Актиномицеты (Actinomycetales, от греч. aktis – луч, mykes – гриб) – это ветвящиеся бактерии, принадлежащие типу Актинобактерии (Actinobacteria). Они являются частью нормальной микрофлоры пищеварительной системы наземных позвоночных и беспозвоночных животных, а также в обилии присутствуют в грунте и играют важнейшую роль в экологии и круговороте веществ в почве.
Эти микроорганизмы являются возбудителями многих оппортунистических патологий – таких, которые возникают в результате снижения функции иммунной системы организма. Актиномицеты широко используются в биотехнологии, так как являются источником целого ряда антибактериальных и противоопухолевых веществ.
Рис. 1. Стрептомицеты синтезируют огромное количество антибактериальных и противоопухолевых препаратов.
Строение актиномицетов: почему все же бактерии, а не грибы?
1. Организация генетического материала
Наследственный материал Актиномицетов заключен в одной молекуле дезоксирибонуклеиновой кислоты, имеющей кольцевую форму и свободно располагающейся в цитоплазме – такая же форма организации генетического материала, называемая нуклеоидом, характерна и для других бактерий. У грибов же генетический материал организован и входит в состав клеточного ядра.
ДНК актиномицетов содержит большое количество ГЦ-пар (65-75% от общего количества нуклеотидов). Этот признак постоянен, не зависит от мутаций и потому используется в систематике микроорганизмов. Такое содержание ГЦ-пар делает ДНК актиномицетов весьма тугоплавкой, потому на анализ ДНК актиномицетов уходит больше времени по сравнению с другими бактериями.

Рис. 2. Схематическое строение клеточной стенки Гр+ бактерий.

Рис. 3. Актиномицеты, окрашенные по Граму.
Актиномицеты имеют плотную бактериальную клеточную стенку, которая расположена снаружи от цитоплазматической мембраны и обуславливает их положительное окрашивание по Грамму. Как и у других Грамм-положительных бактерий, она состоит из нескольких десятков слоев полимера муреина (пептидогликана), который пронизан тейхоевыми и липотейхоевыми кислотами. Липотейхоевые кислоты заякорены в цитоплазматической мембране бактерии и соединяют её с клеточной стенкой. Тейхоевые кислоты придают клеточной стенке отрицательный заряд. Клеточная стенка грибов же состоит из других полимеров – хитина и глюкана.
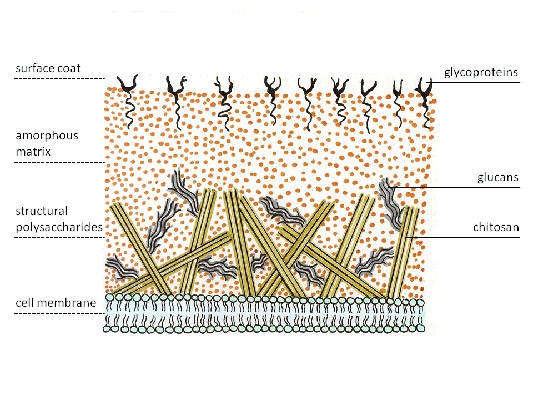
Рис. 4. Клеточная стенка грибов.
3. Клеточные органеллы
Актиномицеты, подобно другим бактериям, не имеют мембранных органелл. У актиномицетов 70S рибосомы, тогда как грибы имеют 80S рибосомы, равно как и остальные эукариотические организмы.
4. Рост колоний
Образование мицелия при росте – это то, что более всего роднит актиномицеты с грибами. Мицелий в случае актиномицетов представляет собой ветвящуюся совокупность гиф. Гифы разделены перегородками на длинные бактериальные клетки, содержащие несколько нуклеоидов. Перегородки у ряда видов могут проходить во взаимно перпендикулярном направлении. Ветвятся гифы путем почкования.
Мицелий, врастающий в субстрат (землю, ил или питательную среду), называется субстратным. Он обеспечивает колонию питательными веществами. Над субстратом возвышается воздушный мицелий, придающий колонии «пушистость» — он образует споры, а также так называемые «вторичные метаболиты» (в отличие от «первичных метаболитов» субстратного мицелия), среди которых много антибактериальных веществ.
Жизненный цикл и физиология актиномицетов
В процессе жизненного цикла большая часть актиномицетов образует споры. Некоторые актиномицеты размножаются путем фрагментации мицелия.
1. Спорообразование
Споры актиномицетов происходят из воздушного мицелия. Это экзоспоры – они развиваются снаружи материнской клетки. Гифы воздушного мицелия, из которых развиваются споры, называются спороносцами. Споры могут содержаться в утолщении на конце спороносца – спорангии (например, у стрептомицетов, актинопланов и плимелий), а могут располагаться цепочкой вдоль спорангиеносца (например, у нокардий и актиномадур).
По числу образующихся спор актиномицеты делятся на:
- Моноспоровые (к примеру, Saccaromonospora, Micromonospora, Thermomonospora) – образуют одиночные споры, чаще путем отпочковывания и последующего отделения перегородкой от материнского гифа;
- Олигоспоровые (например, Actinomadura) – образуют короткие цепочки спор вдоль спороносца;
- Полиспоровые (большинство других актиномицетов, к примеру, Streptomyces, Frankia, Geodermatophilus) – образуют множество спор, заключенных в спорангии.

Рис. 5. Спорангий актиномицетов рода Frankia.
Споры актиномицетов могут быть подвижными – в таком случае, спора имеет жгутик и может передвигаться (споры актинопланов, геодерматофилов и дерматофилов). В большинстве случаев споры неподвижны и распространяются ветром, водой или животными.

Рис. 6. Дерматофилы, световая микроскопия.
Споруляция у актиномицетов особенно активно протекает в неблагоприятных условиях. Устойчивость спор к нагреванию невелика по сравнению со спорами других бактерий, однако высушивание они выдерживают не хуже других, а потому имеют колоссальное приспособительное значение. Актиномицеты доминируют над другими микроорганизмами в сухих пустынных почвах.
Прорастание своры требует определенной влажности внешней среды. В присутствии воды спора набухает, в ней активируются ферменты и запускаются метаболические процессы, сопровождающиеся выходом ростовых трубок (будущих бактериальных тел) и синтезом нуклеиновых кислот.
2. Тип дыхания
Большинство актиномицетов — аэробы (нуждаются в кислороде для поддержания жизнедеятельности). Факультативные анаэробы (бактерии, способные жить как при наличии, так и в отсутствие кислорода) встречаются среди видов с непродолжительной мицелиальной стадией, размножающиеся фрагментацией мицелия.
3.Кислотоустойчивость
Актиномицеты обладают ацидотолерантностью – устойчивостью в кислой среде, которая позволяет им обитать в насыщенных кислотами лесных почвах. Кислотоустойчивость в лаборатории может быть определена окраской препарата, содержащего актиномицеты, по Цилю-Нильсену (фуксином с последующей протравкой серной кислотой и окраской метиленовой синью). Большинство актиномицетов при такой окраске не обесцвечиваются после протравки кислотой и сохраняют красный фуксиновый цвет. Щелочная среда неблагоприятна для этих бактерий: при повышенном рН они склонны к спорообразованию.
4. Особенности метаболизма

Рис. 7. Аэробные актиномицеты образуют пигмент на скошенном агаре. Слева направо: Actinomadura madurae, Nocardia asteroides, Micromonospora.
Выше упоминалось образование воздушным мицелием «вторичных метаболитов». Среди них:
- пигменты, обуславливающие различную расцветку воздушного мицелия при росте на средах;
- летучие пахучие вещества, придающие характерный запах почве после дождя, застоявшейся воде, кожным покровам некоторых животных;
- антибиотики:
a. противогрибковые – полиены;
b. противобактериальные – например, стрептомицин, эритромицин, тетрациклин, ванкомицин;
c. противоопухолевые – антрациклины, блеомицин.
Где обитают актиномицеты?
Актиномицеты в наибольшем количестве обнаруживаются в почвах, притом мицелиальных форм значительно меньше, чем спор. Они играют значительную роль в образовании гумуса, разлагая органические вещества, труднодоступные для утилизации другими бактериями. Актиномицеты в связи с этим используют в качестве санитарно-показательных микроорганизмов в санитарно-эпидемиологическом деле: обнаружение их в большим количестве в почве или воде указывает на наличие компоста в соответствующем субстрате.

Рис. 8. Актиномицеты в компосте.
Актиномицеты являются симбионтами многих растений, помогая им фиксировать азот. В то же время, многие микроорганизмы этого класса являются возбудителями заболеваний растений.

Рис. 9. Стрептомикоз картофеля.
Они также обнаруживаются в составе нормальной микрофлоры пищеварительной системы целого ряда животных, начиная от почвенных кольчатых червей (например, дождевых) и заканчивая крупным домашним скотом.
Эти микроорганизмы помогают расщеплять целлюлозу, в обилии присутствующую в растительной пище. У человека актиномицеты обнаруживаются в полости рта (десна и зубной налет), кишечнике (дистальные отделы толстого кишечника), на коже (лицо, крылья носа, за ушами, между пальцами) и в органах дыхательной системы (преимущественно в верхних дыхательных путях).
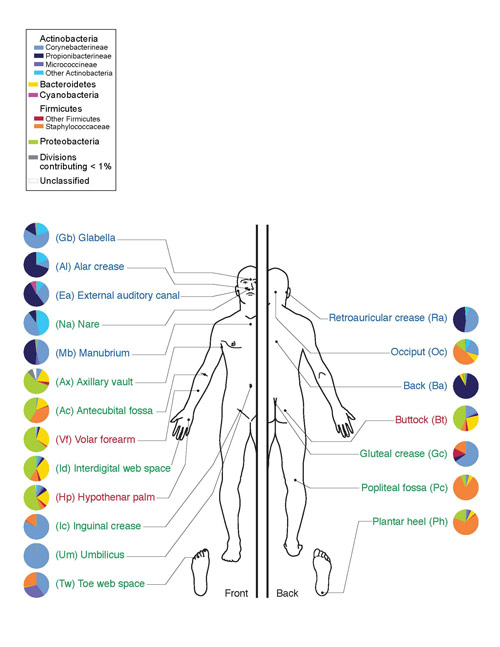
Рис. 10. Микрофлора кожи человека. Тип Актинобактерии обозначены оттенками синего, класс Актиномицеты – ярко-голубым.
Актиномицеты при условии снижения иммунной реактивности организма могут стать причиной актиномикозов – оппортунистических заболеваний, заключающихся в формировании актиномикозных гранулем – скоплений бактериальных тел, напоминающих зерна желтой серы («друз»), окруженных иммунокомпетентными клетками. Воспалительная реакция ведет к расплавлению гранулем, образованию свищей, ведущему к перфорациям органов и разносу бактерий кровью.

Рис. 11. Актиномикозная друза, окраска по Граму.

Рис. 12. Актиномикоз верхней челюсти у коровы.

Рис. 13. Максиллярный актиномикоз человека.
Актиномицеты – удивительные организмы, до сих пор вводящие в заблуждение множество ученых своей схожестью с грибами. Наряду с потенциальной опасностью в виде оппортунистических актиномикозов, эти организмы дарят человеку плодоносную почву и оружие для борьбы с инфекционными и онкологическими заболеваниями – антибиотики и цитостатики.
Химический состав и структура клеточной стенки грибов
Клеточная стенка грибов многослойная, при этом разные слои образованы различающимися по химическому составу структурными углеводами, которые по химическому составу можно разделить на 3 группы:
полимеры глюкозы (глюкан, хитин, целлюлоза). Глюканы составляют наружный слой клеточной стенки большинства грибов. Внутренний слой грибной клеточной стенки образован цепочками хитина, придавая ей жесткость. Хитин замещает целлюлозу, которая у большинства грибов отсутствует, но входит в состав клеточной стенки оомицетов, которые в настоящее время к типичным грибам не относятся. Деацетилированный хитин получил название хитозан, который в комплексе с хитином образует клеточную стенку зигомицетов.
полимеры других моносахаридов (маннозы, галактозы и др.) в отличие от высших растений, где они составляют основу матрикса под общим названием гемицеллюлоза, менее характерны для грибов. Исключение составляют дрожжи, в клеточных стенках которых особенно много полимеров маннозы, называемых маннаны. Предполагают, что такой состав стенки лучше обеспечивает почкование.
полимеры углеводов, ковалентно связанные с пептидами (гликопротеины) формируют срединный слой многослойной клеточной стенки и играют важную роль, как в поддержании структурной целостности клеток, так и в ее обменных процессах с окружающей средой.
К другим специфическим особенностям грибной клетки относятся: отсутствие пластид , что сближает ее с клеткой животных;
Отсутствие крахмала , который у эумицетов замещается полисахаридом, близким к животному крахмалу гликогену, у оомицетов – полисахаридом, близкому к ламинарину бурых водорослей. Вырабатывается и целый ряд специфических для грибов запасных углеводов.
Выработка специфических вторичных метаболитов , из которых большую роль играют антибиотики, фито– и микотоксины, фитогормоны.
К специфическим особенностям грибов относятся также гетерокариоз и парасексуальный процесс.
У грибов очень широко распространено явление гетерокариоза или разноядерности, при котором в одной клетке длительное время сохраняются ядра, гетероаллельные по некоторым генам. Гетерокариоз заменяет гаплоидным грибам гетерозиготность и способствует быстрой адаптации грибов к меняющимся условиям. Наличие разноядерности обусловлено рядом специфических особенностей грибов:
1. наличие более одного ядра в клетке
2. специфическое строение клеточной перегородки, в которой имеется одно или несколько сквозных отверстий, называемых порами, через которые ядра могут мигрировать из одной клетки в другую
3. гифы внутри одной колонии и даже разных близко расположенных колоний, выросших из разных спор одного вида гриба часто срастаются, в результате чего возможен обмен ядрами разных штаммов.
Парасексуальный (псевдополовой) процесс. Если в гетерокариотичных клетках сливаются ядра, гетероаллельные по какому-либо локусу, возникает гетерозиготное диплоидное ядро. Оно может попасть в спору и дать начало диплоидному гетерозиготному клону. В процессе митозов диплоидные ядра могут возвращаться в гаплоидное состояние вследствие потери одного набора хромосом или в них может происходить обмен участками хромосом (митотический кроссинговер). Оба процесса сопровождаются рекомбинацией родительских генов и, следовательно, фенотипов. Парасексуальная (бесполовая) рекомбинация очень редкое явление, не превышающее одного ядра на миллион, но вследствие огромного числа ядер в мицелии постоянно наблюдается в популяциях грибов.
Размножение – вегетативное, бесполое, половое.
Вегетативное – фрагментация таллома, образование хламидоспор, которые после периода покоя прорастают в мицелий, почкование у дрожжей.
Бесполое размножение у разных грибов может осуществляться подвижными и неподвижными спорами. Зооспоры образуют относительно небольшая группа грибов и грибоподобных организмов – водные и некоторые наземные, у которых отчетливо прослеживаются генетические связи с водными грибами и водорослями. Структура жгутиков является важным диагностическим признаком при отнесении к определенному царству. У подавляющего большинства грибов при бесполом размножении образуются неподвижные споры, что указывает на их очень давний выход на сушу. По месту формирования и локализации различают эндогенные, образующиеся в спорангиях спорангиоспоры и экзогенные (конидии), развивающиеся на специальных гифах – конидиеносцах. Конидии образуются у большинства грибов (аскомицеты, базидиомицеты, некоторые зигомицеты) составляя конидиальное спороношение, которое отличается огромным многообразием и широко используется для диагностики грибов.
Половое размножение грибов имеет свою специфику как в морфологии полового процесс, так и в механизмах генетической и физиологической регуляции пола и передачи наследственной информации.
Соматогамия – самый распространенный и наиболее простой тип полового процесса, заключается в слиянии двух не дифференцированных на гаметы соматических клеток. Иногда он протекает даже без слияния клеток – сливаются ядра внутри клетки. Встречается у большинства базидиомицетов, сумчатых дрожжей и некоторых др. таксонов.
Гаметангиогамия – на гаплоидном мицелии обособляются гаметангии, при половом процессе сливается их содержимое. Такой половой процесс характерен для большинства сумчатых грибов. Вариантом гаметангиогамии является зигогамия у зигомицетов.
Гаметогамия в виде изо- гетеро и оогамии у грибов встречается значительно реже, чем у других эукариот. Изо- и гетерогамия встречается лишь у хитридиомицетов. Классическая оогамия с образованием сперматозоидов и яйцеклеток у грибов не выражена, а встречаются сильно измененные варианты.
По особенностям регуляции пола у грибов выделяют несколько типов полового процесса
Гинандромиксис можно рассмотреть на примере двудомных оомицетов, у которых оогонии и антеридии развиваются на разных талломах, например фитофтора или картофельный гриб. Если генетически однородный штамм выращивать в монокультуре, то он размножается только бесполым способом. Если рядом оказываются мицелии двух штаммов, то у них прослеживаются морфогенетические изменения под действием химических выделений стероидной природы – половых феромонов. Антеридиол индуцирует образование антеридиев у партнера, а оогониол –оогониев. При этом регуляция пола имеет относительный характер: будет ли штамм образовывать антеридии или же оогонии, зависит от количественного соотношения соответствующих феромонов у него и его партнера. Отсюда название полового процесса – гинандромиксис.
Димиксис или гетероталлизм. Уже давно было подмечено, что грибы могут быть гомо- или гетероталличными. У гомоталличных видов при половом процессе сливаются генетически идентичные ядра внутри мицелия. У гетероталличных видов для прохождения полового цикла необходимо на каком-то этапе (различном у разных видов грибов) слияние потомков спор (точнее, их ядер). Для половой совместимости двух штаммов необходимо генетическое различие (гетероаллельность) в определенных локусах, называемых локусами спаривания. У большинства грибов (зигомицеты, аскомицеты, часть базидиомицетов) имеется один локус спаривания с двумя аллелями. Локус спаривания состоит из нескольких генов, котрые управляют синтезом половых феромонов. Такой гетероталлизм называется однофакторным или биполярным. Потомство таких грибов после мейоза разделяется на две самонесовместимые, но взаимно совместимые группы в соотношении 1:1, т.е. вероятность родственного(инбридинга) и неродственного(аутбридинга) скрещивания составляет как и у двуполых высших эукариот 50%.
В геноме высших базидиальных грибов имеется два локуса спаривания – A и B, причем совместимы друг с другом только штаммы гетероаллельные по обоим локусам (Ax Bx совместим с Ay By, но не с Ax By и Ay Bx). Такой гетероталлизм называют двухфакторным или тетраполярным. Он снижает вероятность инбридинга до 25 %.
Диафоромиксис – у высших базидиомицетов имеется не два, а много аллелей локуса спаривания, случайно встречающихся у разных штаммов, слагающих популяцию. Такой контроль спаривания обеспечивает 100% вероятность аутбридинга, так как штаммы, имеющие разные аллели, взаимносовместимы, а аллелей очень много. В результате образуются панмиксные гибридные популляции.
Жизненные циклы грибов также разнообразны как и сами грибы. Основные циклы, их принадлежность к отделам грибов
1.Бесполый цикл характерен для огромной группы несовершенных или митогрибов, утративших половое размножение. Деление их ядер исключительно митотические. Подавляющее большинство митогрибов относится к сумчатым грибам, но в связи с утратой полового процесса, составляют формальную группу несовершенных грибов или дейтеромицетов.
2. Гаплоидный цикл. Вегетативный таллом несет гаплоидные ядра. После полового процесса (сингамии) диплоидная зигота (обычно после периода покоя) делится мейотически – зиготический мейоз. Характерен для зигомицетов и многих хитридиомицетов.
3. Гаплоидно-дикариотический цикл характеризуется тем, что после слияния содержимого гаметангиев (гаметангиогамия) или соматических клеток гаплоидного мицелия (соматогамия) ядра образуют дикарионы (пары генетически разных ядер). Они синхронно делятся, образуя дикариотический мицелий. Половой процесс завершается слиянием ядер дикариона, образующаяся зигота делится мейозом без периода покоя. Мейоспоры составляют половое спороношение сумчатых и базидиальных грибов в виде аскоспор и базидиоспор. При их прорастании образуется гаплоидный мицелий. У подавляющего большинства сумчатых грибов (кроме дрожжей и тафриновых грибов) в жизненном цикле преобладает гаплоидная фаза в виде вегетативного мицелия (анаморфа), дикариотическая фаза кратковременна и представлена аскогенными гифами, на которых образуются сумки (телеоморфа). У базидиальных грибов в жизненном цикле преобладает дикариотическая фаза, гаплоидная фаза кратковременна.
4. Гаплоидно-диплоидный цикл в виде изоморфной смены поколений у грибов встречается редко (некоторые дрожжи и водные хитридиомицеты).
5. Диплоидный цикл характерен для оомицетов и некоторых сумчатых дрожжей. Вегетативный таллом диплоидный, мейоз гаметический, наблюдается при формировании гаметангиев или гамет.
3. Экологические группы грибов.
Грибы и грибоподобные организмы входят во все наземные и водные экосистемы, как важнейшая часть гетеротрофного блока, вместе с бактериями занимая трофический уровень редуцентов. Широкое распространение грибов в биосфере определяется рядом важнейших особенностей:
1. Наличие у большинства мицелиальной структуры таллома. (позволяет быстро осваивать субстрат, иметь большую поверхность контакта со средой).
2. Большая скорость роста и размножения, позволяющие в короткие сроки заселять большие массы субстрата, образуя огромное число спор и распространять их на большие расстояния.
3. Высокая метаболическая активность, проявляющаяся в широком диапазоне значений экологических факторов.
4. Высокая скорость генетической рекомбинации, значительная биохимическая изменчивость, экологическая пластичность.
5. Способность быстро переходить к состоянию анабиоза, переживать неблагоприятные условия в течение длительного времени.
Главным экологическим фактором для грибов является пищевой субстрат. По отношению к этому фактору выделяют основные группы грибов, которые называются трофическими группами.
4. Краткая характеристика отделов грибов.
Прокариотическая клетка
Прокариоты - организмы, не обладающие, в отличие от эукариот, оформленным клеточным ядром и другими внутренними мембранными органоидами (за исключением плоских цистерн у фотосинтезирующих видов, например, у цианобактерий). Единственная крупная кольцевая (у некоторых видов - линейная) двухцепочечная молекула ДНК, в которой содержится основная часть генетического материала клетки (так называемый нуклеоид) не образует комплекса с белками-гистонами (так называемого хроматина). К прокариотам относятся бактерии, в том числе цианобактерии (сине-зелёные водоросли), и археи. Потомками прокариотических клеток являются органеллы эукариотических клеток - митохондрии и пластиды. Основное содержимое клетки, заполняющее весь её объём, - вязкая зернистая цитоплазма.
Эукариотическая клетка
Эукариоты - организмы, обладающие, в отличие от прокариот, оформленным клеточным ядром, отграниченным от цитоплазмы ядерной оболочкой. Генетический материал заключён в нескольких линейных двухцепочных молекулах ДНК (в зависимости от вида организмов их число на ядро может колебаться от двух до нескольких сотен), прикреплённых изнутри к мембране клеточного ядра и образующих у подавляющего большинства (кроме динофлагеллят) комплекс с белками-гистонами, называемый хроматином. В клетках эукариот имеется система внутренних мембран, образующих, помимо ядра, ряд других органоидов(эндоплазматическая сеть, аппарат Гольджи и др.). Кроме того, у подавляющего большинства имеются постоянные внутриклеточные симбионты-прокариоты - митохондрии, а у водорослей и растений - также и пластиды.
2) цитоплазма
Цитопла́зма - внутренняя среда живой или умершей клетки, кроме ядра и вакуоли, ограниченная плазматической мембраной. Включает в себя гиалоплазму - основное прозрачное вещество цитоплазмы, находящиеся в ней обязательные клеточные компоненты - органеллы, а также различные непостоянные структуры - включения.
В состав цитоплазмы входят все виды органических и неорганических веществ. В ней присутствуют также нерастворимые отходы обменных процессов и запасные питательные вещества. Основное вещество цитоплазмы - вода.
Цитоплазма постоянно движется, перетекает внутри живой клетки, перемещая вместе с собой различные вещества, включения и органоиды. Это движение называется циклозом. В ней протекают все процессы обмена веществ.
Цитоплазма способна к росту и воспроизведению и при частичном удалении может восстановиться. Однако нормально функционирует цитоплазма только в присутствии ядра. Без него долго существовать цитоплазма не может, так же как и ядро без цитоплазмы.
Важнейшая роль цитоплазмы заключается в объединении всех клеточных структур (компонентов) и обеспечении их химического взаимодействия. Так же цитоплазма поддерживает тургор(объём) клетки, поддержание температуры.
3)Клеточная стенка
Клеточная стенка - жёсткая оболочка клетки, расположенная снаружи от цитоплазматической мембраны и выполняющая структурные, защитные и транспортные функции. Обнаруживается у большинства бактерий, архей, грибов и растений. Животные и многие простейшиене имеют клеточной стенки.
Клеточные стенки прокариот
Клеточные стенки бактерий состоят из пептидогликана (муреина) и бывают двух типов: грамположительного и грамотрицательного. Клеточная стенка грамположительного типа состоит исключительно из толстого слоя пептидогликана, плотно прилегающего к клеточной мембране и пронизанного тейхоевыми илипотейхоевыми кислотами. При грамотрицательном типе слой пептидогликана существенно тоньше, между ним и плазматической мембраной находится периплазматическое пространство, а снаружи клетка окружена ещё одной мембраной, представленной т. н. липополисахаридом и являющаяся пирогенным эндотоксином грамотрицательных бактерий.
Клеточные стенки грибов
Клеточные стенки грибов состоят из хитина и глюканов.
Характерные особенности грибов
К грибам в широком смысле относят гетеротрофные эукариоты с осмотрофным типом питания. С особенностями питания связаны характерные черты строения и образа жизни этих организмов. Наиболее характерные признаки грибов:
– Наиболее распространенной формой вегетативного тела является мицелий, состоящий из многократно ветвящихся нитей, которые пронизывают субстрат и всасывают из него растворенные в воде питательные вещества.
– Органические вещества в субстрате, которые являются источником питания для грибов, в большинстве случаев находятся в форме биополимеров, неспособных проникать через клеточную мембрану. В клетках грибов вырабатываются ферменты деполимеразы, которые выделяются в субстрат и разрушают полимеры до более простых органических соединений, способных транспортироваться в грибную клетку. Такие ферменты принято называть экзоферментами.
– Активное всасывание веществ из субстрата осуществляется благодаря огромному тургорному давлению, которое в грибных клетках намного выше в сравнении с другими эукариотными организмами.
– Для грибов очень характерно образование спор, которые появляются как при бесполом так и половом размножении. Узнавание грибов возможно не по вегетативному телу, а по спороношению. В связи с расположением вегетативного тела внутри субстрата и неподвижным образом жизни, в момент размножения на грибном мицелии образуются разнообразные спороносящие структуры, которые поднимаются над субстратом и образующиеся на них споры распространяются воздушными потоками на очень далекие расстояния.
Современные системы грибов
По современным представлениям, основанных на изучении геномов экоморфа «грибы» представляет собой сборную группу, в которой выделяют истинные грибы, царство Mycota или Fungi, которых большинство и меньшая по количеству видовых таксонов и менее разнообразная группа грибоподобных организмов (псевдомицеты) входит в состав царства Straminopila, объединяющего также окрашенные водоросли (Ochrophyta). Царство истинные грибы объединяет 4 отдела, различающихся строением вегетативного тела и особенностями размножения: Chytridiomycota, Zygomycota, Ascomycota, Basidiomycota.
Таксономически значимые признаки, используемые при построении систем грибов
–Строение вегетативного тела
– Химический состав и структура клеточной стенки
– Тип полового процесса
– Типы спороношений
Типы талломов грибов и грибоподобных организмов
Амебоидный или плазмодиальный таллом представляет собой лишенную клеточной стенки одноядерную или многоядерную клетку, окруженную плазматической мембраной и уплотненным периферическим белковым слоем – перипластом. Для закрепления в субстрате и увеличения всасывающей поверхности могут образовываться корнеподобные выросты, в совокупности образующие ризомицелий. При размножении или весь таллом превращается в единственный спорангий (холокарпический таллом), или образуется несколько спорангиев (эукарпический таллом).
Мицелиальный таллом и его видоизменения. Основной структурный элемент – гифа. Членистый и нечленистый мицелий. Септы, их образование.
Видоизменения мицелия.
Покоящиеся структуры: хламидоспоры и склероции.
Проводящие и поддерживающие структуры: тяжи и ризоморфы.
Инфекционные структуры: апрессории и гаустории
Дрожжеподобный почкующийся таллом представляет собой отдельные, покрытые оболочкой клетки или неустойчивые, легко распадающиеся цепочки клеток, которые образуются при почковании вследствие нерасхождения материнских и дочерних клеток. Постоянное и временное дрожжеподобное состояние талломов.
Химический состав и структура клеточной стенки грибов
Клеточная стенка грибов многослойная, при этом разные слои образованы различающимися по химическому составу структурными углеводами, которые по химическому составу можно разделить на 3 группы:
полимеры глюкозы (глюкан, хитин, целлюлоза). Глюканы составляют наружный слой клеточной стенки большинства грибов. Внутренний слой грибной клеточной стенки образован цепочками хитина, придавая ей жесткость. Хитин замещает целлюлозу, которая у большинства грибов отсутствует, но входит в состав клеточной стенки оомицетов, которые в настоящее время к типичным грибам не относятся. Деацетилированный хитин получил название хитозан, который в комплексе с хитином образует клеточную стенку зигомицетов.
полимеры других моносахаридов (маннозы, галактозы и др.) в отличие от высших растений, где они составляют основу матрикса под общим названием гемицеллюлоза, менее характерны для грибов. Исключение составляют дрожжи, в клеточных стенках которых особенно много полимеров маннозы, называемых маннаны. Предполагают, что такой состав стенки лучше обеспечивает почкование.
полимеры углеводов, ковалентно связанные с пептидами (гликопротеины) формируют срединный слой многослойной клеточной стенки и играют важную роль, как в поддержании структурной целостности клеток, так и в ее обменных процессах с окружающей средой.
К другим специфическим особенностям грибной клетки относятся: отсутствие пластид , что сближает ее с клеткой животных;
Отсутствие крахмала , который у эумицетов замещается полисахаридом, близким к животному крахмалу гликогену, у оомицетов – полисахаридом, близкому к ламинарину бурых водорослей. Вырабатывается и целый ряд специфических для грибов запасных углеводов.
Выработка специфических вторичных метаболитов , из которых большую роль играют антибиотики, фито– и микотоксины, фитогормоны.
К специфическим особенностям грибов относятся также гетерокариоз и парасексуальный процесс.
У грибов очень широко распространено явление гетерокариоза или разноядерности, при котором в одной клетке длительное время сохраняются ядра, гетероаллельные по некоторым генам. Гетерокариоз заменяет гаплоидным грибам гетерозиготность и способствует быстрой адаптации грибов к меняющимся условиям. Наличие разноядерности обусловлено рядом специфических особенностей грибов:
1. наличие более одного ядра в клетке
2. специфическое строение клеточной перегородки, в которой имеется одно или несколько сквозных отверстий, называемых порами, через которые ядра могут мигрировать из одной клетки в другую
3. гифы внутри одной колонии и даже разных близко расположенных колоний, выросших из разных спор одного вида гриба часто срастаются, в результате чего возможен обмен ядрами разных штаммов.
Парасексуальный (псевдополовой) процесс. Если в гетерокариотичных клетках сливаются ядра, гетероаллельные по какому-либо локусу, возникает гетерозиготное диплоидное ядро. Оно может попасть в спору и дать начало диплоидному гетерозиготному клону. В процессе митозов диплоидные ядра могут возвращаться в гаплоидное состояние вследствие потери одного набора хромосом или в них может происходить обмен участками хромосом (митотический кроссинговер). Оба процесса сопровождаются рекомбинацией родительских генов и, следовательно, фенотипов. Парасексуальная (бесполовая) рекомбинация очень редкое явление, не превышающее одного ядра на миллион, но вследствие огромного числа ядер в мицелии постоянно наблюдается в популяциях грибов.
Размножение – вегетативное, бесполое, половое.
Вегетативное – фрагментация таллома, образование хламидоспор, которые после периода покоя прорастают в мицелий, почкование у дрожжей.
Бесполое размножение у разных грибов может осуществляться подвижными и неподвижными спорами. Зооспоры образуют относительно небольшая группа грибов и грибоподобных организмов – водные и некоторые наземные, у которых отчетливо прослеживаются генетические связи с водными грибами и водорослями. Структура жгутиков является важным диагностическим признаком при отнесении к определенному царству. У подавляющего большинства грибов при бесполом размножении образуются неподвижные споры, что указывает на их очень давний выход на сушу. По месту формирования и локализации различают эндогенные, образующиеся в спорангиях спорангиоспоры и экзогенные (конидии), развивающиеся на специальных гифах – конидиеносцах. Конидии образуются у большинства грибов (аскомицеты, базидиомицеты, некоторые зигомицеты) составляя конидиальное спороношение, которое отличается огромным многообразием и широко используется для диагностики грибов.
Половое размножение грибов имеет свою специфику как в морфологии полового процесс, так и в механизмах генетической и физиологической регуляции пола и передачи наследственной информации.
Соматогамия – самый распространенный и наиболее простой тип полового процесса, заключается в слиянии двух не дифференцированных на гаметы соматических клеток. Иногда он протекает даже без слияния клеток – сливаются ядра внутри клетки. Встречается у большинства базидиомицетов, сумчатых дрожжей и некоторых др. таксонов.
Гаметангиогамия – на гаплоидном мицелии обособляются гаметангии, при половом процессе сливается их содержимое. Такой половой процесс характерен для большинства сумчатых грибов. Вариантом гаметангиогамии является зигогамия у зигомицетов.
Гаметогамия в виде изо- гетеро и оогамии у грибов встречается значительно реже, чем у других эукариот. Изо- и гетерогамия встречается лишь у хитридиомицетов. Классическая оогамия с образованием сперматозоидов и яйцеклеток у грибов не выражена, а встречаются сильно измененные варианты.
По особенностям регуляции пола у грибов выделяют несколько типов полового процесса
Гинандромиксис можно рассмотреть на примере двудомных оомицетов, у которых оогонии и антеридии развиваются на разных талломах, например фитофтора или картофельный гриб. Если генетически однородный штамм выращивать в монокультуре, то он размножается только бесполым способом. Если рядом оказываются мицелии двух штаммов, то у них прослеживаются морфогенетические изменения под действием химических выделений стероидной природы – половых феромонов. Антеридиол индуцирует образование антеридиев у партнера, а оогониол –оогониев. При этом регуляция пола имеет относительный характер: будет ли штамм образовывать антеридии или же оогонии, зависит от количественного соотношения соответствующих феромонов у него и его партнера. Отсюда название полового процесса – гинандромиксис.
Димиксис или гетероталлизм. Уже давно было подмечено, что грибы могут быть гомо- или гетероталличными. У гомоталличных видов при половом процессе сливаются генетически идентичные ядра внутри мицелия. У гетероталличных видов для прохождения полового цикла необходимо на каком-то этапе (различном у разных видов грибов) слияние потомков спор (точнее, их ядер). Для половой совместимости двух штаммов необходимо генетическое различие (гетероаллельность) в определенных локусах, называемых локусами спаривания. У большинства грибов (зигомицеты, аскомицеты, часть базидиомицетов) имеется один локус спаривания с двумя аллелями. Локус спаривания состоит из нескольких генов, котрые управляют синтезом половых феромонов. Такой гетероталлизм называется однофакторным или биполярным. Потомство таких грибов после мейоза разделяется на две самонесовместимые, но взаимно совместимые группы в соотношении 1:1, т.е. вероятность родственного(инбридинга) и неродственного(аутбридинга) скрещивания составляет как и у двуполых высших эукариот 50%.
В геноме высших базидиальных грибов имеется два локуса спаривания – A и B, причем совместимы друг с другом только штаммы гетероаллельные по обоим локусам (Ax Bx совместим с Ay By, но не с Ax By и Ay Bx). Такой гетероталлизм называют двухфакторным или тетраполярным. Он снижает вероятность инбридинга до 25 %.
Диафоромиксис – у высших базидиомицетов имеется не два, а много аллелей локуса спаривания, случайно встречающихся у разных штаммов, слагающих популяцию. Такой контроль спаривания обеспечивает 100% вероятность аутбридинга, так как штаммы, имеющие разные аллели, взаимносовместимы, а аллелей очень много. В результате образуются панмиксные гибридные популляции.
Жизненные циклы грибов также разнообразны как и сами грибы. Основные циклы, их принадлежность к отделам грибов
1.Бесполый цикл характерен для огромной группы несовершенных или митогрибов, утративших половое размножение. Деление их ядер исключительно митотические. Подавляющее большинство митогрибов относится к сумчатым грибам, но в связи с утратой полового процесса, составляют формальную группу несовершенных грибов или дейтеромицетов.
2. Гаплоидный цикл. Вегетативный таллом несет гаплоидные ядра. После полового процесса (сингамии) диплоидная зигота (обычно после периода покоя) делится мейотически – зиготический мейоз. Характерен для зигомицетов и многих хитридиомицетов.
3. Гаплоидно-дикариотический цикл характеризуется тем, что после слияния содержимого гаметангиев (гаметангиогамия) или соматических клеток гаплоидного мицелия (соматогамия) ядра образуют дикарионы (пары генетически разных ядер). Они синхронно делятся, образуя дикариотический мицелий. Половой процесс завершается слиянием ядер дикариона, образующаяся зигота делится мейозом без периода покоя. Мейоспоры составляют половое спороношение сумчатых и базидиальных грибов в виде аскоспор и базидиоспор. При их прорастании образуется гаплоидный мицелий. У подавляющего большинства сумчатых грибов (кроме дрожжей и тафриновых грибов) в жизненном цикле преобладает гаплоидная фаза в виде вегетативного мицелия (анаморфа), дикариотическая фаза кратковременна и представлена аскогенными гифами, на которых образуются сумки (телеоморфа). У базидиальных грибов в жизненном цикле преобладает дикариотическая фаза, гаплоидная фаза кратковременна.
4. Гаплоидно-диплоидный цикл в виде изоморфной смены поколений у грибов встречается редко (некоторые дрожжи и водные хитридиомицеты).
5. Диплоидный цикл характерен для оомицетов и некоторых сумчатых дрожжей. Вегетативный таллом диплоидный, мейоз гаметический, наблюдается при формировании гаметангиев или гамет.
3. Экологические группы грибов.
Грибы и грибоподобные организмы входят во все наземные и водные экосистемы, как важнейшая часть гетеротрофного блока, вместе с бактериями занимая трофический уровень редуцентов. Широкое распространение грибов в биосфере определяется рядом важнейших особенностей:
1. Наличие у большинства мицелиальной структуры таллома. (позволяет быстро осваивать субстрат, иметь большую поверхность контакта со средой).
2. Большая скорость роста и размножения, позволяющие в короткие сроки заселять большие массы субстрата, образуя огромное число спор и распространять их на большие расстояния.
3. Высокая метаболическая активность, проявляющаяся в широком диапазоне значений экологических факторов.
4. Высокая скорость генетической рекомбинации, значительная биохимическая изменчивость, экологическая пластичность.
5. Способность быстро переходить к состоянию анабиоза, переживать неблагоприятные условия в течение длительного времени.
Главным экологическим фактором для грибов является пищевой субстрат. По отношению к этому фактору выделяют основные группы грибов, которые называются трофическими группами.
4. Краткая характеристика отделов грибов.
МИКРОБИОЛОГИЯ, 2010, том 79, № 6, с. 723-733
УДК 582.281(047)
КЛЕТОЧНАЯ СТЕНКА ГРИБОВ: СОВРЕМЕННЫЕ ПРЕДСТАВЛЕНИЯ О СОСТАВЕ И БИОЛОГИЧЕСКОЙ ФУНКЦИИ
© 2010 г. Е. П. Феофилова1
Учреждение Российской академии наук Институт микробиологии им. С.Н. Виноградского РАН, Москва
Поступила в редакцию 05.11.2009 г.
Обзор посвящен мало изученной поверхностной структуре клетки мицелиальных грибов - клеточной стенке (КС). Приводятся данные о методах выделения и проверке чистоты на отсутствие цитоплазмати-ческого содержимого во фракции КС и о ее химическом составе. Подробно рассматриваются структурные (каркасные) и внутриструктурные компоненты КС - аминополисахариды, а- и в-глюканы, белки, липиды, уроновые кислоты, гидрофобины, спорополленин и меланины. Особое внимание уделяется хитину, его новой функции в антистрессовой защите клеток, а также отличиям этого аминополисахарида грибов от хитина водорослей и Arthropoda. Обсуждается феномен апикального роста гифы и участие специальных микровезикул в морфогенезе грибной клетки. Приводятся данные о ферментах, участвующих в синтезе и лизисе КС. В заключение обсуждается функциональная значимость КС у грибов по сравнению с поверхностными структурами высших эукариот.
Ключевые слова: мицелиальные грибы, клеточная стенка, методы выделения, химический состав, апикальный рост, физиологические функции, морфогенез.
Одной из ключевых проблем современной биологии является вопрос - каким образом в процессе развития происходит образование морфологически одинаковых организмов, какие биохимические механизмы и какие клеточные структуры участвуют в этом процессе, который уже миллиарды лет протекает на нашей планете. Не менее интересными представляются данные о том, как влияет стресс на морфогенез и какие биополимеры контролируют внешнюю форму клеток. Как показывают результаты исследований последних лет, растущая гифа ми-целиальных грибов является уникальной моделью, позволяющей понять процесс формообразования клеток и то, как сохраняется в онтогенезе постоянство клеточной морфологии. Очень большой вклад в исследование этого феномена внесли данные по изучению состава и биологической функции КС мицелиальных грибов.
КС растений была открыта в 1665 году, а КС грибов только в начале 18 столетия. Однако эта поверхностная клеточная структура длительное время практически не изучалась, так как считалось, что КС выполняет только "каркасную" функцию, и ее сравнивали со стенами дома, несущими опорную нагрузку. Но в начале XX века отношение к КС резко изменилось, ее начали интенсивно исследовать, но в основном у растений и бактерий. Однако интенсивное развитие грибоводства и биотехнологических производств, в которых продуцентами БАВ были мицелиальные грибы, интенсифицировали разви-
1 Адрес для корреспонденции (e-mail: [email protected]).
тие научных исследований по изучению КС грибов. Данные по этой поверхностной структуре за период до 80-х годов прошлого столетия были обобщены в первой в мире книге по КС грибов . В последующие годы основной массив работ пришелся на конец XX и начало XXI веков. Были проведены основополагающие исследования, посвященные систематике, химическому составу КС, апикальному росту гифы, метаболизму хитина, антигрибным препаратам, гидрофобинам, ковалентно связанным белкам, ферментам, участвующим в образовании КС, ветвлении гиф и лизисе КС . Эти данные не были обобщены, но именно обсуждение их в совокупности позволяет показать, что эта поверхностная структура клетки несет полифункциональную нагрузку и выполняет такие важнейшие функции, как, например, защита клетки от действия неблагоприятных факторов, контроль морфогенеза, участие в процессах репродукции, определение антигенных и адгезивных свойств, контроль процессов диморфизма и образования покоящихся клеток грибов, восприятие внешнего сигнала и передача его на мембрану и внутриклеточные мессенджеры. Учитывая сказанное, мы считали целесообразным остановиться на обсуждении следующих данных: о химическом составе КС, биологической функции ее основных биополимеров, строении КС и межклеточных взаимодействиях в гифах грибов, феномене апикального роста гифы, основных ферментах, участвующих в образовании и лизисе КС.
ХИМИЧЕСКИЙ СОСТАВ КЛЕТОЧНОЙ
СТЕНОКИ ГРИБОВ И БИОЛОГИЧЕСКАЯ ФУНКЦИЯ ЕЕ ОСНОВНЫХ КОМПОНЕНТОВ
Изучение химического состава КС грибов начинается с получения чистой фракции этой структуры, т.е. очистки КС от цитоплазматического загрязнения. Первоначальная процедура - разрушение клеток грибов и отмывка от клеточного содержимого - осуществляется при низкой температуре (около 4- 5°С) для того, чтобы предотвратить разрушительную деятельность деградирующих ферментов. Перед разрушением клеток их замораживают при температуре жидкого азота. Разрушение клеток осуществляется либо в специальных гомогенизаторах, либо на прессах методом "твердого давления". Хорошие результаты дает дальнейшее использование ультразвукового дезинтегратора. Отмывка клеточного содержимого осуществляется холодной водой и повторяется 4-5 раз. Для более тщательного удаления цитоплазматического содержимого используют отмывку с использованием №С1, 8 М мочевины, 1 М аммония или 0.5 М уксусной кислоты. В настоящее время разработаны методы выделения КС грибов в процессе их онтогенеза, причем эти методы существенно различаются для мицелия и покоящихся клеток . В зависимости от направления исследований, в частности, при определении полисахарид-ного состава, из КС извлекают липиды, используя органические растворители, например, хлороформ и метанол в соотношении 2: 1, иногда хорошие результаты дает дополнительная обработка КС серным эфиром. Этот растворитель хорошо подсушивает КС и иногда может заменить лиофильную сушку. Следующий этап - определение чистоты полученной фракции КС. С этой целью используют световую и электронную микроскопию и специализированные методы окраски. Наиболее часто применяется реакция, основанная на взаимодействии I3- с хитозаном. Изолированные чистые КС с раствором Люголя окрашиваются в розовый или фиолетовый цвет, в то время как интактные КС имеют красную окраску . Используется также окраска на наличие ядер с DAPI (4,6-ёкт1ёто-2-ркепуЦп-ёо1). Следует особо подчеркнуть, что выделение чистой фракции КС является очень ответственной процедурой, от которой зависят последующие результаты анализа ее химического состава. В этом отношении интересна работа , в которой у представителя мукоровых грибов обнаружен в мицелии глюкан, точнее, хитозан-глюкановый комплекс, хотя все предыдущие исследования свидетельствовали об отсутствии в мицелии Mucorales глюкана .
В настоящее время компоненты КС грибов разделяют на структурные компоненты (хитин, р-(1- 3)-р(1-6)-глюканы, р-(1-4)-глюкан (целлюлоза), и внутриструктурные (их называют матрикс), к которым стали относить маннопротеины, галакто-ман-нопротеины, ксило-маннопротеины, глюкуроно-
маннопротеины и а-(1-3)-глюкан . Наименее изучены а-глюканы, представляющие собой линейный полимер глюкозы (у Schizosaccharomyces pombe такой глюкан содержит приблизительно около 260 остатков глюкозы). Этот полимер состоит из двух связанных линейных цепей, содержащих около 120 (1-3)-а^-глюкозных остатков и (1-4)-a-D-глюкозных остатков на концах молекулы полимера. Предполагают, что этот глюкан необходим для протекания процесса морфогенеза . Из КС мицелия Penicillium roqueforti выделен нерастворимый в воде глюкан, который после высушивания теряет способность растворяться в щелочи, однако его удалось растворить в 10% растворе хлорида лития в диме-тилсульфоксиде. При изучении глюкана методом метилирования были получены два, три-О-метил-производных глюкозы, отвечающие связям 1 - 3 и 1 -»- 4 между моносахаридами в соотношении около 5: 2, и следы тетра-О-метил-производного, отвечающего концевым невосстанавливающим мо-носахаридным остаткам. Такой результат свидетельствует о линейной структуре молекул, содержащих только 1 -»- 3 и 1 -»- 4 связи между остатками глюкозы. Эти данные были подтверждены анализом спектра 13С-ЯМР полисахарида, из которого также следует а-конфигурация всех остатков глюкозы . Такие полисахариды, особенно после химической модификации (сульфатирования или карбосимети-лирования), способной придать им растворимость в воде, могут быть использованы как биологически активные полимеры и представляют интерес для изучения их биологической функции в клеточной стенке грибов и растений.
Нерастворимые в воде (1 -»- 3)-а^-глюканы были выделены ранее из нескольких видов высших и низших грибов, в том числе из Penicillium chrysoge-num , а родственные а^-глюканы с двумя типами связей (1 -3) и (1 -»- 4), различающиеся их соотношением, находили в высших растениях (Aconi-tum kusnezoffii Reichb ).
Структурный компонент - целлюлоза - характерен для оомицетных грибов, которых по современной систематике относят к псевдогрибам . Мо-носахаридный состав КС включает глюкозу, манно-зу, ксилозу, причем, преобладающим сахаром является глюкоза, составляющая до 68%. В состав КС грибов входят также аминокислоты, липиды (не более 3%) и ^ацетил^-глюкозамин. Глюканы, например, 1,3-глюкан, образуют прочный комплекс с хитином, называемый хитин-глюкановым комплексом (ХГК), который составляет скелет грибной клетки. Этот комплекс присутствует в КС практически у всех грибов, за исключением зигомицетов, что служит надежным систематическим признаком. У некоторых грибов на поверхности КС обнаружен слизистый материал, состоящий из полисахаридов, представленных ß-1,3-глюканами, содержащими только глюкозные единицы, соединенные ß-1,6-связями с каждой третьей единицей глюкозы по