Клетки растений окружены плотной полисахаридной оболочкой, выстланной изнутри плазмалеммой.
Образование клеточной стенки происходит в метафазе и телофазе клеточного деления. В экваториальной зоне деления возникает срединная пластинка, состоящая из пектата кальция, которая, нарастая от центра к периферии, отделяет одну новообразованную клетку от другой. Срединная пластинка с той и другой стороны покрывается первичной клеточной стенкой. Рост в толщину происходит за счет наложения новых слоев со стороны содержимого каждой клетки. Рост клетки в длину начинается с разрыхления матрикса. В этом процессе важную роль играют фитогормоны. В образовавшиеся полости поступают новые порции материала, из которого строится клеточная стенка. Синтез и транспорт этих веществ осуществляются главным образом вакуолями аппарата Гольджи.
Клеточную стенку делящихся и растущих растяжением клеток называют первичной. После прекращения роста клетки на первичную клеточную стенку изнутри откладываются новые слои и возникает прочная вторичная клеточная стенка.
В состав клеточной стенки входят структурные компоненты (целлюлоза у растений, хитин у грибов), компоненты матрикса стенки (гемицеллюлозы, пектин, белки), инкрустирующие компоненты (лигнин, суберин) и вещества, откладывающиеся на поверхности стенки (кутин и воска). Клеточные стенки могут содержать также силикаты и карбонаты кальция.
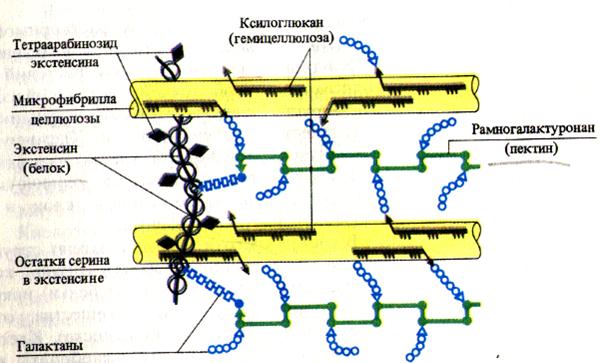
Целлюлоза (полимер b-D-глюкозы), гемицеллюлозы (полимеры гексоз и пентоз) и пектиновые вещества (производные уроновых кислот) являются углеводными компонентами клеточных стенок. Целлюлоза и пектиновые вещества адсорбируют воду, обеспечивая оводненность клеточной стенки. Пектиновые вещества, содержащие много карбоксильных групп, связывают ионы двухвалентных металлов, которые способны обмениваться на другие катионы (Н + , К + и т.д.). Это обусловливает катионообменную способность клеточных стенок растений. Помимо углеводных компонентов в состав матрикса клеточной стенки входит также структурный белок, называемый экстенсином. Это гликопротеин, содержащий более 20% L-оксипролина от суммы аминокислот. По этому признаку белок клеточных стенок растений сходен с межклеточным белком животных - коллагеном.
Целлюлоза: А – структура молекулы целлюлозы; Б – ассоциации молекулы целлюлозы: 1 – мицелла, 2 – микрофибрилла, 3 – макрофибрилла
Основным инкрустирующим веществом клеточной стенки является лигнин. Интенсивная лигнификация клеточных стенок начинается после прекращения роста клетки. Лигнин представляет собой полимер с неразветвленной молекулой, состоящей из ароматических спиртов (п-кумарового, кониферилового, синапового). Разрушение и конденсация лигнина в почве - один из факторов образования гумуса. Интенсивная лигнификация (пропитка слоев целлюлозы лигнином) клеточных оболочек начинается после прекращения роста клетки. Лигнин может откладываться отдельными участками - в виде колец, спиралей или сетки, как это наблюдается в оболочках клеток проводящей ткани - ксилемы, или сплошным слоем, за исключением тех мест, где осуществляются контакты между соседними клетками в виде плазмодесм. Лигнин скрепляет целлюлозные волокна и действует как очень твердый и жесткий каркас, усиливающий прочность клеточных стенок на растяжение и сжатие. Он же обеспечивает клеткам дополнительную защиту от физических и химических воздействий, снижает водопроницаемость. Содержание лигнина в оболочке достигает 30%. Инкрустация им клеточных оболочек приводит к их одревеснению, которое часто влечет за собой отмирание живого содержимого клетки. Лигнин в сочетании с целлюлозой придает особые свойства древесине, которые делают ее незаменимым строительным материалом.
В регуляции водного и теплового режима растений участвуют ткани, стенки клеток которых пропитаны суберином. Отложение суберина делает стенки трудно проницаемыми для воды и растворов (например, в эндодерме, перидерме). Суберин откладывается на оболочку изнутри и делает ее практически непроницаемой для воды и растворов. В результате протопласт клетки отмирает и клетка заполняется воздухом. Такой процесс называется опробковением. Наблюдается опробковение оболочки клеток в покровных тканях многолетних древесных растений - перидерме, корке, а также в эндодерме корня. Суберин - основное вещество пробковых образований, которому эти образования главным образом обязаны своими свойствами: непроницаемостью для воды, для газов и малой теплопроводностью..
Поверхность эпидермальных клеток растений защищена гидрофобными веществами - кутином и восками: Предшественники этих соединений секретируются из цитоплазмы на поверхность, где и происходит их полимеризация. Слой кутина обычно пронизан полисахаридными компонентами стенки (целлюлозой, пектином) и образует кутикулу. Кутикула участвует в регуляции водного режима тканей и защищает клетки от повреждений и проникновения инфекции.
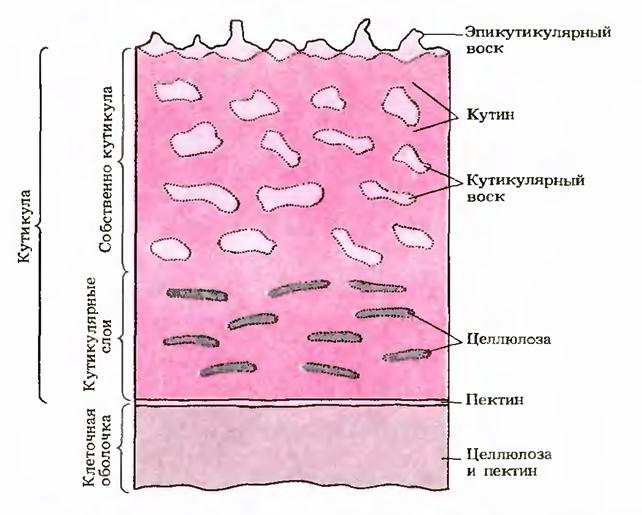
В оболочках эпидермальных клеток некоторых растений (злаков, осок и др.) накапливается большое количество минеральных веществ (минерализация), в первую очередь карбоната кальция и кремнезема. При минерализации листья и стебли растений становятся жесткими, твердыми и в меньшей степени поедаются животными.
В первичных клеточных стенках на долю целлюлозы приходится до 30% сухой массы стенки. Количество гемицеллюлоз и пектиновых веществ меняется в зависимости от объекта. Вместе с белками пектиновые вещества могут составлять около 30% сухой массы клетки, причем количество белка достигает 5 - 10%. Около 40% приходится на долю гемицеллюлоз.
1) Клеточная стенка - структурное образование. Функция: придаёт прочность и форму, защищает протопласт от внешних условий, участвует в проведении и поглощении веществ.
Основа клеточной оболочки(состав)- высокополимерные углеводы (целлюлоза, т.е клетчатка - не переваривается, указывает на низкую продуктивность), молекулы целлюлозы собраны в сложные пучки (мицелии), мицелии объединяются в фибриллы, их промежутки заполнены гемицеллюлозой (полуклетчатка - менее стойкое соединение) и пектином (полезные, набухают в воде, являются источником энергии).
Различают первичную и вторичную клеточные оболочки. Меристематические и молодые растущие клетки имеют первичную клеточную оболочку, тонкую, богатую пектином и гемицеллюлозой; фибриллы целлюлозы в матриксе первичной клеточной оболочки расположены неупорядоченно.
Вторичная клеточная оболочка образуется обычно по достижении клеткой окончательного размера и накладывается слоями на первичную со стороны протопласта. Во вторичной клеточной оболочке преобладает целлюлоза, ее фибриллы располагаются упорядочение, параллельно, но направление их в каждом слое иное, что повышает прочность клеточной оболочки. Во вторичной клеточной оболочке есть отверстия (поры), где клетки разделяют лишь первичная оболочка и плазмодесмы (цитоплазматические мостики, соединяющие соседние клетки растений).
Видоизменения клеточной стенки:
- Одревеснение клеточной оболочки происходит в результате отложения лигнина (неуглеводный компонент в фибриллах), клетки теряет эластичность, но могут пропускать воду. Эти клетки чаще мертвые, нежели живые. Стенки некоторых клеток могут включать: воск, кутину, суберин. Функции: придает клетке форму; отделяет одну клетку от другой, является скелетом для каждой клетки и придает прочность всему растению, выполняет защитную функцию.
- Опробкоеение вызывается особым жироподобным веществом - суберином. Такие оболочки становятся непроницаемыми для воды и газов, также, они не пропускают тепло, содержимое клеток с опробковевшими оболочками отмирает.
- Кутинизация заключается в выделении жироподобного вещества кутина. Обычно кутинизируются наружные стенки кожицы листьев и "травянистых стеблей. Это делает их менее проницаемыми для воды, уменьшает испарение воды у растений, охраняет от перегрева и ультрафиолета. Кутин образует на поверхности органа пленку, называемую кутикулой.
- Минерализация клеточных оболочек - это отложение: кремнезема и солей кальция. Наиболее сильно инкрустируются оболочки клеток кожицы листьев и стеблей злаков, осок, хвощей. Листьями злаков и осок можно поранить руки.
- Ослизнение оболочек - превращение целлюлозы и пектиновых веществ в слизи и камеди. Ослизнение хорошо наблюдается на семенах льна, находившихся в воде. Образование слизей способствует лучшему поглощению воды семенами и прикреплению их к почве.
2) Размножение: способность отдельно взятой особи дать начало целой серии себе подобных.
Делят на: половое и бесполое (собственной бесполое и вегетативное)
Вегетативное : новые особи развиваются из отдельных вегетативных органов или их взаимодействий. Осуществляется благодаря регинираций (св-во восстанавливать из части тела организм). Био значение: новый организм сходен с материнским.
Способы вегетативного размножения:
- размножение черенками (частью растения, которое не заражено, садят в субстрат,спородина),
- размножение методом прививки (путём осращивания частей нескольких растений, применяющийся в садоводстве),
- размножение клубнями (мясистые клубни с пит в-вами садят в землю, живородная гречиха),
- размножение отпрысками (образуют побеги на корнях, осина),
- размножение луковичками (осенью отсаживают от самого растения в землю)
- размножение усами (ползучие побегы,укореняются придат корнями, костяника, земляника)
- размножение корневищами (подземный побег, запас пит в-в, ландыш, фиалка, пырей)
Использование вегетативного размножения человеком. Остальное см в 40.
С давних пор человек, культивируя растения, стал использовать вегетативное размножение. Например, выращивание картофеля, земляники, банана во всех странах мира осуществляется только вегетативным путем – клубнями, усами и корневищами.
Использование вегетативного воспроизведения растений в сельскохозяйственной практике получило название искусственного вегетативного размножения .
Основные приемы искусственного вегетативного размножения сводятся к повторению тех, которые происходят у растений в естественных природных условиях.
Люди часто используют размножение черенками – частями зеленого или одревесневшего побега(виноград, смородина, крыжовник, роза, гвоздика, фикус) , клубнями (картофель, георгина, батат, топинамбур) , листьями (сенполия, глоксиния, бегония) , луковицами (лук, чеснок, тюльпан, нарцисс) , делением куста (смородина, пиретрум) и отводками (крыжовник, жимолость, клематис) , усами (клубника) , корневищами (сахарный тростник, ирисы, флоксы) , корневыми отпрысками(слива, малина, вишня, сирень) .
3) Тыквенные . Форма: травы. Корень стержневой. Стебель: лазающий, стелющий, вьющийся Лист: простой, черешковый, без прилистников.
Формула: раздельнополый
1) правильный женский Ca (5) Co (5) A 0 G (3) околоцветник под завязью
2) правильный мужской Ca (5) Со (5) А 2+2+1 G 0
Соцветие одиночное. Плод: тыквина
Представители: огурец, дыня, тыква, арбуз., кабачок
Значение: пищевое,кормовое
Клеточная оболочка способна к утолщению и видоизменению. В результате этого образуется ве вторичная структура. Утолщение оболочки происходит путем наложения новых слоев на нервпчпуго оболочку. Ввиду того что наложепие идет уже иа твердую оболочку, фибриллы целлюлозы в каждом слое лежат параллельно, а в соседних слоях - под углом друг к другу. Этим достигается значительная прочность и твердость вторичной оболочки. По мере того как число слоев фибрилл целлюлозы становится больше и толщина стенки увеличивается, она теряет эластичность и способность к росту. Во вторичной клеточпой стенке содержание целлюлозы значительно возрастает, в некоторых случаях до 60% и более. По мере дальнейшего старения клеток матрикс оболочки может заполняться различными веществами - лигнином, суберином (одревеснение или опробковение оболочки). Лигнин образуется из гемицеллюлозы н пектиновых веществ.[ ...]
Клеточная оболочка древесного волокна имеет несколько слоев: первичный, который называется наружной оболочкой волокна, и вторичный (стенка, состоящая, в свою очередь, из трех слоев: наружного, среднего и внутреннего). Между первичными стенками клеток находится слой межклеточного вещества, при помощи которого волокна соединяются друг с другом. Вторичная стенка относительно толстая и представляет собой главную массу объема клетки.[ ...]
Во вторичных слоях клеточных стенок древесины сосны накапливались в больших количествах маннаны (22%) и уроновый ангидрид (25%).[ ...]
[ ...]
Фаза утолщения клеточной стенки. Как происходит утолщение. В период разрастания протопласт окружен только первичной стенкой. Когда же древесная клетка достигает своего наибольшего размера по поверхности или вскоре после этого, стенка клетки утолщается. Это вызвано наслаиванием вторичной стенки на первичную, причем этот новый слой возникает в результате дальнейшей деятельности протопласта внутри полости клетки. Естественно, что клетки, в которых протопласт исчез, не могут продолжать утолщать свои стенки. Образование вторичной стенки является признаком необратимого изменения в клетке, дальнейшее разрастание которой уже исключено, но не обязательно исключается дальнейшее деление при условии, что получаемые таким образом дочерние клетки занимают такой же объем, как и первоначальная клетка.[ ...]
М.1ип - ковер, покрывало). Он состоит из таблитчатых тонкостенных клеток с густой цитоплазмой. Обычно он однорядный, но иногда бывает дву рядным или многорядным. Клетки «ноту ма сначала одноядерные, по позднее они часто становятся двухъядерными или даже многоядерными. Тапетум представляет собой физиологически чрезвычайно активную ткань: его клетки содержат ферменты, гормоны, и питательный материал, используемый в процессе микроспорогепеза. Имеются некоторые основания считать секреторный тип в эволюционном отношении первичным, а амебоидный - вторичным.[ ...]
Необходимо, однако, отметить, что эти данные следует рассматривать как приближенные, так как исходные препараты не были тщательно очищены.[ ...]
Трудно определить расположение в клеточной стенке полиуронидных гемицеллюлоз, потому что реагенты, используемые для их выявления, оказывают воздействие и на лигнин . Некоторые исследователи предполагают, что цементирующим веществом между фибриллами и различными слоями клеточной стенки являются гемицеллюлозы. Коэн считает даже, что лигнин вторичной стенки имеет одинаковую природу с гемицеллюлозами. Основанием для такого предположения служит, по-видимому, тот факт, что некоторые углеводы при обработке сильными кислотами могут давать нерастворимые остатки определенного рисунка. Следует подчеркнуть, однако, что участки, как тщательно обработанные реагентами, растворяющими гемицеллюлозы, так и не обработанные ими, дают при воздействии 72%-ной серной кислоты остатки очень похожей структуры .[ ...]
Для выяснения состава отдельных слоев клеточных стенок была сделана попытка количественного определения ксилоуронидов в разных слоях трахеид и либриформа . Измерения производились на волокнах из красной японской сосны, европейской пихты, бука и березы. Для этого волокна осторожно нитровали в среде уксусного ангидрида и четыреххлористого углерода. Затем наружный нитрованный слой удаляли растворением в ацетоне, после чего контролировали содержание пентозанов в остатке по фурфуролу. Было установлено, что пентозаны в древесных волокнах по слоям разделены неравномерно. Наибольшее количество пентозанов найдено в наружных слоях волокон и концентрация их падает от периферии к центру. Так, наружные слои волокон хвойной древесины содержат 50-80% пентозанов, а у лиственных почти 100%. Во вторичных слоях клеточных стенок у хвойных содержание пентозанов оказалось не более 2-4%, а у лиственных 8-10%. Таким образом, химический метод подтвердил результаты, полученные ранее методом сорбции ультрафиолетового света.[ ...]
Различают лигнин первичный, находящийся в одревесненных клеточных стенках (природный лигнин) и вторичный - изолированный лигнин. Последний является в значительной степени веществом, измененным в процессе изолирования и загрязненным примесями посторонних веществ. Изменение лигнина выражается в отщеплении метоксильных групп, внутримолекулярной конденсации и в других признаках.[ ...]
Многие различия между типами тканей обусловлены строением клеточной стенки, особенно вторичной. Как мы уже говорили, образование первичной клеточной стенки происходит в процессе растяжения клетки, и, следовательно, она должна обладать свойством растяжимости, тогда как вторичная стенка формируется уже после того, как удлинение прекратилось.[ ...]
Престон
Одновременно с этими внутренними изменениями наружная твердая стенка ооспоры расщепляется на ее вершине на пять зубцов, давая выход проростку, возникающему из центральной клетки (рис. 269, 3). Первое деление центральной клетки происходит поперечной перегородкой, перпендикулярной к ее длинной оси, и приводит к образованию двух функционально различных клеток. Из одной, более крупной клетки в дальнейшем образуется стеблевой побег, который на начальной стадии развития называют предростком, из другой, меньшей клетки - первый ризоид. Оба они растут путем поперечных клеточных делений. Предросток растет вверх и довольно быстро зеленеет, заполняясь хлоропластами, первый ризоид направляется вниз и остается бесцветным (рис. 269, 4). После ряда клеточных делений, сообщающих им строение однорядных нитей, происходит их дифференцировка на узлы и междоузлия, и дальнейший их верхушечный рост протекает уже так, как было описано выше для стебля. Из узлов предростка возникают вторичные предростки, мутовки листьев и боковые ветви стебля, из узлов первого ризоида - вторичные ризоиды и их мутовчатые волоски. Таким путем и формируется таллом, состоящий из нескольких стеблевых побегов в верхней части и нескольких сложных ризоидов в нижней части (рис. 2G9, 5).[ ...]
Надмолекулярная структура. На рис 6.10 приведена модель структуры клеточной стенки. Она включает 2 основных слоя: первичную стенку Р и вторичную Последняя подразделяется на 3 слоя: 5], 5 , Слой М, срединная пластинка, является межклеточным веществом, соединяющим клетки между собой.[ ...]
В последующих разделах (ем. часть II) будет исчерпывающе рассмотрена химия клеточных стенок, относительные количества лигнина в них и другие родственные темы. Однако заканчивая рассмотрение четвертой и конечной фазы онтогенеза древесной клетки, следует упомянуть о некоторых явлениях, которые тем или иным путем связаны с лигннфикацией, как ее пошшают ботаники. Подобно образованию и разрастанию клеток, а также утолщению клеточных стенок, лигнификация может происходить лишь при жизни клеточного протопласта, так как отмершие клетки не могут лигнифи-цировать свои стенки. Процесс лигнификации может быть закончен в слое межклеточного вещества и в первичной стенке, но может продолжаться во вторичной стенке, даже если этот названный последним слой еще центростремительно увеличивается в толщину. В древесине деревьев лигнификация часто очень скоро заканчивается в слое, примыкающем к внутренней стороне камбия, обычно почти одновременно с тем, когда новые клетки достигли своего наибольшего размера, а вторичные стенки - своей конечной толщины. Это объясняет, почему заболонь при одинаковом содержании влаги так же или почти так же крепка, как ядровая древесина.[ ...]
Детальное исследование распределения лигнина и полисахаридов в одревесневших клеточных стенках древесины ели и березы измерением интенсивности абсорбции тонкого пучка ультрафиолетовых лучей при прохождении их через прозрачный срез подтвердило преимущественное расположение лигнина в срединной пластинке и первичной стенке, а также частично в наружных слоях вторичной стенки . В срединной пластинке еловой древесины содержание лигнина достигает 73%, а во вторичной стенке - не более 16%. Отсюда следует, что полисахариды сосредоточены в основном во вторичном слое. Была сделана попытка измерить этим методом взаимное расположение целлюлозы и гемицеллюлоз. Для этого полисахариды вначале были превращены в окрашенные соединения, абсорбирующие свет.[ ...]
В большинстве клеток ясно различаются чередующиеся зоны большего или меньшего отложения лигнина, которые создают видимость концентрических колец. При противоположном процессе, когда клеточная стенка обрабатывается делигнифицирующими. реагентами, рисунок целлюлозы остается прежним. Это свидетельствует о том, что существуют, по-видимому, две взаимопроникающие системы, состоящие одна из целлюлозы и других полисахаридов, а другая из лигнина. Бейли и Керр показали, что размеры частиц доходят до 0,1 ¡х и меньше. Промежутки или полосы объясняют существование относительно больших «фибрилл», замеченных некоторыми исследователями . Кроме преобладающих концентрических рисунков, в волокнах некоторых видов древесины проявляется расположение радиальных линий или комбинация обоих типов. Клетки сжатой древесины часто имеют жесткие, почти твердые полосы лигнина рядом с полостью клетки и радиально-расположенные пластинки его, отделенные зонами полисахаридного вещества, в средней части стенки клетки.[ ...]
В состав лишайников входят многие элементы и вещества. Все их можно разделить на две большие группы - первичные и вторичные. К первичным относятся те вещества, которые непосредственно принимают участие в клеточном обмене веществ; из них построено тело лишайников. К вторичным относятся конечные продукты обмена веществ, располагающиеся обычно на стенках гиф. Многие из этих вторичных лишайниковых веществ (в более старой литературе их называли лишайниковыми кислотами) специфичны для лишайников и не встречаются в организмах из других систематических групп.[ ...]
Риттер , Людтке и др. сообщили, что при обработке древесных волокон различными реагентами, вызывающими набухание, вторичная стенка (а также, вероятно, и первичная) распадается на нитеобразные фрагменты или фибриллы. Риттер разделил эти фибриллы на веретенообразные тела, а их в свою очередь, на сферические единицы . Значение таких относительно крупных структурных единиц (длина веретенообразных тел примерно 4[х) неясно, ввиду описанной выше тонкопористой структуры вторичной стенки. Ни в остатках лигнина после растворения целлюлозы, ни в остатках целлюлозы после растворения лигнина не обнаруживается заметных промежутков, указывающих на границы названных единиц клеточных стенок. Кроме того, недавно проведенными исследованиями с помощью электронного микроскопа в структуре клеточных стенок не было установлено присутствия подобных сравнительно крупных единиц.[ ...]
При оценке действия различных дереворазрушающих грибов на растительную ткань необходимо учитывать, что отдельные гифы их. движутся в толще клеточных стенок избирательно. Так, грибы белой гнили предпочитают срединную пластинку и первичную оболочку, где сосредоточен главным образом лигнин. Грибы красной или бурой гнили, наоборот, предпочитают проходить по вторичной оболочке, наиболее богатой углеводами. Соответственно различается и окраска поврежденной ими древесины. Более подробно эти вопросы будут рассмотрены в дальнейшем.[ ...]
Исследования трахеид и либриформа с помощью поляризационного и электронного микроскопа, а также рентгенографии позволили установить существование в клеточных стенках пяти концентрических слоев : наружной, или первичной, стенки и вторичной стенки. Вторичная стенка в свою очередь разделяется на три слоя, обычно обозначаемых 81, вг и Бз. Кроме того, между первичными стенками соседних клеток располагается склеивающая их срединная пластинка (рис. 35).[ ...]
Повышение выходов при использовании водяного пара объясняется тем, что ускоряется вынос ценных продуктов из реакционного пространства и задерживается развитие реакций вторичного распада. Кроме того, при соприкосновении водяного пара с капиллярной системой древесины на поверхностных слоях ее возможна конденсация пара, что создает условия для термического разложения в кислой водной среде. При этом реакции разложения происходят в первую очередь в слоях клеточной стенки, которые расположены с внутренних сторон клеточных полостей и состоят преимущественно из нетермостойких гемицеллюлоз, легко отщепляющих ацетильные группы и часть связанных с ними метоксилов, образуя соответственно уксусную кислоту и метиловый спирт.[ ...]
Вряд ли правильно называть клетками сегменты, слагающие нити сфероплеи, и не только потому, что они обладают множеством ядер и хлоропластов (и, следовательно, являются явно вторичными образованиями), но и потому, что отделяющие их поперечные перегородки не похожи на клеточные стенки других многоклеточных зеленых водорослей. Они сильно варьируют по форме, а также по способу и месту образования (рис. 226, 4-6). Часто поперечные перегородки имеют вид кольцевых внутренних утолщений на стенках клетки, которые не смыкаются в центре, так что остается отверстие, через которое проходит цитоплазматический тяж (рис. 226, 4). В других случаях вместо перегородок образуются особые пробки. И, наконец, в любом месте нити могут возникать группы радиально сходящихся тяжей, напоминающих скелетные тяжи каулерпы и играющих механическую роль.[ ...]
Снаружи от плазматической мембраны их клеток нет дополнительной плотной клеточной стенки или она состоит из хитина, редко из целлюлозы. Запасные углеводы обычно в форме гликогена (животного крахмала).[ ...]
Маркс-Фигипи и Пепцел изучали изменение СП хлопковой целлюлозы на различных стадиях созревания хлопка. Они показали, что вязкость растворов хлопковой целлюлозы снижается через несколько часов после открытия коробочки. Целлюлоза вторичной клеточной стенки в волокнах нераскрывшихся коробочек хлопка при небольшой зрелости (выход целлюлозы-18%) имеет единственный максимум на кривой распределения при СП 14 000. Около 10% материала имеет более низкий молекулярный вес (СП 1500-2500), эта целлюлоза содержится в первичной клеточной стенке.[ ...]
Положение мест образования микрофибрилл но отношению к поверхности мембраны цитоплазмы может быть различно. Так, у бактерий этот процесс протекает в среде, значительно удаленной от поверхности клетки и, следовательно, от мембраны. По-видимому, аналогичным образом синтез протекает и в утолщенных первичных стенках клеток эпидермиса колеоптилей овса, поскольку синтез целлюлозы в этом случае осуществляется равномерно но толщине клеточной стенки . В оболочках асцидий отложение целлюлозы происходит, по-видимому, также в местах, удаленных от поверхности секреторных клеток , хотя достаточно убедительных доказательств этого предположения нет. Напротив, микрофибриллы вторичных стенок клеток растений, возможно, образуются на внутренней поверхности стенки, в непосредственной близости от мембраны цитоплазмы . Поскольку целлюлозы во вторичных стенках значительно больше, чем в первичных, можно сделать вывод, что большинство целлюлозных микрофибрилл образуется вблизи мембраны цитоплазмы . Однако это не является обязательным.[ ...]
Одним из методов, основанных на этом принципе, является метод определения реакционной способности целлюлозы по картине набухания ксантогенатов в изо-пропилОвом спирте. Процесс набухания при взаимодействии волокна с растворителем схематически можно представить следующим образом: жидкость проникает внутрь волокна, вследствие чего объем волокна увеличивается. Затем происходит разрыв слабого эластичного наружного слоя вторичной клеточной стенки волокна и в местах разрыва образуются вздутия («бусы»). Остатки этого слоя образуют на набухшем волокне перетяжки и манжеты. Затем наружный слой отделяется и волокно равномерно набухает, на нем образуются поперечные полосы и волокно делится на пакеты дисков и отдельные диски, которые в дальнейшем растворяются.[ ...]
Зависимость прочности древесины от содержания влаги. Так как прочность и жесткость древесины частично определяются силами сцепления, связывающими молекулы, то любой агент, уменьшающий эти силы, меняет ее прочность в целом. Одним из таких агентов является вода, поэтому прочность древесины увеличивается по мере уменьшения содержания влаги не только в результате повышенной плотности, происшедшей от усушки, но также из-за присутствия вторичных валентных сил сцепления1. Так как присутствие воды в количестве, превышающем точку насыщения волокна, не изменяет характера клеточной стенки, то потеря или приобретение капиллярной (свободной) воды практически не влияет на показатели прочности древесины.[ ...]
Структуры, содержащие много лигнина, окрашиваются в темно-коричневый цвет до черного, тогда как слабо лигнифицированные зоны окрашиваются в светло-желтый цвет до янтарного. Результаты этой цветной реакции полностью подтверждают предшествующие работы по исследованию химии клеточной стенки. Вторичные стенки волокнистых элементов у древесины лиственных пород, растущих в умеренном климате, более светлые, следовательно, они менее лигнифицированы, чем вторичные стенки хвойных пород. Стенки сосудов у лиственных пород окрашены в более темный цвет, чем окружающие волокнистые элементы, следовательно, они содержат больше лигнина; мембраны пор также сильно лигнифицированы .[ ...]
Эта операция осуществлялась на одревесневших срезах, предварительно освобожденных от лигнина с помощью хлорита натрия в уксуснокислой среде. Затем срезы были обработаны п-фенилаз-; бензоилхлоридом с целью этерификации полисахаридов. Ярко окрашенные в оранжево-красный цвет срезы после набухания в пиридине фотометрировались. Подвергая такой обработке срезы, со стоящие из холоцеллюлозы, до и после удаления гемицеллюлоз, удалось установить, что основная масса гемицеллюлоз в древесине ели и березы сосредоточена в наружных слоях вторичной стенки. Так, при экстракции среза еловой холоцеллюлозы 16%-ньш едким натром было установлено, что из наружных слоев клетки извлекается до 60-80%, из средины клеточной стенки около 50% и из слоя Бз только 16% растворимых в щелочи гемицеллюлоз от общего количества полисахаридов. Аналогичная картина наблюдалась и для поперечных срезов либриформа из древесины березы.[ ...]
Опыты Риттера , а позднее Бейли и др. показали, что независимо от возможного присутствия пектиновых полиуронидов в срединной пластинке,она состоит главным образом из лигнина, как его понимают химики (нерастворим в холодной 72%-ной серной кислоте, растворим после хлорирования и обработки слабыми основаниями или основными солями). Кроме того, Риттер доказал, что большая часть лигнина находится именно в этом слое. Это утверждение противоречило преобладавшему в то время мнению о присутствии большей части лигнина в других слоях, особенно во вторичной стенке. Позднее было доказано , что в таких случаях кажущаяся широкой и объемистой вторичная стенка в действительности подобна паутине, которая после высыхания съеживается и превращается в разрозненные кусочки. Исли первичные стенки включены в сложную срединную пластинку, то весьма вероятно, что здесь находится и большая часть лигнина.[ ...]
Кальциевые каналы обнаружены и в мембранах растительных клеток. Показана регуляция входа 45Са2+ микросомы, выделенные из колеоп-тилей кукурузы и гипокотилей тыквы, светом, ПУК и зависимость этой реакции от кальмодулина. Для функционирования потенциалзависимых Са2+-каналов (харовая водоросль Ыие11ор,ш) необходимо наличие М§2+. Состояние этих потенциалзависимых каналов контролируется системой ферментов, рейдирующих уровень цАМФ в клетке. Были также получены данные, свидетельствующие о прямом действии экзогенного цАМФ на поглощение 45Са2+ в клетках СМатуёотопт гетскагсШ (мутант без клеточной стенки). Данные, приведенные на рис. 4.1, свидетельствуют о регуляторном действии цАМФ на поглощение Са2+ клетками. Это указывает на возможность взаиморегуляции двух систем вторичных посредников - цАМФ и Са2+. В опытах с животными клетками усиление поглощения Са2+ под действием цАМФ объясняется фосфорилированием белков потенциалзависимых Са2+-каналов и вследствие этого увеличением пребывания их в открытом состоянии.[ ...]
Изучению действия ультразвука на целлюлозные волокна посвящено много исследований . Некоторые исследователи сопоставляли или сочетали влияние ультразвука с различными механическими воздействиями. Так, Яйме, Кронерт и Нейхауз изучали действие ультразвука на целлюлозные волокна по сравнению с высокочастотными механическими колебаниями и показали, что ультразвук с частотой 20-3000 кгц разрыхляет структуру волокна, увеличивает степень его набухания и обезвоживания. Механическая прочность бумаги, изготовленной из таких целлюлоз, повышается, особенно прочность к раздиранию. Аналогично действуют и высокочастотные механические колебания. Ивасаки, Линдберг и Мейер считают, что общая картина изменений структуры волокна под действием ультразвука в водной среде сходна с изменениями структуры волокон при механическом размоле. При этом происходят глубокие изменения морфологической структуры волокон, приводящие к сдвигам во вторичной клеточной стенке, отрыву крупных кусков от первичной стенки, затем к набуханию вторичной стенки и ее дефибриллированию. В работе Сафоновой и Клен-ковой при изучении микрофотографий волокон, подвергнутых ультразвуковому воздействию в воде, показано, что имеются и другие, более глубокие нарушения в структуре волокна, которое становится пронизанным целой сетью многочисленных поперечных каналов. Отмечается , что волокна ранней древесины и волокна, не подвергавшиеся высушиванию, более восприимчивы к действию ультразвука.
Растительные клетки, подобно клеткам прокариот и грибов, заключены в сравнительно жесткую клеточную стенку. Материал для построения этой клеточной стенки секретирует сама заключенная в ней живая клетка (протопласт). По своему химическому составу клеточные стенки растений отличаются от клеточных стенок прокариот и грибов (табл. 2.1), но этим структурам свойственны некоторые общие функции, а именно функции опоры и защиты; кроме того, и те и другие ограничивают подвижность клеток. Клеточная стенка, отлагающаяся во время деления клеток растения, называется первичной клеточной стенкой. Позже в результате утолщения она может превратиться во вторичную клеточную стенку. В этом разделе мы опишем процесс образования первичной клеточной стенки. На рис. 7.21 воспроизведена электронная микрофотография, на которой можно видеть одну из ранних стадий этого процесса.
Строение клеточной стенки
Первичная клеточная стенка состоит из целлюлозных микрофибрилл, погруженных в матрикс, в состав которого входят сложные полисахариды. Целлюлоза тоже представляет собой полисахарид (ее химическое строение описано в разд. 5.2.3). Особо важное значение для той роли, которую целлюлоза выполняет в клеточных стенках, имеют ее волокнистое строение и высокая прочность на разрыв, сравнимая с прочностью стали. Отдельные молекулы целлюлозы - это длинные полисахаридные цепи. Множество таких молекул, сшитых друг с другом поперечными водородными связями, собраны в прочные пучки, называемые микрофибриллами . Погруженные в матрикс микрофибриллы образуют каркас клеточной стенки. Матрикс клеточной стенки состоит из полисахаридов, которые для удобства описания делят обычно на пектины и гемицеллюлозы в зависимости от их растворимости в различных растворителях, употребляемых для экстракции. Пектины , или пектиновые вещества , при экстракции обычно выделяются первыми, поскольку их растворимость выше. Это - смешанная группа кислых полисахаридов (построенных из моносахаридов арабинозы и галактозы, галактуроновой кислоты, принадлежащей к классу сахарных кислот, и метанола). Длинные молекулы пектиновых веществ могут быть линейными или разветвленными. Срединная пластинка , скрепляющая стенки соседних клеток, состоит из клейких студнеобразных пектатов магния и кальция. В клеточных стенках некоторых созревающих плодов нерастворимые пектиновые вещества превращаются снова в растворимые пектины. При добавлении сахара эти последние образуют гели; поэтому их используют как желирующие вещества.
Гемицеллюлозы - это смешанная группа полисахаридов, растворимых в щелочах (к ним относятся полимеры ксилозы, галактозы, маннозы, глюкозы и глюкоманнозы). У гемицеллюлоз, как и у целлюлозы, молекулы имеют форму цепи, однако их цепи короче, менее упорядочены и сильнее разветвлены.
Клеточные стенки гидратированы: 60-70% их массы обычно составляет вода. По свободному пространству клеточной стенки вода перемещается беспрепятственно. Присутствие воды оказывает влияние на химические и физические свойства полисахаридов клеточной стенки.
Материалы с повышенной механической прочностью, подобные материалу клеточной стенки, т. е. состоящие более чем из одного компонента, называются композиционными материалами или композитами ; их прочность обычно выше, чем у каждого из компонентов в отдельности. Системы из волокон и матрицы (в технике основу композиционного материала называют не матриксом, а матрицей. - Прим. перев) находят широкое применение в технике, так что на изучение их свойств как в технике, так и в биологии тратится много усилий. Матрица, работающая на сжатие, передает напряжение волокнам, работающим на растяжение. Она же обеспечивает абразивную стойкость и, по-видимому, стойкость к неблагоприятным химическим воздействиям, возможным в тех или иных условиях. В строительном деле издавна применяется железобетон, т. е. сочетание бетона со стальной арматурой. Позже появился более легкий композиционный материал, в котором роль матрицы играет пластик, а роль арматуры - стеклянное или углеродное волокно. Древесина представляет собой композиционный материал; своей прочностью она обязана клеточным стенкам. Примером жестких композиционных материалов биологического происхождения могут также служить кость, хрящ и покрывающая экзоскелет членистоногих кутикула. Существуют и гибкие композиционные материалы, например соединительная ткань.
У некоторых клеток, например у клеток мезофилла листа, на всем протяжении их жизни имеется только первичная клеточная стенка. Однако у большинства клеток на внутреннюю поверхность первичной клеточной стенки (кнаружи от плазматической мембраны) отлагаются дополнительные слои целлюлозы, т. е. возникает вторичная клеточная стенка. Обычно это происходит после того, как клетка достигнет своего максимального размера, и лишь немногие клетки, например клетки колленхимы, продолжают рост во время этой фазы. Вторичное утолщение клеточных стенок растения не следует путать с вторичным утолщением (вторичным ростом) самого растения, т. е. с увеличением толщины ствола в результате добавления новых клеток.
В любом слое вторичного утолщения целлюлозные волокна располагаются под одним и тем же углом, но в разных слоях этот угол различен, чем и обеспечивается еще большая прочность структуры. Такое расположение целлюлозных волокон показано на рис. 7.27.
Некоторые клетки, такие, как трахеальные элементы ксилемы и клетки склеренхимы, претерпевают интенсивную лигнификацию (одревеснение); при этом все слои целлюлозы (первичный и три вторичных) пропитываются лигнином - сложным полимерным веществом, не относящимся к полисахаридам. В клетках протоксилемы отложения лигнина имеют кольцевую, спиральную или сетчатую форму, как это видно на рис. 8.11. В других случаях лигнификация бывает сплошной, если не считать так называемых поровых полей, т. е. тех участков в первичной клеточной стенке, через которые осуществляются контакт между соседними клетками при помощи группы плазмодесм (разд. 8.1.3 и рис. 8.7). Лигнин скрепляет целлюлозные волокна и удерживает их на месте. Он действует как очень твердый и жесткий матрикс, усиливающий прочность клеточных стенок на растяжение и в особенности на сжатие (предотвращает прогибы). Он же обеспечивает клеткам дополнительную защиту от неблагоприятных физических и химических воздействий. Вместе с целлюлозой, остающейся в клеточных стенках, лигнин придает древесине те особые свойства, которые делают ее незаменимым строительным материалом.
Функции клеточной стенки
Ниже перечисляются основные функции клеточных стенок растений.
1. Клеточные стенки обеспечивают отдельным клеткам и растению в целом механическую прочность и опору. В некоторых тканях прочность усиливается благодаря интенсивной лигнификации клеточных стенок (небольшое количество лигнина присутствует во всех клеточных стенках).
2. Относительная жесткость клеточных стенок и сопротивление растяжению обусловливают тургесцентность клеток, когда в них осмотическим путем поступает вода. Это усиливает опорную функцию во всех растениях и служит единственным источником опоры для травянистых растений и для таких органов, как листья, т. е. там, где отсутствует вторичный рост. Клеточные стенки также предохраняют клетки от разрыва в гипотонической среде.
3. Ориентация целлюлозных микрофибрилл ограничивает и в известной мере регулирует как рост, так и форму клеток, поскольку от расположения этих микрофибрилл зависит способность клеток к растяжению. Если, например, микрофибриллы располагаются поперек клетки, окружая ее как бы обручами, то клетка, в которую путем осмоса поступает вода, будет растягиваться в продольном направлении.
4. Система связанных друг с другом клеточных стенок (апопласт ) служит главным путем, по которому передвигаются вода и минеральные вещества. Клеточные стенки скреплены между собой с помощью срединных пластинок. В стенках имеются небольшие поры, сквозь которые проходят цитоплазматические тяжи, называемые плазмодесмами . Плазмодесмы связывают живое содержимое отдельных клеток - объединяют все протопласты в единую систему, в так называемый симпласт .
5. Наружные клеточные стенки эпидермальных клеток покрываются особой пленкой-кутикулой, состоящей из воскообразного вещества кутина, что снижает потери воды и уменьшает риск проникновения в растение болезнетворных организмов. В пробковой ткани клеточные стенки по завершении вторичного роста пропитываются суберином, выполняющим сходную функцию.
6. Клеточные стенки сосудов ксилемы, трахеид и ситовидных трубок (с ситовидными пластинками) приспособлены для дальнего транспорта веществ по растению. Этот вопрос рассматривается в гл. 8 и 14.
7. Стенки клеток эндодермы корня пропитаны суберином и поэтому служат барьером на пути движения воды (разд. 14.1.5).
8. У некоторых клеток их видоизмененные стенки хранят запасы питательных веществ; таким способом, например, запасаются гемицеллюлозы в некоторых семенах.
9. У передаточных клеток площадь поверхности клеточных стенок увеличена и соответственно увеличена площадь поверхности плазматической мембраны, что повышает эффективность переноса веществ путем активного транспорта (разд. 14.8.6).
Образование клеточной стенки начинается во время деления клетки. В плоскости деления образуется клеточная пластинка, единый слой, общий для двух дочерних клеток. Она состоит из пектиновых веществ, имеющих полужидкую консистенцию; целлюлоза отсутствует. Во взрослой клетке клеточная пластинка сохраняется, но претерпевает изменения, поэтому ее называют срединной , или межклеточной пластинкой (межклеточным веществом) (рис. 2.16 ). Срединная пластинка обычно очень тонка и почти неразличима.
Сразу после образования клеточной пластинки протопласты дочерних клеток начинают откладывать собственную клеточную стенку. Она откладывается изнутри как на поверхности клеточной пластинки, так и на поверхности других клеточных стенок, принадлежавших ранее материнской клетке. После деления клетка вступает в фазу роста растяжением, который обусловлен интенсивным осмотическим поглощением клеткой воды, связанным с образованием и ростом центральной вакуоли. Тургорное давление начинает растягивать стенку, но она не рвется благодаря тому, что в нее постоянно откладываются новые порции микрофибрилл и веществ матрикса. Отложение новых порций материала происходит равномерно по всей поверхности протопласта, поэтому толщина клеточной стенки не уменьшается.
Стенки делящихся и растущих клеток называют первичными . Они содержат много (60-90%) воды. В сухом веществе преобладают матричные полисахариды (60-70%), содержание целлюлозы не превышает 30%, лигнин отсутствует. Толщина первичной стенки очень невелика (0,1-0,5 мкм).
Для многих клеток отложение клеточной стенки прекращается одновременно с прекращением роста клетки. Такие клетки окружены тонкой первичной стенкой до конца жизни ( рис. 2.16).
Рис. 2.16. Паренхимная клетка с первичной стенкой.
У других клеток отложение стенки продолжается и по достижении клеткой окончательного размера. При этом толщина стенки увеличивается, а объем, занимаемый полостью клетки, сокращается. Такой процесс носит название вторичного утолщения стенки, а саму стенку называют вторичной (рис. 2.17 ).
Вторичная стенка может рассматриваться как дополнительная, выполняющая главным образом механическую, опорную функцию. Именно вторичная стенка ответственна за свойства древесины, текстильного волокна, бумаги. Вторичная стенка содержит значительно меньше воды, чем первичная; в ней преобладают микрофибриллы целлюлозы (40-50% от массы сухого вещества), которые располагаются параллельно друг другу. Из полисахаридов матрикса характерны гемицеллюлозы (20-30%), пектиновых веществ очень мало. Вторичные клеточные стенки, как правило, подвергаются одревеснению. В неодревесневших вторичных стенках (лубяные волокна льна, волоски хлопчатника) содержание целлюлозы может достигать 95%. Большое содержание и строго упорядоченная ориентация микрофибрилл определяют высокие механические свойства вторичных стенок. Часто клетки, имеющие вторичную одревесневшую клеточную стенку, после завершения вторичного утолщения отмирают.
Срединная пластинка склеивает соседние клетки. Если ее растворить, стенки клеток теряют связь друг с другом и разъединяются. Этот процесс называется мацерация . Довольно обычна естественная мацерация, при которой пектиновые вещества срединной пластинки переводятся в растворимое состояние с помощью фермента пектиназы и затем вымываются водой (перезрелые плоды груши, дыни, персика, банана). Часто наблюдается частичная мацерация, при которой срединная пластинка растворяется не по всей поверхности, а лишь в углах клеток. Вследствие тургорного давления соседние клетки в этих местах округляются, в результате чего образуются межклетники (рис. 2.16 ). Межклетники образуют единую разветвленную сеть, которая заполняется парами воды и газами. Таким образом, межклетники улучшают газообмен клеток.
Характерная особенность вторичной стенки – ее неравномерное отложение поверх первичной стенки, в результате чего во вторичной стенке остаются неутолщенные участки – поры . Если вторичная стенка не достигает большой толщины, поры выглядят как мелкие углубления. У клеток с мощной вторичной стенкой поры в разрезе имеют вид радиальных каналов, идущих от полости клетки до первичной стенки. По форме порового канала различают поры двух типов – простые и окаймленные (рис. 2.17 ).
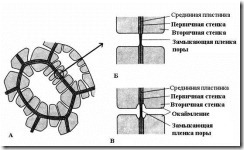
Рис. 2.17. Типы пор : А – клетки с вторичными стенками и многочисленными простыми порами; Б – пара простых пор; В – пара окаймленных пор.
У простых пор диаметр порового канала по всей длине одинаковый и имеет форму узкого цилиндра. Простые поры характерны для паренхимных клеток, лубяных и древесинных волокон.
Поры в двух смежных клетках, как правило, возникают друг против друга. Эти общие поры имеют вид одного канала, разделенного тонкой перегородкой из срединной пластинки и первичной стенки. Такая совокупность двух пор смежных стенок соседних клеток носит название пары пор и функционирует как одно целое. Разделяющий их канал участок стенки называется замыкающей пленкой поры , или поровой мембраной . В живых клетках замыкающая пленка поры пронизана многочисленными плазмодесмами (рис. 2.18 ).
Плазмодесмы присущи только растительным клеткам. Они представляют собой тяжи цитоплазмы, пересекающие стенку смежных клеток. Число плазмодесм в одной клетке очень велико – от нескольких сотен до десятков тысяч, обычно плазмодесмы собраны в группы. Диаметр плазмодесменного канала составляет 30-60 нм. Его стенки выстланы плазмалеммой, непрерывной с плазмалеммой смежных клеток. В центре плазмодесмы проходит мембранный цилиндр – центральный стержень плазмодесмы , непрерывный с мембранами элементов эндоплазматической сети обеих клеток. Между центральным стержнем и плазмалеммой в канале находится гиалоплазма, непрерывная с гиалоплазмой смежных клеток.
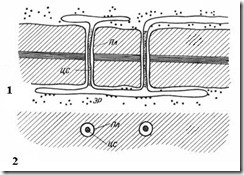
Рис. 2.18. Плазмодесмы под электронным микроскопом (схема ): 1 – на продольном срезе; 2 – на поперечном срезе; Пл – плазмалемма; ЦС – центральный стержень плазмодесмы; ЭР – элемент эндоплазматического ретикулума.
Таким образом, протопласты клеток не полностью изолированы друг от друга, а сообщаются по каналам плазмодесм. По ним происходит межклеточный транспорт ионов и мелких молекул, а также передаются гормональные стимулы. Посредством плазмодесм протопласты клеток в растительном организме образуют единое целое, называемое симпластом , а транспорт веществ через плазмодесмы получил название симпластического в отличие от апопластического транспорта по клеточным стенкам и межклетникам.
У окаймленных пор (рис. 2.17 )канал резко суживается в процессе отложения клеточной стенки, поэтому внутреннее отверстие поры, выходящее в полость клетки, гораздо уже, чем наружное, упирающееся в первичную стенку. Окаймленные поры характерны для рано отмирающих клеток водопроводящих элементов древесины. У них поровый канал по направлению к замыкающей пленке воронковидно расширяется, а вторичная стенка нависает в виде валика над расширенной частью канала, образуя камеру поры. Название окаймленной поры происходит оттого, что при рассмотрении с поверхности внутреннее отверстие имеет вид маленького круга или узкой щели, тогда как наружное отверстие как бы окаймляет внутреннее в виде круга большего диаметра или более широкой щели.
Поры облегчают транспорт воды и растворенных веществ от клетки к клетке, в то же время не снижая прочности клеточной стенки.