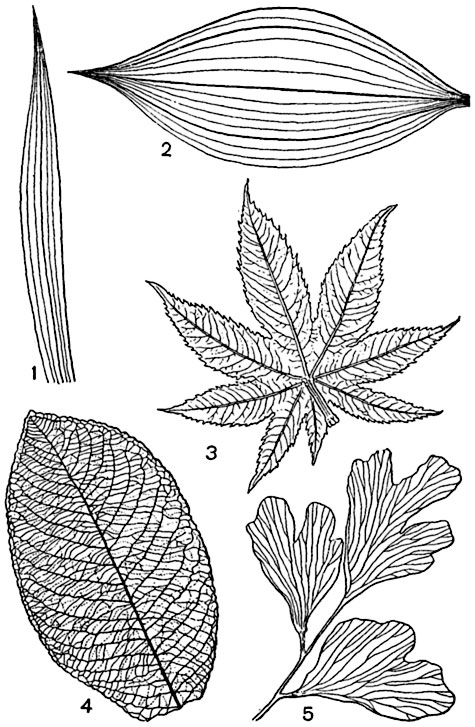
FABRICS. CLASSIFICATION OF FABRICS.
At the heart of the organization higher plants lies the principle of cell specialization, which lies in the fact that each cell of the body does not perform all its inherent functions, but only some, but more fully and perfectly.
Fabrics- stable, naturally repeating complexes of cells, similar in origin, structure and adapted to perform one or more functions.
There are various classifications of fabrics, but they are all quite arbitrary.
Depending on the main function, several groups of plant tissues are distinguished.
1. educational fabrics, or meristems,- have the ability to divide and form all other tissues.
Primary;
Secondary;
Tertiary.
3. Main fabrics- make up most of the plant body. The following main fabrics are distinguished:
Assimilation (chlorophyll-bearing);
Stockers;
Airborne (aerenchyma);
Aquifers.
4. Mechanical fabrics(supporting, skeletal):
Collenchyma;
Sclerenchyma.
5. Conductive fabrics:
Xylem (wood) is an ascending tissue;
Phloem (phloem) is a tissue of descending flow.
6. Excretory tissues:
External:
Glandular hairs;
Hydathodes - water stomata;
Sunbirds;
Internal:
Excretory cells with essential oils, resins, tannins;
Multicellular receptacles for secretions, lacticifers.
Based on the ability of cells to divide, two types of tissues are distinguished: educational, or meristems, And permanent- integumentary, excretory, basic, mechanical, conductive.
The fabric is called simple, if all its cells are identical in shape and function (parenchyma, sclerenchyma, collenchyma). Complex tissues consist of cells that are different in shape, structure and function, but are related by a common origin (for example, xylem, phloem).
There is also a classification of tissues based on their origin (ontogenetic). According to this classification, primary and secondary tissues are distinguished. From the primary meristem located at the top of the shoot and the tip of the root, as well as from the seed embryo, primary constants tissues (epidermis, collenchyma, sclerenchyma, assimilation tissue, epiblema). Cells of permanent tissues are incapable of further division. From cells of a specialized meristem - procambia - are formed primary conductors tissues (primary xylem, primary phloem).
From the secondary meristem - cambium - are formed secondary tissues: secondary xylem, secondary phloem; from phellogen plug, phelloderm, and lentils are formed, which arise when the stem and root thicken. Secondary tissues are typically found in gymnosperms and dicotyledonous angiosperms. The powerful development of secondary tissues - wood and bast - is characteristic of woody plants.
EDUCATIONAL FABRICS
Educational fabrics Thanks to the constant mitotic division of their cells, they ensure the formation of all plant tissues, i.e. actually shape his body. Any cell in its development goes through three stages: embryonic, growth and differentiation stage (that is, the cell acquires a certain function). As the embryo differentiates, the primary meristem is retained only at the tip of the future shoot (in the growth cone) and at the tip of the root - apical (apical) meristems. The embryo of any plant consists of meristem cells.
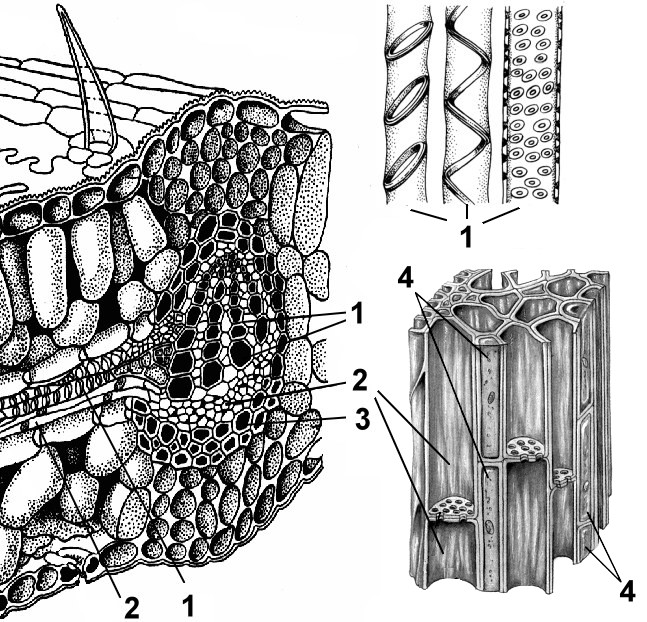
Cytological characteristics of meristems. Typical features are most clearly expressed in the apical meristems. These meristems are composed of isodiametric polyhedral cells not separated by intercellular spaces. Their shells are thin, contain little cellulose and are stretchable.
The cavity of each cell is filled with dense cytoplasm with a relatively large nucleus occupying a central position and intensively dividing by mitosis. The hyaloplasm contains many diffusely scattered ribosomes, proplastids, mitochondria and dictyosomes. There are few vacuoles and they are small. Conducting tissues are formed from a meristem that has a prosenchymal shape and large vacuoles - procambium and cambium. Procambium cells are polygonal in cross section, cambium cells are rectangular.
Cells that retain their meristematic properties continue to divide, forming more and more new cells called initials. Some of the daughter cells differentiate, turning into cells of various tissues, they are called derivatives of initials. Initial cells can divide indefinitely many times, and derivatives of initials divide one or more times and develop into permanent tissues.
Based on their origin, primary and secondary meristems are distinguished.
Primary meristems
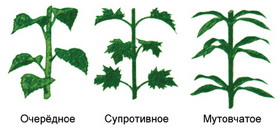
Primary meristems arise directly from the meristem of the embryo and have the ability to divide. According to their position in the plant, primary meristems can be apical (apical), intercalary (intercalary) and lateral (lateral).
Apical (apical) meristems- such meristems that are located in adult plants at the tops of stems and tips of roots and ensure body growth in length. In the growth cone of the stems, two meristematic layers are distinguished: the tunic, from which the integumentary tissue and peripheral part primary cortex, and the corpus from which the inner part of the primary cortex and the central axial cylinder(Fig. 2.3).
Rice. 2.3. Apical meristems of the stem: A- longitudinal section: 1 - growth cone; 2 - leaf primordium; 3 - tubercle of axillary bud;
There are three layers at the root tip:
1) dermatogen, from which the primary integumentary-absorbing tissue - rhizoderm - is formed;
2) periblema, from which the tissues of the primary cortex develop;
3) plerome, forming the tissues of the central axial cylinder.
Lateral (lateral) meristems they can be primary or secondary in origin; on a cross section of the axial organs they look like rings. An example of a primary lateral meristem is the procambium and pericycle. From procambia The cambium and the primary elements of the vascular-fibrous bundles (primary phloem and primary xylem) are formed, while the procambium cells directly differentiate into the cells of the primary conducting tissues.
The lateral meristems are located parallel to the surface of the organ and ensure the growth of the axial organs in thickness.
Intercalary meristems more often primary and preserved in the form of separate areas in zones of active growth in various parts plants (for example, at the base of leaf petioles, at the base of internodes). At the base of internodes in cereals, the activity of this meristem leads to elongation of the internodes, which ensures the growth of the stem in length.
Secondary meristems
Secondary meristems include lateral and wound meristems.
Lateral (lateral) meristems presented cambium And phellogen. They are formed from promeristems (procambium) or permanent tissues by their dedifferentiation. Cambium cells are divided by septa parallel to the surface of the organ (periclinally). Elements of secondary phloem develop from cells deposited outward by the cambium, and elements of secondary xylem develop from cells deposited inward. The cambium, which arose from permanent tissues through dedifferentiation, is called additional In structure and function it does not differ from the cambium, which arose from promeristems. Phellogen is formed from permanent tissues located in the subepidermal layers (under the epidermis). Dividing periclinally, phellogen separates future plug cells (phelleme) outward, and phelloderm cells inward. Thus, phellogen forms secondary integumentary tissue - periderm. The lateral meristems are located parallel to the surface of the organ and ensure the growth of the axial organs in thickness.
Wound meristems are formed when tissues and organs are damaged. Around the damage, living cells dedifferentiate, begin to divide, and thereby transform into a secondary meristem. Their task is to form a dense protective tissue consisting of parenchyma cells - callus. This tissue is whitish or yellowish in color, its cells have large nuclei and fairly thick cell walls. Callus occurs during grafting, ensuring the fusion of the scion with the rootstock, and at the base of the cuttings. It can form adventitious roots and buds, so it is used to obtain isolated tissue cultures.
COVERING TISSUE
Primary integumentary tissue
TO primary integumentary tissues include the epidermis, the epidermal itself, the parastomatal cells, the guard cells of the stomata and the trichomes.
Pectic substances and cellulose included in cell wall, can be subject to mucus formation with the formation slime And gums. They are polymeric carbohydrates related to pectin substances and are characterized by their ability to swell strongly when in contact with water. Gums in a swollen state are sticky and can be pulled out into threads, while mucus is very blurry and cannot be pulled out into threads. Pectic mucilages are found in representatives of the Liliaceae, Cruciferae, Malvaceae, Linden, and Rosaceae families, in contrast to cellulose mucilages, which are much less common (for example, in Orchids).
Stomata
They are highly specialized formations of the epidermis, consisting of two bean-shaped guard cells and a stomatal fissure (a kind of intercellular space between them). They are found mainly in the leaves, but are also found in the stem (Fig. 2.6). 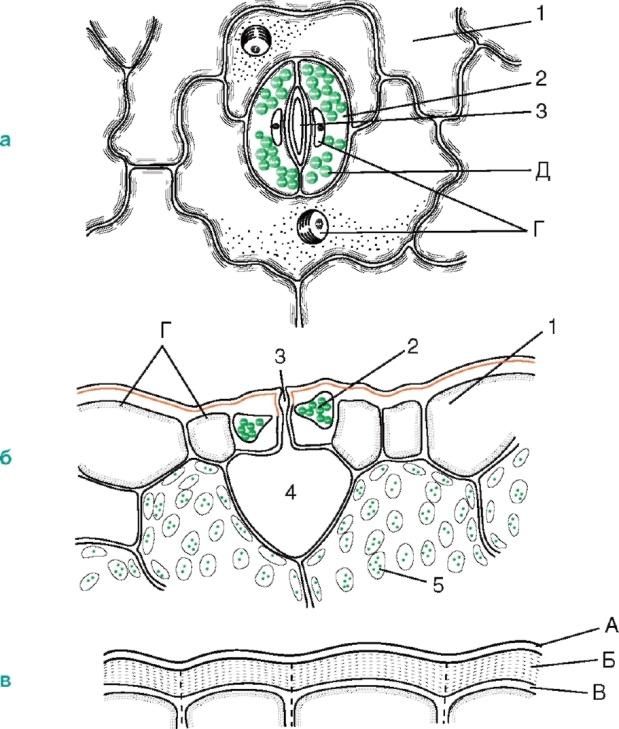
Rice. 2.6. Stomatal structure: a, b- the skin of a thyme leaf (top view and cross section); V- peel from the stem of Cereus (cactus family); 1 - actual epidermal cells; 2 - closing stomatal cells A; 3 - stomatal fissure; 4 - air cavity; 5 - cells of chlorophyll-bearing parenchyma; A - cuticle; B - cuticular layer - shell with suberin and wax; B - cellulose layer of the wall; G - nucleus with nucleolus; D - chloroplasts
The walls of the guard cells are thickened unevenly: the walls directed towards the gap (abdominal) are significantly thicker compared to the walls directed away from the gap (dorsal). The gap can expand and contract, regulating transpiration and gas exchange. Under the gap there is a large respiratory cavity (intercellular space), surrounded by mesophyll cells of the leaf.
The guard cells are surrounded by parastomatal cells, which together form stomatal complex(Fig. 2.7). The following main types of stomatal complexes are distinguished: 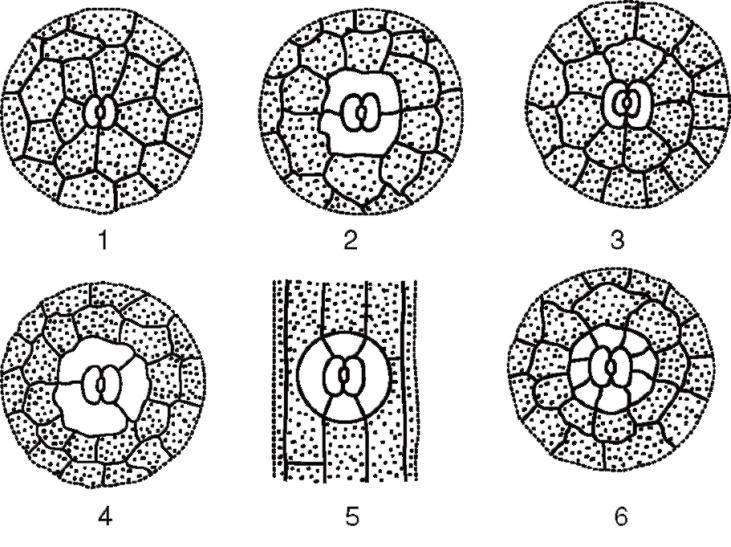
Rice. 2.7. The main types of stomatal apparatus: 1 - anomocytic (in all higher plants, except horsetails); 2 - diacytic (in ferns and flowering plants); 3 - paracytic (in ferns, horsetails, flowering and oppressive); 4 - anisocytic (only in flowering plants); 5 - tetracytic (mainly in monocots); 6 - ancyclocytic (in ferns, gymnosperms and flowering plants)
1) anomocytic(disorderly) - guard cells do not have clearly defined parastomatal cells; characteristic of all higher plants, excluding conifers;
2) anisocytic(unequal cell) - the guard cells of the stomata are surrounded by three parastomatal cells, one of which is much larger (or smaller) than the others;
3) paracytic(parallel cell) - one parastomatal cell (or more) is located parallel to the guard cells;
4) diacytic(cross-cell) - two parastomatal cells are located perpendicular to the guard cells;
5) tetracite(from Greek tetra- four) - mainly in monocots;
Stomata are located on the underside of the leaf, but aquatic plants with floating leaves, they are only on the upper side of the leaf. Based on the shape of the leaf epidermal cells and the location of the stomata, a monocotyledonous plant can be distinguished from a dicotyledonous one (Fig. 2.8). The actual epidermal cells of the leaves of dicotyledonous plants are wavy in outline (Fig. 2.9), while in monocotyledonous plants they are elongated, rhombic in shape. 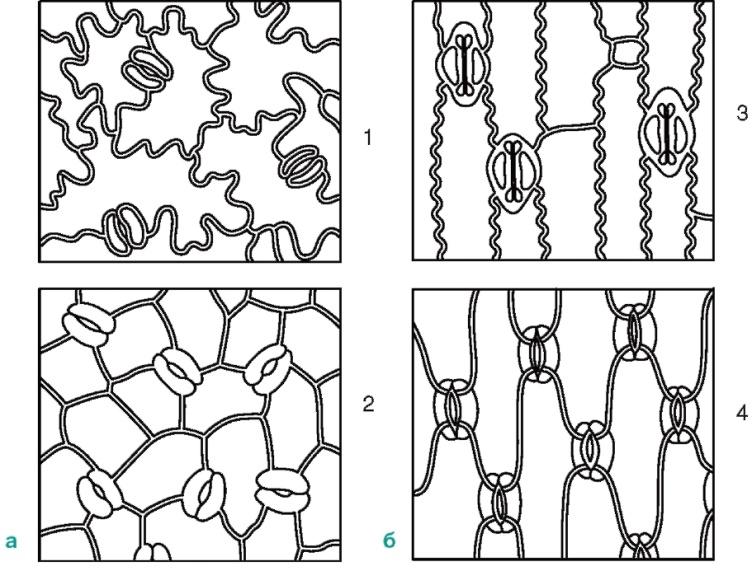
Rice. 2.8. Location of stomata on the epidermis (view from the surface): A-dicotyledonous plants: 1 - initial letter; 2 - watermelon; b-monocots: 3 - corn; 4 - iris
The types of stomata can be divided according to their level of location relative to the surface of the epidermis as follows.
1.7.1. Stomata located in the same plane as the epidermis. The most common type and is usually not indicated in the description of microscopy of medicinal plant materials, i.e. this paragraph is omitted. Diagnostic signs will be either protruding or submerged stomata.
1.7.2. Protruding stomata - stomata located above the epidermis. Usually, when the microscope microscope is rotated (when the lens is lowered), such stomata are first detected, and only then epidermal cells appear, so it is almost impossible to capture them in a photograph from the surface of a leaf, as well as to depict them in a drawing. In the same plane as the epidermis, such stomata can be seen in transverse sections, but for this, the section must pass through the stomata, which is difficult to obtain given their rare location on the leaf. Such stomata are characteristic, for example, of bearberry leaves.
1.7.3. Submerged stomata - stomata immersed in the epidermis. When observed under a microscope by rotating the microscrew (while lowering the lens), the epidermal cells are first clearly detected, then it becomes possible to more clearly see the contours of the stomata. It is also difficult to display them in photographs and drawings of preparations from the surface. Found in lily of the valley leaves, watch leaves, eucalyptus leaves. Sometimes the recesses in which the stomata are located are lined or covered with hairs and are called stomatal crypts.
1.8. Types of stomatal cells
There are 19 types described in the literature; we have selected only those that are used in the analysis of medicinal plant raw materials**.
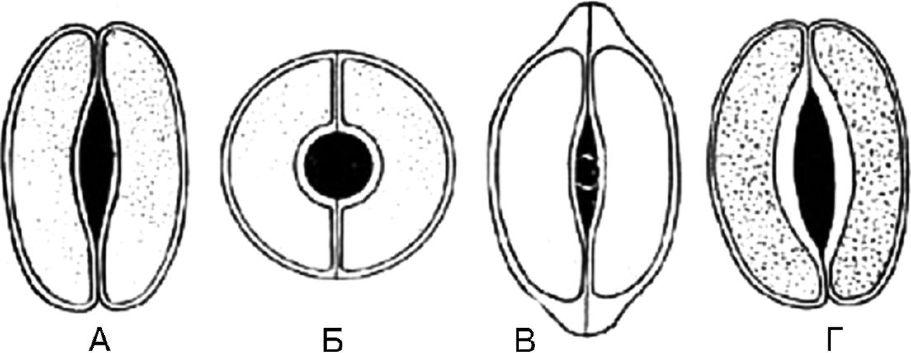
Rice. 63. Types of stomatal cells. A - lentiform; B - spherical; B - cap-shaped; G – scaphoid
1.8.1. Lenticular - 2 identical crescent-shaped cells arranged symmetrically. On the frontal plane, the thickening of the shell is almost uniform. The fissure is fusiform (Fig. 63, A). The type of stomatal cells is characteristic of most plants.
1.8.2. Spheroidal - two identical, strongly circularly curved cells are located symmetrically. On the frontal plane, the thickening of the shell is almost uniform. The slot is round (Fig. 63, B).
1.8.3. Cap-shaped - two identical crescent-shaped cells in the polar parts have thickenings in the form of a cap. The fissure is fusiform (Fig. 63, B). Found in foxgloves.
1.8.4. Scaphoid - the internal walls of stomatal cells are thickened. The fissure is fusiform (Fig. 63, D). Observed in centaury grass and watch leaves.
The mechanism of stomata operation is determined by the osmotic properties of cells. When the leaf surface is illuminated by the sun, an active process of photosynthesis occurs in the chloroplasts of the guard cells. Saturation of cells with photosynthetic products and sugars entails the active entry of potassium ions into the cells, as a result of which the concentration of cell sap in the guard cells increases. There is a difference in the concentration of cell sap of the parastomatal and guard cells. Due to the osmotic properties of the cells, water from the parastomatal cells enters the guard cells, which leads to an increase in the volume of the latter and a sharp increase in turgor. Thickening of the “abdominal” walls of the guard cells facing the stomatal fissure ensures uneven stretching of the cell wall; the guard cells acquire a distinct bean-shaped shape, and the stomatal fissure opens. When the intensity of photosynthesis decreases (for example, in the evening), the formation of sugars in guard cells decreases. The influx of potassium ions stops. The concentration of cell sap in guard cells is reduced compared to parastomatal cells. Water leaves the guard cells by osmosis, lowering their turgor; as a result, the stomatal fissure closes at night.
The cells of the epidermis are tightly closed together, thanks to which the epidermis performs a number of functions:
Prevents the penetration of pathogenic organisms into the plant;
Protects internal tissues from mechanical damage;
Regulates gas exchange and transpiration;
Water and salts are released through it;
Can function as suction tissue;
takes part in the synthesis of various substances, the perception of irritations and the movement of leaves.
Trichomes - outgrowths of epidermal cells of different shape, structure and functions: hairs, scales, bristles, etc. They are divided into covering and glandular. glandular trichomes, unlike coverts, they have cells that secrete secretions. Covering hairs forming a woolly, felt or other cover on the plant, reflecting part of sun rays and thereby reduce transpiration. Sometimes the hairs are found only where the stomata are located, for example, on the underside of a coltsfoot leaf. In some plants, living hairs increase the total evaporating surface, which helps accelerate transpiration.
Trichome sizes vary significantly. The longest trichomes (up to 5-6 cm) cover cotton seeds. Covering trichomes have the form of simple single or multicellular, branched or stellate hairs. Covering trichomes may long time remain alive or quickly die, filling with air.
They differ from trichomes, which arise only with the participation of epidermal cells. emergents, in the formation of which deeper located tissues of the subepidermal layers also participate.
Anatomical and diagnostic signs that have highest value and high variability in the determination of medicinal raw materials. Hairs can be simple or capitate, which in turn can be unicellular or multicellular. Multicellular hairs can be single-row, double-row or branched.
COVERING TISSUE.
- general characteristics and St.
- The structure and functions of the primary integumentary tissue - the epidermis.
Functions of the epidermis
Basic epidermal cells
- The structure of the secondary integumentary tissue - periderm
Education and mortgage
Structure of lentils
- The structure of the crust is retidoma.
General characteristics and saints.
Integumentary tissues cover the plant body from above, i.e. are located on the surface and are in direct contact with external environment. On the one hand, they protect plants from any adverse influences (from the penetration of microorganisms, from the evaporation of moisture), therefore the cells of the integumentary tissues are tightly closed, without intercellular spaces. On the other hand, they must ensure the connection of plants with the external environment (gas exchange, transpiration), therefore they have special structures.
Thus, the structure and properties of integumentary tissues are determined by the functions they perform.
Properties: 1. Multifunctionality, i.e. These are complex tissues, consisting of several types of cells that perform different functions.
2. Provide selective permeability (for gases, H 2 O and other substances), for which there are special structures.
3. Constant shift tissues with age, or changes in their functions.
There are: primary, secondary and tertiary integumentary tissues, which replace each other with age.
2. Primary integumentary tissue – epidermis (skin).
It arose as a result of the evolutionary adaptation of plants to terrestrial living conditions.
(Epi - Greek “above”, “above”, derma - “skin”).
a) Arises from the outer layer of the apical (apical) meristem of the shoot - protoderm, therefore - this is the primary integumentary tissue; protects and covers leaves, young stems, flowers and fruits of higher plants.
Epidermis is the primary covering tissue of a plant.
Usually single-layered, consists of 3 types of cells that perform different functions, i.e. This is a complex fabric.
Includes:
1) tightly closed main cells of the epidermis,
2) guard and secondary cells of stomata,
3) trichomes – derivatives of the epidermis in the form of outgrowths and hairs.
The functions and properties of the epidermis are determined by the structural features of these cells, which are ideally adapted for the terrestrial existence of plants.
(When transitioning to a secondary aquatic lifestyle flowering plants, for example, may lose stomata and protective structures of the main cells of the epidermis).
1. Basic cells of the epidermis.
1) They form 1 layer of living, transparent cells (since they are located on growing organs), therefore they transmit light well to the assemblies located below. cells where photosynthesis occurs.
Protective and integumentary function provide other features:
2) tightly closed, without intercellular spaces
3) thick outer cell walls, thin lateral
4) the side walls of cells often have a wavy shape, which improves their closure and adhesion to each other.
5) The cell membrane is complex: Bottom part consists of cellulose, and the upper one is cutinized.
6) The formation of a cuticle layer on the surface of the shell, which regulates the reduction of evaporation and gas exchange, is very characteristic.
Cuticle- a thin, transparent layer of cutin and plant waxes on the outer walls of the main cells. Wax is a fat-like substance that reduces permeability to water and gases. Consequently, the power of the cuticle, the distribution of waxes and cutin, the number and nature of pores determine the permeability to solutions and gases, chemical resistance, and bactericidal resistance.
In wet in a dry state, the cuticle is more permeable to gases and liquids than in a dry state, so absorption occurs through the cuticle when it rains aqueous solutions(foliar feeding of plants).
7) Well developed ap. Golgi and ER, therefore mainly. epidermal cells go synthesis row in-in(carbohydrates, proteins, etc.) (biosynthetic function of the epidermis)
Guard and secondary cells of stomata.
Stomata or stomatal apparatus- these are two bean-shaped guard cells with differently thickened walls and an intercellular space located between them - the stomatal fissure (the inner walls of the stomatal cells facing the fissure are thick, and the outer ones are thin).
Less often, side cells of the epidermis are found next to them, differing from its main cells.
Below the stomata there is a substomatal air cavity, surrounded by parenchyma cells and associated with the intercellular system of the organ.
Characteristic:
1) There are chloroplasts (photosynthesis of carbohydrates) and mitochondria (synthesis of ATP). Necessary because The opening of the stomatal fissure occurs actively with the expenditure of ATP energy, and the closure is passive.
2) Due to the different thickness of the walls of the guard cells, a gap is formed.
Hence, basic functions:
1 - transpiration(adjustable evaporation of water)
Gas exchange.
(Independently, study the mechanism of stomata).
The mechanism of opening and closing the stomatal fissure is based on osmotic phenomena, and all organelles of the protoplast of guard cells also take part.
- - K-Na – pump(rapid response mechanism). When the water content in the plant is high, K + transport proteins work in the plasmalemma of the guard cells, actively pumping K + ions from the surrounding cells into the cytoplasm, using ATP energy, and then they enter the vacuoles of the guard cells of the stomata. With a further increase in the concentration of K + ions, water follows them passively, along the concentration gradient, into the guard cells of the stomata from the surrounding cells. The volume of vacuoles increases, the osmotic pressure in the guard cells increases, and the cell membranes stretch. Due to the fact that the shells of the guard cells are thickened unevenly, the outer shells stretch more strongly, and the inner shells (facing each other) cannot stretch and diverge, a gap is formed between them - the stomata open. When there is a lack of water in the plant, the K+ ion transport proteins stop working. K-Na – the pump stops. In this case, K + ions passively leave the stomatal cells along a concentration gradient and go into the surrounding cells, followed by water. The guard cells lose turgor and passively close - the stomata close.
Thus, the opening of the stomatal fissure occurs actively, with the expenditure of energy, the source of which is carbohydrates accumulated by chloroplasts. Closing the gap occurs passively, without energy consumption.
- – Daily change in sugar concentration(a lot during the day, little at the end of the night) (because there are chloroplasts). During the day, photosynthesis occurs in the chloroplasts of guard cells. Consequently, in the light, the concentration of glucose in them increases and, as it accumulates, in the evening, water flows into the cells along the concentration gradient. The guard cells acquire an elastic turgor state, the stomata open. At night, photosynthesis stops, glucose is spent on the respiration of guard cells, and its concentration decreases. Because of this, along the concentration gradient, water leaves the guard cells, they lose turgor and collapse. The stomatal fissure closes. In the morning the process is repeated. It is now believed that this mechanism is limited, the main one being the operation of the K-Na pump.
Different plants have developed a certain rhythm of the stomatal apparatus. In most plants, stomata are open both day and night, closing only when the water content decreases. During hot hours, the stomata are usually closed, but in desert plants they are open (using evaporation, they cool the surface of the leaves and absorb water more actively). In some plants, such as Kalanchoe, stomata open at night and close during the day.
Types of stomata:
They are important in establishing the relationship of plant taxa.
1.- anamocytic stomata– have secondary cells that do not differ from the main ones. epidermal cells (characteristic for all groups of plants, except horsetails).
2.- diacite stomata– have two side cells, the common wall of which is perpendicular to the stomatal fissure (a number of flowering plants, the families Lamiaceae and Cloves have them).
3. – paracytic stomata- subsidiary cells are located parallel to the guard cells and stomatal fissure (there are ferns, horsetails and a number of flowering plants).
4. – anisocytic stomata– guard cells are surrounded by three side cells, one of which is either larger or smaller than the others (only in flowering plants).
5. – tetracite stomata- have 4 subsidiary cells (monocots have them).
6. – encyclocytic stomata– subsidiary cells form a narrow ring around the guard cells (found in ferns, gymnosperms and a number of flowering plants).
If the stomata are open, evaporation occurs as if there is no epidermis at all.
The number and location of stomata varies among different plants. On average, a plant has 100-700 stomata per 1 mm2. Usually, at land plants most of stomata are located on the underside of the leaves, on top there are very few or none at all. This is explained by the following reasons: 1) the leaves heat up more strongly from above, therefore, evaporation will be more intense and moisture loss will be greater; 2) carbon dioxide is formed in the soil and rises up, falling directly into the stomata; moreover, it is heavier than air and usually accumulates in the lower air layers.
In plants with vertical leaves (ribs), such as eucalyptus, stomata are distributed on both sides of the leaf. In aquatic plants with floating leaves, the stomata are located on the upper side.
- Trichomes– hairs, outgrowths and derivatives of the epidermis.
Shape and structure are a systematic feature for identifying plant species and genera.
There are: glandular and covering.
Glandular trichomes form and accumulate excreta and secretions. This essential oils(geranium, mint, etc.) or protective ingredients (nettle). A drop of oil is released under the cuticle, accumulates there, and then comes out when the cuticle breaks. The cuticle is restored and a new drop of essential oil accumulates under it.
Functions:
1-protective
2-thermoregulation
3-bactericidal (essential oils – phytoncides)
Covering trichomes consist:
1- often from dead cells air-filled (single or multicellular dead trichomes)
Functions: reflection of sunlight, favorable microclimatic conditions for stomata, reduction of heat. Consequently, a decrease in transpiration and fur. protection (plants of deserts and steppes).
2- rarely from living cells.
Functions: increase total evaporation. surface. Consequently, increased evaporation (plants of the humid tropics and tropical swamps).
Thus, the epidermis, as a multifunctional complex tissue, performs functions only in living form.
1. Regulation of gas exchange and transpiration (between the plant and the environment)
2. Protective (mechanical, bactericidal, against excessive evaporation of water).
3. Excretory (releases salts, water, essential oils).
4. Absorption (suction) (water with a minimum of ---- foliar feeding).
5. Biosynthetic (synthesis of organic substances: cutins, waxes, etc.)
Practical Spanish
- epidermal hairs (cotton --- textile).
- essential oils --- perfumes and medicine.
With further growth of the stem, the fur supply moves to 1st place. strength, protection against water loss and sudden temperature changes, so the epidermis is replaced by a secondary integumentary tissue - the periderm.
3. Secondary integumentary tissue – periderm.
P. is a multilayer, secondary integumentary tissue.
Occurs after the epidermis. Its formation begins with the formation of the secondary meristem - phellogen (cork cambium). It is formed in annual shoots of trees and shrubs by mid-summer.
Its formation can proceed in 3 ways:
1) most often occurs in the cells of the subepidermal layer (under the epidermis)
2) sometimes phellogen is formed in the deeper layers of the bark (raspberries, currants),
3) rarely phellogen forms in the epidermis itself during the tangential division of its cells.
(the upper cell remains epidermal, and the lower one becomes a phellogen cell).
In addition to phellogen, the periderm includes 2 more types of cells, therefore it is also a complex tissue.
Types of periderm cells:
1) phellem (cork) ---protective function
2) phellogen (cork cambium) --- educational function
3) phelloderm --- performs a nutritional function in relation to phellogen.
The structure of the periderm.
1. Base – phellogen, a single-layer meristem arising from living parenchyma cells of permanent tissues, therefore - a secondary tissue.
Its cells divide tangentially and lay out many layers of cells plugs - phellems(main cells), and inside phelloderm cells.
At first, the cork cells are thin-walled, then they become suberized (at inner surface cell membranes, an insulator is deposited - suberin) and their living contents die and are filled with air (hence, such dead cells acquire good insulating properties).
In this state they perform main functions:
1) protective(mechanical, protection against moisture loss, bactericidal protection, because it is resistant to destruction);
2) thermoregulatory(cork is a thermostat, because it protects the plant well from changes in temperature, therefore, it is deposited in woody plants by the end of summer - beginning of autumn (thanks to the cork, the plant gradually freezes and gradually thaws, which prevents the formation of ice crystals in the cytoplasm of the cells, which damage and destroy the cell).
Phellogen heals wounds when the plant is damaged and when favorable conditions constantly forms new layers of cork. Therefore, the periderm is a multilayer tissue. After the development of the periderm, the epidermis is cut off from the tributary nutrients and H 2 O, dies and peels off (the green color of the stem is replaced by brownish).
The living tissues lying under the phellem plug need gas exchange, so they form in the periderm lentils – passage gaps in the cork, through which gas exchange occurs. Lentils are usually laid under the stomata. Instead of a plug, phellogen deposits living parenchyma cells here.
A cross section of a lentil shows that the cork layers alternate with parenchyma cells with well-developed intercellular spaces. Gas exchange occurs through the intercellular spaces. Phellogen underlies the parenchyma tissue, because it contains intercellular spaces and does not interfere with gas exchange.
By winter, phellogen is deposited under the parenchyma layer of cells trailing layer of dead cork-phellem cells – the lentil closes (because in winter the plant sharply reduces gas exchange and metabolic processes to almost 0).
In the spring, under the pressure of new parenchyma cells deposited by phellogen, this layer is torn and gas exchange is restored.
Lentils different types trees and shrubs have their own shape and size and change with age.
Often in trees the periderm is replaced by crust - retid– tertiary cover tissue (for example, pine, apple, ash, maple; plane tree and eucalyptus have no crust).
The crust is very powerful, thick, fissured.
Education.
The crust is formed by repeated laying of layers of phellogen, and then periderm, in the deep tissues of the cortex.
Living cells, caught between these layers, die and become part of the crust.
Thus, the cortex consists of alternating layers of cork and cells of other, dead cortical tissues, i.e. – complex in composition.
Due to the fact that the dead tissues of the plug cannot stretch when the stem grows in thickness, cracks form in the dead areas on the retidome.
1) protective (increase mechanical strength, fire resistance, etc.);
2) thermoregulatory (protects the plant from sudden changes in temperature).
MECHANICAL FABRICS
1. Definition
2. Basic types of fur. fabrics and their functions.
3. Meaning for a person.
4. Distribution of fur. tissues in the body of plants.
Mechanical fabrics
a) collenchyma - living (angular, lamellar, loose)
b) sclerenchyma – dead (fibers, sclereids)
1. Any plant needs support to maintain its integrity. In plants, the evolution of mechanical adaptations is clearly visible when adapting to life in different environments. Primary water algae that lived in dense and inert water have fur. strength is achieved thanks to cell cellulose membranes (i.e. only at the cellular level). When reaching land into a rarefied environment, this becomes insufficient and small terrestrial plants achieve stability thanks to cell membranes and cell turgor ( cellular level) . With increasing linear dimensions the strength of cell membranes and turgor becomes insufficient, and special mechanical tissues that provide support to the plant (tissue level). Subsequently, with adaptive evolution, plants adapt better and better to the air-soil environment and fur. tissues are positioned differently and more optimally in different organs(i.e., strength is ensured at the organ and organism level of organization).
That., mechanical fabrics– play the role of a support system and give strength to plants.
Main functions:
1) supporting(form the internal skeleton of plant organs)
2) protective(provide mechanical strength and resistance to mechanical influences)
They perform functions in combination with other tissues, therefore they play a role fittings and are often called reinforcing.
Mechanical (reinforcing) fabrics- specialized tissues, in combination with others, forming the reinforcement of organs and the entire body of the plant.
There are 2 main types of fur. tissues: collenchyma and sclerenchyma.
1. Collenchyma(“colla” – glue) - consists of alive prosenchymal cells with blunt and beveled ends.
Characteristic:
1. Unevenly thickened shells
2. There is no boundary between the primary and secondary shells, and they do not become lignified.
3. These are living cells that have chloroplasts.
Hence,
4. Can carry out photosynthesis (therefore, collenchyma with chloroplasts is often called
chlorenchyma and is located immediately below the epidermis).
5. Perform their functions only in a state of turgor. If its cells lose water, the plants wither.
Formed in young shoots, in growing leaves directly from apical meristems Therefore, if the collenchyma cells were dead, with secondary uniform thickenings and lignification, then they could not stretch and grow by stretching after other tissues, and therefore would not perform their functions. Therefore, collenchyma is the tissue of young plants.
Types of collenchyma.
Corner room – parts of the shells of 3-5 cells thickened at the corners merge and form 3-5-gons;
Loose k.– with intercellular spaces between fused thickened sections of membranes (in aquatic plants);
Lamellar k.– thickened parts of the shells are arranged in parallel layers.
Collenchyma is mainly developed in dicotyledonous plants.
Sclerenchyma(Greek “skleros” - hard) – a type of fur. tissue that provides strength to the organs and the entire body of the plant. It is found in all flowering plants (both monocots and dicots).
Characteristic:
1) Cells often have uniformly thickened secondary membranes, which most often become lignified (have greater crush strength, but higher fragility).
2) Dead cells. The protoplast usually dies after the shells are formed.
Those. perform the main functions dead cells.
Sclerenchyma presented fibers And sclereids.
Fibers- long prosenchymal, most often lignified cells, pointed at the ends with thick walls and a narrow cavity. Pores are few.
(exceptions, non-lignified bast fibers of a number of plants, for example, flax).
Fibers there are: 1) bast(part of the phloem phloem) (longer and thinner) and 2) wood (libriform fibers)(part of wood).
There may be other fibers that are part of other tissues and are located in groups or singly.
Sclereids– sclerenchyma cells that do not have the shape of fibers.
1. – round – stony cells – brachysclereids.
2. – branched – astrosclereids, etc.
Just as fibers form either continuous groups, layers (in nut shells, cherry pits, plums, peach, apricot), or are located singly or in groups of several cells - idioblasts(for example, idioblasts in the pulp of pear and quince fruits).
Idioblasts(Greek idios - special, blastos - embryo) - tissue cells located singly among arrays of other tissues (characteristic of mechanical and excretory tissues).
Meaning for a person.
First of all, fiber
1) Wood fibers form wood --- building materials, paper, etc.
2) Bast fibers - bast (bast shoes, shingles, etc.)






Stomata- these are openings in the epidermis through which gas exchange occurs. They are found mainly on the leaves, but are also found on the stems. Each stomata is surrounded by two guard cells, which, unlike ordinary epidermal cells, contain chloroplasts. Guard cells control the size of the stomatal opening by changing their turgidity. Appearance stomata and guard cells are clearly visible in micrographs obtained using a scanning electron microscope.
In the article we have already talked about what epidermal cells, guard cells and stomata, when viewed from above under a light microscope. The figure shows a schematic cross-section of the stomata. It can be seen that the walls of the guard cells are unevenly thickened: the wall that is closer to the opening of the stoma, called the ventral one, is thicker than the opposite one, called the dorsal one. In addition, the cellulose microfibrils in the wall are oriented in such a way that the ventral wall is less elastic than the dorsal wall. Some microfibrils form like hoops around guard cells, similar to sausages.
These hoops are not elastic, and as the cell fills water, etc. That is, the growth of its turgor, they do not allow its diameter to increase, allowing it to stretch only in length. But since the guard cells are connected at their ends, and the thin dorsal walls stretch more easily than the thick ventral ones, the cells acquire a semicircular shape. As a result, a gap appears between two adjacent guard cells, called the stomatal fissure. The same effect is observed if you inflate two oblong tubes held together at the ends. balloon, sticking adhesive tape along their touching sides (imitation of an inextensible ventral wall). To complete the picture, you can loosely wrap them with the same tape in a spiral, imitating cellulose hoops.
When guard cells lose water and turgor, the stomatal fissure closes. How the change in cell turgidity occurs is not yet clear.
According to the classical, so-called “ sugar-starch According to the hypothesis, during daylight hours the concentration of water-soluble sugars in the guard cells increases, and consequently, their osmotic potential becomes more negative, which stimulates the flow of water into them by osmosis. However, no one has yet been able to show that enough sugar accumulates in guard cells to cause the observed changes in osmotic potential.
Recently it was found that during the day in light in guard cells there is intense potassium cations accumulate and the anions accompanying them: they play the role previously assigned to sugar. It is still unclear whether their charges are balanced in this case. Some studied plants showed accumulation in the light. large quantity anions organic acids, in particular malate. At the same time, the size of starch grains that appear in the dark in the chloroplasts of guard cells decreases. The fact is that starch in the light (blue rays of the spectrum are required) turns into malate, possibly according to the following scheme:
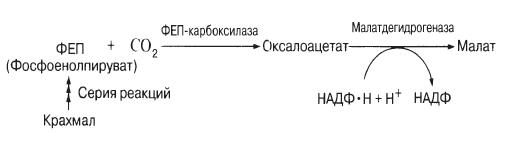
Some species, such as onions, do not have starch in their guard cells. Therefore, with open stomata malate does not accumulate, and cations appear to be absorbed along with inorganic ions such as chloride ions.
In the dark, potassium (K+) leaves the guard cells in surrounding epidermal cells. As a result, the water potential of the guard cells increases, and water from them rushes to where it is lower. The turgor of the guard cells decreases, they change shape, and the stomatal fissure closes.
Some questions for now remain unanswered. For example, why does potassium enter the guard cells? What is the role of chloroplasts other than storing starch? Perhaps potassium is ingested due to the “switching on” of ATPase localized in the plasmalemma. Some evidence suggests that this enzyme is activated by blue light. Perhaps ATPase is needed to pump protons (H+) out of the cell, and potassium cations move into the cell to balance the charge (a similar pump discussed in Section 13.8.4 operates in the phloem). Indeed, as this hypothesis suggests, the pH inside guard cells drops in the light. In 1979, it was shown that the chloroplasts of the guard cells of faba beans (Vtcia faba) do not contain Calvin cycle enzymes, and the thylakoid system is poorly developed, although chlorophyll is present there. Consequently, ordinary C3 photosynthesis does not work; starch is not formed in this way. This probably explains why starch is formed not during the day, as in ordinary photosynthetic cells, but at night.
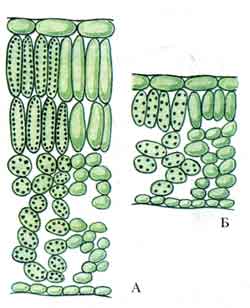
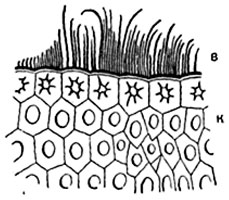
Peel or epidermis. The skin, or epidermis, is primary tissue leaves and stems. The skin consists of tightly closed cells, which in most plants have more or less sinuous outlines in plan (Fig. 86). In cross sections of the organ, skin cells usually have a quadrangular or pentagonal outline. Sometimes the outer and inner walls are slightly convex. Skin cells of elongated organs (leaf petioles, linear and lanceolate leaf blades, stems) are usually greatly elongated in a direction parallel to the longitudinal axis of the organ. In some plants, for example in many cereals, the skin consists of several types of cells (Fig. 87).
The membrane of epidermal cells usually thickens unevenly: in each cell the outer wall is thickest, the side walls are somewhat thinner, and the inner wall is relatively thin. There are exceptions to this rule. The lateral and internal walls of cells usually have pores (Fig. 87), but the outer wall is equipped with them in very large quantities. in rare cases. The cell wall consists mainly of cellulose. In many plants (especially cereals, sedges, horsetails), the outer wall of the skin is impregnated with silica; in some plants, small crystals or grains of calcium oxalate are deposited in the outer wall, less often in the lateral walls (in dracaenas) and even in the internal ones.
The surface of the skin is covered with a film - cuticle (cuticle), consisting of cutin. The cuticle is very thin or completely absent on the surface of the growth cones. Often the cuticle has a smooth surface, in other cases it forms protrusions in the form of tubercles, crossbars, straight or wavy ribs, etc. These protrusions consist entirely of
Rice. 86. Sections of the skin of a buttercup leaf ( Ranunculus repens):
1 - from the top side of the sheet; 2 - from its lower side.
cuticles or are projections of the cellulose wall covered with cuticle.
On cross sections of the skin, it is sometimes visible that the cuticle also penetrates part of the thickness of the walls. The cuticle strongly refracts light and is therefore clearly visible in sections. The isolated cuticle usually represents a colorless, structureless continuous film. Some plants (for example, the golden tree) Aucuba) the cuticle consists of plates, each of which belongs to one epidermal cell; There is no cuticle above the border areas of the skin cells or it is very thin.
The thickness of the cuticle depends on the type and age of the plant and habitat conditions (Fig. 88); on the surface of stems with perennial skin (in some types of viburnum, maple), the cuticle is torn and renewed from the inside by the formation of new layers. The cuticle of some sedge leaves undergoes silicification.
In many plants, skin cells form in the outer walls of the skin cells, sometimes in the lateral walls, and even more rarely in the inner walls. cuticular layers- interlayers in cellulose cell membrane, containing cutin (Fig. 76). The cuticle and cuticular layers are often impregnated with wax; When the preparations are heated in water under a cover glass, it melts in the form of drops. In some plants, wax covers the cuticle in the form of small grains arranged in a uniform layer (on the leaves and stems of many cereals, lilies, such as tulips), thin sticks, often curved and curled at the end (on the stems of some cereals, sugar cane, Fig. 89) , solid crusts, thin - 1μ thick (on thuja leaves, young ones).
Waxy coating, like the cuticle, reduces transpiration
![]()
1 - guard cells of stomata; 2 - side cells; 3 - epidermal cells with numerous pores; in the cell cavity there is wall protoplasm and a nucleus, in some of the cells not drawn.
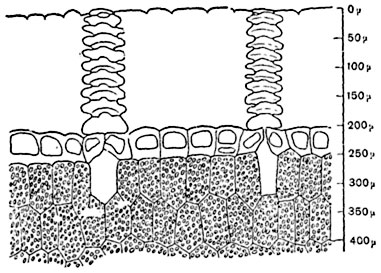
Rice. 88. Part of a cross section through a leaf-shaped tubercle of a cactus Ariocarpus retusus; the extremely thick cuticle is penetrated by figured passages above the stomata. Under the skin there is green parenchyma.
organs. The wax makes the surface of the organs non-wettable: water quickly drains from them, which prevents capillary clogging of the stomata with water and the colonization of the plant surface with small epiphytes.
To- skin: V- wax sticks.
Therefore, it is understandable why the stems and leaves of many underwater plants lack cuticles.
The skin cells contain a thin wall layer of protoplasm with a nucleus and plastids and a large central vacuole. Usually from plastids there are leukoplasts.
In many plants, the vacuoles of skin cells - often only on the underside of the leaf - contain anthocyanin.
In the vacuoles of epidermal cells, tannins and oxalic acid salts are often found in the form of solutions (in sorrel, wood sorrel) or crystals; Less commonly, there are alkaloids, toxic glucosides, gypsum (in capers - in the form of crystals), calcium salts of malic and tartaric acids (in grapes).
Chloroplasts and chromoplasts in the epidermis of flowering plants are very rare, only in some shade and aquatic plants.
Stomata. To carry out gas exchange between internal tissues plants and the external environment and for transpiration (evaporation of water) there are stomata in the skin.
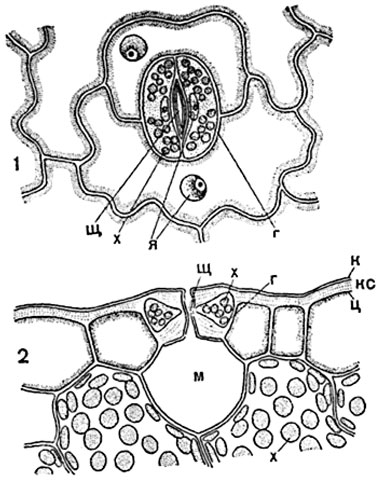
The stomata consists of two specialized guard cells and a slit-like opening between them - the stomatal fissure. When examining a piece of skin laid flat on a glass slide, the stomata are usually seen as a pair of bean-shaped or semilunar cells facing each other with their concave sides (Fig. 90-92). The membrane of each guard cell on inside strongly and unevenly thickened, on the opposite, outer side - thin.
On a cross section drawn through the middle of the stomatal fissure perpendicular to it (Fig. 91), outgrowths of the walls of the guard cells of the stoma, called spurs or beaks, are visible.
The stomatal fissure has a complex outline: the gap between the outer spurs leads into the vestibule, or front yard stomata. Next comes the narrowest part of the stomatal fissure - central slit of stomata, located between thin sections of the internal walls. The central gap widens further into the rear patio. The hole between the spurs of the lower walls of the guard cells
Rice. 90. Hyacinth leaf peel ( Hyacinthus orientalis) with stomata, in terms of:
h- guard cells, O- stomatal slit.
Rice. 91. Stomata with surrounding epidermal cells (thyme leaves Thymus):
1 - in respect of; 2 - in section; sch- central slit of the stomata; A- the thin wall of the guard cell of the stomata, facing the neighboring skin cells; in the guard cells the nucleus is visible ( I) with a nucleolus and chlorophyll grains ( x); To- cuticle; m- intercellular air cavity ("breathing cavity").
Rice. 92. Stomatal apparatus of beet leaf ( Beta vulgaris):
1, 3 - stomata in plan; 2, 4 - in section; 1, 2 - stomata is closed; 3, 4 - the stomata is open.
communicates the backyard with the respiratory cavity (Fig. 91, 2 ; 93).
Returning to the stomata in plan (Fig. 91, 1 ), we note that the stomatal slit in optical sections parallel to the surface of the epidermis has the shape of a narrow biconvex lens; The plan usually shows the outline of the gap at the level of the outer hole and at the level of the middle of the central gap. The outer walls, and in some plants partly also the inner walls, are covered with a cuticle; The edges of the courtyards sometimes consist entirely of cuticle. Functioning guard cells are living; they contain a wall layer of protoplasm with a nucleus and chlorophyll grains.
Typically, stomata are distributed singly and more or less evenly on the surface of a leaf or stem (Fig. 90). In some plants (for example, in cereals) they are located in longitudinal rows, and in others (in oleander, spleen) - in groups.
The formation of stomatal guard cells occurs when the meristematic cells of the epidermis divide unevenly, resulting in the formation of initial stomatal cells. In the simplest cases, the initial cell becomes the producing cell of the stomata. The producing cell forms, by dividing, a pair of guard cells called stomata; splitting of the septum between them leads to the formation of a stomatal fissure. In some plants, the initial cell forms, by division, entire groups of cells - a pair of guard cells and two or more subsidiary cells
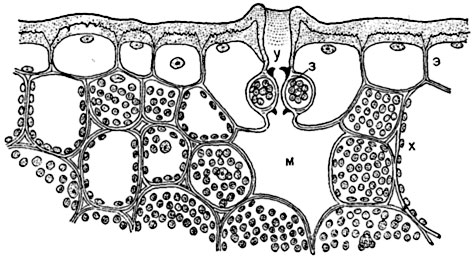
Rice. 93. Part of a cross section of a sheet Aloe disticha with a stomata immersed in the leaf tissue:
uh- skin with thick outer walls covered with a powerful cuticle; at- cavity above the stomata; h- guard cells with cuticular projections (indicated in black); m- pneumatic intercellular space (“respiratory cavity”); X- cells of chlorophyll-bearing parenchyma.
stomata; the latter differ from other skin cells by being smaller in size and having a unique outline (Fig. 94).
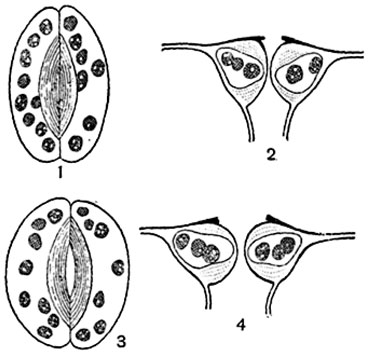
The stomatal apparatus regulates the gas exchange of the plant. The guard cells can move, change volume and shape, as a result of which the outline of the stomatal fissure also changes: it can be more or less widely open or completely closed (Fig. 92). When the stomata open and close, changes occur in the living contents of the guard cells. When the slit of the stomata is open, the protoplasm in them is more viscous than when it is closed. The core changes shape: in fodder beans, when the stomata is open for a long time, it becomes spindle-shaped, when closed, it becomes round-lobed, and in dahlias, on the contrary (Fig. 95).
The opening and closing of stomata is determined by turgor phenomena. Changes in the turgor of guard cells may be a direct consequence of leaf wilting or an increase in its water content.
As the turgor of the guard cells increases, the volume of their cell cavity increases, as a result of which the walls move away from one another and the gap opens. When the turgor of the guard cells decreases, changes of the opposite kind occur: the walls come closer to contact and the stomatal gap closes.
Changes in turgor in guard cells are regulated by the conversion of chloroplast starch into sugar, and vice versa - sugar into starch: when starch is saccharified in guard cells, the concentration of cell sap increases and their suction power increases. As a result, guard cells absorb water (partly at the expense of neighboring cells), which causes a change in their volume and opening of the stomatal fissure (Fig. 96). The conversion of guard cell sugar into starch results in similar closure of the stomatal gap.
Indirect participation in the closing and opening of stomata can take not only cells adjacent to the guard cells, but also more distant secondary cells of the stomata and ordinary cells of the skin. With prolonged rain, all skin cells are saturated with water, their turgor increases greatly and the guard cells of the stomata are compressed; as a result, the stomatal slits passively close. Some time after the rain stops, normal stomatal function is restored.
The turgor theory of the action of stomata, put forward by Mohl in 1856, was supplemented by a number of scientists pointing to possible meaning changes in the permeability of the protoplasm of guard cells.
There is a great variety in the structure of stomata. The shape of the slot can be significantly simplified or complicated compared to the typical one described above. It is most simplified in plants with leaves floating on water (in the white water lily): the gap has the form of a funnel, with a wide end connecting to the respiratory cavity, and a narrow end passing into the outer hole; the degree of opening of the stomatal slit depends on the state of this opening.
In plants with leaves located more or less horizontally, especially in trees and grasses of shady habitats, stomata are located mainly or exclusively on their lower side.
In many plants, mainly those confined to dry, sunny and windy habitats, the stomata are immersed deep in the organ due to highly cutinized projections of neighboring cells (Fig. 93). Sometimes the stomata are located in whole groups deep in the dimples (like in oleander). In some species of Australian banksias ( Banksia) there are long white hairs at the bottom and around the pits with stomata; Thus, shelters are formed with calm from the wind and shading from the sun.

A- three producing stomatal cells ( 1, 1, 1 ) have already been separated by septa from the initial cells; nuclear division occurs in neighboring cells ( 2, 2 ), preceding the formation of a side cell; B- for each of the producing cells of the three stomata, two secondary cells were formed ( 3 ); IN- in the three producing cells of the stomata, nuclear division occurs ( 4, 4 ), preceding the formation of guard cells; G- the producing cell is divided into two cells that differentiate into a pair of stomatal guard cells ( 5, 5 ), with a stomatal gap between them; 6, 6 - side cells; D- formed stomatal apparatus, consisting of two guard cells ( 5, 5 ) and two side cells ( 6, 6 ).
Hairs (trichomes)). In most higher plants, some or many of the skin cells form outgrowths, the so-called hairs, which have different shape, papillae, tubercles, bristles, bushes, stellate scales, etc. In the simplest case, a hair represents a papilla - a short round-conical outgrowth of a skin cell. The papillae

A- the stomata is open, the guard cells contain very small starch grains, the nucleus is rounded-amoeboid in shape, with vacuoles in the center; b- the stomata is closed, in the guard cells there are relatively large grains of starch, cell nucleus spindle-shaped, with large nucleoli in the center.
are hairs of almost all types in early stage their education. There are hairs that are not separated by a septum from the skin cell that produced them, such as the bristles of many borage. In most cases, the hair is separated by a septum from the epidermal cell that produced it - the “mother cell of the hair.” The detached hair rarely remains single-celled; more often it becomes multicellular. Multicellular hairs are very diverse. Among them there are 1) linear, consisting of cells arranged in a row (potato leaves); 2) bushy-branched (mullein, Fig. 97, 6 ); 3) scaly and stellate-squamous ( Elaeagnus, rice. 97, 7, 8 ) and 4) massive, representing, as it were, a bunch of fused thread-like hairs (the hairs of many Lamiaceae). The contents of the hairs of some categories are destroyed in the adult stage, and their cavities are usually filled with air. Such hairs, usually white, or grayish, or yellowish color, can be considered one of the devices that moderates the effect of the hot rays of the sun on a leaf or stem and weakens the drying effect of the winds.
Some hairs can serve as protection for the plant from being eaten by animals; Thus, closely spaced hard, sharp bristles and a felt cover protect leaves and stems from being eaten by slugs and partly from being eaten by herbivorous mammals.
Burning hairs. Stinging nettle hair - large living cell with a vesicular base sitting in a cup-shaped multicellular “stand”. The hair represents in miniature a hollow syringe needle and ends with an obliquely set head. Since the shell of the hair, rich in lime and silica, is very brittle and, moreover, at the point of transition of the “needle” into the “head” is very thin, then at the slightest touch of an animal or person to the head, the latter breaks off, the sharp edge of the “needle” pierces the skin, And cell sap a hair containing a burning substance is injected into it (Fig. 97, 9-12 ).
Touching the leaves of some nettles of tropical flora (for example, Urtica urentissima Indonesia) causes painful or even life-threatening inflammation.
The integumentary tissues also include the periderm and the crust (for a description of them, see pp. 173-175).
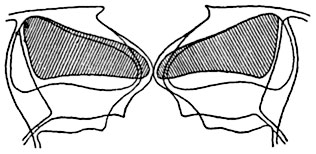
Rice. 96. Schematic optical section of the stomata of the winter plant ( Heieborus sp..) in closed and open state.
The external outlines and cell cavity of the guard cells are shown with the stomatal slit closed and with the slit open. The cell cavity of both guard cells is shaded when the stomata is closed.