1) Покровные ткани
Покровные ткани располагаются на поверхности органов растений. Они состоят из плотно сомкнутых клеток и защищают внутренние части растения от неблагоприятных внешних воздействий. Придает прочность. В соответствии с происхождением из различных меристем выделяют первичные и вторичные покровные ткани:
- Первичные
Ризодерма (эпиблема) – первичная однослойная поверхностная ткань корня. Основная функция ризодермы – всасывание, избирательное поглощение из почвы воды с растворенными в ней элементами минерального питания. Клетки ризодермы тонкостенные, с вязкой цитоплазмой и большим количеством митохондрий. Характерной особенностью ризодермы является образование у части клеток корневых волосков – трубчатых выростов. Корневые волоски увеличивают поглощающую поверхность ризодермы в десять и более раз.
Эпидерма - она покрывает листья, стебли травянистых и молодых побегов древесных растений, цветки, плоды и семена. Основная функция эпидермы – регуляция газообмена и транспирации (испарения воды живыми тканями). Она препятствует проникновению внутрь растения болезнетворных организмов. Эпидерма может функционировать как всасывающая ткань. Она принимает участие в синтезе различных веществ, в восприятии раздражений, в движении листьев.
Состав эпидермы:
Основные клетки эпидермы – живые клетки таблитчатой формы. Клетки плотно сомкнуты, межклетники отсутствуют. Боковые стенки часто извилистые, что повышает прочность их сцепления, реже прямые.
Их внутренний, более мощный, слой состоит из целлюлозы и пектиновых веществ; наружный слой подвергается кутинизации. Поверх наружных стенок выделяется сплошной слой кутина, образующий защитную пленку – кутикулу .
Устьица
– образования для регуляции транспирации и газообмена. Предохраняет от засушливости. Устьице состоит из двух замыкающих
клеток
бобовидной формы, между которыми находится устьичная
щель
, которая может расширяться и сужаться. Эти клетки часто образуют волоски, которые превращаются в шипы.
Чечеви́чки
- образования в виде мелких бугорков, служащие для газообмена.
- Вторичные
Перидерма –покрывает корни вторичного строения и стебли многолетних побегов. Она может возникнуть и в результате залечивания поврежденных тканей раневой меристемой.
Перидерма состоит из:
1)Пробка состоит из нескольких слоев таблитчатых клеток, расположенных плотно, без межклетников. Клетки пробки мертвые, они не имеют протопласта и заполнены воздухом. В полости клеток могут также откладываться вещества (суберин) , повышающие защитные свойства пробки. Главная функция пробки – защита от потери влаги.
2)Пробковый камбий – откладывает клетки пробки наружу и клетки пробковой паренхимы внутрь органа. Пробковая паренхима состоит из живых паренхимных клеток. Однако часто пробковый камбий работает односторонне, откладывая только пробку, а пробковая паренхима остается однослойной.
3) Корка . Возникает в результате многократного заложения новых прослоек перидермы в более глубоких слоях коры. Таким образом, корка состоит из чередующихся слоев пробки и прочих отмерших тканей коры. Толстая корка надежно предохраняет стволы деревьев от механических повреждений и т.д.
Применение: пища животных.
2) Опыление - перенос пыльцы с тычинок на рыльце пестика.
- Ксеногамия - перекрёстное опыление, при котором цветки одного растения опыляются пыльцой цветков других растений того же вида.
Приспособление для перекрестного опыления:
1) Дихогамия - неодновременное созревание в цветках пыльников и рылец.
2) Гетеростилия - неодинаковая длина столбиков цветков на разных растениях одного и того же вида
- Клейстога́мия - тип самоопыления , при котором опыление происходит в закрытых цветках .
- Автогамия - самоопыление, при котором пыльца попадает на рыльце того же цветка.
Особенность самоопыления:
1) тычинки и пестики созревают одновременно
3) пыльники выше пестика
3) передняя пыльца крупная, тяжелая
3) Покрытосеменные образуют один из наиболее крупных отделов царства растений. Они составляют основную часть растительной массы в биосфере. Покрытосеменные растения - это дуб, береза, яблоня, пшеница, рожь, капуста, пальма, подорожник и др. Многие виды покрытосеменных вошли в число культурных растений.
Представители покрытосеменных произрастают повсеместно. Одни живут очень недолго - несколько дней. Например, эфемеры крупка весенняя, проломник Турчанинова живут 35-60 дней и дают семена. Другие живут сотни лет. Например, платан восточный, или чинар, живет до 2000 лет, достигает 50 м в высоту, а его ствол - около 18 м в окружности.
У растений, относящихся к этому отделу, семена покрыты тканями плода, который образуется из завязи пестика цветка. Благодаря этим особенностям отдел получил название Покрытосеменные или Цветковые. Покрытосеменные (цветковые) растения чрезвычайно разнообразны по форме, по требованиям к условиям обитания, но всем им свойственны общие признаки строения, размножения и развития.
Отличие двуд от однод:
Двудольные: стержневая, листья разнообразные, открытые провод пучки, пятичленный цветок. чаще сетчатое жилковаение. семядоли (в них пит в-ве)
однод: простые,линейные,паралельные жилко,без прилистников. влагалищ способ прикрепл, закр провод пучки, трехчленный цветок. в эндосперме пит в-ва.
Покровные ткани первичного тела растений .
I. Эпидерма . Листья и молодые зеленые побеги, как чехлом, покрыты однослойной первичной покровной тканью - эпидермой. Изредка эпидерма многослойна. Такая многослойная эпидерма обнаружена в листьях известного комнатного растения фикуса (Ficus elastica) . Эпидерма возникает из первичной меристемы - протодермы . Это сложная ткань, поскольку ее клетки различаются по форме и отчасти по функциям. В частности, резко отличаются клетки, образующие устьица, и клетки трихом. Наружная поверхность клеток эпидермы часто покрыта слоем кутикулы или, реже, восковым налетом различной толщины. Кутикула может достигать значительной толщины, особенно у растений засушливых мест обитания. Нередко ее поверхность покрыта различного рода складками или бородавчатыми выростами. Исключая устьичные щели, клетки эпидермы плотно сомкнуты, т. е. отсутствуют межклетники. Главная функция эпидермы - регуляция газообмена и транспирации, т. е. испарения воды растением. Газообмен и транспирация осуществляются преимущественно через устьица, но частично и через кутикулу. Кроме того, через поры и тяжи пектиновых веществ в наружных стенках клеточных оболочек эпидермы проникают вода и неорганические питательные вещества, что особенно характерно для водных растений. Иногда эпидерма выполняет необычные для этой ткани функции - такие, как фотосинтез (у части водных растений), запасание воды (у некоторых пустынных растений) или секрецию веществ вторичного метаболизма (ряд эфирномасличных).
Характер клеток эпидермы различен, большинство, получившее название основных клеток эпидермы, отличается разнообразием очертаний. Боковые стенки, как правило, извилисты, что повышает плотность их сцепления друг с другом, реже прямые. Эпидермальные клетки осевых органов и листьев многих однодольных сильно вытянуты вдоль оси органа (рис. 36).
В основных клетках эпидермы обнаруживается тонкий постенный слой протопласта с мелкими редкими лейкопластами и ядром.
Часто всю полость эпидермальной клетки занимает одна крупная вакуоль. Клеточный сок ее бесцветен, но иногда, особенно в эпидерме цветков и плодов, он окрашен. Стенки эпидермальных клеток утолщены неравномерно. Обычно наиболее утолщена наружная стенка, а боковые и внутренние - тонкие. Иногда в клетках эпидермы встречаются кристаллы, клетки многих злаков пропитаны кремнеземом. Клетки эпидермы многих семян содержат полисахариды в виде слизи, которая при увлажнении набухает. Семена при этом легко приклеиваются к движущимся предметам и таким образом распространяются.
У некоторых растений под эпидермой расположена особая ткань - гиподерма . Она отчасти выполняет механическую функцию,
отчасти предохраняет растение от избыточного испарения. Хорошо развитая гиподерма заметна в своеобразных игольчатых листьях - хвоинках сосны.
Устьица представляют собой высокоспециализированные образования эпидермы, состоящие из двух замыкающих клеток, между которыми имеется своеобразный межклетник, или устьичная щель (рис. 37). Щель может расширяться и сужаться, регулируя

транспирацию и газообмен. Под щелью располагается дыхательная, или воздушная, полость, окруженная клетками мякоти листа. Клетки эпидермы, примыкающие к замыкающим, получили название побочных, или околоустьичных. Они участвуют в движении замыкающих клеток. Замыкающие и побочные клетки образуют устьичный аппарат . Число побочных клеток и их расположение относительно устьичной щели позволяют выделять ряд устьичных типов. Их изучением занимается стоматография . Данные стоматографии нередко используются в систематике растений для уточнения систематического положения таксонов. Наиболее часто встречающиеся устьичные типы показаны на рисунке 38.
Аномоцитный тип устьичного аппарата обычен для всех групп высших растений, исключая хвойные. Побочные клетки в этом случае не отличаются от остальных клеток эпидермы. Диацитный тип характеризуется только двумя побочными клетками, общая стенка которых перпендикулярна устьичной щели. Этот тип обнаружен у некоторых цветковых, в частности у большинства губоцветных и гвоздичных. При парацитном типе побочные клетки располагаются параллельно замыкающим и устьичной щели. Он найден у папоротников, хвощей и ряда цветковых растений. Анизоцитный тип обнаружен только у цветковых растений. Здесь замыкающие клетки окружены тремя побочными, одна из которых заметно крупнее или мельче остальных. Тетрацитным типом устьичного аппарата характеризуются преимущественно однодольные. При энциклоцитном типе побочные клетки образуют узкое кольцо вокруг замыкающих клеток. Подобная структура найдена у папоротников, голосеменных и ряда цветковых. Расположение замыкающих клеток
относительно прочих клеток эпидермы у разных видов неодинаково. В одних случаях замыкающие клетки находятся на одном уровне с энидермальными, иногда выступают над ними или, напротив, залегают значительно глубже (погруженные устьица) . Последнее наблюдается у растений, приспособленных к засушливым условиям. Иногда углубления, в которых располагаются устьица, выстланы или прикрыты волосками. Называются они устьичными криптами .
Число и распределение устьиц на листе или побеге варьируют в зависимости от вида растений и условий жизни. Число их обычно колеблется от нескольких десятков до нескольких сотен на 1 мм 2 поверхности.
Механизм движения замыкающих клеток весьма сложен и неодинаков у разных видов. У большинства растений при недостаточном водоснабжении в ночные часы, а иногда и днем тургор в замыкающих клетках понижается и щель замыкается, снижая тем самым уровень транспирации. С повышением тургора устьица открываются. Считают, что главная роль в этих изменениях принадлежит ионам калия. Существенное значение в регуляции тургора имеет присутствие в замыкающих клетках хлоропластов. Первичный крахмал хлоропластов, превращаясь в сахар, повышает концентрацию клеточного сока. Это способствует притоку воды из соседних клеток и переходу замыкающих клеток в упругое состояние.
Общая площадь устьичных отверстий составляет лишь 1-2% площади листа. Несмотря на это, транспирация при открытых устьичных щелях достигает 50-70% испарения, равного по площади открытой водной поверхности.
Трихомы у растений - это различные по форме, строению и функциям выросты клеток эпидермы - волоски, чешуйки, железки, нектарники. Размеры трихом варьируют в значительных пределах. Наиболее длинные трихомы (до 5-6 см) покрывают семена хлопчатника. Трихомы могут быть живыми и отмершими и выполнять различные функции. Они делятся на кроющие и железистые. Кроющие - это одноклеточные, многоклеточные, ветвистые и звездчатые волоски. Железистые трихомы (железки и нектарники) являются элементами секреторных тканей. Разнообразие кроющих трихом довольно велико (рис. 39). Их строение и форму иногда используют в систематике. Кроющие волоски образуют на растении различной густоты опушение, предохраняющее от избыточной транспирации или изредка, напротив, ускоряющее ее. На листьях они чаще возникают с той стороны, где имеются устьица. Обильное опушение многих пустынных растений способствует отражению мощной солнечной радиации. Многие эпифиты тропиков используют трихомы для поглощения воды и минеральных солей.
Помимо волосков на эпидерме ряда видов заметны выросты, называемые эмергенцами . К ним относятся известные жгучие волоски крапивы, шипы розы, малины, ежевики и др. Шипы на плодах многих зонтичных, дурмана, каштана также являются эмергенцами. В формировании эмергенцев принимают участие не только клетки эпидермы, но и слои клеток, лежащие под ней.
II. Эпиблема . Эпиблема, нередко называемая также ризодермой, - первичная однослойная покровная ткань корня. Она возникает из наружных клеток апикальной меристемы этого органа вблизи корневого чехлика и покрывает молодые корневые окончания. Эпиблема - одна из важнейших тканей растения, поскольку именно через нее происходит поглощение воды и минеральных солей из почвы.
В зоне всасывания корня эпиблема пассивно или активно
поглощает элементы минерального питания, затрачивая в последнем случае энергию. В связи с этим эпиблема богата митохондриями. Она недолговечна и, отмирая, передает свои функции новым участкам эпиблемы растущего корня. Особенности клеток эпиблемы соответствуют основной функции ткани. Они тонкостенны, лишены кутикулы и имеют более вязкую цитоплазму. В ней отсутствуют устьица. Каждая клетка эпиблемы потенциально способна к образованию корневого волоска , но чаще корневые волоски формируются лишь из части клеток, получивших специальное название трихобластов . Корневые волоски обычно одноклеточные, развиваются в результате выпячивания наружной стенки трихобласта и достигают в длину 1-2 мм. Обычно они существуют в течение нескольких дней, а затем отмирают.
Первичной покровной тканью листьев и стеблей является кожица (эпидерма). Коленца -- живая ткань, клетки ее содержат тонкий постенный слой протоплазмы с ядром и пластидами (чаще всего с лейкопластами) и крупную центральную вакуолю.
Кожица состоит из плотно сомкнутых клеток, имеющих в плане обычно извилистые очертания. На поперечных разрезах они 4--5-угольны, причем наружная стенка слегка выпукла. Клетки кожицы органов, вытянутых в длину (листовых черешков, листовых пластинок линейного очертания, стеблей), в большинстве удлинены в направлении, параллельном продольной оси органа. В каждой клетке кожицы обычно наиболее толста наружная стенка. Боковые и внутренние стенки более тонки и снабжены порами. Оболочка состоит в основном из целлюлозы. У многих растений (особенно у злаков, осоковых, хвощей) наружная стенка инкрустируется кремнеземом. Поверхность кожицы покрыта пленкой -- кутикулой (надкожицей), состоящей из кутана. На поперечных разрезах через кожицу нередко видны отроги кутикулы, внедряющиеся в боковые стенки клеток кожицы, и более мелкие отроги, входящие в целлюлозную толщу наружных стенок.
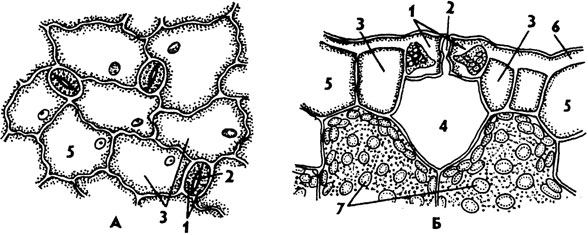
1 -- верхняя кожица. 2 -- кутикула. 3 -- слой в оболочках клеток кожицы с кутикулярными пробойками, 4 - целлюлозный слой, 5 -- клеточное ядро, 6 -- ямки в кожице («наружная дыхательная полость»)
Эпидермис листа - это кожица или покровная ткань листа. Эпидермис состоит из одного слоя уплощенных клеток, плотно прилегающих друг к другу. В каждой из клеток основное пространство внутри занимает центральная вакуоль с клеточным соком. Все содержащиеся в клетке органоиды и ядро оттеснены к оболочке вакуолью. Однако ядро, являющееся хранителем всей наследственной информации, четко определяется в каждой клетке. Основные клетки кожицы листа лишены хлоропластов. Клетки, имеющие отличную от основных форму и расположенные попарно, формируют устьица. Любое устьице имеет характерное строение: состоит из двух смыкающихся клеток, имеющих бобовидную форму, а между этими клетками остается щель, напоминающая своим видом линзу. Это межклеточное пространство называется устьичной щелью. Размер и форма устьичной щели не постоянны. Ее изменчивость обусловлена плотностью прилегания друг к другу замыкающих устьичных клеток. Строение замыкающих устьичных клеток отличается от строения основных. Основное отличие состоит в том, что в устьичных клетках имеются хлоропласты, в которых происходит фотосинтез. Также в них хорошо различимо ядро, как и в других клетках эпидермиса листа. Устьичные щели открываются для доступа нужного для фотосинтеза и дыхания воздуха внутрь листа.
Устьица представляют собой высокоспециализированные образования эпидермы, состоящие из двух замыкающих клеток, между которыми имеется своеобразный межклетник, или устьичная щель. Щель может расширяться и сужаться, регулируя транспирацию и газообмен. Под щелью располагается дыхательная, или воздушная, полость, окруженная клетками мякоти листа. Клетки эпидермы, примыкающие к замыкающим, получили название побочных, или околоустьичных. Они участвуют в движении замыкающих клеток. Замыкающие и побочные клетки образуют устьичный аппарат.
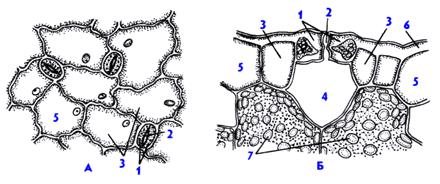
1 - замыкающие клетки,
2 - устьичная щель,
3 - побочные клетки,
4 - дыхательная полость,
5 - эпидермальные клетки,
6 - кутикула
А - вид на эпидерму сверху, Б - поперечный разрез устьичного аппарата
Общая площадь устьичных отверстий составляет лишь 1-2% площади листа. Несмотря на это, транспирация при открытых устьичных щелях достигает 50-70% испарения, равного по площади открытой водной поверхности.
Эпиблема (от греческого epiblema -- покрывало, покрытие), ризодерма, волосконосный слой, первичная поверхностная однослойная всасывающая ткань корня. Формируется вблизи конуса нарастания растущего корня из дерматогена. Клетки эпиблемы (трихобласты) образуют выросты -- корневые волоски, которые во много раз увеличивают поглощающую поверхность корня и закрепляют растение в почве, а также выделяют ряд кислот, действующих на вещества субстрата. Эпиблема -- первая барьерная ткань корня, избирательно поглощающая вещества почвы. Она недолговечна и, отмирая, передаёт свои функции новым участкам эпиблемы растущего корня.
В зоне всасывания корня эпиблема пассивно или активно поглощает элементы минерального питания, затрачивая в последнем случае энергию. В связи с этим эпиблема богата митохондриями. Она недолговечна и, отмирая, передает свои функции новым участкам эпиблемы растущего корня.
Особенности клеток эпиблемы соответствуют основной функции ткани. Они тонкостенны, лишены кутикулы и имеют более вязкую цитоплазму. В ней отсутствуют устьица. Каждая клетка эпиблемы потенциально способна к образованию корневого волоска.
При свободном росте - при развитии корней в воде или во влажном воздухе - волоски имеют форму цилиндра или конуса с закруглением на конце. Между твердыми частицами почвы волосок становится искривленным и кое-где сплющенным или вздутым, что бы извлекать из почвы воду и минеральные вещества.
Корневые волоски играют и механическую роль, давая опору верхушке корня, пробивающейся при росте между частицами почвы, и способствуя заякориванию
1.doc
^3.4. Покровные ткани
Покровные ткани располагаются на поверхности органов растений на границе с внешней средой. Они состоят из плотно сомкнутых клеток и защищают внутренние части растения от неблагоприятных внешних воздействий, излишнего испарения и иссушения, резкой перемены температуры, проникновения микроорганизмов, служат для газообмена и транспирации. В соответствии с происхождением из различных меристем выделяют первичные и вторичные покровные ткани.
К первичным покровным тканям относят: 1) ризодерму , или эпиблему и 2) эпидерму .
Ризодерма (эпиблема) – первичная однослойная поверхностная ткань корня. Образуется из протодермы – наружного слоя клеток апикальной меристемы корня. Основная функция ризодермы – всасывание, избирательное поглощение из почвы воды с растворенными в ней элементами минерального питания. Через ризодерму происходит выделение веществ, действующих на субстрат и преобразующих его. Клетки ризодермы тонкостенные, с вязкой цитоплазмой и большим количеством митохондрий (минеральные ионы поглощаются активно, с затратой энергии, против градиента концентрации). Характерной особенностью ризодермы является образование у части клеток корневых волосков – трубчатых выростов, в отличие от трихомов не отделенных стенкой от материнской клетки (рис. 3.4). Корневые волоски увеличивают поглощающую поверхность ризодермы в десять и более раз. Волоски имеют длину 1-2 (3) мм. Ризодерму часто рассматривают как всасывающую ткань.
Рис. 3.4. Кончик корня ожики многоцветковой: 1 – корневой волосок.
Эпидерма - первичная покровная ткань, образующаяся из протодермы конуса нарастания побега. Она покрывает листья, стебли травянистых и молодых побегов древесных растений, цветки, плоды и семена. Основная функция эпидермы – регуляция газообмена и транспирации (испарения воды живыми тканями). Кроме того, эпидерма выполняет целый ряд других функций. Она препятствует проникновению внутрь растения болезнетворных организмов, защищает внутренние ткани от механических повреждений и придает органам прочность. Через эпидерму могут выделяться наружу эфирные масла, вода, соли. Эпидерма может функционировать как всасывающая ткань. Она принимает участие в синтезе различных веществ, в восприятии раздражений, в движении листьев.
Эпидерма - сложная ткань, в ее состав входят морфологически различные типы клеток: 1) основные клетки эпидермы ; 2) замыкающие и побочные клетки устьиц ; 3) трихомы .
Основные клетки эпидермы
– живые клетки таблитчатой формы. Вид клеток с поверхности различен (рис. 3.5
). Клетки плотно сомкнуты, межклетники отсутствуют. Боковые стенки (перпендикулярные поверхности органа) часто извилистые, что повышает прочность их сцепления, реже прямые. Эпидермальные клетки осевых органов и листьев многих однодольных сильно вытянуты вдоль оси органа. 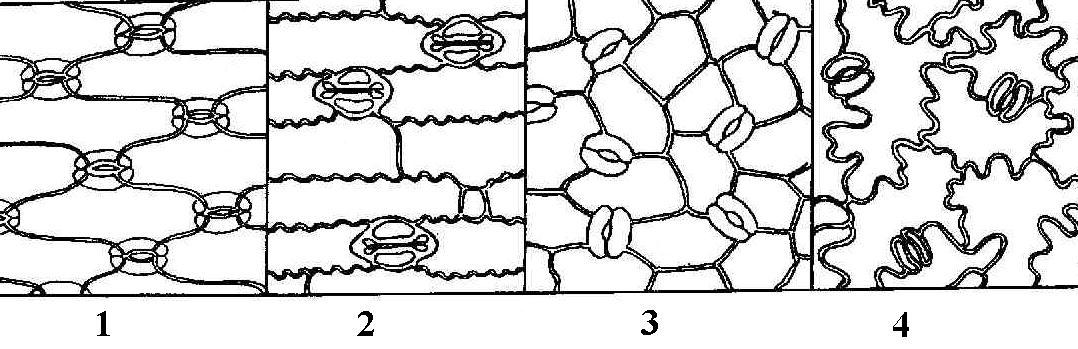
Рис. 3.5. Эпидерма листа различных растений (вид с поверхности): 1 - ирис; 2 - кукуруза; 3 – арбуз; 4 - буквица.
Наружные стенки клеток обычно толще остальных. Их внутренний, более мощный, слой состоит из целлюлозы и пектиновых веществ; наружный слой подвергается кутинизации. Поверх наружных стенок выделяется сплошной слой кутина, образующий защитную пленку – кутикулу . Помимо кутина в ее состав входят вкрапления воска, что еще больше снижает проницаемость кутикулы для воды и для газов. Воск может откладываться в кристаллической форме и на поверхности кутикулы в виде чешуек, палочек, трубочек и других структур, видимых только в электронный микроскоп. Этот сизый, легко стирающийся налет хорошо заметен на листьях капусты, плодах сливы, винограда. Мощность кутикулы, распределение в ней восков и кутина определяют химическую стойкость и проницаемость эпидермы для газов и растворов. В условиях засушливого климата у растений развивается более толстая кутикула. У растений, погруженных в воду, кутикула отсутствует.
Клетки эпидермы имеют живой протопласт, обычно с хорошо развитой эндоплазматической сетью и аппаратом Гольджи. У большинства видов растений в цитоплазме присутствуют лейкопласты. У водных растений, папоротников, обитателей тенистых мест (гибискус) встречаются редкие хлоропласты. Эпидерма чаще всего состоит из одного слоя клеток. Редко встречается двух- или многослойная эпидерма, преимущественно у тропических растений, живущих в условиях непостоянной обеспеченности водой (бегонии, пеперомии, фикусы). Нижние слои многослойной эпидермы функционируют как водозапасающая ткань. У некоторых растений клеточные стенки могут пропитываться кремнеземом (хвощи, злаки, осоки) или содержать слизи (семена льна, айвы, подорожников).
Устьица – образования для регуляции транспирации и газообмена. Устьице состоит из двух замыкающих клеток бобовидной формы, между которыми находится устьичная щель , которая может расширяться и сужаться. Под щелью располагается крупный межклетник – подустьичная полость . Клетки эпидермы, примыкающие к замыкающим клеткам, часто отличаются от остальных клеток, и тогда их называют побочными , или околоустьичными клетками (рис. 3.6 ). Они участвуют в движении замыкающих клеток.
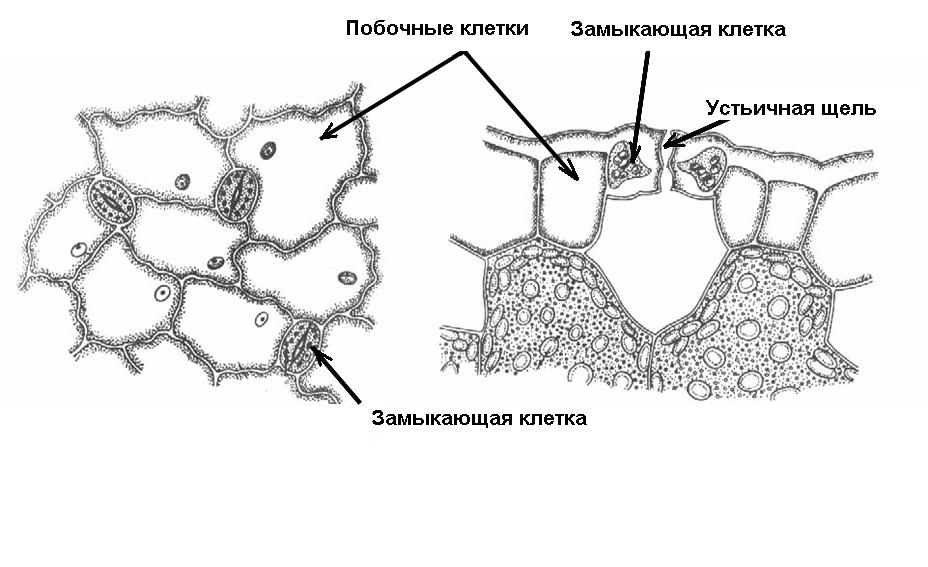
Рис. 3.6. Схема строения устьица.
Замыкающие и побочные клетки образуют устьичный аппарат . В зависимости от числа побочных клеток и их расположения относительно устьичной щели выделяют несколько типов устьичного аппарата (рис. 3.7 ). В фармакогнозии типы устьичного аппарата используются для диагностики лекарственного растительного сырья.
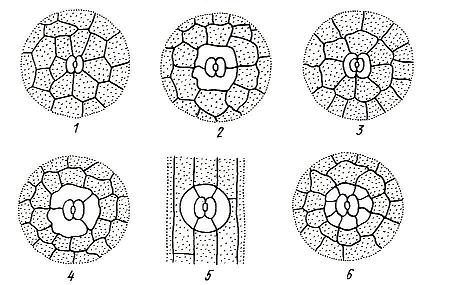
Рис. 3.7. Типы устьичного аппарата : 1 – аномоцитный; 2 – диацитный; 3 – парацитный; 4 – анизоцитный; 5 – тетрацитный; 5 – энциклоцитный.
Аномоцитный тип устьичного аппарата обычен для всех групп растений, исключая хвощи. Побочные клетки в этом случае не отличаются от остальных клеток эпидермы. Диацитный тип характеризуется двумя побочными клетками, которые располагаются перпендикулярно устьичной щели. Этот тип обнаружен у некоторых цветковых растений, в частности, у большинства губоцветных (мята, шалфей, чабрец, душица) и гвоздичных. При парацитном типе две побочные клетки располагаются параллельно замыкающим и устьичной щели. Он найден у папоротников, хвощей и ряда цветковых растений. Анизоцитный тип обнаружен только у цветковых растений, в частности, он встречается у крестоцветных (пастушья сумка, желтушник) и пасленовых (белена, дурман, красавка). В этом случае замыкающие клетки окружены тремя побочными, одна из которых заметно крупнее или мельче остальных. Тетрацитным типом устьичного аппарата характеризуются преимущественно однодольные. При энциклоцитном типе побочные клетки образуют узкое кольцо вокруг замыкающих клеток. Подобная структура найдена у папоротников, голосеменных и некоторых цветковых.
Механизм движения замыкающих клеток основан на том, что стенки их утолщены неравномерно, поэтому форма клеток меняется при изменении их объема. Изменение объема клеток устьичного аппарата происходит вследствие изменения осмотического давления. Увеличение давления происходит за счет активного поступления из соседних клеток ионов калия, а также за счет повышения концентрации сахаров, образующихся в процессе фотосинтеза. За счет поступления воды объем вакуоли увеличивается, тургорное давление растет, и устьичная щель открывается. Отток ионов совершается пассивно, вода выходит из замыкающих клеток, их объем уменьшается, и устьичная щель закрывается. У большинства растений устьица открываются в светлое время суток и закрываются ночью. Это связано с тем, что фотосинтез протекает только на свету, и для него необходим приток из атмосферы углекислого газа.
Число и распределение устьиц очень варьируют в зависимости от вида растения и экологических условий. У большинства растений их число составляет 100-700 на 1мм 2 поверхности листа. С помощью устьиц эпидерма эффективно регулирует газообмен и транспирацию. Если устьица полностью открыты, то транспирация идет с такой же скоростью, как если бы эпидермы не было вовсе (согласно закону Дальтона, при одной и той же суммарной площади отверстий скорость испарения тем выше, чем больше число отверстий). При закрытых устьицах транспирация резко снижается и фактически может идти только через кутикулу.
У многих растений эпидерма образует наружные одно- или многоклеточные выросты различной формы – трихомы . Трихомы отличаются крайним разнообразием, оставаясь вместе с тем вполне устойчивыми и типичными для определенных видов, родов и даже семейств. Поэтому признаки трихомов широко используются в систематике растений и в фармакогнозии в качестве диагностических.
Трихомы делятся на: 1) кроющие и 2) железистые . Железистые трихомы образуют вещества, которые рассматриваются как выделения. Они будут рассмотрены в разделе, посвященном выделительным тканям.
^ Кроющие трихомы имеют вид простых, разветвленных или звездчатых волосков, одно- или многоклеточных (рис. 3.8 ). Кроющие трихомы могут длительное время оставаться живыми, но чаще они быстро отмирают и заполняются воздухом.
Густой слой волосков отражает часть солнечных лучей и уменьшает нагрев, создает затишное пространство около эпидермы, что в совокупности снижает транспирацию. Часто волоски образуют покров только там, где располагаются устьица, например на нижней стороне листьев мать-и-мачехи, багульника. Жесткие, колючие волоски защищают растения от поедания животными, сосочки на лепестках привлекают насекомых.
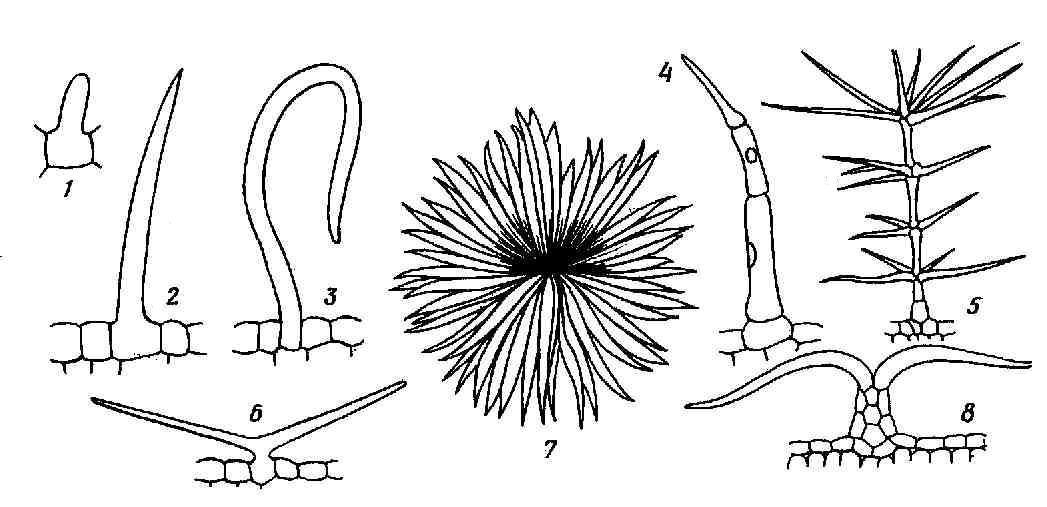
Рис. 3.8. Кроющие трихомы : 1-3 – простые одноклеточные, 4 – простой многоклеточный, 5 – ветвистый многоклеточный, 6 – простой двурогий, 7,8 – звездчатый (в плане и на поперечном разрезе листа).
От трихомов, образующихся только из эпидермальных клеток, следует отличать эмергенцы , в формировании которых принимают участие и более глубоко расположенные ткани. К ним относят шипы розы, малины, ежевики, покрывающие черешки листьев и молодые побеги.
К вторичным покровным тканям относятся: 1) перидерма и 2) корка , или ритидом .
Перидерма – сложная многослойная покровная ткань, которая приходит на смену первичным покровным тканям – ризодерме и эпидерме. Перидерма покрывает корни вторичного строения и стебли многолетних побегов. Она может возникнуть и в результате залечивания поврежденных тканей раневой меристемой.
Перидерма состоит из трех комплексов клеток, различных по строению и функциям. Это: 1) феллема , или пробка , выполняющая главные защитные функции; 2) феллоген , или пробковый камбий , за счет работы которого образуется перидерма в целом; 3) феллодерма , или пробковая паренхима , выполняющая функцию питания феллогена ( рис. 3.9).
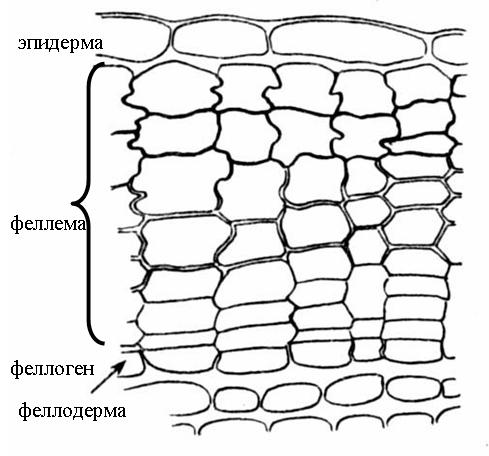
Рис. 3.9. Строение перидермы стебля бузины .
Феллема (пробка) состоит из нескольких слоев таблитчатых клеток, расположенных плотно, без межклетников. Вторичные клеточные стенки состоят из чередующихся слоев суберина и воска, что делает их непроницаемыми для воды и газов. Клетки пробки мертвые, они не имеют протопласта и заполнены воздухом. В полости клеток могут также откладываться вещества, повышающие защитные свойства пробки.
^ Феллоген (пробковый камбий) – вторичная латеральная меристема. Это один слой меристематических клеток, откладывающих клетки пробки наружу и клетки феллодермы внутрь органа. Феллодерма (пробковая паренхима) относится к основным тканям и состоит из живых паренхимных клеток. Однако часто феллоген работает односторонне, откладывая только пробку, а феллодерма остается однослойной (рис. 3.9).
Главная функция пробки – защита от потери влаги. Кроме того, пробка предохраняет растение от проникновения болезнетворных организмов, а также дает механическую защиту стволам и ветвям деревьев, а феллоген залечивает нанесенные повреждения, образуя новые слои пробки. Поскольку клетки пробки заполнены воздухом, пробковый футляр обладает малой теплопроводностью и хорошо предохраняет от резких колебаний температуры.
У большинства деревьев и кустарников феллоген закладывается в однолетних побегах уже в середине лета. Чаще всего он возникает из паренхимных клеток, лежащих сразу под эпидермой (рис. 3.9 ). Иногда феллоген образуется в более глубоких слоях коры (смородина, малина). Редко эпидермальные клетки, делясь, превращаются в феллоген (ива, айва, олеандр).
Газообмен и транспирация в органах, покрытых перидермой, происходят через чечевички (рис. 3.10 ). В местах чечевичек пробковые слои разорваны и чередуются с паренхимными клетками, рыхло соединенными между собой. По межклетникам этой выполняющей ткани циркулируют газы. Феллоген подстилает выполняющую ткань и, по мере ее отмирания, дополняет новыми слоями. С наступлением холодного сезона феллоген откладывает под выполняющей тканью замыкающий слой , состоящий из клеток пробки. Весной этот слой под напором новых клеток разрывается. В замыкающих слоях имеются небольшие межклетники, так что живые ткани ветвей деревьев даже зимой не отграничены наглухо от окружающей среды.
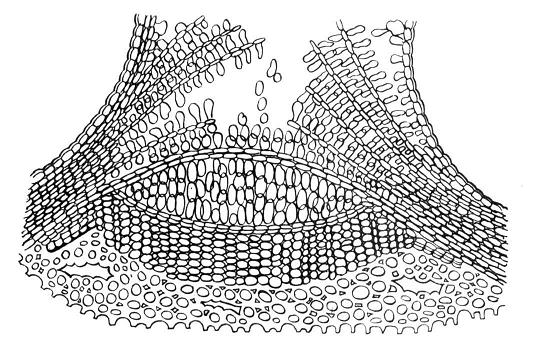
Рис. 3.10. Строение чечевички бузины на поперечном разрезе.
На молодых побегах чечевички выглядят как небольшие бугорки. По мере утолщения ветвей их форма меняется. У березы они растягиваются по окружности ствола и образуют характерный рисунок из черных черточек на белом фоне. У осины чечевички принимают форму ромбов.
У большинства древесных растений на смену гладкой перидерме приходит трещиноватая корка (ритидом) . У сосны это происходит на 8-10-м году, у дуба – в 25-30 лет, у граба – в 50 лет. Лишь у некоторых деревьев (осина, бук, платан, эвкалипт) корка вообще не образуется.
Корка возникает в результате многократного заложения новых прослоек перидермы во все более глубоких слоях коры. Живые клетки, заключенные между этими прослойками, погибают. Таким образом, корка состоит из чередующихся слоев пробки и прочих отмерших тканей коры (рис. 3.11 ).
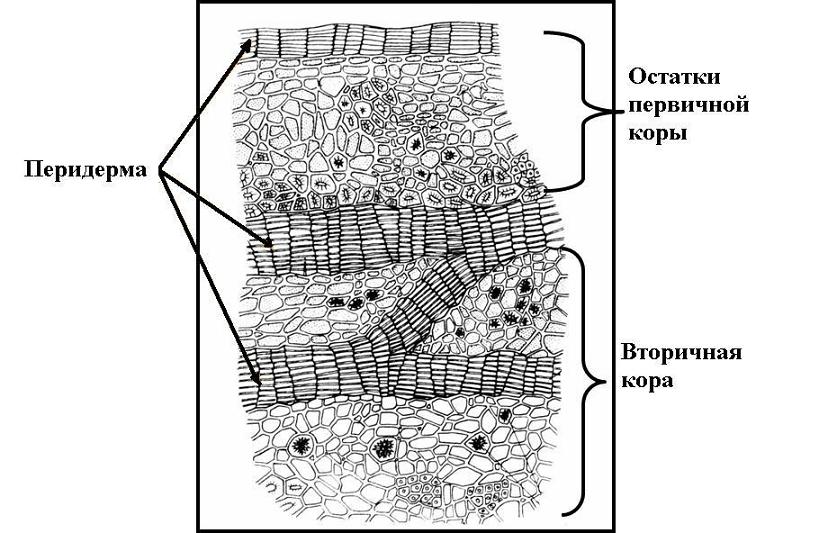
Рис. 3.11. Корка дуба на поперечном разрезе .
Мертвые ткани корки не могут растягиваться, следуя за утолщением ствола, поэтому на стволе появляются трещины, не доходящие, однако, до глубинных живых тканей. Граница между перидермой и коркой внешне заметна по появлению этих трещин, особенно ясна эта граница у березы, у которой белая береста (перидерма) сменяется черной трещиноватой коркой. Толстая корка надежно предохраняет стволы деревьев от механических повреждений, лесных пожаров, резкой смены температур.