Genetic engineering is a set of methods, techniques and technologies for isolating genes from cells or an organism, obtaining recombinant RNA and DNA, performing various manipulations with genes, as well as introducing them into other organisms. This discipline helps to obtain the desired characteristics of the modified organism.
Genetic engineering is not a science in the broad sense, but is considered a biotechnological tool. It uses research from sciences such as genetics and molecular microbiology.
The created methods of genetic engineering related to the management of heredity were one of the most striking events in the development of science.
Scientists, molecular biologists, and biochemists have learned to change, modify genes and create completely new ones by combining genes from different organisms. They also learned how to synthesize material in accordance with given patterns. Scientists began to introduce artificial material into organisms, forcing them to work. Genetic engineering is based on all this work.
However, there are some limitations of "biological material". Scientists are trying to solve this problem with the help of and Experts note that this path is quite promising. Over the past few decades, scientists have developed techniques by which certain plant or plant cells can be forced to develop and reproduce independently, separately from the organism.
The achievements of genetic engineering are of great importance. used in experiments, as well as in the industrial production of certain substances that cannot be obtained using bacterial cultures. However, there are difficulties in this area as well. For example, the problem is the lack of ability in animal cells to divide as infinitely many times as
During the experiments, fundamental discoveries were made. Thus, for the first time, a “chemically pure” isolated gene was bred. Subsequently, scientists discovered ligase and restriction enzymes. With the help of the latter, it became possible to cut the gene into pieces - nucleotides. And with the help of ligases, on the contrary, you can connect, “glue” these pieces together, but in a new combination, creating, constructing a different gene.
Scientists have also made significant progress in the process of “reading” biological information. For many years, W. Gilbert and F. Sanger, American and English scientists, have been deciphering the data contained in genes.
Experts note that genetic engineering over the entire period of its existence has not had a negative impact on the researchers themselves, has not caused harm to humans and has not caused damage to nature. Scientists note that the results achieved both in the process of studying the functioning of the mechanisms that ensure the life of organisms and in the applied industry are very impressive. At the same time, the prospects seem truly fantastic.
Despite the great importance of genetics and genetic engineering in agriculture and medicine, its main results have not yet been achieved.
Scientists face quite a lot of challenges. It is necessary to determine not only the functions and purpose of each gene, but also the conditions under which its activation occurs, in what periods of life, under the influence of what factors, in what parts of the body it turns on and provokes the synthesis of the corresponding protein. In addition, it is important to find out the role of this protein in the life of the body, what reactions it triggers, whether it goes beyond cellular limits, and what information it carries. The problem of protein folding is quite complex. The solution to these and many other problems is carried out by scientists within the framework of genetic engineering.
Genetic engineering
Modern biology fundamentally differs from traditional biology not only in the greater depth of development of cognitive ideas, but also in a closer connection with the life of society and with practice. We can say that in our time biology has become a means of transforming the living world in order to satisfy the material needs of society. This conclusion is illustrated primarily by the close connection of biology with biotechnology, which has become the most important area of material production, an equal partner of mechanical and chemical technologies created by man, as well as with medicine.
Since their inception, biology and biotechnology have always developed together, with biology being the scientific basis of biotechnology from the very beginning. However, for a long time, the lack of its own data did not allow biology to have a very large influence on biotechnology. The situation changed dramatically with the creation in the second half of the 20th century. genetic engineering methodology, which is understood as genetic manipulation for the purpose of constructing new and reconstructing existing genotypes. Being by its nature a methodological achievement, genetic engineering did not lead to a break in existing ideas about biological phenomena, did not affect the fundamental principles of biology, just as radio astronomy did not shake the fundamental principles of astrophysics, the establishment of the “mechanical equivalent of heat” did not lead to a change in the laws of thermal conductivity, but the proof the atomic theory of matter did not change the relationships between thermodynamics, hydrodynamics and the theory of elasticity (A.A. Baev).
Nevertheless, genetic engineering has opened a new era in biology for the reason that new opportunities have emerged for penetrating into the depths of biological phenomena in order to further characterize the forms of existence of living matter, more effectively study the structure and function of genes at the molecular level, and understand the subtle mechanisms of their operation. genetic apparatus. The success of genetic engineering means a revolution in modern
natural science. They determine the criteria for the value of modern ideas about the structural and functional features of the molecular and cellular levels of living matter. Modern data on living things are of enormous educational importance, because they provide an understanding of one of the most important aspects of the organic world and thereby make an invaluable contribution to the creation of a scientific picture of the world. Thus, by dramatically expanding its cognitive base, biology through genetic engineering also had a leading influence on the rise of biotechnology.
Genetic engineering creates groundwork on the path to understanding the methods and ways of “constructing” new or improving existing organisms, giving them greater economic value and the ability to sharply increase the productivity of biotechnological processes. However, genetic engineering has created new horizons for medicine in the diagnosis and treatment of many diseases, both non-hereditary and hereditary. It has opened new avenues in the search for new drugs and materials used in medicine. Genetic engineering and biotechnology have stimulated the development of bionanotechnology techniques.
Within the framework of genetic engineering there are genetic And cellular engineering. Genetic engineering refers to manipulations to create recombinant DNA molecules. This methodology is often referred to as molecular cloning, gene cloning, recombinant DNA technology, or simply genetic manipulation. It is important to emphasize that the objects of genetic engineering are DNA molecules and individual genes. In contrast, cell engineering refers to the genetic manipulation of isolated individual cells or groups of cells of plants and animals.
GENETIC ENGINEERING AND ITS TOOLS
Genetic engineering is a set of various experimental techniques (techniques) that provide design (reconstruction) and cloning of DNA molecules and genes for specified purposes.
Genetic engineering methods are used in a certain sequence (Fig. 127), and several stages are distinguished in the implementation.
not a typical genetic engineering experiment aimed at cloning a gene, namely:
1. Isolation of plasmidial DNA from the cells of the organism of interest (initial) and isolation of the DNA vector.
2. Cutting (restriction) of the DNA of the original organism into fragments containing genes of interest using one of the restriction enzymes and isolating these genes from the restriction mixture. At the same time, the vector DNA is cut (restricted), transforming it from a circular structure into a linear one.
3. Connecting the DNA segment of interest (gene) with the vector DNA in order to obtain hybrid DNA molecules.
4. Introduction of recombinant DNA molecules by transformation into some other organism, for example into E. coli or somatic cells.
5. Sowing bacteria into which hybrid DNA molecules were introduced onto nutrient media that allow the growth of only cells containing hybrid DNA molecules.
6. Identification of colonies consisting of bacteria containing hybrid DNA molecules.
7. Isolation of cloned DNA (cloned genes) and its characterization, including sequencing of nitrogenous bases in the cloned DNA fragment.
Rice. 127.Consecutive stages of a genetic engineering experiment
During evolution, bacteria developed the ability to synthesize so-called restriction enzymes (endonucleases), which became part of the cellular (bacterial) restriction modification system. In bacteria, restriction-modification systems are an intracellular immune system for protecting against foreign DNA. Unlike higher organisms, in which the recognition and destruction of viruses, bacteria and other pathogens occurs extracellularly, in bacteria, protection from foreign DNA (DNA of plants and animals in whose bodies they live) occurs intracellularly, i.e. when foreign DNA penetrates the cytoplasm of bacteria. In order to protect themselves, bacteria have also evolved the ability to “tag” their own DNA with methylation bases on certain sequences. For the same reason, foreign DNA, due to the absence of methyl groups on the same sequences, is melted (cut) into fragments by various bacterial restriction enzymes, and then degraded by bacterial exonucleases to zerootides. We can say that in this way bacteria protect themselves from the DNA of plants and animals, in whose bodies they live temporarily (as pathogens) or permanently (as saprophytes).
Restriction enzymes were first isolated from E. coli in 1968. It turned out that they are capable of cutting (melting) DNA molecules at different restriction sites (locations). These enzymes were called class I endonucleases. Then class II endonucleases were discovered in bacteria, which specifically recognize restriction sites in foreign DNA and also carry out restriction at these sites. It is enzymes of this class that began to be used in genetic engineering. At the same time, class III enzymes were discovered that melt DNA near recognition sites, but these enzymes are not important in genetic engineering.
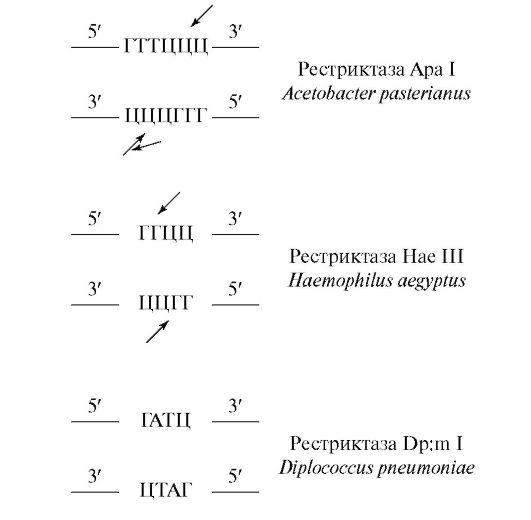
The action of the restriction-modification system is “rationalized” by the so-called palindromic (recognition) sequences of nitrogenous bases, which are DNA restriction sites. Palindromic sequences are sequences of bases that read the same way forward and backward, such as a sequence of letters radar. Since DNA strands have an antiparallel direction, a sequence is considered to be palindromic if it is identical when read in the direction from the 5" to the 3" end on the top and on the bottom strand from the 3" to the 5" end, namely :

Palindromes can be of any size, but most palindromes that are used as restriction enzyme recognition sites consist of 4, 5, 6, and rarely 8 bases.
Restriction enzymes are an absolutely necessary tool in genetic engineering for cutting out fragments of interest (genes) from large DNA molecules. Since more than 100 restriction enzymes are known, this allows the selection of restriction enzymes and selective excision of fragments from the original DNA.
A remarkable feature of restriction enzymes is that they cut molecules into several fragments (restrictions) of DNA with steps, as a result of which at the resulting ends one chain is longer than the other, forming a kind of tail. Such ends (tails) are called “sticky” ends, since they are capable of self-complementarity.
Let us consider the results of restriction using the example of one of the most well-known restriction enzymes Eco RI from the restriction modification system E. coI. Instead of melting the DNA at the center of the palindromic recognition sequence, this enzyme melts the DNA outside the center and produces 4 self-complementary (“sticky”) ends consisting of different numbers of nucleotides, namely:

These sticky ends are useful in genetic engineering experiments because they can be rejoined complementarily at low temperatures, allowing efficient closure of DNA fragments.
Recognition sites and melting sites in the case of other restriction enzymes have different contents, namely:
Following DNA restriction, restriction DNA fragments (DNA restriction fragments) are isolated from the restriction mixture, which are then necessary for combining with the vector. To isolate DNA restriction enzymes, electrophoresis is used, since with this method it is very easy to fractionate restricted DNA due to the size of the restriction fragments and constant electric charge-mass ratios. Fragments in an electric field migrate during electrophoresis at a frequency dependent on their size (mass). The larger (longer) the fragment, the slower it migrates in the electric field. The material used for electrophoresis is non-chargeable agarose or polyacrylamide. To identify fragments, ethidium bromide is used, which colors the fragments, which makes them easier to detect.
The efficiency of electrophoresis is very high, since it can be used to separate fragments whose sizes range from 2 to 50,000 bases.
After electrophoresis, fragments are isolated from agarose using various methods. Based on size comparison results
of restriction enzymes of the same DNA obtained using different restriction enzymes, restriction maps are constructed, which show the restriction sites of each of the restriction enzymes used. In practical terms, restriction maps make it possible to determine not only the size of restriction sites, but also to determine the location of loci of certain genes in DNA molecules.
Since in higher organisms, heterogeneous DNA is synthesized during transcription, which is corrected by processing, genetic engineering usually uses complementary DNA (cDNA), which is obtained by using mRNA as a template, on which reverse transcriptase synthesizes single-stranded DNA (cDNA), which is a copy of mRNA. These single-stranded DNAs are subsequently converted into double-stranded DNA. cDNA is considered to contain continuous nucleotide sequences (transcribed and translated). It is cDNA that is used for restriction.
DNA fragments (restrictions) isolated after electrophoresis from agarose gels can be preliminarily subjected to sequencing, i.e. determine their nucleotide sequence. For this purpose, chemical and enzymatic sequencing methods are used. The chemical method is based on obtaining fragments labeled with radioactive phosphorus (32 P) and removing one of the bases from these fragments, followed by taking into account the results of autoradiography of gels containing these fragments. The enzymatic method is based on the introduction of a nucleotide at the end of the analyzed fragment, which is then used in the synthesis of different fragments in vitro, analyzed for nucleotide sequence electrophoretically. To study specific nucleotide sequences in a DNA molecule, use
also hybridization of DNA-DNA, RNA-RNA, DNA-RNA, Northern
and Southern blots.
Genetic vectors. The DNA segment (gene) that is intended for molecular cloning must have the ability to replicate when transferred into a bacterial cell, i.e. be a replicon. However, he does not have such an ability. Therefore, to ensure the transfer and detection of cloned genes in cells, they are combined with so-called genetic vectors. The latter must have at least two properties. First, the vectors must be capable of replication
in cells, and at several ends. Secondly, they must provide the possibility of selecting cells containing the vector, i.e. have a marker that can be used to counter-select cells containing the vector along with the cloned gene (recombinant DNA molecules). Plasmids and phages meet these requirements. Plasmids are good vectors because they are replicons and can contain genes for resistance to any antibiotic, which allows the selection of bacteria for resistance to this antibiotic and, therefore, easy detection of recombinant DNA molecules
(Fig. 128).
Rice. 128. Vector pBRl
Since there are no natural plasmid vectors, all plasmid vectors known to date have been constructed artificially. The starting material for the creation of a number of genetic vectors was R-plasmids, in which excess DNA sequences, including those with multiple restriction sites, were removed using restriction enzymes. This deletion was determined by the fact that the plasmid vector should have only one recognition site for one restriction enzyme, and this site should lie in a functionally unimportant region of the plasmid genome. For example, the plasmid vector pBR 322, which has resistance genes to ampicillin and tetracycline, which makes it very convenient
for the selection of bacteria containing the cloned DNA segment, it has single restriction sites for more than 20 restriction enzymes, including such well-known restriction enzymes as Eco RI, Hind III, Pst I, Pva II and Sal I.
Phage vectors also have a number of advantages. They may include larger (longer) cloned DNA fragments compared to plasma vectors. Further, the transfer of the cloned fragment by phages into cells as a result of their infection of the latter is more effective than DNA transformation. Finally, phage vectors allow more efficient screening (recognition) on the agar surface of colonies containing cells carrying the gene being cloned. Many phage vectors are based on lambda phage.
In addition to phage ones, other viral vectors constructed on the basis of the herpes virus, as well as vectors constructed on the basis of yeast DNA, are also used.
If gene cloning is carried out using mammalian or plant cells, then the requirements for vectors are the same as in the case of cloning in bacterial cells.
Construction of recombinant DNA molecules. The direct construction of recombinant DNA molecules follows after the restrictions of the studied DNA and vector DNA are obtained. It consists in joining the restriction segments of the DNA under study with the restriction of the vector DNA, which, as a result of restriction, is converted from circular to linear DNA.
To connect fragments of the DNA under study with the vector DNA, DNA ligase is used (Fig. 129). Ligation will be successful if the interlocking structures have 3"-hydroxyl and 5"-phosphate groups and if these groups are positioned appropriately relative to one another. The fragments combine through their sticky ends as a result of self-complementarity. At high concentrations of fragments, the latter from time to time become in the correct position (opposite each other). Many restriction enzymes, such as EcoRI, produce sticky ends consisting of four bases. The process of ligation of “sticky” ends, consisting of four bases, occurs at a low temperature (up to 12? C).
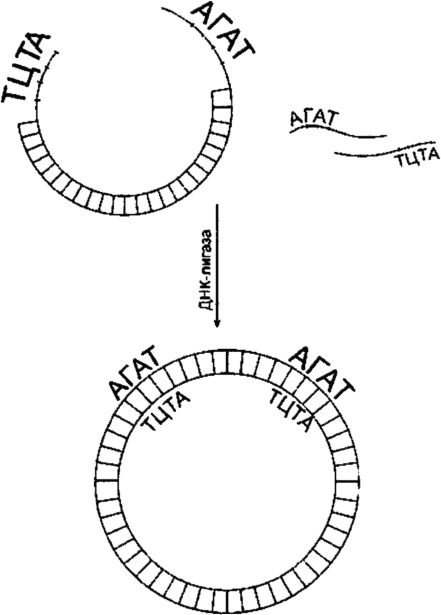
Rice. 129. DNA ligation
If restriction digestion produces fragments without sticky ends, they are “forcibly” converted into molecules with sticky ends using the enzyme transferase. This enzyme adds nucleotides to the 3" end of DNA. A poly-A tail may be added on one fragment and a poly-T tail on the other. Polymerase chain reaction (PCR) is also used to generate any desired DNA ends. The principle of PCR is based on on the denaturation of DNA isolated from cells and “annealing” it with the addition of DNA oligonucleotides consisting of 15-20 nucleotides each to the renaturating chains. These oligonucleotides must be complementary to the sequences in the chains separated by distances of 50-2000 nucleotides. Being a “seed” for DNA synthesis in vitro, they allow DNA polymerase to copy those sections that are located between the “primers”. This copying produces a large number of copies of the DNA fragment being studied.
Introduction of recombinant DNA molecules into cells. After the DNA fragment of interest (gene) is fused with a genetic vector using DNA ligase, the resulting recombinant molecules are introduced into cells in order to achieve their replication (due to the genetic vector) and increase the number of copies. The most popular way of introducing recombinant DNA molecules into cells, in which a plasmid serves as a vector, is transformation E. coli. For this purpose, bacterial cells are pre-treated with calcium or rubidium (ions), in order
so that they become “competent” in the perception of recombinant DNA. To increase the frequency of DNA penetration into cells, the electroporation method is used, which involves briefly exposing cells to an intense electric field. This treatment creates cavities in cell membranes, which allows cells to better perceive DNA. After introducing recombinant DNA molecules into bacteria, the latter are plated on MPA (meat peptone agar) enriched with antibiotics to select the desired cells, i.e. cells containing recombinant DNA molecules. The transformation frequency is low. Typically, one transformant appears per 10 5 seeded cells. If the vector is phage, then they resort to transfection of cells (bacteria or yeast) with the phage. As for animal somatic cells, they are transfected with DNA in the presence of chemicals that facilitate the passage of DNA through plasma membranes. Direct microinjections of DNA into oocytes, cultured somatic cells and mammalian embryos are also possible.
The most important point associated with molecular cloning is the search for a way to determine whether the cloned fragment is actually included in the vector and, together with the vector, forming a recombinant DNA molecule, enters the cells. If we are talking about bacterial cells, then one of the methods is based on taking into account insertional inactivation of the plasmid (vector) resistance gene. For example, in the plasmid vector pBR 322, which determines resistance to ampicillin and tetracycline, the only site for the Pst I restriction enzyme is located in the locus occupied by the ampicillin resistance gene. PstI fusion at this site generates sticky ends, allowing ligation of the cloned fragment to vector DNA. However, in this case, the plasmid (vector) ampicillin resistance gene is inactivated, while the tetracycline resistance gene on the vector remains intact. It is the tetracycline resistance gene that is used for the selection of cells transformed by recombinant DNA molecules. This makes it possible to ensure that the cells of colonies grown on a medium with tetracycline actually contain recombinant DNA molecules; they are checked using the so-called “spot test” on a pair of dishes with solid medium, one of which contains ampicillin, while the other is devoid of this antibiotic. The DNA to be cloned is
only in transformants resistant to tetracycline. As for transformants that are simultaneously resistant to ampicillin and tetracycline (ArTc), they contain plasmid (vector) molecules that spontaneously acquired a circular shape without the inclusion of foreign (clonable) DNA.
Another method for detecting the insertion of foreign (clonable) fragments into a plasmid vector is based on the use of a vector containing the β-galactosidase gene. Insertion of foreign DNA into this gene inevitably inactivates β-galactosidase synthesis, which can be detected by plating transformed cells on media that contains β-galactosidase substrates. This medium allows the selection of colored cell colonies. There are other methods.
As already noted, linear restriction fragments of vector DNA are capable of restoring the circular structure without including cloned segments. To reduce the frequency of spontaneous formation of such circular vector DNA molecules, vector DNA restrictors are treated with phosphatase. As a result of this, the formation of circular DNA molecules becomes impossible, since the 5"-PO 4 ends necessary for the action of the ligase will be absent.
The set of transformant colonies grown on a selective medium is a set of cells containing clones of different fragments (genes) of the cloned genomic or cDNA. Collections of these clones form so-called DNA libraries, widely used in genetic engineering work.
The final stage of gene cloning is the isolation and study of cloned DNA, including sequencing. Promising strains of bacteria or somatic cells containing recombinant DNA molecules that control the synthesis of proteins of interest that have commercial value are transferred to industry.
CELL ENGINEERING
As noted at the beginning of the chapter, cell engineering refers to the genetic manipulation of isolated animal and plant cells. These manipulations are often carried out in vitro, and their main goal is to obtain genotypes of these organisms with specified properties, primarily economically useful. As for-
Since the human being, cell engineering turned out to be applicable to his germ cells.
A prerequisite for the development of cell engineering in humans and animals was the development of methods for cultivating their somatic cells on artificial nutrient media, as well as obtaining hybrids of somatic cells, including interspecific hybrids. In turn, advances in the cultivation of somatic cells have influenced the study of germ cells and fertilization in humans and animals. Since the 60s. XX century In several laboratories around the world, numerous experiments were carried out on the transplantation of somatic cell nuclei into eggs artificially devoid of nuclei. The results of these experiments were often contradictory, but in general they led to the discovery of the ability of cell nuclei to ensure the normal development of the egg (see Chapter IV).
Based on the results of studying the development of fertilized eggs in the 60s. XX century Research was also begun to determine the possibility of fertilizing eggs outside the mother's body. Very quickly, these studies led to the discovery of the possibility of fertilizing eggs with sperm in vitro and the further development of embryos formed in this way when implanted in a woman’s uterus. Further improvement of the methods developed in this area has led to the fact that the birth of “test tube” children has become a reality. Already by 1981, 12 children were born in the world, whose life was given in the laboratory, in a test tube. Currently, this section of cell engineering has become widespread, and the number of “test tube” children is already tens of thousands (Fig. 130). In Russia, work on obtaining “test tube” children began only in 1986.
In 1993, a technique for producing monozygotic human twins was developed in vitro by dividing embryos into blastomeres and growing the latter to 32 cells, after which they could be implanted into a woman’s uterus.
Influenced by the results associated with the production of “test tube” children, a technology was also developed in animals, called transplantation embryos. It is associated with the development of a method for inducing polyovulation, methods for artificial fertilization of eggs and implantation of embryos into the body of animals - adoptive mothers. The essence of this technology comes down to the following:
shyu. A highly productive cow is injected with hormones, resulting in polyovulation, which involves the maturation of 10-20 cells at once. The eggs are then artificially fertilized with male reproductive cells in the oviduct. On the 7th-8th day, the embryos are washed out of the uterus and transplanted into the uteruses of other cows (foster mothers), who then give birth to twin calves. Calves inherit the genetic status of their original parents.

Rice. 130.Test tube children
Another area of animal cell engineering is the creation of transgenic animals. The simplest way to obtain such animals is to introduce linear DNA molecules into the eggs of the original animals. Animals that develop from eggs fertilized in this way will contain a copy of the introduced gene on one of their chromosomes and, in addition, they will pass on this gene to inheritance. A more complex method for producing transgenic animals was developed on mice that differ in coat color, and boils down to the following. First, four-day-old embryos are removed from the body of a pregnant gray mouse and crushed into individual cells. Then the nuclei are removed from the embryonic cells and transferred to the eggs of black mice, previously deprived of nuclei. Eggs of black mice containing foreign nuclei are placed in test tubes
with a nutrient solution for further development. The embryos developed from the eggs of black mice are implanted into the uteruses of white mice. Thus, in these experiments, it was possible to obtain a clone of mice with a gray coat color, i.e. clone embryonic cells with specified properties. In Chapter IV, we examined the results of fertilization of artificially denucleated sheep eggs with nuclear material from somatic cells of animals of the same species. In particular, the nuclei were removed from sheep eggs, and then the nuclei of somatic cells (embryonic, fetal or adult cells) were injected into such eggs, after which the eggs thus fertilized were injected into the uteri of adult sheep. The lambs born were identical to the donor ewe. An example is Dolly the sheep. Clonal calves, mice, rabbits, cats, mules and other animals were also obtained. Such construction of transgenic animals is a direct way of cloning animals with economically useful traits, including individuals of a certain sex.
Transgenic animals are also obtained using starting material belonging to different species. In particular, there is a known method of transferring the gene that controls growth hormone from rats into mouse eggs, as well as a method of combining sheep blastomeres with goat blastomeres, which led to the emergence of hybrid animals (sheep). These experiments indicate the possibility of overcoming species incompatibility at the earliest stages of development. Particularly attractive prospects open up (if species incompatibility is completely overcome) in the way of fertilization of eggs of one species with the nuclei of somatic cells of another species. We are talking about the real prospect of creating economically valuable animal hybrids that cannot be obtained by crossing.
It should be noted that nuclear transplantation work is not yet very effective. Experiments performed on amphibians and mammals have generally shown that their effectiveness is low, and it depends on the incompatibility between the donor nuclei and the recipient oocytes. In addition, chromosomal aberrations that form in the transplanted nuclei during further development, which are accompanied by the death of transgenic animals, are also an obstacle to success.
At the intersection of studies of cell hybridization and immunological research, a problem arose related to the production and study of so-called monoclonal antibodies. As noted above, antibodies produced by the body in response to the introduction of an antigen (bacteria, viruses, red blood cells, etc.) are proteins called immunoglobulins and form a fundamental part of the body's defense system against pathogens. But any foreign body introduced into the body is a mixture of different antigens that will stimulate the production of different antibodies. For example, human red blood cells possess antigens not only for blood groups A (II) and B (III), but also many other antigens, including the Rh factor. Further, proteins in the cell wall of bacteria or the capsid of viruses can also act as different antigens, causing the formation of different antibodies. At the same time, lymphoid cells of the body's immune system are usually represented by clones. This means that even for this reason alone, antibodies in the blood serum of immunized animals are always a mixture consisting of antibodies produced by cells of different clones. Meanwhile, for practical needs, antibodies of only one type are needed, i.e. so-called monospecific sera containing antibodies of only one type or, as they are called, monoclonal antibodies.
In search of methods for producing monoclonal antibodies, Swiss researchers in 1975 discovered a method of hybridization between lymphocytes of mice immunized with a particular antigen and cultured tumor cells of the bone marrow. Such hybrids are called “hybridoma”. From the “lymphocytic” part, represented by a lymphocyte of one clone, a single hybridoma inherits the ability to cause the formation of the necessary antibodies, of one type, and thanks to the “tumor (myeloma)” part, it becomes capable, like all tumor cells, of indefinitely multiplying on artificial nutrient media, giving a large population of hybrids. In Fig. 131 shows a diagram of the isolation of cell lines that synthesize monoclonal antibodies. Mouse monoclonal antibody cell lines are isolated by fusing myeloma cells with lymphocytes from the spleen of a mouse immunized five days previously.
desired antigen. Cell fusion is achieved by mixing them in the presence of polyethylene glycol, which induces the fusion of cell membranes, and then seeding them on a nutrient medium that allows the growth and reproduction of only hybrid cells (hybridoma). Hybridomas are propagated in a liquid medium, where they grow further and secrete antibodies into the culture liquid, of only one type, and in unlimited quantities. These antibodies are called monoclonal. To increase the frequency of antibody formation, they resort to cloning hybridomas, i.e. to the selection of individual hybridoma colonies capable of causing the formation of the largest number of antibodies of the desired type. Monoclonal antibodies have found widespread use in medicine for the diagnosis and treatment of a number of diseases. However, the most important advantage of monoclonal technology is that it can produce antibodies against materials that cannot be purified. On the contrary, monoclonal antibodies can be obtained against the cell (plasma) membranes of animal neurons. To do this, mice are immunized with isolated neuronal membranes, after which their splenic lymphocytes are combined with myeloma cells, and then proceed as described above.

Rice. 131. Obtaining monoclonal antibodies
GENETIC ENGINEERING AND MEDICINE
Genetic engineering has proven to be very promising for medicine, primarily in the creation of new technologies for the production of physiologically active proteins used as medicines (insulin, somatostatin, interferons, somatotropin, etc.).
Insulin is used to treat patients with diabetes, which is the third most common cause of death (after heart disease and cancer). The global need for insulin is several tens of kilograms. Traditionally, it is obtained from the pancreatic glands of pigs and cows, but the hormones of these animals are slightly different from human insulin. Pig insulin differs in one amino acid, while cow insulin differs in three. It is believed that animal insulin often causes side effects. Although the chemical synthesis of insulin has been carried out for a long time, until now the industrial production of hormones has remained very expensive. Now cheap insulin is produced using the genetic engineering method by chemical-enzymatic synthesis of the insulin gene, followed by the introduction of this gene into Escherichia coli, which then synthesizes the hormone. This insulin is more “biological”, since it is chemically identical to the insulin produced by human pancreatic cells.
Interferons are proteins synthesized by cells mainly in response to infection of the body by viruses. Interferons are characterized by species specificity. For example, in humans there are three groups of interferons produced by different cells under the control of the corresponding genes. Interest in interferons is determined by the fact that they are widely used in clinical practice to treat many human diseases, especially viral ones.
Being large in size, interferon molecules are not easily accessible for synthesis. Therefore, most interferons are now obtained from human blood, but the yield from this method of production is small. Meanwhile, the need for interferon is extremely high. This set the task of finding an effective method for producing interferon in industrial quantities. Genetic engineering underlies the modern production of “bacterial” interferon.
The influence of genetic engineering on the technology of those medicinal substances that have long been created using biological technology has increased. Back in the 40-50s. XX century was created
biological industry for the production of antibiotics, which constitute the most effective part of the medicinal arsenal of modern medicine. However, in recent years there has been a significant increase in bacterial drug resistance, especially to antibiotics. The reason is the widespread distribution in the microbial world of plasmids that determine the drug resistance of bacteria. This is why many previously famous antibiotics have lost their former effectiveness. The only way to overcome bacterial resistance to antibiotics so far is to search for new antibiotics. According to experts, about 300 new antibiotics are created every year in the world. However, most of them are either ineffective or toxic. Only a few antibiotics are introduced into practice every year, which forces us not only to maintain, but also to increase the capacity of the antibiotic industry based on genetic engineering developments.
The main tasks of genetic engineering in those technologies of medicinal substances in which microorganisms are drug producers are determined by the need for genetic engineering reconstruction of the latter in order to increase their activity. At the same
Since then, the idea of creating drugs in the form of small molecules has begun to be implemented, which contributes to their greater effectiveness.
Immune biotechnology is primarily associated with the production of new generation vaccines for the prevention of infectious diseases in humans and animals. The first commercial products created using genetic engineering were vaccines against human hepatitis, animal foot-and-mouth disease, and some others. An extremely important direction in this area is associated with the production of monoclonal antibodies, reagents necessary for the diagnosis of pathogens, as well as for the purification of hormones, vitamins, proteins of various natures (enzymes, toxins, etc.).
Of significant practical interest is the method of producing artificial hemoglobin by introducing hemoglobin genes into tobacco plants, where, under the control of these genes, α- and β-chains of globin are produced, which are combined into hemoglobin. Hemoglobin synthesized in the cells of tobacco plants is fully functional (binds oxygen). Cellular engineering, as applied to humans, is associated not only with solving fundamental problems of human biology, but also with overcoming, first of all, female infertility. Since the frequency of positive cases of implantation of embryos obtained into the uterus of women in vitro, is small, then obtaining monozygotic twin embryos in vitro also matters, since the possibilities of repeated implantations due to “spare” embryos increase. Of particular interest are the prospects for using stem cells as a source of cell and tissue replacement in the treatment of diseases such as diabetes, spinal cord injuries, heart pain, osteoarthritis, and Parkinson's disease. But to realize these prospects, an in-depth study of stem cell biology is necessary.
In the use of genetic engineering in relation to medical problems, the task of developing genetic engineering methods for the radical treatment of hereditary diseases, which, unfortunately, cannot yet be treated with existing methods, has acquired particular importance. The content of this task is to develop ways to correct (normalize) mutations that result in hereditary diseases, and to ensure the transmission of “corrections” by inheritance. It is believed that the successful development of genetic engineering methods for the treatment of hereditary diseases will be
contribute to data on the human genome obtained as a result of the international scientific program “Human Genome”.
ECOLOGICAL PROBLEMS OF GENETIC ENGINEERING
Taking biotechnology to a new level, genetic engineering has also found application in developing ways to identify and eliminate environmental pollutants. In particular, strains of bacteria have been constructed that are unique indicators of the mutagenic activity of chemical contaminants. On the other hand, bacterial strains have been genetically engineered to contain plasmids, under the control of which the synthesis of enzymes occurs that are capable of destroying many chemical compounds that pollute the environment. In particular, some plasmid-containing bacteria are capable of decomposing oil and petroleum products into harmless compounds that have ended up in the environment as a result of various accidents or other unfavorable reasons.
However, genetic engineering is the transformation of genetic material that does not exist in nature. Consequently, genetic engineering products are completely new products that do not exist in nature. Therefore, due to the unknown nature of its products, it itself is fraught with danger both for nature and the environment, and for personnel working in laboratories where they use genetic engineering methods or work with structures created during genetic engineering work.
Since the possibilities of gene cloning are limitless, even at the very beginning of these studies, questions arose among scientists about the nature of the organisms being created. At the same time, a number of undesirable consequences of this methodology were suggested, and these assumptions also found support among the general public. In particular, disagreements have arisen about the properties of bacteria that received animal genes in genetic engineering experiments. For example, do bacteria retain E. coli their species identity due to the content of introduced genes of animal origin (for example, the insulin gene) or should they be considered a new species? Further, how durable are such bacteria, in what ecological niches can they
exist? But the most important thing began to be the emergence of concerns that during the production and manipulation of recombinant DNA molecules, genetic structures with properties that were unforeseen and dangerous to human health could be created for the historically established ecological balance. At the same time, calls for a moratorium on genetic engineering began. These calls caused an international outcry and led to an international conference, which took place in 1975 in the United States, at which the possible implications of research in this area were widely discussed. Then, in countries where genetic engineering began to develop, rules for working with recombinant DNA molecules were developed. These rules are aimed at preventing the products of genetic engineering laboratories from entering the environment.
Another aspect of the undesirable consequences of genetic engineering work is associated with the danger to the health of personnel working in laboratories where genetic engineering methods are used, since such laboratories use phenol, ethidium bromide, and UV radiation, which are factors harmful to health. In addition, in these laboratories there is the possibility of contamination by bacteria containing recombinant DNA molecules that control undesirable properties, such as drug resistance of bacteria. These and other points determine the need to improve the level of safety in genetic engineering work.
Finally, the problems of the dangers of genetically modified products (genetically modified tomatoes, potatoes, corn, soybeans), as well as products such as bread, pastes, candies, ice cream, cheese, vegetable oil, meat products, which in some countries, especially in the USA, have become widespread. For 12,000 years of agriculture, humans have consumed foods that come from naturally occurring sources. Therefore, it is assumed that genetically modified food will introduce new toxins, allergens, bacteria, and carcinogens into the human body, which will lead to completely new diseases for future generations. This raises the question of a truly scientific assessment of genetically modified food.
ISSUES FOR DISCUSSION
1. What is meant by genetic, cellular and genetic engineering? Is there a difference between these concepts and molecular cloning?
2. What is the progressiveness of genetic engineering compared to other methods used in biology?
3. List the main “tools” of genetic engineering.
4. What are restriction enzymes, what are their properties and their role in genetic engineering?
5. Do all restriction enzymes form “sticky” ends of the DNA being studied, and does the structure of the “sticky” ends depend on the type of restriction enzyme?
6. Define genetic vectors. Are there natural vectors?
7. How are genetic vectors obtained in the laboratory? What biological objects are the starting material for obtaining vectors?
8. What is the maximum length of sequences of DNA nitrogenous bases that can still be included in a genetic vector? Do vectors differ in “power”?
9. Characterize the properties of DNA ligase and determine its role in genetic engineering.
10. How is the cloned DNA segment (gene) connected to the genetic vector?
11. What is the frequency of introduction of recombinant DNA molecules into bacterial cells?
12. On what principle is the selection of bacterial cells containing recombinant DNA molecules based? Give one example of such selection.
14. Many strains of bacteria have the same enzymes that ensure their metabolism in almost the same way. Meanwhile, the nucleotide specificity of bacterial restriction-modification systems is different. Can you explain this phenomenon?
15. Why cannot DNA sequences representing restriction enzyme recognition sites contain more than eight base pairs?
16. How many times will the sequence HGC, recognized by the restriction enzyme Hae III, occur in a 50,000 base pair DNA segment with 30, 50, and 70 percent GC content?
17. Restriction enzymes Bam HI and Bgl I melt the sequences G GATCC and T GATCA, respectively. Is it possible to include DNA fragments produced by Bgl I restriction into the Bam HI site? If so, why? If the plasmid (vector) used contains one Bgl I restriction site, then on what nutrient medium can this plasmid be selected for bacteria?
18. Calculate the frequency of bacterial transformation per DNA molecule if 5-10 5 transformants are formed per 5000 plasmid base pairs?
19. Is it possible to clone DNA replication point 0? E. coli and if so, how?
20. Is it possible to determine how many DNA molecules are needed to transform one cell? E. coli?
21. Is it possible to determine the splice site on mRNA using polymerase chain reaction?
22. How can polymerase chain reaction be used to introduce a desired restriction site into a location of interest on a DNA fragment to be cloned?
23. Name the methods of cell engineering as applied to animals. What is the economic value of animals produced by these methods?
24. Define the concepts “transgenic plants” and “transgenic animals”. Do transgenic organisms retain their species identity or can they be considered organisms of new species?
25. What are hybridomas and monoclonal antibodies? How do you get them?
26. Is cell engineering applicable to humans?
27. Let us assume that the injection of foreign DNA into a mouse egg and the implantation of the egg fertilized in this way into the mouse’s body resulted in pregnancy and the birth of mice containing copies of the injected DNA in the genome. However, the little mice turned out to be mosaics, i.e. Some of their cells contain copies of the injected DNA, others lack this DNA. Can you explain the nature of this phenomenon?
28. Do you consider food prepared from genetically modified products to be genetically dangerous?
29. Is scientific testing of genetically modified food necessary?
Knowledge is determined by what we affirm as Truth.
P.A. Florensky, 1923
Genetic (genetic) engineering
Genetic (genetic) engineering– artificial construction of genetic structures and hereditarily modified organisms. Genetic engineering is a section (applied branch) of molecular genetics associated with the targeted creation of new DNA molecules capable of multiplying in a host cell. In this case, an artificial, purposeful change in the genotype of the organism (microorganism) occurs and the formation of new characteristics and properties. Genetic engineering deals with decoding the structure of genes, their synthesis and cloning, and the insertion of genes isolated from the cells of living organisms into the cells of plants and animals in order to specifically change their genetic characteristics.
Well-developed methods of genetic engineering are transgenesis, microbiological synthesis, etc.
Transgenesis– transfer of genes from one type of organism to another. Transgenesis is carried out by cutting and stitching sections of DNA with the participation of enzymes - restriction enzymes and ligases.
Stages of transgenesis:
a) isolation of genes (DNA fragments) from bacterial, plant or animal cells using an enzyme restriction enzymes;
b) connection (linking) of genes (DNA fragments) with a plasmid using an enzyme ligases;
c) introduction of a hybrid plasmid DNA containing the desired gene into the host cell;
d) copying (cloning) this gene in the host cell and ensuring its operation according to the scheme: “DNA code – transcription – translation – protein”
Genetic engineering tools are enzymes discovered in 1974 - restriction enzymes (restriction endonucleases). Restriction enzymes recognize sections (sites) of DNA and make cuts in DNA strands. At the ends of each fragment, single-stranded tails are formed, called “ sticky ends" since they can, as it were, stick together due to complementarity.
Restriction enzymes recognize a specific sequence of DNA nucleotides in double-stranded DNA. The restriction enzyme then attaches to the recognized nucleotide site and cuts it at the site of attachment. More often, restriction enzymes recognize regions of 4–6 nucleotide pairs in a DNA molecule and cut both DNA strands in the middle of these regions or usually with an offset. Examples of restriction enzymes: restriction enzyme Eco RI, which recognizes a DNA fragment of six nucleotides GAATTC (the cut site between nucleotides G and A of both DNA strands); restriction enzyme Hind III recognizes the AAGCTT region (the cut site between nucleotides A and A of both DNA strands); restriction enzyme Bam I recognizes the GGATCC region (the cut site between nucleotides G and G of both DNA strands); restriction enzyme Hae III recognizes the GGC site (the cut site between the G and C nucleotides of both DNA strands); restriction enzyme Hpa II recognizes the CCGG region (the site of the cut between the C and C nucleotides of both DNA strands).
Next, to construct a genetically modified organism, it is necessary to introduce the desired gene into the cell of this organism. The introduction of foreign genes into the body is carried out using plasmid vector. The vector is plasmid – small circular DNA molecule which is extracted from the cytoplasm of a bacterial cell. Plasmids– factors of heredity located outside the chromosomes, representing extrachromosomal DNA.

Rice. 37.
A– Scheme for introducing foreign DNA into a bacterial plasmid using enzymes (restriction endonuclease and ligase).
B– Scheme of human gene transfer responsible for the synthesis of the hormone insulin and the formation of vector DNA.
Properties of the plasmid: 1) has the ability for autonomous replication; 2) contains genes encoding antibiotics; 3) are able to integrate into the chromosome of the recipient cell; 4) recognizes sections of DNA that can be cut by restriction enzymes; 5) a restriction enzyme can cut the plasmid and transfer it to a linear state. Researchers use these properties of the plasmid to obtain recombinant (hybrid) DNA.
The sequence of introducing DNA into a plasmid (plasmid vector) using a restriction enzyme(Fig. 37 A):
1) restriction– cutting of the DNA molecule with a restriction enzyme, the formation of DNA fragments and isolation of the required gene;
2) inclusion of the isolated gene into a plasmid, i.e., obtaining recombinant (hybrid) DNA by introducing a fragment of foreign DNA into a plasmid;
3) ligation– enzyme cross-linking ligase plasmid (vector) and foreign DNA fragments; in this case, the ends of the vector and foreign DNA (the so-called “sticky ends”) are complementary to each other;
4) transformation– introduction of a recombinant plasmid into the genome of another cell (recipient cell), in particular a bacterial cell.
It should be noted that plasmids penetrate only a portion of the treated bacteria. Transformed bacteria, together with plasmids, acquire resistance to a specific antibiotic, which makes it possible to separate them from non-transformed bacteria that die on a medium containing an antibiotic. Each of the transformed bacteria, placed on a nutrient medium, multiplies and forms a colony of many thousands of descendants - a clone.
5) screening– selection among transformed bacteria of those that contain plasmids with the desired gene.
Transgenic animals and plants
Cloned genes are introduced into mammalian eggs or plant protoplasts (an isolated cell without a cell wall) using microinjection, and then animals or plants are grown from them, in whose genome foreign genes operate. Plants and animals whose genomes have been altered through genetic engineering operations are called transgenic organizations (transgenic plants and animals), because it contains foreign genes. Transgenic mice, rabbits, pigs, and sheep were obtained. Their genome contains genes from bacteria, mammals, and humans. Transgenic plants (corn, peppers, tomatoes, wheat, rye, legumes, potatoes, etc.) containing genes from unrelated species have been obtained. Transgenic plants are resistant to herbicides, insects, unfavorable rain conditions, etc. The problem of changing the heredity of many agricultural plants is being gradually solved.
Genetic map of chromosomes. Gene therapy
A genetic map of chromosomes is a diagram of the relative arrangement of genes located in the same linkage group. Such maps are compiled for each pair of homologous chromosomes. The genetic map shows the order of genes on the chromosome and the distance between them (the percentage of crossing over between certain genes). Thus, the creation of new strains of microorganisms capable of synthesizing hormones, proteins, and drugs is based on knowledge of the genetic maps of microorganisms. Human genetic maps are essential for medical genetics. Knowledge about the localization of a gene on a specific chromosome is used in the diagnosis of a number of hereditary diseases, as well as in gene therapy to correct the structure and function of genes.
Gene therapy – replacing defective genes with intact ones, or correcting their structure.
To combat hereditary, oncological and age-related diseases, gene therapy methods that are safe for human cells are being developed. Using gene therapy methods, it is possible to replace defective genes in the body in which point mutations have occurred with intact ones. Nowadays, scientists are mastering methods human biosafety: introduction of the necessary genes into the cells of the human body. This will allow you to get rid of many hereditary diseases.
Microbiological synthesis
Genetic engineering methods have made it possible to implement microbiological synthesis(Fig. 37 B). Using genetic engineering methods, microbiologists were able to obtain strains of bacteria, thanks to which microbiological synthesis is successfully carried out. To do this, the necessary bacterial cells that do not contain plasmids are selected. DNA molecules with a given sequence of nucleotides are isolated, which determine the development of the desired trait. A plasmid with an integrated DNA section (genome) is introduced into a bacterial cell, in which the built-in DNA section begins to work (replication, transcription, translation processes take place), and the necessary protein (interferon, geneferon, immunoglobulin, insulin, somatotropin, etc.) is synthesized in the bacterial cell. ). Hormones (insulin, somatotropin), many amino acids, antibiotics, vaccines, etc. are obtained in industrial quantities. Such bacteria are multiplied on an industrial scale and produce the necessary protein.
Using genetic methods, a strain of the microorganism Pseudomonas denitrificans was obtained, which produces tens of times more vitamin C and B vitamins than the original form; a new strain of the bacterium Micrococcus glutamicus secretes hundreds of times more of the amino acid lysine than the original (wild) culture of the lysine-producing bacterium.
Cell engineering
Cell engineering– cultivation of individual cells or tissues in special artificial media, development of methods for creating a new type of cells by hybridizing them, replacing chromosomes and growing hybrids from them.
1. Tissue culture method
The method consists of cultivating isolated cells or pieces of tissue on an artificial nutrient medium under appropriate microclimatic conditions. As a result of cultivation, plant cells or pieces of tissue are regenerated into a whole plant. By microclonal propagation of individual cells or pieces of tissue (usually the apical meristem of a stem or root), many useful plants can be obtained. Microclimatic conditions and nutrient media for the regeneration of ornamental, cultural, and medicinal plants are selected experimentally. Tissue culture is also used to produce diploid plants after treating the original haploid forms with colchicine.
2. Somatic hybridization
Somatic hybridization includes the production of hybrid cells, and from them - new forms; artificial fertilization of eggs.
Obtaining new hybrid plants by fusion of protoplasts (nucleus and cytoplasm) of various cells in tissue culture. To fuse protoplasts, the plant cell wall is destroyed with the help of enzymes and an isolated protoplast is obtained. When such protoplasts of different plant species are cultivated, they merge and form forms with new useful characteristics. Artificial fertilization of eggs is carried out using the method of in vitro fertilization (IVF), which allows fertilization of eggs in vitro with subsequent implantation of the embryo at an early stage of development, and to overcome some forms of infertility in humans.
3. Chromosome engineering– replacement of individual chromosomes in plant cells or addition of new ones. Diploids have pairs of homologous chromosomes, and such organisms are called disomics. If one chromosome is left in any one pair, then a monosomy is formed. If you add a third homologous chromosome to any pair, a trisomic is formed, etc. It is possible to replace individual chromosomes of one species with chromosomes of another species. Received forms are called substituted.
Genetic engineering serves to obtain the desired qualities of a changeable or genetically modified organism. Unlike traditional selection, during which the genotype is subject to changes only indirectly, genetic engineering allows direct intervention in the genetic apparatus using the technique of molecular cloning. Examples of the application of genetic engineering are the production of new genetically modified varieties of grain crops, the production of human insulin using genetically modified bacteria, the production of erythropoietin in cell culture or new breeds of experimental mice for scientific research.
The first experiments are being conducted on the use of bacteria with rearranged DNA to treat patients.
The basis of the microbiological, biosynthetic industry is the bacterial cell. The cells necessary for industrial production are selected according to certain characteristics, the most important of which is the ability to produce, synthesize, in the maximum possible quantities, a certain compound - an amino acid or an antibiotic, a steroid hormone or an organic acid. Sometimes you need to have a microorganism that can, for example, use oil or wastewater as “food” and process it into biomass or even protein quite suitable for feed additives. Sometimes we need organisms that can develop at elevated temperatures or in the presence of substances that are certainly lethal to other types of microorganisms.
The task of obtaining such industrial strains is very important; for their modification and selection, numerous methods of actively influencing the cell have been developed - from treatment with potent poisons to radioactive irradiation. The goal of these techniques is one - to achieve changes in the hereditary, genetic apparatus of the cell. Their result is the production of numerous mutant microbes, from hundreds and thousands of which scientists then try to select the most suitable for a particular purpose. The creation of techniques for chemical or radiation mutagenesis was an outstanding achievement in biology and is widely used in modern biotechnology.
But their capabilities are limited by the nature of the microorganisms themselves. They are not able to synthesize a number of valuable substances that accumulate in plants, primarily in medicinal and essential oil plants. They cannot synthesize substances that are very important for the life of animals and humans, a number of enzymes, peptide hormones, immune proteins, interferons, and many simpler compounds that are synthesized in the bodies of animals and humans. Of course, the possibilities of microorganisms are far from exhausted. Of the entire abundance of microorganisms, only a tiny fraction has been used by science, and especially by industry. For the purposes of selection of microorganisms, of great interest are, for example, anaerobic bacteria that can live in the absence of oxygen, phototrophs that use light energy like plants, chemoautotrophs, thermophilic bacteria that can live at temperatures, as recently discovered, about 110 ° C, etc.
And yet the limitations of “natural material” are obvious. They have tried and are trying to get around the restrictions with the help of cell and tissue cultures of plants and animals. This is a very important and promising path, which is also being implemented in biotechnology. Over the past few decades, scientists have developed methods by which individual tissue cells of a plant or animal can be made to grow and reproduce separately from the body, like bacterial cells. This was an important achievement - the resulting cell cultures are used for experiments and for the industrial production of certain substances that cannot be obtained using bacterial cultures.
Another direction of research is the removal from DNA of genes that are unnecessary for coding proteins and the functioning of organisms and the creation of artificial organisms with a “truncated set” of genes based on such DNA. This makes it possible to dramatically increase the resistance of modified organisms to viruses.
History of development and methods
In the second half of the 20th century, several important discoveries and inventions were made that underlie genetic engineering. Many years of attempts to “read” the biological information that is “written” in genes have been successfully completed. This work was started by the English scientist Frederick Sanger and the American scientist Walter Gilbert (Nobel Prize in Chemistry 1980). As is known, genes contain information-instructions for the synthesis of RNA molecules and proteins, including enzymes, in the body. To force a cell to synthesize new substances that are unusual for it, it is necessary that the corresponding sets of enzymes be synthesized in it. And for this it is necessary to either purposefully change the genes located in it, or introduce new, previously absent genes into it. Changes in genes in living cells are mutations. They occur under the influence, for example, of mutagens - chemical poisons or radiation. But such changes cannot be controlled or directed. Therefore, scientists have focused their efforts on trying to develop methods for introducing new, very specific genes needed by humans into cells.
All methods of genetic engineering Genetic engineering techniques ) are used to implement one of the following stages of solving a genetic engineering problem:
- Obtaining an isolated gene.
- Introduction of a gene into a vector for transfer into the body.
- Transfer of a vector with a gene into the modified organism.
- Transformation of body cells.
- Selection of genetically modified organisms ( GMO) and eliminating those that were not successfully modified.
The process of gene synthesis is now very well developed and even largely automated. There are special devices equipped with computers, in the memory of which programs for the synthesis of various nucleotide sequences are stored. This apparatus synthesizes DNA segments up to 100-120 nitrogen bases in length (oligonucleotides). A technique has become widespread that makes it possible to use the polymerase chain reaction to synthesize DNA, including mutant DNA. A thermostable enzyme, DNA polymerase, is used in it for template DNA synthesis, for which artificially synthesized pieces of nucleic acid - oligonucleotides - are used as seeds. The enzyme reverse transcriptase allows, using such primers, to synthesize DNA on a template of RNA isolated from cells. The DNA synthesized in this way is called complementary DNA (RNA) or cDNA. An isolated, "chemically pure" gene can also be obtained from a phage library. This is the name of a bacteriophage preparation, into the genome of which random fragments from the genome or cDNA are built in, reproduced by the phage along with all its DNA.
The technique of introducing genes into bacteria was developed after Frederick Griffith discovered the phenomenon of bacterial transformation. This phenomenon is based on a primitive sexual process, which in bacteria is accompanied by the exchange of small fragments of non-chromosomal DNA, plasmids. Plasmid technologies formed the basis for the introduction of artificial genes into bacterial cells.
Significant difficulties were associated with the introduction of a ready-made gene into the hereditary apparatus of plant and animal cells. However, in nature there are cases when foreign DNA (of a virus or bacteriophage) is included in the genetic apparatus of a cell and, with the help of its metabolic mechanisms, begins to synthesize “its” protein. Scientists studied the features of the introduction of foreign DNA and used it as a principle for introducing genetic material into a cell. This process is called transfection.
If unicellular organisms or multicellular cell cultures are subject to modification, then at this stage cloning begins, that is, the selection of those organisms and their descendants (clones) that have undergone modification. When the task is to obtain multicellular organisms, cells with an altered genotype are used for vegetative propagation of plants or introduced into the blastocysts of a surrogate mother when it comes to animals. As a result, cubs are born with a changed or unchanged genotype, among which only those that exhibit the expected changes are selected and crossed with each other.
Application in scientific research
Although on a small scale, genetic engineering is already being used to give women with some types of infertility a chance to get pregnant. For this purpose, eggs from a healthy woman are used. As a result, the child inherits the genotype from one father and two mothers.
However, the possibility of making more significant changes to the human genome faces a number of serious ethical problems. In 2016, in the United States, a group of scientists received approval for clinical trials of a method of treating cancer using the patient’s own immune cells, subjected to genetic modification using CRISPR / Cas9 technology.
At the end of 2018, two children were born in China, whose genome was artificially changed (the CCR5 gene was turned off) at the embryonic stage using the CRISPR/Cas9 method, as part of research conducted since 2016 to combat HIV. One of the parents (father) was HIV-infected , and the children, according to the statement, were born healthy. Since the experiment was unauthorized (previously, all such experiments on human embryos were allowed only in the early stages of development with the subsequent destruction of the experimental material, that is, without implantation of the embryo into the uterus and the birth of children), the scientist responsible for it did not provide evidence for his statements that were made at the international conference on genome editing. At the end of January 2019, the Chinese authorities officially confirmed the facts of this experiment. Meanwhile, the scientist was prohibited from engaging in scientific activities and he was arrested.
Cell engineering
Cellular engineering is based on the cultivation of plant and animal cells and tissues capable of producing substances necessary for humans outside the body. This method is used for clonal (asexual) propagation of valuable plant forms; to obtain hybrid cells that combine the properties of, for example, blood lymphocytes and tumor cells, which makes it possible to quickly obtain antibodies.
Genetic engineering in Russia
It is noted that after the introduction of state registration of GMOs, the activity of some public organizations and individual State Duma deputies, trying to prevent the introduction of innovative biotechnologies in Russian agriculture, has noticeably increased. More than 350 Russian scientists signed an open letter from the Society of Scientific Workers in support of the development of genetic engineering in the Russian Federation. The open letter notes that a ban on GMOs in Russia will not only harm healthy competition in the agricultural market, but will lead to a significant lag in food production technologies, increased dependence on food imports, and will undermine the prestige of Russia as a state in which a course towards innovative development has been officially announced [ significance of the fact? ] .
see also
Notes
- Alexander Panchin Beating God // Popular Mechanics. - 2017. - No. 3. - P. 32-35. - URL: http://www.popmech.ru/magazine/2017/173-issue/
- Olga Volkova. 12 methods in pictures: genetic engineering. Part I, historical (Russian). Biomolecule. Retrieved March 25, 2019.
- Michael Waldholtz Transformers // In the world of science. - 2017. - No. 5-6. - P. 126 - 135.
- In vivo genome editing using a high-efficiency TALEN system(English) . Nature. Retrieved January 10, 2017.
- Elements - science news: Monkeys cured of color blindness using gene therapy (undefined) (September 18, 2009). Retrieved January 10, 2017.
- Transgenic monkeys give birth to first offspring (undefined) . membrana (May 29, 2009). Retrieved January 10, 2017.
- Genetically altered babies born (undefined) . BBC. Retrieved April 26, 2008. Archived August 22, 2011.
- B. Alberts, A. Johnson, J. Lewis, M. Raff, K. Roberts, P. Walter, 2008. “Molecular biology of the cell,” 5th ed., Garland Science, USA, pp. 1302-1303
- Kimmelman J. (2009) “Ethics of cancer gene transfer clinical research,” Methods in Molecular Biology 542, 423-445
- Wagner AM, Schoeberlein A, Surbek D. (2009) "Fetal gene therapy: opportunities and risks", Advanced Drug Delivery Reviews 61, 813-821
- Gatzidou E, Gatzidou G, Theocharis SE. (2009) “Genetically transformed world records: a reality or in the sphere of fantasy?”, Medical Science Monitor 15, RA41-47
- Lowenstein PR. (2008) “Clinical trials in gene therapy: ethics of informed consent and the future of experimental medicine,” Current Opinion in Molecular Therapy 10, 428-430
1. Possibilities of genetic engineering. 4
2. History of genetic engineering. 6
3. Genetic engineering as a science. Genetic engineering methods. 10
4. Areas of application of genetic engineering. 12
5. Scientific facts about the dangers of genetic engineering. 18
Conclusion. 22
References.. 23
Introduction
The topic of genetic engineering has recently become increasingly popular. Most attention is paid to the negative consequences that the development of this branch of science can lead to, and very little coverage is given to the benefits that genetic engineering can bring.
The most promising area of application is the production of drugs using genetic engineering technologies. Recently, it has become possible to obtain useful vaccines based on transgenic plants. Of no less interest is the production of food products using the same technologies.
Genetic engineering is the science of the future. At the moment, all over the world, millions of hectares of land are sown with transgenic plants, unique medical preparations and new producers of useful substances are being created. Over time, genetic engineering will make it possible to achieve new advances in medicine, agriculture, the food industry and animal husbandry.
The purpose of this work is to study the features of the possibility, history of development and areas of application of genetic engineering.
1. Possibilities of genetic engineering
An important part of biotechnology is genetic engineering. Born in the early 70s, she has achieved great success today. Genetic engineering techniques transform bacterial, yeast and mammalian cells into “factories” for the large-scale production of any protein. This makes it possible to analyze in detail the structure and functions of proteins and use them as medicines. Currently, Escherichia coli (E. coli) has become a supplier of such important hormones as insulin and somatotropin. Previously, insulin was obtained from animal pancreatic cells, so its cost was very high. To obtain 100 g of crystalline insulin, 800-1000 kg of pancreas is required, and one gland of a cow weighs 200 - 250 grams. This made insulin expensive and difficult to access for a wide range of diabetics. In 1978, researchers from Genentech first produced insulin in a specially engineered strain of Escherichia coli. Insulin consists of two polypeptide chains A and B, 20 and 30 amino acids long. When they are connected by disulfide bonds, native double-chain insulin is formed. It has been shown that it does not contain E. coli proteins, endotoxins and other impurities, does not produce side effects like animal insulin, and has no biological activity
is different. Subsequently, proinsulin was synthesized in E. coli cells, for which a DNA copy was synthesized on an RNA template using reverse transcriptase. After purifying the resulting proinsulin, it was split into native insulin, while the stages of extraction and isolation of the hormone were minimized. From 1000 liters of culture fluid, up to 200 grams of the hormone can be obtained, which is equivalent to the amount of insulin secreted from 1600 kg of the pancreas of a pig or cow.
Somatotropin is a human growth hormone secreted by the pituitary gland. A deficiency of this hormone leads to pituitary dwarfism. If somatotropin is administered in doses of 10 mg per kg of body weight three times a week, then in a year a child suffering from its deficiency can grow 6 cm. Previously, it was obtained from cadaveric material, from one corpse: 4 - 6 mg of somatotropin in terms of final pharmaceutical product. Thus, the available quantities of the hormone were limited, in addition, the hormone obtained by this method was heterogeneous and could contain slow-growing viruses. In 1980, the Genentec company developed a technology for the production of somatotropin using bacteria, which was devoid of these disadvantages. In 1982, human growth hormone was obtained in culture of E. coli and animal cells at the Pasteur Institute in France, and in 1984, industrial production of insulin began in the USSR. In the production of interferon, both E. coli, S. cerevisae (yeast), and a culture of fibroblasts or transformed leukocytes are used. Safe and cheap vaccines are also obtained using similar methods.
Recombinant DNA technology is based on the production of highly specific DNA probes, which are used to study the expression of genes in tissues, the localization of genes on chromosomes, and identify genes with related functions (for example, in humans and chicken). DNA probes are also used in the diagnosis of various diseases.
Recombinant DNA technology has made possible an unconventional protein-gene approach called reverse genetics. In this approach, a protein is isolated from a cell, the gene for this protein is cloned, and it is modified, creating a mutant gene encoding an altered form of the protein. The resulting gene is introduced into the cell. If it is expressed, the cell carrying it and its descendants will synthesize the altered protein. In this way, defective genes can be corrected and hereditary diseases can be treated.
If the hybrid DNA is introduced into a fertilized egg, transgenic organisms can be produced that express the mutant gene and pass it on to their offspring. Genetic transformation of animals makes it possible to establish the role of individual genes and their protein products both in the regulation of the activity of other genes and in various pathological processes. With the help of genetic engineering, lines of animals resistant to viral diseases have been created, as well as breeds of animals with traits beneficial to humans. For example, microinjection of recombinant DNA containing the bovine somatotropin gene into a rabbit zygote made it possible to obtain a transgenic animal with hyperproduction of this hormone. The resulting animals had pronounced acromegaly.
The carriers of the material basis of genes are chromosomes, which include DNA and proteins. But the genes of formation are not chemical, but functional. From a functional point of view, DNA consists of many blocks that store a certain amount of information - genes. The action of the gene is based on its ability to determine protein synthesis through RNA. The DNA molecule contains, as it were, information that determines the chemical structure of protein molecules. A gene is a section of a DNA molecule that contains information about the primary structure of any one protein (one gene - one protein). Because there are tens of thousands of proteins in organisms, there are tens of thousands of genes. The totality of all the genes of a cell makes up its genome. All cells of the body contain the same set of genes, but each of them implements a different part of the stored information. Therefore, for example, nerve cells differ from liver cells in both structural, functional and biological characteristics.
Now, it is even difficult to predict all the possibilities that will be realized in the next few decades.
2. History of genetic engineering
The history of high biomedical technologies, genetic research methods, as well as genetic engineering itself, is directly related to the eternal desire of man to improve the breeds of domestic animals and cultivated plants cultivated by people. By selecting certain individuals from groups of animals and plants and crossing them with each other, man, without having a correct idea of the inner essence of the processes occurring inside living beings, nevertheless, for many hundreds and thousands of years, created improved breeds of animals and varieties of plants that had certain useful and necessary properties for people.
In the 18th and 19th centuries, many attempts were made to find out how traits are passed on from generation to generation. One important discovery was made in 1760 by the botanist Koelreuther, who crossed two types of tobacco, transferring pollen from the stamens of one species to the pistils of another species. Plants obtained from hybrid seeds had characteristics intermediate between those of both parents. Koelreuter drew the logical conclusion from this that parental characteristics are transmitted both through pollen (seed cells) and through ovules (ovules). However, neither he nor his contemporaries, who were engaged in the hybridization of plants and animals, were able to reveal the nature of the mechanism of transmission of heredity. This is partly explained by the fact that at that time the cytological basis of this mechanism was not yet known, but mainly by the fact that scientists tried to study the inheritance of all plant characteristics simultaneously.
The scientific approach to studying the inheritance of certain traits and properties was developed by the Austrian Catholic monk Gregor Mendel, who in the summer of 1865 began his experiments in plant hybridization (crossing different varieties of peas) on the territory of his monastery. He was the first to discover the basic laws of genetics. Gregor Mendel achieved success because he studied the inheritance of individual, clearly distinct (contrasting) traits, counted the number of offspring of each type, and carefully kept detailed records of all his crossing experiments. Familiarity with the basics of mathematics allowed him to correctly interpret the data obtained and put forward the assumption that each trait is determined by two hereditary factors. A talented monk-researcher was later able to clearly show that hereditary properties are not mixed, but are transmitted to offspring in the form of certain units. This brilliant conclusion was subsequently fully confirmed when it was possible to see chromosomes and find out the characteristics of different types of cell division: mitosis (somatic cells - body cells), meiosis (sexual, reproductive, germinal) and fertilization.
Mendel reported the results of his work at a meeting of the Brunn Society of Naturalists and published them in the proceedings of this society. The significance of his results was not understood by his contemporaries, and these studies did not attract the attention of plant breeders and natural scientists for almost 35 years.
In 1900, after the details of cell division by type of mitosis, meiosis and fertilization itself became known, three researchers - de Vries in Holland, Correns in Germany and Chermak in Austria - conducted a series of experiments and, independently of each other, rediscovered the laws of heredity, previously described by Mendel. Later, having discovered an article by Mendel in which these laws were clearly formulated 35 years earlier, these scientists unanimously paid tribute to the monk scientist by naming the two basic laws of heredity after him.
In the first decade of the 20th century, experiments were carried out with a wide variety of plants and animals, and numerous observations were made regarding the inheritance of characters in humans, which clearly showed that in all these organisms heredity obeys the same basic laws. It was found that the factors described by Mendel that determine an individual trait are located in the chromosomes of the cell nucleus. Subsequently, in 1909, these units were called genes by the Danish botanist Johansen (from the Greek word “ge-nos” - genus, origin), and the American scientist William Sutton noticed a surprising similarity between the behavior of chromosomes during the formation of gametes (sex cells), their fertilization and transmission of Mendelian hereditary factors - genes. Based on these ingenious discoveries, the so-called chromosomal theory of heredity was created.
As a matter of fact, genetics itself, as the science of heredity and variability of living organisms and methods of controlling them, arose at the beginning of the 20th century. The American geneticist T. Morgan, together with his collaborators, conducted numerous experiments that made it possible to reveal the genetic basis of sex determination and explain a number of unusual forms of inheritance in which the transmission of a trait depends on the sex of the individual (the so-called sex-linked traits). The next major step forward was made in 1927, when G. Möller established that by irradiating the Drosophila fruit fly and other organisms with X-rays, it was possible to artificially induce gene changes in them, that is, mutations. This made it possible to obtain many new mutant genes - additional material for the study of heredity. Data on the nature of mutations served as one of the keys to understanding and the structure of the genes themselves.
In the 20s of our century, Soviet scientists of the school of A.S. Serebrovsky carried out the first experiments that showed how complex the gene is. These ideas were used by J. Watson and F. Crick, who managed in 1953 in England to create a DNA model and decipher the genetic code. The subsequent research work related to the targeted creation of new combinations of genetic material led to the emergence of genetic engineering itself.
At the same time, in the 40s, an experimental study of the relationships between genes and enzymes began. For this purpose, another object was widely used - the mold Neurospora, from which it was possible to artificially obtain and study a number of biochemical mutations associated with the loss of one or another special enzyme (protein). Over the past two decades, the most common targets of genetic research have been Escherichia coli and certain bacteriophages that infect this bacterium.
Since the very beginning of the 20th century, there has been continued interest in the study of the inheritance of certain (specific) traits in humans and in the hereditary transmission of desirable and undesirable traits in domestic animals and cultivated plants. Based on an ever-increasing knowledge of genetic patterns, geneticists and breeders have learned, almost to order, to breed livestock breeds that can survive in hot climates, cows that produce a lot of milk with a high fat content, chickens that lay large eggs with thin shells, and varieties of corn and wheat, which are highly resistant to certain diseases.
In 1972, the first hybrid (recombinant) DNA was obtained in the USA in the laboratory of P. Berg. Exciting ideas in the field of human genetics and genetic research methods have begun to be widely developed and applied in medicine itself. In the 70s, decoding of the human genome began. For more than decades, there has been a project called the Human Genome. Of the 3 billion nucleotide pairs arranged in continuous continuous passages, only about 10 million characters have been read so far. At the same time, new genetic techniques are being created that increase the speed of DNA reading. Director of the Medical Genetic Center of the Russian Academy of Medical Sciences V.I. Ivanov definitely believes that “the entire genome will be read around 2020.”
3. Genetic engineering as a science. Genetic engineering methods
Genetic engineering is the in vitro construction of functionally active genetic structures (recombinant DNA), or in other words, the creation of artificial genetic programs (Baev A.A.). According to E.S. Piruzyan genetic engineering is a system of experimental techniques that makes it possible to construct artificial genetic structures in the laboratory (in vitro) in the form of so-called recombinant or hybrid DNA molecules.
We are talking about the directed, according to a predetermined program, construction of molecular genetic systems outside the body with their subsequent introduction into a living organism. In this case, recombinant DNA becomes an integral part of the genetic apparatus of the recipient organism and imparts to it new unique genetic, biochemical, and then physiological properties.
The goal of applied genetic engineering is to design such recombinant DNA molecules that, when introduced into the genetic apparatus, would give the body properties useful to humans.
Recombinant DNA technology uses the following methods:
Specific cleavage of DNA by restriction nucleases, accelerating the isolation and manipulation of individual genes;
Rapid sequencing of all nucleotides in a purified DNA fragment, which makes it possible to determine the boundaries of the gene and the amino acid sequence encoded by it;
Construction of recombinant DNA;
Nucleic acid hybridization, which allows the detection of specific RNA or DNA sequences with greater accuracy and sensitivity, based on their ability to bind complementary nucleic acid sequences;
DNA cloning: in vitro amplification using a polymerase chain reaction or introduction of a DNA fragment into a bacterial cell, which, after such transformation, reproduces this fragment in millions of copies;
Introduction of recombinant DNA into cells or organisms.
4. Areas of application of genetic engineering
The current scientific discoveries in the field of human genetics are actually of revolutionary significance, since we are talking about the possibility of creating a “map of the human genome,” or “pathological anatomy of the human genome.” This genetic map will make it possible to determine the location of genes on the long DNA helix that are responsible for certain hereditary diseases. According to genetic scientists, these unlimited possibilities formed the basis for the idea of using so-called gene therapy in clinical practice, which is a direction of treating patients that involves replacing affected genes using high biomedical technologies and genetic engineering. Invasion into the composition of human gene systems and ensuring their vital activity is possible both at the level of somatic (all bodily cells with certain structural and functional differences) cells of the body, and at the level of reproductive, reproductive (germinal) and germinal (embryonic) cells.
Genetic engineering as a type of therapy - the treatment of a specific genetically determined disease - is associated with the supply of a corresponding non-defective DNA molecule for the purpose of replacing it with the help of a gene - a section of a chromosome that contains a defect, or for integration into human genetic material by merging with so-called somatic cells of the human body that have a genetic defect. The task of genetic engineering in relation to a person is to provide an appropriate targeted effect on a specific gene to correct it towards proper functioning and provide a person suffering from a hereditary disease with a normal, unaltered version of the gene. In contrast to drug therapy, this therapy, called genetic engineering, will apparently be able to provide the patient with long-term, prolonged, highly effective treatment that brings great relief and benefit.
However, all modern methods of introducing DNA into living organisms are not able to direct and deliver it to a specific population of cells containing an altered and therefore malfunctioning gene. In other words, the so-called directed transfer, the transport of genes in the body (in the “in vivo” model) is currently impossible.
Another methodological approach, based on extracting from the patient’s body a certain population of cells containing the affected gene, and manipulating the genetic material by replacing defective genes in the cells using genetic engineering (in the “in vitro” model) and returning them to that place in the body, where they were taken from the patient is currently possible in medical genetic centers. This method of gene therapy through genetic engineering has already been used in an experimental attempt to cure two patients suffering from a rare genetic disease called beta thalassemia, which, like sickle cell anemia, is also caused by the presence of a malformed and therefore incorrectly functioning protein in red blood cells. The essence of the manipulation was that so-called stem cells were isolated from the bone marrow of these patients, into the chromosomes of which the DNA section responsible for the production of the normal hemoglobulin protein was introduced - the gene. After the malfunctioning stem cells remaining in the patients' bone marrow were almost completely destroyed, the patients were injected with genetically engineered stem cells. Unfortunately, these two attempts were clinically unsuccessful, as the patients died. This first case of genetic engineering in a hospital setting was not authorized or approved by the relevant review committees, and its participants were strongly condemned for gross violation of the rules of research in the field of human genetics.
Genetic engineering of reproductive (reproductive) cells can lead to completely different consequences, since the introduction of DNA into these cells differs from correcting a genetic defect in somatic (bodily, non-reproductive) cells. It is known that the introduction of other genes into the chromosomes of germ cells leads to their transmission to subsequent generations. In principle, one can imagine adding certain sections of DNA to replace defective sections to the genetic material of each reproducing cell of a certain person who is affected by one or another genetically determined disease.
Indeed, this has been achieved in mice. Thus, an egg was obtained from the ovary of a female, which was subsequently fertilized in vitro (in vitro), and then a foreign DNA section was introduced into the chromosome of the fertilized egg. The fertilized egg itself with an altered genome was implanted (introduced) into the maternal uterus of a female mouse. The source of foreign DNA in one experiment was rabbit genetic material, and in the other, human genetic material.
In order to detect during the period of fetal development the likelihood of having a child with certain genetic abnormalities, such as Down syndrome or Tay-Sachs disease, a research technique called amniocentesis is used - a prenatal analysis, during which a sample of biological fluid containing germ cells, taken from the amniotic sac early in the second trimester of pregnancy. In addition, the technique of extracting various fetal cells from a sample of the mother’s placental blood was further developed. The uterine cells obtained in this way can currently only be used to identify a limited number of genetically determined diseases in which there are pronounced, gross disturbances in the DNA structure and changes determined using biochemical tests. Genetic engineering using recombinant DNA during prenatal research opens up the possibility of correctly diagnosing various and numerous hereditary diseases.
In this case, techniques are being developed to create so-called gene “probes”, using which it is possible to determine whether a chromosome contains a normal, unchanged gene or an abnormal, defective gene. In addition, genetic engineering associated with the use of recombinant DNA, which is at one of the stages of its formation, will in the future make it possible to carry out the so-called “planning” of human genes, so that a certain gene that carries distorted, pathological information and is therefore of interest for geneticists, could be identified on time and quickly enough by analogy with the method of using another “tagged” gene. This complex medical and biological technique should help in determining the location of any gene in uterine cells, and not just in those in which various disorders are likely to be detected using the amniocentesis technique.
In this regard, in recent years, new sections of biomedical sciences have emerged, such as, for example, high DNA technologies, embryo therapy and cell therapy (cytotherapy), that is, intrauterine diagnosis and treatment of a genetically determined disease both at the educational stage and the development of the embryo (embryo), and at the stage of fetal maturation. Invasion and manipulation of embryonic material has a direct impact on the inheritance of genetic changes, since they have the ability to be transmitted from generation to generation. Moreover, genetic diagnosis itself begins to develop into genetic forecasting, that is, into determining the future fate of a person, consolidating the main revolutionary changes in medicine itself, which, as a result of complex medical-genetic experiments and techniques, gained the opportunity long before the appearance of the “clinical picture of the disease” , sometimes even before a person’s birth, to determine what hereditary ailments threaten him. Thus, thanks to the efforts of geneticists and specialists in the field of genetic engineering, the so-called “predictive medicine” was born in the depths of the biomedical sciences, that is, medicine that “makes predictions for the future.”
At the same time, various technologies and methods of genetic engineering make it possible to predict in the prenatal period of a child’s development, before his birth, not only the presence of a certain hereditary disease, but also to describe in detail the medical and genetic properties of the growing embryo and fetus.
With the accumulation of new data on the genetic mapping of the human genome and the description (sequencing) of its DNA, and also because the modern methods of studying DNA polymorphisms being developed make it possible to make available genetic information about certain structural and functional (including pathological) features of the human body, which, apparently, will appear in the future, but are not yet noticeable now, it becomes possible to obtain, with the help of medical genetic diagnostics, all genetic information about the child not only preclinically, that is, before the manifestation of a certain hereditary disease, and prenatally, that is, before his birth , but also preceptively, that is, even before its conception.
In the foreseeable future, thanks to the success and progress in the field of medical genetic diagnostics, it will be possible, based on DNA diagnostic data, to fairly confidently judge, for example, what a person’s height, mental abilities, predisposition to certain diseases (in particular, cancer) will be. or mental), doomed to the manifestation and development of any hereditary diseases.
Modern medical and biological technologies make it possible to detect various disorders in genes that can manifest themselves and cause certain ailments, not only at the stage of a clinically pronounced disease, but also when there are no signs of pathology yet and the disease itself will not manifest itself so soon. Examples of this include Alzheimer's disease and Huntington's chorea, which affect a person over 40 years of age, or even 70 years of age. However, even in these cases, it is possible to detect genes that can cause similar diseases in humans, even before the patient’s conception. It is also known that diabetes mellitus can be classified as one of these diseases. Predisposition to this disease and the genetically determined pathology itself are inherited and can manifest themselves in case of non-compliance with a certain lifestyle in adulthood or old age. We can say with reasonable certainty that if both parents or one of them suffer from diabetes, then the likelihood of inheriting the diabetes gene or a combination of such genes is passed on to children.
In this case, it turns out to be possible to conduct appropriate medical and biological studies and make a correct diagnosis in the presence of microscopically small quantities of biological material. Sometimes a few individual cells are enough for this, which will be multiplied in culture in vitro, and from them a “genetic portrait” of the tested person will be obtained, of course, not for all the genes of his genome (there are tens of thousands of them!), but for those of them , in respect of which there are reasonable grounds to suspect the presence of certain defects. The simultaneous development of methods of cellular and genetic engineering will make it possible, at subsequent stages of knowledge of the genome, to open up the practical possibility of arbitrarily, and, above all, for therapeutic purposes, changing the sequence and order of genes, their composition and structure.
Medicine is not the only area of application of genetic engineering. There are genetic engineering of plants and genetic engineering of bacteriological cells.
Recently, new opportunities have emerged in obtaining “edible” vaccines based on transgenic plants.
Great progress has been made in transgenic plants in the world. They are largely due to the fact that the problem of obtaining an organism from a cell, a group of cells or an immature embryo in plants is now not very difficult. Cell technologies, tissue culture and the creation of regenerants are widely used in modern science.
Let's consider the achievements in the field of plant growing that were obtained at the Siberian Institute of Plant Physiology and Biochemistry of the Siberian Branch of the Russian Academy of Sciences.
Thus, in recent years, a number of transgenic plants have been obtained by transferring into their genome the genes ugt, acp, acb, accc and others isolated from various plant objects.
As a result of the introduction of these genes, transgenic plants of wheat, potatoes, tomatoes, cucumbers, soybeans, peas, rapeseed, strawberries, aspen and some others appeared.
The introduction of genes was carried out either by “targeting” tissues from a “gene gun” (the design of which was developed at our institute), or by a genetic vector based on an agrobacterial plasmid with built-in target genes and corresponding promoters.
As a result, a number of new transgenic forms were formed. Here are some of them.
Transgenic wheat (2 varieties), which has significantly more intensive growth and tillering, is presumably more resistant to drought and other unfavorable environmental factors. Its productivity and inheritance of acquired properties are being studied.
Transgenic potatoes, which have been monitored for three years. It consistently produces a yield that is 50-90 percent higher than the control, has acquired almost complete resistance to auxin herbicides and, in addition, its tubers “blacken” significantly less on cuts due to a decrease in polyphenol oxidase activity.
Transgenic tomato (several varieties), characterized by greater bushiness and yield. In a greenhouse, its yield is up to 46 kg per square meter (more than two times higher than the control).
Transgenic cucumber (several varieties) produces a larger number of fertile flowers and, consequently, fruits with a yield of up to 21 kg per square meter versus 13.7 in the control.
There are transgenic forms of other plants, many of which also have a number of useful economic traits.
Genetic engineering is the science of today and tomorrow. Already, tens of millions of hectares are being sown with transgenic plants around the world, new medicines and new producers of useful substances are being created. Over time, genetic engineering will become an increasingly powerful tool for new advances in medicine, veterinary medicine, pharmacology, the food industry and agriculture.
5. Scientific facts about the dangers of genetic engineering
It should be noted that along with the progress that the development of genetic engineering brings, there are also some facts about the dangers of genetic engineering, the main ones of which are presented below.
1. Genetic engineering is fundamentally different from the development of new varieties and breeds. The artificial addition of foreign genes greatly disrupts the finely regulated genetic control of a normal cell. Gene manipulation is fundamentally different from the combination of maternal and paternal chromosomes that occurs in natural crossings.
2. Currently, genetic engineering is technically imperfect, since it is not able to control the process of inserting a new gene. Therefore, it is impossible to predict the insertion site and the effects of the added gene. Even if the location of a gene can be determined once it has been inserted into the genome, the available DNA information is very incomplete to predict the results.
3. As a result of the artificial addition of a foreign gene, hazardous substances may unexpectedly be formed. In the worst case, these may be toxic substances, allergens or other substances harmful to health. Information about these types of possibilities is still very incomplete.
4. There are no completely reliable methods for testing for harmlessness. More than 10% of serious side effects of new drugs cannot be detected, despite carefully conducted safety studies. The risk that the dangerous properties of new genetically modified foods will go undetected is likely to be much greater than in the case of drugs.
5. The current requirements for testing for harmlessness are extremely insufficient. They are clearly designed to simplify the approval process. They allow the use of extremely insensitive harmlessness testing methods. There is therefore a significant risk that hazardous food products will be able to pass inspection undetected.
6. Food products created to date using genetic engineering do not have any significant value for humanity. These products satisfy mainly commercial interests only.
7. Knowledge about the effects of genetically modified organisms introduced into the environment is completely insufficient. It has not yet been proven that organisms modified by genetic engineering will not have harmful effects on the environment. Environmentalists have suggested various potential environmental complications. For example, there are many opportunities for the uncontrolled spread of potentially harmful genes used by genetic engineering, including gene transfer by bacteria and viruses. Complications caused by the environment are likely to be impossible to correct because the released genes cannot be taken back.
8. New and dangerous viruses may emerge. It has been experimentally shown that viral genes embedded in the genome can combine with the genes of infectious viruses (so-called recombination). These new viruses may be more aggressive than the original ones. Viruses may also become less species specific. For example, plant viruses can become harmful to beneficial insects, animals, and also humans.
9. Knowledge of the hereditary substance, DNA, is very incomplete. The function of only three percent of DNA is known. It is risky to manipulate complex systems about which knowledge is incomplete. Extensive experience in the fields of biology, ecology and medicine shows that this can cause serious unpredictable problems and disorders.
10. Genetic engineering will not help solve the problem of world hunger. The claim that genetic engineering can make a significant contribution to solving the problem of world hunger is a scientifically unfounded myth.
Conclusion
Genetic engineering is a method of biotechnology that deals with research into the restructuring of genotypes. The genotype is not just a mechanical sum of genes, but a complex system that has developed during the evolution of organisms. Genetic engineering makes it possible to transfer genetic information from one organism to another through in vitro operations. Gene transfer makes it possible to overcome interspecies barriers and transfer individual hereditary characteristics of one organism to another.
Rearrangement of genotypes, when performing genetic engineering tasks, represents qualitative changes in genes not associated with changes in the structure of chromosomes visible in a microscope. Gene changes are primarily associated with the transformation of the chemical structure of DNA. Information about the structure of a protein, written as a sequence of nucleotides, is implemented as a sequence of amino acids in the synthesized protein molecule. A change in the sequence of nucleotides in chromosomal DNA, the loss of some and the inclusion of other nucleotides, changes the composition of the RNA molecules formed on DNA, and this, in turn, determines a new sequence of amino acids during synthesis. As a result, a new protein begins to be synthesized in the cell, which leads to the appearance of new properties in the body. The essence of genetic engineering methods is that individual genes or groups of genes are inserted into or excluded from the genotype of an organism. As a result of inserting a previously absent gene into the genotype, the cell can be forced to synthesize proteins that it had not previously synthesized.
Bibliography
2. Lee A., Tinland B. Integration of t-DNA into the plant genome: prototype and reality // Plant Physiology. 2000. - Volume 47. - No. 3.
3. Lutova L. A., Provorov N. A., Tikhodeev O. N. et al. Genetics of plant development. - St. Petersburg: Nauka, 2000. - 539 p.
4. Lyadskaya M. Genetic engineering can do anything - even grow a vaccine in the garden // Pharmaceutical Bulletin. - 2000. - No. 7.
5. Romanov G. A. Genetic engineering of plants and ways to solve the problem of biosafety // Plant Physiology, 2000. - Volume 47. - No. 3.
6. Salyaev R. Myths and realities of genetic engineering // Science in Siberia. - 2002. - No. 7.
7. Favorova O. O. Treatment with genes - fiction or reality? // Pharmaceutical Bulletin. - 2002. - No. 5.
Kuzmina N.A. Fundamentals of biotechnology: textbook. - Omsk: OGPU, 2001. - 256 p.
Lutova L. A., Provorov N. A., Tikhodeev O. N. et al. Genetics of plant development. - St. Petersburg: Nauka, 2000. - 539 p.
Lyadskaya M. Genetic engineering can do anything - even grow a vaccine in the garden // Pharmaceutical Bulletin. - 2000. - No. 7.
Kuzmina N.A. Fundamentals of biotechnology: textbook. - Omsk: OGPU, 2001. - 256 p.
Favorova O. O. Treatment with genes - fiction or reality? // Pharmaceutical Bulletin. - 2002. - No. 5.
Salyaev R. Myths and realities of genetic engineering // Science in Siberia. - 2002. - No. 7.
Kuzmina N.A. Fundamentals of biotechnology: textbook. - Omsk: OGPU, 2001. - 256 p.