Chapter 1
BASICS OF CELL PHYSIOLOGY
I. Dudel
Plasma membrane . Animal cells are bounded by a plasma membrane (Figure 1.1). We will dwell on its structure, which is very similar to the structure of many intracellular membranes, in a little more detail. The main matrix of the membrane consists of lipids, mainly phosphatidylcholine. These lipids consist of a hydrophilic head group to which long hydrophobic hydrocarbon chains are attached. In water, such lipids spontaneously form a bilayer film 4–5 nm thick, in which the hydrophilic groups face the aqueous medium, and the hydrophobic hydrocarbon chains are arranged in two rows, forming an anhydrous lipid phase. Cell membranes are lipid bilayers of this type and contain glycolipids, cholesterol and phospholipids (Fig. 1.2). The hydrophilic part of glycolipids is formed by oligosaccharides. Glycolipids are always located on the outer surface of the plasma membrane, with the oligosaccharide part of the molecule oriented like a hair immersed in the environment. Scattered among the phospholipids in almost equal quantities, cholesterol molecules stabilize the membrane. The distribution of various lipids in the inner and outer layers of the membrane is not the same, and even within one layer there are areas in which certain types of lipids are concentrated. This uneven distribution
Rice. 1.1. Schematic drawing of a cell showing the most important organelles
probably has some, as yet unclear, functional significance.
The main functional elements embedded in the relatively inert lipid matrix of the membrane are squirrels(Fig. 1.2). Protein by weight accounts for 25 to 75% in various membranes, but since protein molecules are much larger than lipid molecules, 50% by weight is equivalent to the ratio of 1 protein molecule to 50 lipid molecules. Some proteins penetrate the membrane from its outer to inner surface, while others are fixed in one layer. Protein molecules are usually oriented so that their hydrophobic groups are embedded in the lipid membrane and the polar hydrophilic groups on the surface of the membrane are embedded in the aqueous phase. Many proteins on the outer surface of the membrane are glycoproteins; their hydrophilic saccharide groups face the extracellular environment.
Membrane systems of intracellular organelles .
Approximately half of the cell volume is occupied by organelles isolated from the cytosol by membranes. The total surface area of the membranes of intracellular organelles is at least 10 times greater than the surface of the plasma membrane. The most widely used membrane system is endoplasmic reticulum, representing a network

Rice. 1.2.Schematic representation of the plasma membrane. Proteins are embedded in the phospholipid bilayer, with some of them penetrating the bilayer, while others are only anchored to the outer or inner layer
highly convoluted tubes or sac-like elongated structures; large areas of the endoplasmic reticulum are studded with ribosomes; such a reticulum is called granular or rough (Fig. 1.1). Golgi apparatus also consists of membrane-bound lamellae, from which vesicles or vesicles break off (Fig. 1.1). Lysosomes and peroxisomes- These are small specialized vesicles. In all these various organelles, the membrane and the space it encompasses contain specific sets of enzymes; Inside the organelles, special metabolic products accumulate, used to carry out various functions of the organelles.
CoreAnd mitochondria differ in that each of these organelles is surrounded by two membranes. The nucleus is responsible for the kinetic control of metabolism; the folded inner membrane of the mitochondria is the site of oxidative metabolism; here, due to the oxidation of pyruvate or fatty acids, the high-energy compound adenosine triphosphate (ATP, or ATP) is synthesized.
Cytoskeleton . The cytoplasm surrounding the organelles cannot in any way be considered amorphous; it is permeated by a cytoskeletal network. The cytoskeleton consists of microtubules, actin filaments and intermediate filaments (Fig. 1.1). Microtubules have an outer diameter of about 25 nm; they are formed, like a regular polymer, as a result of the assembly of tubulin protein molecules. Actin filaments– contractile fibers, located in the near-membrane layer and throughout the cell, mainly take part in processes associated with movement. Intermediate filaments consist of blocks of different chemical composition in different types of cells; they form a variety of connections between the two other cytoskeletal elements mentioned above. Organelles and the plasma membrane are also associated with the cytoskeleton, which not only maintains the shape of the cell and the position of the organelles in it, but also determines changes in the shape of the cell and its mobility.
Cytosol . About half of the cell volume is occupied by the cytosol. Since it is approximately 20% protein by weight, it is more of a gel than an aqueous solution. Small molecules, including organic and inorganic ions, dissolved in the aqueous phase. An exchange of ions occurs between the cell and the environment (extracellular space); these metabolic processes will be discussed in the next section. The concentration of ions in the extracellular space is maintained with considerable accuracy at a constant level; the intracellular concentration of each ion also has a specific level, different from that outside the cell (Table 1.1). The most abundant cation in the extracellular environment is Na+ in the cell its concentration is more than 10 times lower. On the contrary, the concentration of K + is highest inside the cell; outside the cell it is lower by more than an order of magnitude. The greatest gradient between extracellular and intracellular concentrations exists for Ca 2+, the concentration of free ions inside the cell is at least 10,000 times lower than outside it. Not all ions are dissolved in the cytosol; some of them are adsorbed on proteins or deposited in organelles. For example, in the case of Ca 2+, bound ions are much more numerous than free ones. Most of the cytosolic proteins are enzymes, with the participation of which many processes of intermediate metabolism are carried out: glycolysis and gluconeogenesis, synthesis or destruction of amino acids, protein synthesis on ribosomes (Fig. 1.1). The cytosol also contains lipid droplets and glycogen granules, which serve as reserves of important molecules.
Table 1.1.Intra- and extracellular ion concentrations in muscle cells of homeothermic animals. A – – “high molecular weight cellular anions”
|
Intracellular concentration |
Extracellular concentration |
||
|
Na+ |
12 mmol/l |
Na+ |
145 mmol/l |
|
155 mmol/l |
K+ |
4 mmol/l |
|
|
Ca 2+ |
10 –7 10 –8 mmol/l |
Ca 2+ |
2 mmol/l |
|
C l – |
4 mmol/l |
C1 – |
120 mmol/l |
|
HCO 3 – |
8 mmol/l |
HCO 3 – |
27 mmol/l |
|
A – |
155 mmol/l |
Others cations |
5 mmol/l |
|
Resting potential –90 mV |
|||
1.2. Exchange of substances between the cell and the environment
We have briefly described the structure of the cell so that we can use this description to review the basics of cellular physiology. In no case can a cell be considered a static entity, since there is a constant exchange of substances between various intracellular compartments, as well as between compartments and the environment. Cell structures are in dynamic equilibrium, and the interactions of cells with each other and with the external environment are a necessary condition for maintaining the life of a functioning organism. In this chapter we will look at the fundamental mechanisms of such exchange. In subsequent chapters these mechanisms will be considered as applied to the nerve cell and its functions;
however, the same mechanisms underlie the functioning of all other organs.
Diffusion.The simplest process of moving matter is diffusion. In solutions (or gases), atoms and molecules move freely, and differences in concentrations are balanced due to diffusion. Let's consider two volumes filled with liquid or gas (Fig. 1.3), in which substances have concentrations c 1 and c 2 and separated by a layer with surface area A and thickness d. Flow of matter m during time t described Fick's first law of diffusion:
dm/ dt= DA/ d ( C 1 –C 2)=DA/ dD C(1)
where D is the diffusion coefficient, constant for a given substance, solvent and temperature. In a more general form for the concentration difference dc at a distance dx
dm/dt= –D A dc/dx,(2)
the flow through section A is proportional to the concentration gradient dc/dx . The minus sign appears in the equation because the change in concentration in the x direction is negative.
Diffusion is the most important process due to which most of the molecules in aqueous solutions move over short distances. This also applies to their movement in the cell insofar as diffusion is not impeded by membranes. Many substances can diffuse freely through lipid membranes, especially water and dissolved gases such as O 2 and CO 2 . Fat soluble

Rice. 1.3.Quantitative diffusion scheme. The two spaces are separated by a layer of thicknessdand area A. C; – high concentration of particles in the left part of the volume, C: – low concentration of particles in the right parts, pink surface– concentration gradient in the diffusion layer. Diffusion flow dm/dt – see equation (1)
substances also diffuse well through membranes; This also applies to polar molecules of a fairly small size, such as ethanol and urea, while sugars pass through the lipid layer with difficulty. At the same time, lipid layers are practically impermeable to charged molecules, including even inorganic ions. For non-electrolytes, the diffusion equation (1) is usually transformed by combining the characteristics of the membrane and the diffusing substance into one parameter-permeability (P):
dm/dt=P AD c.(3)
In Fig. 1.4 are compared permeability (P) of the lipid membrane for various molecules.
Diffusion through membrane pores . The plasma membrane (and other cell membranes) is permeable not only to substances diffusing through the lipid layer, but also to many ions, sugars, amino acids and nucleotides. These substances cross the membrane through pores formed transport proteins, immersed in the membrane. Inside such proteins there is a water-filled channel with a diameter of less than 1 nm, through which small molecules can diffuse. They move along a concentration gradient, and if they carry a charge, then their movement through the channels is also regulated by the membrane potential. Membrane channels are relatively selective

Rice. 1.4.Permeability of artificial lipid bilayers to various substances
in relation to the type of molecules that can pass through them, there are, for example, potassium, sodium and calcium channels, each of which is impermeable to almost any ion except a specific one. Such selectivity is determined by the charge or structure of binding sites in the channel walls, which facilitates the transport of a specific molecule and prevents the penetration of other substances through the channel (Fig. 1.5, A) .
Behind the behavior membrane ion channels easy to observe, since the current arising from the movement of ions can be measured, even for a single channel. It is shown that the channels spontaneously and with high frequency change their state from open to closed. The potassium channel is characterized by current pulses with an amplitude of about 2 pA (2 10 –12 A) and a duration of several milliseconds (see Fig. 2.12, p. 37) [3]. During this period, tens of thousands of ions pass through it. The transition of proteins from one conformation to another is studied using X-ray diffraction, Mössbauer spectroscopy and nuclear magnetic resonance (NMR). Proteins are thus very dynamic, mobile structures, and the channel passing through the protein is not just a rigid, water-filled tube (Fig. 1.5, A), but a labyrinth of rapidly moving molecular groups and charges. This dynamic characteristic of the channel is reflected in energy profile of the channel, shown in Fig. 1.5, B. Here, the abscissa axis represents the length of the channel from the external solution with ion concentration C 0 and potential 0 to the internal solution with concentration C 1 and potential E. The ordinate axis

Rice. 1.5.A. Schematic of a protein forming a potassium channel embedded in the lipid bilayer of the plasma membrane. Four negative charges are fixed on the “wall” of the channel. B. Schematic energy profile of the channel shown in Fig. A. The ordinate axis shows the values of kinetic energy required for passage of the channel; along the abscissa axis – the distance between the inner and outer surfaces of the membrane. The energy minima correspond to the binding sites of positively charged ions with fixed negative charges in the channel wall. The energy maxima correspond to diffusion obstacles in the channel. The conformation of the channel protein is assumed to spontaneously oscillate; energy profile options are shown with solid and dashed lines; these oscillations greatly facilitate the binding of ions when overcoming the energy barrier (modified)
the ion energy levels at the channel binding sites are presented; the peak in the graph represents the permeability barrier that the ion energy must exceed to penetrate the channel, and the “dip” in the graph represents a relatively stable state (binding). Despite the obstacle of the energy peak, the ion can penetrate the channel if the energy profile changes spontaneously; the ion may thus suddenly find itself “on the other side” of the energy peak and may continue to move into the cell. Depending on the charge, size and degree of hydration of the ion and its ability to bind to channel wall structures, the energy profile of the channel varies for different ions, which may explain the selectivity of individual channel types.
Diffusion equilibrium of ions . The diffusion of various ions through membrane channels should lead to the elimination of differences in concentrations between the extra- and intracellular environments. As, however, can be seen from table. 1.1, such differences remain, therefore, there must be some equilibrium between diffusion and other transport processes across the membrane. The next two sections deal with the ways in which this equilibrium is achieved. In the case of ions, the diffusion equilibrium is affected by their charge. Diffusion of uncharged molecules is ensured by the difference in concentrations dc , and when the concentrations are equalized, the transport itself stops. Charged particles are additionally affected by the electric field. For example, when a potassium ion leaves a cell along its concentration gradient, it carries a single positive charge. Thus, the intracellular environment becomes more negatively charged, resulting in a potential difference across the membrane. The intracellular negative charge prevents new potassium ions from leaving the cell, and those ions that do leave the cell will further increase the charge on the membrane. The flow of potassium ions stops when the action of the electric field compensates for the diffusion pressure due to the difference in concentration. Ions continue to pass through the membrane, but in equal amounts in both directions. Therefore, for a given difference in ion concentrations on the membrane, there is equilibrium potential E ion at which the flow of ions through the membrane stops. The equilibrium potential can be easily determined using Nernst equations:
Eion= RT/ zF* lnC out/ C in(4)
where R – gas constant, T – absolute temperature, z – ion valence (negative for anions) C out – extracellular ion concentration, C in – intracellular ion concentration, F Faraday number. If we substitute constants into the equation, then at body temperature (T = 310 K) the equilibrium potential for potassium ions E K is equal to:
Ek= –61 mB log /(5)
If [ K + out ]/[ K + in ] = 39, as follows from table. 1.1 then
Ek= –61 m B log 39= –97 mV.
Indeed, it was found that all cells have membrane potential; in mammalian muscle cells its level is about -90 mV. Depending on conditions and relative ion concentrations, cells can have a membrane potential ranging from -40 to -120 mV. For the cell in the example above (Table 1.1) resting potential, equal to approximately -90 mV, indicates that the fluxes of potassium ions through membrane channels are approximately in equilibrium. This is not surprising, since in a resting membrane the open state of potassium channels is most likely, i.e. the membrane is most permeable to potassium ions. The membrane potential, however, is determined by the flows of other ions.
The ease with which uncharged particles can diffuse through the membrane is quantified in equation (3). Permeability to charged particles is described by a slightly more complex equation:
P= m RT/ dF(6)
Where m– ion mobility in the membrane, d – membrane thickness, a R, T and F – known thermodynamic constants. The permeability values for various ions determined in this way can be used to calculate the membrane potential Em , when potassium, sodium and chlorine ions pass through the membrane simultaneously (with permeability P K, P Na and P Cl respectively). It is assumed that the potential drops uniformly in the membrane, so that the field strength is constant. In this case it applies Goldman equation, or constant field equation :
Em= R T/ F * ln(P K + P Na + P Cl )/ (P K + P Na + P Cl )(7)
For most cell membranes P K approximately 30 times higher than P Na (see also section 1.3). Relative value P Cl varies greatly; for many membranes P Cl small compared to R K , however for others (for example, in skeletal muscles) P Cl , significantly higher than P K.
Active transport, sodium pump . The previous section describes the passive diffusion of ions and the resulting membrane potential at given intra- and extracellular ion concentrations. However, as a result of this process, the concentration of ions inside the cell is not automatically stabilized, since the membrane
potential is slightly more electronegative than EK, and much compared to E Na (about +60 mV). Due to diffusion, intracellular ion concentrations, at least potassium and sodium, must be equalized with extracellular ones. The stability of the ion gradient is achieved through active transport: membrane proteins transport ions across the membrane against electrical and/or concentration gradients, consuming metabolic energy for this. The most important process of active transport is the work Na/K – a pump that exists in almost all cells;
the pump pumps sodium ions out of the cell while simultaneously pumping potassium ions into the cell. This ensures a low intracellular concentration of sodium ions and a high concentration of potassium ions (Table 1.1). The concentration gradient of sodium ions on the membrane has specific functions related to the transmission of information in the form of electrical impulses (see Section 2.2), as well as the maintenance of other active transport mechanisms and regulation of cell volume (see below). Therefore, it is not surprising that more than 1/3 of the energy consumed by a cell is spent on the Na/K pump, and in some of the most active cells up to 70% of energy is spent on its operation.
The Na/K transport protein is an ATPase. On the inner surface of the membrane, it breaks down ATP into ADP and phosphate (Fig. 1.6). The energy of one ATP molecule is used to transport three sodium ions from the cell and simultaneously two potassium ions into the cell, i.e., in total, one positive charge is removed from the cell in one cycle. Thus, the Na/K pump is electrogenic(creates an electrical current across the membrane), which leads to an increase in electronegativity of the membrane potential by approximately 10 mV. The transport protein performs this operation at high speed: from 150 to 600 sodium ions per second. The amino acid sequence of the transport protein is known, but the mechanism of this complex metabolic transport is not yet clear. This process is described using energy profiles of the transfer of sodium or potassium ions by proteins (Fig. 1.5,5). By the nature of the changes in these profiles, associated with constant changes in the conformation of the transport protein (a process that requires energy), one can judge the stoichiometry of the exchange: two potassium ions are exchanged for three sodium ions.
Na/K pump, same as isolated Na+ /K + -dependent membrane ATPase, specifically inhibited by the cardiac glycoside ouabain (strophanthin). Since the operation of the Na/K pump is a multi-stage chemical reaction, it, like all chemical reactions, is largely dependent on temperature, which

Rice. 1.6.Diagram of a Na/K pump-ATPase (immersed in the lipid bilayer of the plasma membrane), which in one cycle removes three Na + ions from the cell against potential and concentration gradients and brings two K ions into the cell + . During this process, one ATP molecule is broken down into ADP and phosphate. The diagram shows the ATPase as a dimer consisting of a large (functional) and small subunits; in the membrane it exists as a tetramer formed by two large and two small subunits
shown in Fig. 1.7. Here, the flux of sodium ions from muscle cells is shown relative to time; this is practically equivalent to the flow of sodium ions mediated by the operation of the Na/K pump, because the passive flow of sodium ions against concentration and potential gradients is extremely small. If the drug is cooled by approximately 18°C, the flow of sodium ions from the cell will quickly decrease by 15 times, and immediately after heating it will be restored to its original level. This decrease in the flow of sodium ions from the cell is several times greater than that which would correspond to the temperature dependence of the diffusion process or a simple chemical reaction. A similar effect is observed when metabolic energy is depleted as a result of dinitrophenol (DNP) poisoning (Fig. 1.7,5). Consequently, the flow of sodium ions from the cell is ensured by an energy-dependent reaction - an active pump. Another characteristic of the pump, along with significant temperature and energy dependence, is the presence of a saturation level (like all other chemical reactions); this means that the speed of the pump cannot increase indefinitely as the concentration of transported ions increases (Fig. 1.8). In contrast, the flux of a passively diffusing substance increases in proportion to the concentration difference in accordance with the law of diffusion (equations 1 and 2).

Rice. 1.7. A, B. Active Na transport + . Y axis: flux of radioactive 24 Na + from the cell (imp./min). Abscissa axis: time since the start of the experiment. A. The cell is cooled from 18.3 °C to 0.5 °C; flow Na+ from the cell during this period is inhibited. B. Suppression of Na + flux from the cell with dinitrophenol (DNP) at a concentration of 0.2 mmol/l (modified)
In addition to the Na/K pump, the plasma membrane contains at least one more pump - calcium; this pump pumps calcium ions (Ca 2+) out of the cell and is involved in maintaining their intracellular concentration at an extremely low level (Table 1.1). The calcium pump is present in very high density in the sarcoplasmic reticulum of muscle cells, which accumulate calcium ions as a result of the breakdown of ATP molecules (see Chapter 4).
Effect of the Na/K pump on membrane potential and cell volume . In Fig. Figure 1.9 shows the various components of the membrane current and shows the intracellular concentrations of ions that

Rice. 1.8.The relationship between the rate of transport of molecules and their concentration (at the entrance to the channel or at the binding site of the pump) during diffusion through the channel or during pumping transport. The latter becomes saturated at high concentrations (maximum speed, Vmax ) value on the x-axis corresponding to half the maximum pump speed ( Vmax /2), is the equilibrium concentration TO m

Rice. 1.9.Diagram showing Na+ concentrations , K+ and Cl – inside and outside the cell and the pathways for the penetration of these ions through the cell membrane (through specific ion channels or using a Na/K pump. At given concentration gradients, equilibrium potentials E Na, E K and E С l – equal to those indicated, membrane potential Em = – 90 mV
ensure their existence. An outward current of potassium ions is observed through potassium channels, since the membrane potential is slightly more electropositive than the equilibrium potential for potassium ions. The overall conductance of sodium channels is much lower than that of potassium channels, i.e. sodium channels are open much less frequently than potassium channels at resting potential; however, approximately the same number of sodium ions enter the cell as potassium ions exit the cell, because large concentration and potential gradients are required for sodium ions to diffuse into the cell. The Na/K pump provides ideal compensation for passive diffusion currents, as it transports sodium ions out of the cell and potassium ions into it. Thus, the pump is electrogenic due to the difference in the number of charges transferred into and out of the cell, which at normal speed of its operation creates a membrane potential of approximately 10 mV is more electronegative than if it were produced by passive ion fluxes alone (see Equation 7). As a result, the membrane potential approaches the potassium equilibrium potential, which reduces the leakage of potassium ions. Na activity The /K pump is regulated by the intracellular concentration of sodium ions. The speed of the pump slows down as the concentration of sodium ions to be removed from the cell decreases (Fig. 1.8), so that the pump operation and the flow of sodium ions into the cell balance each other, maintaining the intracellular concentration of sodium ions at a level of approximately 10 mmol/L.
To maintain equilibrium between pumping and passive membrane currents, many more Na/K pump molecules are needed than channel proteins for potassium and sodium ions. When the channel is open, tens of thousands of ions pass through it in a few milliseconds (see above), and since the channel usually opens several times per second, in total more than 10 5 ions pass through it during this time. A single pump protein moves several hundred sodium ions per second, so the plasma membrane must contain about 1000 times more pump molecules than channel molecules. Measurements of channel currents at rest showed the presence on average of one potassium and one sodium open channel per 1 μm 2 membrane; It follows from this that about 1000 molecules of the Na/K pump should be present in the same space, i.e. the distance between them is on average 34 nm; The diameter of the pump protein, as a channel protein, is 8–10 nm. Thus, the membrane is quite densely saturated with pumping molecules
The fact that the flow of sodium ions into the cell, and potassium ions out of the cell, is compensated by the operation of the pump, there is another consequence, which consists in maintaining a stable osmotic pressure and constant volume. Inside the cell there is a high concentration of large anions, mainly proteins (A - in Table 1.1), which are not able to penetrate the membrane (or penetrate through it very slowly) and therefore are a fixed component inside the cell. To balance the charge of these anions, an equal number of cations is needed. Due to the action of the Na/K pump, these cations are mainly potassium ions. A significant increase in the intracellular ion concentration could only occur with an increase in the concentration of anions due to the flow of C1 - along the concentration gradient into the cell (Table 1.1), but the membrane potential counteracts this. Incoming current Cl – observed only until the equilibrium potential for chlorine ions is reached; this is observed when the chlorine ion gradient is almost opposite to the potassium ion gradient, since chlorine ions are negatively charged (Equation 4). Thus, a low intracellular concentration of chlorine ions is established, corresponding to a low extracellular concentration of potassium ions. The result is a limitation of the total number of ions in the cell. If the membrane potential drops when the Na/K pump is blocked, for example during anoxia, then the equilibrium potential for chlorine ions decreases, and the intracellular concentration of chloride ions increases accordingly. Restoring the balance of charges, potassium ions also enter the cell; the total concentration of ions in the cell increases, which increases osmotic pressure; this forces water into the cell. The cell swells. This swelling is observed in vivo in conditions of lack of energy.
Concentration gradient Na + as a driving force for membrane transport . The importance of the Na/K pump for the cell is not limited to the stabilization of normal K + and Na + gradients on the membrane. Energy stored in membrane gradient Na+ , is often used to facilitate membrane transport of other substances. For example, in Fig. 1.10 shows “simport” Na+ and sugar molecules into the cell. The membrane transport protein transports the sugar molecule into the cell even against a concentration gradient, while at the same time Na + moves along the concentration and potential gradient, providing energy for transport of sugars. This transport of sugars is entirely dependent on the existence of a high gradient Na+ ; if intracellular concentration Na+ increases significantly, the transport of sugars stops. For various c akharov, there are different simport systems. Amino acid transport into a cell similar to transport c akharov shown in Fig. 1.10; it is also provided by the gradient Na+ , There are at least five different symport systems, each specialized for one group of related amino acids.
In addition to simport systems, there are also "anti-porters". One of them, for example, transfers one calcium ion out of the cell in one cycle in exchange for three incoming sodium ions (Fig. 1.10). Energy for Ca 2+ transport is formed due to the entry of three sodium ions along a concentration and potential gradient. This energy is sufficient (at resting potential) to maintain a high calcium ion gradient (from less than 10–7 mol/L inside the cell to approximately 2 mmol/L outside the cell).
Endo- and exocytosis . For certain substances that enter the cell or must be excreted

Rice. 1.10.Proteins embedded in the lipid bilayer of the membrane mediate the symport of glucose and Na + into the cell, as well as Ca2+/Na+ –antiport, in which the driving force is the Na + gradient on the cell membrane
from it, there are no transport channels; These substances include, for example, proteins and cholesterol. They can pass through the plasma membrane into vesicles, or bubbles, through endo- and exocytosis. In Fig. Figure 1.11 shows the main mechanisms of these processes. In exocytosis, certain organelles (see below) form vesicles filled with a substance that needs to be removed from the cell, such as hormones or extracellular enzymes. When such vesicles reach the plasma membrane, their lipid membrane fuses with it, thus allowing the contents to escape into the external environment. In the opposite process, endocytosis, the plasma membrane invaginates, forming a pit, which then deepens and closes, forming an intracellular vesicle filled with extracellular fluid and some macromolecules. To ensure this membrane fusion and closure of the vesicle, the contractile elements of the cytoskeleton act in conjunction with the membranes themselves (see below). Endocytosis does not always simply involve the capture of the extracellular environment into the cell. The cell membrane often contains specific receptors for macromolecules, such as insulin or antigens, often organized into specialized groups. After these macromolecules bind to their receptors, endocytosis occurs in the membrane area surrounding the receptor, and the macromolecule is selectively transported into the cell (Fig. 1.12, B).
Endo- and exocytosis occur continuously in cells. The amount of membrane material turnover is significant; within 1 hour, the macrophage absorbs double the surface area of its cytoplasmic membrane in the form of vesicles. In most cells, the turnover of membrane material does not occur as intensively, but it should still be significant.

Rice. 1.11.Exocytosis and endocytosis. Up: the intracellular vesicle fuses with the lipid bilayer of the plasma membrane and opens into the extracellular space. This process is called exocytosis. At the bottom: the plasma membrane invaginates in a small area and releases a vesicle filled with extracellular material. This process is called endocytosis
1.3. Transport of substances within the cell
Endo- and exocytosis are not only processes of transport of substances across the cell membrane, but also processes of exchange of membranes - the structural components of the cell itself. The subject of consideration in this section are other similar transport processes in the cell and its organelles.

Rice. 1.12. A–B. Scheme of processes including exo- and endocytosis. A. The protein synthesized in the granular endoplasmic reticulum is transported through the Golgi apparatus to the plasma membrane, where it is secreted by exocytosis. B. Cholesterol bound to LDL (low-density lipoprotein) particles attaches to the plasma membrane, induces the formation of an endocytic vesicle in this region of the membrane and is transported to lysosomes, where it is released. IN. Extracellular material captured during endocytosis (in the figure on right), transported through the cell in vesicles, or vesicles, and released through exocytosis (in the figure left)
Diffusion . Naturally, in the cytosol the difference in concentrations is eliminated due to diffusion; the same is true for the fluids contained in the organelles. Due to the high concentration of dissolved protein, diffusion here is much slower than in water. Lipid membranes—around cells and within organelles—are two-dimensional fluids in which diffusion occurs. Lipids in a membrane bilayer diffuse within their own layer, rarely moving from one to another. The proteins immersed in them are also quite mobile; they rotate around an axis perpendicular to the membrane or diffuse laterally with very different diffusion constants, 2–10,000 times slower than phospholipids. So, if some proteins move freely in the lipid layer and at the same speed as the lipid molecules themselves, then others are anchored, i.e. quite firmly connected to the cytoskeleton. There are “permanent” aggregates of specific proteins in the membrane, for example, pre- and postsynaptic structures of nerve cells. Freely moving proteins can be demonstrated by binding them to fluorescent dyes, the luminescence of which is induced by briefly illuminating a small area of the membrane. Such experiments show that in less than 1 min, proteins bound to the dye are uniformly distributed over the membrane over distances of up to 10 μm.
Active transport in organelle membranes .
Active transport processes, which play a vital role in the functioning of the plasma membrane, also occur inside the cell, in the membranes of organelles. The specific contents of various organelles are created partly due to internal synthesis, and partly due to active transport from the cytosol. One example of the latter is the above-mentioned Ca 2+ pump in the sarcoplasmic reticulum of muscle cells. It is especially interesting that in the case of ATP synthesis in mitochondria, the opposite principle applies to what occurs in the ATPase pumps of the plasma membrane (Fig. 1.6). During ATP synthesis, oxidative metabolism leads to the formation of a steep gradient H+ on internal membranes. This gradient is the driving force for the reverse process of the pumping cycle of active transport of molecules: H + ions move across the membrane along the gradient, and the energy released as a result provides the synthesis of ATP from ADP and phosphate. The resulting ATP, in turn, provides energy to the cell, including for active transport.
Transport in vesicles . The cell has a large number of organelles and associated vesicles (Fig. 1.1). These organelles, and especially the vesicles, are in constant motion, transporting their contents to other organelles or to the plasma membrane. Vesicles can also migrate from the cell membrane to organelles, as in endocytosis.
Process protein secretion shown in Fig. 1.12, A. The protein is synthesized near the cell nucleus on ribosomes associated with the endoplasmic reticulum (the so-called granular, or rough, endoplasmic reticulum); Once in the endoplasmic reticulum, the protein is packaged into transport vesicles, which separate from the organelle and migrate to the Golgi apparatus. Here they merge with the cisternae of the Golgi apparatus, where the protein is modified (i.e., converted into a glycoprotein). At the ends of the cisternae, the vesicles separate again. Secretory vesicles carrying the modified protein move to the plasma membrane and release their contents by exocytosis.
Another example of a transport pathway in a cell is shown in Fig. 1.12, B; This is the absorption of cholesterol by the cell. Cholesterol transported in the blood is mainly bound to proteins, such as particles "low density lipoprotein"(LDL). Such particles attach to specific membrane areas containing LDL receptors, where endocytosis occurs and LDL is transported into the cell in “bordered” vesicles. These vesicles fuse to form endosomes and lose their “edging” during this process. Endosomes, in turn, merge with primary lysosomes, containing predominantly hydrolytic enzymes, and form secondary, larger lysosomes. In them, cholesterol is released from LDL particles and diffuses into the cytosol, where it becomes available, for example, for the synthesis of lipid membranes. Vesicles that do not contain LDL are also separated from endosomes, which move in a special way to the plasma membrane and merge with it, returning membrane material and, probably, receptors for LDL. From the moment the LDL particle binds to the membrane until cholesterol is released from the secondary lysosome, 10-15 minutes pass. Impairments in the binding and absorption of LDL, i.e., in the supply of cholesterol to the cell, play a decisive role in the development of a serious and widespread disease, atherosclerosis (“hardening” of the arteries).
There are many other transport routes similar to those shown in Fig. 1.11 and 1.12, A, with the help of which specific vesicles move in the cell. It is not known exactly how they move, but cytoskeletal elements are probably involved in this process. Vesicles can slide along microtubules, in which case the energy for movement is apparently provided by the protein associated with the vesicles, ATPase (see below). It remains completely unclear how many different vesicles, moving one after another in all directions, reach their destination. They obviously need to be “marked” in a way that is recognized by the transport system and converted into targeted movement.
Transport by formation and destruction of organelles . So far, we have considered endo- and exocytosis as processes of transporting the contents of vesicles. There is another aspect of these processes, which lies in the fact that the targeted removal of the plasma membrane on one part of the cell surface by endocytosis and, on the contrary, its addition on another by exocytosis moves large areas of the membrane (Fig. 1.12.E), giving the cell the opportunity, for example , form an outgrowth or move.
Similar rearrangements are also typical for the cytoskeleton, especially for microfilaments and microtubules (Fig. 1.1). Microfilaments consist primarily of F-actin protein, which is capable of assembly into fibrous bundles as a result of polymerization of the monomer from the cytosol. The bundles are polarized, that is, they often grow only at one end, accumulating new actin molecules, while the other end is inert or disassembly occurs here. Due to this polarized growth, microfilaments move efficiently and the structure of their network can change. The transition of actin from a depolymerized state (sol) to an organized state (gel) can occur very quickly under the influence of other proteins or changes in ion concentration (see below). There are also proteins that cause actin filaments to break down to form short fragments. The thin projections of many cells - filopodia - contain a central bundle of actin (Fig. 1.1), and the various movements of the filopodia are probably due to actin transitions: polymerization - depolymerization.
Microtubulesalso often undergo similar movements. The mechanism of these movements is similar - polymerization of tubulin from the cytosol in such a way that one of the ends of the microtubule grows, while the other either does not change, or disassembly occurs there. Thus, a microtubule, by appropriate addition or removal of material, can move throughout the cytosol.
Active movements of the cytoskeleton . Changes in cytoskeletal structures can occur as a result of both active movements and rearrangements described above. In many cases, the movement of microtubules and actin filaments is driven by contractile proteins that bind the filaments or tubules and can move them relative to each other. Squirrels myosin and dynein present in the cytosol of all cells in relatively high concentrations; they are the elements that convert energy into movement in specialized cells (muscle) and organelles (cilia). In muscle cells, myosin forms thick filaments oriented parallel to actin filaments. The myosin molecule with its “head” attaches to the actin filament and, using the energy of ATP, displaces myosin along the actin molecule. Myosin then detaches from actin. The combination of many such connection-disconnection cycles leads to macroscopic contraction of muscle fibers(chapter 4). Dynein plays a similar role in the movement of microtubules during cilia (Fig. 1.1). In the cytoplasm of unspecialized cells, myosin and dynein do not form regular fibers, but in most cases small groups of molecules. Even in such small aggregates, they are capable of moving actin filaments or microtubules. Rice. Figure 1.13 illustrates this process when oppositely polarized myosin molecules are also attached to two actin filaments, polarized in different directions. The myosin headgroups bend toward the tail of the molecule, consuming ATP in the process, and the two actin filaments move in the opposite direction, after which the myosin detaches from them. Movements of this kind, during which ATP energy is converted into mechanical work, can change the shape of the cytoskeleton and, consequently, the cell, and also provide transport of organelles associated with the cytoskeleton.
The processes of intracellular transport can be most clearly demonstrated on the axon of a nerve cell. Axon transport is discussed here in detail to illustrate events that are likely to occur in similar ways in most cells. An axon that is only a few microns in diameter can reach a length of one meter or more, and the movement of proteins by diffusion from the nucleus to the distal end of the axon would take years. It has long been known that when any part of the axon undergoes constriction, the part of the axon located more proximally expands. It looks as if centrifugal flow is blocked in the axon. Such flow–fast axon transport can be demonstrated by the movement of radioactive markers, as in the experiment shown in Fig. 1.14. Radiolabeled leucine was injected into the dorsal root ganglion, and then radioactivity was measured in the sciatic nerve at a distance of 166 mm from the neuronal cell bodies from the 2nd to the 10th hour. Over 10 hours, the peak of radioactivity at the injection site changed slightly. But the wave of radioactivity spread along the axon at a constant speed of about 34 mm in 2 hours, or 410 mm/day. It has been shown that in all neurons of homeothermic animals, fast axonal transport occurs at the same speed, and no noticeable differences are observed between thin, unmyelinated fibers and the thickest axons, as well as between motor and sensory fibers. The type of radioactive marker also does not affect the rate of fast axonal transport; markers can serve as a variety of radioactive

Rice. 1.13.The non-muscle myosin complex, with a certain orientation, can bind to actin filaments of different polarity and, using the energy of ATP, displace them relative to each other
molecules, such as various amino acids, that are included in the proteins of the cell body of the neuron. If we analyze the peripheral part of the nerve to determine the nature of the carriers of radioactivity transported here, then such carriers are found mainly in the protein fraction, but also in the composition of mediators and free amino acids. Knowing that the properties of these substances are different and the sizes of their molecules are especially different, we can explain the constant speed of transport only by a transport mechanism common to all of them.
Described above fast axon transport is anterograde, i.e., directed away from the cell body. It has been shown that some substances move from the periphery to the cell body using retrograde transport. For example, acetylcholinesterase is transported in this direction at a speed 2 times slower than the speed of fast axonal transport. A marker often used in neuroanatomy, horseradish peroxidase, also moves by retrograde transport. Retrograde transport probably plays an important role in the regulation of protein synthesis in the cell body. A few days after the axon is cut, chromatolysis is observed in the cell body, which indicates a disruption of protein synthesis. The time required for chromatolysis correlates with the duration of retrograde transport from the site of axon transection to the cell body. This result also suggests an explanation for this disorder - the transmission from the periphery of the “signal substance” that regulates protein synthesis is disrupted. It is obvious that the main "vehicles" used for fast axonal

Rice. 1.14.Experiment demonstrating rapid axonal transport in sensory fibers of the cat sciatic nerve. Tritium-labeled leucine is injected into the dorsal root ganglion and radioactivity in the ganglion and sensory fibers is measured at 2, 4, 6, 8 and 10 hours after injection. (bottom of the picture). By x-axis The distance from the ganglion to the areas of the sciatic nerve where the measurement is made is plotted. On the ordinate axis only for the upper and lower curves, radioactivity (pulses/min) is plotted on a logarithmic scale. "Wave" of increased radioactivity (arrows) moves at a speed of 410 mm/day (along)
transport are vesicles (vesicles) and organelles, such as mitochondria, which contain substances that need to be transported. The movement of the largest vesicles or mitochondria can be observed using a microscope in vivo . Such particles make short, rapid movements in one direction, stop, often move slightly backward or to the side, stop again, and then jerk in the main direction. 410 mm/day corresponds to an average speed of anterograde movement of approximately 5 μm/s; the speed of each individual movement must, therefore, be much higher, and if we take into account the sizes of organelles, filaments and microtubules, then these movements are indeed very fast. Fast axonal transport requires a significant concentration of ATP. Poisons such as colchicine, which disrupts microtubules, also block fast axonal transport. It follows from this that in the transport process we are considering, vesicles and organelles move along microtubules and actin filaments; this movement is provided by small aggregates of dynein and myosin molecules, acting as shown in Fig. 1.13, using ATP energy.
Fast axon transport may also be involved in pathological processes. Some neurotropic viruses (for example, herpes or polio viruses) enter the axon at the periphery and move via retrograde transport to the neuron body, where they multiply and exert their toxic effects. Tetanus toxin, a protein produced by bacteria that enter the body when the skin is damaged, is captured by nerve endings and transported to the neuron body, where it causes characteristic muscle spasms. There are known cases of toxic effects on axon transport itself, for example, exposure to the industrial solvent acrylamide. In addition, it is believed that the pathogenesis of vitamin deficiency beriberi and alcoholic polyneuropathy involves a violation of fast axonal transport.
In addition to fast axonal transport in the cell, there is also quite intense slow axon transport. Tubulin moves along the axon at a speed of about 1 mm/day, and actin moves faster—up to 5 mm/day. Other proteins migrate with these cytoskeletal components; for example, the enzymes appear to be associated with actin or tubulin. The rates of movement of tubulin and actin are roughly consistent with the growth rates found for the mechanism described previously, where the molecules are incorporated into the active end of a microtubule or microfilament. Therefore, this mechanism may underlie slow axonal transport. The rate of slow axonal transport also approximately corresponds to the rate of axon growth, which apparently indicates restrictions imposed by the structure of the cytoskeleton on the second process.
Concluding this section, it should be emphasized that cells are by no means static structures, as they appear, for example, in electron microscopic photographs. Plasma membrane and especially organelles are in constant rapid movement and constant restructuring; that is why they are able to function. Further, these are not simple chambers in which chemical reactions take place, but highly organized conglomerates of membranes and fibers, in which reactions occur in an optimally organized sequence.
1.4. Regulation of cellular functions
The maintenance of the individual cell as a functional unit is primarily regulated by the nucleus; the study of such regulatory mechanisms is the subject of cell biology and biochemistry. At the same time, cells must modify their functions in accordance with environmental conditions and the needs of other cells of the body, i.e. they serve as objects of functional regulation. Below we will briefly consider how these regulatory influences act on the plasma membrane and how they reach intracellular organelles.
Regulatory effects on the cell membrane
Membrane potential . In many cases, the regulation of cellular functions is carried out by changing the membrane potential. Local changes in potential are possible when: 1) current from an adjacent area of the cell or generated by another cell flows through the membrane; 2) the extracellular concentration of ions changes (often [K + ] out ); 3) membrane ion channels open. Changes in membrane potential can affect the conformation of membrane proteins, causing, in particular, channels to open or close. As described above, the functioning of some membrane pumps depends on the membrane potential. Nerve cells are specialized to perceive changes in membrane potential as information that must be processed and transmitted (see Chapter 2).
Extracellular regulatory substances . The most important regulatory mechanism involving extracellular substances is their interaction with specific receptors on the plasma membrane or inside the cell. These substances include synaptic transmitters, which transmit information between nerve cells, local agents, and substances that circulate in the blood and reach all cells of the body, such as hormones and antigens. Synaptic transmitters are small molecules released from nerve endings at the synapse;
when they reach the plasma membrane of a neighboring, postsynaptic cell, they trigger electrical signals or other regulatory mechanisms. This issue is discussed in detail in Chap. 3.
Local chemical agents often secreted by specialized cells. They diffuse freely in the extracellular space, but their action is limited to a small group of cells due to the rapid destruction of these substances, either spontaneously or under the action of enzymes. One example of the release of such agents is the release histamine mast cells due to damage or immune response. Histamine causes relaxation of vascular smooth muscle cells, increases the permeability of the vascular endothelium and stimulates sensory nerve endings that mediate the sensation of itching. Other local chemical agents are released by many other cells. Typical local agents are prostaglandins, making up a group of approximately 20 fatty acid derivatives. They are released continuously from widely distributed cells, but act only locally, as they are quickly destroyed by membrane phospholipases. Various prostaglandins have a wide spectrum of action: they can trigger the contraction of smooth muscle cells, cause aggregation of blood platelets (platelets), or suppress the development of the corpus luteum in the ovaries.
Other local agents serve growth factors. The best known is nerve growth factor (NGF) for sympathetic neurons, which is necessary for the growth and survival of these neurons during development in vivo or in cell culture. Apparently, the target cells for this class of neurons secrete NGF and thereby ensure proper innervation. When forming organs, cells often need to “find their way” to target cells, which can be located at considerable distances. Accordingly, there must be many specialized growth factors similar to NGF.
Hormones and antigens carried by blood to all cells. Antigens trigger an immune response from cells bearing specific antibodies. However, antigens, as a rule, are foreign substances that are not produced in the reacting organism (for more details, see Chapter 18). Some hormones, such as insulin or thyroxine, affect a wide variety of cell types, while others, such as sex hormones, affect only a specific type of cell. Hormones are either peptides, whose action is triggered by binding to a receptor on the cell membrane, or steroids and thyroxine, which diffuse through the lipid membrane and bind to intracellular receptors. Steroid hormones bind to nuclear chromatin, resulting in the transcription of certain genes. The resulting proteins cause changes in cellular functions, which is the specific effect of hormones. Issues related to the release and action of hormones are discussed in detail in Chapter. 17.
Intracellular communication involving second messengers
The regulatory functions described above include effects on the cell membrane. Information received by the cell membrane must often cause organelles to react and is transferred to them by various substances known as second messengers (as opposed to first messengers, which come to the cell from external sources). The study of second mediators is evolving rapidly, and there is no guarantee that the current level of understanding of the problem will be sufficiently complete. Here we will touch on three well-studied mediators: Ca 2+ , cAMP and inositol triphosphate.
Calcium.The simplest intracellular mediator is the Ca 2+ ion. Its free concentration in a resting cell is very low and amounts to 10_–8–10–7 mol/l. It can enter the cell through specific membrane channels when they are in an open state, for example, when the membrane potential changes (see Chapter 2). The resulting increase in Ca 2+ concentration triggers important reactions in the cell, such as the contraction of myofibrils, which is the basis of muscle contraction (see Chapter 4), or the release of vesicles containing neurotransmitters from nerve endings (see Chapter 3) . Both reactions require a Ca 2+ concentration of approximately 10 –5 mol/L. Ca 2+, which has a regulatory effect, can also be released from intracellular stores, such as the endoplasmic reticulum. The release of Ca 2+ from the depot requires the participation of other intermediaries (see, for example, Fig. 1.16).
Cyclic adenosine monophosphate, cAMP. Recently, it has been proven that cyclic adenosine monophosphate (cAMP), a derivative of the body's main energy source, ATP, is an important second messenger. The complex chain of reactions shown in Fig. 1.15, starts with receptor R s on the outer surface of the plasma membrane, which can serve as a specific binding site for various mediators and hormones. After binding to a specific "stimulating" molecule R s changes its conformation; these changes affect the protein G s on the inner surface of the membrane in such a way that activation of the latter by intracellular guanosine triphosphate (GTP) becomes possible. Activated protein G s , in turn, stimulates the enzyme on the inner surface of the membrane, adenylate cyclase (AC), which catalyzes the formation of cAMP from ATP. Water-soluble cAMP is the mediator that transmits the effect

Rice. 1.15.A chain of reactions involving the intracellular messenger cAMP (cyclic adenosine monophosphate). Excitatory or inhibitory external signals activate membrane receptors R s or Ri . These receptors regulate the binding process G – proteins with intracellular GTP (guanosine triphosphate), thereby stimulating or inhibiting intracellular adenylate cyclase (AC). The amplifying enzyme AC converts adenosine triphosphate (ATP) into cAMP, which is then broken down into AMP by phosphodiesterase. Free cAMP diffuses into the cell and activates adenylate kinase (A-kinase), releasing its catalytic subunit C, which catalyzes the phosphorylation of intracellular proteins, i.e. forms the final effect of an extracellular stimulus. The diagram also shows pharmacological drugs and toxins that trigger (+) or inhibit (-) certain reactions (modified)
extracellular receptor stimulation R s to the internal structures of the cell.
In parallel with the stimulatory chain of reactions involving R s binding of inhibitory mediators and hormones to the corresponding receptor is possible R i which again through GTP-activated protein G , inhibits AC and thus the production of cAMP. Diffusing into the cell, cAMP reacts with adenylate kinase (A-kinase); this releases subunit C, which catalyzes the phosphorylation of protein P. This phosphorylation converts proteins into an active form, and they can now exert their specific regulatory effects (for example, cause glycogen degradation). This complex regulatory system is extremely efficient because the end result is the phosphorylation of many proteins, i.e., the regulatory signal passes through the circuit with a high gain. External mediators that bind to receptors R s and R i specific to each of them, extremely diverse. Adrenaline, connecting with R s or R i participates in the regulation of lipid and glycogen metabolism, as well as in enhancing the contraction of the heart muscle and in other reactions (see Chapter 19). Thyroid-stimulating hormone, activating R s , stimulates the release of the hormone thyroxine by the thyroid gland, and prostaglandin I inhibits the aggregation of blood platelets. Inhibitory effects, including adrenaline, mediated through R i are expressed in slowing down lipolysis. Thus, the cAMP system is a multifunctional intracellular regulatory system, which can be precisely controlled by extracellular stimulatory and inhibitory signaling substances.
Inositol phosphate "IF" h " The intracellular system of the second messenger, inositol phosphate, was discovered only recently (Fig. 1.16). In this case, there is no inhibitory pathway, but there is a similarity to the cAMP system, in which the effect of stimulation of the R receptor is transferred to the GTP-activated G protein on the inner surface of the membrane. At the next stage, the usual membrane lipid phosphatidylinositol (PI), having previously received two additional phosphate groups, is converted into PI-diphosphate (PIP 2), which is cleaved by activated phosphodiesterase (PDE) into inositol triphosphate(IFz) and lipid diacylglycerol(DAG). Inositol triphosphate is a water-soluble second messenger that diffuses into the cytosol. It acts primarily by releasing Ca 2+ from the endoplasmic reticulum. Ca 2+ in turn acts as a mediator, as described above; for example, it activates Ca 2+ -dependent phosphokinase, which phosphorylates enzymes. The lipid subunit of DAG (Fig. 1.16) also carries the signal, diffusing in the lipid phase of the plasma membrane to C-kinase located on its inner surface, which is activated with the participation of phosphatidylserine as a cofactor. Then C-kinase triggers phosphorylation of proteins, converting them into an active form.
The intracellular second messenger system IF 3 can also be controlled by a variety of external mediators and hormones, including acetylcholine, serotonin, vasopressin, and thyroid-stimulating hormone; like the cAMP system, it is characterized by a variety of intracellular effects. It is possible that this system is also activated by light in the visual receptor of the eye and plays a central role in phototransduction (see Chapter 11). For the first time in the individual development of the organism, the receptor of the IG system is activated by sperm, as a result of which IG takes part in the regulatory reactions accompanying the fertilization of the egg.
The cAMP and IFz-DAG systems are highly effective biological enhancers. They

Rice. 1.16.A chain of reactions involving the intracellular mediator IPG (inositol triphosphate). As in the cAMP system, the extracellular signal is mediated through a proteinG, which in this case activates phosphodiesterase (PDE). This enzyme breaks down phosphatidylinosine diphosphate (PIP 2 ) in the plasma membrane before IF h and diacylglycerol (DAG); IF h diffuses into the cytoplasm. Here it causes the release of Ca 2+ from the endoplasmic reticulum; increase in Ca concentration 2+ in the cytoplasm ([Ca 2+ ] i ) activates protein kinase, which phosphorylates and therefore activates enzymes. Another product, DAG, remains in the membrane and activates protein kinase C (phosphatidylserine cofactor, PS). Protein kinase C also phosphorylates enzymes that mediate specific actions associated with extrinsic receptor stimulation R . Branches of the chain of reactions involving IF h and DAG can be activated independently by ionomycin and phorbol ester, respectively (modified)
convert the reaction between a transmitter and an outer membrane receptor into phosphorylation of a variety of intracellular proteins, which can then influence various cell functions. One of the significant aspects of the problem is that, as far as is known today, there are only these two closely related regulatory systems of this type, used by numerous external mediators to regulate a variety of intracellular processes. At the same time, these regulatory systems, including Ca 2+, closely interact with each other, which allows them to carry out fine regulation of cellular functions.
Tutorials and Guides
1. Alberts IN., Bray D., Lewis J., Raff M., Roberts TO., Watson J.D.Molecular Biology of the Cell, New York and London, Garland Publishing Inc., 1983.
2. Czihak G., Longer H., Ziegler H.(eds.). Biology. Berlin, Heidelberg, New York, Springer, 1983.
3. Hille IN. Ionic channels of excitable membranes. Sunderland, Mass., Sinauer Assoc., 1984.
4. Hoppe W., Lohmann W.. Marki H., Ziegler H.(eds.). Biophysics. Berlin, Heidelberg, New York, Springer, 1984.
5. Jungermann TO., Mahler H.Biochemie. Berlin, Heidelberg, New York, Springer, 1980.
6. Kandel E. R., Schwartz– J. H.,(eds.). Principles of neural science, New York, Amsterdam, Oxford, Elsevier, 1985.
7. Schiebler T. H., Schmidt W.Anatomic des Menschen. Berlin, Heidelberg, New York, Tokyo, Springer, 1983.
Original articles and reviews
8. Berridge M. J. The molecular basis of communication within the cell, Sci. Amer, 253, 124-134 (1985).
9. Berridge M. J., Irvine R. F. Inositol triphosphate, a novel second messenger in cellular signal transduction. Nature, 312, 315 321 (1984).
10. Bretscher M.S. The molecules of the cell membrane, Sci. Amer., 253, 124–134 (1985).
11. Daut J. The living cell as an energy–transducing machine. A minimal model of myocardial metabolism, Biochem. et Biophys. Acta, 895, 41–62 (1987).
12. Hodgkin A.L., Katz IN. The effect of sodium ions on the electrical activity of the giant axon of the squid. J. Physiol. (Lond.), 108, 37–77 (1949).
13. Hodgkin A.L., Keynes R.D. Active transport of cations in giant axons from Sepia and Loligo, J. Physiol. (Lond.), 128, 28–42 (1955).
14. Longer P. Ionic channels with conformational substates, Biophys. J., 47, 581–590 (1985).
15. Ochs S., Worth P.M. Exoplasmic transport in normal and pathologic systems. In: Physiology and Pathology of Axons, S.G. Waxmann, Ed. New York, Raven Press, 1978.
Theory for task 5 from the Unified State Exam in biology
Cell structure. The relationship between the structure and functions of the parts and organelles of a cell is the basis of its integrity
Cell structure
Structure of prokaryotic and eukaryotic cells
The main structural components of cells are the plasma membrane, cytoplasm and hereditary apparatus. Depending on the characteristics of the organization, two main types of cells are distinguished: prokaryotic and eukaryotic. The main difference between prokaryotic cells and eukaryotic cells is the organization of their hereditary apparatus: in prokaryotes it is located directly in the cytoplasm (this area of the cytoplasm is called nucleoid) and is not separated from it by membrane structures, whereas in eukaryotes most of the DNA is concentrated in the nucleus, surrounded by a double membrane. In addition, the genetic information of prokaryotic cells, located in the nucleoid, is written in a circular DNA molecule, while in eukaryotes the DNA molecules are open.
Unlike eukaryotes, the cytoplasm of prokaryotic cells also contains a small number of organelles, while eukaryotic cells are characterized by a significant variety of these structures.
Structure and functions of biological membranes
The structure of the biomembrane. The cell-bounding membranes and membrane organelles of eukaryotic cells have a common chemical composition and structure. They include lipids, proteins and carbohydrates. Membrane lipids are mainly represented by phospholipids and cholesterol. Most membrane proteins are complex proteins, such as glycoproteins. Carbohydrates do not occur independently in the membrane; they are associated with proteins and lipids. The thickness of the membranes is 7-10 nm.
According to the currently generally accepted fluid mosaic model of membrane structure, lipids form a double layer, or lipid bilayer, in which the hydrophilic “heads” of lipid molecules face outward, and the hydrophobic “tails” are hidden inside the membrane. These “tails,” due to their hydrophobicity, ensure the separation of the aqueous phases of the internal environment of the cell and its environment. Proteins are associated with lipids through various types of interactions. Some proteins are located on the surface of the membrane. Such proteins are called peripheral, or superficial. Other proteins are partially or completely immersed in the membrane - these are integral, or submerged proteins. Membrane proteins perform structural, transport, catalytic, receptor and other functions.
Membranes are not like crystals; their components are constantly in motion, as a result of which gaps appear between lipid molecules - pores through which various substances can enter or leave the cell.
Biological membranes differ in their location in the cell, chemical composition and functions. The main types of membranes are plasma and internal. Plasma membrane contains about 45% lipids (including glycolipids), 50% proteins and 5% carbohydrates. Chains of carbohydrates, which are part of complex proteins-glycoproteins and complex lipids-glycolipids, protrude above the surface of the membrane. Plasmalemma glycoproteins are extremely specific. For example, they are used for mutual recognition of cells, including sperm and egg.
On the surface of animal cells, carbohydrate chains form a thin surface layer - glycocalyx. It is detected in almost all animal cells, but its degree of expression varies (10-50 µm). The glycocalyx provides direct communication between the cell and the external environment, where extracellular digestion occurs; Receptors are located in the glycocalyx. In addition to the plasmalemma, the cells of bacteria, plants and fungi are also surrounded by cell membranes.
Internal membranes eukaryotic cells delimit different parts of the cell, forming peculiar “compartments” - compartments, which promotes the separation of various metabolic and energy processes. They may differ in chemical composition and functions, but their general structural plan remains the same.
Membrane functions:
- Limiting. The idea is that they separate the internal space of the cell from the external environment. The membrane is semi-permeable, that is, only those substances that the cell needs can freely pass through it, and there are mechanisms for transporting the necessary substances.
- Receptor. It is primarily associated with the perception of environmental signals and the transfer of this information into the cell. Special receptor proteins are responsible for this function. Membrane proteins are also responsible for cellular recognition according to the “friend or foe” principle, as well as for the formation of intercellular connections, the most studied of which are the synapses of nerve cells.
- Catalytic. Numerous enzyme complexes are located on the membranes, as a result of which intensive synthetic processes occur on them.
- Energy transforming. Associated with the formation of energy, its storage in the form of ATP and consumption.
- Compartmentalization. Membranes also delimit the space inside the cell, thereby separating the starting materials of the reaction and the enzymes that can carry out the corresponding reactions.
- Formation of intercellular contacts. Despite the fact that the thickness of the membrane is so small that it cannot be distinguished with the naked eye, it, on the one hand, serves as a fairly reliable barrier for ions and molecules, especially water-soluble ones, and on the other, ensures their transport into and out of the cell.
- Transport.

Membrane transport. Due to the fact that cells, as elementary biological systems, are open systems, to ensure metabolism and energy, maintain homeostasis, growth, irritability and other processes, the transfer of substances through the membrane - membrane transport - is required. Currently, the transport of substances across the cell membrane is divided into active, passive, endo- and exocytosis.
Passive transport- This is a type of transport that occurs without energy consumption from higher to lower concentrations. Lipid-soluble small non-polar molecules (O 2, CO 2) easily penetrate the cell by simple diffusion. Those insoluble in lipids, including charged small particles, are picked up by carrier proteins or pass through special channels (glucose, amino acids, K +, PO 4 3-). This type of passive transport is called facilitated diffusion. Water enters the cell through pores in the lipid phase, as well as through special channels lined with proteins. The transport of water through a membrane is called by osmosis.

Osmosis is extremely important in the life of a cell, because if it is placed in a solution with a higher concentration of salts than in the cell solution, then water will begin to leave the cell and the volume of living contents will begin to decrease. In animal cells, the cell as a whole shrinks, and in plant cells, the cytoplasm lags behind the cell wall, which is called plasmolysis. When a cell is placed in a solution less concentrated than the cytoplasm, water transport occurs in the opposite direction - into the cell. However, there are limits to the extensibility of the cytoplasmic membrane, and an animal cell eventually ruptures, while a plant cell does not allow this to happen due to its strong cell wall. The phenomenon of filling the entire internal space of a cell with cellular contents is called deplasmolysis. The intracellular concentration of salts should be taken into account when preparing medications, especially for intravenous administration, as this can lead to damage to blood cells (for this, saline solution with a concentration of 0.9% sodium chloride is used). This is no less important when cultivating cells and tissues, as well as animal and plant organs.

Active transport proceeds with the expenditure of ATP energy from a lower concentration of a substance to a higher one. It is carried out using special pump proteins. Proteins pump K + , Na + , Ca 2+ and other ions through the membrane, which promotes the transport of essential organic substances, as well as the emergence of nerve impulses, etc.
Endocytosis- this is an active process of absorption of substances by the cell, in which the membrane forms invaginations and then forms membrane vesicles - phagosomes, which contain the absorbed objects. Then the primary lysosome fuses with the phagosome and forms secondary lysosome, or phagolysosome, or digestive vacuole. The contents of the vesicle are digested by lysosome enzymes, and the breakdown products are absorbed and assimilated by the cell. Undigested residues are removed from the cell by exocytosis. There are two main types of endocytosis: phagocytosis and pinocytosis.
Phagocytosis is the process of capture by the cell surface and absorption of solid particles by the cell, and pinocytosis- liquids. Phagocytosis occurs mainly in animal cells (single-celled animals, human leukocytes), it provides their nutrition and often protects the body. By pinocytosis, proteins, antigen-antibody complexes are absorbed during immune reactions, etc. However, many viruses also enter the cell by pinocytosis or phagocytosis. In plant and fungal cells, phagocytosis is practically impossible, since they are surrounded by durable cell membranes.

Exocytosis- a process reverse to endocytosis. In this way, undigested food remains are released from the digestive vacuoles, and substances necessary for the life of the cell and the body as a whole are removed. For example, the transmission of nerve impulses occurs due to the release of chemical messengers by the neuron sending the impulse - mediators, and in plant cells this is how auxiliary carbohydrates of the cell membrane are secreted.
Cell walls of plant cells, fungi and bacteria. Outside the membrane, the cell can secrete a strong framework - cell membrane, or cell wall.
In plants, the basis of the cell wall is cellulose, packed in bundles of 50-100 molecules. The spaces between them are filled with water and other carbohydrates. The plant cell wall is permeated with tubules - plasmodesmata, through which the membranes of the endoplasmic reticulum pass. Plasmodesmata carry out the transport of substances between cells. However, transport of substances, such as water, can also occur along the cell walls themselves. Over time, various substances, including tannins or fat-like substances, accumulate in the cell wall of plants, which leads to lignification or suberization of the cell wall itself, displacement of water and death of cellular contents. Between the cell walls of neighboring plant cells there are jelly-like spacers - middle plates that hold them together and cement the plant body as a whole. They are destroyed only during the process of fruit ripening and when the leaves fall.

The cell walls of fungal cells are formed chitin- a carbohydrate containing nitrogen. They are quite strong and are the external skeleton of the cell, but still, like in plants, they prevent phagocytosis.
In bacteria, the cell wall contains carbohydrates with peptide fragments - murein, however, its content varies significantly among different groups of bacteria. Other polysaccharides can also be secreted on top of the cell wall, forming a mucous capsule that protects bacteria from external influences.
The membrane determines the shape of the cell, serves as a mechanical support, performs a protective function, provides the osmotic properties of the cell, limiting the stretching of the living contents and preventing rupture of the cell, which increases due to the entry of water. In addition, water and substances dissolved in it overcome the cell wall before entering the cytoplasm or, conversely, when leaving it, while water is transported through the cell walls faster than through the cytoplasm.
Cytoplasm
Cytoplasm- This is the internal contents of the cell. All cell organelles, the nucleus and various waste products are immersed in it.
The cytoplasm connects all parts of the cell to each other, and numerous metabolic reactions take place in it. The cytoplasm is separated from the environment and divided into compartments by membranes, that is, cells have a membrane structure. It can be in two states - sol and gel. Sol- this is a semi-liquid, jelly-like state of the cytoplasm, in which vital processes proceed most intensively, and gel- a denser, gelatinous state that impedes the occurrence of chemical reactions and the transport of substances.
The liquid part of the cytoplasm without organelles is called hyaloplasm. Hyaloplasm, or cytosol, is a colloidal solution in which there is a kind of suspension of fairly large particles, for example proteins, surrounded by dipoles of water molecules. Precipitation of this suspension does not occur due to the fact that they have the same charge and repel each other.
Organoids
Organoids- These are permanent components of the cell that perform specific functions.
Depending on their structural features, they are divided into membrane and non-membrane. Membrane organelles, in turn, are classified as single-membrane (endoplasmic reticulum, Golgi complex and lysosomes) or double-membrane (mitochondria, plastids and nucleus). Non-membrane The organelles are ribosomes, microtubules, microfilaments and the cell center. Of the listed organelles, only ribosomes are inherent in prokaryotes.
Structure and functions of the nucleus. Core- a large double-membrane organelle lying in the center of the cell or at its periphery. The dimensions of the nucleus can range from 3-35 microns. The shape of the nucleus is most often spherical or ellipsoidal, but there are also rod-shaped, fusiform, bean-shaped, lobed and even segmented nuclei. Some researchers believe that the shape of the nucleus corresponds to the shape of the cell itself.
Most cells have one nucleus, but, for example, in the cells of the liver and heart there can be two of them, and in a number of neurons - up to 15. Skeletal muscle fibers usually contain many nuclei, but they are not cells in the full sense of the word, since they are formed in the result of the fusion of several cells.
The core is surrounded nuclear envelope, and its internal space is filled nuclear juice, or nucleoplasm (karyoplasm), in which they are immersed chromatin And nucleolus. The nucleus performs such important functions as storing and transmitting hereditary information, as well as controlling the life of the cell.

The role of the nucleus in the transmission of hereditary information was convincingly proven in experiments with the green alga Acetabularia. In a single giant cell, reaching a length of 5 cm, a cap, a stalk and a rhizoid are distinguished. Moreover, it contains only one nucleus located in the rhizoid. In the 1930s, I. Hemmerling transplanted the nucleus of one species of acetabularia with a green color into the rhizoid of another species, with a brown color, from which the nucleus had been removed. After some time, the plant with the transplanted nucleus grew a new cap, like the nucleus donor algae. At the same time, the cap or stalk, separated from the rhizoid and not containing a nucleus, died after some time.

Nuclear envelope formed by two membranes - outer and inner, between which there is space. The intermembrane space communicates with the cavity of the rough endoplasmic reticulum, and the outer membrane of the nucleus can carry ribosomes. The nuclear envelope is permeated with numerous pores lined with special proteins. Transport of substances occurs through the pores: the necessary proteins (including enzymes), ions, nucleotides and other substances enter the nucleus, and RNA molecules, spent proteins, and subunits of ribosomes leave it. Thus, the functions of the nuclear envelope are the separation of the contents of the nucleus from the cytoplasm, as well as the regulation of metabolism between the nucleus and the cytoplasm.
Nucleoplasm called the contents of the nucleus, in which chromatin and the nucleolus are immersed. It is a colloidal solution, chemically reminiscent of cytoplasm. Enzymes of the nucleoplasm catalyze the exchange of amino acids, nucleotides, proteins, etc. The nucleoplasm is connected to the hyaloplasm through nuclear pores. The functions of the nucleoplasm, like the hyaloplasm, are to ensure the interconnection of all structural components of the nucleus and to carry out a number of enzymatic reactions.
Chromatin called a collection of thin filaments and granules immersed in the nucleoplasm. It can only be detected by staining, since the refractive indices of chromatin and nucleoplasm are approximately the same. The filamentous component of chromatin is called euchromatin, and granular - heterochromatin. Euchromatin is weakly compacted, since hereditary information is read from it, while more spiralized heterochromatin is genetically inactive.
Chromatin is a structural modification of chromosomes in a non-dividing nucleus. Thus, chromosomes are constantly present in the nucleus; only their state changes depending on the function that the nucleus performs at the moment.
The composition of chromatin mainly includes nucleoprotein proteins (deoxyribonucleoproteins and ribonucleoproteins), as well as enzymes, the most important of which are associated with the synthesis of nucleic acids, and some other substances.
The functions of chromatin consist, firstly, in the synthesis of nucleic acids specific to a given organism, which direct the synthesis of specific proteins, and secondly, in the transfer of hereditary properties from the mother cell to the daughter cells, for which purpose the chromatin threads are packaged into chromosomes during the division process.
Nucleolus- a spherical body, clearly visible under a microscope, with a diameter of 1-3 microns. It is formed on sections of chromatin in which information about the structure of rRNA and ribosomal proteins is encoded. There is often only one nucleolus in the nucleus, but in those cells where intensive vital processes occur, there may be two or more nucleoli. The functions of the nucleoli are the synthesis of rRNA and the assembly of ribosomal subunits by combining rRNA with proteins coming from the cytoplasm.
Mitochondria- double-membrane organelles of round, oval or rod-shaped form, although spiral-shaped ones are also found (in sperm). The diameter of mitochondria is up to 1 µm, and the length is up to 7 µm. The space inside the mitochondria is filled with matrix. Matrix- This is the main substance of mitochondria. A circular DNA molecule and ribosomes are immersed in it. The outer membrane of mitochondria is smooth and impermeable to many substances. The inner membrane has projections - cristas, increasing the surface area of membranes for chemical reactions to occur. On the surface of the membrane there are numerous protein complexes that make up the so-called respiratory chain, as well as mushroom-shaped ATP synthetase enzymes. The aerobic stage of respiration occurs in mitochondria, during which ATP is synthesized.

Plastids- large double-membrane organelles, characteristic only of plant cells. The internal space of the plastids is filled stroma, or matrix. The stroma contains a more or less developed system of membrane vesicles - thylakoids, which are collected in piles - grains, as well as its own circular DNA molecule and ribosomes. There are four main types of plastids: chloroplasts, chromoplasts, leucoplasts and proplastids.
Chloroplasts- these are green plastids with a diameter of 3-10 microns, clearly visible under a microscope. They are found only in the green parts of plants - leaves, young stems, flowers and fruits. Chloroplasts are generally oval or ellipsoidal in shape, but can also be cup-shaped, spiral-shaped, or even lobed. The number of chloroplasts in a cell averages from 10 to 100 pieces. However, for example, in some algae it may be one, have significant dimensions and a complex shape - then it is called chromatophore. In other cases, the number of chloroplasts can reach several hundred, while their sizes are small. The color of chloroplasts is due to the main pigment of photosynthesis - chlorophyll, although they also contain additional pigments - carotenoids. Carotenoids only become noticeable in the fall, when the chlorophyll in aging leaves breaks down. The main function of chloroplasts is photosynthesis. Light reactions of photosynthesis occur on thylakoid membranes, on which chlorophyll molecules are attached, and dark reactions take place in the stroma, where numerous enzymes are contained.

Chromoplasts- These are yellow, orange and red plastids containing carotenoid pigments. The shape of chromoplasts can also vary significantly: they can be tubular, spherical, crystalline, etc. Chromoplasts give color to the flowers and fruits of plants, attracting pollinators and distributors of seeds and fruits.
Leukoplasts- These are white or colorless plastids, mostly round or oval in shape. They are common in non-photosynthetic parts of plants, for example in the skin of leaves, potato tubers, etc. They store nutrients, most often starch, but in some plants it can be proteins or oil.
Plastids are formed in plant cells from proplastids, which are already present in the cells of educational tissue and are small double-membrane bodies. At the early stages of development, different types of plastids are capable of transforming into each other: when exposed to light, the leucoplasts of a potato tuber and the chromoplasts of a carrot root turn green.
Plastids and mitochondria are called semi-autonomous organelles of the cell, since they have their own DNA molecules and ribosomes, carry out protein synthesis and divide independently of cell division. These features are explained by their origin from single-celled prokaryotic organisms. However, the “independence” of mitochondria and plastids is limited, since their DNA contains too few genes for free existence, while the rest of the information is encoded in the chromosomes of the nucleus, which allows it to control these organelles.
Endoplasmic reticulum (ER), or endoplasmic reticulum (ER), is a single-membrane organelle, which is a network of membrane cavities and tubules occupying up to 30% of the contents of the cytoplasm. The diameter of the EPS tubules is about 25-30 nm. There are two types of EPS - rough and smooth. Rough XPS carries ribosomes, where protein synthesis occurs. Smooth XPS lacks ribosomes. Its function is the synthesis of lipids and carbohydrates, as well as the transport, storage and neutralization of toxic substances. It is especially developed in those cells where intensive metabolic processes occur, for example in liver cells - hepatocytes - and skeletal muscle fibers. Substances synthesized in the ER are transported to the Golgi apparatus. The assembly of cell membranes also occurs in the ER, but their formation is completed in the Golgi apparatus.
Golgi apparatus, or Golgi complex, is a single-membrane organelle formed by a system of flat cisterns, tubules and vesicles detached from them. The structural unit of the Golgi apparatus is dictyosome- a stack of tanks, at one pole of which substances from the EPS come, and from the opposite pole, having undergone certain transformations, they are packed into vesicles and sent to other parts of the cell. The diameter of the tanks is about 2 microns, and the diameter of small bubbles is about 20-30 microns. The main functions of the Golgi complex are the synthesis of certain substances and modification (change) of proteins, lipids and carbohydrates coming from the ER, the final formation of membranes, as well as the transport of substances throughout the cell, renewal of its structures and the formation of lysosomes. The Golgi apparatus received its name in honor of the Italian scientist Camillo Golgi, who first discovered this organelle (1898).

Lysosomes- small single-membrane organelles up to 1 μm in diameter, which contain hydrolytic enzymes involved in intracellular digestion. The membranes of lysosomes are poorly permeable to these enzymes, so the lysosomes perform their functions very accurately and targetedly. Thus, they take an active part in the process of phagocytosis, forming digestive vacuoles, and in case of starvation or damage to certain parts of the cell, they digest them without affecting others. The role of lysosomes in cell death processes has recently been discovered.
Vacuole is a cavity in the cytoplasm of plant and animal cells, bounded by a membrane and filled with liquid. Digestive and contractile vacuoles are found in protozoan cells. The former take part in the process of phagocytosis, as they break down nutrients. The latter ensure the maintenance of water-salt balance due to osmoregulation. In multicellular animals, digestive vacuoles are mainly found.
In plant cells, vacuoles are always present; they are surrounded by a special membrane and filled with cell sap. The membrane surrounding the vacuole is similar in chemical composition, structure and functions to the plasma membrane. Cell sap is an aqueous solution of various inorganic and organic substances, including mineral salts, organic acids, carbohydrates, proteins, glycosides, alkaloids, etc. The vacuole can occupy up to 90% of the cell volume and push the nucleus to the periphery. This part of the cell performs storage, excretory, osmotic, protective, lysosomal and other functions, since it accumulates nutrients and waste products, ensures the supply of water and maintains the shape and volume of the cell, and also contains enzymes for the breakdown of many cell components. In addition, the biologically active substances of the vacuoles can prevent many animals from eating these plants. In a number of plants, due to the swelling of vacuoles, cell growth occurs by elongation.
Vacuoles are also present in the cells of some fungi and bacteria, but in fungi they perform only the function of osmoregulation, while in cyanobacteria they maintain buoyancy and participate in the process of assimilation of nitrogen from the air.
Ribosomes- small non-membrane organelles with a diameter of 15-20 microns, consisting of two subunits - large and small. Eukaryotic ribosomal subunits are assembled in the nucleolus and then transported into the cytoplasm. Ribosomes in prokaryotes, mitochondria, and plastids are smaller in size than ribosomes in eukaryotes. Ribosomal subunits include rRNA and proteins.

The number of ribosomes in a cell can reach several tens of millions: in the cytoplasm, mitochondria and plastids they are in a free state, and on the rough ER - in a bound state. They take part in protein synthesis, in particular, they carry out the process of translation - the biosynthesis of a polypeptide chain on an mRNA molecule. Free ribosomes synthesize the proteins of hyaloplasm, mitochondria, plastids, and their own ribosomal proteins, while ribosomes attached to the rough ER carry out the translation of proteins for removal from cells, membrane assembly, and the formation of lysosomes and vacuoles.
Ribosomes can be found singly in the hyaloplasm or assembled in groups during the simultaneous synthesis of several polypeptide chains on one mRNA. Such groups of ribosomes are called polyribosomes, or polysomes.

Microtubules- These are cylindrical hollow non-membrane organelles that penetrate the entire cytoplasm of the cell. Their diameter is about 25 nm, wall thickness is 6-8 nm. They are formed by numerous protein molecules tubulin, which first form 13 threads resembling beads and then assemble into a microtubule. Microtubules form a cytoplasmic reticulum, which gives the cell shape and volume, connects the plasma membrane with other parts of the cell, ensures the transport of substances throughout the cell, takes part in the movement of the cell and intracellular components, as well as in the division of genetic material. They are part of the cell center and organelles of movement - flagella and cilia.
Microfilaments, or microthreads, are also non-membrane organelles, however, they have a filamentous shape and are formed not by tubulin, but actin. They take part in the processes of membrane transport, intercellular recognition, division of the cell cytoplasm and in its movement. In muscle cells, the interaction of actin microfilaments with myosin filaments mediates contraction.
Microtubules and microfilaments form the internal skeleton of the cell - cytoskeleton. It is a complex network of fibers that provide mechanical support for the plasma membrane, determines the shape of the cell, the location of cellular organelles and their movement during cell division.

Cell center- a non-membrane organelle located in animal cells near the nucleus; it is absent in plant cells. Its length is about 0.2-0.3 microns, and its diameter is 0.1-0.15 microns. The cell center is formed by two centrioles, lying in mutually perpendicular planes, and radiant sphere from microtubules. Each centriole is formed by nine groups of microtubules, collected in groups of three, i.e., triplets. The cellular center takes part in the processes of microtubule assembly, division of the cell's hereditary material, as well as in the formation of flagella and cilia.

Organelles of movement. Flagella And cilia They are cell outgrowths covered with plasmalemma. The basis of these organelles is made up of nine pairs of microtubules located along the periphery and two free microtubules in the center. Microtubules are interconnected by various proteins, ensuring their coordinated deviation from the axis - oscillation. Oscillations are energy-dependent, that is, the energy of high-energy ATP bonds is spent on this process. Restoration of lost flagella and cilia is a function basal bodies, or kinetosomes located at their base.
The length of cilia is about 10-15 nm, and the length of flagella is 20-50 µm. Due to strictly directed movements of flagella and cilia, not only the movement of single-celled animals, sperm, etc. occurs, but also the cleaning of the respiratory tract and the movement of the egg through the fallopian tubes, since all these parts of the human body are lined with ciliated epithelium.

Inclusions
Inclusions- These are non-permanent components of the cell that are formed and disappear during its life. These include both reserve substances, for example, grains of starch or protein in plant cells, glycogen granules in the cells of animals and fungi, volutin in bacteria, drops of fat in all types of cells, and waste products, in particular, food residues undigested as a result of phagocytosis , forming so-called residual bodies.
The relationship between the structure and functions of the parts and organelles of a cell is the basis of its integrity
Each of the parts of the cell, on the one hand, is a separate structure with a specific structure and functions, and on the other, a component of a more complex system called a cell. Most of the hereditary information of a eukaryotic cell is concentrated in the nucleus, but the nucleus itself is not able to ensure its implementation, since this requires at least the cytoplasm, which acts as the main substance, and ribosomes, on which this synthesis occurs. Most ribosomes are located on the granular endoplasmic reticulum, from where proteins are most often transported to the Golgi complex, and then, after modification, to those parts of the cell for which they are intended, or are excreted. Membrane packaging of proteins and carbohydrates can be embedded in the membranes of organelles and the cytoplasmic membrane, ensuring their constant renewal. Lysosomes and vacuoles, which perform important functions, also detach from the Golgi complex. For example, without lysosomes, cells would quickly turn into a kind of dumping ground for waste molecules and structures.
The occurrence of all these processes requires energy produced by mitochondria, and in plants, by chloroplasts. And although these organelles are relatively autonomous, since they have their own DNA molecules, some of their proteins are still encoded by the nuclear genome and synthesized in the cytoplasm.
Thus, the cell is an inextricable unity of its constituent components, each of which performs its own unique function.
Metabolism and energy conversion are properties of living organisms. Energy and plastic metabolism, their relationship. Stages of energy metabolism. Fermentation and respiration. Photosynthesis, its significance, cosmic role. Phases of photosynthesis. Light and dark reactions of photosynthesis, their relationship. Chemosynthesis. The role of chemosynthetic bacteria on Earth
Metabolism and energy conversion - properties of living organisms
A cell can be likened to a miniature chemical factory in which hundreds and thousands of chemical reactions occur.
Metabolism- a set of chemical transformations aimed at the preservation and self-reproduction of biological systems.
It includes the intake of substances into the body during nutrition and respiration, intracellular metabolism, or metabolism, as well as the isolation of final metabolic products.
Metabolism is inextricably linked with the processes of converting one type of energy into another. For example, during the process of photosynthesis, light energy is stored in the form of the energy of chemical bonds of complex organic molecules, and during the process of respiration it is released and spent on the synthesis of new molecules, mechanical and osmotic work, dissipated in the form of heat, etc.
The occurrence of chemical reactions in living organisms is ensured thanks to biological catalysts of a protein nature - enzymes, or enzymes. Like other catalysts, enzymes accelerate the occurrence of chemical reactions in a cell by tens and hundreds of thousands of times, and sometimes even make them possible, but do not change the nature or properties of the final product(s) of the reaction and do not change themselves. Enzymes can be both simple and complex proteins, which, in addition to the protein part, also include a non-protein part - cofactor (coenzyme). Examples of enzymes are salivary amylase, which breaks down polysaccharides during prolonged chewing, and pepsin, which ensures the digestion of proteins in the stomach.
Enzymes differ from non-protein catalysts in their high specificity of action, a significant increase in the reaction rate with their help, as well as the ability to regulate the action by changing the conditions of the reaction or the interaction of various substances with them. In addition, the conditions under which enzymatic catalysis occurs differ significantly from those under which non-enzymatic catalysis occurs: the optimal temperature for the functioning of enzymes in the human body is $37°C$, the pressure should be close to atmospheric, and the $pH$ of the environment can significantly hesitate. Thus, amylase requires an alkaline environment, and pepsin requires an acidic environment.
The mechanism of action of enzymes is to reduce the activation energy of substances (substrates) that enter into a reaction due to the formation of intermediate enzyme-substrate complexes.

Energy and plastic metabolism, their relationship
Metabolism consists of two processes occurring simultaneously in the cell: plastic and energy metabolism.
Plastic metabolism (anabolism, assimilation) is a set of synthesis reactions that involve the expenditure of ATP energy. In the process of plastic metabolism, organic substances necessary for the cell are synthesized. Examples of plastic exchange reactions are photosynthesis, protein biosynthesis, and DNA replication (self-duplication).
Energy metabolism (catabolism, dissimilation) is a set of reactions that break down complex substances into simpler ones. As a result of energy metabolism, energy is released and stored in the form of ATP. The most important processes of energy metabolism are respiration and fermentation.
Plastic and energy exchanges are inextricably linked, since in the process of plastic exchange organic substances are synthesized and this requires ATP energy, and in the process of energy exchange organic substances are broken down and energy is released, which will then be spent on synthesis processes.
Organisms receive energy during the process of nutrition, and release it and convert it into an accessible form mainly during the process of respiration. According to the method of nutrition, all organisms are divided into autotrophs and heterotrophs. Autotrophs capable of independently synthesizing organic substances from inorganic ones, and heterotrophs use exclusively prepared organic substances.
Stages of energy metabolism
Despite the complexity of energy metabolism reactions, it is conventionally divided into three stages: preparatory, anaerobic (oxygen-free) and aerobic (oxygen).
On preparatory stage molecules of polysaccharides, lipids, proteins, nucleic acids break down into simpler ones, for example, glucose, glycerol and fatty acids, amino acids, nucleotides, etc. This stage can occur directly in the cells or in the intestines, from where the broken down substances are delivered through the bloodstream.
Anaerobic stage energy metabolism is accompanied by further breakdown of monomers of organic compounds into even simpler intermediate products, for example, pyruvic acid, or pyruvate. It does not require the presence of oxygen, and for many organisms living in the mud of swamps or in the human intestines, it is the only way to obtain energy. The anaerobic stage of energy metabolism occurs in the cytoplasm.
Various substances can undergo oxygen-free cleavage, but quite often the substrate of the reactions is glucose. The process of its oxygen-free splitting is called glycolysis. During glycolysis, a glucose molecule loses four hydrogen atoms, i.e., it is oxidized, and two molecules of pyruvic acid, two molecules of ATP and two molecules of the reduced hydrogen carrier $NADH + H^(+)$ are formed:
$C_6H_(12)O_6 + 2H_3PO_4 + 2ADP + 2NAD → 2C_3H_4O_3 + 2ATP + 2NADH + H^(+) + 2H_2O$.
The formation of ATP from ADP occurs due to the direct transfer of phosphate anion from pre-phosphorylated sugar and is called substrate phosphorylation.
Aerobic stage energy exchange can occur only in the presence of oxygen, while intermediate compounds formed during oxygen-free cleavage are oxidized to the final products (carbon dioxide and water) and most of the energy stored in the chemical bonds of organic compounds is released. It turns into the energy of high-energy bonds of 36 ATP molecules. This stage is also called tissue respiration. In the absence of oxygen, intermediate compounds are converted into other organic substances, a process called fermentation.
Breath
The mechanism of cellular respiration is schematically depicted in Fig.

Aerobic respiration occurs in mitochondria, with pyruvic acid first losing one carbon atom, which is accompanied by the synthesis of one reducing equivalent of $NADH + H^(+)$ and a molecule of acetyl coenzyme A (acetyl-CoA):
$C_3H_4O_3 + NAD + H~CoA → CH_3CO~CoA + NADH + H^(+) + CO_2$.
Acetyl-CoA in the mitochondrial matrix is involved in a chain of chemical reactions, the totality of which is called Krebs cycle (tricarboxylic acid cycle, citric acid cycle). During these transformations, two ATP molecules are formed, acetyl-CoA is completely oxidized to carbon dioxide, and its hydrogen ions and electrons are added to the hydrogen carriers $NADH + H^(+)$ and $FADH_2$. The carriers transport hydrogen protons and electrons to the inner membranes of mitochondria, forming cristae. With the help of carrier proteins, hydrogen protons are pumped into the intermembrane space, and electrons are transmitted through the so-called respiratory chain of enzymes located on the inner membrane of mitochondria and discharged onto oxygen atoms:
$O_2+2e^(-)→O_2^-$.
It should be noted that some respiratory chain proteins contain iron and sulfur.
From the intermembrane space, hydrogen protons are transported back into the mitochondrial matrix with the help of special enzymes - ATP synthases, and the energy released in this case is spent on the synthesis of 34 ATP molecules from each glucose molecule. This process is called oxidative phosphorylation. In the mitochondrial matrix, hydrogen protons react with oxygen radicals to form water:
$4H^(+)+O_2^-→2H_2O$.
The set of reactions of oxygen respiration can be expressed as follows:
$2C_3H_4O_3 + 6O_2 + 36H_3PO_4 + 36ADP → 6CO_2 + 38H_2O + 36ATP.$
The overall breathing equation looks like this:
$C_6H_(12)O_6 + 6O_2 + 38H_3PO_4 + 38ADP → 6CO_2 + 40H_2O + 38ATP.$
Fermentation
In the absence of oxygen or its deficiency, fermentation occurs. Fermentation is an evolutionarily earlier method of obtaining energy than respiration, but it is energetically less beneficial because fermentation produces organic substances that are still rich in energy. There are several main types of fermentation: lactic acid, alcoholic, acetic acid, etc. Thus, in skeletal muscles in the absence of oxygen during fermentation, pyruvic acid is reduced to lactic acid, while the previously formed reducing equivalents are consumed, and only two ATP molecules remain:
$2C_3H_4O_3 + 2NADH + H^(+) → 2C_3H_6O_3 + 2NAD$.
During fermentation with the help of yeast, pyruvic acid in the presence of oxygen is converted into ethyl alcohol and carbon monoxide (IV):
$C_3H_4O_3 + NADH + H^(+) → C_2H_5OH + CO_2 + NAD^(+)$.
During fermentation with the help of microorganisms, acetic, butyric, formic acids, etc. can also be formed from pyruvic acid.
ATP, obtained as a result of energy metabolism, is spent in the cell for various types of work: chemical, osmotic, electrical, mechanical and regulatory. Chemical work involves the biosynthesis of proteins, lipids, carbohydrates, nucleic acids and other vital compounds. Osmotic work includes the processes of absorption by the cell and removal from it of substances that are in the extracellular space in concentrations greater than in the cell itself. Electrical work is closely interrelated with osmotic work, since it is as a result of the movement of charged particles through membranes that a membrane charge is formed and the properties of excitability and conductivity are acquired. Mechanical work involves the movement of substances and structures inside the cell, as well as the cell as a whole. Regulatory work includes all processes aimed at coordinating processes in the cell.
Photosynthesis, its significance, cosmic role
Photosynthesis is the process of converting light energy into the energy of chemical bonds of organic compounds with the participation of chlorophyll.
As a result of photosynthesis, about 150 billion tons of organic matter and approximately 200 billion tons of oxygen are produced annually. This process ensures the carbon cycle in the biosphere, preventing carbon dioxide from accumulating and thereby preventing the greenhouse effect and overheating of the Earth. Organic substances formed as a result of photosynthesis are not completely consumed by other organisms; a significant part of them over the course of millions of years has formed deposits of minerals (hard and brown coal, oil). Recently, rapeseed oil (“biodiesel”) and alcohol obtained from plant residues have also begun to be used as fuel. Ozone is formed from oxygen under the influence of electrical discharges, which forms an ozone screen that protects all life on Earth from the destructive effects of ultraviolet rays.
Our compatriot, the outstanding plant physiologist K. A. Timiryazev (1843-1920), called the role of photosynthesis “cosmic”, since it connects the Earth with the Sun (space), providing an influx of energy to the planet.
Phases of photosynthesis. Light and dark reactions of photosynthesis, their relationship
In 1905, the English plant physiologist F. Blackman discovered that the rate of photosynthesis cannot increase indefinitely; some factor limits it. Based on this, he hypothesized that there are two phases of photosynthesis: light And dark. At low light intensity, the rate of light reactions increases in proportion to the increase in light intensity, and, in addition, these reactions do not depend on temperature, since they do not require enzymes to occur. Light reactions occur on thylakoid membranes.
The rate of dark reactions, on the contrary, increases with increasing temperature, however, upon reaching a temperature threshold of $30°C$, this increase stops, which indicates the enzymatic nature of these transformations occurring in the stroma. It should be noted that light also has a certain effect on dark reactions, despite the fact that they are called dark reactions.

The light phase of photosynthesis occurs on thylakoid membranes carrying several types of protein complexes, the main of which are photosystems I and II, as well as ATP synthase. Photosystems include pigment complexes, which, in addition to chlorophyll, also contain carotenoids. Carotenoids capture light in areas of the spectrum where chlorophyll does not, and also protect chlorophyll from destruction by high-intensity light.
In addition to pigment complexes, photosystems also include a number of electron acceptor proteins, which sequentially transfer electrons from chlorophyll molecules to each other. The sequence of these proteins is called electron transport chain of chloroplasts.
A special complex of proteins is also associated with photosystem II, which ensures the release of oxygen during photosynthesis. This oxygen-releasing complex contains manganese and chlorine ions.
IN light phase light quanta, or photons, falling on chlorophyll molecules located on thylakoid membranes, transfer them to an excited state, characterized by higher electron energy. In this case, excited electrons from the chlorophyll of photosystem I are transferred through a chain of intermediaries to the hydrogen carrier NADP, which attaches hydrogen protons, always present in an aqueous solution:
$NADP + 2e^(-) + 2H^(+) → NADPH + H^(+)$.
The reduced $NADPH + H^(+)$ will subsequently be used in the dark stage. Electrons from the chlorophyll of photosystem II are also transferred along the electron transport chain, but they fill the “electron holes” of the chlorophyll of photosystem I. The lack of electrons in the chlorophyll of photosystem II is filled by taking away water molecules, which occurs with the participation of the oxygen-releasing complex already mentioned above. As a result of the decomposition of water molecules, which is called photolysis, hydrogen protons are formed and molecular oxygen is released, which is a by-product of photosynthesis:
$H_2O → 2H^(+) + 2e^(-) + (1)/(2)O_2$.
Genetic information in a cell. Genes, genetic code and its properties. Matrix nature of biosynthesis reactions. Biosynthesis of protein and nucleic acids
Genetic information in a cell
Reproduction of one's own kind is one of the fundamental properties of living things. Thanks to this phenomenon, there is similarity not only between organisms, but also between individual cells, as well as their organelles (mitochondria and plastids). The material basis of this similarity is the transfer of genetic information encrypted in the DNA nucleotide sequence, which is carried out through the processes of DNA replication (self-duplication). All the characteristics and properties of cells and organisms are realized thanks to proteins, the structure of which is primarily determined by the sequence of DNA nucleotides. Therefore, the biosynthesis of nucleic acids and proteins plays paramount importance in metabolic processes. The structural unit of hereditary information is the gene.
Genes, genetic code and its properties
Hereditary information in a cell is not monolithic; it is divided into separate “words” - genes.
Gene is an elementary unit of genetic information.
Work on the “Human Genome” program, which was carried out simultaneously in several countries and was completed at the beginning of this century, gave us an understanding that a person has only about 25-30 thousand genes, but information from most of our DNA is never read, since it contains a huge number of meaningless sections, repeats and genes encoding traits that have lost meaning for humans (tail, body hair, etc.). In addition, a number of genes responsible for the development of hereditary diseases, as well as drug target genes, have been deciphered. However, the practical application of the results obtained during the implementation of this program is postponed until the genomes of more people are deciphered and it becomes clear how they differ.
Genes that encode the primary structure of protein, ribosomal or transfer RNA are called structural, and genes that provide activation or suppression of reading information from structural genes - regulatory. However, even structural genes contain regulatory regions.
The hereditary information of organisms is encrypted in DNA in the form of certain combinations of nucleotides and their sequence - genetic code. Its properties are: tripletity, specificity, universality, redundancy and non-overlapping. In addition, there are no punctuation marks in the genetic code.
Each amino acid is encoded in DNA by three nucleotides - triplet, for example, methionine is encoded by the TAC triplet, that is, the code is triplet. On the other hand, each triplet encodes only one amino acid, which is its specificity or unambiguity. The genetic code is universal for all living organisms, that is, hereditary information about human proteins can be read by bacteria and vice versa. This indicates the unity of origin of the organic world. However, 64 combinations of three nucleotides correspond to only 20 amino acids, as a result of which one amino acid can be encoded by 2-6 triplets, that is, the genetic code is redundant or degenerate. Three triplets do not have corresponding amino acids, they are called stop codons, since they indicate the end of the synthesis of the polypeptide chain.
The sequence of bases in DNA triplets and the amino acids they encode

*Stop codon, indicating the end of the synthesis of the polypeptide chain.
Abbreviations for amino acid names:
Ala - alanine
Arg - arginine
Asn - asparagine
Asp - aspartic acid
Val - valine
His - histidine
Gly - glycine
Gln - glutamine
Glu - glutamic acid
Ile - isoleucine
Leu - leucine
Liz - lysine
Meth - methionine
Pro - proline
Ser - serine
Tyr - tyrosine
Tre - threonine
Three - tryptophan
Fen - phenylalanine
Cis - cysteine
If you start reading genetic information not from the first nucleotide in the triplet, but from the second, then not only will the reading frame shift, but the protein synthesized in this way will be completely different not only in the nucleotide sequence, but also in structure and properties. There are no punctuation marks between the triplets, so there are no obstacles to shifting the reading frame, which opens up space for the occurrence and maintenance of mutations.
Matrix nature of biosynthesis reactions
Bacterial cells are capable of doubling every 20-30 minutes, and eukaryotic cells - every day and even more often, which requires high speed and accuracy of DNA replication. In addition, each cell contains hundreds and thousands of copies of many proteins, especially enzymes, therefore, the “piecemeal” method of their production is unacceptable for their reproduction. A more progressive method is stamping, which allows you to obtain numerous exact copies of the product and also reduce its cost. For stamping, a matrix is required from which the impression is made.
In cells, the principle of template synthesis is that new molecules of proteins and nucleic acids are synthesized in accordance with the program embedded in the structure of pre-existing molecules of the same nucleic acids (DNA or RNA).
Biosynthesis of protein and nucleic acids
DNA replication. DNA is a double-stranded biopolymer, the monomers of which are nucleotides. If DNA biosynthesis occurred on the principle of photocopying, then numerous distortions and errors in hereditary information would inevitably arise, which would ultimately lead to the death of new organisms. Therefore, the process of DNA doubling occurs differently, in a semi-conservative way: the DNA molecule unwinds, and a new chain is synthesized on each of the chains according to the principle of complementarity. The process of self-reproduction of a DNA molecule, ensuring accurate copying of hereditary information and its transmission from generation to generation, is called replication(from lat. replicationo- repetition). As a result of replication, two absolutely exact copies of the mother DNA molecule are formed, each of which carries one copy of the mother DNA molecule.

The replication process is actually extremely complex, since a number of proteins are involved in it. Some of them unwind the double helix of DNA, others break the hydrogen bonds between the nucleotides of complementary chains, others (for example, the enzyme DNA polymerase) select new nucleotides based on the principle of complementarity, etc. Two DNA molecules formed as a result of replication diverge into two during division newly formed daughter cells.
Errors in the replication process occur extremely rarely, but if they do occur, they are very quickly eliminated by both DNA polymerases and special repair enzymes, since any error in the nucleotide sequence can lead to an irreversible change in the structure and functions of the protein and, ultimately, adversely affect the viability of a new cell or even an individual.
Protein biosynthesis. As the outstanding philosopher of the 19th century F. Engels figuratively put it: “Life is a form of existence of protein bodies.” The structure and properties of protein molecules are determined by their primary structure, i.e., the sequence of amino acids encoded in DNA. Not only the existence of the polypeptide itself, but also the functioning of the cell as a whole depends on the accuracy of the reproduction of this information, so the process of protein synthesis is of great importance. It appears to be the most complex synthesis process in the cell, since it involves up to three hundred different enzymes and other macromolecules. In addition, it flows at high speed, which requires even greater precision.
There are two main stages in protein biosynthesis: transcription and translation.
Transcription(from lat. transcription- rewriting) is the biosynthesis of mRNA molecules on a DNA matrix.

Since the DNA molecule contains two antiparallel chains, reading information from both chains would lead to the formation of completely different mRNAs, therefore their biosynthesis is possible only on one of the chains, which is called coding, or codogenic, in contrast to the second, non-coding, or non-codogenic. The rewriting process is ensured by a special enzyme, RNA polymerase, which selects RNA nucleotides according to the principle of complementarity. This process can occur both in the nucleus and in organelles that have their own DNA - mitochondria and plastids.
The mRNA molecules synthesized during transcription undergo a complex process of preparation for translation (mitochondrial and plastid mRNAs can remain inside the organelles, where the second stage of protein biosynthesis occurs). During the process of mRNA maturation, the first three nucleotides (AUG) and a tail of adenyl nucleotides are attached to it, the length of which determines how many copies of the protein can be synthesized on a given molecule. Only then do mature mRNAs leave the nucleus through nuclear pores.
In parallel, the process of amino acid activation occurs in the cytoplasm, during which the amino acid joins the corresponding free tRNA. This process is catalyzed by a special enzyme and requires ATP.
Broadcast(from lat. broadcast- transfer) is the biosynthesis of a polypeptide chain on an mRNA matrix, during which genetic information is translated into the amino acid sequence of the polypeptide chain.
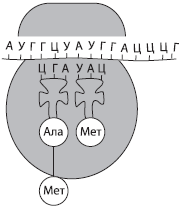
The second stage of protein synthesis most often occurs in the cytoplasm, for example on the rough ER. For its occurrence, the presence of ribosomes, activation of tRNA, during which they attach the corresponding amino acids, the presence of Mg2+ ions, as well as optimal environmental conditions (temperature, pH, pressure, etc.) are necessary.
To start broadcasting ( initiation) a small ribosomal subunit is attached to an mRNA molecule ready for synthesis, and then, according to the principle of complementarity to the first codon (AUG), a tRNA carrying the amino acid methionine is selected. Only after this does the large ribosomal subunit attach. Within the assembled ribosome there are two mRNA codons, the first of which is already occupied. A second tRNA, also carrying an amino acid, is added to the codon adjacent to it, after which a peptide bond is formed between the amino acid residues with the help of enzymes. The ribosome moves one codon of the mRNA; the first tRNA freed from an amino acid returns to the cytoplasm after the next amino acid, and a fragment of the future polypeptide chain hangs, as it were, on the remaining tRNA. The next tRNA is attached to the new codon that finds itself within the ribosome, the process is repeated and step by step the polypeptide chain lengthens, i.e. elongation.
End of protein synthesis ( termination) occurs as soon as a specific nucleotide sequence is encountered in the mRNA molecule that does not code for an amino acid (stop codon). After this, the ribosome, mRNA and polypeptide chain are separated, and the newly synthesized protein acquires the appropriate structure and is transported to the part of the cell where it will perform its functions.
Translation is a very energy-intensive process, since the energy of one ATP molecule is consumed to attach one amino acid to tRNA, and several more are used to move the ribosome along the mRNA molecule.
To speed up the synthesis of certain protein molecules, several ribosomes can be successively attached to an mRNA molecule, which form a single structure - polysome.
A cell is the genetic unit of a living thing. Chromosomes, their structure (shape and size) and functions. The number of chromosomes and their species constancy. Somatic and germ cells. Cell life cycle: interphase and mitosis. Mitosis is the division of somatic cells. Meiosis. Phases of mitosis and meiosis. Development of germ cells in plants and animals. Cell division is the basis for the growth, development and reproduction of organisms. The role of meiosis and mitosis
A cell is the genetic unit of a living thing.
Despite the fact that nucleic acids are the carrier of genetic information, the implementation of this information is impossible outside the cell, which is easily proven by the example of viruses. These organisms, often containing only DNA or RNA, cannot reproduce independently; to do this, they must use the hereditary apparatus of the cell. They cannot even penetrate a cell without the help of the cell itself, except through the use of membrane transport mechanisms or due to cell damage. Most viruses are unstable; they die after just a few hours of exposure to the open air. Consequently, a cell is a genetic unit of a living thing, which has a minimum set of components for preserving, changing and implementing hereditary information, as well as its transmission to descendants.
Most of the genetic information of a eukaryotic cell is located in the nucleus. The peculiarity of its organization is that, unlike the DNA of a prokaryotic cell, the DNA molecules of eukaryotes are not closed and form complex complexes with proteins - chromosomes.
Chromosomes, their structure (shape and size) and functions
Chromosome(from Greek chromium- color, coloring and soma- body) is the structure of the cell nucleus, which contains genes and carries certain hereditary information about the characteristics and properties of the organism.
Sometimes the circular DNA molecules of prokaryotes are also called chromosomes. Chromosomes are capable of self-duplication; they have structural and functional individuality and retain it over generations. Each cell carries all the hereditary information of the body, but only a small part works in it.
The basis of a chromosome is a double-stranded DNA molecule packed with proteins. In eukaryotes, histone and non-histone proteins interact with DNA, whereas in prokaryotes, histone proteins are absent.
Chromosomes are best seen under a light microscope during cell division, when, as a result of compaction, they take on the appearance of rod-shaped bodies separated by a primary constriction - centromere — on shoulders. On a chromosome there may also be secondary constriction, which in some cases separates the so-called satellite. The ends of chromosomes are called telomeres. Telomeres prevent the ends of chromosomes from sticking together and ensure their attachment to the nuclear membrane in a non-dividing cell. At the beginning of division, the chromosomes are doubled and consist of two daughter chromosomes - chromatid, fastened at the centromere.

According to their shape, chromosomes are divided into equal-armed, unequal-armed and rod-shaped chromosomes. The sizes of chromosomes vary significantly, but the average chromosome has dimensions of 5 $×$ 1.4 microns.
In some cases, chromosomes, as a result of numerous DNA duplications, contain hundreds and thousands of chromatids: such giant chromosomes are called polytene. They are found in the salivary glands of Drosophila larvae, as well as in the digestive glands of roundworms.
The number of chromosomes and their species constancy. Somatic and germ cells
According to cellular theory, a cell is a unit of structure, vital activity and development of an organism. Thus, such important functions of living things as growth, reproduction and development of the organism are provided at the cellular level. Cells of multicellular organisms can be divided into somatic and reproductive cells.
Somatic cells- these are all the cells of the body formed as a result of mitotic division.
The study of chromosomes has made it possible to establish that the somatic cells of the body of each biological species are characterized by a constant number of chromosomes. For example, a person has 46 of them. The set of chromosomes of somatic cells is called diploid(2n), or double.

Sex cells, or gametes, are specialized cells used for sexual reproduction.
Gametes always contain half as many chromosomes as somatic cells (in humans - 23), therefore the set of chromosomes of germ cells is called haploid(n), or single. Its formation is associated with meiotic cell division.
The amount of DNA in somatic cells is designated as 2c, and in sex cells - 1c. The genetic formula of somatic cells is written as 2n2c, and sexual cells - 1n1c.
In the nuclei of some somatic cells, the number of chromosomes may differ from their number in somatic cells. If this difference is greater than one, two, three, etc. haploid sets, then such cells are called polyploid(tri-, tetra-, pentaploid, respectively). In such cells, metabolic processes usually proceed very intensively.
The number of chromosomes in itself is not a species-specific feature, since different organisms can have an equal number of chromosomes, but related ones can have a different number. For example, the malarial plasmodium and the horse roundworm each have two chromosomes, while humans and chimpanzees have 46 and 48, respectively.
Human chromosomes are divided into two groups: autosomes and sex chromosomes (heterochromosomes). Autosome in human somatic cells there are 22 pairs, they are the same for men and women, and sex chromosomes only one pair, but it is this that determines the sex of the individual. There are two types of sex chromosomes - X and Y. Women's body cells carry two X chromosomes, and men's - X and Y.
Karyotype- this is a set of characteristics of the chromosome set of an organism (the number of chromosomes, their shape and size).
The conditional record of a karyotype includes the total number of chromosomes, sex chromosomes and possible deviations in the set of chromosomes. For example, the karyotype of a normal man is written as 46, XY, and the karyotype of a normal woman is 46, XX.
Cell life cycle: interphase and mitosis
Cells do not arise anew every time, they are formed only as a result of the division of mother cells. After division, the daughter cells require some time to form organelles and acquire the appropriate structure that would ensure the performance of a specific function. This period of time is called maturation.
The period of time from the appearance of a cell as a result of division until its division or death is called life cycle of a cell.
In eukaryotic cells, the life cycle is divided into two main stages: interphase and mitosis.
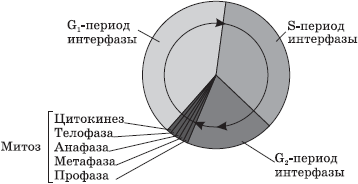
Interphase- this is a period of time in the life cycle during which the cell does not divide and functions normally. Interphase is divided into three periods: G 1 -, S- and G 2 -periods.
G 1 -period(presynthetic, postmitotic) is a period of cell growth and development during which active synthesis of RNA, proteins and other substances necessary for the complete life support of the newly formed cell occurs. Towards the end of this period, the cell may begin to prepare to duplicate its DNA.
IN S-period(synthetic) the process of DNA replication itself occurs. The only part of the chromosome that does not undergo replication is the centromere, so the resulting DNA molecules do not diverge completely, but remain held together in it, and at the beginning of division the chromosome has an X-shaped appearance. The genetic formula of a cell after DNA doubling is 2n4c. Also in the S-period, the centrioles of the cell center are doubled.
G 2 -period(postsynthetic, premitotic) is characterized by intensive synthesis of RNA, proteins and ATP necessary for the process of cell division, as well as the separation of centrioles, mitochondria and plastids. Until the end of interphase, chromatin and the nucleolus remain clearly distinguishable, the integrity of the nuclear envelope is not disrupted, and the organelles do not change.
Some of the body's cells are able to perform their functions throughout the life of the body (neurons of our brain, muscle cells of the heart), while others exist for a short time, after which they die (intestinal epithelial cells, epidermal cells of the skin). Consequently, the body must constantly undergo processes of cell division and the formation of new ones that would replace dead ones. Cells capable of dividing are called stem. In the human body they are found in the red bone marrow, in the deep layers of the epidermis of the skin and other places. Using these cells, you can grow a new organ, achieve rejuvenation, and also clone the body. The prospects for using stem cells are absolutely clear, but the moral and ethical aspects of this problem are still being discussed, since in most cases embryonic stem cells obtained from human embryos killed during abortion are used.
The duration of interphase in plant and animal cells averages 10-20 hours, while mitosis takes about 1-2 hours.
During successive divisions in multicellular organisms, daughter cells become increasingly diverse as they read information from an increasing number of genes.
Some cells stop dividing over time and die, which may be due to the completion of certain functions, as in the case of epidermal skin cells and blood cells, or due to damage to these cells by environmental factors, in particular pathogens. Genetically programmed cell death is called apoptosis, while accidental death - necrosis.
Mitosis is the division of somatic cells. Phases of mitosis
Mitosis- a method of indirect division of somatic cells.
During mitosis, the cell goes through a series of successive phases, as a result of which each daughter cell receives the same set of chromosomes as in the mother cell.
Mitosis is divided into four main phases: prophase, metaphase, anaphase and telophase. Prophase- the longest stage of mitosis, during which chromatin condenses, resulting in X-shaped chromosomes consisting of two chromatids (daughter chromosomes) becoming visible. In this case, the nucleolus disappears, the centrioles diverge to the poles of the cell, and an achromatin spindle (division spindle) from microtubules begins to form. At the end of prophase, the nuclear membrane disintegrates into separate vesicles.

IN metaphase The chromosomes are lined up along the equator of the cell with their centromeres, to which the microtubules of the fully formed spindle are attached. At this stage of division, the chromosomes are most compacted and have a characteristic shape, which makes it possible to study the karyotype.
IN anaphase Rapid DNA replication occurs at centromeres, as a result of which chromosomes are split and chromatids diverge to the poles of the cell, stretched by microtubules. The distribution of chromatids must be absolutely equal, since it is this process that ensures the maintenance of a constant number of chromosomes in the cells of the body.
On the stage telophases daughter chromosomes gather at the poles, despiral, nuclear membranes form around them from vesicles, and nucleoli appear in the newly formed nuclei.
After nuclear division, cytoplasmic division occurs - cytokinesis, during which a more or less uniform distribution of all organelles of the mother cell occurs.
Thus, as a result of mitosis, two daughter cells are formed from one mother cell, each of which is a genetic copy of the mother cell (2n2c).
In sick, damaged, aging cells and specialized tissues of the body, a slightly different division process can occur - amitosis. Amitosis called direct division of eukaryotic cells, in which the formation of genetically equivalent cells does not occur, since the cellular components are distributed unevenly. It is found in plants in the endosperm, and in animals - in the liver, cartilage and cornea of the eye.
Meiosis. Phases of meiosis
Meiosis is a method of indirect division of primary germ cells (2n2c), which results in the formation of haploid cells (1n1c), most often germ cells.
Unlike mitosis, meiosis consists of two successive cell divisions, each of which is preceded by interphase. The first division of meiosis (meiosis I) is called reductionist, since in this case the number of chromosomes is halved, and the second division (meiosis II) - equational, since in its process the number of chromosomes is preserved.

Interphase I proceeds like interphase of mitosis. Meiosis I is divided into four phases: prophase I, metaphase I, anaphase I and telophase I. B prophase I Two important processes occur: conjugation and crossing over. Conjugation- This is the process of fusion of homologous (paired) chromosomes along the entire length. The pairs of chromosomes formed during conjugation are preserved until the end of metaphase I.
Crossing over- mutual exchange of homologous regions of homologous chromosomes. As a result of crossing over, the chromosomes received by the body from both parents acquire new combinations of genes, which causes the appearance of genetically diverse offspring. At the end of prophase I, as in the prophase of mitosis, the nucleolus disappears, the centrioles diverge to the poles of the cell, and the nuclear membrane disintegrates.

IN metaphase I pairs of chromosomes are aligned along the equator of the cell, and spindle microtubules are attached to their centromeres.
IN anaphase I Whole homologous chromosomes, consisting of two chromatids, diverge to the poles.
IN telophase I Nuclear membranes are formed around clusters of chromosomes at the poles of the cell, and nucleoli are formed.
Cytokinesis I ensures separation of the cytoplasms of daughter cells.
The daughter cells (1n2c) formed as a result of meiosis I are genetically heterogeneous, since their chromosomes, randomly dispersed to the cell poles, contain different genes.
Comparative characteristics of mitosis and meiosis
| Sign | Mitosis | Meiosis | |
| Which cells begin to divide? | Somatic (2n) | Primary germ cells (2n) | |
| Number of divisions | 1 | 2 | |
| How many and what kind of cells are formed during division? | 2 somatic (2n) | 4 sexual (n) | |
| Interphase | Preparing the cell for division, DNA doubling | Very short, DNA doubling does not occur | |
| Phases | Meiosis I | Meiosis II | |
| Prophase | Chromosome condensation, disappearance of the nucleolus, disintegration of the nuclear membrane, conjugation and crossing over may occur | Chromosome condensation, disappearance of the nucleolus, disintegration of the nuclear membrane | |
| Metaphase | Pairs of chromosomes are located along the equator, a spindle is formed | Chromosomes line up along the equator, a spindle is formed | |
| Anaphase | Homologous chromosomes from two chromatids move towards the poles | Chromatids move towards the poles | |
| Telophase | Chromosomes despiral, new nuclear membranes and nucleoli are formed | Chromosomes despiral, new nuclear membranes and nucleoli are formed | |
Interphase II very short, since DNA doubling does not occur in it, that is, there is no S-period.
Meiosis II also divided into four phases: prophase II, metaphase II, anaphase II and telophase II. IN prophase II the same processes occur as in prophase I, with the exception of conjugation and crossing over.
IN metaphase II chromosomes are located along the equator of the cell.
IN anaphase II chromosomes are split at centromeres and chromatids are stretched towards the poles.
IN telophase II Nuclear membranes and nucleoli are formed around clusters of daughter chromosomes.
After cytokinesis II The genetic formula of all four daughter cells is 1n1c, but they all have a different set of genes, which is the result of crossing over and the random combination of chromosomes of the maternal and paternal organisms in the daughter cells.
Development of germ cells in plants and animals
Gametogenesis(from Greek gamete- wife, gametes- husband and genesis- origin, emergence) is the process of formation of mature germ cells.
Since sexual reproduction most often requires two individuals - a female and a male, producing different sex cells - eggs and sperm, then the processes of formation of these gametes must be different.
The nature of the process depends to a significant extent on whether it occurs in a plant or animal cell, since in plants only mitosis occurs during the formation of gametes, and in animals both mitosis and meiosis occur.
Development of germ cells in plants. In angiosperms, the formation of male and female reproductive cells occurs in different parts of the flower - the stamens and pistils, respectively.
Before the formation of male reproductive cells - microgametogenesis(from Greek micros- small) - happens microsporogenesis, that is, the formation of microspores in the anthers of stamens. This process is associated with the meiotic division of the mother cell, which results in four haploid microspores. Microgametogenesis is associated with mitotic division of the microspore, giving a male gametophyte from two cells - a large vegetative(siphonogenic) and shallow generative. After division, the male gametophyte becomes covered with dense membranes and forms a pollen grain. In some cases, even during the process of pollen maturation, and sometimes only after transfer to the stigma of the pistil, the generative cell divides mitotically to form two immobile male germ cells - sperm. After pollination, a pollen tube is formed from the vegetative cell, through which sperm penetrate into the ovary of the pistil for fertilization.

The development of female germ cells in plants is called megagametogenesis(from Greek megas- big). It occurs in the ovary of the pistil, which is preceded by megasporogenesis, as a result of which four megaspores are formed from the mother cell of the megaspore lying in the nucellus through meiotic division. One of the megaspores divides mitotically three times, giving the female gametophyte - an embryo sac with eight nuclei. With the subsequent separation of the cytoplasms of the daughter cells, one of the resulting cells becomes an egg, on the sides of which lie the so-called synergids, at the opposite end of the embryo sac three antipodes are formed, and in the center, as a result of the fusion of two haploid nuclei, a diploid central cell is formed.
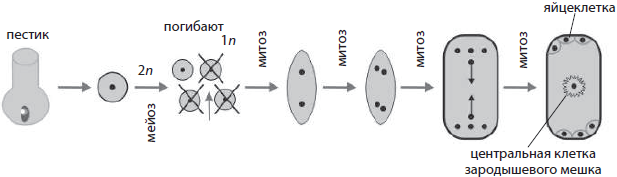
Development of germ cells in animals. In animals, there are two processes of formation of germ cells - spermatogenesis and oogenesis.
Spermatogenesis(from Greek sperm, spermatos- seed and genesis- origin, occurrence) is the process of formation of mature male germ cells - sperm. In humans, it occurs in the testes, or testicles, and is divided into four periods: reproduction, growth, maturation and formation.

IN breeding season primordial germ cells divide mitotically, resulting in the formation of diploid spermatogonia. IN growth period spermatogonia accumulate nutrients in the cytoplasm, increase in size and turn into primary spermatocytes, or 1st order spermatocytes. Only after this do they enter meiosis ( maturation period), as a result of which first two are formed secondary spermatocyte, or 2nd order spermatocyte, and then - four haploid cells with a fairly large amount of cytoplasm - spermatids. IN formation period they lose almost all their cytoplasm and form a flagellum, turning into sperm.
Sperm, or livelies, - very small mobile male reproductive cells with a head, neck and tail.

IN head, in addition to the core, is acrosome- a modified Golgi complex that ensures the dissolution of the egg membranes during fertilization. IN cervix are the centrioles of the cell center, and the base ponytail form microtubules that directly support sperm movement. It also contains mitochondria, which provide the sperm with ATP energy for movement.
Oogenesis(from Greek UN- egg and genesis- origin, occurrence) is the process of formation of mature female germ cells - eggs. In humans, it occurs in the ovaries and consists of three periods: reproduction, growth and maturation. Periods of reproduction and growth, similar to those in spermatogenesis, occur during intrauterine development. In this case, diploid cells are formed from primary germ cells as a result of mitosis. oogonia, which then turn into diploid primary oocytes, or 1st order oocytes. Meiosis and subsequent cytokinesis occurring in maturation period, are characterized by uneven division of the cytoplasm of the mother cell, so that in the end, at first one is obtained secondary oocyte, or 2nd order oocyte, And first polar body, and then from the secondary oocyte - the egg, which retains the entire supply of nutrients, and the second polar body, while the first polar body is divided into two. Polar bodies take up excess genetic material.
In humans, eggs are produced with an interval of 28-29 days. The cycle associated with the maturation and release of eggs is called menstrual.
Egg- a large female reproductive cell that carries not only a haploid set of chromosomes, but also a significant supply of nutrients for the subsequent development of the embryo.

The egg in mammals is covered with four membranes, which reduce the likelihood of damage by various factors. The diameter of the egg in humans reaches 150-200 microns, while in an ostrich it can be several centimeters.
Cell division is the basis for the growth, development and reproduction of organisms. The role of mitosis and meiosis
If in unicellular organisms cell division leads to an increase in the number of individuals, i.e., reproduction, then in multicellular organisms this process can have different meanings. Thus, the division of embryonic cells, starting from the zygote, is the biological basis of the interconnected processes of growth and development. Similar changes are observed in humans during adolescence, when the number of cells not only increases, but also a qualitative change in the body occurs. The reproduction of multicellular organisms is also based on cell division, for example, in asexual reproduction, thanks to this process, a whole part of the organism is restored, and in sexual reproduction, in the process of gametogenesis, sex cells are formed, which subsequently give rise to a new organism. It should be noted that the main methods of division of a eukaryotic cell - mitosis and meiosis - have different meanings in the life cycles of organisms.
As a result of mitosis, there is an even distribution of hereditary material between daughter cells - exact copies of the mother. Without mitosis, the existence and growth of multicellular organisms developing from a single cell, the zygote, would be impossible, since all cells of such organisms must contain the same genetic information.
During the process of division, daughter cells become more and more diverse in structure and functions, which is associated with the activation of more and more new groups of genes in them due to intercellular interaction. Thus, mitosis is necessary for the development of the organism.
This method of cell division is necessary for the processes of asexual reproduction and regeneration (restoration) of damaged tissues, as well as organs.
Meiosis, in turn, ensures the constancy of the karyotype during sexual reproduction, since it halves the set of chromosomes before sexual reproduction, which is then restored as a result of fertilization. In addition, meiosis leads to the emergence of new combinations of parental genes due to crossing over and random combination of chromosomes in daughter cells. Thanks to this, the offspring turns out to be genetically diverse, which provides material for natural selection and is the material basis for evolution. A change in the number, shape and size of chromosomes, on the one hand, can lead to the appearance of various deviations in the development of the organism and even its death, and on the other hand, it can lead to the appearance of individuals more adapted to the environment.
Thus, the cell is the unit of growth, development and reproduction of organisms.
We invite you to familiarize yourself with the materials and.
: cellulose membrane, membrane, cytoplasm with organelles, nucleus, vacuoles with cell sap.The presence of plastids is the main feature of a plant cell.
Functions of the cell membrane- determines the shape of the cell, protects against environmental factors.
Plasma membrane- a thin film, consisting of interacting molecules of lipids and proteins, delimits the internal contents from the external environment, ensures the transport of water, minerals and organic substances into the cell by osmosis and active transport, and also removes waste products.
Cytoplasm- the internal semi-liquid environment of the cell, in which the nucleus and organelles are located, provides connections between them, and participates in basic life processes.
Endoplasmic reticulum- a network of branching channels in the cytoplasm. It is involved in the synthesis of proteins, lipids and carbohydrates, and in the transport of substances. Ribosomes are bodies located on the ER or in the cytoplasm, consisting of RNA and protein, and are involved in protein synthesis. EPS and ribosomes are a single apparatus for the synthesis and transport of proteins.
Mitochondria- organelles delimited from the cytoplasm by two membranes. Organic substances are oxidized in them and ATP molecules are synthesized with the participation of enzymes. Increase in the surface of the inner membrane on which enzymes are located due to cristae. ATP is an energy-rich organic substance.
Plastids(chloroplasts, leucoplasts, chromoplasts), their content in the cell is the main feature of the plant organism. Chloroplasts are plastids containing the green pigment chlorophyll, which absorbs light energy and uses it to synthesize organic substances from carbon dioxide and water. Chloroplasts are separated from the cytoplasm by two membranes, numerous outgrowths - grana on the inner membrane, in which chlorophyll molecules and enzymes are located.
Golgi complex- a system of cavities delimited from the cytoplasm by a membrane. The accumulation of proteins, fats and carbohydrates in them. Carrying out the synthesis of fats and carbohydrates on membranes.
Lysosomes- bodies delimited from the cytoplasm by a single membrane. The enzymes they contain accelerate the breakdown of complex molecules into simple ones: proteins into amino acids, complex carbohydrates into simple ones, lipids into glycerol and fatty acids, and also destroy dead parts of the cell and entire cells.
Vacuoles- cavities in the cytoplasm filled with cell sap, a place of accumulation of reserve nutrients and harmful substances; they regulate the water content in the cell.
Core- the main part of the cell, covered on the outside with a two-membrane, pore-pierced nuclear envelope. Substances enter the core and are removed from it through the pores. Chromosomes are carriers of hereditary information about the characteristics of an organism, the main structures of the nucleus, each of which consists of one DNA molecule combined with proteins. The nucleus is the site of DNA, mRNA, and rRNA synthesis.

The presence of an outer membrane, cytoplasm with organelles, and a nucleus with chromosomes.
Outer or plasma membrane- delimits the contents of the cell from the environment (other cells, intercellular substance), consists of lipid and protein molecules, ensures communication between cells, transport of substances into the cell (pinocytosis, phagocytosis) and out of the cell.
Cytoplasm- the internal semi-liquid environment of the cell, which provides communication between the nucleus and organelles located in it. The main life processes take place in the cytoplasm.
Cell organelles:
1) endoplasmic reticulum (ER)- a system of branching tubules, participates in the synthesis of proteins, lipids and carbohydrates, in the transport of substances in the cell;
2) ribosomes- bodies containing rRNA are located on the ER and in the cytoplasm and participate in protein synthesis. EPS and ribosomes are a single apparatus for protein synthesis and transport;
3) mitochondria- “power stations” of the cell, delimited from the cytoplasm by two membranes. The inner one forms cristae (folds), increasing its surface. Enzymes on the cristae accelerate the oxidation of organic substances and the synthesis of energy-rich ATP molecules;
4) Golgi complex- a group of cavities delimited by a membrane from the cytoplasm, filled with proteins, fats and carbohydrates, which are either used in vital processes or removed from the cell. The membranes of the complex carry out the synthesis of fats and carbohydrates;
5) lysosomes- bodies filled with enzymes accelerate the breakdown of proteins into amino acids, lipids into glycerol and fatty acids, polysaccharides into monosaccharides. In lysosomes, dead parts of the cell, whole cells, are destroyed.
Cellular inclusions- accumulations of reserve nutrients: proteins, fats and carbohydrates.
Core- the most important part of the cell. It is covered with a double-membrane shell with pores, through which some substances penetrate into the nucleus, and others enter the cytoplasm. Chromosomes are the main structures of the nucleus, carriers of hereditary information about the characteristics of the organism. It is transmitted during the division of the mother cell to daughter cells, and with germ cells to daughter organisms. The nucleus is the site of DNA, mRNA, and rRNA synthesis.
Exercise:
Explain why organelles are called specialized cell structures?
Answer: organelles are called specialized cell structures, since they perform strictly defined functions, hereditary information is stored in the nucleus, ATP is synthesized in mitochondria, photosynthesis occurs in chloroplasts, etc.
If you have questions about cytology, you can contact
summary of other presentations“Methods of teaching biology” - School zoology. Introducing students to the use of scientific zoological data. Moral education. Additional blessing of the chicken coop. Selection of methods. Life processes. Aquarium fish. Nutrition. Environmental education. Materiality of life processes. Negative results. Attention students. Mandatory form. Looking at small animals. Goals and objectives of biology. Story.
“Problem-based learning in biology lessons” - Knowledge. New textbooks. The path to the solution. Problem. Seminars. What is a task? Albrecht Durer. Problem-based learning in biology lessons. Non-standard lessons. What is meant by problem-based learning? The quality of life. Biology as an academic subject. Question. Problem solving lesson. Decreased interest in the subject. Problem-based laboratory classes.
“Critical thinking in biology lessons” - Technology of “critical thinking”. Using the technology of “development of critical thinking”. Table for the lesson. Motivation to study. Ecosystems. The meaning of “developing critical thinking.” Signs of technology. RKM technology. Lesson structure. Main directions. History of technology. Pedagogical technologies. Rules of technology. Biology assignments. Photosynthesis. Techniques used at different stages of the lesson.
“Biology lessons with an interactive whiteboard” - Electronic textbooks. Benefits for students. An interactive whiteboard helps convey information to every student. Didactic tasks. Solving biological problems. Advantages of working with interactive whiteboards. Working with presentations. Work on comparing objects. Moving objects. Using spreadsheets. Using an interactive whiteboard in the process of teaching schoolchildren. Benefits for teachers.
“System-activity approach in biology” - Seminar questions. Activity method. Dryopithecus. Extraterrestrial path of human origin. Lysosomes. Chemical organization. Gymnosperms. Metabolism. Analyzers. System-activity approach to teaching biology. Chromosomes. Cytoplasm. Blindness. Ear length. Classification of a person. Skeleton of a mammal. Paths of human evolution. Mitosis. Superficial complex. Problematic question. Nucleolus. Nuclear envelope.
“Computer in Biology” - Collaborative activity of students. Angiosperm families. Interactive training. Models of learning. An example of an assessment system. Instruction card questions. Example of an instruction card. Researchers. Microgroups. Interactive learning technologies. Carousel. Interactive learning technologies. Interactive approaches in biology lessons. Group form of work. Tasks for groups of “researchers”.
The connection of the organism with the environment, from a physicochemical point of view, is an open system, that is, a system where biochemical processes are ongoing. The starting substances come from the environment, and the substances that are also continuously formed are carried outside. The equilibrium between the speed and concentration of products of multidirectional reactions in the body is conditional, imaginary, since the intake and removal of substances does not stop. Continuous connection with the environment allows us to consider a living organism as an open system.
For all living cells, the source of energy is the Sun. Plant cells capture energy from sunlight with the help of chlorophyll, using it for assimilation reactions during the process of photosynthesis. Cells of animals, fungi, and bacteria use solar energy indirectly, during the breakdown of organic substances synthesized by earthly plants.
Some of the cell's nutrients are broken down during cellular respiration, thus supplying the energy necessary for various types of cellular activity. This process takes place in organelles called mitochondria. Mitochondria consists of two membranes: the outer one, separating the organelle from the cytoplasm, and the inner one, forming numerous folds. The main product of respiration is ATP. It leaves the mitochondria and is used as an energy source for many chemical reactions in the cytoplasm and cell membrane. If oxygen is required for cellular respiration, then respiration is called aerobic, but if reactions occur in the absence of oxygen, then we speak of anaerobic respiration.
For any type of work performed in a cell, energy is used in one and only form - in the form of energy from the phosphate bonds of ATP. ATP is an easily mobile compound. The formation of ATP occurs on the inner membrane of mitochondria. ATP is synthesized in all cells during respiration due to the energy of oxidation of carbohydrates, fats and other organic substances. In green plant cells, the main amount of ATP is synthesized in chloroplasts due to solar energy. During photosynthesis, they produce many times more ATP than mitochondria. ATP decomposes with the rupture of phosphorus-oxygen bonds and the release of energy. This occurs under the action of the enzyme ATPase during the hydrolysis of ATP - the addition of water with the elimination of a phosphoric acid molecule. As a result, ATP is converted into ADP, and if two molecules of phosphoric acid are split off, then into AMP. The reaction of elimination of each gram-molecule of acid is accompanied by the release of 40 kJ. This is a very large energy output, which is why the phosphorus-oxygen bonds of ATP are usually called macroergistic (high-energy).
The use of ATP in plastic exchange reactions is carried out by coupling them with ATP hydrolysis. Molecules of various substances are charged with energy by attaching the phosphorus group released during hydrolysis from the ATP molecule, i.e. by phosphorylation.
The peculiarity of phosphate derivatives is that they cannot leave the cell, although their “discharged” forms freely pass through the membrane. Thanks to this, phosphorylated molecules remain in the cell until they are used in appropriate reactions.
The reverse process of converting ADP into ATP occurs by adding a phosphoric acid molecule to ADP, releasing water and absorbing a large amount of energy.
Thus, ATP is a universal and direct source of energy for cell activity. This creates a single cellular pool of energy and makes it possible to redistribute and transport it from one area of the cell to another.
The transfer of the phosphate group plays an important role in chemical reactions such as the assembly of macromolecules from monomers. For example, amino acids can be combined into peptides only after being previously phosphorylated. Mechanical processes of contraction or movement, transport of a dissolved substance against a concentration gradient and other processes involve the consumption of energy stored in ATP.
The process of energy metabolism can be represented as follows. High-molecular organic substances in the cytoplasm are enzymatically, by hydrolysis, converted into simpler ones from which they consist: proteins - into amino acids, poly- and disaccharides - into monosaccharides (+ glucose), fats into glycerol and fatty acids. There are no oxidative processes, little energy is released, which is not used and goes into thermal form. Most cells use carbohydrates first. Polysaccharides (starch in plants and glycogen in animals) are hydrolyzed to glucose. Glucose oxidation occurs in three phases: glycolysis, oxidative decarboxylation (Krebs cycle - citric acid cycle) and oxidative phosphorylation (respiratory chain). Glycolysis, as a result of which one molecule of glucose is split into two molecules of pyruvic acid with the release of two molecules of ATP, occurs in the cytoplasm. In the absence of oxygen, pyruvic acid is converted to either ethanol (fermentation) or lactic acid (anaerobic respiration).
When glycolysis occurs in animal cells, the six-carbon molecule of glucose breaks down into two molecules of lactic acid. This process is multi-stage. It is carried out sequentially by 13 enzymes. During alcoholic fermentation, two molecules of ethanol and two molecules of CO2 are formed from a glucose molecule.
Glycolysis is a phase common to anaerobic and aerobic respiration; the other two occur only under aerobic conditions. The process of oxygen-free oxidation, in which only part of the energy of metabolites is released and used, is final for anaerobic organisms. In the presence of oxygen, pyruvic acid passes into the mitochondria, where, as a result of a number of sequential reactions, it is completely oxidized aerobically to H2O and CO2 with simultaneous phosphorylation of ADP to ATP. In this case, two ATP molecules are produced by glycolysis, two by the Krebs cycle, and 34 by the respiratory chain. The net yield for the complete oxidation of one glucose molecule to H2O and CO2 is 38 molecules.
Thus, in aerobic organisms, the final decomposition of organic substances is carried out by oxidizing them with atmospheric oxygen to simple inorganic substances: CO2 and H2O. This process takes place on the cristae of mitochondria. In this case, the maximum amount of free energy is released, a significant part of which is reserved in ATP molecules. It is easy to see that aerobic oxidation provides the cell with free energy to the greatest extent.
As a result of catabolism, energy-rich ATP molecules accumulate in the cell, and CO2 and excess water are released into the external environment.
Sugar molecules not required for respiration can be stored in the cell. Excess lipids are either broken down, after which the products of their breakdown enter the mitochondria as a substrate for respiration, or are deposited as reserves in the cytoplasm in the form of fat droplets. Proteins are built from amino acids entering the cell. Protein synthesis occurs in organelles called ribosomes. Each ribosome consists of two subparticles - large and small: both subparticles include protein molecules and RNA molecules.
Ribosomes are often attached to a special membrane system consisting of cisterns and vesicles - the so-called endoplasmic reticulum (ER); in cells that produce a lot of protein, the endoplasmic reticulum is often very well developed and covered with ribosomes. Some enzymes are only effective if they are attached to a membrane. Most of the enzymes involved in lipid synthesis are located here. Thus, the endoplasmic reticulum is like a kind of cell workbench.
In addition, the ER divides the cytoplasm into separate compartments, i.e., it separates various chemical processes occurring simultaneously in the cytoplasm, and thereby reduces the likelihood that these processes will interfere with each other.
Products produced by a given cell are often used outside the cell. In such cases, proteins synthesized on ribosomes pass through the membranes of the endoplasmic reticulum and are packaged into membrane vesicles that form around them, which are then detached from the ER. These vesicles, flattened and stacked on top of each other, like stacked pancakes, form a characteristic structure called the Golgi complex, or Golgi apparatus. During their stay in the Golgi apparatus, proteins undergo certain changes. When the time comes for them to leave the cell, the membrane vesicles merge with the cell membrane and are emptied, pouring their contents out, i.e., secretion occurs by exocytosis.
The Golgi apparatus also produces lysosomes - membrane sacs containing digestive enzymes. Finding out how a cell makes, packages, and exports certain proteins, and how it “knows” which proteins it should keep for itself, is one of the most fascinating branches of modern cytology.
The membranes of any cell are constantly moving and changing. ER membranes move slowly throughout the cell. Individual sections of these membranes separate and form vesicles, which temporarily become part of the Golgi apparatus, and then, through the process of exocytosis, merge with the cell membrane.
Later, the membrane material is returned to the cytoplasm, where it is used again.