This article will acquaint the reader with the structure of the simplest organisms, namely, it will focus on the structure of the contractile vacuole, which performs an excretory (and other) function, talk about the significance of the simplest organisms and describe the ways of their existence in the environment.
Contractile vacuole. Concept
Vacuole (from the French vacuole, from the Latin word vacuus - empty), spherical-shaped small cavities in plant and animal cells or single-celled organisms. Contractile vacuoles are primarily common among simple organisms that live in freshwater, such as protists such as the amoeba proteus and the ciliate slipper, which received its original name due to the shape of the body, similar to the shape of the sole of a shoe. In addition to the listed protozoa, identical structures were also found in the cells of various freshwater sponges that belong to the Badyagov family.

The structure of the contractile vacuole. Its features
The contractile vacuole is a membrane organelle that releases excess fluid from the cytoplasm. The localization and structure of this apparatus varies among different microorganisms. From a complex of vesicular or tubular vacuoles called spongia, fluid enters the contractile vacuole. Thanks to the constant operation of this system, a stable cell volume is maintained. Protozoa have contractile vacuoles, which are an apparatus that regulates osmotic pressure and also serves to excrete waste products from the body. The body of protozoa consists of only one cell, which, in turn, carries out all the necessary vital functions. Representatives of this subkingdom, such as the slipper ciliate, common amoeba, and other single-celled organisms, have all the properties of an independent organism.
The role of simple organisms
The cell performs all vital functions: excretion, respiration, irritability, movement, reproduction, metabolism. Protozoa are ubiquitous. The largest number of species live in marine and fresh waters, many inhabit moist soil, can infect plants, and live in the bodies of multicellular animals and humans. In nature, protozoa perform a sanitary role; they also participate in the cycle of substances and are food for many animals.

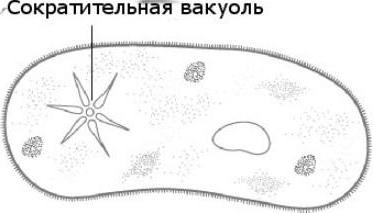
Contractile vacuole in amoeba vulgaris
The common amoeba is a representative of the rhizopod class; unlike other representatives, it does not have a constant body shape. Movement is carried out with the help of pseudopods. Now let’s figure out what function the contractile vacuole performs in an amoeba. This is the regulation of the level of osmotic pressure inside its cell. It can form in any part of the cell. Through the outer membrane, water from the environment enters osmotically. The concentration of dissolved substances in the amoeba cell is higher than in the environment. Thus, a pressure difference is created inside the protozoan cell and outside it. The functions of the contractile vacuole in an amoeba are a kind of pumping apparatus that removes excess water from the cell of a simple organism. Amoeba Proteus can release accumulated liquid into the environment anywhere on the surface of the body.
In addition to osmoregulatory, it performs the function of respiration in life, since as a result of osmosis, incoming water delivers oxygen dissolved in it. What other function does the contractile vacuole perform? It also performs an excretory function, namely, metabolic products are removed together with water into their environment.

Respiration, excretion, osmoregulation in the ciliate slipper
The body of protozoa is covered with a dense shell, which has a constant shape. and algae, including some protozoa. The body of a ciliate has a more complex structure than that of an amoeba. In the slipper cell, there are two contractile vacuoles located in front and behind. In this device, a reservoir and several small tubules are distinguishable. Contractile vacuoles are constantly located, thanks to this structure (from microtubules), in a permanent place in the cell.
The main function of the contractile vacuole in the life activity of this representative of the protozoa is osmoregulation; it also removes excess water from the cell, which penetrates into the cell due to osmosis. First, the afferent channels swell, then the water from them is pumped into a special reservoir. The reservoir contracts, separates from the supply channels, and water is thrown out through the pores. In a ciliate cell there are two contractile vacuoles, which, in turn, act in antiphase. Due to the operation of two such devices, a continuous process is ensured. In addition, water continuously circulates due to the activity of contractile vacuoles. They compress alternately, and the frequency of contractions depends on the ambient temperature.
Thus, at room temperature (+18 - +20 degrees Celsius), the frequency of vacuole contractions is, according to some data, 10-15 seconds. And given that the natural habitat of the slipper is any fresh water body with stagnant water and the presence of decaying organic matter in it, the temperature of this environment changes by several degrees depending on the time of year and, therefore, the frequency of contractions can reach 20-25 seconds. In an hour, the contractile vacuole of a simple organism is capable of releasing large amounts of water from the cell. commensurate with its size. They accumulate nutrients, undigested food debris, end products of metabolism, and oxygen and nitrogen can also be found.
Wastewater treatment with protozoa
The influence of protozoa on the cycle of substances in nature is of great importance. In reservoirs, due to the discharge of wastewater, bacteria multiply in large numbers. As a result, various simple organisms appear, which use these bacteria as food and thus contribute to the natural

Conclusion
Despite the simple structure of these unicellular organisms, the body of which consists of one cell, but performs the functions of a whole organism, amazingly adapted to the environment. This can be observed even in the example of the structure of the contractile vacuole. Today, the enormous importance of the simplest in nature and their participation in the cycle of substances has already been proven.
CONTRACTIVE VACUOL, see VACUOL... Scientific and technical encyclopedic dictionary
Contractile vacuole contractile vacuole. A type of vacuole in some groups of protists, involved in the removal of water (solutions) from the cell during contraction and in the absorption of water by the cell during expansion, which serves to regulate osmotic pressure.... ... Molecular biology and genetics. Dictionary.
Structure of a eukaryotic cell. The vacuole is listed as number 10. The vacuole is a single-membrane organelle found in some eukaryotic cells and ... Wikipedia
Or Heliozoa order of the class Sarcodica (q.v.) type of protozoa (q.v.) animals. Morphological properties. They are distinguished by a spherical protoplasmic body, from which thin, thread-like, non... ... extend in all directions, like rays. Encyclopedic Dictionary F.A. Brockhaus and I.A. Efron
Or Ciliata, a detachment of the class Ciliates, or ciliates (q.v.), a type of protozoa (q.v.). CILIATED CILATES. I (Aspirotricha). Meaning of letters: a powder; al alveolar layer of ectoplasm; ad.Z adoral row of cilia; b tactile seta; cl cilia; ... Encyclopedic Dictionary F.A. Brockhaus and I.A. Efron
- (Lobosea), a class of the most simply organized protozoa above the class of rhizomes. Deprived of internal skeleton and external shells. The body shape is variable, sizes usually range from 20 to 700 microns, rarely slightly more. The shape and size of pseudopodia are characteristic of... ... Biological encyclopedic dictionary
- (Flagellata s. Mastigophora, see table. Flagellates, Flagellata) class of protozoa (Protozoa). Like all other representatives of this type, they have a body consisting of only one cell, representing protoplasm and a nucleus with a nucleolus.... ... Encyclopedic Dictionary F.A. Brockhaus and I.A. Efron
Or Protozoa. Contents of the article: Characteristics and classification. Historical sketch. Morphology; protoplasm with inclusions (trichocysts, nucleus, contractile vacuoles, chromatophores, etc.). Covers and skeleton. Movement P.; pseudopodia, flagella and... Encyclopedic Dictionary F.A. Brockhaus and I.A. Efron
A. CONTRACTIVE VACUOLES
1. Morphology of contractile vacuoles
Most protozoa (except Sporozoa) They have a special, more or less strictly localized, sometimes very complex excretory apparatus, most of which has the character of so-called contractile, or pulsating, vacuoles.
As a general rule, all freshwater animals have vacuoles. Mastigophora And Sarcodina and in all ciliates.
U Mastigophora for the most part there is only 1 contractile vacuole, rarely 2 (Phytomastigina).
The dimensions of the vacuolar apparatus are generally directly proportional to the size of the body and inversely proportional to the number of vacuoles in the body of the animal. So, in large ciliates Heterotricha (Stentor, Spirostomum) there is only 1 vacuole, but it (together with its channels) reaches very large sizes. U Trachelius ovum up to 30 vacuoles, but they are very small.
Sometimes an increase in the number of vacuoles is observed as the animal grows, as in Collinia branchiarum and some others Astomata. That multiplication of the number of vacuoles that Collinia (Anoplophrya) occurs during an individual's life, apparently took place during the evolution of the entire group Astomata(Fig. 124). Indeed, among them one can identify a number of forms with a gradually more complex vacuolar apparatus. Yes, y Dogielella there is only 1 subterminal located vacuole, in the genus Anoplophrya 1 longitudinal row of a small number of vacuoles; at birth Butschliella And Mopodontophrya the number of vacuoles in this row increases greatly, and in Radiophrya vacuoles form 2 longitudinal rows. Finally, the kind Haptophrya a number of numerous vacuoles merge into a common longitudinal vacuolar canal, the method of origin of which is evident from the presence along it of a number of separate outlet openings.
U Sarcodina And Mastigophora the entire excretory apparatus is limited to the vacuole itself. In a few flagellates (e.g. Euglena) the contractile vacuole is surrounded by a rim of small educational vacuoles connected to them by thin tubules.
Pappas and Brandt (1958) think that water in the vacuole Amoeba enters as a result of the outpouring of small vesicles lying around into it, by fusion of the membranes of these vesicles with the central vacuole. In ciliates, contractile vacuoles are usually supplied with one or more afferent canals that pulsate. U Spirostomum - 1 such channel, Stentor- 2, y Frontonia- IO long channels, etc. The channels apparently diverge in the cytoplasm to a much greater distance than is visible under a microscope. At least during osmosis Paramecium the continuation of the peripheral ends of the canals into long, thin end sections becomes noticeable. Educational vacuoles and afferent canals collect fluid (i.e., mainly water) from the cytoplasm and deliver it to the contractile vacuole. The entire vacuolar apparatus of ciliates lies deep in the endoplasm and does not change its position in the body.
Recently, Schneider (1960) published interesting data on the ultramicroscopic structure of the contractile vacuole and adductor canals Paramecium caudatum. According to D. N. Nasonov (1924), the adductor canals are surrounded by an osmiophilic coupling, and according to
observations of Gelei (Gelei, 1925a, 1925b), spongy plasma (Nierenplasma or Nephridialplasma).
According to Schneider's research, this area of plasma surrounding the adductor channels consists of numerous thin and convoluted

Rice. 125. Scheme of the structure of a contractile vacuole with an adductor canal and the surrounding cytoplasm in Paramecium according to electron microscopy. (After Schneider, 1960). A - radial canal in systole, contractile vacuole in diastole; B - radial canal in diastole, contractile vacuole in systole. AK - vacuole exit channel; Amp - peripheral canal ampulla; EK - channel connecting the ampulla with the contractile vacuole; EPR - endoplasmic reticulum channels; FB - contractile fibrils of the vacuole; KV - contractile vacuole; NK - contractile vacuole adductor channel; NT - network of “nephridial channels” associated with endoplasmic reticulum channels; RS - cluster of "nephridial canals" in cross section.
tubes with osmiophilic walls (Fig. 125). The internal lumen of these tubes during systole is 100 Å, and during diastole 150 Å. The entire layer of nephridial plasma has a thickness of 1-1.5 μ. At its periphery, the tubes pass directly into the tubules of the endoplasmic reticulum, which penetrate the entire body of the paramecium. In addition, nephridial plasma is surrounded by special tubes with osmiophilic walls. They are sometimes arranged in bunches. Their diameter reaches 500 Å. The length of these tubes is about 1 μ.
The adductor canal has an osmiophilic membrane. During systole, this channel becomes slit-like and has a lumen of about 300 Å; in diastole it expands to 4000 Å. The tubules surrounding the adductor canal open into it only during diastole, when they increase in diameter, while during systole these tubules are closed and not connected to the canal. The afferent canal at its proximal end directly passes into the ampulla, which opens into the vacuole reservoir through the terminal canaliculus. The wall of the ampulla and the terminal canal contains tubular contractile fibrils arranged in several bundles. From the walls of the terminal canal they pass directly to the wall of the contractile vacuole reservoir, distributing there also in the form of separate bundles.
According to Rudzinska (1957), in the cytoplasm around the vacuole Tokophrya infusionum Small ergastoplasma bubbles are concentrated and mitochondria accumulate. In addition, structures that can be compared to dictyosomes are found around the vacuole. They appear to be involved in the secretion and accumulation of fluid coming from the cytoplasm.

A. A. Strelkov (1939) found around the excretory canal at Cycloposthium a clearly visible annular fiber, which is, in all likelihood, a myonema that regulates the exit of fluid through the canal to the outside. In all ciliates, the excretory opening of the contractile vacuole is a preformed pore. It is usually round or oval and has a completely constant position among the rows of cilia for each species. In the family Conchophthirus the hole looks like a narrow slit with a thickened edge. Perhaps this pore shape is Conchophthirus has a known functional significance when closing it. Namely, it is very likely that here the closure occurs not through an annular constriction of the outlet channel, but by the collapse of the edges of the gap like two lips. The pore opens into the excretory canal, usually very short, but sometimes (at Paramecium trichium) forming several loop-like bends.
Electron microscopic examination of the contractile vacuole Tokophrya infusionum confirmed the presence of a permanently existing pore and excretory canal, around which fibrils (180 A thick) are located that compress it (Rudzinska, 1957). Similar fibrils were found in Paramecium(Schneider, 1960).
2. Functional significance of contractile vacuoles
1) The process of contraction of pulsating vacuoles
Vacuoles work rhythmically, alternately expanding (diastole stage) and slowly filling with fluid, then suddenly contracting (systole stage) and pushing their contents out through the excretory canal. Where there are special adductor channels, the contraction cycle of the entire apparatus becomes more complicated; namely, it begins with the expansion of the channels, which occurs while the vacuole is in a state of systole (channel diastole); this is followed by contraction of the channels (systole), pouring their contents into the contractile vacuole, which causes its systole; at the same time, the channels begin to expand again. In fact, corresponding stages are also present in vacuoles that lack a canal system, but there they are little or not visible, since during the diastole of the vacuole, very small and irregularly located vacuoles located in the surrounding plasma are poured into it (cf. Ophryoscolecidae, rice. 126).
The rate of pulsation varies among different protozoa, and also changes among the same species under the influence of changes in environmental factors.
So, for different freshwater Protozoa the intervals between pulsations at different temperatures are shown in table. 6.
TABLE 6
Changes in the rate of pulsation of the contractile vacuole in ciliates(per second) at different temperatures

Oligochaeta. It turned out that the rate of pulsation of the contractile vacuole is not the same for everyone Astomata and is in connection with the osmotic conditions of the environment in which the owner lives. U Mesnilella clavataw. Hoplitophrya secans among freshwater oligochaetes, pulsation is as frequent as in free-living freshwater ciliates: the pulsation rate does not exceed 1 minute. (20-30 sec.).
U Radiophrya from marine oligochaete Enchytraeus sp., living in the littoral zone of the Barents Sea, pulsation occurs every 6-8 minutes.
Finally, at Metaradiophrya lumbrici And Anoplophrya lumbrici from the intestines of a soil oligochaete Eisenia foetida and at Mesnilella fastigata And Radiophrya prolijera from soil Enchytraeus sp. the pulsation rate is slower (1.5-4 min.) than that of freshwater ciliates, and somewhat faster than that of Astomata from marine oligochaetes. Thus, in closely related species of ciliates Astomata a significant physiological difference is revealed due to adaptation to an osmotic environment. For ciliates from freshwater worms, the isotonic medium is 0.4% ringer, while for ciliates of the intestines of soil worms, isotonicity corresponds to a 0.75-0.8% ringer solution, and for marine forms - a 3% solution.
The influence of changes in the amount of salts in the environment on the rate of vacuole pulsation was demonstrated by Zuelzer (1910). She showed that Amoeba verrucosa when accustomed to life in sea water, it reveals, starting with a salt concentration of 0.3%, a gradual slowdown in the pulsation of the vacuole, and when the salt concentration reaches 1.5%, the vacuole completely disappears, only to reappear when the culture is gradually diluted with fresh water. Similarly, Herfs (1922) obtained for Paramecium, accustomed to life in NaGl solutions and in fresh water, the following data:
| Salt content in water (in%) | 0 | 0.25 | 0.5 | 0.75 | 1 |
| Interval between pulsations (in sec.) | 6.2 | 9.3 | 18.4 | 24.8 | 163 |
| The amount of fluid secreted in 1 hour, expressed in body volumes | 4.8 | 2.82 | 1.38 | 1.08 | 0.16 |
In general, the amount of fluid excreted through the vacuoles in a certain period of time, other things being equal, is proportional to the rate of pulsation and for different Protozoa can be quite different. For example, a contractile vacuole Uronema nigricans at a temperature of 28° for 2 minutes. releases a volume of water equal to the volume of the body of the ciliate, and Euplotes patella this requires 14 minutes, Paramecium aurelia- 46 min.
2) Physiological significance of contractile vacuoles
The functions of vacuoles are apparently diverse. First of all, there is reason to think that the vacuoles are responsible for the removal from the body of the final metabolic products that accumulate in it.
Suggestions about this function were made in the last century by Stein (1878) and other researchers, but Howland (1924a, 1924b) was the first to indirectly show that uric acid is secreted by protozoa. Her cultures of paramecium and amoebae contained uric acid, the concentration of which increased approximately in proportion to the length of time the culture was kept. M. Novikov (1908), Shumway (1917) and Flather (1919) showed that when feeding ciliates of the thyroid gland, under the action of epinephrine and pineal gland extract, the pulsation of vacuoles becomes more frequent, and the vacuoles themselves increase, i.e. stimulation is obtained excretion under the influence of the same drugs that cause diuresis in vertebrates.
Weatherby (1927) discovered urea in the fluid of the contractile vacuole Spirostomum, extracted using a micropipette. However, judging by the calculations of this author, the vacuole removes only 1% of the total excreted by mass cultures. Spirostomum urea.
In general, data on the nature of nitrogenous excretion products are very contradictory. In addition to uric acid and urea, some authors indicated ammonia as excretion products instead of these substances (for example, in Glaucoma And Spirostomum). The only circumstance that does not entirely agree with the excretory function of contractile vacuoles is that they never participate in the excretion of intravital dyes introduced into the plasma, which are very often excreted in higher animals through the kidneys and other excretory organs.
The opinion has been repeatedly expressed that contractile vacuoles take a certain part in the respiratory metabolism of protozoa, facilitating the removal of carbonic acid from the cytoplasm; however, there is no actual evidence of specific activity of the vacuole in this direction. Only indirect data from Ludwig (Ludwig, 1928) on the correspondence between the actual amount of carbonic acid excreted through the vacuole and the estimated amount of carbonic acid secreted by animals are interpreted by this author in favor of the respiratory function of the vacuole.
Most authors currently highlight the important role that contractile vacuoles play as an osmoregulatory apparatus. Even Degen (1905) pointed out that the plasma of freshwater protozoa, due to the presence of salts in it, has a higher osmotic pressure than the environment; and this circumstance leads to the constant absorption of water by the cytoplasm. The contractile vacuole serves to remove excess water continuously entering the plasma. The correctness of this view is confirmed both by the nature of the distribution of contractile vacuoles among protozoa, and by experiments in acclimating freshwater protozoa to water with increasing salt concentrations (Firiley, 1930; Loefer, 1939, etc.).
Recently, an experimental study of Jl. H. Seravini (1958) over the pulsation of the vacuole Paramecium caidatum, which provides data showing that when ciliates are transferred to saline solutions of increased concentration, during the first 15-30 minutes. There is a several-fold decrease in the rate of pulsation of the vacuole, then the rate of pulsation gradually accelerates and approaches the speed characteristic of control ciliates, but usually does not reach its level. For example, in 0.2% NaCl after 30 min. The pulsation rate was on average 40.5 seconds, and after 48 hours. reached 13.1 sec., while in the control group the pulsation proceeded at a speed of 10.3 sec. The same was observed in solutions of 0.1% LiCi, 0.5% CaCl 2, 1% sucrose, and urea solution. The slowdown occurred 7-15 times, but after 2 days
the pulsation rate in these solutions was slowed down by only 1.5-3 times. At lower salt concentrations, a complete restoration of the vacuole pulsation rate was observed. Substances such as HGl, NaOH, GaGl 2, formalin, in negligible concentrations, caused a slight slowdown in pulsation, which then quickly returned to normal. Adrenaline at a concentration of 0.0005% even increased water exchange, but over time the pulsation also returned to normal. Thus, osmotically active and inactive substances cause similar changes in the activity of the contractile vacuole. In this case, there is a general tendency to regulate the water flow passing through the cytoplasm of the ciliate by changing the rate of pulsation of the vacuole. In addition, ciliates adapt to changing conditions of osmotic pressure of the external environment. This was also noted by Kitching (1952) regarding Archesium And Podophrya. In these ciliates, in high concentrations of ethylene glycol and sucrose, the activity of the vacuole completely stops, but after some time the pulsation of the vacuole and water exchange are restored.
All this indicates a complex mechanism of action of the contractile vacuole, which plays an undoubted role in the water exchange of ciliates. Seravin believes, however, based on the fact that osmotically inactive substances cause a delay in vacuole pulsation, it is possible to assume that water exchange in ciliates is not determined by osmotic patterns and therefore the role of the contractile vacuole in osmoregulation cannot be considered proven.
To understand the function of the vacuole, it is, of course, also necessary to understand the mechanism of water circulation from the cytoplasm into the vacuole reservoir and the mechanism of contraction of the vacuole itself. I. Gelei and G. Gelei (Gelei, 1928; Gelei, 1939), like a number of other researchers, believe that only the turgor tension created in the cytoplasm, and, consequently, osmotic conditions determines the reduction of the vacuole reservoir. It is possible that there is also an active mechanism for the secretion of water into the vacuole (Kitching, 1956). According to Schneider (Schneider, 1950), the pulsation of the vacuole is caused by the activity of contractile fibrils located in the wall of the vacuole, and the filling of the afferent canals with water occurs as follows. The liquid collects in the ergastoplasmic tubules and then, due to the larger diameter of these tubules than the nephridial plasma tubules, a flow of water is created from the ergastoplasmic tubules into the nephridial plasma tubules and from them into the lumen of the channel. Of course, further research is needed to more accurately understand the mechanism of these processes. Excretory and respiratory activity of the contractile vacuole seems very possible, but not conclusively proven.
3) Contractile vacuole and Golgi apparatus
The question of the morphological significance of the contractile vacuole entered a new phase after the work of D. N. Nasonov (1924, 1925), who expressed the idea that the contractile vacuole of protozoa is homologous to the Golgi apparatus in cells Metazoa. Nasonov (1924) and almost simultaneously with him I. Gelei (J. Gelei, 1925a, 1925b), applying the osmation technique to ciliates, discovered important, new features in the structure of contractile vacuoles (Fig. 127). Nasonov found that around the contractile vacuole of many ciliates (Campanella, Lionotus, Parameciuin, Nassula), as well as Chilomonas is strongly blackened by
reduced osmium shell, with a smooth surface or surrounded by a layer of small, osmiophilic grains. The shell either surrounds the entire vacuole entirely (in the above species), or has the form of a mesh ring encircling the vacuole along its equator (Chilodon, Dogielella). In forms with a system of adductor channels (Paramecium), the shell continues to the latter; in this case, the channels can be traced in the endoplasm much further than with any other research method, and their distal parts are surrounded by a gray mesh sleeve. Nasonov considers this section of the canals to be the actual excretory part of the apparatus, while the proximal part of the canals and the vacuole itself are only the excretory part of the vacuolar system. U Dogielella Nasonov could state that during the systole of the vacuole, the black ring surrounding it does not follow the latter during its contraction, but remains in its original place. Gelei (1925a, 1925b) tried to compare them with the nephridial system Metazoa, equating the contractile vacuole itself to the bladder, and other parts of the apparatus to other sections of the nephridia. Nasonov focused on the similarity of the osmotic membrane of vacuoles with the Golgi apparatus in Metazoa. Indeed, the drawings he provides of different types of contractile vacuoles and Golgi apparatuses taken from different types of cells in Metazoa, extremely convincing in their similarity.

Based on this, Nasonov considers it possible to look for adductor canals in the osmiophilic spongy membrane Parameciurn and mesh ring Dogitlella not only morphological, but also functional parallelism between these formations and the Golgi apparatus. The network structure of the ring substance depends on the presence of small educational vacuoles in it - the stage of bound secretion. These vacuoles then merge to form a central vacuole lying in the center of the ring - the stage of free secretion secreted outward. According to Nasonov’s concept, the Golgi apparatus serves to collect known substances from the plasma, which are prepared in it for their removal from the body through a vacuole. The entire factual side of Nasonov’s findings was fully confirmed by later research by a number of authors: A. A. Strelkov (1939) Cycloposlhiidae Fauré-Fremiet, 1925) Vorlicella, King (1928) Paramecium trichium And Euplotes etc. True, in some protozoa it was not possible to detect a special osmiophilic membrane around the vacuole, which could depend on methodological issues.
in everyone's cells Metazoa in the form of a mesh formation or consisting of individual dictyosomes. Therefore, we can think that it is also present in all unicellular organisms. Indeed, studies by Hirschler (1927), Hovasse (1937), Joyet-Lavererie (1926) and others showed the presence of many protozoa in the plasma (Gregarina, Coccidia, Flagellata, Sarcodina) small rods, rings, balls with a strongly osmiophilic edge and cells very similar to dictyosomes Metazoa.
Signs of the Golgi apparatus Metazoa First of all, this device is able to specifically reduce certain compounds of heavy metals - silver and osmium, which is expressed in the blackening of the structures of the device. These structures are not stained intravitally with neutral red and methylene blue. Often the characteristic structures of the Golgi apparatus are dictyosomes, which consist of two parts - chromophilic and chromophobic. Typically, secretory granules are immersed in the chromophobe part. As a rule, the Golgi apparatus is localized precisely in the secretory zone of the cell.
Electron microscope studies have shown the unity of the structures of the Golgi apparatus in different cells Metazoa. In all studied cells, double membranes (or γ-cytomembranes) were identified, located parallel to each other and forming a system of 3-7 or more pairs. Each pair encloses a lighter space 50-200 Å wide and forms a sort of flat sac. Sometimes it expands to a large vacuole. The membranes are immersed in a fine-grained base material. In some cases, membranes may surround vacuoles of varying sizes. All these structures are quite stable.
It is significant that there is complete correspondence between the pictures of the Golgi apparatus described using a light microscope by Nasonov and other researchers in cells Metazoa, and modern images obtained with an electron microscope.
Dalton and Felix (1957) also note that the contractile vacuole Chlamidomonas bounded by membranes similar to those of the Golgi apparatus of cells Metazoa, and thus the authors support Nasonov's hypothesis about the homology of these organelles. Around the contractile vacuole in ciliates Campanella And Ophridium there are numerous tubes with dense osmiophilic walls, which can be to some extent comparable to the ultrastructures of the Golgi apparatus (Fauré-Fremiet et Rouiller, 1959). The above data from Schneider on the structure of nephridial plasma around the adductor channels of the contractile vacuole Parameciiiim make, however, difficult to homologize its structure with the ultrastructure of dictyosomes.
Even earlier, French authors (Duboscq et Grassé, 1933; Grassé et Hollande, 1941) also rejected the homology of the osmiophilic membranes of contractile vacuoles with the Golgi apparatus. They believe that the osmiophilic walls of vacuoles, like the contractile vacuoles themselves, are formations completely independent of the Golgi apparatus.
It should be noted that sickle-shaped or ring-shaped dictyosomes were found in some ciliates, such as in Balantidium elongatum, out of connection, with vacuoles (Villeneuve-Brachori, 1940). Noirot-Timothee (1957) identified with an electron microscope the typical structures of the Golgi apparatus in the endoplasm of some Ophryoscolecidae, not associated with the contractile vacuole.
French authors homologize the parabasal body with the Golgi apparatus (see p. 115), since this formation, especially when examined with an electron microscope, turned out to be similar to dictyosomes of cells Metazoa(Fig. 80).
Further comparative studies of protozoa are undoubtedly needed to resolve the issue of the homology of certain structures with the Golgi apparatus of cells Metazoa.