Educational fabrics educational tissues
(meristems), plant tissues consisting of cells that retain the ability to divide for a long time. Thanks to the repeated division of these cells, plants grow throughout their entire life (for some trees this is hundreds or even thousands of years). The second important property of meristem cells is that they give rise to specialized cells that form permanent tissues - integumentary, basal, conductive, mechanical, excretory. Depending on the distribution on the body of the developing plant, four types of meristems are distinguished. Apical, or apical, meristems ensure the growth of shoots and roots in length. Lateral, or lateral, meristems determine the growth of stems and roots in thickness and are called cambium. Intercalary, or intercalary, meristems are temporarily stored in the internodes of the stem and at the bases of young leaves, ensuring the growth of these areas, but then turn into permanent tissues. Wound, or traumatic, meristems arise in places where the plant is damaged, where they form a protective callus. The question is how different tissues are formed from identical meristematic cells, i.e. how cell differentiation occurs remains unresolved.
.(Source: “Biology. Modern illustrated encyclopedia.” Chief editor A. P. Gorkin; M.: Rosman, 2006.)
See what “educational fabrics” are in other dictionaries:
EDUCATIONAL FABRICS- see meristematic tissues... Dictionary of botanical terms
secondary educational tissues- Synonyms: secondary meristems are educational tissues arising either from primary meristems or from permanent tissues. Represented by cambium, phellogen and wound meristems...
primary educational tissues- Synonyms: primary meristems, promeristems, educational tissues derived directly from the meristems of the embryo; Their cells initially have the ability to divide. To P. o. t. include apical meristems of shoot and root, procambium, ... ... Anatomy and morphology of plants
PLANT TISSUE- groups or complexes of cells related by common structure, origin, functions and location. In accordance with this, educational, integumentary, basic, mechanical, conductive and excretory T. r. are distinguished. Educational fabrics, or... ... Agricultural Encyclopedic Dictionary
plant tissue- plant tissues, groups or complexes of cells related by common structure, origin, functions and location. In accordance with this, educational, integumentary, basic, mechanical, conductive and excretory T. r. are distinguished... ... Agriculture. Large encyclopedic dictionary
Cell systems, similar in structure, origin and function, differ in size, shape and location. The composition of the tissue includes tissue fluid (fills the intercellular spaces) and substances located between the cells, for example. calcium salts... ... Biological encyclopedic dictionary
Model of the apical meristem “tunica corpus”. Meristems, or educational tissues, or meristematic tissues (ancient Greek ... Wikipedia
Tunica body Model Meristems, or Educational tissues, or Meristematic tissues (Greek meristos divisible) is a general name for plant tissues consisting of intensively dividing and maintaining physiological activity throughout ... ... Wikipedia
Tunica body Model Meristems, or Educational tissues, or Meristematic tissues (Greek meristos divisible) is a general name for plant tissues consisting of intensively dividing and maintaining physiological activity throughout ... ... Wikipedia
Plant organs are made up of tissues. Plant tissue is a collection of cells that have a similar structure, homogeneous origin and perform the same function. The tissues of higher land plants have reached special development. In plants, educational tissues are distinguished: integumentary, mechanical, conductive, basic and excretory.
Educational tissues, or meristems, serve in the plant for growth; they are of primary origin and secondary. The primary educational tissue is located in the growth cones of roots and stems. Let's look first at root growth cone(Fig. 1, I).
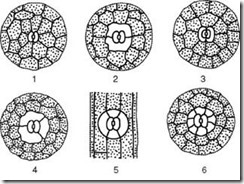
Fig.1. Primary educational tissue in the root growth cone:
I - root growth cone: 1 - cells of educational tissue; 2 - cells of the root cap. II - successive phases of complex nuclear and cell division(karyokinesis, or mitosis): 1 - interphase; 2 - prophase; 3 - metaphase; 4-5 - anaphase; 6-7 - telophase; 8 - cytokinesis.
The tip of the root has a cone-shaped shape, covered with a root cap, under which the primary educational tissue is located. The cells of educational tissue are tetrahedral or multifaceted, have dense protoplasm, large nuclei and thin cellulose membranes. Their vacuoles are not developed or poorly developed. These cells have the ability to divide vigorously. Cell division is preceded by complex division of the nucleus, in which peculiar changes occur. On ready-made, well-prepared and colored preparations, nuclei are visible that are in different phases of complex division, otherwise called karyokinesis (“karyon” - nucleus, “kinesis” - movement) (Fig. 1, II). Numerous observations have established that in the nucleus, thin chromatin filaments first arise from the chromatin substance, which shorten, thicken and turn into individual particles - chromosomes (“chromo” - color, “soma” - body), often curved in the shape of a horseshoe (prophase of division) . Each chromosome in prophase seems to be split lengthwise into two halves, but in reality chromosome doubling (reduplication) occurs. The nucleolus and nuclear envelope gradually disappear. The chromosomes are located in the middle part of the nucleus in one plane in the form of a plate. The halves of each chromosome (“daughter” chromosomes) fit tightly to one another, and in this phase (metaphase) on transverse sections of the root, you can easily count the number of chromosomes and examine their shape. At the poles of the nucleus (even before the disappearance of the nuclear membrane), a system of thin achromatic (non-staining) filaments appears, arranged in the form of two caps. The threads grow and take the form of a spindle. Following this, the divergence of the halves of the chromosomes (daughter chromosomes) begins towards the two opposite poles of the cell (anaphase). The divergence of chromosomes ends with the daughter chromosomes, gathered at the poles of the cell, intertwining with each other, turning into two balls (telophase). At the same time, pectin thickenings appear on the achromatin threads in the middle part of the cell - a phragmoplast is formed, that is, the rudiment of the transverse membrane. Finally, the nuclei are surrounded by a membrane, nucleoli (one or more) are formed in them, and the protoplasm is divided by a transverse membrane (cytokinesis), resulting in the appearance of two new cells.
The process of nuclear and cell division occurs more often at night, its speed depends on temperature and humidity. The division process lasts on average 75-120 minutes, but sometimes lasts several hours, with the first phase (prophase) spending the most time. During cell division, the cell membrane arises as a product of the vital activity of protoplasm and nucleus. First, the median plate appears. It consists of an intercellular substance that glues together the primary cellulose membranes belonging to two cells.
For the first time, the divergence of chromosomes towards the poles of the cell, as well as other details of the division of the nucleus and cell, was established in 1875 by Moscow University professor I. D. Chistyakov.
The essence of the complex division of the nucleus and cell, also called mitosis, apparently lies in the fact that by uniform splitting of chromosomes into two halves (or, more precisely, doubling of chromosomes), daughter cells receive from the mother more or less equal amounts of chromatin substance (deoxyribonucleic acid). acids - DNA), which transmits heredity from the mother cell to the daughter cells. (“Mitos” is a thread. It is usually accepted that mitosis = karyokinesis + cytokinesis). The shape and number of chromosomes are specific characteristics for each plant species (soft wheat 2n = 42 chromosomes, durum wheat 2n = 28, barley 2n = 14, etc.). However, sometimes there is variability in the number of chromosomes, and cells with different chromosomes can be observed in one organism.
In addition to mitosis (or karyokinesis), direct division of the cell nucleus (amitosis) occurs in plants - a simple pulling of the nucleus, and sometimes the entire cell, into two halves.
Along with mitosis, reduction nuclear division is common in plants - meiosis, in which the number of chromosomes in new cells is reduced by half compared to the mother cell. This division occurs in plants during sporulation, after which male and female sex cells (gametes) arise and develop. The latter have half the number of chromosomes in their nuclei, and such nuclei are called haploid in contrast to the diploid nuclei of vegetative cells that make up the vegetative organs of plants. The sexual process of plants consists of the fusion of two gametes (male and female). If there were no reduction of the chromatin substance, that is, a halving of its mass, then in each new sexual generation this mass of substance, and therefore the number of chromosomes, would inevitably increase. In germ cells (gametes), the nuclei are haploid, and only after the fertilization process do the cell nuclei become diploid again.
Reduction division was discovered by scientist V.I. Belyaev, and scientist S.G. Navashin discovered double fertilization in flowering plants and is the founder of a whole school of scientists studying all the details of nuclear and cell division.
Primary educational tissue gives rise to other tissues, since its cells, changing, differentiate. The ability of cells to rapidly divide in the stems and roots of dicotyledonous plants is maintained for life in meristematic cells called cambium, due to which the stem and root thicken. Secondary educational tissues include those that arise from other tissues. An example is phellogen, which often arises from the ground tissue.
FABRICS. CLASSIFICATION OF FABRICS.
The organization of higher plants is based on the principle of cell specialization, which lies in the fact that each cell of the body does not perform all its inherent functions, but only some, but more fully and perfectly.
Fabrics- stable, naturally repeating complexes of cells, similar in origin, structure and adapted to perform one or more functions.
There are various classifications of fabrics, but they are all quite arbitrary.
Depending on the main function, several groups of plant tissues are distinguished.
1. educational fabrics, or meristems,- have the ability to divide and form all other tissues.
2. Covering tissues:
Primary;
Secondary;
Tertiary.
3. Main fabrics- make up most of the plant body. The following main fabrics are distinguished:
Assimilation (chlorophyll-bearing);
Stockers;
Airborne (aerenchyma);
Aquifers.
4. Mechanical fabrics(supporting, skeletal):
Collenchyma;
Sclerenchyma.
5. Conductive fabrics:
Xylem (wood) is an ascending tissue;
Phloem (phloem) is a tissue of descending flow.
6. Excretory tissues:
External:
Glandular hairs;
Hydathodes are water stomata;
Sunbirds;
Internal:
Excretory cells with essential oils, resins, tannins;
Multicellular receptacles for secretions, lacticifers.
Based on the ability of cells to divide, two types of tissues are distinguished: educational, or meristems, And permanent- integumentary, excretory, basic, mechanical, conductive.
The fabric is called simple, if all its cells are identical in shape and function (parenchyma, sclerenchyma, collenchyma). Complex tissues consist of cells that are different in shape, structure and function, but are related by a common origin (for example, xylem, phloem).
There is also a classification of tissues based on their origin (ontogenetic). According to this classification, primary and secondary tissues are distinguished. From the primary meristem located at the top of the shoot and the tip of the root, as well as from the seed embryo, primary constants tissues (epidermis, collenchyma, sclerenchyma, assimilation tissue, epiblema). Cells of permanent tissues are incapable of further division. From cells of a specialized meristem - procambia - are formed primary conductors tissues (primary xylem, primary phloem).
From the secondary meristem - cambium - are formed secondary tissues: secondary xylem, secondary phloem; from phellogen plug, phelloderm, and lentils are formed, which arise when the stem and root thicken. Secondary tissues are typically found in gymnosperms and dicotyledonous angiosperms. The powerful development of secondary tissues - wood and bast - is characteristic of woody plants.
EDUCATIONAL FABRICS
Educational fabrics Thanks to the constant mitotic division of their cells, they ensure the formation of all plant tissues, i.e. actually shape his body. Any cell in its development goes through three stages: embryonic, growth and differentiation stage (that is, the cell acquires a certain function). As the embryo differentiates, the primary meristem is preserved only at the tip of the future shoot (in the growth cone) and at the tip of the root - apical (apical) meristems. The embryo of any plant consists of meristem cells.
Cytological characteristics of meristems. Typical features are most clearly expressed in the apical meristems. These meristems are composed of isodiametric polyhedral cells not separated by intercellular spaces. Their shells are thin, contain little cellulose and are stretchable.
The cavity of each cell is filled with dense cytoplasm with a relatively large nucleus occupying a central position and intensively dividing by mitosis. The hyaloplasm contains many diffusely scattered ribosomes, proplastids, mitochondria and dictyosomes. There are few vacuoles and they are small. Conducting tissues are formed from a meristem that has a prosenchymal shape and large vacuoles - procambium and cambium. Procambium cells are polygonal in cross section, cambium cells are rectangular.
Cells that retain their meristematic properties continue to divide, forming more and more new cells called initials. Some of the daughter cells differentiate, turning into cells of various tissues, they are called derivatives of initials. Initial cells can divide indefinitely many times, and derivatives of initials divide one or more times and develop into permanent tissues.
Based on their origin, primary and secondary meristems are distinguished.
Primary meristems
Primary meristems arise directly from the meristem of the embryo and have the ability to divide. According to their position in the plant, primary meristems can be apical (apical), intercalary (intercalary) and lateral (lateral).
Apical (apical) meristems- such meristems that are located in adult plants at the tops of stems and tips of roots and ensure body growth in length. In the stems, in the growth cone, two meristematic layers are distinguished: the tunica, from which the integumentary tissue and the peripheral part of the primary cortex are formed, and the corpus, from which the inner part of the primary cortex and the central axial cylinder are formed (Fig. 2.3).
Rice. 2.3. Apical meristems of the stem: A- longitudinal section: 1 – growth cone; 2 – leaf primordium; 3 – tubercle of axillary bud;
There are three layers at the root tip:
1) dermatogen, from which the primary integumentary-absorbing tissue - rhizoderm - is formed;
2) periblema, from which the tissues of the primary cortex develop;
3) plerome, forming the tissues of the central axial cylinder.
by origin they can be primary and secondary; on a cross section of the axial organs they look like rings. An example of a primary lateral meristem is the procambium and pericycle. From procambia The cambium and the primary elements of the vascular-fibrous bundles (primary phloem and primary xylem) are formed, while the procambium cells directly differentiate into the cells of the primary conducting tissues.
The lateral meristems are located parallel to the surface of the organ and ensure the growth of the axial organs in thickness.
Intercalary meristems more often they are primary and are preserved in the form of separate areas in zones of active growth in various parts of the plant (for example, at the base of leaf petioles, at the bases of internodes). At the base of internodes in cereals, the activity of this meristem leads to elongation of the internodes, which ensures the growth of the stem in length.
Secondary meristems
Secondary meristems include lateral and wound meristems.
Lateral (lateral) meristems presented cambium And phellogen. They are formed from promeristems (procambium) or permanent tissues by their dedifferentiation. Cambium cells are divided by septa parallel to the surface of the organ (periclinally). Elements of secondary phloem develop from cells deposited outward by the cambium, and elements of secondary xylem develop from cells deposited inward. The cambium, which arose from permanent tissues through dedifferentiation, is called additional In structure and function it does not differ from the cambium, which arose from promeristems. Phellogen is formed from permanent tissues located in the subepidermal layers (under the epidermis). Dividing periclinally, phellogen separates future plug cells (phelleme) outward, and phelloderm cells inward. Thus, phellogen forms a secondary integumentary tissue - periderm. The lateral meristems are located parallel to the surface of the organ and ensure the growth of the axial organs in thickness.
Wound meristems are formed when tissues and organs are damaged. Around the damage, living cells dedifferentiate, begin to divide, and thereby transform into a secondary meristem. Their task is to form a dense protective tissue consisting of parenchyma cells - callus. This tissue is whitish or yellowish in color, its cells have large nuclei and fairly thick cell walls. Callus occurs during grafting, ensuring the fusion of the scion with the rootstock, and at the base of the cuttings. It can form adventitious roots and buds, so it is used to obtain isolated tissue cultures.
COVERING TISSUE
Primary integumentary tissue
TO primary integumentary tissues include the epidermis, the epidermal itself, the parastomatal cells, the guard cells of the stomata and the trichomes.
Pectic substances and cellulose included in the cell wall can be subject to mucus formation with the formation slime And gums. They are polymeric carbohydrates related to pectin substances and are characterized by their ability to swell strongly when in contact with water. Gums in a swollen state are sticky and can be pulled out into threads, while mucus is very blurry and cannot be pulled out into threads. Pectic mucilages are found in representatives of the Liliaceae, Cruciferae, Malvaceae, Linden, and Rosaceae families, in contrast to cellulose mucilages, which are much less common (for example, in Orchids).
 Stomata
They are highly specialized formations of the epidermis, consisting of two bean-shaped guard cells and a stomatal fissure (a kind of intercellular space between them). They are found mainly in the leaves, but are also found in the stem (Fig. 2.6).
Stomata
They are highly specialized formations of the epidermis, consisting of two bean-shaped guard cells and a stomatal fissure (a kind of intercellular space between them). They are found mainly in the leaves, but are also found in the stem (Fig. 2.6).
Rice. 2.6. Stomatal structure: a, b- the skin of a thyme leaf (top view and cross section); V- peel from the stem of Cereus (cactus family); 1 – actual epidermal cells; 2 – guard cells of the stomata; 3 – stomatal fissure; 4 – air cavity; 5 – cells of chlorophyll-bearing parenchyma; A – cuticle; B – cuticular layer – shell with suberin and wax; B – cellulose layer of the wall; G – nucleus with nucleolus; D – chloroplasts
The walls of the guard cells are thickened unevenly: the walls directed towards the gap (abdominal) are significantly thicker compared to the walls directed away from the gap (dorsal). The gap can expand and contract, regulating transpiration and gas exchange. Under the gap there is a large respiratory cavity (intercellular space), surrounded by mesophyll cells of the leaf.
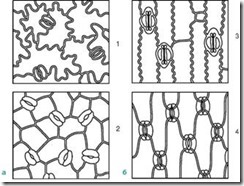
 The guard cells are surrounded by parastomatal cells, which together form stomatal complex(Fig. 2.7). The following main types of stomatal complexes are distinguished:
The guard cells are surrounded by parastomatal cells, which together form stomatal complex(Fig. 2.7). The following main types of stomatal complexes are distinguished:
Rice. 2.7. The main types of stomatal apparatus: 1 – anomocytic (in all higher plants, except horsetails); 2 – diacytic (in ferns and flowering plants); 3 – paracytic (in ferns, horsetails, flowering and oppressive); 4 – anisocytic (only in flowering plants); 5 – tetracytic (mainly in monocots); 6 – ancyclocytic (in ferns, gymnosperms and flowering plants)
1) anomocytic(disorderly) – guard cells do not have clearly defined parastomatal cells; characteristic of all higher plants, excluding conifers;
2) anisocytic(unequal cell) – the guard cells of the stomata are surrounded by three parastomatal cells, one of which is much larger (or smaller) than the others;
3) paracytic(parallel cell) - one peristomatal cell (or more) is located parallel to the guard cells;
4) diacytic(cross-cell) - two parastomatal cells are located perpendicular to the guard cells;
5) tetracite(from Greek tetra- four) – mainly in monocots;
 Stomata are located on the underside of the leaf, but in aquatic plants with floating leaves they are found only on the upper side of the leaf. Based on the shape of the leaf epidermal cells and the location of the stomata, a monocotyledonous plant can be distinguished from a dicotyledonous one (Fig. 2.8). The actual epidermal cells of the leaves of dicotyledonous plants are wavy in outline (Fig. 2.9), while in monocotyledonous plants they are elongated, rhombic in shape.
Stomata are located on the underside of the leaf, but in aquatic plants with floating leaves they are found only on the upper side of the leaf. Based on the shape of the leaf epidermal cells and the location of the stomata, a monocotyledonous plant can be distinguished from a dicotyledonous one (Fig. 2.8). The actual epidermal cells of the leaves of dicotyledonous plants are wavy in outline (Fig. 2.9), while in monocotyledonous plants they are elongated, rhombic in shape.
Rice. 2.8. Location of stomata on the epidermis (view from the surface): A- dicotyledonous plants: 1 - initial letter; 2 – watermelon; b- monocots: 3 – corn; 4 – iris
The types of stomata can be divided according to their level of location relative to the surface of the epidermis as follows.
1.7.1. Stomata located in the same plane as the epidermis. The most common type and is usually not indicated in the description of microscopy of medicinal plant materials, i.e. this paragraph is omitted. Diagnostic signs will be either protruding or submerged stomata.
1.7.2. Protruding stomata - stomata located above the epidermis. Usually, when the microscope microscope is rotated (when the lens is lowered), such stomata are first detected, and only then epidermal cells appear, so it is almost impossible to capture them in a photograph from the surface of a leaf, as well as to depict them in a drawing. In the same plane as the epidermis, such stomata can be seen in transverse sections, but for this, the section must pass through the stomata, which is difficult to obtain given their rare location on the leaf. Such stomata are characteristic, for example, of bearberry leaves.
1.7.3. Submerged stomata - stomata immersed in the epidermis. When observed under a microscope by rotating the microscrew (while lowering the lens), the epidermal cells are first clearly detected, then it becomes possible to more clearly see the contours of the stomata. It is also difficult to display them in photographs and drawings of preparations from the surface. Found in lily of the valley leaves, watch leaves, eucalyptus leaves. Sometimes the recesses in which the stomata are located are lined or covered with hairs and are called stomatal crypts.
1.8. Types of stomatal cells
There are 19 types described in the literature; we have selected only those that are used in the analysis of medicinal plant raw materials**.
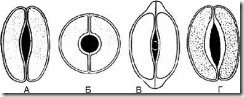
Rice. 63. Types of stomatal cells. A – lentiform; B – spherical; B – cap-shaped; G – scaphoid
1.8.1. Lenticular - 2 identical crescent-shaped cells arranged symmetrically. On the frontal plane, the thickening of the shell is almost uniform. The fissure is fusiform (Fig. 63, A). The type of stomatal cells is characteristic of most plants.
1.8.2. Spheroidal - two identical, strongly circularly curved cells are located symmetrically. On the frontal plane, the thickening of the shell is almost uniform. The slot is round (Fig. 63, B).
1.8.3. Cap-shaped - two identical crescent-shaped cells in the polar parts have thickenings in the form of a cap. The fissure is fusiform (Fig. 63, B). Found in foxgloves.
1.8.4. Scaphoid - the internal walls of stomatal cells are thickened. The fissure is fusiform (Fig. 63, D). Observed in centaury grass and watch leaves.
The mechanism of stomata operation is determined by the osmotic properties of cells. When the leaf surface is illuminated by the sun, an active process of photosynthesis occurs in the chloroplasts of the guard cells. Saturation of cells with photosynthetic products and sugars entails the active entry of potassium ions into the cells, as a result of which the concentration of cell sap in the guard cells increases. There is a difference in the concentration of cell sap of the parastomatal and guard cells. Due to the osmotic properties of the cells, water from the parastomatal cells enters the guard cells, which leads to an increase in the volume of the latter and a sharp increase in turgor. Thickening of the “abdominal” walls of the guard cells facing the stomatal fissure ensures uneven stretching of the cell wall; the guard cells acquire a distinct bean-shaped shape, and the stomatal fissure opens. When the intensity of photosynthesis decreases (for example, in the evening), the formation of sugars in guard cells decreases. The influx of potassium ions stops. The concentration of cell sap in guard cells is reduced compared to parastomatal cells. Water leaves the guard cells by osmosis, lowering their turgor; as a result, the stomatal fissure closes at night.
The cells of the epidermis are tightly closed together, thanks to which the epidermis performs a number of functions:
Prevents the penetration of pathogenic organisms into the plant;
Protects internal tissues from mechanical damage;
Regulates gas exchange and transpiration;
Water and salts are released through it;
Can function as suction tissue;
takes part in the synthesis of various substances, the perception of irritations and the movement of leaves.
Trichomes - outgrowths of epidermal cells of different shape, structure and functions: hairs, scales, bristles, etc. They are divided into covering and glandular. glandular trichomes, unlike coverts, they have cells that secrete secretions. Covering hairs forming a woolly, felt or other cover on the plant, they reflect part of the sun's rays and thereby reduce transpiration. Sometimes the hairs are found only where the stomata are located, for example, on the underside of a coltsfoot leaf. In some plants, living hairs increase the total evaporating surface, which helps accelerate transpiration.
Trichome sizes vary significantly. The longest trichomes (up to 5-6 cm) cover cotton seeds. Covering trichomes have the form of simple single or multicellular, branched or stellate hairs. Covering trichomes can remain alive for a long time or quickly die, filling with air.
They differ from trichomes, which arise only with the participation of epidermal cells. emergents, in the formation of which deeper located tissues of the subepidermal layers also participate.
Anatomical and diagnostic features that are of greatest importance and high variability in determining medicinal raw materials. Hairs can be simple or capitate, which in turn can be unicellular or multicellular. Multicellular hairs can be single-row, double-row or branched.
1.9.1. Simple hairs.
A. Simple unicellular hairs
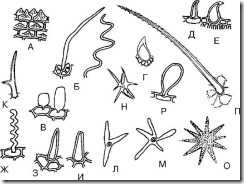
Rice. 68. Simple unicellular hairs. A – papillary; B – filamentous; B – vesicular; G – spinous; D – hook-shaped; E – retort-shaped; F – blunt thread-like corrugated; Z – sharp-cone-shaped; I – blunt-cone-shaped; K – two-ended; L – three-pointed; M, N, O – multi-pointed; P – lumpy; P – club-shaped
1. Papillary (Fig. 68, A; Fig. 69-74) - low but wide outgrowths of epidermal cells. Most often formed on the petals. They can be blunt-apical or acute-apical. They are found on thyme leaves, lily of the valley flowers, tricolor violet flowers, along the edges of centaury leaves, in bearded gentian grass, etc.
2. Cone-shaped (Fig. 68, H, I; Fig. 75-79) - straight hairs inclined at an angle to the surface, ending in a blunt (obtuse-cone-shaped) or sharp (sharp-cone-shaped) end. The most common hairs. Usually they are straight. Hairs inclined towards the surface are observed on thyme leaves. Pointed cone-shaped hairs are found on senna leaves, lingonberry leaves, in tricolor violet grass (on leaves), in thyme grass, etc. Blunt cone-shaped hairs are found on the epidermis of anise fruits and on the petals of tricolor violet flowers.
3. Filiform (Fig. 68, B, G; Fig. 80, 81) – thin and long hairs. Can be straight or corrugated. Straight thread-like hairs are found on the epidermis of hawthorn and raspberry fruits. Corrugated thread-like hairs are observed on the epidermis of violet petals (Fig. 82).
4. Retort-shaped (Fig. 68, E; Fig. 83) - hairs with an expanded base and a narrow curved or straight end. Found in hop fruits and grass.
5. Bladder-shaped (Fig. 68,B; Fig. 84) – hairs in the form of small bubbles. They can be found, for example, in immortelle flowers.
6. Hook-shaped (Fig. 68, E; Fig. 85-87) - hairs, pointed at the top and curved in the form of a hook. These hairs can be found at the base of bearberry leaves, on the surface of lingonberry leaves, senna leaves, and anise fruits.
7. Fringed (Fig. 88, 89) - long outgrowths of epidermal cells located along the edge of a leaf, petal or sepal. Such hairs are found, for example, along the edges of the petals of lily of the valley and violet.
8. Spine-shaped (Fig. 68, D; Fig. 90) – hairs are almost round in shape with a pointed end. Tricolor violets are observed in the grass.
9. Club-shaped (Fig. 68, P; Fig. 91) - hairs with an expanded end part, reminiscent of a club. Found on thyme petals and tricolor violet petals.
10. Double-ended (Fig. 68, K) - hairs branched into two ends. Observed in the grass of shepherd's purse.
11. Three-pointed (multi-pointed) (Fig. 68, L-O) - hairs branched into three (or more) ends. Observed in the grass of shepherd's purse.
12. Lumpy (Fig. 68, P) - cone-shaped hairs with protrusions (bumps) on their surface.
B. Simple multicellular single-row hairs
The contour of a multicellular hair may coincide with that of a unicellular hair, but contain two or more cells and, accordingly, be called a similar unicellular hair indicating the number of cells, for example, a 2-cell retort-shaped hair, an 11-cell filamentous hair, a 10-15-cell cone-shaped hair.

Rice. 92. Simple multicellular hairs. A – cone-shaped; B – bubble-shaped; B – articular; G – whip-shaped; D – feathery; E – T-shaped; F – double row; Z – scaly; I – beam; L – parallel; M – forked; K – bristly
1. Cone-shaped (Fig. 92, A; 93-96). The most widely occurring hairs. There are straight and inclined to the surface, the latter are rare (for example, in thyme herb). Cone-shaped hairs can be sharp- or blunt-cone-shaped. Sharp-cone-shaped hairs are more common (in the herb of oregano, in the leaves of mint, sage, etc.). Blunt cones are found in marigold flowers.
2. Retort-shaped - hairs with an expanded base and a narrow curved or straight end.
3. Bladder-shaped (Fig. 92, B; Fig. 97) – hairs in the form of small bubbles. There are immortelle ovaries on the surface.
4. Filiform - thin and long multicellular hairs.
5. Hook-shaped - hairs pointed at the top and curved in the form of a hook.
6. Caterpillars (Fig. 98, 99) - hairs of almost the same thickness throughout their entire length, consisting of almost identical short cells, with a blunt end and reminiscent of a caterpillar. They are observed in the grass of tricolor violets and in the grass of succession.
7. Fringed (Fig. 100) - long multicellular outgrowths of epidermal cells located along the edge of a leaf, petal or sepal. Sequences are found in the grass.
8. Scourgeous (Fig. 92, D; Fig. 101-104) - hairs that have a multicellular base, consisting of a chain of short cells, and a long filamentous tortuous terminal cell. In the literature, such hairs are described as whip-like, cord-like, thread-like, or felt-like. The hair type is characteristic of the Asteraceae family. Found in yarrow grass, immortelle flowers, coltsfoot leaves, tansy flowers, etc.
9. Articular (Fig. 92, B; Fig. 105, 106) - hairs that have expanded cell bases at the places of their articulation (resembling a joint). Similar hairs are found in motherwort grass, on oregano petals, and rarely in thyme grass.
10. Lumpy - hairs that have protrusions (bumps) on their surface. Two-celled tuberous fibers are observed, for example, in the foliage of thermopsis.
B. Simple multicellular branched hairs
1. Parallel (Fig. 92, L; Fig. 107, 108) - hairs, consisting of two long straight cells (hairs), fused at the bases. Found in linden flowers, rarely in rosehips and hawthorns.
2. Forked (Fig. 92, M; Fig. 109, 110) - hairs, consisting of two long winding cells (hairs), fused at the bases. Observed in linden flowers, very rarely in hawthorn flowers and fruits.
3. Star-shaped (Fig. 111) - hairs, consisting of 3 or more long convoluted cells (hairs), fused at the bases. Found in linden flowers.
4. Scaly (Fig. 92, H) - hairs consisting of a multicellular plate (in the form of a rosette) and a short stalk (the stalk may be absent). Such hairs are found in sea buckthorn. (This name for the hairs is given in accordance with modern botanical terminology; according to other sources, these hairs are called stellate or corymbose scaly).
5. T-shaped (Fig. 92, E) is a two-pointed hair with a single- or multicellular stalk. In the literature they are also described as horn-shaped, rocker-shaped.
6. Cirrus (Fig. 92, E) - multicellular hairs, reminiscent of a branching tree. Found, for example, in mullein.
D. Simple multicellular multirow hairs
1. Double row (Fig. 92, G; Fig. 112, 113) – hairs, the cells of which are arranged in two rows. They can be observed in calendula flowers and dried grass.
2. Beams (Fig. 92, I) - hairs consisting of two or more cells tightly fused with each other, forming a bundle.
3. bristly (Fig. 92 K, Fig. 114) - multicellular hairs, consisting of hairs of different lengths fused with each other and having free pointed ends. Such hairs are usually hairs of the fly (family Asteraceae). In the literature they are also called jagged complex, awl-shaped.
1.9.2. Capitate hairs. Capitate hairs are also called glandular hairs. They can be unicellular or multicellular.

Rice. 115. Capitate hairs. A – with a single-celled stalk and a single-celled head; B – with a two-row leg and a single-cell head; B – with a double-row leg and a double-row head; G – with a unicellular stalk and a multicellular head; D – with a multicellular stalk and a unicellular head; F – with a multicellular stalk and a multicellular head; Z – unicellular (burning); I – thyroid
A. Unicellular capitate hairs
Representatives of this type of hairs can be cone-shaped or retort-shaped, but always have a head at the end. Such hairs are found, for example, on nettle leaves. B. Multicellular capitate (glandular) hairs
1. Hairs with a multicellular head and a unicellular stalk (Fig. 115, D; Fig. 116-120). Found in motherwort grass and bearberry leaves.
2. Hairs that have a single-celled head and a single-celled stalk (Fig. 115, A; Fig. 121-123). Found in motherwort grass, sage leaves, elderberry flowers, mint leaves.
3. Hairs with a unicellular head and a multicellular stalk (Fig. 115, D; Fig. 124). Found in motherwort grass, sage leaves, calendula flowers and chamomile pedicels.
4. Hairs having a multicellular head and a multicellular stalk (Fig. 115, G; Fig. 125). Found in motherwort grass, bearberry leaves, linden flowers, calendula flowers, violet grass.
5. Hairs with a single-celled head and a two-row stalk (Fig. 115, B; Fig. 126). Found in flowers of calendula and immortelle.
6. Hairs having a two-row head and a two-row stalk (Fig. 115, B; Fig. 127-130). They are found in the flowers of calendula and immortelle, and in the grass of marsh grass.
7. Club-shaped multicellular glandular hairs (Fig. 131-133; see Fig. 155, E) - hairs that have a contour with an expanded end part, reminiscent of a club. They can be observed, for example, in lingonberry leaves and violet grass.
8. Thyroid multicellular glandular hairs (Fig. 115, I; Fig. 134) are a shield of polygonal thin-walled cells sitting on a one- or two-cell short stalk. The cuticle of the scutellum moves away from the cells with the essential oil secreted underneath it. They are found in hop fruits.
1.10. The nature of the thickening of cell walls and hairs covering the cuticle
A. Character of thickening of cell walls
1.10.1. Thin-walled (Fig. 135-137). Most hairs are thin-walled. The cell walls of multicellular long thin-walled hairs sometimes collapse, disrupting the even contour of the hair. Such hairs, for example, are found on the lower epidermis of the kakali leaf.
1.10.2. Thick-walled (Fig. 138, 139). They are found in rose hips, string grass, Dahurian lollipop grass, etc.
1.10.3. Unevenly thickened. You can see motherwort in the grass (Fig. 140). Rosehip hairs with oblique transverse pores also belong to this category (Fig. 141, 142).
B. The nature of the hairs covering the cuticle
1.10.4. Smooth surface (Fig. 143). Hairs with such a surface are observed, for example, in plantain leaves, celandine grass, etc.
1.10.5. warty surface (see Fig. 1, G; Fig. 144, 145) - the epidermis forms protrusions in the form of tubercles (warts). There are hairs, for example, in violet grass, in thyme grass, in motherwort grass, in mint leaves, in senna leaves, etc. In this case, the surface of the hair can be slightly warty when small protrusions of the cuticle are formed, and coarsely warty when significant protrusions are formed. In the first case, hairs in violet grass can be cited as an example, in the second - hairs from senna leaves and thyme grass.
1.10.6. Coarse surface (see Fig. 68, P) - the cuticle forms very large protrusions, for example, the hairs of the Thermopsis lanceolata grass.
10.7. Wrinkled surface - folds or waves of the hair cuticle from the base to its apex (Fig. 146). It is observed more often in papillae, for example, in centaury grass and elderberry flowers. However, ordinary hairs can also have a wrinkled surface, for example, in string grass. At the base, the hairs of mint leaves have a wrinkled surface (Fig. 147).
1.10.8. Hatched surface - the epidermis forms short protrusions in the form of streaks (tubercles, warts). It is rare and is an intermediate option between a warty and wrinkled surface. It is observed, for example, on the hairs of elder flowers (Fig. 148).
1.11. Features of hair attachment sites
1.11.1. Common attachment sites (Fig. 149, 150) - hairs are attached to the cell or between the cells of the epidermis. The most common type is found, for example, in coltsfoot leaves, mint leaves, sage leaves, motherwort herb, violet herb, etc.
1.11.2. Outgrowths from epidermal cells (see Fig. 68, A; 69-74). Observed in elderberry flowers, centaury grass, motherwort petals, bearded gentian grass, epidermis of anise fruits, etc.
1.11.3. At the base of the hair a rosette of epidermal cells is formed (Fig. 151, 152). Found in senna leaves, plantain leaves, oregano herb, etc.
1.11.4. Extended hair base (Fig. 153). Found in plantain leaves.
1.11.5. The expanded part of the base of the hair is immersed in the base of subepidermal tissues - emergents (see Fig. 115, 3). It can be observed, for example, in nettle leaves.
1.11.6. Multicellular hair base (Fig. 154). You can observe, for example, strings in the grass.
Often the hairs break off, leaving their attachment points on the epidermis, which also needs to be noted as an anatomical and diagnostic sign of medicinal plant materials.
SECONDARY COVERING TISSUE
The secondary integumentary tissue is called periderma. This is a complex covering tissue of the stems, roots and rhizomes of perennial plants. It replaces the epidermis of the axial organs, which gradually dies and sloughs off. The periderm is formed from phellogen (secondary meristem). Phellogen is formed in the epidermis, subepidermal layer, and even in the deep layers of the axial organs. Phellogen cells divide as follows: plug cells are laid outward, and living parenchyma cells of phelloderm are laid inward. The phelloderm cells of the stems contain chloroplasts.
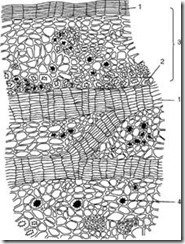
 The plug is made up of dead cells whose cell wall is impregnated with a fat-like substance called suberin. The cells are arranged in even rows, have a rectangular shape (on a cross section), and fit tightly to each other, forming a multilayer case. Cork protects internal living tissues from moisture loss, sudden temperature fluctuations and the penetration of microorganisms. Living tissues lying under the plug need gas exchange and removal of excess moisture. That is why, under the stomata, as a result of the division of the subepidermal layers (even before the appearance of the periderm), and subsequently the phellogen, living, loosely located parenchyma cells with many intercellular spaces, called performing weavingnew, which breaks the epidermis and creates the possibility of gas exchange and transpiration with the external environment. This structural formation is called lentils(Fig. 2.12).
The plug is made up of dead cells whose cell wall is impregnated with a fat-like substance called suberin. The cells are arranged in even rows, have a rectangular shape (on a cross section), and fit tightly to each other, forming a multilayer case. Cork protects internal living tissues from moisture loss, sudden temperature fluctuations and the penetration of microorganisms. Living tissues lying under the plug need gas exchange and removal of excess moisture. That is why, under the stomata, as a result of the division of the subepidermal layers (even before the appearance of the periderm), and subsequently the phellogen, living, loosely located parenchyma cells with many intercellular spaces, called performing weavingnew, which breaks the epidermis and creates the possibility of gas exchange and transpiration with the external environment. This structural formation is called lentils(Fig. 2.12).
Rice. 2.12. The structure of the periderm with the lentil: 1 – the supporting tissue of the lentil; 2 – remains of the epidermis; 3 – cork (phellema); 4 – phellogen; 5 – phelloderm
Lentils, which look like small tubercles, are clearly visible on the surface of shoots of trees and shrubs (Fig. 2.13).
On birch trunks their remains are observed in the form of characteristic transverse black stripes; on aspen they take the form of diamonds.
Tertiary integumentary tissue
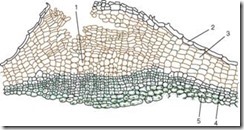
 Crust (rhytide) is a tertiary integumentary tissue that is formed in perennial plants in the root, stem, and rhizome. Every year, in the deeper layers, a new layer of phellogen is laid down and the periderm is formed. The outer layer of the periderm - the cork - isolates all overlying tissues, as a result of which they die. Thus, the collection of numerous periderms with dead tissues between them is the crust (Fig. 2.14).
Crust (rhytide) is a tertiary integumentary tissue that is formed in perennial plants in the root, stem, and rhizome. Every year, in the deeper layers, a new layer of phellogen is laid down and the periderm is formed. The outer layer of the periderm - the cork - isolates all overlying tissues, as a result of which they die. Thus, the collection of numerous periderms with dead tissues between them is the crust (Fig. 2.14).
Rice. 2.14. Oak peel: 1 – layers of cork; 2 – fibers; 3 – remains of the primary crust; 4 – drusen of calcium oxalate
(meristematic cells), and are found in the parts of plants where growth occurs.
1. Cytological features of meristems
Differentiated plant cells cannot divide at all or transform into other types of cells. Therefore, cell division in meristems is necessary to provide new cells for the growth of other tissues, the formation of new organs, and the structure of the plant body. The function of meristematic cells is similar to animal stem cells, which do not differentiate or differentiate slightly, and are capable of continuous cell division. Meristematic cells are small, and the cytoplasm and nucleus completely fill the cell. The vacuoles are extremely small and the cytoplasm does not contain differentiated plastids (chloroplasts or chromoplasts), although they are present in vestigial form (protoplastids). Meristematic cells are tightly packed, with almost no intercellular space. The cell wall is a very thin primary cell wall.
The physiological activity of meristems requires a balance between two antagonistic processes: the formation of new tissues and support for the renewal of the population of meristematic cells.
Meristem- forming plant tissue, from which all other tissues can be formed, meristematic cells for a long time retain the ability to divide with the formation of new unspecialized cells.
The cells of the meristem are flattened, small, densely located next to each other, without intercliniforms, the nucleus is located in the center of the cells, the vacuoles are small, numerous, the cell membrane is primary, there are only plastid precursors.
The cytological features of the croup meristems are typically expressed in the apical meristems. The cells are isodiametric polygons, not separated by intercellular spaces. Cell walls are thin, with little cellulose content. The cytoplasm is dense, the nucleus is large, located in the center. In the cytoplasm there is a large number of ribosomes and mitochondria (intensive synthesis of proteins and other substances occurs). Numerous vacuoles are very small.
The cells of the lateral meristems are unequal in size and shape. This is due to the difference in the cells of permanent tissues from which they are formed. For example, in the cambium there are parenchymal and prosenchymal cells. The parenchyma of the leading complexes is formed from the parenchymal initials, and the conductive elements themselves are formed from the prosenchymal initials.
Meristems are classified according to their position in the plant:
And by origin:
- primary
- secondary.
Primary meristems are in close connection with the apical meristem and directly from them are formed, in particular, the protoderm, the main meristem, and the procambium. Protoderm is a single-layer layer of poorly differentiated cells that covers the apex from the outside; the epidermis is subsequently formed from it. The cortex is formed from the main meristem, and the central cylinder is formed from the procambium, the cells of which are elongated along the axis of the stem.
Secondary meristems penetrate from specialized cells of mature plant organs and ensure their lateral growth (phelogen, pericycle, cambium). Tissues formed from the primary meristem are called primary, and those formed from the secondary meristem are called secondary. Secondary meristems should also include the so-called traumatic meristems, which are formed in places where the plant body is damaged and provide regeneration.
Among the listed types of meristems, the first one to appear in ontogenesis is the apical meristem, from which differentiation of all other tissues occurs. It is located at the tops (APEX) of the stem and root and their lateral branches. A feature of the life activity of a plant organism is, in particular, the preservation of formative activity in the apical meristems throughout the entire ontogenesis, therefore they say that plants are capable of unlimited growth.
Structure of the apical meristem (growth point)
1 - Central part
2 - Peripheral part
2. Apical meristem
4. Intercalary meristem
Intercalary meristem (synonym - intercalary meristem) - cells are located between areas of differentiated tissues, providing intercalary growth.
The intercalary meristem, for example, is located in the nodes of cereals, at the base of petioles, and stamen filaments. These are residual primary meristems. They arise from rope meristems, but their transformation into permanent tissues is delayed compared to other stem tissues. These delicate meristems are especially noticeable in cereals. In the case of bread, they ensure the raising of the stems due to the uneven division of cells from the lower and upper sides of the straw.
5. Wound meristem
Formed when tissues and organs are injured. Living cells surrounding the affected areas dedifferentiate and begin to divide, that is, they turn into a secondary meristem. Wound meristems form Kalus- dense tissue of a bluish or yellowish color, consisting of parenchyma cells of different sizes, arranged in a disorderly manner. Callus cells have large nuclei and relatively thick cell walls. The callus can give rise to any plant tissue or organ. A plug is formed at the periphery; differentiation of callus cells into other tissues is possible. Additional roots and buds can be formed in Kalus. Callus occurs when plants are grafted and ensures the growth of the scion and rootstock; at the base of the cuttings. Calus is also used to obtain isolated tissue cultures.