СОКРАТИТЕЛЬНАЯ ВАКУОЛЬ, см. ВАКУОЛЬ … Научно-технический энциклопедический словарь
Contractile vacuole сократительная вакуоль. Tип вакуоли у некоторых групп протистов, участвующей в выведении воды (растворов) из клетки при сокращении и в поглощении воды клеткой при расширении, что служит для регуляции осмотического давления.… … Молекулярная биология и генетика. Толковый словарь.
Структура эукариотической клетки. Вакуоль указана под номером 10 Вакуоль одномембранный органоид, содержащийся в некоторых эукариотических клетках и … Википедия
Или Heliozoa отряд класса саркодиковых (см.) типа простейших (см.) животных. Морфологические свойства. Отличаются шаровидным протоплазматическим телом, от которого по всем направлениям, наподобие лучей, отходят тонкие, нитевидные, не… … Энциклопедический словарь Ф.А. Брокгауза и И.А. Ефрона
Или Ciliata отряд класса наливочных, или инфузорий (см.), типа простейших (см.). РЕСНИЧНЫЕ ИНФУЗОРИИ. I (Aspirotricha). Значение букв: а порошица; al альвеолярный слой эктоплазмы; ad.Z адоральный ряд ресничек; b осязательная щетинка; cl реснички; … Энциклопедический словарь Ф.А. Брокгауза и И.А. Ефрона
- (Lobosea), класс наиболее просто организованных простейших над класса корненожек. Лишены внутр. скелета и наруж. раковины. Форма тела непостоянна, размеры обычно от 20 до 700 мкм, реже несколько более. Форма и размеры псевдоподий характерны для… … Биологический энциклопедический словарь
- (Flagellata s. Mastigophora, см. табл. Биченосцы, Flagellata) класс простейших животных (Protozoa). Как и все прочие представители этого типа, они имеют тело, состоящее только из одной клеточки, представляющей протоплазму и ядро с ядрышком.… … Энциклопедический словарь Ф.А. Брокгауза и И.А. Ефрона
Или Protozoa. Содержание статьи: Характеристика и классификация. Исторический очерк. Морфология; протоплазма с включениями (трихоцисты, ядро, сократительные вакуоли, хроматофоры и др.). Покровы и скелет. Движение П.; псевдоподии, жгутики и… … Энциклопедический словарь Ф.А. Брокгауза и И.А. Ефрона
А. СОКРАТИТЕЛЬНЫЕ ВАКУОЛИ
1. Морфология сократительных вакуолей
Большинство простейших (кроме Sporozoa) обладают особым, более или менее строго локализованным, иногда весьма сложно устроенным экскреторным аппаратом, большей частью имеющим характер так называемых сократительных, или пульсирующих, вакуолей.
Как общее правило, вакуоли имеются у всех пресноводных Mastigophora и Sarcodina и у всех инфузорий.
У Mastigophora большей частью имеется всего 1 сократительная вакуоль, редко 2 (Phytomastigina).
Размеры вакуолярного аппарата в общем прямо пропорциональны размерам тела и обратно пропорциональны количеству вакуолей в теле животного. Так, у крупных инфузорий Heterotricha (Stentor, Spirostomum) имеется всего 1 вакуоль, но зато она (вместе с ее каналами) достигает очень больших размеров. У Trachelius ovum до 30 вакуолей, но зато они очень мелки.
Иногда замечается увеличение числа вакуолей по мере роста животного, как например у Collinia branchiarum и некоторых других Astomata. То умножение числа вакуолей, которое у Collinia (Anoplophrya) происходит в течение индивидуальной жизни, имело, по-видимому, место в течение эволюции всей группы Astomata (рис. 124). Действительно, среди них можно наметить ряд форм с постепенно усложняющимся вакуолярным аппаратом. Так, у Dogielella имеется всего 1 субтерминально расположенная вакуоль, у рода Anoplophrya 1 продольный ряд из небольшого числа вакуолей; у родов Butschliella и Мопоdontophrya число вакуолей в этом ряду сильно увеличивается, а у Radiophrya вакуоли образуют 2 продольных ряда. Наконец, у рода Haptophrya совершается слияние ряда многочисленных вакуолей в общий продольный вакуолярный канал, способ происхождения которого виден из наличия вдоль него целого ряда отдельных выводных отверстий.
У Sarcodina и Mastigophora весь экскреторный аппарат ограничивается самой вакуолью. У немногих жгутиконосцев (например, Euglena) сократительная вакуоль окружена венчиком мелких образовательных вакуолей, соединенных с пей тонкими канальцами.
Паппас и Бранд (Pappas a. Brandt, 1958) думают, что вода в вакуоль у Amoeba поступает вследствие излияния в нее мелких пузырьков, лежащих вокруг, путем слияния мембран этих пузырьков с центральной вакуолью. У инфузорий обычно сократительные вакуоли снабжаются одним или несколькими приводящими каналами, которые пульсируют. У Spirostomum - 1 такой канал, у Stentor - 2, у Frontonia - IO длинных каналов и т. д. Каналы, по-видимому, расходятся в цитоплазме на гораздо более далекое расстояние, чем это видно под микроскопом. По крайней мере, при осмировании Paramecium становится заметным продолжение периферических концов каналов в длинные, тонкие концевыe отделы. Образовательные вакуоли и приводящие каналы собирают жидкость (т. е. главным образом воду) из цитоплазмы и доставляют ее в сократительную вакуоль. Весь вакуолярный аппарат инфузорий залегает в глубине эндоплазмы и не меняет своего положения в теле.
Недавно Шнейдер (Schneider, 1960) опубликовал интересные данные по ультрамикроскопическому строению сократительной вакуоли и приводящих каналов Paramecium caudatum. По данным Д. H. Насонова (1924), приводящие каналы окружены осмиофильной муфтой, а по
наблюдениям Гелея (Gelei, 1925а, 1925b), губчатой плазмой (Nierenplasma или Nephridialplasma).
По исследованиям Шнейдера, этот участок плазмы, окружающий приводящие каналы, состоит из многочисленных тонких и извитых

Рис. 125. Схема строения сократительной вакуоли с приводящим каналом и окружающей его цитоплазмой у Paramecium по данным электронной микроскопии. (По Schneider, 1960). А - радиальный канал в стадии систолы, сократительная вакуоль в стадии диастолы; В - радиальный канал в диастоле, сократительная вакуоль в систоле. AK - выводной канал вакуоли; Amp - ампула периферического канала; EK - канал, соединяющий ампулу с сократительной вакуолью; EPR - каналы эндоплазматической сети; FB - сократительные фибриллы вакуоли; KV - сократительная вакуоль; NK -приводящий канал сократительной вакуоли; NT - сеть "нефридиальных каналов", связанная с каналами эндоплазматической сети; RS - скопление "нефридиальных каналов" в поперечном разрезе.
трубочек с осмиофильными стенками (рис. 125). Внутренний просвет этих трубочек во время систолы равен 100 Å, а во время диастолы 150 Å. Весь слой нефридиальной плазмы имеет толщину 1-1,5 μ. На его периферии трубочки переходят непосредственно в канальцы эндоплазматической сети, которые пронизывают все тело парамеции. Кроме того, нефридиальная плазма окружена еще особыми трубочками с осмиофильными стенками. Они располагаются иногда пучками. Их диаметр достигает 500 Å. Длина этих трубочек около 1 μ.
Приводящий канал имеет осмиофильную оболочку. Во время систолы этот канал становится щелевидным и имеет просвет около 300 Å, в диастоле он расширяется до 4000 Å. Трубочки, окружающие приводящий канал, открываются в него только во время диастолы, когда они увеличиваются в диаметре, тогда как во время систолы эти трубочки замкнуты и не связаны с каналом. Приводящий канал на своем проксимальном конце непосредственно переходит в ампулу, которая через концевой каналец открывается в резервуар вакуоли. В стенке ампулы и концевого канала залегают трубковидные сократимые фибриллы, расположенные несколькими пучками. Со стенок концевого канала они переходят непосредственно на стенку резервуара сократительной вакуоли, распределяясь там также в виде отдельных пучков.
По данным Руджинской (Rudzinska, 1957), в цитоплазме вокруг вакуоли Tokophrya infusionum сосредотачиваются мелкие пузырьки эргастоплазмы и скапливаются митохондрии. Кроме того, вокруг вакуоли обнаруживаются структуры, которые можно сравнить с диктиосомами. Они, по-видимому, участвуют в секреции и накоплении жидкости, поступающей из цитоплазмы.

А. А. Стрелков (1939) нашел вокруг выводного канала у Cycloposthium хорошо заметное кольцевое волокно, представляющее собой, по всей вероятности, мионему, регулирующую выход жидкости через канал наружу. У всех инфузорий экскреторное отверстие сократительной вакуоли представляет собой преформированную пору. Обычно оно округлое или овальное и имеет совершенно постоянное для каждого вида положение среди рядов ресничек. У рода Conchophthirus отверстие имеет вид узкой щели с утолщенным краем. Быть может, такая форма поры у Conchophthirus имеет известное функциональное значение при ее закрывании. А именно, весьма вероятно, что здесь закрывание происходит не посредством кольцевой перетяжки выводного канала, а спадением краев щели наподобие двух губ. Пора открывается в выводной канал, обычно очень короткий, но иногда (у Paramecium trichium) образующий несколько петлевидных изгибов.
Электронномикроскопическое исследование сократительной вакуоли Tokophrya infusionum подтвердило наличие постоянно существующей поры и экскреторного канала, вокруг которого располагаются фибриллы (180 А толщиной), осуществляющие его сжатие (Rudzinska, 1957). Аналогичные фибриллы обнаружены у Paramecium (Schneider, 1960).
2. Функциональное значение сократительных вакуолей
1) Процесс сокращения пульсирующих вакуолей
Вакуоли работают ритмически, попеременно то расширяясь (стадия диастолы) и медленно наполняясь жидкостью, то внезапно сокращаясь (стадия систолы) и выталкивая свое содержимое через выводной канал наружу. Там, где имеются особые приводящие каналы, цикл сокращения всего аппарата усложняется; а именно, он начинается с расширения каналов, происходящего в то время, когда вакуоль находится в состоянии систолы (диастола каналов); за этим следует сокращение каналов (систолы), изливающих свое содержимое в сократительную вакуоль, что вызывает ее систолу; в то же время каналы начинают вновь расширяться. Фактически соответствующие стадии имеются и у вакуолей, лишенных системы каналов, но там они мало заметны или совсем не видны, так как при диастоле вакуоли в нее изливаются находящиеся в окружающей плазме очень мелкие и неправильно расположенные вакуольки (ср. Ophryoscolecidae, рис. 126).
Темп пульсации варьирует у разных простейших, а также изменяется и у одного и того же вида под влиянием изменения факторов внешней среды.
Так, для разных пресноводных Protozoa промежутки между пульсациями при разных температурах показаны в табл. 6.
ТАБЛИЦА 6
Изменение темпа пульсации сократительной вакуоли у инфузорий (в сек.) при разных температурах

Oligochaeta. При этом оказалось, что темп пульсации сократительной вакуоли не является одинаковым у всех Astomata и находится в связи с осмотическими условиями той среды, в которой обитает хозяин. У Mesnilella clavataw. Hoplitophrya secans из пресноводных олигохет пульсация столь же часта, как и у свободноживущих пресноводных инфузорий: темп пульсации не превышает 1 мин. (20-30 сек.).
У Radiophrya из морской олигохеты Enchytraeus sp., обитающей на литорали Баренцева моря, пульсация совершается через 6-8 мин.
Наконец, у Metaradiophrya lumbrici и Anoplophrya lumbrici из кишечника почвенной олигохеты Eisenia foetida и у Mesnilella fastigata и Radiophrya prolijera из почвенных Enchytraeus sp. темп пульсации более медленный (1.5-4 мин.), чем у инфузорий из пресноводных, и несколько более быстрый, чем у Astomata из морских олигохет. Таким образом, у близких видов инфузорий Astomata обнаруживается существенное физиологическое различие вследствие приспособленности к разной в осмотическом отношении среде. Для инфузорий из пресноводных червей изотонической средой является 0.4%-й рингер, тогда как для инфузорий кишечника почвенных червей изотония соответствует 0.75-0.8%-му раствору рингера, а для морских форм - 3%-му раствору.
Влияние изменения количества солей в окружающей среде на темп пульсации вакуоли было продемонстрировано еще Цюльцер (Zuelzer, 1910). Она показала, что Amoeba verrucosa при приучении к жизни в морской воде обнаруживает, начиная с концентрации соли в 0.3%, постепенное замедление пульсации вакуоли, а когда концентрация соли достигает 1.5%, вакуоль совсем исчезает, чтобы возникнуть вновь при постепенном разбавлении культуры пресной водой. Точно так же Херфс (Herfs, 1922) получил для Paramecium, приучаемых к жизни в растворах NaGl и в пресной воде, следующие данные:
| Содержание соли в воде (в %) | 0 | 0.25 | 0.5 | 0.75 | 1 |
| Промежуток между пульсациями (в сек.) | 6.2 | 9.3 | 18.4 | 24.8 | 163 |
| Количество выделяемой в 1 час жидкости, выраженное в объемах тела | 4.8 | 2.82 | 1.38 | 1.08 | 0.16 |
Вообще количество жидкости, выводимой через вакуоли в известный срок, при прочих равных условиях пропорционально темпу пульсации и у разных Protozoa может быть весьма различным. Например, сократительная вакуоль Uronema nigricans при температуре 28° за 2 мин. выделяет объем воды, равный объему тела инфузории, а у Euplotes patella для этого требуется 14 мин., у Paramecium aurelia - 46 мин.
2) Физиологическое значение сократительных вакуолей
Функции вакуолей, по-видимому, многообразны. Прежде всего имеются основания думать, что на долю вакуолей падает выведение из тела накопляющихся в нем конечных продуктов обмена веществ.
Предположения об этой функции высказывались еще в прошлом веке Штейном (Stein, 1878) и другими исследователями, но Хоуланд (Howland, 1924а, 1924b) первая косвенно показала, что простейшими выделяется мочевая кислота. В ее культурах парамеций и амеб имелась мочевая кислота, концентрация которой увеличивалась приблизительно пропорционально сроку содержания культуры. M. Новиков (1908), Шумвей (Shumway, 1917) и Флатер (Flather, 1919) показали, что при скармливании инфузориям щитовидной железы, при действии эпинефрина и экстракта эпифиза пульсация вакуолей учащается, а сами вакуоли увеличиваются, т. е. получается стимуляция экскреции под влиянием тех же средств, которые у позвоночных вызывают диурез.
Везерби (Weatherby, 1927) обнаружил мочевину в жидкости сократительной вакуоли Spirostomum, добытой при помощи микропипетки. Однако, судя по подсчетам этого автора, вакуоль выводит лишь 1 % всей выделяемой массовыми культурами Spirostomum мочевины.
В общем данные о характере азотистых продуктов выделения весьма разноречивы. Помимо мочевой кислоты и мочевины, некоторыми авторами вместо этих веществ в качестве продуктов выделения указывался аммиак (например, у Glaucoma и Spirostomum). He совсем согласуется с экскреторной функцией сократительных вакуолей лишь то обстоятельство, что они никогда не участвуют в выведении наружу интравитальных красок, введенных в плазму, которые очень часто выводятся у высших животных через почки и другие выделительные органы.
Неоднократно высказывалось мнение, что сократительные вакуоли принимают известное участие в дыхательном обмене простейших, содействуя выведению из цитоплазмы угольной кислоты; однако фактических доказательств специфической деятельности вакуоли в этом направлении не имеется. Только косвенные данные Людвига (Ludwig, 1928) о соответствии действительного количества выводимой через вакуоль и предположительного количества выделяемой животным угольной кислоты толкуются этим автором в пользу дыхательной функции вакуоли.
Большинством авторов в настоящее время выдвигается на первый план та важная роль, которую сократительные вакуоли играют в качестве осморегуляторного аппарата. Еще Деген (Degen, 1905) указал, что плазма пресноводных простейших благодаря присутствию в ней солей обладает более высоким осмотическим давлением, чем окружающая среда; а это обстоятельство ведет к постоянному впитыванию цитоплазмой воды. Для удаления непрерывно поступающего в плазму избытка воды и служит сократительная вакуоль. Правильность этого взгляда подтверждается как характером распространения сократительных вакуолей среди простейших, так и опытами приучения пресноводных простейших к воде с возрастающей концентрацией солей (Firiley, 1930; Loefer, 1939, и др.).
Недавно появилось экспериментальное исследование Jl. H. Серавини (1958) над пульсацией вакуоли Paramecium саиdatum, в котором приводятся данные, показывающие, что при переводе инфузорий в солевые растворы повышенной концентрации у них на протяжении первых 15- 30 мин. наблюдается снижение скорости пульсации вакуоли в несколько раз, затем темп пульсации постепенно ускоряется и приближается к скорости, свойственной контрольным инфузориям, но обычно не достигает его уровня. Например, в 0.2% NaCl через 30 мин. темп пульсации был в среднем 40.5 сек., а через 48 час. достиг 13.1 сек., тогда как у контрольных пульсация протекала со скоростью 10.3 сек. То же наблюдалось и в растворах 0.1 % LiCi, 0.5% CaCl 2 , в 1%-й сахарозе, в растворе мочевины. Замедление происходило в 7-15 раз, но через 2 суток
темп пульсации в этих растворах был замедлен всего лишь в 1.5-3 раза. В более низких концентрациях солей наблюдалось полное восстановление темпа пульсации вакуоли. Такие вещества, как HGl, NaOH, GaGl 2 , формалин, в ничтожных концентрациях вызывали слабое замедление пульсации, которая затем быстро приходила к норме. Адреналин в концентрации 0.0005% даже увеличил водообмен, по со временем пульсация также возвратилась к норме. Таким образом, осмотически активные и неактивные вещества вызывают сходные изменения деятельности сократительной вакуоли. При этом наблюдается общая тенденция регуляции водного потока, проходящего через цитоплазму инфузории, путем изменения темпа пульсации вакуоли. Кроме того, происходит приспособление инфузорий к изменяющимся условиям осмотического давления внешней среды. Это же отмечал и Китчинг (Ritching, 1952) в отношении Саrchesium и Podophrya. У этих инфузорий в высоких концентрациях этиленгликоля и сахарозы происходит полная остановка деятельности вакуоли, но через некоторое время пульсация вакуоли и водообмен восстанавливаются.
Все это свидетельствует о сложном механизме действия сократительной вакуоли, играющей несомненную роль в водообмене инфузорий. Серавин считает, однако, на основании того, что осмотически неактивные вещества вызывают задержку пульсации вакуоли, возможным предположить, что водообмен у инфузорий не определяется осмотическими закономерностями и поэтому роль сократительной вакуоли в осморе-гуляции не может считаться доказанной.
Для понимания функции вакуоли необходимо, конечно, понять также и механизм циркуляции воды из цитоплазмы в резервуар вакуоли и механизм сокращения самой вакуоли. И. Гелей и Г. Гелей (Gelei, 1928; Gelei, 1939), как и ряд других исследователей, считают, что только тургорным напряжением, создающимся в цитоплазме, и, следовательно, осмотическими условиями определяется сокращение резервуара вакуоли. Возможно, что имеется и активный механизм секреции воды в вакуоль (Kitching, 1956). По мнению Шнейдера (Schneider, 1950), пульсация вакуоли обусловлена деятельностью сократимых фибрилл, расположенных в стенке вакуоли, а наполнение приводящих каналов водой происходит следующим образом. Жидкость собирается в канальца эргастоплазмы и затем вследствие большего диаметра этих канальцев, нежели трубочек нефридиальной плазмы, создается ток воды из канальцев эргастоплазмы в трубочки нефридиальной плазмы и из них уже в просвет канала. Нужны, конечно, дальнейшие исследования, чтобы более точно разобраться в механизме этих процессов. Экскреторная и дыхательная деятельность сократительной вакуоли кажутся весьма возможными, но не окончательно доказанными.
3) Сократительная вакуоль и аппарат Гольджи
Вопрос о морфологическом значении сократительной вакуоли вступил в новую фазу после работ Д. H. Насонова (1924, 1925), который высказывает мысль о том, что сократительная вакуоль простейших гомологична аппарату Гольджи в клетках Metazoa. Насонов (1924) и почти одновременно с ним И. Гелей (J. Gelei, 1925а, 1925b), применяя технику осмирования к инфузориям, обнаружили важные, новые черты в строении сократительных вакуолей (рис. 127). Насонов нашел, что вокруг сократительной вакуоли многих инфузорий (Campanella, Lionotus, Parameciuin, Nassula), а также у Chilomonas находится сильно чернящаяся от
восстановленного осмия оболочка, с гладкой поверхностью или окруженная слоем мелких, осмиофильных зерен. Оболочка или окружает всю вакуоль целиком (у вышеперечисленных видов), или же имеет вид сетчатого кольца, опоясывающего вакуоль по ее экватору (Chilodon, Dogielella). У форм, обладающих системой приводящих каналов (Paramecium), оболочка продолжается и на последние; при этом каналы могут быть прослежены в эндоплазме гораздо дальше, чем при любом ином методе исследования, и дистальные части их окружены сетчатой муфтой серого цвета. Этот отдел каналов Насонов считает собственно экскреторной частью аппарата, тогда как проксимальную часть каналов и саму вакуоль лишь выводной частью вакуолярной системы. У Dogielella Насонов мог констатировать, что во время систолы вакуоли окружающее ее черное кольцо не следует за последней при ее сокращении, но остается на своем прежнем месте. Гелей (Gelei, 1925a, 1925b) пробовал сравнивать их с нефридиальной системой Metazoa, приравнивая саму сократительную вакуоль мочевому пузырю, другие части аппарата - иным разделам нефридиев. Насонов остановился на сходстве осмирующейся оболочки вакуолей с аппаратом Гольджи у Metazoa. Действительно, приводимые им рисунки разных типов сократительных вакуолей и аппаратов Гольджи, взятых из разных типов клеток у Metazoa, чрезвычайно убедительны по своему сходству.

На основании этого Насонов считает возможным искать в осмиофильной губчатой оболочке приводящих каналов у Parameciurn и сетчатого кольца у Dogitlella не только морфологического, но и функционального параллелизма между этими образованиями и аппаратом Гольджи. Сетчатость вещества кольца зависит от наличия в нем маленьких образовательных вакуолек - стадия связанного секрета. Эти вакуольки затем, сливаясь, образуют центральную вакуоль, лежащую в центре кольца - стадия свободного секрета, выделяемого наружу. По концепции Насонова, аппарат Гольджи служит для собирания из плазмы известных веществ, которые подготовляются в нем для своего выведения вон из тела через вакуоль. Вся фактическая сторона находок Насонова вполне подтвердилась позднейшими исследованиями ряда авторов: А. А. Стрелкова (1939) у Cycloposlhiidae, Форе-Фремье (Fauré-Fremiet, 1925) у Vorlicella, Кинг (King, 1928) у Paramecium trichium и Euplotes и др. Правда, у некоторых простейших не удавалось обнаружить вокруг вакуоли особой осмиофильной оболочки, что могло зависеть от методических моментов.
в клетках всех Metazoa в виде сетчатого образования или состоящего из отдельных диктиосом. Поэтому можно думать, что он имеется также и у всех одноклеточных организмов. Действительно, исследования Хиршлер (Hirschler, 1927), Овас (Hovasse, 1937), Жуае-Лавернь (Joyet-Lavererie, 1926) и других показали наличие в плазме многих простейших (Gregarina, Coccidia, Flagellata, Sarcodina) мелких палочек, колечек, шариков с сильно осмиофильным краем и весьма сходных с диктиосомами клеток Metazoa.
Признаками аппарата Гольджи у Metazoa является прежде всего способность этого аппарата специфически восстанавливать некоторые соединения тяжелых металлов - серебра и осмия, что выражается в почернении структур аппарата. Эти структуры не окрашиваются прижизненно нейтральным красным и метиленовым синим. Нередко характерными структурами аппарата Гольджи являются диктиосомы, которые состоят из двух частей - хромофильной и хромофобной. Обычно секреторные гранулы погружены в хромофобную часть. Как правило, аппарат Гольджи локализуется именно в секреторной зоне клетки.
Исследования с электронным микроскопом показали единство структур аппарата Гольджи в разных клетках Metazoa. Bo всех исследованных клетках выявлены двойные мембраны (или γ-цитомембраны), расположенные параллельно друг другу и образующие систему из 3-7 и более пар. Каждая пара ограничивает более светлое пространство шириной 50-200 Å и образует нечто вроде плоского мешочка. Иногда оно расширяется до большой вакуоли. Мембраны погружены в мелкозернистое основное вещество. В ряде случаев мембраны могут окружать разной величины вакуоли. Все эти структуры являются в достаточной степени стабильными.
Существенным является тот факт, что имеется полное соответствие между картинами аппарата Гольджи, описанного с помощью светового микроскопа Насоновым π другими исследователями в клетках Metazoa, и современными изображениями, полученными с электронным микроскопом.
Дальтон и Феликс (Dalton a. Felix, 1957) отмечают также, что сократительная вакуоль Chlamidomonas ограничена мембранами, сходными с таковыми аппарата Гольджи клеток Metazoa, и, таким образом, авторы поддерживают гипотезу Насонова о гомологии этих органелл. Вокруг сократительной вакуоли у инфузорий Campanella и Ophridium располагаются многочисленные трубочки с плотными осмиофильными стенками, которые могут быть до некоторой степени сравнимы с ультраструктурами аппарата Гольджи (Fauré-Fremiet et Rouiller, 1959). Приведенные выше данные Шнейдера о строении нефридиальной плазмы вокруг приводящих каналов сократительной вакуоли Parameciiim делают, однако, затруднительным гомологизирование ее структуры с ультраструктурой диктиосом.
Еще раньше французские авторы (Duboscq et Grassé, 1933; Grassé et Hollande, 1941) также отвергли гомологию осмиофильных оболочек сократительных вакуолей с аппаратом Гольджи. Они считают, что осмиофильные стенки вакуолей, как и сами сократительные вакуоли, являются совершенно независимыми от аппарата Гольджи образованиями.
Необходимо отметить, что у некоторых инфузорий обнаружены серповидные или кольцевидные диктиосомы, как например у Balantidium elongatum, вне связи, с вакуолями (Villeneuve-Brachori, 1940). Нуаро-Тимоте (Noirot-Timothee, 1957) выявила с электронным микроскопом типичные структуры аппарата Гольджи в эндоплазме некоторых Ophryoscolecidae, не связанные с сократительной вакуолью.
Французские авторы гомологизируют парабазальное тело с аппаратом Гольджи (см. стр. 115), так как это образование особенно при исследовании с электронным микроскопом оказалось сходным с диктиосомами клеток Metazoa (рис. 80).
Нужны несомненно дальнейшие сравнительные исследования простейших, чтобы решить вопрос о гомологии тех или иных структур с аппаратом Гольджи клеток Metazoa.
Данная статья ознакомит читателя со строением простейших организмов, а именно - акцентирует внимание на строении сократительной вакуоли, выполняющей выделительную (и не только) функцию, расскажет о значении простейших и опишет способы их существования в окружающей среде.
Сократительная вакуоль. Понятие
Вакуоль (от франц. vacuole, от латинского слова vacuus - пустой), шаровидной формы небольшие полости в растительных и животных клетках или одноклеточных организмах. Сократительные вакуоли в первую очередь распространены среди простейших организмов, которые обитают в пресноводной воде, например, среди протистов, таких как амеба протей и инфузория туфелька, которая получила такое оригинальное название из-за формы тела, схожего с формой подошвы туфли. Помимо перечисленных простейших, идентичные структуры также были обнаружены и в клетках различных пресноводных губок, которые принадлежат к семейству Бадяговые.

Строение сократительной вакуоли. Ее особенности
Сократительная вакуоль является мембранным органоидом, который осуществляет выброс лишней жидкости из цитоплазмы. Локализация и строение этого аппарата варьируется у различных микроорганизмов. Из комплекса пузыревидных или трубчатых вакуолей, называемых спонгиями, жидкость попадает в сократительную вакуоль. Благодаря постоянной работе этой системы поддерживается стабильный объём клетки. У простейших имеются сократительные вакуоли, которые представляют собой аппарат, регулирующий осмотическое давление, а также служащий для выделения из организма продуктов распада. Тело простейших состоит всего лишь из одной клетки, которая, в свою очередь, осуществляет все необходимые жизненные функции. Представители этого подцарства, такие как инфузория туфелька, амеба обыкновенная, другие одноклеточные обладают всеми свойствами самостоятельного организма.
Роль простейших организмов
Клетка выполняет все жизненные функции: выделение, дыхание, раздражимость, движение, размножение, обмен веществ. Простейшие распространены повсеместно. Наибольшее количество видов обитает в морских и пресных водах, многие населяют влажную почву, могут поражать растения, жить в телах многоклеточных животных и человека. В природе простейшие выполняют санитарную роль, также они участвуют в круговороте веществ, являются пищей для многих животных.

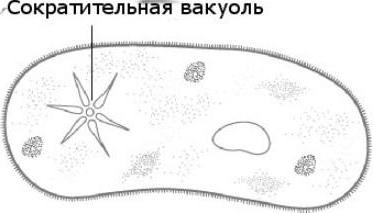
Сократительная вакуоль у амебы обыкновенной
Амеба обыкновенная - представитель класса корненожки, не имеет в отличие от других представителей постоянной формы тела. Передвижение осуществляет с помощью ложноножек. Теперь разберемся с тем, какую функцию выполняет сократительная вакуоль у амебы. Это регуляция уровня осмотического давления внутри ее клетки. Она у может образоваться в любом участке клетки. Через наружную мембрану вода из окружающей среды поступает внутрь осмотически. Концентрация растворенных веществ в клетке амебы выше, чем в окружающей среде. Таким образом, создается разность давления внутри клетки простейшего и за ее пределами. Функции сократительной вакуоли у амебы - это своеобразный откачивающий аппарат, который выводит избыток воды из клетки простейшего организма. Выбрасывать в окружающую среду накопившуюся жидкость амеба протей может в любом участке поверхности тела.
Помимо осморегуляторной, выполняет функцию дыхания в жизнедеятельности, так как в результате осмоса поступающая вода доставляет растворенный в ней кислород. Какую же еще функцию выполняет сократительная вакуоль? Так же выполняет выделительную функцию, а именно вместе с водой выводятся продукты обмена веществ в окружающую их среду.

Дыхание, выделение, осморегуляция у инфузории туфельки
Тело простейших покрыто плотной оболочкой, которое имеет постоянную форму. так и водорослями, в том числе и некоторыми простейшими. Организм инфузории имеет более сложное, чем у амебы строение. В клетке туфельки спереди и сзади расположены две сократительные вакуоли. В этом аппарате различимы резервуар и несколько небольших канальцев. Сократительные вакуоли постоянно находятся, благодаря такому строению (из микротрубочек), на постоянном месте в клетке.
Главная функция сократительной вакуоли в жизнедеятельности данного представителя простейших - осморегуляция, также она выводит из клетки лишнюю воду, которое проникает в клетку за счет осмоса. Сначала происходит набухание приводящих каналов, потом вода из них перекачивается в специальный резервуар. Резервуар сокращается, отделяется от приводящих каналов, через поры вода выбрасывается наружу. В клетке инфузории находится две сократительные вакуоли, которые, в свою очередь, действуют в противофазе. За счет работы двух таких аппаратов обеспечивается непрерывный процесс. Помимо этого, вода непрерывно циркулирует благодаря деятельности сократительных вакуолей. Они сжимаются поочередно, и частота сокращений зависит от температуры окружающей среды.
Так, при комнатной температуре (+18 - +20 градусов по Цельсию) частота сокращений вакуолей составляет, по некоторым данным, 10-15 секунд. А учитывая, что естественной средой обитания туфельки являются любые пресные водоемы со стоячей водой и наличием в ней разлагающихся органических веществ, температура этой среды на несколько градусов меняется в зависимости от времени года и, следовательно, частота сокращений может достигать 20-25 секунд. За час сократительная вакуоль простейшего организма способна выбросить из клетки воду в количестве. соизмеримом с ее размерами. В них скапливаются питательные вещества, непереваренные остатки пищи, конечные продукты обмена веществ, также можно обнаружить кислород и азот.
Очистка сточных вод простейшими
Влияние простейших на круговорот веществ в природе имеет огромное значение. В водоемах, вследствие спуска сточных вод, размножаются в большом количестве бактерии. В результате появляются различные простейшие организмы, которые и используют в пищу эти бактерии и таким образом способствуют естественной

Заключение
Несмотря на простое строение этих одноклеточных организмов, тело которых состоит из одной клетки, но выполняет функции целого организма, удивительным образом приспособленного к окружающей среде. Это можно наблюдать даже на примере строения сократительной вакуоли. На сегодняшний день уже доказано огромное значение простейших в природе и участие их в круговороте веществ.