од мртвите клетки, во која клеточниот ѕид е импрегниран со супстанцијата слична на маснотии суберин. Клетките се наредени во рамномерни редови и имаат правоаголна форма(на пресек), цврсто се вклопуваат еден до друг, формирајќи повеќеслојна кутија. Плута ги штити внатрешните живи ткива од губење на влага, ненадејни температурни флуктуации и пенетрација на микроорганизми. На живите ткива што лежат под приклучокот им е потребна размена на гасови и отстранување на вишокот на влага. Токму затоа, под стоматите, поради поделбата на субепидермалните слоеви (дури и пред појавата на перидермот), а потоа и фелогените, живи, лабаво лоцирани паренхимски клетки со многу меѓуклеточни простори, т.н. изведување ткаењенов, со што се крши епидермисот и се создава можност за размена на гасови и транспирација со надворешната средина. Ова структурно образованиеповикани леќата(Сл. 2.12).
Плута се состои од мртвите клетки, во која клеточниот ѕид е импрегниран со супстанцијата слична на маснотии суберин. Клетките се наредени во рамномерни редови и имаат правоаголна форма(на пресек), цврсто се вклопуваат еден до друг, формирајќи повеќеслојна кутија. Плута ги штити внатрешните живи ткива од губење на влага, ненадејни температурни флуктуации и пенетрација на микроорганизми. На живите ткива што лежат под приклучокот им е потребна размена на гасови и отстранување на вишокот на влага. Токму затоа, под стоматите, поради поделбата на субепидермалните слоеви (дури и пред појавата на перидермот), а потоа и фелогените, живи, лабаво лоцирани паренхимски клетки со многу меѓуклеточни простори, т.н. изведување ткаењенов, со што се крши епидермисот и се создава можност за размена на гасови и транспирација со надворешната средина. Ова структурно образованиеповикани леќата(Сл. 2.12).
Ориз. 2.12.Структура на перидермот со леќата: 1 – потпорното ткиво на леќата; 2 – остатоци од епидермисот; 3 – плута (фелема); 4 – фелоген; 5 – фелодерм
Леќата, која изгледа како мали туберкули, е јасно видлива на површината на ластарите од дрвја и грмушки (сл. 2.13).
На стеблата од бреза, нивните остатоци се забележани во форма на карактеристични попречни црни ленти, на трепетлика тие имаат форма на дијаманти.
Терциерно интегрално ткиво
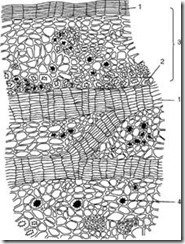 Кора (ритид)е терциерно интегрално ткиво кое се формира кај повеќегодишни растенија во коренот, стеблото и ризомот. Секоја година во подлабоки слоеви се поставува нов слојфелоген и се формира перидермот. Надворешниот слој на перидермот - плута - ги изолира сите прекриени ткива, како резултат на што тие умираат. Така, збирката на бројни перидерми со мртви ткива меѓу нив е кората (сл. 2.14).
Кора (ритид)е терциерно интегрално ткиво кое се формира кај повеќегодишни растенија во коренот, стеблото и ризомот. Секоја година во подлабоки слоеви се поставува нов слојфелоген и се формира перидермот. Надворешниот слој на перидермот - плута - ги изолира сите прекриени ткива, како резултат на што тие умираат. Така, збирката на бројни перидерми со мртви ткива меѓу нив е кората (сл. 2.14).
Ориз. 2.14.Кора од даб: 1 – слоеви плута; 2 – влакна; 3 – остатоци од примарната кора; 4 – друзен на калциум оксалат