Actinomycetes (Actinomycetales មកពីភាសាក្រិច aktis - កាំរស្មី mykes - ផ្សិត) គឺជាបាក់តេរីដែលបែកខ្ញែកជាកម្មសិទ្ធិរបស់ phylum Actinobacteria ។ ពួកវាជាផ្នែកមួយនៃ microflora ធម្មតា។ ប្រព័ន្ធរំលាយអាហារសត្វឆ្អឹងខ្នង និងសត្វឆ្អឹងខ្នង ហើយក៏មានវត្តមាននៅក្នុងដីដ៏សម្បូរបែប និងដើរតួយ៉ាងសំខាន់ក្នុងបរិស្ថានវិទ្យា និងវដ្តនៃសារធាតុនៅក្នុងដី។
អតិសុខុមប្រាណទាំងនេះគឺជាភ្នាក់ងារមូលហេតុនៃរោគសាស្ត្រឱកាសនិយមជាច្រើន - ដែលកើតឡើងជាលទ្ធផលនៃការថយចុះមុខងារ។ ប្រព័ន្ធភាពស៊ាំរាងកាយ។ Actinomycetes ត្រូវបានគេប្រើយ៉ាងទូលំទូលាយក្នុងបច្ចេកវិជ្ជាជីវសាស្ត្រ ព្រោះវាជាប្រភពនៃសារធាតុប្រឆាំងនឹងបាក់តេរី និងសារធាតុប្រឆាំងដុំសាច់។
អង្ករ។ 1. Streptomycetes សំយោគបរិមាណដ៏ច្រើននៃថ្នាំប្រឆាំងនឹងបាក់តេរី និងថ្នាំប្រឆាំងដុំសាច់។
រចនាសម្ព័ន្ធនៃ actinomycetes: ហេតុអ្វីបានជាពួកវាជាបាក់តេរីហើយមិនមែនជាផ្សិត?
1. ការរៀបចំសម្ភារៈហ្សែន
សម្ភារៈតំណពូជរបស់ Actinomycetes មាននៅក្នុងម៉ូលេគុលមួយនៃអាស៊ីត deoxyribonucleic ដែលមានរាងជារង្វង់ ហើយមានទីតាំងនៅដោយសេរីនៅក្នុង cytoplasm - ទម្រង់ដូចគ្នានៃការរៀបចំសម្ភារៈហ្សែនដែលហៅថា nucleoid គឺជាលក្ខណៈនៃបាក់តេរីផ្សេងទៀត។ នៅក្នុងផ្សិតសម្ភារៈហ្សែនត្រូវបានរៀបចំនិងរួមបញ្ចូលនៅក្នុង ស្នូលកោសិកា.
Actinomycete DNA មាន មួយចំនួនធំនៃគូ GC (65-75% នៃចំនួនសរុបនៃ nucleotides) ។ លក្ខណៈនេះគឺថេរ មិនអាស្រ័យលើការបំប្លែងទេ ហើយដូច្នេះវាត្រូវបានគេប្រើនៅក្នុងការធ្វើវចនានុក្រមនៃអតិសុខុមប្រាណ។ ខ្លឹមសារនៃគូ GC នេះធ្វើឱ្យ DNA នៃ actinomycetes មានភាពច្របូកច្របល់ខ្លាំង ដូច្នេះការវិភាគ DNA នៃ actinomycetes ត្រូវការពេលវេលាច្រើនជាងបើប្រៀបធៀបទៅនឹងបាក់តេរីដទៃទៀត។

អង្ករ។ ២. រចនាសម្ព័ន្ធគ្រោងការណ៍ជញ្ជាំងកោសិកានៃបាក់តេរី Gr+ ។

អង្ករ។ 3. Actinomycetes, Gram ប្រឡាក់។
Actinomycetes មានជញ្ជាំងកោសិកាបាក់តេរីក្រាស់ដែលមានទីតាំងនៅខាងក្រៅ ភ្នាស cytoplasmicនិងបណ្តាលឱ្យមានស្នាមប្រឡាក់ Gram វិជ្ជមានរបស់ពួកគេ។ ដូចបាក់តេរី Gram-positive ផ្សេងទៀតដែរ វាមានស្រទាប់ជាច្រើននៃ murein polymer (peptidoglycan) ដែលត្រូវបានជ្រាបចូលទៅក្នុងអាស៊ីត teichoic និង lipoteichoic ។ អាស៊ីត Lipoteichoic ត្រូវបានបោះយុថ្កានៅក្នុងភ្នាស cytoplasmic នៃបាក់តេរី ហើយភ្ជាប់វាទៅនឹងជញ្ជាំងកោសិកា។ អាស៊ីត Teichoic ចូលទៅក្នុងជញ្ជាំងកោសិកា បន្ទុកអវិជ្ជមាន. ជញ្ជាំងកោសិកានៃផ្សិតមានសារធាតុប៉ូលីម៊ែរផ្សេងទៀត - chitin និង glucan ។
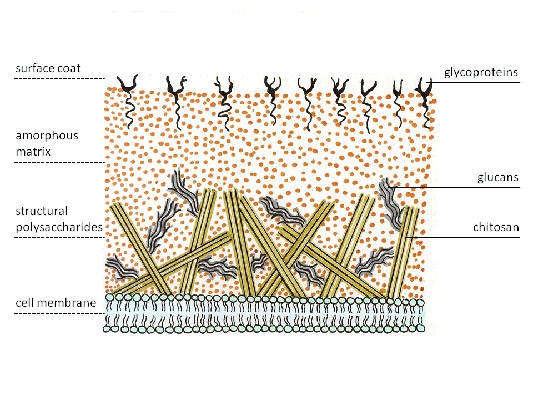
អង្ករ។ 4. ជញ្ជាំងកោសិកាផ្សិត។
3. សរីរាង្គកោសិកា
Actinomycetes ដូចជាបាក់តេរីផ្សេងទៀតមិនមានទេ។ សរីរាង្គភ្នាស. Actinomycetes មាន 70S ribosomes ខណៈពេលដែលផ្សិតមាន 80S ribosomes ដូចទៅនឹងសារពាង្គកាយ eukaryotic ដទៃទៀតដែរ។
4. ការរីកចម្រើននៃអាណានិគម
ការបង្កើត mycelium ក្នុងអំឡុងពេលលូតលាស់គឺជាអ្វីដែល actinomycetes ស្រដៀងនឹងផ្សិត។ mycelium ក្នុងករណី actinomycetes គឺជាបណ្តុំនៃ hyphae ។ hyphae ត្រូវបានបែងចែកដោយ Septa ទៅជាកោសិកាបាក់តេរីវែងដែលមាន nucleoids ជាច្រើន។ ភាគថាសនៅក្នុងប្រភេទមួយចំនួនអាចដំណើរការក្នុងទិសដៅកាត់កែងគ្នាទៅវិញទៅមក។ សាខា Hyphae ដោយ budding ។
Mycelium លូតលាស់ទៅក្នុងស្រទាប់ខាងក្រោម (ដី ដីល្បាប់ ឬ សារធាតុចិញ្ចឹមមធ្យម) ដែលហៅថាស្រទាប់ខាងក្រោម។ វាផ្តល់សារធាតុចិញ្ចឹមដល់អាណានិគម។ mycelium ពីលើអាកាសកើនឡើងពីលើស្រទាប់ខាងក្រោមដោយផ្តល់ឱ្យអាណានិគមនូវ "ភាពរលោង" - វាបង្កើតជាស្ពែរក៏ដូចជាអ្វីដែលគេហៅថា "មេតាបូលីតបន្ទាប់បន្សំ" (ផ្ទុយទៅនឹង "មេតាបូលីតបឋម" នៃស្រទាប់ខាងក្រោម mycelium) ដែលក្នុងនោះមានច្រើន សារធាតុប្រឆាំងបាក់តេរី។
វដ្តជីវិត និងសរីរវិទ្យានៃ actinomycetes
ក្នុងអំឡុងពេលវដ្តជីវិត ភាគច្រើន actinomycetes បង្កើត spores ។ actinomycetes ខ្លះបន្តពូជដោយការបែកខ្ញែកនៃ mycelium ។
1. Sporulation
Actinomycete spores មានប្រភពចេញពី mycelium ពីលើអាកាស។ ទាំងនេះគឺជា exospores - ពួកវាវិវត្តនៅខាងក្រៅកោសិកាម្តាយ។ hyphae នៃ mycelium ពីលើអាកាសដែល spores លូតលាស់ត្រូវបានគេហៅថា spore carriers ។ spores អាចមាននៅក្នុងការក្រាស់នៅចុងបញ្ចប់នៃ sporangium - sporangium (ឧទាហរណ៍នៅក្នុង streptomycetes, actinoplanes និង plymelia) ឬអាចស្ថិតនៅក្នុងខ្សែសង្វាក់តាមបណ្តោយ sporangium (ឧទាហរណ៍នៅក្នុង nocardia និង actinomadura) ។
យោងតាមចំនួននៃ spores ដែលបានបង្កើតឡើង actinomycetes ត្រូវបានបែងចែកជា:
- Monosporous (ឧទាហរណ៍ Saccaromonospora, Micromonospora, Thermomonospora) - បង្កើត spores តែមួយ, ជាញឹកញាប់ដោយការចេញផ្កានិងការបំបែកជាបន្តបន្ទាប់ពី hypha ម្តាយដោយ septum មួយ;
- Oligosporous (ឧទាហរណ៍ Actinomadura) - បង្កើតជាខ្សែសង្វាក់ខ្លីនៃ spores តាមបណ្តោយអ្នកដឹកជញ្ជូន spores;
- Polysporous (ភាគច្រើន actinomycetes ឧទាហរណ៍ Streptomyces, Frankia, Geodermatophilus) - បង្កើត spores ជាច្រើនដែលរុំព័ទ្ធនៅក្នុង sporangia ។

អង្ករ។ 5. Sporangium នៃ actinomycetes នៃ genus Frankia ។
ស្ពែម Actinomycete អាចជាចលនា - ក្នុងករណីនេះ spores មាន flagellum ហើយអាចផ្លាស់ទី (spores នៃ actinoplanes, geodermatophiles និង dermatophiles) ។ ក្នុងករណីភាគច្រើន ស្ពឺគឺមិនមានចលនាទេ ហើយត្រូវបានរីករាលដាលដោយខ្យល់ ទឹក ឬសត្វ។

អង្ករ។ 6. Dermatophils មីក្រូទស្សន៍ពន្លឺ។
Sporulation នៅក្នុង actinomycetes គឺសកម្មជាពិសេសនៅក្រោមលក្ខខណ្ឌមិនអំណោយផល។ ភាពធន់នៃ spores ទៅនឹងកំដៅគឺទាបបើប្រៀបធៀបទៅនឹង spores នៃបាក់តេរីផ្សេងទៀត ប៉ុន្តែពួកគេអាចទប់ទល់នឹងការស្ងួតមិនអាក្រក់ជាងអ្នកដទៃទេ ហើយដូច្នេះវាមានសារៈសំខាន់ក្នុងការសម្របខ្លួនយ៉ាងសម្បើម។ Actinomycetes គ្រប់គ្រងអតិសុខុមប្រាណដទៃទៀតនៅក្នុងដីវាលខ្សាច់ស្ងួត។
ដំណុះនៃ swarm ទាមទារសំណើមជាក់លាក់មួយ។ បរិស្ថានខាងក្រៅ. នៅក្នុងវត្តមាននៃទឹក, spores ហើម, អង់ស៊ីមត្រូវបានធ្វើឱ្យសកម្មនិងដំណើរការមេតាប៉ូលីសត្រូវបានចាប់ផ្តើម, អមដោយការបញ្ចេញបំពង់លូតលាស់ (សាកសពបាក់តេរីនាពេលអនាគត) និងការសំយោគនៃអាស៊ីត nucleic ។
2. ប្រភេទនៃការដកដង្ហើម
actinomycetes ភាគច្រើនគឺជា aerobes (ពួកគេត្រូវការអុកស៊ីសែនដើម្បីរក្សាជីវិត) ។ facultative anaerobes (បាក់តេរីដែលអាចរស់នៅក្នុងវត្តមាន និងអវត្ដមាននៃអុកស៊ីសែន) ត្រូវបានរកឃើញក្នុងចំណោមប្រភេទសត្វដែលមានដំណាក់កាល mycelial ខ្លី បន្តពូជដោយការបែកខ្ញែកនៃ mycelium ។
3. ធន់នឹងអាស៊ីត
Actinomycetes មានភាពធន់នឹងអាស៊ីត - ភាពធន់នឹងបរិយាកាសអាស៊ីតដែលអនុញ្ញាតឱ្យពួកគេរស់នៅក្នុងដីព្រៃដែលមានជាតិអាស៊ីត។ ភាពធន់នឹងអាស៊ីតនៅក្នុងមន្ទីរពិសោធន៍អាចត្រូវបានកំណត់ដោយការប្រឡាក់នូវការរៀបចំដែលមានសារធាតុ actinomycetes យោងទៅតាម Ziehl-Neelsen (fuchsin បន្តដោយការឆ្លាក់ជាមួយនឹងអាស៊ីតស៊ុលហ្វួរិក និងស្នាមប្រឡាក់ជាមួយនឹងពណ៌ខៀវមេទីល)។ actinomycetes ភាគច្រើនដែលមានពណ៌នេះមិនប្រែពណ៌ទេបន្ទាប់ពីឆ្លាក់ដោយអាស៊ីត និងរក្សាពណ៌ស្វាយក្រហម។ បរិស្ថានអាល់កាឡាំងមិនអំណោយផលសម្រាប់បាក់តេរីទាំងនេះ: នៅកម្រិតខ្ពស់ pH ពួកគេងាយនឹង sporulation ។
4. លក្ខណៈពិសេសនៃការរំលាយអាហារ

អង្ករ។ 7. Aerobic actinomycetes បង្កើតជាសារធាតុពណ៌នៅលើ agar slants ។ ពីឆ្វេងទៅស្តាំ៖ Actinomadura madurae, Nocardia asteroides, Micromonospora ។
ការបង្កើត "មេតាបូលីតបន្ទាប់បន្សំ" ដោយ mycelium ពីលើអាកាសត្រូវបានរៀបរាប់ខាងលើ។ ក្នុងចំណោមពួកគេ:
- សារធាតុពណ៌ដែលបណ្តាលឱ្យមានពណ៌ផ្សេងគ្នានៃ mycelium ពីលើអាកាសនៅពេលរីកលូតលាស់នៅលើប្រព័ន្ធផ្សព្វផ្សាយ;
- សារធាតុក្លិនដែលងាយនឹងបង្កជាហេតុដែលផ្តល់ក្លិនលក្ខណៈដល់ដីបន្ទាប់ពីភ្លៀង ទឹកនៅទ្រឹង និងស្បែករបស់សត្វមួយចំនួន;
- ថ្នាំអង់ទីប៊ីយោទិច៖
ក. ថ្នាំសំលាប់មេរោគ - ប៉ូលីយូធ្យូន;
ខ. ថ្នាំសំលាប់មេរោគ - ឧទាហរណ៍ streptomycin, erythromycin, tetracycline, vancomycin;
គ. ថ្នាំប្រឆាំងនឹងដុំសាច់ - anthracyclines, bleomycin ។
តើ actinomycetes រស់នៅទីណា?
Actinomycetes នៅក្នុង ចំនួនធំបំផុតត្រូវបានរកឃើញនៅក្នុងដី ហើយមានទម្រង់ mycelial តិចជាង spores ។ ពួកវាដើរតួយ៉ាងសំខាន់ក្នុងការបង្កើត humus បំបែកសារធាតុសរីរាង្គដែលពិបាកសម្រាប់បាក់តេរីផ្សេងទៀតក្នុងការប្រើប្រាស់។ ក្នុងន័យនេះ actinomycetes ត្រូវបានគេប្រើជាមីក្រូសរីរាង្គសូចនាករអនាម័យនៅក្នុងបញ្ហាអនាម័យនិងរោគរាតត្បាត: ការរកឃើញរបស់ពួកគេនៅក្នុង បរិមាណដ៏ច្រើន។នៅក្នុងដី ឬទឹកបង្ហាញពីវត្តមានរបស់ជីកំប៉ុសនៅក្នុងស្រទាប់ខាងក្រោមរៀងៗខ្លួន។

អង្ករ។ 8. Actinomycetes នៅក្នុងជីកំប៉ុស។
Actinomycetes គឺជាសារធាតុផ្សំនៃរុក្ខជាតិជាច្រើន ជួយជួសជុលអាសូត។ ក្នុងពេលជាមួយគ្នានោះ microorganisms ជាច្រើននៃថ្នាក់នេះគឺជាភ្នាក់ងារបង្កជំងឺនៃរុក្ខជាតិ។

អង្ករ។ 9. ដំឡូង streptomycosis ។
ពួកគេក៏ត្រូវបានរកឃើញផងដែរនៅក្នុង microflora ធម្មតានៃប្រព័ន្ធរំលាយអាហាររបស់សត្វមួយចំនួនចាប់ពីដី annelids(ឧទាហរណ៍ទឹកភ្លៀង) និងបញ្ចប់ដោយសត្វពាហនៈធំ ៗ ។
អតិសុខុមប្រាណទាំងនេះជួយបំបែកសែលុយឡូសដែលមានច្រើននៅក្នុងអាហាររុក្ខជាតិ។ ចំពោះមនុស្ស សារធាតុ actinomycetes ត្រូវបានរកឃើញនៅក្នុងប្រហោងមាត់ (អញ្ចាញធ្មេញ និងបន្ទះ) ពោះវៀន (ពោះវៀនធំ) ស្បែក (មុខ ច្រមុះ ខាងក្រោយត្រចៀក ចន្លោះម្រាមដៃ) និងក្នុងសរីរាង្គ។ ប្រព័ន្ធផ្លូវដង្ហើម(ជាចម្បងនៅក្នុងផ្លូវដង្ហើមខាងលើ) ។
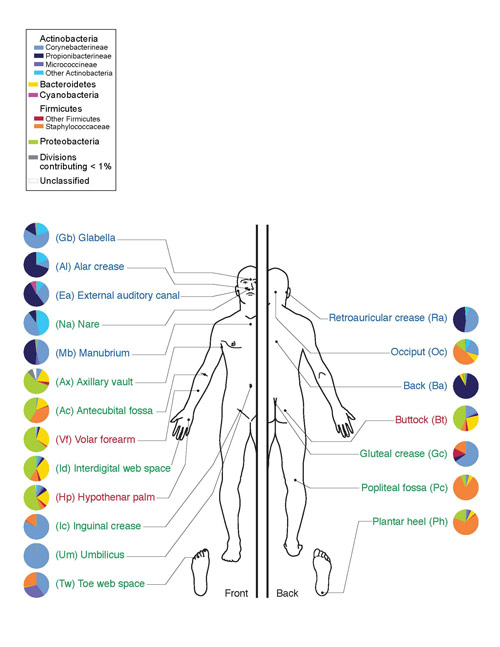
អង្ករ។ 10. microflora នៃស្បែកមនុស្ស។ phylum Actinobacteria ត្រូវបានចង្អុលបង្ហាញជាពណ៌ខៀវ ថ្នាក់ Actinomycetes ត្រូវបានចង្អុលបង្ហាញជាពណ៌ខៀវភ្លឺ។
Actinomycetes ប្រសិនបើប្រតិកម្មនៃភាពស៊ាំរបស់រាងកាយត្រូវបានកាត់បន្ថយអាចបណ្តាលឱ្យ actinomycosis - ជំងឺឱកាសនិយមដែលមាននៅក្នុងការបង្កើត actinomycosis granulomas - ការប្រមូលផ្តុំនៃសាកសពបាក់តេរីដែលស្រដៀងនឹងធញ្ញជាតិនៃស្ពាន់ធ័រពណ៌លឿង ("drusen") ហ៊ុំព័ទ្ធដោយកោសិកា immunocompetent ។ ប្រតិកម្មរលាកនាំទៅដល់ការរលាយនៃ granulomas ការបង្កើត fistulas ដែលនាំឱ្យ perforation នៃសរីរាង្គនិងការរីករាលដាលនៃបាក់តេរីដោយឈាម។

អង្ករ។ 11. Actinomycosis drusen, ស្នាមប្រឡាក់ក្រាម។

អង្ករ។ 12. Actinomycosis នៃថ្គាមខាងលើនៅក្នុងគោមួយ។

អង្ករ។ 13. Human maxillary actinomycosis ។
Actinomycetes - សារពាង្គកាយដ៏អស្ចារ្យនៅតែធ្វើឱ្យអ្នកវិទ្យាសាស្ត្រជាច្រើនយល់ច្រឡំជាមួយនឹងភាពស្រដៀងគ្នាទៅនឹងផ្សិត។ រួមជាមួយនឹងគ្រោះថ្នាក់ដែលអាចកើតមានក្នុងទម្រង់នៃ actinomycosis ឱកាសនិយម សារពាង្គកាយទាំងនេះផ្តល់ឱ្យមនុស្សនូវដីមានជីជាតិ និងអាវុធសម្រាប់ប្រយុទ្ធប្រឆាំងនឹងជំងឺឆ្លង និងជំងឺ oncological - ថ្នាំអង់ទីប៊ីយោទិច និង cytostatics ។
សមាសភាពគីមីនិងរចនាសម្ព័ន្ធនៃជញ្ជាំងកោសិកានៃផ្សិត
ជញ្ជាំងកោសិកានៃផ្សិតមានច្រើនស្រទាប់ ដោយមានស្រទាប់ផ្សេងៗគ្នាដែលបង្កើតឡើងដោយកាបូអ៊ីដ្រាតរចនាសម្ព័ន្ធដែលខុសគ្នានៅក្នុងសមាសធាតុគីមីរបស់ពួកគេ ដែលអាចត្រូវបានបែងចែកជា 3 ក្រុមដោយផ្អែកលើសមាសធាតុគីមីរបស់ពួកគេ៖
ប៉ូលីមែរគ្លុយកូស(glucan, chitin, សែលុយឡូស) ។ Glucans បង្កើតស្រទាប់ខាងក្រៅនៃជញ្ជាំងកោសិកានៃផ្សិតភាគច្រើន។ ស្រទាប់ខាងក្នុងនៃជញ្ជាំងកោសិកាផ្សិតត្រូវបានបង្កើតឡើងដោយច្រវាក់នៃ chitin ដែលផ្តល់ឱ្យវានូវភាពរឹង។ Chitin ជំនួស cellulose ដែលអវត្តមាននៅក្នុងផ្សិតភាគច្រើន ប៉ុន្តែជាផ្នែកមួយនៃជញ្ជាំងកោសិកានៃ oomycetes ដែលបច្ចុប្បន្នមិនមែនជារបស់ផ្សិតធម្មតាទេ។ ឈីទីនដែលខូចត្រូវបានគេហៅថា chitosan ដែលរួមបញ្ចូលគ្នាជាមួយ chitin បង្កើតជាជញ្ជាំងកោសិកានៃ zygomycetes ។
ប៉ូលីមែរនៃ monosaccharides ផ្សេងទៀត (mannose, galactose ជាដើម)ផ្ទុយទៅនឹងរុក្ខជាតិខ្ពស់ជាងដែលពួកគេបង្កើតជាមូលដ្ឋាននៃម៉ាទ្រីសក្រោម ឈ្មោះទូទៅ hemicellulose ដែលមិនសូវមានលក្ខណៈធម្មតាសម្រាប់ផ្សិត។ ករណីលើកលែងមួយគឺផ្សិត ដែលជញ្ជាំងកោសិការបស់វាសម្បូរទៅដោយសារធាតុប៉ូលីម៊ូសដែលហៅថា ម៉ាន់ណាន។ វាត្រូវបានសន្មត់ថាសមាសភាពជញ្ជាំងនេះល្អប្រសើរជាងមុនធានាការចេញផ្កា។
ប៉ូលីមែរនៃកាបូអ៊ីដ្រាតដែលត្រូវបានផ្សារភ្ជាប់ទៅនឹង peptides (glycoproteins)បង្កើតស្រទាប់កណ្តាលនៃជញ្ជាំងកោសិកាពហុស្រទាប់ ហើយលេង តួនាទីសំខាន់ទាំងក្នុងការរក្សាបាននូវភាពសុចរិតនៃរចនាសម្ព័ន្ធនៃកោសិកា និងនៅក្នុងដំណើរការមេតាបូលីសរបស់វាជាមួយនឹងបរិស្ថាន។
លក្ខណៈពិសេសផ្សេងទៀតនៃកោសិកាផ្សិតរួមមាន: អវត្តមាននៃផ្លាស្ទិចដែលនាំវាទៅជិតកោសិកាសត្វ។
គ្មានម្សៅដែលនៅក្នុង eumycetes ត្រូវបានជំនួសដោយ polysaccharide នៅជិត glycogen ម្សៅសត្វ ក្នុង oomycetes ដោយ polysaccharide ជិត laminarin សារាយពណ៌ត្នោត. ចំនួននៃកាបូអ៊ីដ្រាតបម្រុងជាក់លាក់ចំពោះផ្សិតក៏ត្រូវបានផលិតផងដែរ។
ការអភិវឌ្ឍន៍ជាក់លាក់ សារធាតុរំលាយអាហារបន្ទាប់បន្សំ, ក្នុងចំនោមនោះ តួនាទីធំអង់ទីប៊ីយ៉ូទិក phyto- និង mycotoxins និង phytohormones ដើរតួនាទី។
លក្ខណៈជាក់លាក់នៃផ្សិតក៏រួមបញ្ចូលផងដែរ។ heterokaryosis និងដំណើរការ parasexual ។
នៅក្នុងផ្សិត បាតុភូតនៃ heterokaryosis ឬ heteronuclearity គឺរីករាលដាលយ៉ាងខ្លាំង ដែលនៅក្នុងកោសិកាមួយ យូរស្នូលដែលមានលក្ខណៈ heteroalllelic សម្រាប់ហ្សែនមួយចំនួនត្រូវបានរក្សាទុក។ Heterokaryosis ជំនួស heterozygosity នៅក្នុងផ្សិត haploid និងជំរុញការសម្របខ្លួនយ៉ាងឆាប់រហ័សនៃផ្សិតទៅនឹងការផ្លាស់ប្តូរលក្ខខណ្ឌ។ វត្តមាននៃ heteronuclearity គឺដោយសារតែចំនួននៃ លក្ខណៈជាក់លាក់ផ្សិត៖
1. វត្តមាននៃស្នូលច្រើនជាងមួយនៅក្នុងកោសិកាមួយ។
2. រចនាសម្ព័ន្ធជាក់លាក់នៃកោសិកា septum ដែលក្នុងនោះមានមួយ ឬច្រើនតាមរយៈរន្ធ ហៅថា រន្ធញើស ដែលតាមរយៈនោះ ស្នូលអាចធ្វើចំណាកស្រុកពីកោសិកាមួយទៅកោសិកាមួយទៀត។
3. hyphae នៅក្នុងអាណានិគមមួយ និងសូម្បីតែអាណានិគមដែលមានគម្លាតយ៉ាងជិតស្និទ្ធខុសៗគ្នា ដែលដុះចេញពី spores ផ្សេងគ្នានៃផ្សិតប្រភេទដូចគ្នា ជារឿយៗលូតលាស់ជាមួយគ្នា ដែលជាលទ្ធផលដែលការផ្លាស់ប្តូរនៃ nuclei នៃប្រភេទផ្សេងគ្នាគឺអាចធ្វើទៅបាន។
Parasexual (pseudosexual)ដំណើរការ។ ប្រសិនបើនៅក្នុងកោសិកា heterokaryotic nuclei ដែលមាន heteroallelic នៅទីតាំងណាមួយនោះ nucleus diploid heterozygous លេចឡើង។ វាអាចចូលទៅក្នុង spore និងបង្កើតជាក្លូន heterozygous diploid ។ ក្នុងអំឡុងពេលដំណើរការនៃ mitosis ស្នូល diploid អាចត្រលប់ទៅរដ្ឋ haploid ដោយសារតែការបាត់បង់ក្រូម៉ូសូមមួយឈុត ឬការផ្លាស់ប្តូរផ្នែកក្រូម៉ូសូមអាចកើតឡើងនៅក្នុងពួកវា (ការឆ្លងកាត់ mitotic) ។ ដំណើរការទាំងពីរនេះត្រូវបានអមដោយការផ្សំឡើងវិញនៃហ្សែនឪពុកម្តាយ ហើយជាលទ្ធផល phenotypes ។ Parasexual ( asexual ) recombination គឺជាបាតុភូតដ៏កម្រមួយ មិនលើសពីមួយ nucleus ក្នុងមួយលាន ប៉ុន្តែដោយសារតែចំនួន nuclei ដ៏ច្រើននៅក្នុង mycelium វាត្រូវបានគេសង្កេតឃើញជានិច្ចនៅក្នុងពពួកផ្សិត។
ការបន្តពូជ- លូតលាស់, ផ្លូវភេទ, ផ្លូវភេទ។
បន្លែ- ការបែកខ្ចាត់ខ្ចាយនៃ thallus ការបង្កើត chlamydospores ដែលបន្ទាប់ពីរយៈពេលនៃការ dormancy លូតលាស់ទៅជា mycelium, budding នៅក្នុង yeast ។
ផ្លូវភេទការបន្តពូជនៃផ្សិតផ្សេងៗគ្នាអាចត្រូវបានអនុវត្តដោយ spores ចល័ត និង immobile ។ Zoospores បង្កើតជាក្រុមតូចមួយនៃផ្សិត និងសារពាង្គកាយដែលស្រដៀងនឹងផ្សិត - ទឹក និងដីមួយចំនួនដែលក្នុងនោះ ទំនាក់ទំនងហ្សែនជាមួយផ្សិតក្នុងទឹក និងសារាយ។ រចនាសម្ព័ន្ធរបស់ flagella គឺសំខាន់ សញ្ញារោគវិនិច្ឆ័យនៅពេលប្រគល់ឱ្យនគរជាក់លាក់។ ភាគច្រើននៃផ្សិត ការបន្តពូជផ្លូវភេទ spores immobile ត្រូវបានបង្កើតឡើងដែលបង្ហាញពីការលេចឡើងបុរាណបំផុតរបស់ពួកគេនៅលើដី។ យោងទៅតាមកន្លែងនៃការបង្កើតនិងការធ្វើមូលដ្ឋានីយកម្ម sporangiospores endogenous ត្រូវបានសម្គាល់ដែលបង្កើតឡើងនៅក្នុង sporangia និង exogenous (conidia) ការអភិវឌ្ឍនៅលើ hyphae ពិសេស - conidiophores ។ Conidia ត្រូវបានបង្កើតឡើងនៅក្នុងផ្សិតភាគច្រើន (ascomycetes, basidiomycetes, zygomycetes មួយចំនួន) បង្កើតបានជា sporulation conidial ដែលមានភាពចម្រុះខ្លាំង ហើយត្រូវបានគេប្រើយ៉ាងទូលំទូលាយសម្រាប់ការធ្វើរោគវិនិច្ឆ័យផ្សិត។
ការបន្តពូជផ្លូវភេទនៃផ្សិតមានភាពជាក់លាក់របស់វាទាំងនៅក្នុង morphology នៃដំណើរការផ្លូវភេទ និងនៅក្នុងយន្តការនៃហ្សែន និង បទប្បញ្ញត្តិសរីរវិទ្យាយេនឌ័រ និងការបញ្ជូនព័ត៌មានតំណពូជ។
សូម៉ាតូហ្គាមី- ប្រភេទទូទៅបំផុត និងសាមញ្ញបំផុតនៃដំណើរការផ្លូវភេទ រួមមានការបញ្ចូលគ្នានៃ gametes ពីរដែលមិនខុសគ្នា។ កោសិកា somatic. ពេលខ្លះវាកើតឡើងសូម្បីតែដោយគ្មានការលាយកោសិកា - ស្នូលនៅខាងក្នុងកោសិកាបញ្ចូលគ្នា។ ត្រូវបានរកឃើញនៅក្នុង basidiomycetes ភាគច្រើន yeasts marsupial និង taxa មួយចំនួនទៀត។
ល្បែងផ្គុំរូប- gametangia ត្រូវបានញែកដាច់ពីគេនៅលើ haploid mycelium ហើយមាតិការបស់វាត្រូវបានបង្ហូរក្នុងអំឡុងពេលដំណើរការផ្លូវភេទ។ ដំណើរការផ្លូវភេទនេះគឺជាលក្ខណៈនៃផ្សិត marsupial ភាគច្រើន។ វ៉ារ្យ៉ង់នៃ gametangiogamy គឺ zygogamy នៅក្នុង zygomycetes ។
ការលេងហ្គេមនៅក្នុងទម្រង់នៃ iso-hetero និង oogamy វាកើតឡើងតិចជាញឹកញាប់នៅក្នុងផ្សិតជាង eukaryotes ផ្សេងទៀត។ Iso- និង heterogamy កើតឡើងតែនៅក្នុង chytridiomycetes ប៉ុណ្ណោះ។ oogamy បុរាណជាមួយនឹងការបង្កើតមេជីវិតឈ្មោលនិងស៊ុតមិនត្រូវបានបង្ហាញនៅក្នុងផ្សិតទេប៉ុន្តែវ៉ារ្យ៉ង់ដែលបានកែប្រែយ៉ាងខ្លាំងត្រូវបានរកឃើញ។
យោងតាមលក្ខណៈនៃបទបញ្ជាផ្លូវភេទនៅក្នុងផ្សិតប្រភេទជាច្រើននៃដំណើរការផ្លូវភេទត្រូវបានសម្គាល់។
Gynandromixisអាចត្រូវបានពិចារណាដោយប្រើឧទាហរណ៍នៃ omycetes dioecious ដែលក្នុងនោះ oogonia និង antheridia មានការរីកចម្រើននៅលើ thalli ផ្សេងគ្នា ឧទាហរណ៍ blight ចុងឬផ្សិតដំឡូង។ ប្រសិនបើប្រភេទហ្សែនដែលមានលក្ខណៈដូចគ្នាត្រូវបានដាំដុះនៅក្នុង monoculture នោះវាបន្តពូជតែប៉ុណ្ណោះ។ តាមរបៀបផ្លូវភេទ. ប្រសិនបើ mycelia នៃប្រភេទពីរគឺនៅជិតនោះ ការផ្លាស់ប្តូរ morphogenetic អាចត្រូវបានតាមដាននៅក្នុងពួកវាក្រោមឥទ្ធិពលនៃអាថ៌កំបាំងគីមីនៃធម្មជាតិ steroid - pheromones ផ្លូវភេទ។ Antheridiol ជំរុញការបង្កើត antheridia នៅក្នុងដៃគូ ហើយ oogoniol ជំរុញការបង្កើត oogonia ។ ក្នុងករណីនេះបទប្បញ្ញត្តិនៃការរួមភេទគឺទាក់ទងគ្នា៖ ថាតើសំពាធនឹងបង្កើតជា antheridia ឬ oogonia អាស្រ័យលើសមាមាត្របរិមាណនៃសារធាតុ pheromones ដែលត្រូវគ្នានៅក្នុងវា និងដៃគូរបស់វា។ ដូច្នេះឈ្មោះនៃដំណើរការផ្លូវភេទ - gynandromixis ។
Dimixis ឬ heterothallism ។វាត្រូវបានគេកត់សម្គាល់ជាយូរមកហើយថាផ្សិតអាចជា homo- ឬ heterothallic ។ នៅក្នុងប្រភេទ homothallic ស្នូលដូចគ្នាបេះបិទតាមហ្សែននៅក្នុង fuse mycelium អំឡុងពេលដំណើរការផ្លូវភេទ។ នៅក្នុងប្រភេទ heterothallic ដើម្បីឆ្លងកាត់វដ្តផ្លូវភេទវាចាំបាច់នៅដំណាក់កាលខ្លះ (ខុសគ្នានៅក្នុង ប្រភេទផ្សេងគ្នាផ្សិត) ការបញ្ចូលគ្នានៃកូនចៅនៃ spores (ច្បាស់ជាងនេះទៅទៀតស្នូលរបស់ពួកគេ) ។ ដើម្បីឱ្យប្រភេទទាំងពីរត្រូវគ្នានឹងការរួមភេទ ត្រូវតែមានភាពខុសគ្នានៃហ្សែន (heteroalllelicity) នៅទីតាំងជាក់លាក់ដែលហៅថា mating loci ។ ផ្សិតភាគច្រើន (zygomycetes, ascomycetes និង basidiomycetes មួយចំនួន) មានទីតាំងមិត្តរួមមួយជាមួយ alleles ពីរ។ ទីតាំងមិត្តរួមមានហ្សែនជាច្រើនដែលគ្រប់គ្រងការសំយោគ pheromones ផ្លូវភេទ។ heterothallism បែបនេះត្រូវបានគេហៅថា unifactorial ឬ bipolar ។ ពូជនៃផ្សិតបែបនេះបន្ទាប់ពី meiosis ត្រូវបានបែងចែកជាពីរក្រុមដែលមិនឆបគ្នាប៉ុន្តែត្រូវគ្នាទៅវិញទៅមកក្នុងសមាមាត្រ 1: 1 ពោលគឺឧ។ ប្រូបាប៊ីលីតេនៃការឆ្លងដែលទាក់ទង (ការបង្កាត់ពូជ) និងមិនទាក់ទងគ្នា (ការបង្កាត់ពូជ) គឺដូចជានៅក្នុង eukaryotes ខ្ពស់ជាង bisexual 50% ។
នៅក្នុង genome នៃ basidiomycetes ខ្ពស់មានទីតាំងមិត្តពីរគឺ A និង B ហើយមានតែប្រភេទ heteroallelic នៅ loci ទាំងពីរប៉ុណ្ណោះដែលអាចប្រើបានជាមួយគ្នាទៅវិញទៅមក (Ax Bx គឺត្រូវគ្នាជាមួយ Ay By ប៉ុន្តែមិនមែនជាមួយ Ax By និង Ay Bx) ។ heterothallism បែបនេះត្រូវបានគេហៅថា two-factor ឬ tetrapolar ។ វាកាត់បន្ថយឱកាសនៃការបង្កាត់ពូជរហូតដល់ 25% ។
Diaphoromixis- basidiomycetes ខ្ពស់មិនមានពីរទេប៉ុន្តែ alleles ជាច្រើននៃទីតាំងមិត្តរួមដែលត្រូវបានរកឃើញដោយចៃដន្យនៅក្នុងប្រភេទផ្សេងគ្នាដែលបង្កើតជាចំនួនប្រជាជន។ ការគ្រប់គ្រងការរួមផ្សំនេះធានាបាននូវឱកាស 100% នៃការបង្កាត់ពូជ ដោយសារពូជដែលមានអាឡែរផ្សេងគ្នាគឺត្រូវគ្នាទៅវិញទៅមក ហើយមានអាឡែសច្រើន។ ជាលទ្ធផល ប្រជាជនកូនកាត់ panmix ត្រូវបានបង្កើតឡើង។
វដ្តជីវិតនៃផ្សិតមានភាពចម្រុះដូចជាផ្សិតខ្លួនឯង។ វដ្តសំខាន់, របស់ពួកគេជាកម្មសិទ្ធិរបស់នាយកដ្ឋាននៃផ្សិត
1. វដ្ដផ្លូវភេទគឺជាលក្ខណៈនៃក្រុមដ៏ធំនៃភាពមិនល្អឥតខ្ចោះ ឬ mitofungi ដែលបានបាត់បង់ការបន្តពូជផ្លូវភេទ។ ការបែងចែកស្នូលរបស់ពួកគេគឺ mitotic ទាំងស្រុង។ ភាគច្រើននៃ mitofungi ជាកម្មសិទ្ធិរបស់ផ្សិត marsupial ប៉ុន្តែដោយសារតែការបាត់បង់ដំណើរការផ្លូវភេទ ពួកវាបង្កើតបានជា ក្រុមផ្លូវការផ្សិតមិនល្អឥតខ្ចោះ ឬ deuteromycetes ។
2. វដ្ត Haploid ។ thallus លូតលាស់មានស្នូល haploid ។ បន្ទាប់ពីដំណើរការផ្លូវភេទ (ការរួមផ្សំគ្នា) ហ្សីហ្គោតឌីផូឡូអ៊ីត (ជាធម្មតាបន្ទាប់ពីសម្រាកមួយរយៈ) បែងចែកតាមបែប meiotically - zygotic meiosis ។ លក្ខណៈនៃ zygomycetes និង chytridiomycetes ជាច្រើន។
3. វដ្តនៃ haploid-dicaryotic ត្រូវបានកំណត់លក្ខណៈដោយការពិតដែលថាបន្ទាប់ពីការលាយបញ្ចូលគ្នានៃមាតិកានៃ gametangia (gametangiogamy) ឬកោសិកា somatic នៃ haploid mycelium (somatogamy) ស្នូលបង្កើតជា dikaryons (គូនៃ nuclei ខុសគ្នាហ្សែន) ។ ពួកគេបែងចែកស្របគ្នាបង្កើតជា dikaryotic mycelium ។ ដំណើរការផ្លូវភេទបញ្ចប់ដោយការលាយបញ្ចូលគ្នានៃ nuclei dikaryon ដែលជាលទ្ធផល zygote បែងចែកដោយ meiosis ដោយគ្មានពេលសម្រាក។ Meiospores បង្កើតជា sporulation ផ្លូវភេទនៃ marsupial និង basidiomycetes ក្នុងទម្រង់ ascospores និង basidiospores ។ នៅពេលដែលពួកវាដុះពន្លក haploid mycelium ត្រូវបានបង្កើតឡើង។ នៅក្នុងភាគច្រើននៃផ្សិត marsupial (លើកលែងតែផ្សិតដំបែ និងផ្សិត taffrin) វដ្ដជីវិតដំណាក់កាល haploid គ្របដណ្ដប់ក្នុងទម្រង់នៃ mycelium លូតលាស់ (anamorph) ដំណាក់កាល dikaryotic គឺមានរយៈពេលខ្លីហើយត្រូវបានតំណាងដោយ hyphae ascogenic ដែលថង់ត្រូវបានបង្កើតឡើង (teleomorph) ។ នៅក្នុង basidiomycetes ដំណាក់កាល dikaryotic គ្របដណ្តប់លើវដ្តជីវិត ដំណាក់កាល haploid គឺមានរយៈពេលខ្លី។
4. វដ្ត haploid-diploid ក្នុងទម្រង់នៃការផ្លាស់ប្តូរ isomorphic នៃជំនាន់គឺកម្រមាននៅក្នុងផ្សិត (ផ្សិតខ្លះ និង chytridiomycetes ក្នុងទឹក)។
5. វដ្ត diploid គឺជាលក្ខណៈនៃ omycetes និង yeasts marsupial មួយចំនួន។ thallus លូតលាស់គឺ diploid, gametic meiosis ត្រូវបានគេសង្កេតឃើញក្នុងអំឡុងពេលនៃការបង្កើត gametangia ឬ gametes ។
3. ក្រុមបរិស្ថានផ្សិត
ពពួកផ្សិត និងសារពាង្គកាយដែលស្រដៀងនឹងផ្សិតមាននៅគ្រប់លើដី និង ប្រព័ន្ធអេកូឡូស៊ីទឹក។, របៀប ផ្នែកសំខាន់បំផុតប្លុក heterotrophic, កាន់កាប់រួមគ្នាជាមួយបាក់តេរី កម្រិត trophicអ្នកបំបែក។ ការប្រើប្រាស់ទូលំទូលាយផ្សិតនៅក្នុង biosphere ត្រូវបានកំណត់ដោយចំនួននៃ លក្ខណៈពិសេសសំខាន់បំផុត:
1. ភាគច្រើនមានរចនាសម្ព័ន្ធ mycelial នៃ thallus ។ (អនុញ្ញាតឱ្យអ្នកធ្វើជាម្ចាស់ស្រទាប់ខាងក្រោមបានយ៉ាងឆាប់រហ័ស និងមានផ្ទៃធំនៃទំនាក់ទំនងជាមួយបរិស្ថាន)។
2. ល្បឿនខ្ពស់នៃការលូតលាស់ និងការបន្តពូជ អនុញ្ញាត រយះពេលខ្លីប្រជាជន ម៉ាស់ធំស្រទាប់ខាងក្រោមបង្កើតបានជាចំនួនដ៏ច្រើននៃ spores និងចែកចាយពួកវាក្នុងចម្ងាយឆ្ងាយ។
3. សកម្មភាពមេតាបូលីសខ្ពស់ដែលបង្ហាញឱ្យឃើញក្នុងជួរដ៏ធំទូលាយនៃកត្តាបរិស្ថាន។
4. អត្រាខ្ពស់នៃការផ្សំហ្សែនឡើងវិញ ការប្រែប្រួលជីវគីមីដ៏សំខាន់ ភាពប្លាស្ទិកបរិស្ថាន។
5. សមត្ថភាពក្នុងការផ្លាស់ប្តូរយ៉ាងលឿនទៅស្ថានភាពនៃចលនាផ្អាកមួយ, បទពិសោធន៍ លក្ខខណ្ឌមិនអំណោយផលក្នុងអំឡុងពេលដ៏យូរ។
មេ កត្តាបរិស្ថានសម្រាប់ផ្សិតគឺជាស្រទាប់ខាងក្រោមអាហារ។ ទាក់ទងទៅនឹងកត្តានេះក្រុមសំខាន់ៗនៃផ្សិតត្រូវបានសម្គាល់ដែលត្រូវបានគេហៅថាក្រុម trophic ។
4. ការពិពណ៌នាសង្ខេបអំពីនាយកដ្ឋានផ្សិត។
កោសិកា Prokaryotic
ប្រូការីយ៉ូត- សារពាង្គកាយដែលមិនដូច eukaryotes មិនមានកោសិកាបង្កើត និងខាងក្នុងផ្សេងទៀត។ សរីរាង្គភ្នាស(លើកលែងតែ cisternae ផ្ទះល្វែងក្នុងប្រភេទរស្មីសំយោគ ដូចជា cyanobacteria)។ រាងជារង្វង់ធំតែមួយគត់ (នៅក្នុងប្រភេទខ្លះ - លីនេអ៊ែរ) ម៉ូលេគុល DNA ពីរខ្សែដែលមានផ្ទុកនូវសារធាតុហ្សែននៃកោសិកា (ដែលគេហៅថា nucleoid) មិនបង្កើតជាស្មុគស្មាញជាមួយប្រូតេអ៊ីនអ៊ីស្តូនទេ (ហៅថាក្រូម៉ាទីន។ ) Prokaryotes រួមមានបាក់តេរីរួមទាំង cyanobacteria (សារាយពណ៌ខៀវបៃតង) និង archaea ។ កូនចៅនៃកោសិកា prokaryotic គឺជាសរីរាង្គនៃកោសិកា eukaryotic - mitochondria និង plastids ។ ខ្លឹមសារសំខាន់នៃកោសិកាដែលបំពេញបរិមាណទាំងមូលរបស់វាគឺ viscous granular cytoplasm ។
កោសិកា Eukaryotic
Eukaryotes- សារពាង្គកាយដែលមិនដូច prokaryotes មានស្នូលកោសិកាដែលកំណត់ពី cytoplasm ដោយភ្នាសនុយក្លេអ៊ែរ។ សម្ភារៈហ្សែនមាននៅក្នុងម៉ូលេគុល DNA ពីរជួរលីនេអ៊ែរជាច្រើន (អាស្រ័យលើប្រភេទនៃសារពាង្គកាយ ចំនួនរបស់វាក្នុងមួយស្នូលអាចមានចាប់ពីពីរទៅច្រើនរយ) ដែលភ្ជាប់ពីខាងក្នុងទៅភ្នាសនៃស្នូលកោសិកា ហើយបង្កើតបានជាទំហំធំ។ ភាគច្រើន (លើកលែងតែ dinoflagellates) ស្មុគស្មាញដែលមានប្រូតេអ៊ីនអ៊ីស្តូនហៅថា chromatin ។ កោសិកា Eukaryotic មានប្រព័ន្ធនៃភ្នាសខាងក្នុងដែលបន្ថែមពីលើស្នូលបង្កើតជាសរីរាង្គផ្សេងទៀត (endoplasmic reticulum, Golgi apparatus ជាដើម)។ លើសពីនេះ ភាគច្រើនមានសារធាតុ prokaryotic intracellular symbionts - mitochondria ហើយសារាយ និងរុក្ខជាតិក៏មាន plastids ផងដែរ។
2) cytoplasm
ស៊ីតូប្លាស្មា - បរិស្ថានខាងក្នុងកោសិកាដែលនៅរស់ ឬស្លាប់ លើកលែងតែស្នូល និងវ៉ាកូអូល មានកំណត់ ភ្នាសប្លាស្មា. វារួមបញ្ចូលទាំង hyaloplasm - សារធាតុថ្លាសំខាន់នៃ cytoplasm សមាសធាតុកោសិកាសំខាន់ៗដែលមាននៅក្នុងវា - សរីរាង្គក៏ដូចជារចនាសម្ព័ន្ធមិនអចិន្រ្តៃយ៍ផ្សេងៗ - ការដាក់បញ្ចូល។
សមាសភាពនៃ cytoplasm រួមមានគ្រប់ប្រភេទនៃសរីរាង្គនិង សារធាតុអសរីរាង្គ. វាក៏ផ្ទុកនូវកាកសំណល់ដែលមិនអាចរលាយបាននៃដំណើរការមេតាបូលីស និងកន្លែងទំនេរ សារធាតុចិញ្ចឹម. សារធាតុសំខាន់នៃ cytoplasm គឺទឹក។
Cytoplasm មានចលនាឥតឈប់ឈរ ហូរនៅខាងក្នុងកោសិការស់ ផ្លាស់ទីជាមួយវា។ សារធាតុផ្សេងៗការរួមបញ្ចូល និងសរីរាង្គ។ ចលនានេះត្រូវបានគេហៅថា cyclosis ។ ដំណើរការមេតាប៉ូលីសទាំងអស់កើតឡើងនៅក្នុងវា។
cytoplasm មានសមត្ថភាពលូតលាស់ និងបន្តពូជ ហើយប្រសិនបើត្រូវបានដកចេញដោយផ្នែកអាចត្រូវបានស្ដារឡើងវិញ។ ទោះជាយ៉ាងណាក៏ដោយ cytoplasm ដំណើរការជាធម្មតាតែនៅក្នុងវត្តមាននៃស្នូលប៉ុណ្ណោះ។ បើគ្មានវាទេ cytoplasm មិនអាចមានរយៈពេលយូរដូចស្នូលដែលមិនមាន cytoplasm នោះទេ។
តួនាទីដ៏សំខាន់បំផុតនៃ cytoplasm គឺដើម្បីបង្រួបបង្រួមរចនាសម្ព័ន្ធកោសិកាទាំងអស់ (សមាសធាតុ) និងផ្តល់ឱ្យពួកគេនូវ អន្តរកម្មគីមី. cytoplasm ក៏រក្សា turgor (បរិមាណ) នៃកោសិកា និងរក្សាសីតុណ្ហភាព។
3) ជញ្ជាំងកោសិកា
ជញ្ជាំងកោសិកា- ភ្នាសកោសិការឹងដែលមានទីតាំងនៅខាងក្រៅភ្នាស cytoplasmic និងអនុវត្តមុខងាររចនាសម្ព័ន្ធ ការពារ និងដឹកជញ្ជូន។ ត្រូវបានរកឃើញនៅក្នុងបាក់តេរី archaea ផ្សិត និងរុក្ខជាតិភាគច្រើន។ សត្វពាហនៈ និងប្រូតូហ្សូអាជាច្រើនមិនមានជញ្ជាំងកោសិកាទេ។
ជញ្ជាំងកោសិកាប្រូការីយ៉ូត
ជញ្ជាំងកោសិកាបាក់តេរីត្រូវបានផ្សំឡើងដោយសារធាតុ peptidoglycan (murein) ហើយមានពីរប្រភេទគឺក្រាមវិជ្ជមាន និងក្រាមអវិជ្ជមាន។ ជញ្ជាំងកោសិកានៃប្រភេទក្រាមវិជ្ជមានមានទាំងស្រុងនៃស្រទាប់ក្រាស់នៃ peptidoglycan ដែលនៅជាប់នឹងភ្នាសកោសិកា និងជ្រាបចូលទៅក្នុងអាស៊ីត teichoic ឬ poteichoic ។ នៅក្នុងប្រភេទក្រាមអវិជ្ជមាន ស្រទាប់ peptidoglycan គឺស្តើងជាង រវាងវា និងភ្នាសប្លាស្មាមានចន្លោះ periplasmic ហើយនៅខាងក្រៅកោសិកាត្រូវបានហ៊ុំព័ទ្ធដោយភ្នាសមួយទៀត ដែលតំណាងដោយអ្វីដែលគេហៅថា។ lipopolysaccharide និងជា pyrogenic endotoxin នៃបាក់តេរីក្រាមអវិជ្ជមាន។
ជញ្ជាំងកោសិកាផ្សិត
ជញ្ជាំងកោសិកានៃផ្សិតមាន chitin និង glucans ។
លក្ខណៈពិសេសនៃផ្សិត
ទៅផ្សិត ក្នុងន័យទូលំទូលាយរួមបញ្ចូល eukaryotes heterotrophic ជាមួយនឹងប្រភេទអាហារូបត្ថម្ភ osmotrophic ។ ទាក់ទងនឹងទម្លាប់នៃរបបអាហារ ចរិតលក្ខណៈរចនាសម្ព័ន្ធ និងរបៀបរស់នៅរបស់សារពាង្គកាយទាំងនេះ។ ភាគច្រើន លក្ខណៈផ្សិត៖
- ទម្រង់ទូទៅបំផុតនៃរាងកាយលូតលាស់គឺ mycelium ដែលមានសរសៃអំបោះបន្តបន្ទាប់គ្នាដែលជ្រាបចូលទៅក្នុងស្រទាប់ខាងក្រោម និងស្រូបយកសារធាតុចិញ្ចឹមដែលរលាយក្នុងទឹកពីវា។
– បញ្ហាសរិរាង្គនៅក្នុងស្រទាប់ខាងក្រោម ដែលជាប្រភពអាហារូបត្ថម្ភសម្រាប់ផ្សិត ក្នុងករណីភាគច្រើនមានទម្រង់ biopolymers ដែលមិនអាចជ្រាបចូលបាន។ ភ្នាសកោសិកា. កោសិកាផ្សិតផលិតអង់ស៊ីម depolymerase ដែលត្រូវបានបញ្ចេញទៅក្នុងស្រទាប់ខាងក្រោម និងបំបែកសារធាតុប៉ូលីម៊ែរទៅជាសារធាតុសាមញ្ញជាង។ សមាសធាតុសរីរាង្គមានសមត្ថភាពដឹកជញ្ជូនទៅក្នុងកោសិកាផ្សិត។ អង់ស៊ីមបែបនេះជាធម្មតាត្រូវបានគេហៅថា exoenzymes ។
- ការស្រូបយកសារធាតុសកម្មពីស្រទាប់ខាងក្រោមត្រូវបានអនុវត្តដោយសារតែសម្ពាធ turgor ដ៏ធំសម្បើមដែលនៅក្នុងកោសិកាផ្សិតគឺខ្ពស់ជាងច្រើនបើប្រៀបធៀបជាមួយសារពាង្គកាយ eukaryotic ផ្សេងទៀត។
- ផ្សិតត្រូវបានកំណត់លក្ខណៈយ៉ាងខ្លាំងដោយការបង្កើត spores ដែលលេចឡើងក្នុងអំឡុងពេលបន្តពូជទាំងភេទនិងផ្លូវភេទ។ ការទទួលស្គាល់ផ្សិតគឺអាចធ្វើទៅបានមិនមែនដោយរាងកាយលូតលាស់នោះទេប៉ុន្តែដោយ sporulation ។ ដោយសារតែទីតាំងនៃរាងកាយលូតលាស់នៅខាងក្នុងស្រទាប់ខាងក្រោម និងរបៀបរស់នៅមិនចល័ត នៅពេលបន្តពូជ រចនាសម្ព័ន្ធផ្ទុកមេរោគផ្សេងៗត្រូវបានបង្កើតឡើងនៅលើផ្សិត mycelium ដែលឡើងពីលើស្រទាប់ខាងក្រោម ហើយ spores ដែលបង្កើតឡើងនៅលើពួកវាត្រូវបានចែកចាយដោយចរន្តខ្យល់។ លើចម្ងាយឆ្ងាយណាស់។
ប្រព័ន្ធទំនើបផ្សិត
ដោយ គំនិតទំនើបដោយផ្អែកលើការសិក្សាអំពីហ្សែននៃ "ផ្សិត" ecomorph គឺជាក្រុមសមូហភាពដែលផ្សិតពិតត្រូវបានសម្គាល់ នគរ Mycota ឬ Fungi ដែលភាគច្រើន និងចំនួនតិចនៃប្រភេទសត្វ និងតិច។ ក្រុមចម្រុះសារពាង្គកាយដូចផ្សិត (pseudomycetes) គឺជាផ្នែកមួយនៃព្រះរាជាណាចក្រ Straminopila ដែលរួមបញ្ចូលសារាយពណ៌ (Ochrophyta) ផងដែរ។ នគរនៃផ្សិតពិតបង្រួបបង្រួម 4 ផ្នែកដែលខុសគ្នានៅក្នុងរចនាសម្ព័ន្ធនៃរាងកាយលូតលាស់និងលក្ខណៈនៃការបន្តពូជ: Chytridiomycota, Zygomycota, Ascomycota, Basidiomycota ។
តាមនិតិវិធី សញ្ញាសំខាន់ៗប្រើក្នុងការសាងសង់ប្រព័ន្ធផ្សិត
- រចនាសម្ព័ន្ធនៃសរីរាង្គលូតលាស់
- សមាសធាតុគីមី និងរចនាសម្ព័ន្ធនៃជញ្ជាំងកោសិកា
- ប្រភេទនៃដំណើរការផ្លូវភេទ
- ប្រភេទនៃ sporulation
ប្រភេទនៃ thalli នៃផ្សិតនិងសារពាង្គកាយដូចផ្សិត
Amoeboid ឬ plasmodial thallus គឺជាកោសិកា mononuclear ឬ multinucleate ដែលមិនមានជញ្ជាំងកោសិកាដែលព័ទ្ធជុំវិញដោយភ្នាសប្លាស្មានិងស្រទាប់ប្រូតេអ៊ីនគ្រឿងកុំព្យូទ័របង្រួម - periplast ។ ដើម្បីបោះយុថ្កានៅក្នុងស្រទាប់ខាងក្រោម និងបង្កើនផ្ទៃស្រូបយក ការរីកដុះដាលដូចឫសអាចត្រូវបានបង្កើតឡើង ដែលរួមគ្នាបង្កើតជា rhizomycelium ។ ក្នុងអំឡុងពេលបន្តពូជ ទាំង thallus ទាំងមូលប្រែទៅជា sporangium តែមួយ (holocarpic thallus) ឬ sporangia ជាច្រើនត្រូវបានបង្កើតឡើង (eucarpic thallus) ។
mycelial thallusនិងការកែប្រែរបស់វា។ មូលដ្ឋាន ធាតុរចនាសម្ព័ន្ធ- ហាយហ្វា។ mycelium ចម្រៀកនិងមិនចម្រៀក។ Septa និងការបង្កើតរបស់ពួកគេ។
ការកែប្រែនៃ mycelium ។
រចនាសម្ព័ន្ធសម្រាក៖ chlamydospores និង sclerotia ។
រចនាសម្ព័ន្ធនិងការគាំទ្រ: strands និង rhizomorphs ។
រចនាសម្ព័ន្ធឆ្លង៖ appressoria និង haustoria
ដុះពន្លកដូចផ្សិតតំណាងឱ្យកោសិកាដែលគ្របដណ្ដប់ដោយភ្នាស ឬមិនស្ថិតស្ថេរ ងាយបំបែកខ្សែសង្វាក់នៃកោសិកាដែលត្រូវបានបង្កើតឡើងកំឡុងពេលពន្លកដោយសារតែការមិនខុសគ្នានៃកោសិកាម្តាយ និងកូនស្រី។ សភាពដូចផ្សិតអចិន្ត្រៃយ៍ និងបណ្តោះអាសន្ននៃ thalli ។
សមាសភាពគីមីនិងរចនាសម្ព័ន្ធនៃជញ្ជាំងកោសិកានៃផ្សិត
ជញ្ជាំងកោសិកានៃផ្សិតមានច្រើនស្រទាប់ ដោយមានស្រទាប់ផ្សេងៗគ្នាដែលបង្កើតឡើងដោយកាបូអ៊ីដ្រាតរចនាសម្ព័ន្ធដែលខុសគ្នានៅក្នុងសមាសធាតុគីមីរបស់ពួកគេ ដែលអាចត្រូវបានបែងចែកជា 3 ក្រុមដោយផ្អែកលើសមាសធាតុគីមីរបស់ពួកគេ៖
ប៉ូលីមែរគ្លុយកូស(glucan, chitin, សែលុយឡូស) ។ Glucans បង្កើតស្រទាប់ខាងក្រៅនៃជញ្ជាំងកោសិកានៃផ្សិតភាគច្រើន។ ស្រទាប់ខាងក្នុងនៃជញ្ជាំងកោសិកាផ្សិតត្រូវបានបង្កើតឡើងដោយច្រវាក់នៃ chitin ដែលផ្តល់ឱ្យវានូវភាពរឹង។ Chitin ជំនួស cellulose ដែលអវត្តមាននៅក្នុងផ្សិតភាគច្រើន ប៉ុន្តែជាផ្នែកមួយនៃជញ្ជាំងកោសិកានៃ oomycetes ដែលបច្ចុប្បន្នមិនមែនជារបស់ផ្សិតធម្មតាទេ។ ឈីទីនដែលខូចត្រូវបានគេហៅថា chitosan ដែលរួមបញ្ចូលគ្នាជាមួយ chitin បង្កើតជាជញ្ជាំងកោសិកានៃ zygomycetes ។
ប៉ូលីមែរនៃ monosaccharides ផ្សេងទៀត (mannose, galactose ជាដើម)មិនដូចរុក្ខជាតិខ្ពស់ទេដែលពួកគេបង្កើតជាមូលដ្ឋាននៃម៉ាទ្រីសក្រោមឈ្មោះទូទៅ hemicellulose ពួកវាមិនសូវជាលក្ខណៈនៃផ្សិតទេ។ ករណីលើកលែងមួយគឺផ្សិត ដែលជញ្ជាំងកោសិការបស់វាសម្បូរទៅដោយសារធាតុប៉ូលីម៊ូសដែលហៅថា ម៉ាន់ណាន។ វាត្រូវបានសន្មត់ថាសមាសភាពជញ្ជាំងនេះល្អប្រសើរជាងមុនធានាការចេញផ្កា។
ប៉ូលីមែរនៃកាបូអ៊ីដ្រាតដែលត្រូវបានផ្សារភ្ជាប់ទៅនឹង peptides (glycoproteins)បង្កើតជាស្រទាប់កណ្តាលនៃជញ្ជាំងកោសិកាពហុស្រទាប់ ហើយដើរតួនាទីយ៉ាងសំខាន់ទាំងក្នុងការរក្សាភាពសុចរិតនៃរចនាសម្ព័ន្ធរបស់កោសិកា និងក្នុងដំណើរការមេតាបូលីសរបស់វាជាមួយនឹងបរិស្ថាន។
លក្ខណៈពិសេសផ្សេងទៀតនៃកោសិកាផ្សិតរួមមាន: អវត្តមាននៃផ្លាស្ទិចដែលនាំវាទៅជិតកោសិកាសត្វ។
គ្មានម្សៅដែលនៅក្នុង eumycetes ត្រូវបានជំនួសដោយ polysaccharide ជិតទៅនឹង glycogen ម្សៅសត្វ នៅក្នុង omycetes ដោយ polysaccharide ជិត laminarin នៃសារាយពណ៌ត្នោត។ ចំនួននៃកាបូអ៊ីដ្រាតបម្រុងជាក់លាក់ចំពោះផ្សិតក៏ត្រូវបានផលិតផងដែរ។
ការអភិវឌ្ឍន៍ជាក់លាក់ សារធាតុរំលាយអាហារបន្ទាប់បន្សំដែលក្នុងនោះថ្នាំអង់ទីប៊ីយោទិច phyto- និង mycotoxins និង phytohormones ដើរតួយ៉ាងសំខាន់។
លក្ខណៈជាក់លាក់នៃផ្សិតក៏រួមបញ្ចូលផងដែរ។ heterokaryosis និងដំណើរការ parasexual ។
នៅក្នុងផ្សិត បាតុភូតនៃ heterokaryosis ឬ heteronuclearity គឺរីករាលដាលយ៉ាងខ្លាំង ដែលក្នុងនោះ nuclei ដែលមាន heteroallelic សម្រាប់ហ្សែនមួយចំនួននៅតែមាននៅក្នុងកោសិកាមួយក្នុងរយៈពេលយូរ។ Heterokaryosis ជំនួស heterozygosity នៅក្នុងផ្សិត haploid និងជំរុញការសម្របខ្លួនយ៉ាងឆាប់រហ័សនៃផ្សិតទៅនឹងការផ្លាស់ប្តូរលក្ខខណ្ឌ។ វត្តមាននៃស្នូលផ្សេងគ្នាគឺដោយសារតែលក្ខណៈពិសេសមួយចំនួននៃផ្សិត:
1. វត្តមាននៃស្នូលច្រើនជាងមួយនៅក្នុងកោសិកាមួយ។
2. រចនាសម្ព័ន្ធជាក់លាក់នៃកោសិកា septum ដែលក្នុងនោះមានមួយ ឬច្រើនតាមរយៈរន្ធ ហៅថា រន្ធញើស ដែលតាមរយៈនោះ ស្នូលអាចធ្វើចំណាកស្រុកពីកោសិកាមួយទៅកោសិកាមួយទៀត។
3. hyphae នៅក្នុងអាណានិគមមួយ និងសូម្បីតែអាណានិគមដែលមានគម្លាតយ៉ាងជិតស្និទ្ធខុសៗគ្នា ដែលដុះចេញពី spores ផ្សេងគ្នានៃផ្សិតប្រភេទដូចគ្នា ជារឿយៗលូតលាស់ជាមួយគ្នា ដែលជាលទ្ធផលដែលការផ្លាស់ប្តូរនៃ nuclei នៃប្រភេទផ្សេងគ្នាគឺអាចធ្វើទៅបាន។
Parasexual (pseudosexual)ដំណើរការ។ ប្រសិនបើនៅក្នុងកោសិកា heterokaryotic nuclei ដែលមាន heteroallelic នៅទីតាំងណាមួយនោះ nucleus diploid heterozygous លេចឡើង។ វាអាចចូលទៅក្នុង spore និងបង្កើតជាក្លូន heterozygous diploid ។ ក្នុងអំឡុងពេលដំណើរការនៃ mitosis ស្នូល diploid អាចត្រលប់ទៅរដ្ឋ haploid ដោយសារតែការបាត់បង់ក្រូម៉ូសូមមួយឈុត ឬការផ្លាស់ប្តូរផ្នែកក្រូម៉ូសូមអាចកើតឡើងនៅក្នុងពួកវា (ការឆ្លងកាត់ mitotic) ។ ដំណើរការទាំងពីរនេះត្រូវបានអមដោយការផ្សំឡើងវិញនៃហ្សែនឪពុកម្តាយ ហើយជាលទ្ធផល phenotypes ។ Parasexual ( asexual ) recombination គឺជាបាតុភូតដ៏កម្រមួយ មិនលើសពីមួយ nucleus ក្នុងមួយលាន ប៉ុន្តែដោយសារតែចំនួន nuclei ដ៏ច្រើននៅក្នុង mycelium វាត្រូវបានគេសង្កេតឃើញជានិច្ចនៅក្នុងពពួកផ្សិត។
ការបន្តពូជ- លូតលាស់, ផ្លូវភេទ, ផ្លូវភេទ។
បន្លែ- ការបែកខ្ចាត់ខ្ចាយនៃ thallus ការបង្កើត chlamydospores ដែលបន្ទាប់ពីរយៈពេលនៃការ dormancy លូតលាស់ទៅជា mycelium, budding នៅក្នុង yeast ។
ផ្លូវភេទការបន្តពូជនៃផ្សិតផ្សេងៗគ្នាអាចត្រូវបានអនុវត្តដោយ spores ចល័ត និង immobile ។ Zoospores បង្កើតបានជាក្រុមតូចមួយនៃផ្សិត និងសារពាង្គកាយដែលស្រដៀងនឹងផ្សិត - ក្នុងទឹក និងនៅលើដីមួយចំនួន ដែលទំនាក់ទំនងហ្សែនជាមួយផ្សិតក្នុងទឹក និងសារាយអាចមើលឃើញយ៉ាងច្បាស់។ រចនាសម្ព័នរបស់ flagella គឺជាលក្ខណៈវិនិច្ឆ័យដ៏សំខាន់មួយនៅពេលដែលត្រូវបានចាត់ឱ្យទៅនគរជាក់លាក់មួយ។ នៅក្នុងភាគច្រើននៃផ្សិត កំឡុងពេលបន្តពូជដោយភេទដូចគ្នា ស្ពែមដែលមិនមានចលនាត្រូវបានបង្កើតឡើង ដែលបង្ហាញពីការកើតមាននៅលើដី។ យោងទៅតាមកន្លែងនៃការបង្កើតនិងការធ្វើមូលដ្ឋានីយកម្ម sporangiospores endogenous ត្រូវបានសម្គាល់ដែលបង្កើតឡើងនៅក្នុង sporangia និង exogenous (conidia) ការអភិវឌ្ឍនៅលើ hyphae ពិសេស - conidiophores ។ Conidia ត្រូវបានបង្កើតឡើងនៅក្នុងផ្សិតភាគច្រើន (ascomycetes, basidiomycetes, zygomycetes មួយចំនួន) បង្កើតបានជា sporulation conidial ដែលមានភាពចម្រុះខ្លាំង ហើយត្រូវបានគេប្រើយ៉ាងទូលំទូលាយសម្រាប់ការធ្វើរោគវិនិច្ឆ័យផ្សិត។
ការបន្តពូជផ្លូវភេទនៃផ្សិតមានភាពជាក់លាក់របស់វាទាំងនៅក្នុង morphology នៃដំណើរការផ្លូវភេទ និងនៅក្នុងយន្តការនៃបទបញ្ជាហ្សែន និងសរីរវិទ្យានៃការរួមភេទ និងការបញ្ជូនព័ត៌មានតំណពូជ។
សូម៉ាតូហ្គាមី- ប្រភេទទូទៅបំផុត និងសាមញ្ញបំផុតនៃដំណើរការផ្លូវភេទ រួមមានការបញ្ចូលគ្នានៃកោសិកា somatic ពីរដែលមិនខុសគ្នាទៅជា gametes ។ ពេលខ្លះវាកើតឡើងសូម្បីតែដោយគ្មានការលាយកោសិកា - ស្នូលនៅខាងក្នុងកោសិកាបញ្ចូលគ្នា។ ត្រូវបានរកឃើញនៅក្នុង basidiomycetes ភាគច្រើន yeasts marsupial និង taxa មួយចំនួនទៀត។
ល្បែងផ្គុំរូប- gametangia ត្រូវបានញែកដាច់ពីគេនៅលើ haploid mycelium ហើយមាតិការបស់វាត្រូវបានបង្ហូរក្នុងអំឡុងពេលដំណើរការផ្លូវភេទ។ ដំណើរការផ្លូវភេទនេះគឺជាលក្ខណៈនៃផ្សិត marsupial ភាគច្រើន។ វ៉ារ្យ៉ង់នៃ gametangiogamy គឺ zygogamy នៅក្នុង zygomycetes ។
ការលេងហ្គេមនៅក្នុងទម្រង់នៃ iso-hetero និង oogamy វាកើតឡើងតិចជាញឹកញាប់នៅក្នុងផ្សិតជាង eukaryotes ផ្សេងទៀត។ Iso- និង heterogamy កើតឡើងតែនៅក្នុង chytridiomycetes ប៉ុណ្ណោះ។ oogamy បុរាណជាមួយនឹងការបង្កើតមេជីវិតឈ្មោលនិងស៊ុតមិនត្រូវបានបង្ហាញនៅក្នុងផ្សិតទេប៉ុន្តែវ៉ារ្យ៉ង់ដែលបានកែប្រែយ៉ាងខ្លាំងត្រូវបានរកឃើញ។
យោងតាមលក្ខណៈនៃបទបញ្ជាផ្លូវភេទនៅក្នុងផ្សិតប្រភេទជាច្រើននៃដំណើរការផ្លូវភេទត្រូវបានសម្គាល់។
Gynandromixisអាចត្រូវបានពិចារណាដោយប្រើឧទាហរណ៍នៃ omycetes dioecious ដែលក្នុងនោះ oogonia និង antheridia មានការរីកចម្រើននៅលើ thalli ផ្សេងគ្នា ឧទាហរណ៍ blight ចុងឬផ្សិតដំឡូង។ ប្រសិនបើប្រភេទហ្សែនដែលមានលក្ខណៈដូចគ្នាត្រូវបានដាំដុះនៅក្នុងពហុវប្បធម៍ នោះវាបន្តពូជតែតាមភេទប៉ុណ្ណោះ។ ប្រសិនបើ mycelia នៃប្រភេទពីរគឺនៅជិតនោះ ការផ្លាស់ប្តូរ morphogenetic អាចត្រូវបានតាមដាននៅក្នុងពួកវាក្រោមឥទ្ធិពលនៃអាថ៌កំបាំងគីមីនៃធម្មជាតិ steroid - pheromones ផ្លូវភេទ។ Antheridiol ជំរុញការបង្កើត antheridia នៅក្នុងដៃគូ ហើយ oogoniol ជំរុញការបង្កើត oogonia ។ ក្នុងករណីនេះបទប្បញ្ញត្តិនៃការរួមភេទគឺទាក់ទងគ្នា៖ ថាតើសំពាធនឹងបង្កើតជា antheridia ឬ oogonia អាស្រ័យលើសមាមាត្របរិមាណនៃសារធាតុ pheromones ដែលត្រូវគ្នានៅក្នុងវា និងដៃគូរបស់វា។ ដូច្នេះឈ្មោះនៃដំណើរការផ្លូវភេទ - gynandromixis ។
Dimixis ឬ heterothallism ។វាត្រូវបានគេកត់សម្គាល់ជាយូរមកហើយថាផ្សិតអាចជា homo- ឬ heterothallic ។ នៅក្នុងប្រភេទ homothallic ស្នូលដូចគ្នាបេះបិទតាមហ្សែននៅក្នុង fuse mycelium អំឡុងពេលដំណើរការផ្លូវភេទ។ នៅក្នុងប្រភេទសត្វ heterothallic ដើម្បីឆ្លងកាត់វដ្តផ្លូវភេទនៅដំណាក់កាលខ្លះ (ខុសគ្នាសម្រាប់ប្រភេទផ្សិតផ្សេងៗគ្នា) ការលាយបញ្ចូលគ្នានៃកូនចៅនៃ spores (កាន់តែច្បាស់ជាងនេះទៅទៀតស្នូលរបស់វា) គឺចាំបាច់។ ដើម្បីឱ្យប្រភេទទាំងពីរត្រូវគ្នានឹងការរួមភេទ ត្រូវតែមានភាពខុសគ្នានៃហ្សែន (heteroalllelicity) នៅទីតាំងជាក់លាក់ដែលហៅថា mating loci ។ ផ្សិតភាគច្រើន (zygomycetes, ascomycetes និង basidiomycetes មួយចំនួន) មានទីតាំងមិត្តរួមមួយជាមួយ alleles ពីរ។ ទីតាំងមិត្តរួមមានហ្សែនជាច្រើនដែលគ្រប់គ្រងការសំយោគ pheromones ផ្លូវភេទ។ heterothallism បែបនេះត្រូវបានគេហៅថា unifactorial ឬ bipolar ។ ពូជនៃផ្សិតបែបនេះបន្ទាប់ពី meiosis ត្រូវបានបែងចែកជាពីរក្រុមដែលមិនឆបគ្នាប៉ុន្តែត្រូវគ្នាទៅវិញទៅមកក្នុងសមាមាត្រ 1: 1 ពោលគឺឧ។ ប្រូបាប៊ីលីតេនៃការឆ្លងដែលទាក់ទង (ការបង្កាត់ពូជ) និងមិនទាក់ទងគ្នា (ការបង្កាត់ពូជ) គឺដូចជានៅក្នុង eukaryotes ខ្ពស់ជាង bisexual 50% ។
នៅក្នុង genome នៃ basidiomycetes ខ្ពស់មានទីតាំងមិត្តពីរគឺ A និង B ហើយមានតែប្រភេទ heteroallelic នៅ loci ទាំងពីរប៉ុណ្ណោះដែលអាចប្រើបានជាមួយគ្នាទៅវិញទៅមក (Ax Bx គឺត្រូវគ្នាជាមួយ Ay By ប៉ុន្តែមិនមែនជាមួយ Ax By និង Ay Bx) ។ heterothallism បែបនេះត្រូវបានគេហៅថា two-factor ឬ tetrapolar ។ វាកាត់បន្ថយឱកាសនៃការបង្កាត់ពូជរហូតដល់ 25% ។
Diaphoromixis- basidiomycetes ខ្ពស់មិនមានពីរទេប៉ុន្តែ alleles ជាច្រើននៃទីតាំងមិត្តរួមដែលត្រូវបានរកឃើញដោយចៃដន្យនៅក្នុងប្រភេទផ្សេងគ្នាដែលបង្កើតជាចំនួនប្រជាជន។ ការគ្រប់គ្រងការរួមផ្សំនេះធានាបាននូវឱកាស 100% នៃការបង្កាត់ពូជ ដោយសារពូជដែលមានអាឡែរផ្សេងគ្នាគឺត្រូវគ្នាទៅវិញទៅមក ហើយមានអាឡែសច្រើន។ ជាលទ្ធផល ប្រជាជនកូនកាត់ panmix ត្រូវបានបង្កើតឡើង។
វដ្តជីវិតនៃផ្សិតមានភាពចម្រុះដូចជាផ្សិតខ្លួនឯង។ វដ្តសំខាន់, របស់ពួកគេជាកម្មសិទ្ធិរបស់នាយកដ្ឋាននៃផ្សិត
1. វដ្ដផ្លូវភេទគឺជាលក្ខណៈនៃក្រុមដ៏ធំនៃភាពមិនល្អឥតខ្ចោះ ឬ mitofungi ដែលបានបាត់បង់ការបន្តពូជផ្លូវភេទ។ ការបែងចែកស្នូលរបស់ពួកគេគឺ mitotic ទាំងស្រុង។ ភាគច្រើននៃ mitofungi ជាកម្មសិទ្ធិរបស់ផ្សិត marsupial ប៉ុន្តែដោយសារតែការបាត់បង់ដំណើរការផ្លូវភេទ ពួកវាបង្កើតបានជាក្រុមផ្លូវការនៃផ្សិតមិនល្អឥតខ្ចោះ ឬ deuteromycetes ។
2. វដ្ត Haploid ។ thallus លូតលាស់មានស្នូល haploid ។ បន្ទាប់ពីដំណើរការផ្លូវភេទ (ការរួមផ្សំគ្នា) ហ្សីហ្គោតឌីផូឡូអ៊ីត (ជាធម្មតាបន្ទាប់ពីសម្រាកមួយរយៈ) បែងចែកតាមបែប meiotically - zygotic meiosis ។ លក្ខណៈនៃ zygomycetes និង chytridiomycetes ជាច្រើន។
3. វដ្តនៃ haploid-dicaryotic ត្រូវបានកំណត់លក្ខណៈដោយការពិតដែលថាបន្ទាប់ពីការលាយបញ្ចូលគ្នានៃមាតិកានៃ gametangia (gametangiogamy) ឬកោសិកា somatic នៃ haploid mycelium (somatogamy) ស្នូលបង្កើតជា dikaryons (គូនៃ nuclei ខុសគ្នាហ្សែន) ។ ពួកគេបែងចែកស្របគ្នាបង្កើតជា dikaryotic mycelium ។ ដំណើរការផ្លូវភេទបញ្ចប់ដោយការលាយបញ្ចូលគ្នានៃ nuclei dikaryon ដែលជាលទ្ធផល zygote បែងចែកដោយ meiosis ដោយគ្មានពេលសម្រាក។ Meiospores បង្កើតជា sporulation ផ្លូវភេទនៃ marsupial និង basidiomycetes ក្នុងទម្រង់ ascospores និង basidiospores ។ នៅពេលដែលពួកវាដុះពន្លក haploid mycelium ត្រូវបានបង្កើតឡើង។ នៅក្នុងភាគច្រើននៃផ្សិត marsupial (លើកលែងតែផ្សិតដំបែនិងផ្សិត taphrin) វដ្តជីវិតត្រូវបានគ្រប់គ្រងដោយដំណាក់កាល haploid ក្នុងទម្រង់ជា mycelium លូតលាស់ (anamorph) ដំណាក់កាល dikaryotic គឺមានរយៈពេលខ្លីហើយត្រូវបានតំណាងដោយ hyphae ascogenous ដែលថង់។ ត្រូវបានបង្កើតឡើង (teleomorph) ។ នៅក្នុង basidiomycetes ដំណាក់កាល dikaryotic គ្របដណ្តប់លើវដ្តជីវិត ដំណាក់កាល haploid គឺមានរយៈពេលខ្លី។
4. វដ្ត haploid-diploid ក្នុងទម្រង់នៃការផ្លាស់ប្តូរ isomorphic នៃជំនាន់គឺកម្រមាននៅក្នុងផ្សិត (ផ្សិតខ្លះ និង chytridiomycetes ក្នុងទឹក)។
5. វដ្ត diploid គឺជាលក្ខណៈនៃ omycetes និង yeasts marsupial មួយចំនួន។ thallus លូតលាស់គឺ diploid, gametic meiosis ត្រូវបានគេសង្កេតឃើញក្នុងអំឡុងពេលនៃការបង្កើត gametangia ឬ gametes ។
3. ក្រុមអេកូឡូស៊ីនៃផ្សិត។
ពពួកផ្សិត និងសារពាង្គកាយដែលស្រដៀងនឹងផ្សិតត្រូវបានរួមបញ្ចូលនៅក្នុងប្រព័ន្ធអេកូឡូស៊ីលើដី និងក្នុងទឹកទាំងអស់ ដែលជាផ្នែកសំខាន់បំផុតនៃប្លុក heterotrophic រួមជាមួយនឹងបាក់តេរីដែលកាន់កាប់កម្រិត trophic នៃ decomposers ។ ការចែកចាយយ៉ាងទូលំទូលាយនៃផ្សិតនៅក្នុងជីវមណ្ឌលត្រូវបានកំណត់ដោយលក្ខណៈសំខាន់ៗមួយចំនួន៖
1. ភាគច្រើនមានរចនាសម្ព័ន្ធ mycelial នៃ thallus ។ (អនុញ្ញាតឱ្យអ្នកធ្វើជាម្ចាស់ស្រទាប់ខាងក្រោមបានយ៉ាងឆាប់រហ័ស និងមានផ្ទៃធំនៃទំនាក់ទំនងជាមួយបរិស្ថាន)។
2. ល្បឿនខ្ពស់នៃការលូតលាស់ និងការបន្តពូជ អនុញ្ញាតឱ្យក្នុងរយៈពេលដ៏ខ្លីដើម្បីធ្វើអាណានិគមលើស្រទាប់ខាងក្រោមដ៏ធំ បង្កើតជាចំនួនដ៏ច្រើននៃ spores និងចែកចាយពួកវាក្នុងចម្ងាយឆ្ងាយ។
3. សកម្មភាពមេតាបូលីសខ្ពស់ដែលបង្ហាញឱ្យឃើញក្នុងជួរដ៏ធំទូលាយនៃកត្តាបរិស្ថាន។
4. អត្រាខ្ពស់នៃការផ្សំហ្សែនឡើងវិញ ការប្រែប្រួលជីវគីមីដ៏សំខាន់ ភាពប្លាស្ទិកបរិស្ថាន។
5. សមត្ថភាពក្នុងការផ្លាស់ប្តូរយ៉ាងឆាប់រហ័សទៅកាន់ស្ថានភាពនៃចលនាដែលត្រូវបានផ្អាក និងរស់រានមានជីវិតពីលក្ខខណ្ឌមិនអំណោយផលក្នុងរយៈពេលយូរ។
កត្តាបរិស្ថានចម្បងសម្រាប់ផ្សិតគឺស្រទាប់ខាងក្រោមអាហារ។ ទាក់ទងទៅនឹងកត្តានេះក្រុមសំខាន់ៗនៃផ្សិតត្រូវបានសម្គាល់ដែលត្រូវបានគេហៅថាក្រុម trophic ។
4. ការពិពណ៌នាសង្ខេបនៃផ្នែកផ្សិត។
មីក្រូជីវវិទ្យា, ឆ្នាំ ២០១០, ភាគ ៧៩, លេខ ៦, ទំ។ ៧២៣-៧៣៣
UDC 582.281(047)
ជញ្ជាំងកោសិកាផ្សិត៖ គំនិតទំនើបអំពីសមាសភាព និងមុខងារជីវសាស្ត្រ
© 2010 E. P. Feofilova1
ការបង្កើត បណ្ឌិតសភារុស្ស៊ីវិទ្យាស្ថានវិទ្យាសាស្ត្រមីក្រូជីវវិទ្យាដាក់ឈ្មោះតាម។ S.N. Vinogradsky RAS ទីក្រុងម៉ូស្គូ
បានទទួលដោយអ្នកកែសម្រួល 05.11.2009
ការពិនិត្យឡើងវិញគឺផ្តោតលើរចនាសម្ព័ន្ធផ្ទៃដែលបានសិក្សាតិចតួចនៃកោសិកានៃផ្សិត filamentous - ជញ្ជាំងកោសិកា (CW) ។ ទិន្នន័យត្រូវបានផ្តល់ជូនលើវិធីសាស្រ្តឯកោ និងការធ្វើតេស្តភាពបរិសុទ្ធសម្រាប់អវត្តមាននៃមាតិកាស៊ីតូប្លាសម៉ិចនៅក្នុងប្រភាគ CS និងលើសមាសធាតុគីមីរបស់វា។ រចនាសម្ព័ន្ធ (ក្របខ័ណ្ឌ) និងសមាសធាតុខាងក្នុងនៃ CS - aminopolysaccharides, a- និង b-glucans, ប្រូតេអ៊ីន, lipid, អាស៊ីតអ៊ុយរិច, hydrophobins, sporopollenin និង melanins - ត្រូវបានពិភាក្សាលម្អិត។ ការយកចិត្តទុកដាក់ពិសេសត្រូវបានផ្តល់ឱ្យ chitin, របស់វា។ មុខងារថ្មី។នៅក្នុងការការពារប្រឆាំងនឹងភាពតានតឹងនៃកោសិកាក៏ដូចជាភាពខុសគ្នារវាង aminopolysaccharide នៃផ្សិតនិង chitin នៃសារាយនិង Arthropoda ។ បាតុភូតនៃការលូតលាស់ hyphal apical និងការចូលរួមនៃ microvesicles ពិសេសនៅក្នុង morphogenesis នៃកោសិកាផ្សិតត្រូវបានពិភាក្សា។ ទិន្នន័យត្រូវបានផ្តល់នៅលើអង់ស៊ីមដែលពាក់ព័ន្ធនឹងការសំយោគ និងលីសស៊ីអេស។ ទីបំផុតសារៈសំខាន់មុខងារនៃ SC នៅក្នុងផ្សិតបើប្រៀបធៀបជាមួយនឹងរចនាសម្ព័ន្ធផ្ទៃនៃ eukaryotes ខ្ពស់ជាងត្រូវបានពិភាក្សា។
ពាក្យគន្លឹះ: ផ្សិត filamentous, ជញ្ជាំងកោសិកា, វិធីសាស្រ្តឯកោ, សមាសភាពគីមី, កំណើន apical, មុខងារសរីរវិទ្យា, morphogenesis ។
បញ្ហាសំខាន់មួយ។ ជីវវិទ្យាទំនើបសំណួរគឺថាតើក្នុងអំឡុងពេលនៃដំណើរការអភិវឌ្ឍ ការបង្កើតសារពាង្គកាយដូចគ្នាបេះបិទកើតឡើង យន្តការជីវគីមី និងរចនាសម្ព័ន្ធកោសិកាអ្វីខ្លះដែលពាក់ព័ន្ធនឹងដំណើរការនេះ ដែលបានកើតឡើងនៅលើភពផែនដីរបស់យើងរាប់ពាន់លានឆ្នាំមកហើយ។ មិនគួរឱ្យចាប់អារម្មណ៍តិចជាងនេះគឺទិន្នន័យអំពីរបៀបដែលភាពតានតឹងប៉ះពាល់ដល់ morphogenesis និងដែល biopolymers គ្រប់គ្រង ទម្រង់ខាងក្រៅកោសិកា។ ដូចដែលលទ្ធផលស្រាវជ្រាវបានបង្ហាញ ឆ្នាំថ្មីៗនេះការរីកលូតលាស់ hypha នៃផ្សិត filamentous គឺជាគំរូតែមួយគត់ដែលអនុញ្ញាតឱ្យយើងយល់ពីដំណើរការនៃការបង្កើតកោសិកា និងរបៀបដែលភាពស្ថិតស្ថេរនៃ morphology កោសិកាត្រូវបានរក្សាកំឡុងពេល ontogenesis ។ ការរួមចំណែកយ៉ាងធំធេងក្នុងការសិក្សាអំពីបាតុភូតនេះត្រូវបានធ្វើឡើងដោយទិន្នន័យស្តីពីការសិក្សាសមាសភាពនិង មុខងារជីវសាស្រ្ត CS នៃផ្សិត filamentous ។
CS នៃរុក្ខជាតិត្រូវបានគេរកឃើញនៅឆ្នាំ 1665 ហើយ CS នៃផ្សិតតែនៅដើមសតវត្សទី 18 ប៉ុណ្ណោះ។ ទោះយ៉ាងណាក៏ដោយ នេះគឺមានលក្ខណៈស្រពិចស្រពិល រចនាសម្ព័ន្ធកោសិកាអស់រយៈពេលជាយូរមកហើយវាមិនត្រូវបានគេសិក្សាទេព្រោះវាត្រូវបានគេជឿថា CS អនុវត្តមុខងារ "ស៊ុម" ហើយវាត្រូវបានគេប្រៀបធៀបជាមួយនឹងជញ្ជាំងផ្ទះដែលផ្ទុកបន្ទុក។ ប៉ុន្តែនៅដើមសតវត្សទី 20 អាកប្បកិរិយាចំពោះ CS បានផ្លាស់ប្តូរយ៉ាងខ្លាំង វាបានចាប់ផ្តើមត្រូវបានសិក្សាយ៉ាងយកចិត្តទុកដាក់ ប៉ុន្តែភាគច្រើននៅក្នុងរុក្ខជាតិ និងបាក់តេរី។ ទោះជាយ៉ាងណាក៏ដោយ ការអភិវឌ្ឍន៍ដែលពឹងផ្អែកខ្លាំងលើការដាំដុះផ្សិត និងផលិតកម្មជីវបច្ចេកវិទ្យា ដែលនៅក្នុងនោះផ្សិត filamentous គឺជាអ្នកផលិតសារធាតុសកម្មជីវសាស្រ្ត បានធ្វើឱ្យការអភិវឌ្ឍន៍កាន់តែខ្លាំង។
1 អាស័យដ្ឋានសម្រាប់ការឆ្លើយឆ្លង (អ៊ីមែល៖ [អ៊ីមែលការពារ]).
ចង ការស្រាវជ្រាវវិទ្យាសាស្ត្រលើការសិក្សាអំពីផ្សិត CS ។ ទិន្នន័យលើរចនាសម្ព័ន្ធផ្ទៃនេះសម្រាប់រយៈពេលរហូតដល់ទសវត្សរ៍ឆ្នាំ 1980 ត្រូវបានសង្ខេបនៅក្នុងសៀវភៅដំបូងបង្អស់របស់ពិភពលោកស្តីពីផ្សិត CS ។ ក្នុងឆ្នាំបន្តបន្ទាប់ តួសំខាន់នៃការងារបានកើតឡើងនៅចុងបញ្ចប់នៃថ្ងៃទី 20 និង ការចាប់ផ្តើមនៃ XXIសតវត្ស។ ការសិក្សាជាមូលដ្ឋានត្រូវបានធ្វើឡើងលើប្រព័ន្ធ, សមាសធាតុគីមីនៃ CS, ការលូតលាស់ hyphal apical, ការរំលាយអាហារ chitin, ថ្នាំប្រឆាំងនឹងផ្សិត, hydrophobins, covalent ប្រូតេអ៊ីនជាប់, អង់ស៊ីមដែលចូលរួមក្នុងការបង្កើត KS, សាខា hyphal និង lysis នៃ KS ។ ទិន្នន័យទាំងនេះមិនត្រូវបានគេធ្វើជាទូទៅទេ ប៉ុន្តែវាជាការពិភាក្សារួមគ្នាដែលធ្វើឱ្យវាអាចបង្ហាញថារចនាសម្ព័ន្ធផ្ទៃក្រឡានេះផ្ទុកបន្ទុកច្រើនមុខងារ និងដំណើរការដូចជា មុខងារសំខាន់ៗដូចជាការការពារកោសិកាពីសកម្មភាព កត្តាមិនអំណោយផលការគ្រប់គ្រងនៃ morphogenesis ការចូលរួមនៅក្នុងដំណើរការបន្តពូជ ការកំណត់លក្ខណៈសម្បត្តិ antigenic និង adhesive ការគ្រប់គ្រងដំណើរការនៃ dimorphism និងការបង្កើតកោសិកាផ្សិតដែលសម្រាក ការយល់ឃើញនៃសញ្ញាខាងក្រៅ និងការបញ្ជូនរបស់វាទៅភ្នាស និង intracellular messengers ។ ដោយពិចារណាលើចំណុចខាងលើ យើងបានចាត់ទុកថាវាសមស្របក្នុងការពិភាក្សាអំពីទិន្នន័យខាងក្រោម៖ សមាសភាពគីមីនៃ CS មុខងារជីវសាស្រ្តនៃជីវប៉ូលីមេរចម្បងរបស់វា រចនាសម្ព័ន្ធនៃ CS និងអន្តរកោសិកាក្នុង hyphae ផ្សិត បាតុភូតនៃការលូតលាស់ hyphal apical និង អង់ស៊ីមសំខាន់ៗដែលពាក់ព័ន្ធនឹងការបង្កើត និង លីហ្សីស៊ីស។
សមាសធាតុគីមីនៃកោសិកា
ជញ្ជាំងនៃផ្សិត និងមុខងារជីវសាស្ត្រនៃសមាសធាតុចម្បងរបស់វា
កំពុងសិក្សា សមាសធាតុគីមី CS នៃផ្សិតចាប់ផ្តើមជាមួយនឹងការទទួលបានប្រភាគសុទ្ធនៃរចនាសម្ព័ន្ធនេះ i.e. ការបន្សុតនៃ CS ពីការចម្លងរោគ cytoplasmic ។ នីតិវិធីដំបូង - ការបំផ្លាញកោសិកាផ្សិតនិងការលាងកោសិកា - ត្រូវបានអនុវត្តនៅសីតុណ្ហភាពទាប (ប្រហែល 4-5 អង្សាសេ) ដើម្បីការពារសកម្មភាពបំផ្លិចបំផ្លាញនៃអង់ស៊ីមដែលបំផ្លាញ។ មុនពេលកោសិកាត្រូវបានបំផ្លាញពួកវាត្រូវបានកកនៅសីតុណ្ហភាពមួយ។ អាសូតរាវ. ការបំផ្លិចបំផ្លាញកោសិកាត្រូវបានអនុវត្តទាំងនៅក្នុងឧបករណ៍ homogenizers ពិសេស ឬនៅក្នុងការចុចដោយប្រើ " សម្ពាធរឹងមាំ"ការប្រើប្រាស់ឧបករណ៍បំបែក ultrasonic បន្ថែមទៀតផ្តល់នូវលទ្ធផលល្អ។ មាតិកាកោសិកាត្រូវបានលាងសម្អាត ទឹកត្រជាក់ហើយត្រូវបានធ្វើម្តងទៀត 4-5 ដង។ សម្រាប់ការដកយកចេញនូវមាតិកាស៊ីតូប្លាសស៊ីមឱ្យបានហ្មត់ចត់បន្ថែមទៀត សូមលាងសម្អាតជាមួយ NaCl, 8 M អ៊ុយ, 1 M ammonium ឬ 0.5 M អាស៊ីតអាសេទិច. បច្ចុប្បន្ននេះវិធីសាស្រ្តត្រូវបានបង្កើតឡើងសម្រាប់ការញែក CSs ផ្សិតក្នុងអំឡុងពេល ontogenesis របស់ពួកគេ ហើយវិធីសាស្រ្តទាំងនេះមានភាពខុសគ្នាយ៉ាងខ្លាំងសម្រាប់ mycelium និងកោសិកាសម្រាក។ អាស្រ័យលើទិសដៅនៃការស្រាវជ្រាវ ជាពិសេសនៅពេលកំណត់សមាសភាព polysaccharide សារធាតុ lipids ត្រូវបានស្រង់ចេញពី CS ដោយប្រើសារធាតុរំលាយសរីរាង្គ ឧទាហរណ៍ chloroform និង methanol ក្នុងសមាមាត្រ 2: 1 ជួនកាល លទ្ធផលល្អ។ផ្តល់ការព្យាបាលបន្ថែមនៃ CS ជាមួយនឹងអេធើរស៊ុលហ្វួរី។ សារធាតុរំលាយនេះសម្ងួត CS បានយ៉ាងល្អ ហើយជួនកាលអាចជំនួសការស្ងួតដោយបង្កក។ ដំណាក់កាលបន្ទាប់- ការកំណត់ភាពបរិសុទ្ធនៃប្រភាគ CS លទ្ធផល។ ចំពោះគោលបំណងនេះពន្លឺនិង មីក្រូទស្សន៍អេឡិចត្រុងនិងបច្ចេកទេសគំនូរពិសេស។ ប្រតិកម្មដែលប្រើជាទូទៅបំផុតគឺផ្អែកលើអន្តរកម្មនៃ I3- ជាមួយ chitosan ។ CS សុទ្ធដាច់ដោយឡែកជាមួយនឹងដំណោះស្រាយរបស់ Lugol មានពណ៌ផ្កាឈូកឬ ពណ៌ស្វាយខណៈពេលដែល CS នៅដដែលមានពណ៌ក្រហម។ ស្នាមប្រឡាក់សម្រាប់វត្តមាននៃស្នូលជាមួយ DAPI (4,6-ekt1to-2-rkepyTsn-yo1) ក៏ត្រូវបានគេប្រើផងដែរ។ វាគួរតែត្រូវបានសង្កត់ធ្ងន់ជាពិសេសថាភាពឯកោនៃប្រភាគសុទ្ធនៃ CS គឺជានីតិវិធីដែលមានការទទួលខុសត្រូវយ៉ាងខ្លាំងដែលលទ្ធផលជាបន្តបន្ទាប់នៃការវិភាគនៃសមាសធាតុគីមីរបស់វាអាស្រ័យ។ ក្នុងន័យនេះ ការងារគួរឱ្យចាប់អារម្មណ៍មួយគឺនៅក្នុងនោះ សារធាតុ glucan ឬជាក់លាក់ជាងនេះទៅទៀត សារធាតុ chitosan-glucan complex ត្រូវបានរកឃើញនៅក្នុង mycelium របស់អ្នកតំណាងនៃផ្សិត mucor ទោះបីជាទាំងអស់ក៏ដោយ។ ការសិក្សាពីមុនបង្ហាញពីអវត្តមាននៃ glucan នៅក្នុង mycelium នៃ Mucorales ។
បច្ចុប្បន្ននេះសមាសធាតុនៃផ្សិត CS ត្រូវបានបែងចែកទៅជាសមាសធាតុរចនាសម្ព័ន្ធ (chitin, p-(1-3)-p(1-6)-glucans, p-(1-4)-glucan (cellulose) និង intrastructural (ពួកវាគឺ ហៅថាម៉ាទ្រីស) ដែលបានចាប់ផ្តើមរួមបញ្ចូល mannoproteins, galacto-mannoproteins, xylo-mannoproteins, glucurono-
mannoproteins និង a-(1-3)-glucan ។ ការសិក្សាតិចតួចបំផុតគឺ a-glucan ដែលជាវត្ថុធាតុ polymer លីនេអ៊ែរនៃគ្លុយកូស (នៅក្នុង Schizosaccharomyces pombe glucan នេះមានសំណល់គ្លុយកូសប្រហែល 260) ។ វត្ថុធាតុ polymer នេះមានខ្សែសង្វាក់លីនេអ៊ែរភ្ជាប់ពីរដែលមានសំណល់ប្រហែល 120 (1-3)-a-D-glucose និង (1-4)-a-D-glucose សំណល់នៅចុងនៃម៉ូលេគុលវត្ថុធាតុ polymer ។ វាត្រូវបានគេជឿថា glucan នេះគឺចាំបាច់សម្រាប់ដំណើរការនៃ morphogenesis ។ Glucan ដែលមិនរលាយក្នុងទឹកត្រូវបានញែកចេញពី CS នៃ Penicillium roqueforti mycelium ដែលបន្ទាប់ពីការស្ងួតបាត់បង់សមត្ថភាពក្នុងការរំលាយនៅក្នុងអាល់កាឡាំង ប៉ុន្តែវាអាចរំលាយបានក្នុងដំណោះស្រាយ 10% នៃលីចូមក្លរួនៅក្នុង dimethyl sulfoxide ។ នៅពេលសិក្សា glucan ដោយ methylation ពីរ, tri-O-methyl ដេរីវេនៃគ្លុយកូសត្រូវបានទទួលដែលត្រូវគ្នាទៅនឹងចំណង 1 - 3 និង 1 - "- 4 រវាង monosaccharides ក្នុងសមាមាត្រប្រហែល 5: 2 និងដាននៃ tetra-O ។ - ដេរីវេមេទីលដែលត្រូវគ្នាទៅនឹងស្ថានីយដែលមិនកាត់បន្ថយសំណល់ម៉ូណូ - ណូសាក់។ លទ្ធផលនេះបង្ហាញ រចនាសម្ព័ន្ធលីនេអ៊ែរម៉ូលេគុលដែលមានតែ 1 -»- 3 និង 1 -»- 4 ចំណងរវាងសំណល់គ្លុយកូស។ ទិន្នន័យទាំងនេះត្រូវបានបញ្ជាក់ដោយការវិភាគនៃវិសាលគម 13C-NMR នៃសារធាតុ polysaccharide ដែលការកំណត់រចនាសម្ព័ន្ធនៃសំណល់គ្លុយកូសទាំងអស់ក៏ធ្វើតាមផងដែរ។ polysaccharides បែបនេះជាពិសេសបន្ទាប់ពីការកែប្រែគីមី (sulfation ឬ carbosimethylation) ដែលអាចធ្វើឱ្យពួកវារលាយក្នុងទឹកអាចត្រូវបានប្រើជាប៉ូលីមេដែលមានសកម្មជីវសាស្រ្តនិងមានចំណាប់អារម្មណ៍សម្រាប់ការសិក្សាមុខងារជីវសាស្រ្តរបស់ពួកគេនៅក្នុងជញ្ជាំងកោសិកានៃផ្សិតនិងរុក្ខជាតិ។
មិនរលាយក្នុងទឹក (1 -»- 3)-α-glucans ពីមុនត្រូវបានញែកចេញពីប្រភេទផ្សិតខ្ពស់ និងទាប រួមទាំង Penicillium chrysogenum និង α-glucans ដែលពាក់ព័ន្ធជាមួយនឹងចំណងពីរប្រភេទ (1-3) និង (1 -»។ - 4) ភាពខុសគ្នានៅក្នុងសមាមាត្ររបស់ពួកគេត្រូវបានរកឃើញនៅក្នុង រុក្ខជាតិខ្ពស់ជាង(Aconitum kusnezoffii Reichb) ។
ធាតុផ្សំនៃរចនាសម្ព័ន្ធ- សែលុយឡូស - លក្ខណៈនៃផ្សិត omycete ដែលយោងទៅតាមការធ្វើចំណាកស្រុកទំនើបត្រូវបានចាត់ថ្នាក់ជា pseudofungi ។ សមាសធាតុ monosaccharide នៃ CS រួមមានគ្លុយកូស mannose xylose ហើយជាតិស្ករដែលលេចធ្លោជាងគេគឺគ្លុយកូសដែលមានរហូតដល់ 68% ។ សមាសភាពនៃផ្សិត CS ក៏រួមបញ្ចូលអាស៊ីតអាមីណូ lipid (មិនលើសពី 3%) និង N-acetyl-glucosamine ។ Glucans ដូចជា 1,3-glucan បង្កើតជាស្មុគស្មាញដ៏រឹងមាំជាមួយ chitin ដែលហៅថា chitin-glucan complex (CGC) ដែលបង្កើតជាគ្រោងឆ្អឹងនៃកោសិកាផ្សិត។ ស្មុគ្រស្មាញនេះមានវត្តមាននៅក្នុង CS នៃផ្សិតស្ទើរតែទាំងអស់ លើកលែងតែ zygomycetes ដែលបម្រើជាលក្ខណៈប្រព័ន្ធដែលអាចទុកចិត្តបាន។ នៅក្នុងផ្សិតខ្លះ សារធាតុ mucous ដែលមានសារធាតុ polysaccharides តំណាងដោយ ß-1,3-glucans ដែលមានតែឯកតាគ្លុយកូសដែលតភ្ជាប់ដោយចំណង ß-1,6 ទៅគ្រប់ឯកតាគ្លុយកូសទីបី ត្រូវបានរកឃើញនៅលើផ្ទៃ CS ។