VAKUOLER AF PLANTECELLER
Organoider generel betydning, der har et enkeltmembranstrukturprincip og forekommer kun i planteceller.
Størrelse og antal: bestemmes af cellens alder. I unge celler opstår vakuoler fra små vesikler, der brækker af fra EPS. Efterhånden som celler vokser og differentierer, smelter små vakuoler sammen og danner en eller flere store vakuoler, der optager op til 80 % af hele cellens volumen. Som et resultat skubbes cytoplasmaet med kernen og organellerne til periferien.
Struktur: centrale vakuoler er adskilt fra cytoplasmaet af en enkelt membran - tonoplast, som i tykkelse ligner plasmalemmaet. Vakuolhulrummet fyldes cellesaft. Cellesaft indeholder: uorganiske salte, sukkerarter, organiske syrer og deres salte, andre lavmolekylære stoffer, såvel som nogle højmolekylære forbindelser (f.eks. proteiner).
Funktioner: 1) osmoregulering: på grund af tonoplastens og plasmalemmas semipermeabilitet opretholdes den tilsvarende molekylære koncentration af cellesaft, dvs. vakuolen fungerer som et osmometer;
2) udskillelsesorganer: alle vandopløselige stofskifteprodukter (alkaloider - nikotin, koffein; polyfenoler) kan fjernes gennem tonoplasten;
3) opbevaring: fosfater K + , Na + , Ca 2+ , salte ophobes i cellesaften organiske syrer(oxalater, citrater osv.), sukkerarter og proteiner.
RIBOSOMER
En organel, der ikke har en membranstruktur. Det er den eneste organel af generel betydning, der er til stede i cellerne i både prokaryoter og eukaryoter. Ribosomer blev først beskrevet i 1955 G. J. Palade (Palade granulat), som beviste, at de er ribonukleoproteinkomplekser (RNP).
Kemisk karakter: RNP= rRNA+protein.
Ribosomer står for 85% RNA, præsenteret i en celle.
Form: Ribosomet har en svampeform, fordi det består af to underenheder: stor Og lille, mellem dem er placeret funktionelle centrum af ribosomet (FCR) , hvori under proteinbiosyntesen (oversættelsesperioden) er mRNA placeret i dets to tripletter, og et enzymatisk kompleks fungerer, hvilket sikrer samlingen af et proteinmolekyle fra aminosyrer.
Dimensioner: 15 – 35 nm. Størrelsen af et komplet ribosom i prokaryote celler er 20x17x17 nm, i eukaryote celler - 25x20x20 nm.
Uddannelsessted: dannelsen af ribosomale underenheder sker i nukleolerne i kernen. Samlingen af underenheder til et komplet ribosom sker i cytoplasmaet, når koncentrationen af magnesiumioner (Mg 2+) når 0,001 M; hvis denne koncentration falder, opstår dissociation af underenhederne. Når Mg 2+ koncentrationen stiger ti gange og når en værdi på 0,01 M, interagerer de to ribosomer med hinanden og danner en dimer.
FOREDRAG: Træk af en plantecelle
Den samlede kemiske sammensætning er næsten den samme.
Ligheder med dyreceller: tilstedeværelsen af en membran, cytosol, kerne og specifikke organeller.
Men mellem disse typer eukaryoter er der 2 forskelle i struktur: tilstedeværelsen af specifikke organeller og metabolisme.
I en plantecelle er der central vakuole. Vakuoler planteceller absorbere overskydende vand, svulme op, hvilket fører til cellestrækning, vakuolen optager det meste af plantecellens volumen, og derfor forbliver kun tynde tråde af cytoplasma i plantecellen, der passerer gennem hele cellen. Dette tjener som en økonomisk måde for en plantecelle at øge sin størrelse. I dette tilfælde forbruges mindre energi end ved fyldning af samme volumen med proteinrig cytoplasma.
Unge planteceller kan have flere vakuoler, som, efterhånden som cellen vokser, smelter sammen og danner en eller flere store vakuoler, som fylder op til 90 % af hele cellens volumen. I dette tilfælde skubbes organellerne til periferien af cellen, og også kernen. Vakuolens membran er tonoplasten. Vacuolens hulrum er fyldt med cellesaft, dvs vandopløsning uorganiske salte, sukkerarter, organiske syrer og deres salte, proteiner, lavmolekylære forbindelser.
Centralvakuolens hovedfunktion– opretholdelse af turgortryk. Det bestemmes af molekyler opløst i vakuolens cellesaft, som opretholder osmotisk koncentration. Koncentrationen sikrer, at vakuolen fungerer som en struktur, der fastholder cellens indre tryk og giver den styrke og spænding. Tonoplasten har en ATP-afhængig protonpumpe, der sikrer transporten af sukkerarter, og der er også ionkanaler, der sørger for udskillelse (fjernelse) fra vakuolen af forskellige stofskifte. Protonkoncentrationen i cellesaft svarer til et surt miljø på 2-5pH. Den centrale vakuole sikrer ophobning af reserve næringsstoffer– sukkerarter og proteiner. Tilførslen af disse stoffer er forbundet med membranernes evne til at interagere med tonoplasten. Fordøjelsesprocesser kan også foregå inde i cellesaftvakuolen, pga indeholder hydrolytiske enzymer, mens tonoplasten danner sekretoriske vesikler, de spaltes fra vakuolen, interagerer med eksocetotiske vesikler, hvilket sikrer fordøjelsen af komponenterne indeholdt i dem. De udfører også reaktionerne i den autofatiske cyklus og deltager i hydrolysen af defekt fyldning.
Plantecellevæg.
Dannet med deltagelse af plasmalemmaet. Det er en ekstracellulær flerlagsformation, der beskytter cellens overflade, i det væsentlige dens ydre skelet. Den består af to strukturer: en ramme (lavet af cellulosefibriltråde) og en gel-lignende matrix. Matrixen indeholder polysaccharider: hemicelluloser og pektinstoffer. Hemicelluloser er forgrenede polymerkæder bestående af forskellige hexoser (glucose, manose, galactose), der kan også være pentoser (xylose, arabinose) og uronsyrer (glucuron og galacturonsyre). Disse komponenter af hemicelluloser kombineres med hinanden i forskellige kvantitative forhold og danner forskellige kombinationer. Men kæderne af hemicellulosemolekyler krystalliserer aldrig, og deres fibriller er ikke fikserede (ikke opdaget). Indeholder store mængder vand.
Pektiner. En heterogen gruppe, der omfatter forgrenede, stærkt hydrogenerede polymerer, der bærer negative ladninger på grund af tilstedeværelsen af mange galacturonsyrerester.
Takket være disse komponenter er matrixen en blød plastmasse, der tjener som base (ramme).
Individuelle cellulosepolymerer pakkes i mikrofibriller ved hjælp af hydrogenbindinger. Giver stivhed og styrke. Mellem dem er der ledbånd, der gør cellevæggen til en monolit. Sådanne bundne cellulosemikrofibriller er omgivet af pektiner. De kan interagere med calcium og silicium. Hvad giver den stivhed. På grund af den gel-lignende struktur giver matrixen diffus transmission af vand og små molekyler.
Cellevægge er opdelt i 3 typer: primær, sekundær, tertiær.
Den primære celle består af 90% kulhydrater. Under celledeling opstår et bundt af mikrotubuli i cellernes ækvatorialplan, placeret mellem de divergerende kromosomer (phragmoplast). Blandt mikrotubulierne er der mange små vesikler i det vakuolære system, CG, i den centrale del begynder phragmoplasterne at smelte sammen med hinanden. Danner en fladtrykt skive eller midterplade. Består af et polysaccharid kaldet callose. Mere elastisk sammenlignet med cellulose (ikke indeholdt i CG-bobler, dannet på plasmalemmaet). CG-vesiklerne indeholder de komponenter, der er nødvendige for at bygge membranerne af to datterceller. Processen med fusion af små vakuoler sker fra midten af cellen til periferien og fortsætter, indtil membranvesiklerne smelter sammen med overfladen af cellens laterale overflade.
Den voksende primære cellevæg består allerede af 3 lag (den midterste plade, bestående af en amorf matrix, og 2 perifere, indeholdende hemicellulose og fibriller). Den primære væg er dannet ved sekretion af hemicellulose og cellulosefibriller af to nye cellulære strukturer. Forøgelse af tykkelsen mellem cellevæg vil opstå på grund af aktiviteten af datterceller, som hver på sin side vil udskille stoffer fra cellemembranen, som fortykkes ved at lægge flere og flere nye lag. Ved plasmamembranen syntetiseres og polymeriseres cellulosefibriller, hvilket resulterer i dannelsen af en sekundær cellevæg (der giver cellen dens endelige form). Så snart fortykkelsen af væggen er afsluttet, modificeres den med lignin, en hydrofob polymer, syntetiseret gennem oxidativ polymerisering af op til tre rester af aromatiske alkoholer og fungerer som hovedbestanddelen af træ (20-35% af træet er lignin) ). På grund af sin hydrofobe natur skaber lignin vandtætning i vægge og fungerer som et ekstra forstærkningsmateriale. Cellevæggen bliver lignificeret eller suberiseret (på grund af cutin og suberin). I epidermale celler udskilles voks på overfladen af cellemembranerne.
Svampe har kitin (N-acetylglucosamin).
Plastider.
Fælles egenskab for alle plastider:
De er omgivet af to membraner, der tæt lukker langs hele overfladen af organellerne.
Det indre indhold er stroma. Den indeholder membranskiver dannet som følge af invagination og knopskydning af dele af den indre membran. Disse skiver kaldes thylakoider og har evnen til at foldes til stakke (granas). De interagerer med den fjerde type membran - lamellen.
Plastider har deres eget genom, hvor hvert plastid har flere kopier af cirkulært DNA, der indeholder omkring 100 gener. Genomet koder for proteiner, der er nødvendige for at udføre specialiserede funktioner. RNA-proteiner involveret i transkription og translation er også placeret her. De stammer fra plastider, men de fleste andre proteiner afhænger af kernegenomet. Disse proteiner syntetiseres i cytoplasmaet og skal importeres til plastidet.
Plastider er ikke forbundet med andre organeller ved vesikulære bindinger.
Plastider repræsenterer en række gensidige transformationer.
Proplastid -> leukoplast -> kloroplast -> kromoplast.
Leukoplaster kan omdannes til: amyloplaster, elaioplaster, aleuronkorn og kromoplaster.
Kloroplast.
De har en aflang form, størrelse 2-4 mikron, og længden kan være 10 mikron. Kromatoforer (store kloroplaster).
Kloroplastens indre rum er lumen. 2 vigtige fænomener: fotosyntese, energisyntese. Thylakoidmembranen er karakteriseret ved tilstedeværelsen af klorofyl; den er i stand til at absorbere mængder af sollys og omdanne solenergi til kemisk energi. Absorption af lys af en bestemt bølgelængde forårsager ændringer i klorofyl. Han går i en ophidset tilstand. Og den frigivne energi af aktiveret klorofyl overføres gennem en række mellemstadier til elektrontransportkæden, hvilket fører til syntesen af ATP og genoprettelse af bæreren, som er 2 mononukleotider (NAD og FAD). Der bruges energi på syntese af kuldioxid og syntese af sukkerarter.
Fotosyntese: lys og mørk fase.
Den lette fase forekommer kun i lyset og er forbundet med absorptionen af lys af pigmentet og udførelsen af en kemisk reaktion - Hill-reaktionen.
I den mørke fase fikseres og reduceres kuldioxid (fra atmosfæren), hvilket fører til syntese af kulstof, AMK og andre stoffer. Som et resultat af den lette fase opstår fosforyleringsprocessen, syntesen af ADP og ATP. Samt restaurering af bæreren, der sker under hydrolyse og fotolyse af vand.
I lysfasen af fotosyntesen exciterer sollysets energi elektronerne i klorofyl. De transporteres langs komponenter i den oxidative kæde i thylakoidmembranen, svarende til hvordan elektroner transporteres langs respirationskæden i mitokondrier. I kloroplaster pumpes protoner fra stroma (pH = 8) til lumen (pH = 5), hvilket skaber en gradient på 3 pH-enheder. Denne protongradient skaber en protonmotorkraft på 200 mW ved protonmembranen, men dette skyldes næsten udelukkende pH-gradienten. Sammenlignet med den mitokondrielle respirationskæde bevæger elektroner sig i en anden retning. Elektroner tages fra vand under dets fotolyse og overføres til bæreren med energiforbrug. At. i den lette fase har vi: syntetiserede ATP-molekyler og en reduceret transportør. Begge produkter bruges i den mørke fase.
I det mørke stadie dannes kulhydrater på grund af reduceret NAD og ATP. Denne proces er flertrins. Et stort antal enzymer er involveret i det. Calvin cyklus. En stigning i antallet af kloroplaster og dannelsen af andre former for plastider sker fra forstadiestrukturer (proplastider).
Proplastider er små dobbeltmembran-vesikler uden særpræg. De adskiller sig fra cytoplasmatiske vakuoler i deres tættere indhold og tilstedeværelsen af to membraner. Proplastider er placeret i dele plantevæv. Antallet af proplastider stiger ved deling og knopskydning. Under normal belysning bliver proplastider til kloroplaster (stigning i størrelse, dannelsen af langsgående membranfolder forekommer. Nogle danner lameller, andre - thylakoider). Hvis de udvikler sig i mørke, er der først en stigning i volumenet af plastider, men der dannes ikke et system af indre membraner; der dannes en masse små vesikler, som akkumuleres i separate zoner og danner komplekse gitterstrukturer - prolamellære kroppe (etioplaster). Deres membraner indeholder protochlorophyll (gul), under påvirkning af lys dannes kloroplaster fra dem, og protochlorophyll kan blive til ægte klorofyl (membraner, komponenter i overførselskæden syntetiseres).
Leukoplaster. Forskellig fra kloroplaster. De findes i lagervævsceller. Deres usikre morfologi gør dem vanskelige at skelne fra proplastider. De er synlige som lameller, men er i stand til at danne normale thylakoidstrukturer. I mørke ophober de forskellige reservestoffer. Hvis det er et kulhydrat, så dannes leukoplaster til amyloplaster (der kan også være proteiner i form af aleuronkorn). Og hvis fedtstoffer er elaioplaster.
Vi observerer processen med misfarvning og ændringer i kloroplaster under udviklingen af kronblade og modning af frugter. Yderligere membraner ophobes i plastider, klorofyl og stivelse ødelægges. Når lamellerne ødelægges, frigives lipiddråber, hvori pigmenter (carotenoider) opløses godt, og en farveændring opstår. Kromoplaster er en degenererende form for plastider, som er forbundet med nedbrydning af lipoproteinkomplekser.
Plastid genom.
Dets eget genetiske system sikrer syntesen af proteiner, der er lokaliseret inde i selve plastiderne (thylakoidmembranproteiner). De har en længde på op til 60 mikron. 1,3*10 8 Ja. Cyklusvarigheden og replikationshastigheden stemmer ikke overens. DNA har ingen fikserende strukturer. Alle karakteristika ved ringmolekylet af kloroplaster er tæt på prokaryoternes. Ligheden mellem chloroplast-DNA manifesteres i det faktum, at de grundlæggende regulatoriske processer for transkription er de samme. Syntese af alle typer RNA. Kloroplast-DNA koder for ribosomalt RNA. 70S ribosomsegmenteringskonstanter. De indeholder 17S og 23S. Følsom over for antibiotika (chloramphenicol).
FOREDRAG: Intercellulære kontakter
Symbiotisk teori.
Komintsin og Mereshkovsky.
Denne teori understøttes af den fantastiske lighed i strukturen af kloroplaster og blågrønalger. Lighed med deres hovedfunktioner, næsten identiske evner til fotosyntetiske processer. Som et resultat af symbiosen mellem forskellige prokaryote celler opstod nukleare (eukaryote) celler. Ifølge denne hypotese gik en eukaryot celle i sin udvikling igennem flere stadier af symbiose med andre celler.
I den første fase blev celler såsom anaerobe heterotrofe prokaryoter værter for aerobe bakterier, hvilket førte til dannelsen af aerobe prokaryoter (mitokondrier, i disse prokaryote organismer). Parallelt med denne proces, i værtscellen, dannes det prokaryote kerneapparat (genophor) til en kerne isoleret fra cytoplasmaet. Sådan kunne heterotrofe eukaryote celler opstå. En sådan celle indgår i gentagen symbiose. For eksempel med spiroketter eller flagellignende bakterier. Inddragelsen af disse symbionter førte til fremkomsten af flageller og cilia i heterotrofe eukaryote celler. Og som et resultat af yderligere absorption. Dette førte til fremkomsten af eukaryote dyreceller med flageller. En sådan organisme kunne også optage blågrønalger og det førte til dannelsen af plastider og dannelsen af en plantecelle. Hypotesen om den endosymbiotiske oprindelse af intramembrane organeller er uden tvivl. Talrige fakta om ægte endosymbiose af blågrønne alger og celler er kendt lavere planter og protozoer, hvor symbionterne fungerer perfekt og forsyner værtscellen med fotosynteseprodukter. For eksempel. En hjemmesko ciliat, den indeholder algen chlorella. Kloroplaster kan udvælges af nogle celler og bruges som endosymbionter. Rotifer lever af alger. Kloroplaster befinder sig inde i cellerne i fordøjelseskirtlerne og fortsætter med at udføre deres funktioner.
Funktioner af plantecellernes funktioner. Metoden til syntese af organiske stoffer er autotrofisk ernæring.
Planteceller, på grund af tilstedeværelsen af en cellevæg, tillader ikke plantecellen at ændre sin form og bevæge sig. Og planteceller kompenserer for den manglende bevægelse ved at syntetisere deres egen mad, dvs. er autotrofer. Planter vokser hele deres liv, meristem deler sig konstant og danner nye celler; alle planter tilhører modulære organismer. Deres endelige livsform afhænger af deres miljø. Planteceller har også karakteristika i mitoseforløbet:
Placering af den nye cellevæg. Det bestemmes længe før dannelsen af spindlen, mens spindlen selv ikke deltager i placeringen af denne celle. Allerede før mitosen, i G 2-fasen af cellecyklussen, dannes en specifik struktur i plantecellen, som kaldes præprofaseringen. Det er dannet af mikrotubuli i cortex (cytoskelettet) i form af en bred strimmel, der omgiver cellen. Kernen forbliver intakt (fast). I området af denne ring samles aktinfilamenter, som sikrer dens indsnævring, indtil den bliver til et velformet tæt bundt af mikrotubuli rundt om kernen. Den resulterende struktur ligner et hjul, hvor fælgen og egerne er sammensat af mikrotubuli og aktin, og kernen danner navet. Ringens struktur er beriget med EPR- og KG-elementer. På det tidlige stadium af mitose depolymeriseres og ødelægges mikrotubulierne i præprofaseringen. Og i telofase dannes en ny cellevæg præcis kl det fly, som blev udpeget af preprophase-ringen.
Kernen i en plantecelle bevæger sig til midten af cellen, hvor deling vil ske. Smalle lag (strenge) af cytoplasma strækker sig fra kernen i alle retninger. Efterhånden som mitosen opstår, smelter lagene sammen med hinanden og danner et plan, hvor præprofaseringen plejede at være, og dette plan kaldes fragmosomet.
Hos planter indeholder polerne på den mitotiske spindel ikke centrioler og er mere diffuse i naturen. De ser med andre ord ud til at være slørede. Indledningsvis dannes en profasespindel af mikrotubuli. Fra den, efter ødelæggelsen af kernemembranen, dannes en rigtig spindel, og med dannelsen af denne ægte mitotiske spindel forbliver ingen andre cytoplasmatiske mikrotubuli i cellen. Hvad styrer denne spindel, hvad dannes? Kromosomer. For det første sker polymerisering af mikrotubuli nær kromosomerne, som er orienteret tilfældigt. Derefter sorterer motorproteiner (dynein) mikrotubulierne, deres positive ender sendes til kromosomerne, hvilket giver dem mulighed for at binde sig til kromosomernes kinetochorer, og andre interagerer med de negative ender af mikrotubulierne. Som et resultat af polymerisation, sortering og fastgørelse dannes en bipolær spindel. Efter nuklear deling sker cytoplasmatisk deling. Dette falder sammen med tidspunktet for dannelsen af en ny celleseptum, hvis position er bestemt af præprofaseringen. Forholdet mellem spindelorientering og nyt septum er ikke tæt, som i dyreceller.
Golgi kompleks af planteceller. Funktioner: glykosylering (primær), sortering (separation), syntese af cellevægspolysaccharider (undtagen cellulose og callose), levering af enzymkomplekser (hexagonale rosetter, cellulosesyntese) til membranen, rettet transport af vesikler ind i vakuoler.
KG er lokaliseret i planteceller tættere på plasmaet, længere fra kernen, og er spredt i form af talrige stakke af tanke. Alle elementer i komplekset er forbundet af aktinfilamenter, som danner et netværk, der sikrer intracellulær transport. Aktinstrukturer giver mobilitet (sammen med myosin). Planteceller har et særligt myosin - myosin 8. Dette myosin 8 danner ikke filamenter og akkumuleres i nydannede primære vægge, CG-vesikler indeholder polysaccharider og strukturelle proteiner, der danner den sekundære cellevæg.
Intercellulære kontakter.
Membranernes kontaktfunktion sikrer kontakt af celler med hinanden og med andre. Alle celler i væv er i kontakt med den ekstracellulære matrix. Den ekstracellulære matrix er involveret i at opretholde vævsintegritet og danner en ordnet ramme, inden for hvilken celler bevæger sig og interagerer med hinanden. Interaktionen mellem celler udføres på grund af kontaktområder, som kaldes intercellulære kontakter.
Der er flere klassifikationer af dem, de er struktureret forskelligt. Alberts og Gray (1986) klassifikation. Ifølge det: kontakter kan opdeles i 3 grupper.
Mekanisk (= klæbemiddel). Forbundet med cellers evne til at klæbe til hinanden på grund af tilstedeværelsen af ekstracellulære matrix-glycoproteiner i cellemembranen. Når der sker adhæsion mellem plasmamembraner, forbliver der altid et mellemrum (20 nm) fyldt med glycocalyx. Specielle CAM-proteiner (cam) er direkte ansvarlige for at forbinde celler med hinanden: fibronectin, cadheriner, selectiner, integriner og andre proteiner. Nogle af dem forbinder celler med hinanden gennem intermolekylære interaktioner. Andre danner særlige intercellulære forbindelser. Interaktionerne mellem disse proteiner kan være homofile, dvs. naboceller i dette tilfælde kommunikerer med hinanden ved hjælp af homogene molekyler. Eller heterofile, når de deltager i adhæsion forskellige slags proteiner på naboceller. Der er flere komplekse forbindelser, når intercellulær binding er sikret af linker-mellemmolekyler. Ud over sådanne enkle forbindelser Der er en række specielle strukturer, der udfører specifikke funktioner (desmosomer). De interagerer med elementer i cytoskelettet. Der er 3 typer desmosomer: punktformede, omkransende og hemidesmosomer.
Stiplede forbinder celler med hinanden i form af knapper. Afstanden mellem membranerne af kontaktceller varierer fra 22 til 35 nm. Mellem cellerne, på grund af modifikationer af supramembrankomplekset, dannes en fibrøs matrix, og i dens centrale del er der en "plade" bestående af proteinkugler. De er repræsenteret af interagerende integrerede membranproteiner cadderiner og desmogleins. Denne plade interagerer med cellemembraner gennem et system af tværgående snore. På siden af cytoplasmaet dannes en anden plade af proteinet desmoplakin, hvorfra fibrillære strukturer relateret til 10 nm filamenter (keratin filamenter) strækker sig i den tværgående retning ind i dybet. Derfor findes punktformede desmosomer i epitelet, hjertemusklerne og endotelkar.
Bæltedesmosomer. Afstanden mellem naboceller er 15-20 nm. Centralpladen er ikke så lys. Men der er trans-membran glycoproteiner, der klæber til hinanden og giver mekanisk forbindelse af membraner. Tynde actinfilamenter (6-7 nm) akkumuleres på den cytoplasmatiske side. De ligger langs plasmalemmaet og løber langs hele cellens omkreds. Funktion: mekanisk adhæsion af celler til hinanden; når aktinfilamenter trækker sig sammen, kan cellens form ændre sig; kan forårsage ændringer i cellegeometri; overføre mekanisk belastning til cellen. Forårsaget af en ændring i cellevolumen.
Hemidesmosomer. Forbindelse af celler med den ekstracellulære matrix (For eksempel med basalmembranen). Den funktionelle rolle er mekanisk, hvilket gør det muligt for epitellagene at modstå store mekaniske påvirkninger. Desmosomer er dele af celler, men de er ikke statiske formationer. De er dynamiske. I stand til at forsvinde og dukke op igen. De ødelægges af en fagocentral reaktion.
Lukning (tætte kontakter). De er karakteristiske for enkeltlags epitel. Den zone, hvor de ydre lag af de to plasmamembraner er så tæt som muligt. Derfor, når det ses gennem et elektronmikroskop, er en tre-lags struktur synlig. De 2 ydre lag af begge membraner smelter sammen til et 2-3 nm tykt. Ikke over hele området med tæt kontakt, men repræsenterer en række punktkontakter af integrerede membranproteiner. Mere end 24 typer af proteiner blev fundet i tight junction zone. Sådanne proteiner stikker ud fra membranerne i det bilipide lag, krydser hinanden og danner et netværk (gitter). På siden af den tætte forbindelse er der i selve cytoplasmaet adskillige fibriller (7 nm), som er placeret parallelt med overfladen af plasmalemmaet. Tight junctions er karakteristiske for kirtel- og tarmepitel. De forbinder celler med hinanden. Desuden dette molekylære strukturer regulering af paracellulær transport, dvs. transport af stoffer i rummet mellem celler. Fysiske barrierer for iontransport har en anden karakter. Ioner passerer øjeblikkeligt, men andre komponenter tager længere tid (selv timer) at passere gennem den tætte kontakt.
Permeabilitetsbarrieren i en tæt kontakt er skabt af ladningsbærende lag, der danner en netværksstruktur af fibrillære trådlignende elementer. Ioner kan passere gennem sådanne porer; deres størrelse er lille. For passage af opløselige komponenter skal integriteten af disse tråde imidlertid forstyrres. Opløselige grundstoffer bryder disse tråde, så genforenes de efter passage af molekyler. Og så bevæger molekylet sig gennem kontaktbarrieren. Strukturelt og funktionelt opdeler tight junctions plasmalemmaet i 2 sektioner (apikale og basale), og tight junction fungerer som en septum, på grund af hvilken den molekylære sammensætning holdes inden for grænserne.
Ledende kontakter. Det adskiller sig i dyr og planter.
Der er 2 typer dyreceller: gap og kemisk syntese. Gennem gap junctions kan molekyler bevæge sig fra en celle til en anden. Men i kemisk syntese har celler ikke en direkte forbindelse. Mellemrumsforbindelser anses for at være cellers kommunikationsforbindelser. Dette er en struktur, der er involveret i den direkte overførsel af kemikalier. En karakteristisk kontakt af denne type er sammenføringen af plasmamembranerne fra to naboceller til en afstand på 2-3 nm. Selve spaltekontakten har en størrelse på 0,5-5 µm. Den består af 2 halvdele, størrelsen af hver halvdel er 7-8 nm, de er placeret omkring en kanal 1-2 nm bred. Og disse 2 halvdele kaldes connexons (halvkanaler). De består af 6 connectin-proteinunderenheder. Proteinets molekylvægt er omkring 30 tusind. Ved at kombinere med hinanden danner proteinerne en cylindrisk struktur, og i to celler er størrelsen, antallet, størrelsen og arrangementet af connexons strengt symmetriske. Mellemrumskontakter tillader fri diffusion af molekyler, der vejer 1200 Dalton, undtagen molekyler, hvis masse når 2000 Dalton. Permeabiliteten af spalteforbindelser styres af åbning og lukning af kanalportene. Denne proces kaldes portmekanisme (portmekanisme). Det styres af ændringer i intracellulær pH, calciumion flux eller direkte phosphorylering af connexinproteinunderenheder. Funktioner af spaltekontakter: at udføre organisk og uorganiske stoffer. (For eksempel i myokardiemuskelceller).
Plasmodesmata er en type ledende kontakter i planter. Disse er tynde rørformede cytoplasmatiske kanaler, der forbinder 2 naboceller. Diameteren af disse kanaler varierer fra 20 til 40 nm. Funktioner af disse kanaler: plasma membran fra en celle går over i en anden celles plasmalemma. Langs aksen af denne kanal strækker et cylindrisk rør af desmotulen sig fra en celle til en anden. Dens lumen kommunikerer med ER-cellerne. Rummet mellem desmotubule og plasmodesmata-membranen er fyldt med cytosol. Plasmalemmaet, der grænser op til disse celler, passerer jævnt ind i plasmodesmata og andre cellers membran. Som et resultat er hyaloplasmaet af naboceller forbundet til et enkelt system (symplast). Og formelt er der ingen adskillelse (en celle fra en anden). Fri transport gennem plasmodesmata er begrænset til en molekylvægt på 800 Dalton. Plasmodesmata dannes på stadiet af den primære dannelse af medianpladen fra elementerne i ER. I en nyopdelt celle når antallet af plasmodesmata op til 1000 pr. celle. Med aldring falder deres antal, men deres tykkelse øges. Funktionel rolle: at sikre intercellulær cirkulation af opløsninger (ioner, sukkerarter, næringsstoffer). Den negative side er, at det er lettere at blive syg.
Vakuoler og cellesaft
De fleste modne planteceller er karakteriseret ved en stor central vakuole, der optager op til 70-90% af cellevolumenet. I dette tilfælde er protoplasten med alle organellerne placeret i form af et meget tyndt væglag, der forer cellevæggen. Små cytoplasmatiske vakuoler findes normalt i vægprotoplasten. Nogle gange er kernen placeret i midten af cellen i atomlomme cytoplasma, som er forbundet med væglaget ved at de tyndeste cytoplasmatiske tråde krydser den centrale vakuole.
Cellesaft er en vandig opløsning forskellige stoffer, som er produkter af protoplastens vitale aktivitet, hovedsageligt reservestoffer og affald. Reaktionen af cellesaft er normalt let sur eller neutral, sjældnere basisk. De stoffer, der udgør cellesaften, er ekstremt forskellige. Disse er kulhydrater, proteiner, organiske syrer og deres salte, aminosyrer, mineralioner, alkaloider, glycosider, tanniner, pigmenter og andre vandopløselige forbindelser. De fleste af dem tilhører gruppen af ergastiske stoffer - produkter af protoplastmetabolisme, som kan opstå og forsvinde i forskellige perioder celleliv. Mange cellesaft-stoffer dannes kun i planteceller.
Kulhydrater i plantecellesaft er repræsenteret af monosaccharider (glucose, fructose), disaccharider (saccharose) og polysaccharider (slim, inulin).
Glucose (druesukker) og fructose (frugtsukker) ophobes i store mængder i saftige frugter. Saccharose (roesukker) ophobes i store mængder i sukkerroerødder og sukkerrørsstilke. En række plantefamilier (cactaceae, Crassulaceae, orkideer) er karakteriseret ved ophobning af slim i cellesaften, som holder på vandet. Inulin er et reservepolysaccharid, aflejret som en kolloid opløsning i cellesaften af de underjordiske organer af Asteraceae i stedet for stivelse.
Proteiner akkumuleres i form af en kolloid opløsning i vakuolerne af celler af modne frø. Når frø er dehydreret i de senere stadier af deres udvikling, fjernes vand fra vakuolerne, proteinkoncentrationen i cellesaften stiger, og det bliver til en fast geltilstand. Dehydrerede vakuoler af modne frø kaldes aleurone korn.
Af de organiske syrer i cellesaft er de mest almindelige citronsyre, æblesyre, ravsyre og oxalsyre. Disse syrer findes i store mængder i cellesaften af umodne frugter, hvilket giver dem en sur smag. Når frugter modnes, kan organiske syrer bruges som respirationssubstrater, så frugtens syrlige smag forsvinder som regel. Salte af organiske syrer spiller sammen med mineralioner en vigtig rolle i osmotiske processer.
Tanniner (tanniner) er polymere phenolforbindelser med en astringerende smag. De har antiseptiske egenskaber og beskytter plantevæv mod infektioner og forfald. Cellerne i barken af stængler og rødder (eg, pil), umodne frugter (valnødder), blade (te) og nogle patologiske vækster - galle er særligt rige på tanniner. Tannider bruges i medicin, til garvning af læder og farvning af stoffer mørkebrune.
Alkaloider – forskellige i kemisk nitrogenholdige organiske stoffer, der har en bitter smag. De har egenskaber som baser og findes i cellesaft, normalt i form af salte. Mange alkaloid-bærende planter er giftige og spises ikke af planteædere. I celler, der indeholder alkaloider, udvikles sporer og bakterier af mikroorganismer ikke, og planter påvirkes ikke af svampe- og bakteriesygdomme. Repræsentanter for familierne Solanaceae, Poppy, Rubiaceae, Ranunculaceae osv. er særligt rige på alkaloider.
Glykosider - en bred gruppe naturlige stoffer, forbindelser af sukkerarter med alkoholer, aldehyder, phenoler og andre stoffer. En række planteglykosider bruges i medicin. Glykosider inkluderer også cellesaftpigmenter - flavonoider. En af dem - anthocyaniner– giv cellesaft rød, blå eller lilla; Andet - flavoner- gul. Anthocyaniner er forbundet med farven på blomster af mange planter. Farveskemaet bestemmes af cellesaftens reaktion: hvis det er surt, dominerer røde toner, hvis det er neutralt, dominerer lilla toner, og med en let alkalisk reaktion dominerer blå toner. Udseendet af nuancer er også påvirket af dannelsen af anthocyaninkomplekser med forskellige metaller. Flavoner bestemmer gul kronblade af en række planter.
Betydningen af organiske syrer, tanniner, alkaloider og glykosider af cellesaft i cellemetabolismen er ikke blevet tilstrækkeligt belyst. Tidligere blev de betragtet som endelige bytteprodukter. Det har nu vist sig, at mange af dem kan geninddrages i metaboliske processer og derfor kan betragtes som reservestoffer.
Ud over funktionen med at akkumulere reservestoffer og affald udfører vakuoler i planteceller en anden vigtig funktion– opretholdelse af turgor. Koncentrationen af ioner og sukker i centralvakuolens cellesaft er normalt højere end i cellevæggen; Tonoplasten bremser betydeligt diffusionen af disse stoffer fra vakuolen og er samtidig let gennemtrængelig for vand. Derfor vil vand komme ind i vakuolen. Denne ensrettede proces med vanddiffusion gennem en selektivt permeabel membran kaldes osmose. Vand, der kommer ind i cellesaften, udøver pres på vægprotoplasten og gennem den på cellevæggen, hvilket forårsager dens spændte, elastiske tilstand eller turgor celler. Turgor sikrer, at ikke-lignificerede planteorganer bevarer deres form og position i rummet, såvel som deres modstandsdygtighed over for mekaniske faktorer.
Hvis en celle anbringes i en hypertonisk opløsning af et eller andet ikke-toksisk salt eller sukker (dvs. i en opløsning med en højere koncentration end koncentrationen af cellesaft), så sker der en osmotisk frigivelse af vand fra vakuolen. Som et resultat af dette reduceres dens volumen, den elastiske vægprotoplast bevæger sig væk fra cellevæggen, turgor forsvinder og plasmolyse celler( Fig.2.9).
Ris. 2.9. Plasmolyse skema: 1 – celle i en tilstand af turgor; 2 - begyndelsen af plasmolyse; 3 - fuldstændig plasmolyse.
Plasmolyse er normalt reversibel. Når en celle placeres i vand eller en hypotonisk opløsning, absorberes vand igen kraftigt af centralvakuolen, protoplasten presses igen mod cellevæggen, og turgor genoprettes. Plasmolyse kan tjene som en indikator for en celles levende tilstand; en død celle plasmolyseres ikke, da den ikke har selektivt permeable membraner.
Tab af turgor får planten til at visne. Når de visner i luft under forhold med utilstrækkelig vandforsyning, krymper de tynde cellevægge samtidig med protoplasten og bliver foldet.
Turgortryk bevarer ikke kun formen af ikke-lignificerede plantedele, det er også en af cellevækstfaktorerne, højde celler udstrækning på grund af absorptionen af vand og en stigning i størrelsen af vakuolen. Dyreceller har ikke en central vakuole; deres vækst sker hovedsageligt på grund af en stigning i mængden af cytoplasma, derfor er størrelsen af dyreceller normalt mindre end planteceller.
Den centrale vakuole opstår fra sammensmeltningen af adskillige små vakuoler, der er til stede i meristematiske (embryonale) celler. Disse cytoplasmatiske vakuoler menes at være dannet af membraner endoplasmatisk retikulum eller Golgi-apparat.
Inklusioner
Dannelsen af indeslutninger er forårsaget af overdreven ophobning af visse metaboliske produkter i visse dele af cellen - i vakuolen, hyaloplasma, forskellige organeller og sjældnere i cellevæggen. Disse stoffer udfælder ofte i amorf eller krystallinsk form - indeslutninger. Inklusioner har en bestemt form og er tydeligt synlige under et lysmikroskop. Ved tilstedeværelsen af visse indeslutninger, deres form og fordeling kan en art, slægt og plantefamilier skelnes fra andre, derfor tjener de ofte som vigtige diagnostisk tegn ved analyse af medicinske plantematerialer.
Inklusioner er enten reservestoffer(forbindelser midlertidigt fjernet fra stofskiftet), eller endelige produkter udveksle. Den første kategori af indeslutninger omfatter stivelseskorn,lipid dråber Og proteinaflejringer; til den anden - krystaller nogle stoffer.
Stivelseskorn– de mest almindelige indeslutninger af planteceller. Polysaccharid stivelse– hovedtypen af reservenæringsstoffer til planter. Han er også den mest vigtig forbindelse brugt som føde af planteædere. Stivelse fra korn, kartoffelknolde og bananfrugter er den vigtigste kilde til menneskelig ernæring. Hvedemel består af næsten 75% stivelseskorn, i kartoffelknolde udgør stivelse 20-30%. Kemisk er stivelse en alfa-1,4-D-glucan, molekylerne har form af forgrenede kæder, i stivelseskornet er de placeret langs radier.
Stivelseskorn dannes i plastiders stroma. I kloroplaster aflejres korn i lys assimilativ (primær) stivelse, dannet, når der er et overskud af sukker - produkter af fotosyntese. Dannelsen af osmotisk inaktiv stivelse forhindrer en stigning i osmotisk tryk i kloroplasten. Om natten, når fotosyntese ikke forekommer, hydrolyseres assimilativ stivelse til sukker ved hjælp af enzymer og transporteres til andre dele af planten. Reserve (sekundær) stivelse aflejret i celle amyloplaster forskellige organer planter (rødder, underjordiske skud, frø) fra sukkerarter, der strømmer fra fotosyntetiske celler. Om nødvendigt omdannes lagerstivelse også til sukkerarter.
Dannelsen af stivelseskorn begynder på visse punkter i plastidens stroma, kaldet uddannelsescentre. Kornvækst sker ved successiv aflejring af lag af stivelse omkring uddannelsescenter. Tilstødende lag i ét korn kan have anden indikator brydning af lys, og så er de synlige under et mikroskop - lagdelt stivelseskorn. Arrangementet af lag kan være koncentrisk(hvede) eller excentrisk(kartoffel) ( ris. 2.10). Hvis der er ét uddannelsescenter i amyloplasten, omkring hvilket lag af stivelse er aflejret, så enkel, hvis to eller flere, så dannes det kompleks korn, bestående af flere simple. semi-sammensat korn dannes, når stivelse først aflejres omkring flere punkter, og derefter, efter kontakt med simple korn, opstår der almindelige lag omkring dem ( ris. 2.10).
Formen, størrelsen, antallet i amyloplasten og strukturen (placering af uddannelsescentret, lagdeling, tilstedeværelse eller fravær af revner) af stivelseskorn er ofte specifikke for plantearten ( ris. 2.10). Typisk er stivelseskorn sfæriske, ægformede eller linseformede, men i kartofler er de uregelmæssige. De største korn (op til 100 mikron) er karakteristiske for kartoffelknoldceller; i hvedekorn er de af to størrelser - små (2-9 mikron) og større (30-45 mikron). Majskornceller er karakteriseret ved små korn (5-30 mikron). Ris, havre og boghvede har komplekse stivelseskorn.
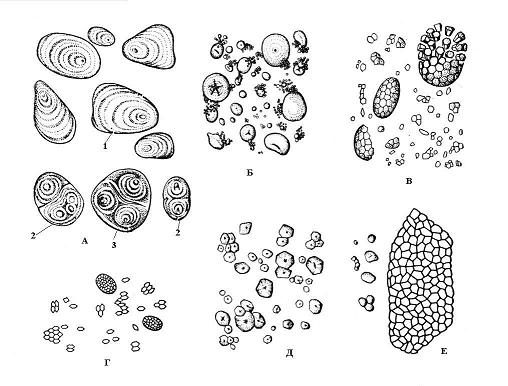
Ris. 2.10. Stivelseskorn forskellige typer planter: A – kartofler; B - hvede; B - havre; G - ris; D - majs; E – boghvede; 1 - simpelt korn; 2 - kompleks korn; 3 – semi-kompleks korn.
Reagenset til stivelse er en opløsning af jod i en opløsning af kaliumiodid - Lugols reagens. Den farver stivelseskorn blåviolette.
Stivelsesaflejringer er udbredte i alle planteorganer, men frø, underjordiske skud (knolde, løg, jordstængler) og parenkym i det ledende væv i rødder og stængler af træagtige planter er særligt rige på det.
Lipid dråber findes i næsten alle planteceller. Fede olier ophobes i et stort antal planter og er den næstvigtigste form for reservenæringsstoffer efter stivelse. Frø og frugter er særligt rige på dem. Frøene fra nogle planter (solsikke, bomuld, jordnødder) kan indeholde op til 40 vægtprocent olie af tørstof. Derfor opnås vegetabilsk fedt hovedsageligt fra frø.
Lipiddråber akkumuleres direkte i hyaloplasmaet. De ligner små sfæriske legemer, hver dråbe er adskilt fra hyaloplasmaet af en membran. Nogle gange kaldes lipiddråber sfærosomer.
Reagenset til fed olie er et farvestof SudanIII, er lipiddråber farvet orange-røde.
Protein indeslutninger i form af forskellige amorfe eller krystallinske aflejringer dannes i forskellige organeller i cellen. Oftest kan proteinkrystaller findes i kernen, sjældnere - i hyaloplasma, plastidstroma, i forlængelserne af de endoplasmatiske retikulum-cisterner, matrixen af peroxisomer og mitokondrier. Størrelsen af proteinkrystaller er oftest over opløsningen af et lysmikroskop.
Opbevaringsproteiner tilhører kategorien simple proteiner - proteiner i modsætning til komplekse proteiner - proteiner, der danner grundlaget for protoplasten. De aflejres i de største mængder i lagervævet af tørre frø i formen aleurone korn, eller proteinlegemer.
Aleuronkorn er sædvanligvis kugleformede og varierer i størrelse (0,2-20 µm). De er omgivet af en membran og indeholder en amorf proteinmatrix, hvori krystallinske indeslutninger er nedsænket - en (mindre ofte, 2-3) proteinkrystal med romboedrisk form og afrundet globoider(fra én til mange) ( ris. 2.11). Globoider består af phytin (et salt af inositolhexaphosphorsyre) og er et lagersted for reservefosfor. Aleuronkorn indeholdende krystaller kaldes kompleks. De er karakteristiske for opbevaringsceller af oliefrø (hør, solsikke, græskar, sennep, ricinusolie osv.). Mindre almindelig enkel aleuronkorn, der ikke indeholder krystaller, men kun amorft protein (bælgfrugter, ris, majs) ( ris. 2.12).
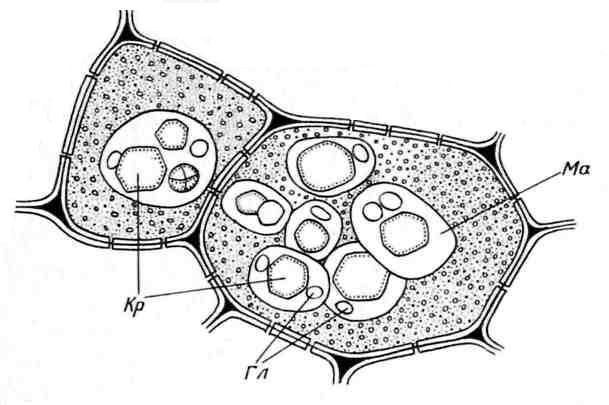
Ris. 2.11. Aleuronkorn i endospermceller fra ricinusbønnefrø:Kr- proteinkrystaller; Gl– globoider; Ma– proteinmatrix.
Under frøudvikling aflejres lagerproteiner i vakuoler. Når frø modnes, ledsaget af deres dehydrering, tørrer proteinvakuoler ud, protein og phytin bundfældes ud af opløsningen og kan krystallisere. Under frøspiring absorberer aleuronkorn vand, svulmer op og omdannes gradvist til typiske vakuoler. Proteiner og stoffer fra globoider bruges på vækst og udvikling af frøplanten.
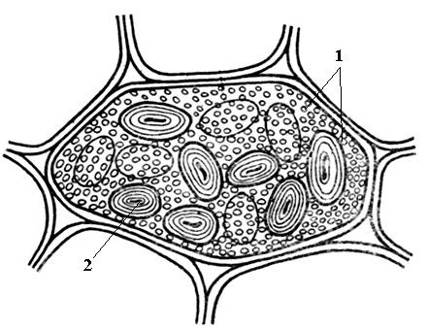
Ris. 2.12. Simpelt aleuron- og stivelseskorn i en kimbladscelle med bønnefrø: 1 – simple aleuronkorn; 2 – stivelseskorn.
Proteinindeslutninger kan farves gyldengule med Lugols reagens.
Calciumoxalatkrystaller findes ofte i planteceller. De deponeres kun i vakuoler. Formen af calciumoxalatkrystaller er ret forskelligartet ( ris. 2,13) og er ofte specifik for visse planter, som bruges til diagnosticering af medicinske plantematerialer. Det kan være enkeltkrystaller romboedrisk, oktaedrisk eller aflang form (hønebaneblade), drusere– stjerneformede sammenvoksninger af sfæriske krystaller (blade af knotweed, datura, senna, rabarberrødder) Rapids– små nåleformede krystaller samlet i bundter (liljekonvalblade, rhizomer) styloider– større, stavformede krystaller (liljekonvalblade) og krystal sand– klynger af mange små enkeltkrystaller (belladonna-blade). De mest almindelige er drusen. 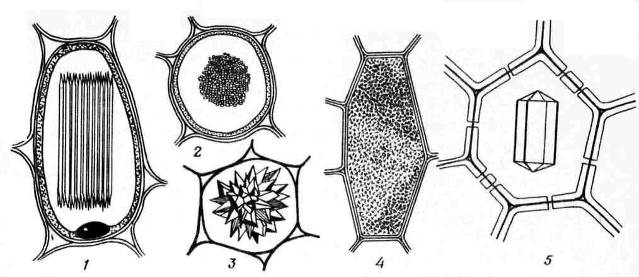
Ris. 2.13. Calciumoxalat krystal dannes: 1,2 – raphidas (1 – sidebillede, 2 – tværsnit); 3 – drusen; 4 - krystallinsk sand; 5 – enkelt krystal.
Langs fibrene i barken eller langs bladårerne på en række planter (egebark, lakridsrødder, sennablade) krystalbærende foring- befinde sig parallelle rækker celler med enkeltkrystaller af calciumoxalat ( Fig.2.14).
![]()
Ris. 2.14. En åre med en krystallinsk foring i et sennablad.
I modsætning til dyr, som frigiver overskydende ioner under ydre miljø sammen med urin, planter, der ikke har udviklede organer sekreter tvinges til at akkumulere dem i vævene. Det antages normalt, at calciumoxalatkrystaller er slutproduktet af celleliv, beregnet til at fjerne overskydende calcium. Faktisk dannes krystaller i store mængder i de organer og væv, som planter fra tid til anden udskiller (blade og bark). Der er dog tegn på, at krystaller kan forsvinde fra vakuoler. I dette tilfælde kan de betragtes som et sted for aflejring af reservekalcium.
Den kemiske natur af calciumoxalatkrystaller kan bekræftes ved virkningen af koncentreret mineralske syrer. Under påvirkning af saltsyre opløses krystallerne. Når det udsættes for svovlsyre, omdannes calciumoxalat til uopløseligt calciumsulfat (gips) og danner talrige nåleformede krystaller.
Tæt på krystallinske indeslutninger cystolitter. De består oftest af calciumcarbonat eller silica og er klyngeformede formationer, der opstår på fremspringene af cellevæggen, der rager ind i cellen ( ris. 2.15). Cystolitter er karakteristiske for planter af nælde- og morbærfamilien. Betydningen af cystolitter er endnu ikke klarlagt.
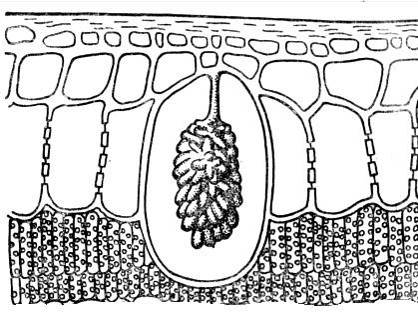
Ris. 2.15. Cystolith i epidermal celle af et ficus blad.
Cellevæg
Cellevæg ( celle membran) – karakteristisk træk plantecelle, der adskiller den fra en dyrecelle. Cellevæggen giver cellen sin specifikke form. Planteceller dyrket på særlige næringsmedier, hvor væggen er enzymatisk fjernet, antager altid en kugleformet form. Cellevæggen giver cellen styrke og beskytter protoplasten, den afbalancerer turgortrykket og forhindrer dermed brud på plasmalemmaet. Samlingen af cellevægge danner det indre skelet, som understøtter plantekroppen og giver den mekanisk styrke.
Cellevæggen er farveløs og gennemsigtig, let permeabel sollys. Normalt er væggene mættede med vand. Cellevægssystemet transporterer vand og lavmolekylære forbindelser opløst i det (transport langs apoplasten).
Cellevæggen består hovedsageligt af polysaccharider, som kan opdeles i skeletstoffer Og matrix stoffer.
Skeletsubstans plantecellevæg er cellulose (fiber), som er en beta-1,4-D-glucan. Dette er den mest almindelige organisk stof biosfære. Cellulosemolekyler er meget lange uforgrenede kæder; de er placeret parallelt med hinanden i grupper på flere dusin og holdes sammen af talrige hydrogenbindinger. Som resultat, mikrofibriller, som skaber væggens strukturelle ramme og bestemmer dens styrke. Cellulosemikrofibriller er kun synlige med et elektronmikroskop, deres diameter er 10-30 nm, og deres længde når flere mikrometer.
Cellulose er uopløselig og kvælder ikke i vand. Det er meget kemisk inert og opløses ikke i organiske opløsningsmidler, koncentrerede alkalier og fortyndede syrer. Cellulose mikrofibriller er elastiske og meget strækbare (ligner stål). Disse egenskaber bestemmer den udbredte anvendelse af cellulose og dets produkter. Verdensproduktionen af bomuldsfibre, som næsten udelukkende består af cellulose, er 1,5 10 7 tons om året. Røgfrit krudt, acetatsilke og viskose, cellofan og papir fremstilles af cellulose. Kvalitativ reaktion udføres på cellulose med et reagens klor-zink-jod, cellulosecellevæggen bliver blåviolet.
Hos svampe er cellevæggens skeletsubstans kitin– et polysaccharid bygget af glucosaminrester. Kitin er endnu mere holdbart end cellulose.
Mikrofibriller er nedsænket i amorfe matrix, normalt en vandmættet plastikgel. Matrixen er en kompleks blanding af polysaccharider, hvis molekyler består af rester af flere forskellige sukkerarter og er kortere og forgrenede kæder end cellulose. Matrixpolysaccharider bestemmer sådanne egenskaber af cellevæggen som stærk hævelse, høj permeabilitet for vand og lavmolekylære forbindelser opløst i den, og kationbytteregenskaber. Matrix polysaccharider er opdelt i to grupper - pektinstoffer Og hemicelluloser.
Pektiske stoffer kraftigt svulme op eller opløses i vand. De ødelægges let af alkalier og syrer. De enkleste repræsentanter for pektinstoffer er vandopløselige pektinsyrer– polymerisationsprodukter af alfa-D-galacturonsyre (op til 100 enheder), forbundet med 1,4-bindinger til lineære kæder (alfa-1,4-D-galacturonan). Pektinsyrer (pektiner)– disse er polymere forbindelser med højere molekylvægt (100-200 enheder) af alfa-D-galacturonsyre, hvor carboxylgrupperne er delvist methylerede. Pectates Og pektinater– calcium- og magnesiumsalte af pektin- og pektinsyrer. Pektinsyrer, pectater og pectinater er opløselige i vand i nærværelse af sukkerarter og organiske syrer for at danne tætte geler.
Planternes cellevægge indeholder hovedsageligt protopektiner– højmolekylære polymerer af methoxyleret polygalacturonsyre med arabinaner og galactaner; i tokimbladede planter indeholder galacturonan-kæderne en lille mængde rhamnose. Protopectiner er uopløselige i vand.
Hemicelluloser er forgrenede kæder bygget af neutrale sukkerrester, glucose, galactose, mannose, xylose er mere almindelige; polymerisationsgrad 50-300. Hemicelluloser er kemisk mere stabile end pektinstoffer, de er sværere at hydrolysere og kvælder mindre let i vand. Hemicelluloser kan aflejres i frøs cellevægge som reservestoffer (daddelpalme, persimmon). Pektinstoffer og hemicelluloser er forbundet ved indbyrdes overgange. Ud over polysaccharider er et særligt strukturelt protein til stede i cellevægsmatrixen. Det er bundet til arabinosesukkerrester og er derfor et glykoprotein.
Matrixpolysaccharider gør mere end blot at fylde mellemrummene mellem cellulosemikrofibriller. Deres kæder er arrangeret på en velordnet måde og danner talrige bindinger både med hinanden og med mikrofibriller, hvilket øger cellevæggens styrke markant.
Plantecellevægge gennemgår ofte kemiske modifikationer. Lignificering, eller lignificering opstår, når det aflejres i matrixen lignin– en polymerforbindelse af phenolisk natur, uopløselig i vand. Den lignificerede cellevæg mister sin elasticitet, dens hårdhed og trykstyrke øges kraftigt, og dens permeabilitet for vand falder. Reagenser for lignin er: 1) phloroglucinol Og koncentreret saltsyre eller svovlsyre(lignificerede vægge får en kirsebærrød farve) og 2) sulfatanilin, under hvilken indflydelse de lignificerede vægge bliver citrongule. Lignificering er karakteristisk for cellevæggene i det ledende væv af xylem (træ) og det mekaniske væv af sclerenchyma.
Prøveudtagning, eller suberinisering opstår som følge af aflejring fra inde cellevæg af hydrofobe polymerer - suberina Og voks. Suberin er en blanding af estere af polymere fedtsyrer. Voksmonomerer er fedtalkoholer og voksestere. Voksen ekstraheres let af organiske opløsningsmidler og smelter hurtigt og danner krystaller. Suberin er en amorf forbindelse, der ikke smelter eller opløses i organiske opløsningsmidler. Suberin og voks, der danner vekslende parallelle lag, beklæder hele cellehulrummet indefra i form af en film. Suberin-filmen er praktisk talt uigennemtrængelig for vand og gasser, så efter dannelsen dør cellen normalt. Suberisering er karakteristisk for cellevægge dækkevæv trafikpropper. Reagenset for suberiserede cellevægge er Sudan III, orange-rød farve.
Cotinization De ydre vægge af de epidermale vævsceller er blotlagt. Kutin Og voks aflejret i skiftende lag på den ydre overflade af cellevæggen i form af en film - neglebånd. Cutin er en fedtlignende polymerforbindelse, der ligner kemisk natur og egenskaber af suberin. Kutikula beskytter planten mod overdreven fordampning af vand fra plantens overflade. Du kan farve det med et reagens Sudan III i orange-rød farve.
Mineralisering cellevæg opstår på grund af aflejring i matrixen stor mængde mineralske stoffer, oftest silica (siliciumoxid), sjældnere calciumoxalat og carbonat. Mineraler give væggen hårdhed og skrøbelighed. Silicaaflejring er karakteristisk for de epidermale celler i padderok, kværn og græs. Stivheden af stængler og blade erhvervet som følge af silicificering tjener som et beskyttende middel mod snegle og reducerer også betydeligt planternes smag og næringsværdi.
Nogle specialiserede celler har slim cellevæg. I dette tilfælde aflejres amorfe, stærkt hydrerede sure polysaccharider i stedet for en sekundær cellulosevæg i form af slim Og tandkød, tæt i kemisk natur på pektinstoffer. Slim opløses godt i vand for at danne slimopløsninger. Gummi er klistret og strækker sig i tråde. Når de er tørre, har de en liderlig konsistens. Når slim aflejres, skubbes protoplasten gradvist mod midten af cellen, dens volumen og volumen af vakuolen falder gradvist. Til sidst kan cellehulen blive fuldstændig fyldt med slim, og cellen dør. I nogle tilfælde kan slim passere gennem den primære cellevæg til overfladen. Golgi-apparatet spiller en stor rolle i syntesen og udskillelsen af slim.
Slim udskilt af planteceller udfører forskellige funktioner. Således fungerer rodkappens slim som et smøremiddel, der letter væksten af rodspidsen i jorden. Slimkirtlerne fra insektædende planter (solduge) udskiller fangende slim, som insekter klæber til. Slimet, der udskilles af frøskallens ydre celler (hør, kvæde, plantain) sikrer frøet til jordoverfladen og beskytter frøplanten mod at tørre ud. Slim farves med et reagens methylenblåt i blå farve.
Frigivelsen af tandkød sker normalt, når planter bliver såret. For eksempel observeres tyggegummi fra sårede områder af stammer og grene ofte i kirsebær og blommer. Kirsebærlim er en hærdet tyggegummi. Gum optræder beskyttende funktion, lukker såret fra overfladen. Gummier dannes hovedsageligt i træagtige planter fra bælgplantefamilierne (akacie, tragant astragalus) og Rosaceae fra blommeunderfamilien (kirsebær, blomme, abrikos). Tandkød og slimhinder bruges i medicin.
Cellevæggen er et produkt af protoplastens vitale aktivitet. Matrixpolysaccharider, vægglykoprotein, lignin og slim dannes i Golgi-apparatet. Cellulosesyntese, dannelse og orientering af mikrofibriller udføres af plasmalemmaet. Stor rolle i orienteringen af mikrofibriller hører det til mikrotubuli, som er placeret parallelt med de aflejrede mikrofibriller nær plasmalemmaet. Hvis mikrotubuli ødelægges, dannes der kun isodiametriske celler.
Cellevægsdannelse begynder under celledeling. I delingsplanet dannes en celleplade, et enkelt lag fælles for de to datterceller. Den består af pektinstoffer med en halvflydende konsistens; der er ingen cellulose. I en voksencelle er cellepladen bevaret, men undergår ændringer, hvorfor den kaldes median, eller intercellulær plade (intercellulært stof)(ris. 2.16). Medianpladen er normalt meget tynd og næsten ikke til at skelne.
Umiddelbart efter dannelsen af cellepladen begynder dattercellernes protoplaster at nedlægge deres egen cellevæg. Det aflejres indefra både på overfladen af cellepladen og på overfladen af andre cellevægge, der tidligere tilhørte modercellen. Efter deling går cellen ind i forlængelsesvækstfasen, som er forårsaget af intens osmotisk absorption af vand af cellen i forbindelse med dannelsen og væksten af den centrale vakuole. Turgortryk begynder at strække væggen, men det rives ikke på grund af det faktum, at nye dele af mikrofibriller og matrixstoffer konstant aflejres i den. Aflejringen af nye dele af materiale sker jævnt over hele overfladen af protoplasten, så tykkelsen af cellevæggen ikke falder.
Væggene i celler, der deler og vokser, kaldes primær. De indeholder meget (60-90%) vand. Tørstoffet er domineret af matrixpolysaccharider (60-70%), celluloseindholdet overstiger ikke 30%, og der er ingen lignin. Tykkelsen af den primære væg er meget lille (0,1-0,5 mikron).
For mange celler ophører cellevægsaflejring samtidig med ophør af cellevækst. Sådanne celler er omgivet af en tynd primær væg indtil livets afslutning ( ris. 2.16).
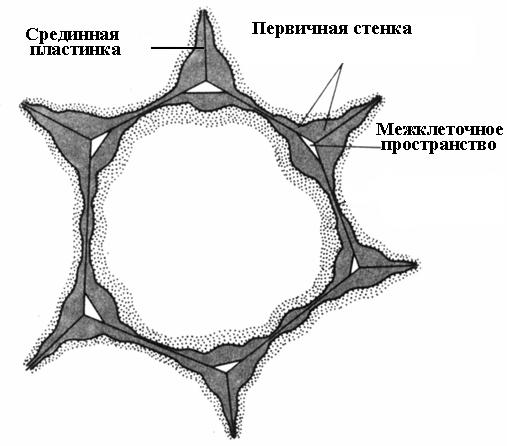
Ris. 2.16. Parenkymcelle med en primær væg.
I andre celler fortsætter vægaflejring, selv efter at cellen når sin endelige størrelse. I dette tilfælde øges vægtykkelsen, og volumenet optaget af cellehulrummet falder. Denne proces kaldes sekundær fortykkelse vægge, og selve væggen hedder sekundær(ris. 2.17).
Den sekundære væg kan betragtes som en ekstra, der hovedsageligt udfører en mekanisk støttefunktion. Nemlig sekundær væg ansvarlig for egenskaberne ved træ, tekstilfiber, papir. Den sekundære væg indeholder væsentligt mindre vand end den primære væg; den er domineret af cellulosemikrofibriller (40-50 % af tørstofvægten), som er placeret parallelt med hinanden. Af matrixpolysacchariderne er hemicelluloser (20-30%) typiske, og der er meget få pektinstoffer. Sekundære cellevægge gennemgår normalt lignificering. I ikke-lignificerede sekundære vægge (hørbastfibre, bomuldshår) kan celluloseindholdet nå op på 95%. Fantastisk indhold og strengt ordnet orientering af mikrofibriller bestemmer de høje mekaniske egenskaber af de sekundære vægge. Ofte dør celler, der har en sekundær lignificeret cellevæg, efter at den sekundære fortykkelse er fuldstændig.
Median lamina limer naboceller sammen. Hvis det opløses, mister cellevæggene kontakten med hinanden og adskilles. Denne proces kaldes maceration. Naturlig maceration er ret almindelig, hvor pektinstofferne i den midterste plade omdannes til en opløselig tilstand ved hjælp af enzymet pectinase og derefter vaskes væk med vand (overmodne frugter af pære, melon, fersken, banan). Ofte observeres delvis maceration, hvor midterpladen ikke opløses over hele overfladen, men kun i hjørnerne af cellerne. På grund af turgortryk er naboceller på disse steder afrundet, hvilket resulterer i dannelsen intercellulære rum(ris. 2.16). De intercellulære rum danner et enkelt forgrenet netværk, som er fyldt med vanddamp og gasser. Således forbedrer intercellulære rum gasudveksling af celler.
Feature sekundær væg - dens ujævne aflejring oven på den primære væg, som et resultat af hvilket utykkede områder forbliver i den sekundære væg - porer. Hvis den sekundære væg ikke når en stor tykkelse, ser porerne ud som små fordybninger. I celler med en stærk sekundær væg ser porerne i tværsnit ud som radiale kanaler, der strækker sig fra cellehulrummet til den primære væg. Baseret på formen af porekanalen er der to typer porer: enkel og om kantet(Fig. 2.17).
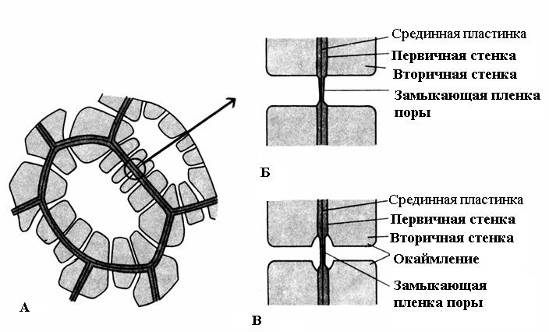
Ris. 2.17. Pore typer: A – celler med sekundære vægge og talrige simple porer; B - et par simple porer; B – par afgrænsede porer.
U simple porer Diameteren af porekanalen er den samme i hele dens længde og har form som en smal cylinder. Simple porer er karakteristiske for parenkymceller, bast- og træfibre.
Porer i to tilstødende celler har en tendens til at dukke op overfor hinanden. Disse almindelige porer ligner en kanal, adskilt af en tynd skillevæg mellem midterpladen og den primære væg. Denne kombination af to porer af tilstødende vægge af naboceller kaldes par porer og fungerer som én helhed. Den del af væggen, der adskiller dem, kaldes lukkefilm af poren, eller poremembran. I levende celler er porens lukkefilm gennemtrængt med talrige plasmodesmata(ris. 2.18).
Plasmodesmata findes kun i planteceller. De er strenge af cytoplasma, der krydser væggen af tilstødende celler. Antallet af plasmodesmata i en celle er meget stort - fra flere hundrede til titusinder; normalt samles plasmodesmata i grupper. Diameteren af den plasmodesmale kanal er 30-60 nm. Dens vægge er foret med plasmalemma, kontinuerligt med plasmalemmaet fra tilstødende celler. I midten af plasmodesmata er der en membrancylinder - central stang af plasmadesmata kontinuert med membranerne af elementerne i det endoplasmatiske reticulum af begge celler. Mellem den centrale stang og plasmamembranen i kanalen er der hyaloplasma, kontinuerligt med hyaloplasmaet af tilstødende celler.
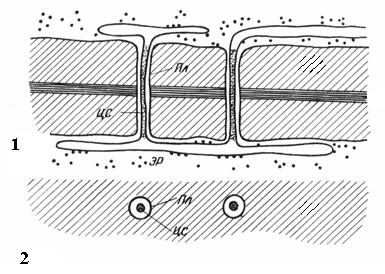
Ris. 2.18. Plasmodesmata under et elektronmikroskop (diagram): 1 – på et længdesnit; 2 - på et tværsnit; Pl– plasmalemma; CA– central stang af plasmodesmata; ER– element i det endoplasmatiske retikulum.
Således er celleprotoplaster ikke fuldstændigt isolerede fra hinanden, men kommunikerer gennem plasmodesmata-kanaler. De bærer intercellulær transport af ioner og små molekyler og transmitterer også hormonelle stimuli. Gennem plasmodesmata, protoplaster af celler i plante organisme danne en enkelt helhed kaldet simplast, og transporten af stoffer gennem plasmodesmata kaldes symplastisk I modsætning til apoplastisk transport langs cellevægge og intercellulære rum.
U afgrænsede porer(ris. 2.17) kanalen indsnævres kraftigt under aflejringen af cellevæggen, så den indre åbning af poren, der åbner ind i cellehulen, er meget smallere end den ydre, der støder op til den primære væg. Afgrænsede porer er karakteristiske for tidligt døende celler af vandledende elementer af træ. I dem udvider porekanalen sig tragtformet mod lukkefilmen, og sekundærvæggen hænger i form af en rulle over den udvidede del af kanalen og danner et porekammer. Navnet afgrænset pore kommer af, at set fra overfladen, ligner den indre åbning en lille cirkel eller snævert mellemrum, mens det ydre hul så at sige grænser op til det indre i form af en cirkel med større diameter eller en bredere slids.
Porer letter transport ind