Vách tế bào. Nó là lớp vỏ đa lớp gồm 9...10 lớp có mật độ electron khác nhau. Một hệ thống các vi sợi được nhúng trong một ma trận vô định hình tạo thành bộ xương tế bào. Các sợi nhỏ, tùy thuộc vào loài, có thể bao gồm cellulose, glucone và chitin. Các polysacarit, protein, sắc tố, lipid khác đóng vai trò là chất kết dính hình thành liên kết hóa học với phần vi sợi của thành tế bào. Sự hiện diện của các phức hợp như vậy mang lại tính thấm chọn lọc cho một số chất và phong tỏa các chất khác.
Các vi sợi hỗ trợ của thành tế bào và ma trận của nó khác nhau về cơ chế hình thành và sinh tổng hợp. Sự hình thành các sợi và ma trận xảy ra không đồng bộ; trước hết, bộ xương sợi của tường được tái tạo. Quá trình sinh tổng hợp hai phần này của thành tế bào được thực hiện với sự tham gia của các enzyme.
Quá trình hình thành thành tế bào xảy ra theo hai cách: vật liệu mới có thể được đưa vào thành tế bào theo cách phân cực hoặc được phủ đều lên toàn bộ bề mặt của nó. Trong trường hợp đầu tiên, các tế bào hình trụ được hình thành, trong trường hợp thứ hai - hình cầu.
Thành tế bào đóng vai trò như một thiết bị bảo vệ và bảo vệ tế bào nấm khỏi tác động của các yếu tố môi trường khác nhau, ví dụ, hàng rào thẩm thấu xác định tính thấm chọn lọc đối với các chất khác nhau. Nó mang lại hình dạng cho các tế bào sinh dưỡng của sợi nấm và cơ quan sinh sản. Trên bề mặt thành tế bào và màng tế bào chất tập trung các enzyme thực hiện quá trình biến đổi các polyme không được tế bào đồng hóa (không hòa tan trong nước).
Kết quả của quá trình ly giải, thành tế bào của nấm có thể bị phá hủy dưới tác động của các enzyme do các tế bào khác tiết ra và hình thành trong chính tế bào của nấm.
Thành phần chính của thành tế bào nấm là chitin, glucans, protein và chất béo. Các polysacarit chứa nitơ và không chứa nitơ tạo thành các phức chất hòa tan và không hòa tan với các chất béo. Thành tế bào được cấu tạo từ 4...6 monosaccharide, tỷ lệ của chúng thay đổi đôi chút giữa các loại nấm khác nhau. Thành phần của các phân đoạn polysaccharide bao gồm glucosamine, mannose, glucose, xyloza, v.v. Cần nhấn mạnh rằng thành phần màng tế bào của các tế bào khác nhau của cùng một loại nấm là không giống nhau.
nguyên sinh chất- Thành phần tế bào được bao bọc trong thành tế bào: Có màng tế bào chất, mạng lưới nội chất, một hoặc nhiều nhân với nucleoli, cũng như ty thể, ribosome với RNA, lysosome, bộ máy Golgi, không bào, phức hợp lamellar, hạt bài tiết, cũng như các loại khác cấu trúc và các tạp chất khác nhau.
Màng tế bào chất. Một màng ba lớp mỏng nằm ngay dưới thành tế bào và ngăn cách nó với tế bào chất. Màng tế bào chất có khả năng thấm chọn lọc các chất vào và ra khỏi tế bào. Màng tế bào chất chứa tới 40% lipid và tới 38% protein. Các hình thức xâm lấn và xâm phạm màng tế bào chất khác nhau được gọi là mesosome.
Mục đích chức năng chính của màng tế bào chất như sau: đưa các chất khác nhau vào tế bào, xử lý enzyme và giải phóng các sản phẩm trao đổi chất. Các chất được xử lý trong màng tế bào chất sẽ đi vào nguyên sinh chất của tế bào và tham gia vào quá trình trao đổi chất.
Lưới nội chất. Nó bao gồm các túi, ống và không bào, đóng vai trò như một loại kho chứa chất dinh dưỡng.
Ty thể. Vô số khối khép kín di động có hình elip, có vách ngăn, được bao phủ bởi lớp vỏ một hoặc hai lớp. Người ta cho rằng ty thể, do có cấu trúc vòng DNA của riêng chúng, nên có khả năng sinh sản. Ty thể được bao quanh bởi một màng trên đó tập trung các enzyme: pyruvate oxidase, succindehydrogenase, phosphatase kiềm và axit, peroxidase, v.v. Ty thể đóng vai trò là nguồn tạo năng lượng trong tế bào. Tùy thuộc vào điều kiện nuôi cấy và trạng thái sinh lý của tế bào, hình dạng của ty thể và số lượng của chúng trong tế bào sẽ khác nhau.
Ribosome. Các hạt tròn có tính chất ribonucleoprotein, kích thước lên tới 200, tham gia vào quá trình tổng hợp protein của tế bào. Số lượng ribosome thay đổi đáng kể giữa các loại nấm khác nhau và phụ thuộc vào các yếu tố bên ngoài, tuổi nuôi cấy, v.v..
bộ máy Golgi. Nó được thể hiện bằng một nhóm bong bóng có đường kính rất nhỏ (0,000 002...O,OOO 01 micron) hoặc các tấm hình đĩa song song. Cơ quan này nằm trong tế bào ở khu vực không có ribosome.
Lysosome. Các dẫn xuất của bộ máy Golgi nằm giữa màng tế bào và màng tế bào chất. Chúng là những dạng hạt được bao quanh bởi màng lipoprotein một lớp. Chúng chứa một loại enzyme thủy phân protein và thực hiện chức năng bảo vệ tế bào khỏi tác động bất lợi của các chất độc hại có nguồn gốc ngoại sinh và nội sinh.
Liposome. Các giọt chất béo được bao quanh bởi màng đơn lớp.
Cốt lõi. Nằm ở trung tâm hoặc ở hai cực của tế bào. Tế bào nấm có thể có một hoặc nhiều nhân. Họ chịu trách nhiệm về các chức năng di truyền. Hình dạng của hạt là tròn hoặc thon dài. Mỗi nhân được bao quanh bởi một màng nhân xốp hai lớp với nhân gồm các hạt dày đặc và các sợi mỏng. Các nucleoli chứa DNA như một phần của nhiễm sắc thể. Thông qua các đường nối, nhân có thể di chuyển từ tế bào này sang tế bào khác.
Bao gồm. Tế bào nấm chứa nhiều thể vùi: volutin, glycogen, lipid, sắc tố, hình thành tủy, muối của axit hữu cơ, axit amin, v.v. Người ta tin rằng glycogen chịu trách nhiệm cho hô hấp nội sinh và volutin đóng vai trò là chất dinh dưỡng dự trữ liên quan đến các quá trình năng lượng.
Cần lưu ý rằng trong quá trình sống, các sản phẩm trao đổi chất khác nhau tích tụ trong tế bào nấm - kháng sinh, enzyme, độc tố, vitamin, v.v.
Tất cả các yếu tố hình thái của nấm cực nhỏ được chia thành hai nhóm: sợi nấm và bào tử. Chúng có nhiều hình dạng và kích cỡ khác nhau. Sự khác biệt về hình thái giữa bào tử và sợi nấm đóng vai trò là đặc điểm phân biệt quan trọng khi xác định loại nấm.
Sợi nấm. Nó là một ống tròn hẹp, đường kính của nó khác nhau giữa các vi nấm từ một đến vài micron.
Với sự phân nhánh dồi dào, các sợi nấm khi tiếp xúc với nhau có thể hình thành sự kết hợp giữa các tế bào - sự nối liền. Nếu có số lượng lớn, sợi nấm sẽ có hình dạng dạng lưới đặc trưng. Sự phát triển của các lỗ nối được quan sát thấy ở nhiều loại nấm có sợi nấm đa bào. Nhờ chúng, có thể di chuyển nhân tế bào từ tế bào này sang tế bào khác và chuyển từ sợi nấm đơn bội sang sợi nấm lưỡng bội. Tuy nhiên, trong hầu hết các trường hợp, chúng thực hiện chức năng sinh dưỡng và phát triển dưới nhiều hình thức do thiếu dinh dưỡng. Chiều dài của các tế bào sợi nấm dao động từ vài micron đến hàng chục và ít hơn là hàng trăm micron.
Sợi nấm được bao quanh bởi lớp vỏ kép, lớp vỏ này mỏng manh hơn ở những cây non. Trong các vách ngăn chia sợi nấm thành các tế bào riêng lẻ, có các lỗ mà qua đó tế bào chất chảy qua trong quá trình phát triển và cùng với đó là các chất dinh dưỡng. Có nhiều thể vùi khác nhau trong tế bào: ở tế bào già, tế bào chất trở nên dạng hạt do có nhiều không bào. Sợi nấm non bao gồm các tế bào hình chữ nhật thon dài, những tế bào già - có hình tròn ngắn hoặc nhiều mặt. Sợi nấm có các phân vùng được gọi là vách ngăn. Tuy nhiên, ở một số loại nấm bậc thấp, sợi nấm bao gồm các sợi nấm không có vách ngăn ngang và giống như một tế bào khổng lồ phân nhánh cao với nhiều nhân và được gọi là sợi nấm không có vách ngăn.
Sợi nấm phát triển như thế nào? Một ống mầm nhô ra từ bào tử, ống này dài ra và sau đó được ngăn cách bởi một vách ngăn với phần giữa, bao gồm cả bào tử. Sau đó, các ống tăng trưởng dài hơn nữa và nhận được một vách ngăn mới, phân chia thành tế bào ở xa hoặc ở đỉnh và tế bào ở gần hoặc ở bên trong. Sau đó, tế bào đỉnh dài ra và phân chia trở lại, tách ra một tế bào thứ hai, bên trong trẻ hơn tế bào đầu tiên. Điều này tiếp tục xảy ra. Trong quá trình này, các tế bào bên trong chỉ dài ra, sự phân chia theo chiều ngang của chúng hiếm khi xảy ra mà các nhánh bên phát triển từ chúng. Ở đầu xa của tế bào bên trong, một phần nhô ra bên trong được hình thành, có dạng hình trụ và sau đó được ngăn cách bởi một vách ngăn với tế bào tạo ra nó. Tế bào mới sau đó sẽ phát triển thành một nhánh phụ, phát triển và phân nhánh giống như tế bào chính. Do sự phát triển của các cành dọc theo sợi nấm chính nên chúng càng già và phát triển hơn, cành của chúng càng nằm gần gốc - phân nhánh acropetal.
Sự phát triển của sợi nấm không có vách ngăn nhìn chung diễn ra theo cách tương tự nhưng không có sự hình thành các phân chia ngang. Sự tăng trưởng xảy ra ở đầu sợi nấm, nơi tích tụ nhiều nguyên sinh chất, lấp đầy toàn bộ lòng và ở các phần sau có sự phát triển đáng kể của không bào trung tâm. Trong môi trường đồng nhất, ví dụ trên bề mặt gelatin dinh dưỡng, sợi nấm (cả không tế bào và đa bào) phát triển đồng đều và tỏa tròn, do đó sợi nấm có dạng vòng tròn mọc ra từ các cạnh. Phần trung tâm trong đó là phần già nhất, thậm chí có khi đã chết, còn phần ngoại vi là phần trẻ nhất.
Mặc dù sự phát triển sợi nấm có tính đồng nhất chung, có thể được gọi là điển hình, nhưng trong một số trường hợp, một số đặc điểm cụ thể vẫn được quan sát thấy, cả về hình thức vĩ mô và mô hình phát triển chung cũng như cấu trúc vi mô. Hình dạng vĩ mô của sợi nấm được xác định chủ yếu bởi sợi nấm trên không. Trong một số trường hợp, chúng hình thành trên chính bề mặt của chất nền và một phần bên trong nó, sau đó sợi nấm có dạng một vòng tròn phẳng được ép vào chất nền; Ngoài ra, trong các trường hợp khác, sợi nấm ít nhiều phát triển, bay lên không khí và tạo cho sợi nấm một số điểm giống, chẳng hạn như một mảnh bông gòn nổi lên trên bề mặt. Kiểu phát triển của cùng một loại nấm có thể khác nhau tùy thuộc vào độ ẩm, dinh dưỡng, v.v. Tuy nhiên, một số dạng nấm có những đặc điểm riêng, chẳng hạn như hình thành sợi nấm trên không tươi tốt - kẻ phá hoại gỗ.
Màu sắc của sợi nấm thường có màu trắng như tuyết, nhưng theo tuổi tác nó sẽ chuyển sang màu nâu với các sắc thái khác nhau. Điều này là do sự lắng đọng sắc tố trong thành tế bào và ít phổ biến hơn là bên trong tế bào.
Có sợi nấm thật và sợi nấm giả. Loại thứ hai được đặc trưng bởi thực tế là các tế bào riêng lẻ không được kết nối với nhau và không có màng chung. Thay vì phân nhánh thực sự, người ta quan sát thấy sự sắp xếp các tế bào giống như cây ở đây.
Để gắn vào chất nền và chiết xuất chất dinh dưỡng từ nó, trong quá trình tiến hóa, một số loại nấm đã hình thành các cơ quan được thiết kế đặc biệt cho việc này: rhizoids và appressoria, những cơ quan này được tính đến khi xác định nấm. Rhizoids có hình dạng rễ, và appressoria là sự phát triển ngắn, mở rộng, đôi khi hình thùy của sợi nấm.
hạch cứng, sợi, rhizomorphs và chlamydospores cũng là những biến đổi của sự phát triển sợi nấm.
Sclerotia là sợi nấm có vách ngăn tạo thành các cơ thể đặc biệt. Khi hạch nấm hình thành, màng sợi nấm dày lên và có màu sẫm. Thành sợi nấm của lớp ngoài của xơ cứng dày lên rất nhiều, nhưng sợi nấm bên trong có thành mỏng hơn và thường không có màu. Xơ cứng là cơ quan thích nghi bảo vệ cho phép nấm tồn tại lâu dài trong môi trường và đảm bảo khả năng chống lại các yếu tố bên ngoài khác nhau: nhiệt độ, ánh sáng mặt trời, v.v. Xơ cứng trưởng thành chứa ít độ ẩm hơn so với sợi nấm và nhiều chất dự trữ - lipid, glycogen .
Kích thước của hạch nấm dao động từ vài mm đến vài chục cm và hình dạng có thể rất đa dạng: hình cầu, không đều, ở dạng sừng thẳng hoặc cong, v.v.
Cấu trúc của các tế bào hạch nấm và cơ chế hình thành của chúng là khác nhau, nhưng sự hình thành của chúng xảy ra bằng cách tăng sự phân nhánh của sợi nấm và sự phân chia của sợi nấm. Có hai cách hình thành hạch nấm được biết đến: đầu cuối - ở đầu sợi nấm; xen kẽ - trong các đoạn riêng biệt của sợi nấm chính.
Ở nhiều loại nấm, trong quá trình phát triển quả thể và một số cấu trúc sinh dưỡng, mô giả được hình thành - plectenchyma (giả nhu mô). Không giống như mô nhu mô thực sự, là kết quả của sự phân chia tế bào theo ba hướng, mô mềm được hình thành bởi đám rối và sự hợp nhất. Nếu nó bao gồm các tế bào có đường kính ít nhiều đẳng hướng thì nó được gọi là paraplectenchyma; nếu nó có cấu trúc giống sợi nấm rõ ràng (tế bào thon dài) thì được gọi là mô tuyến tiền liệt.
Sợi nấm- cấu trúc thực vật của sợi nấm tập hợp tuyến tính. Đường kính của sợi nấm phụ thuộc vào số lượng sợi nấm tập trung xung quanh gốc trung tâm.
Trong trường hợp đơn giản nhất, một số lượng nhỏ sợi nấm song song được dán vào nhau bằng màng nhầy bên ngoài hoặc tạo thành một kết nối bền hơn bằng cách hình thành. hình thành nhiều đường nối ngắn. Trong các trường hợp khác Ở các loại trà, khi dây lớn, sợi nấm của chúng có sự phân hóa nhất định. Các phần bên ngoài mỏng hơn, tạo thành một loại vỏ bao quanh thân cây dày ở trung tâm.
Rhizomorphs- sợi nấm có sự tập hợp phức tạp hơn, khác nhau giữa các loại nấm khác nhau về cường độ phát triển của sợi nấm trung tâm, phạm vi của các nhánh bên và mức độ biệt hóa của các tế bào sợi nấm.
Các phần bên ngoài của rhizomorpha thường có màu sẫm và có nét giống nhất định với rễ của thực vật bậc cao. Chúng phổ biến rộng rãi ở các loại nấm có quả thể lớn: basidiomycetes, thú có túi, v.v.
Mục đích chính của sợi nấm và thân rễ là đảm bảo sự lây lan của nấm trong chất nền và sự di chuyển của các chất dinh dưỡng dọc theo sợi nấm.
bào tử hậu- đây là những thay đổi trong sợi nấm ở các nền văn hóa trưởng thành và già ở phần cuối hoặc dọc theo quá trình của nó. Chức năng chính của chlamydospores không phải là sinh sản mà là bảo tồn loài. Hình dạng của chúng thường có hình tròn, hình bầu dục hoặc hơi thon dài, đường kính vượt quá đường kính của sợi nấm. Một số loại nấm có thành mạch kép, bề mặt nhẵn hoặc thô. Bào tử hậu có thể xuất hiện ở các đầu của sợi nấm, sau đó chúng được gọi là đầu cuối, dọc theo quá trình của sợi nấm - liên cực (trung gian).
Trong các nền văn hóa cổ xưa, các cụm bào tử hậu lớn thường được quan sát thấy có hình dạng kỳ quái, gợi nhớ đến một chuỗi tràng hạt hoặc một chiếc vòng cổ. Chlamydospores trẻ và trưởng thành có khả năng nảy mầm. Tế bào già bị thoái hóa.
Tranh cãi. Với sự trợ giúp của bào tử, nấm không chỉ sinh sản mà còn lây lan khắp môi trường. Điều này được tạo điều kiện thuận lợi bởi khả năng chống chịu cao của vỏ bào tử đối với các yếu tố xâm thực. Bào tử được chia thành nội bào tử, được hình thành bên trong các thùng chứa đặc biệt - bào tử (túi) và ngoại bào tử, nằm trên sợi nấm.
Ở nấm hoàn hảo, bào tử được chia thành bào tử trứng, bào tử hợp tử, bào tử túi, bào tử đảm, bào tử nội bào, bào tử phialospores và bào tử chlamydospore. Bào tử của nấm không hoàn hảo cũng được chia thành nhiều nhóm theo kích thước và nguồn gốc. Nội bào tử được hình thành bên trong sợi nấm bằng cách phân chia các bào tử sau bao gồm thallospores, bao gồm bào tử đốt, bào tử chlamydospore và bào tử phôi. Ngoài ra, nấm không hoàn hảo được đặc trưng bởi sự hình thành conidia, macroconidia, aleuria (microconidia) và hemispores, được coi là conidia không hoàn hảo.
Bán bào tử liên kết chặt chẽ hơn với sợi nấm và đại diện cho một hoặc hai đoạn bị tách ra sau khi phân chia theo chiều ngang của sợi nấm. Hình dạng của chúng là hình trụ, đôi khi tròn hoặc nhiều mặt, vỏ có mạch kép.
Thành phần hóa học và cấu trúc của thành tế bào nấm
Thành tế bào của nấm có nhiều lớp, với các lớp khác nhau được hình thành bởi các carbohydrate cấu trúc khác nhau về thành phần hóa học, có thể chia thành 3 nhóm dựa trên thành phần hóa học của chúng:
polyme glucose(glucan, chitin, xenlulozơ). Glucans tạo nên lớp ngoài của thành tế bào của hầu hết các loại nấm. Lớp bên trong của thành tế bào nấm được hình thành bởi các chuỗi chitin, tạo nên độ cứng. Chitin thay thế cellulose, chất không có ở hầu hết các loại nấm, nhưng là một phần của thành tế bào của oomycetes, hiện không thuộc về các loại nấm điển hình. Chitin đã khử acetyl được gọi là chitosan, kết hợp với chitin tạo thành thành tế bào của hợp tử.
polyme của các monosacarit khác (mannose, galactose, v.v.) Không giống như thực vật bậc cao, nơi chúng tạo thành nền tảng của ma trận dưới tên chung là hemicellulose, chúng ít đặc trưng hơn của nấm. Ngoại lệ là nấm men, thành tế bào của chúng đặc biệt giàu polyme mannose gọi là mannan. Người ta cho rằng thành phần tường này đảm bảo tốt hơn cho sự nảy chồi.
polyme của carbohydrate liên kết cộng hóa trị với peptide (glycoprotein) tạo thành lớp giữa của thành tế bào nhiều lớp và đóng vai trò quan trọng trong việc duy trì tính toàn vẹn cấu trúc của tế bào và trong quá trình trao đổi chất của nó với môi trường.
Các đặc điểm cụ thể khác của tế bào nấm bao gồm: không có plastid, đưa nó đến gần hơn với tế bào động vật;
Không có tinh bột, trong eumycetes được thay thế bằng một polysaccharide gần với glycogen của tinh bột động vật, trong oomycetes bằng một polysaccharide gần với laminarin của tảo nâu. Một số carbohydrate dự trữ dành riêng cho nấm cũng được sản xuất.
Phát triển cụ thể chất chuyển hóa thứ cấp, trong đó kháng sinh, phyto- và mycotoxin và phytohormone đóng vai trò chính.
Các đặc điểm cụ thể của nấm còn bao gồm dị nhân và quá trình lưỡng tính.
Ở nấm, hiện tượng dị nhân hoặc dị nhân rất phổ biến, trong đó các hạt nhân dị thể đối với một số gen vẫn tồn tại trong một tế bào trong một thời gian dài. Heterokaryosis thay thế dị hợp tử ở nấm đơn bội và thúc đẩy sự thích nghi nhanh chóng của nấm với các điều kiện thay đổi. Sự hiện diện của các hạt nhân khác nhau là do một số đặc điểm cụ thể của nấm:
1. sự hiện diện của nhiều hơn một nhân trong một tế bào
2. cấu trúc cụ thể của thành tế bào, trong đó có một hoặc nhiều lỗ xuyên qua, gọi là lỗ chân lông, qua đó nhân có thể di chuyển từ tế bào này sang tế bào khác
3. sợi nấm trong một khuẩn lạc và thậm chí cả các khuẩn lạc có khoảng cách gần nhau khác nhau được trồng từ các bào tử khác nhau của cùng một loại nấm thường phát triển cùng nhau, do đó có thể trao đổi nhân của các chủng khác nhau.
Parasex (giả tình dục) quá trình. Nếu trong các tế bào dị nhân có nhân dị hợp ở bất kỳ locus nào hợp nhất thì nhân lưỡng bội dị hợp tử sẽ xuất hiện. Nó có thể xâm nhập vào bào tử và tạo ra một dòng vô tính dị hợp tử lưỡng bội. Trong quá trình nguyên phân, các hạt nhân lưỡng bội có thể trở lại trạng thái đơn bội do mất một bộ nhiễm sắc thể hoặc sự trao đổi các phần nhiễm sắc thể có thể xảy ra trong chúng (giao thoa phân bào). Cả hai quá trình đều đi kèm với sự tái tổ hợp các gen của bố mẹ và do đó là các kiểu hình. Tái tổ hợp ký sinh trùng (vô tính) là một hiện tượng rất hiếm gặp, không vượt quá một nhân trên một triệu, nhưng do số lượng nhân trong sợi nấm rất lớn nên nó liên tục được quan sát thấy trong quần thể nấm.
sinh sản- sinh dưỡng, vô tính, hữu tính.
thực vật– sự phân mảnh của thallus, hình thành bào tử hậu, sau một thời gian ngủ sẽ nảy mầm thành sợi nấm, nảy chồi trong nấm men.
Vô tính Việc sinh sản ở các loại nấm khác nhau có thể được thực hiện bằng bào tử di động và bất động. Động bào tử tạo thành một nhóm tương đối nhỏ các loại nấm và sinh vật giống nấm - sống dưới nước và một số trên cạn, trong đó có thể thấy rõ mối liên hệ di truyền với nấm và tảo thủy sinh. Cấu trúc của tiên mao là một đặc điểm chẩn đoán quan trọng khi được gán cho một giới cụ thể. Ở phần lớn các loại nấm, trong quá trình sinh sản vô tính, các bào tử bất động được hình thành, điều này cho thấy chúng đã xuất hiện rất lâu đời trên đất liền. Theo nơi hình thành và nội địa hóa, bào tử nội sinh được phân biệt, hình thành ở bào tử và ngoại sinh (conidia), phát triển trên các sợi nấm - conidiophores đặc biệt. Conidia được hình thành ở hầu hết các loại nấm (ascomycetes, basidiomycetes, một số zygomycetes) tạo nên bào tử conidial, cực kỳ đa dạng và được sử dụng rộng rãi để chẩn đoán nấm.
Sinh sản hữu tính của nấm có đặc điểm riêng cả về hình thái của quá trình tình dục cũng như cơ chế điều hòa di truyền và sinh lý của giới tính cũng như việc truyền tải thông tin di truyền.
hôn nhân Somatogamy- loại quá trình sinh dục phổ biến nhất và đơn giản nhất, bao gồm sự hợp nhất của hai tế bào soma không biệt hóa thành giao tử. Đôi khi nó xảy ra ngay cả khi không có sự hợp nhất tế bào - các hạt nhân bên trong tế bào hợp nhất. Được tìm thấy ở hầu hết các loài nấm basidiomycetes, nấm men có túi và một số loài phân loại khác.
giao tử– giao tử được phân lập trên sợi nấm đơn bội và nội dung của chúng bị cạn kiệt trong quá trình giao phối. Quá trình tình dục này là đặc trưng của hầu hết các loại nấm có túi. Một biến thể của gametangiogamy là hợp tử ở hợp tử.
giao tửở dạng iso-hetero và oogamy, nó xảy ra ở nấm ít thường xuyên hơn so với các sinh vật nhân chuẩn khác. Sự đồng giao và dị giao tử chỉ xảy ra ở chytridiomycetes. Giao phối cổ điển với sự hình thành tinh trùng và trứng không được biểu hiện ở nấm, nhưng đã tìm thấy các biến thể biến đổi cao.
Theo đặc điểm điều hòa giới tính ở nấm, người ta phân biệt một số loại quá trình sinh dục.
bệnh phụ khoa có thể được xem xét bằng cách sử dụng ví dụ về oomycetes khác gốc, trong đó oogonia và antheridia phát triển trên các loại thalli khác nhau, ví dụ như bệnh mốc sương hoặc nấm khoai tây. Nếu một chủng đồng nhất về mặt di truyền được trồng trong môi trường độc canh thì nó chỉ sinh sản vô tính. Nếu sợi nấm của hai chủng ở gần nhau, thì những thay đổi về hình thái có thể được theo dõi ở chúng dưới tác động của sự tiết ra hóa chất có tính chất steroid - pheromone giới tính. Antheridiol gây ra sự hình thành antheridia ở đối tác và oogoniol gây ra sự hình thành oogonium. Trong trường hợp này, sự điều hòa giới tính chỉ mang tính tương đối: một chủng sẽ hình thành antheridia hay oogonia phụ thuộc vào tỷ lệ định lượng của các pheromone tương ứng trong đó và đối tác của nó. Do đó tên của quá trình tình dục - gynandromixis.
Dimixis hoặc chủ nghĩa dị thể. Từ lâu người ta đã lưu ý rằng nấm có thể là đồng thể hoặc dị thể. Ở các loài đồng tản, trong quá trình sinh sản hữu tính, các hạt nhân giống hệt nhau về mặt di truyền trong sợi nấm sẽ hợp nhất. Ở các loài dị hợp, để trải qua chu kỳ sinh dục, ở một giai đoạn nào đó (khác nhau đối với các loại nấm khác nhau), sự hợp nhất của các thế hệ bào tử (chính xác hơn là hạt nhân của chúng) là cần thiết. Để hai chủng có thể tương thích về mặt giới tính, phải có sự khác biệt về di truyền (tính dị hợp) ở một số locus nhất định được gọi là locus giao phối. Hầu hết các loại nấm (hợp tử, nấm ascomycetes và một số nấm basidiomycetes) có một locus giao phối với hai alen. Vị trí giao phối bao gồm một số gen kiểm soát sự tổng hợp pheromone giới tính. Chủ nghĩa dị thể như vậy được gọi là đơn yếu tố hoặc lưỡng cực. Con cái của những loại nấm này sau quá trình giảm phân được chia thành hai nhóm tự không tương thích nhưng tương thích lẫn nhau theo tỷ lệ 1:1, tức là xác suất lai có quan hệ họ hàng (cận huyết) và không họ hàng (giao phối cận huyết), giống như ở sinh vật nhân chuẩn lưỡng tính cao hơn, là 50%.
Trong bộ gen của nấm basidiomycetes cao hơn có hai locus giao phối - A và B, và chỉ những chủng dị hình ở cả hai locus mới tương thích với nhau (Ax Bx tương thích với Ay By, nhưng không tương thích với Ax By và Ay Bx). Chủ nghĩa dị thể như vậy được gọi là hai yếu tố hoặc tứ cực. Nó làm giảm nguy cơ cận huyết tới 25%.
Diaphoromixis– các basidiomycetes cao hơn không phải có hai mà có nhiều alen của locus giao phối, được tìm thấy ngẫu nhiên ở các chủng khác nhau tạo nên quần thể. Việc kiểm soát giao phối này đảm bảo 100% cơ hội cận huyết vì các chủng có các alen khác nhau tương thích lẫn nhau và có rất nhiều alen. Kết quả là quần thể lai panmix được hình thành.
Vòng đời của nấm cũng đa dạng như chính nấm. Các chu kỳ chính, thuộc về các bộ phận của nấm
1. Chu kỳ vô tính là đặc trưng của một nhóm lớn các nấm không hoàn hảo hoặc mitofungi đã mất khả năng sinh sản hữu tính. Sự phân chia hạt nhân của chúng chỉ là phân bào. Phần lớn mitofungi thuộc về nấm có túi, nhưng do mất quá trình sinh sản hữu tính nên chúng tạo thành một nhóm chính thức gồm các loại nấm không hoàn hảo hoặc deuteromycetes.
2. Chu kỳ đơn bội. Thallus thực vật mang nhân đơn bội. Sau quá trình sinh dục (syngamy), hợp tử lưỡng bội (thường là sau một thời gian nghỉ ngơi) phân chia theo phương pháp giảm phân - giảm phân hợp tử. Đặc điểm của zygomycetes và nhiều chytridiomycetes.
3. Chu trình đơn bội-dicaryotic được đặc trưng bởi thực tế là sau khi hợp nhất các thành phần của giao tử (gametangiogamy) hoặc tế bào soma của sợi nấm đơn bội (somatogamy), các hạt nhân hình thành các dikaryon (cặp hạt nhân khác nhau về mặt di truyền). Chúng phân chia đồng bộ, tạo thành sợi nấm dikaryotic. Quá trình sinh dục kết thúc bằng sự hợp nhất của các hạt nhân dikaryon, hợp tử thu được sẽ phân chia bằng phương pháp giảm phân mà không có thời gian nghỉ ngơi. Meiospores tạo thành bào tử hữu tính của thú có túi và basidiomycetes dưới dạng ascospores và basidiospores. Khi chúng nảy mầm, sợi nấm đơn bội được hình thành. Ở phần lớn các loại nấm có túi (trừ nấm men và nấm taphrin), vòng đời bị chi phối bởi giai đoạn đơn bội ở dạng sợi nấm thực vật (anamorph), giai đoạn dikaryotic tồn tại trong thời gian ngắn và được biểu hiện bằng sợi nấm ascogenous trên đó túi được hình thành (teleomorph). Ở basidiomycetes, giai đoạn dikaryotic chiếm ưu thế trong vòng đời; giai đoạn đơn bội tồn tại trong thời gian ngắn.
4. Chu trình đơn bội-lưỡng bội ở dạng biến đổi đẳng hình thế hệ hiếm gặp ở nấm (một số nấm men và chytridiomycetes thủy sinh).
5. Chu trình lưỡng bội là đặc trưng của nấm trứng và một số loài nấm có túi. Thallus sinh dưỡng là lưỡng bội, giảm phân giao tử được quan sát thấy trong quá trình hình thành giao tử hoặc giao tử.
3. Nhóm sinh thái của nấm.
Nấm và các sinh vật giống nấm được bao gồm trong tất cả các hệ sinh thái trên cạn và dưới nước như là phần quan trọng nhất của khối dị dưỡng, cùng với vi khuẩn chiếm bậc dinh dưỡng của sinh vật phân hủy. Sự phân bố rộng rãi của nấm trong sinh quyển được xác định bởi một số đặc điểm quan trọng:
1. Phần lớn có cấu trúc sợi nấm của thallus. (cho phép bạn nhanh chóng làm chủ chất nền và có bề mặt tiếp xúc lớn với môi trường).
2. Tốc độ tăng trưởng và sinh sản cao, cho phép xâm chiếm khối lượng lớn chất nền trong một thời gian ngắn, hình thành một số lượng lớn bào tử và phân phối chúng trên khoảng cách xa.
3. Hoạt động trao đổi chất cao, biểu hiện ở nhiều yếu tố môi trường.
4. Tỷ lệ tái tổ hợp di truyền cao, biến đổi sinh hóa đáng kể, độ dẻo môi trường.
5. Khả năng chuyển nhanh sang trạng thái hoạt động lơ lửng và tồn tại trong điều kiện bất lợi trong thời gian dài.
Yếu tố môi trường chính của nấm là chất nền thức ăn. Liên quan đến yếu tố này, các nhóm nấm chính được phân biệt, được gọi là nhóm chiến lợi phẩm.
4. Mô tả ngắn gọn các phần nấm.
VI SINH, 2010, tập 79, số 6, tr. 723-733
UDC 582.281(047)
TƯỜNG TẾ BÀO NẤM: KHÁI NIỆM HIỆN ĐẠI VỀ THÀNH PHẦN VÀ CHỨC NĂNG SINH HỌC
© 2010 E. P. Feofilova1
Viện thuộc Viện Hàn lâm Khoa học Nga Viện Vi sinh được đặt theo tên. S.N. Vinogradsky RAS, Mátxcơva
Nhận bởi biên tập viên 05.11.2009
Bài tổng quan này tập trung vào cấu trúc bề mặt ít được nghiên cứu của tế bào nấm sợi - thành tế bào (CW). Dữ liệu được cung cấp về các phương pháp phân lập và kiểm tra độ tinh khiết để phát hiện sự vắng mặt của hàm lượng tế bào chất trong phần CS và thành phần hóa học của nó. Các thành phần cấu trúc (khung) và nội bộ của CS - aminopolysaccharides, a- và b-glucans, protein, lipid, axit uronic, hydrophobin, sporopollenin và melanin - sẽ được thảo luận chi tiết. Đặc biệt chú ý đến chitin, chức năng mới của nó trong việc bảo vệ tế bào chống lại căng thẳng, cũng như sự khác biệt giữa aminopolysacarit nấm này với chitin của tảo và Arthropoda. Hiện tượng phát triển sợi nấm ở đỉnh và sự tham gia của các vi hạt đặc biệt vào quá trình hình thành hình thái của tế bào nấm sẽ được thảo luận. Dữ liệu được cung cấp về các enzyme liên quan đến quá trình tổng hợp và ly giải CS. Cuối cùng, ý nghĩa chức năng của SC ở nấm so với cấu trúc bề mặt của sinh vật nhân chuẩn cao hơn sẽ được thảo luận.
Từ khóa: nấm sợi, thành tế bào, phương pháp phân lập, thành phần hóa học, sinh trưởng ngọn, chức năng sinh lý, hình thái.
Một trong những vấn đề chính của sinh học hiện đại là câu hỏi làm thế nào mà trong quá trình phát triển lại xảy ra sự hình thành các sinh vật giống hệt nhau về mặt hình thái, cơ chế sinh hóa và cấu trúc tế bào nào tham gia vào quá trình này, vốn đã xảy ra trên hành tinh của chúng ta trong hàng tỷ năm. năm. Không kém phần thú vị là dữ liệu về mức độ căng thẳng ảnh hưởng đến sự hình thành hình thái và loại polyme sinh học nào kiểm soát hình dạng bên ngoài của tế bào. Như kết quả của các nghiên cứu gần đây cho thấy, sợi nấm đang phát triển của nấm sợi là một mô hình độc đáo cho phép chúng ta hiểu quá trình hình thành tế bào và cách duy trì sự ổn định của hình thái tế bào trong quá trình hình thành tế bào. Đóng góp rất lớn cho việc nghiên cứu hiện tượng này là dữ liệu về thành phần và chức năng sinh học của CS của nấm sợi.
CS của thực vật được phát hiện vào năm 1665, còn CS của nấm chỉ được phát hiện vào đầu thế kỷ 18. Tuy nhiên, cấu trúc tế bào bề mặt này thực tế đã không được nghiên cứu trong một thời gian dài, vì người ta tin rằng CS chỉ thực hiện chức năng “khung” và nó được so sánh với các bức tường của một ngôi nhà chịu tải trọng hỗ trợ. Nhưng vào đầu thế kỷ 20, thái độ đối với CS đã thay đổi đáng kể; nó bắt đầu được nghiên cứu chuyên sâu, nhưng chủ yếu ở thực vật và vi khuẩn. Tuy nhiên, sự phát triển mạnh mẽ của nghề trồng nấm và sản xuất công nghệ sinh học, trong đó nấm sợi là nhà sản xuất các hoạt chất sinh học, đã thúc đẩy sự phát triển
1 Địa chỉ liên lạc (e-mail: [email được bảo vệ]).
công trình nghiên cứu khoa học về nghiên cứu nấm CS. Dữ liệu về cấu trúc bề mặt này trong khoảng thời gian cho đến những năm 1980 đã được tóm tắt trong cuốn sách đầu tiên trên thế giới về nấm CS. Trong những năm tiếp theo, nội dung công việc chính diễn ra vào cuối thế kỷ 20 và đầu thế kỷ 21. Các nghiên cứu cơ bản đã được tiến hành về hệ thống, thành phần hóa học của KS, sự phát triển của sợi nấm ở đỉnh, chuyển hóa chitin, thuốc chống nấm, hydrophobin, protein liên kết cộng hóa trị, các enzyme liên quan đến sự hình thành KS, sự phân nhánh của sợi nấm và quá trình ly giải KS. Những dữ liệu này chưa được khái quát hóa, nhưng việc thảo luận chúng cùng nhau cho phép chúng tôi chỉ ra rằng cấu trúc bề mặt này của tế bào mang tải đa chức năng và thực hiện các chức năng quan trọng như bảo vệ tế bào khỏi tác động của các yếu tố bất lợi , kiểm soát hình thái, tham gia vào quá trình sinh sản, xác định tính chất kháng nguyên và kết dính, kiểm soát quá trình lưỡng hình và hình thành tế bào nấm không hoạt động, nhận thức tín hiệu bên ngoài và sự truyền tín hiệu đến màng và các chất truyền tin nội bào. Xem xét những điều trên, chúng tôi cho rằng cần thảo luận về các dữ liệu sau: thành phần hóa học của CS, chức năng sinh học của các polyme sinh học chính của nó, cấu trúc của CS và các tương tác giữa các tế bào trong sợi nấm, hiện tượng tăng trưởng sợi nấm ở đỉnh và các enzyme chính tham gia vào quá trình hình thành và ly giải CS.
THÀNH PHẦN HÓA HỌC CỦA TẾ BÀO
TƯỜNG NẤM VÀ CHỨC NĂNG SINH HỌC CỦA CÁC THÀNH PHẦN CHÍNH CỦA NẤM
Nghiên cứu về thành phần hóa học của CS nấm bắt đầu bằng việc thu được một phần nguyên chất của cấu trúc này, tức là. thanh lọc CS khỏi ô nhiễm tế bào chất. Quy trình ban đầu - tiêu diệt tế bào nấm và rửa nội dung tế bào - được thực hiện ở nhiệt độ thấp (khoảng 4-5°C) để ngăn chặn hoạt động phá hủy của các enzyme phân hủy. Trước khi các tế bào bị phá hủy, chúng được đông lạnh ở nhiệt độ nitơ lỏng. Việc phá hủy tế bào được thực hiện trong các thiết bị đồng nhất đặc biệt hoặc trong máy ép sử dụng phương pháp “áp suất rắn”. Việc sử dụng thêm máy phân hủy siêu âm cho kết quả tốt. Việc rửa nội dung tế bào được thực hiện bằng nước lạnh và lặp lại 4-5 lần. Để loại bỏ triệt để hơn các chất trong tế bào chất, rửa bằng NaCl, urê 8 M, amoni 1 M hoặc axit axetic 0,5 M. Hiện nay, các phương pháp đã được phát triển để phân lập CS nấm trong quá trình phát sinh của chúng và các phương pháp này khác nhau đáng kể đối với sợi nấm và tế bào nghỉ. Tùy thuộc vào hướng nghiên cứu, cụ thể, khi xác định thành phần polysacarit, lipid được chiết từ CS bằng dung môi hữu cơ, ví dụ, cloroform và metanol theo tỷ lệ 2: 1, đôi khi xử lý bổ sung CS bằng ete sulfuric cho kết quả tốt; kết quả. Dung môi này làm khô CS tốt và đôi khi có thể thay thế sấy thăng hoa. Bước tiếp theo là xác định độ tinh khiết của phần CS thu được. Với mục đích này, kính hiển vi ánh sáng và điện tử cũng như các phương pháp nhuộm chuyên dụng được sử dụng. Phản ứng được sử dụng phổ biến nhất dựa trên sự tương tác của I3- với chitosan. CS nguyên chất cô lập với dung dịch Lugol có màu hồng hoặc tím, trong khi CS nguyên vẹn có màu đỏ. Nhuộm để phát hiện sự hiện diện của hạt nhân bằng DAPI (4,6-ekt1to-2-rkepyTsn-yo1) cũng được sử dụng. Cần đặc biệt nhấn mạnh rằng việc phân lập một phần CS nguyên chất là một quy trình rất có trách nhiệm, phụ thuộc vào kết quả phân tích thành phần hóa học tiếp theo của nó. Về vấn đề này, một công trình thú vị là trong đó glucan, hay chính xác hơn là phức hợp chitosan-glucan, được tìm thấy trong sợi nấm đại diện của nấm mucorales, mặc dù tất cả các nghiên cứu trước đây đều chỉ ra sự vắng mặt của glucan trong sợi nấm Mucorales.
Hiện nay, các thành phần của CS nấm được chia thành các thành phần cấu trúc (chitin, p-(1-3)-p(1-6)-glucans, p-(1-4)-glucan (cellulose) và thành phần bên trong (chúng là gọi là ma trận), bắt đầu bao gồm mannoprotein, galacto-mannoprotein, xylo-mannoprotein, glucurono-
mannoprotein và a-(1-3)-glucan. Loại ít được nghiên cứu nhất là a-glucans, là một polyme tuyến tính của glucose (ở Schizosaccharomyces pombe, glucan này chứa khoảng 260 gốc glucose). Polyme này bao gồm hai chuỗi tuyến tính liên kết với nhau chứa khoảng 120 gốc (1-3)-a-D-glucose và (1-4)-a-D-glucose ở hai đầu của phân tử polymer. Người ta tin rằng glucan này cần thiết cho quá trình hình thành hình thái. Một glucan không tan trong nước được phân lập từ CS của sợi nấm Penicillium roqueforti, sau khi sấy khô sẽ mất khả năng hòa tan trong kiềm, nhưng nó có thể hòa tan trong dung dịch lithium clorua 10% trong dimethyl sulfoxide. Khi nghiên cứu glucan bằng phương pháp methyl hóa thu được hai dẫn xuất tri-O-methyl của glucose tương ứng với liên kết 1 - 3 và 1 - "- 4 giữa các monosacarit theo tỷ lệ khoảng 5:2 và dấu vết của một tetra-O- dẫn xuất metyl tương ứng với các gốc mono-nosacarit không khử cuối cùng. Kết quả này cho thấy cấu trúc tuyến tính của các phân tử chỉ chứa 1 -»- 3 và 1 -»- 4 liên kết giữa các gốc glucose. Những dữ liệu này được xác nhận bằng cách phân tích phổ 13C-NMR của polysaccharide, từ đó cấu hình a của tất cả các gốc glucose cũng tuân theo. Các polysacarit như vậy, đặc biệt là sau khi biến đổi hóa học (sulfat hóa hoặc carbosymetyl hóa), có thể làm cho chúng hòa tan trong nước, có thể được sử dụng làm polyme có hoạt tính sinh học và được quan tâm để nghiên cứu chức năng sinh học của chúng trong thành tế bào của nấm và thực vật.
Các loại nấm không tan trong nước (1 -»- 3)-α-glucans trước đây được phân lập từ một số loài nấm bậc cao và bậc thấp, bao gồm Penicillium chrysogenum và các α-glucans liên quan với hai loại liên kết (1-3) và (1 - » - 4), khác nhau về tỷ lệ, được tìm thấy ở thực vật bậc cao (Aconitum kusnezoffii Reichb).
Thành phần cấu trúc - cellulose - là đặc trưng của nấm oomycete, theo phân loại hiện đại, được phân loại là giả nấm. Thành phần monosaccharide của CS bao gồm glucose, mannose, xyloza và đường chiếm ưu thế là glucose, chiếm tới 68%. Thành phần của nấm CS còn bao gồm axit amin, lipid (không quá 3%) và N-acetyl-glucosamine. Glucans, chẳng hạn như 1,3-glucan, tạo thành một phức hợp mạnh với chitin, được gọi là phức hợp chitin-glucan (CGC), tạo nên bộ xương của tế bào nấm. Phức hợp này có mặt trong CS của hầu hết các loại nấm, ngoại trừ hợp tử, đóng vai trò như một đặc điểm hệ thống đáng tin cậy. Ở một số loại nấm, một chất nhầy bao gồm các polysaccharide đại diện bởi ß-1,3-glucans chỉ chứa các đơn vị glucose được kết nối bằng liên kết ß-1,6 với mỗi đơn vị glucose thứ ba đã được tìm thấy trên bề mặt của CS.
Một tỷ lệ khá lớn khối lượng sợi nấm khô là thành tế bào của chúng, cụ thể là 5 đến 15%. Thành phần của nó rất khác nhau và thường rất đặc trưng đối với một số nhóm nấm phân loại nhất định. Điều này có thể được nhìn thấy từ các ví dụ về thành phần của nó trong các đại diện của nấm men, chytrid và mucor - zygomycetes, được đưa ra trong Bảng. 1.3 (Aronson, 1965).
* (Lượng tro không chứa photphat.)
Về mặt cấu trúc, vỏ nấm được xây dựng trên cơ sở hệ thống hai pha, trong đó có các vi sợi nằm trong khối vô định hình của ma trận. Theo kính hiển vi điện tử, nó bao gồm ít nhất hai lớp với các sợi nhỏ định hướng khác nhau. Lớp bên trong thường được định hướng dọc theo trục chính của ô, lớp bên ngoài - nghiêng một góc với nó (Hình 1.1). Vỏ men thường có nhiều lớp, trong đó mannan nằm ở lớp dày bên ngoài và glucan ở lớp mỏng bên trong. Ở nấm thủy sinh, chẳng hạn như ở Allomyces, vỏ tạo thành các vách ngăn giả - pseudosepta, giống như nan hoa của bánh xe. Ở thú có túi và nấm basidiomycetes, người ta quan sát thấy các phân vùng thực sự - vách ngăn. Ở thú có túi, vách ngăn giữa các tế bào thường có một lỗ chân lông đơn giản, ở cả hai bên của lỗ chân lông trong quá trình hình thành có thể nhìn thấy một cặp cơ thể Voronin thẩm thấu. Ở basidiomycetes, các lỗ này thường rất phức tạp, được trang bị các mũ - cha mẹ (Hình 1.2). Những lỗ chân lông như vậy được tìm thấy trong thể đậu quả và thể rhizomorphs của một số loài nấm basidiomycetes bậc cao (Moore, 1965; Burnett, 1968). Tuy nhiên, cho đến nay vẫn chưa hoàn toàn rõ ràng liệu sự khác biệt trong cấu trúc các lỗ vách ngăn của asco- và basidiomycetes là do sự liên kết phân loại của chúng hay do cấu trúc đơn bội và đồng nhân của bộ gen thú có túi và do bản chất di- và dị nhân của basidiomycetes. Chưa có nghiên cứu nào như vậy, nhưng tầm quan trọng của chúng đối với các công trình tiến hóa và phân loại ở vương quốc nấm là rất cơ bản.
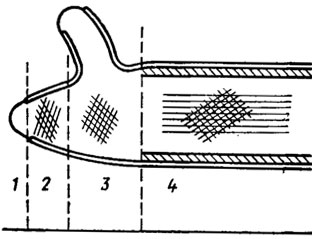

Nhiều thông tin mới đã được bổ sung vào nghiên cứu về lỗ vách ngăn của nấm bằng các nghiên cứu ở cấp độ vi điện tử (Flegler và cộng sự, 1976; Kamaletdinova và Vasiliev, 1982). Đầu tiên, người ta biết rằng dolipores của basidiomycetes là những cấu trúc dường như cách ly hoàn toàn các tế bào sợi nấm với nhau cho đến khi chúng bắt đầu hình thành thể đậu quả (Flegler et al., 1976). Sự phân lập này được thực hiện bằng các enzyme phân giải protein hòa tan và đồng thời các nút hai bên ưa thẩm thấu (chứa protein và lipid), chúng sẽ biến mất vào thời điểm quả thể được hình thành. Sự biến mất của chúng đi kèm với sự thủng của các thể đơn bào và sự liên lạc giữa các tế bào sợi nấm được phục hồi.
Những quan sát gần đây về vách ngăn trong thể quả của các loài nấm đĩa (ví dụ, Peziza badia - Kamaletdinova, Vasiliev, 1982) cho thấy các cấu trúc tương tự ngăn cách các tế bào sợi nấm cũng tồn tại ở lớp nấm có túi. Các thể voronin tham gia vào quá trình hình thành của chúng, được hình thành theo sự xâm lấn hình bình của màng tế bào, các chất thẩm thấu trong đó (thể Voronin) được giải phóng gần vách ngăn và nằm ở lỗ vách ngăn, dần dần thâm nhập vào đó, tạo ra một thiết bị đóng nó lại. Sự xâm nhập sâu hơn của cơ thể Voronin vào lớp màng trinh phía trên dường như bị ngăn chặn bởi một cấu trúc đục lỗ đặc biệt nằm phía trên lỗ vách ngăn trong tế bào mẹ của bao hoạt dịch trong tương lai, cấu trúc này hoàn toàn tách biệt khỏi sự xâm nhập của các bào quan dưới kính hiển vi từ lớp dưới màng trinh . Các cấu trúc cách điện tương tự cũng được quan sát thấy ở các paraphyses hình thành trong quả thể.
Cấu trúc tương tự cũng hiện diện trong sợi nấm hình thành bào tử của nấm không hoàn hảo chứa thể Voronin, ví dụ như ở loài Arthrobotrys conoides. Ở loài deuteromycete Penicillium chrysogenum sản xuất penicillin, một cấu trúc được tìm thấy ở vách ngăn tương ứng chính xác với cấu trúc được tìm thấy bởi Kamaletdinova và Vasiliev ở discomycete Peziza badia (Kurilovich et al., 1980).
Nền tảng khung xương của vỏ nấm bao gồm các polysaccharide tinh thể: cellulose, chitin, chitosan, mannan, glucans và các loại khác. Tất cả chúng đều có cấu trúc tuyến tính với các liên kết β-1,4 của các thành phần ban đầu - monome hexose, amino- và acetaminohexose. Dựa trên kết quả xét nghiệm vi hóa học (nhuộm đỏ ruthenium), trước đây người ta tin rằng thành tế bào của nấm có chứa pectin. Tuy nhiên, kết quả phân tích hóa học không khẳng định monome axit galacturonic có trong cấu trúc của pectin (Aronson, 1965).
Chitin và chitosan đối với hầu hết các loại nấm có đặc điểm vỏ là polyme chứa nitơ. Đồng thời, chitin của nấm rất giống với chitin của côn trùng và động vật giáp xác, điều này đã được xác nhận bằng mẫu nhiễu xạ tia X của chúng. Tuy nhiên, chitin của nấm có ít nitơ hơn so với chitin của động vật và methylpentose, được gọi là mycetose, được tìm thấy trong số các thành phần của nó. Chitin trong nấm có thể được phát hiện vi mô bằng phương pháp Van Wisseling, sử dụng phương pháp khử acetyl một phần bằng kiềm và phản ứng sau đó với chitosan và bằng nhiễu xạ tia X. Nó không chỉ được tìm thấy ở các loài nấm oomycetes, chẳng hạn như nấm saprolegnia và nấm peronosporous. Trước đây người ta tin rằng chitin không có trong nấm men mà nó được chứa trong các vách ngăn của tế bào - vách ngăn của Saccharomycetes (Kulaev, 1975).
Hiện nay người ta đã xác định rằng chitin có thể được tìm thấy ở Chitrydiales, Monoblepharidales, Protomycetales, Hyphochyiridiales, ở tất cả các loài Endomycetales, Blastocladiales, Mucorales, Entomophthorales, ở tất cả các loài thú có túi và basidiomycetes cũng như các loài nấm bất toàn (deuteromycetes) có nguồn gốc từ chúng. Ngoại lệ là Oomycetes, có cellulose trong số các polysaccharide trong vỏ của chúng, chất này hoàn toàn không có ở các đại diện của nấm men.
Gần đây, liên quan đến việc xác định khả năng sử dụng thực tế chitin nấm để tổng hợp polyme, đã xuất hiện khá nhiều nghiên cứu với số liệu nghiên cứu chi tiết hơn về chitin này so với chitin giáp xác (Bảng 1.4). Acetate và D-glucosamine có ít nhất trong Aspergillus niger, điều này được giải thích là do hàm lượng pentose và glucose trong β-glucan và hai α-glucans của vỏ nấm cao hơn. Phân tích các mẫu nhiễu xạ cho thấy cấu trúc tinh thể chitin giống hệt nhau ở nấm và cua, với trật tự lớn hơn một chút ở loài sau. Ngoài ra, chitin của nấm, trái ngược với cấu trúc lớp chitin của cua, có cấu trúc dạng sợi (Feofilova et al., 1980).

Chitosan, chất thay thế chitin trong một số loại nấm nhầy và được tìm thấy trong quá trình thủy phân axit yếu bằng phản ứng với chitosan sulfate, ngoài Mucor rouxii, còn được tìm thấy trong Phycomyces blakesleeanus. Số lượng nhóm acetyl trong đó hóa ra là khác nhau và thay đổi tới 0 ở M. rouxii. Trong số các aminopolysacarit khác, một polyme galactosamine có nhóm amino tự do có khả năng liên kết phốt phát, có cấu trúc kiểu chitosan, được phân lập từ thành tế bào của Neurospora crassa, N. sitphitla, A. niger và Botrytls drierea (Aronson, 1965). Một số loại nấm có chứa polyme của các loại đường amin liên kết với mannan, glucan và protein.
Cellulose trong thành tế bào nấm thường không xuất hiện đồng thời với chitin. Ngoại lệ là một loài Rhizidiomyces thuộc bộ nấm Hyphochytrid, chúng được tìm thấy đồng thời. Cellulose được phát hiện trong nấm từ các tế bào trứng thuộc bộ Acrasiales, Lagenidiales, Saprolegniales, Leptomitales và Peronosporales (Aronson, 1965).
Trong số các loại nấm sống ở môi trường nước, cellulose thường chỉ được tìm thấy ở nhóm có bào tử biflagellate. Blastocladiales và Monoblepharidales, những loài có bào tử đơn roi, không có nó. Ngoại lệ là Rhizidiomyces thuộc bộ Hyphochytridiales, có cả chitin và cellulose và được Nable coi là dạng chuyển tiếp giữa Chytridiales và Blastocladiales mang chitin và oomycetes chứa cellulose, là điều dễ hiểu. Dạng roi đơn này có roi bào tử có lông thay vì dạng roi như ở Blastocladiales và Monoblepharidales. Điều gây tò mò là cấu trúc lông nhung của roi lông của oomycetes giống với cấu trúc của roi của vi khuẩn, trong khi những roi giống như roi hoàn toàn giống với roi của roi.
Cellulose trong nấm có thể dễ dàng được phát hiện bằng phương pháp vi hóa bằng cách sử dụng thuốc thử Speitzer hoặc thuốc thử bao gồm dung dịch iốt trong kali iodua với việc bổ sung dung dịch axit sulfuric 70%.
Glucan trong nấm rất phong phú và khác với cellulose về cấu trúc. Monome của chúng cũng là glucose. Tuy nhiên, glucan nấm men được nghiên cứu nhiều nhất có cấu trúc tinh thể kém hơn cellulose. Độ kết tinh của glucan tăng lên do sự hình thành hydroglucan khi chúng được xử lý bằng axit sulfuric. Các glucan tương tự liên kết với protein đã được tìm thấy trong nấm men và Penicilliunt notatum (Aronson, 1965). Một glucan khác là một phần của vỏ nấm là callose, tương tự như chất được tìm thấy trong ống rây của thực vật bậc cao và được nhuộm mạnh bằng thuốc nhuộm cơ bản, tức là, nó có tính axit và trái ngược với cellulose, có β-1, liên kết 3-glucosidic. Một loại glucan tương tự có thể nhuộm bằng thuốc nhuộm cơ bản đã được tìm thấy ở Sclerofinia. Glucans cũng được tìm thấy trong Aspergillus fischeri, Allomyces macrogynus và Neurospora crassa. Nấm cũng chứa các dị thể polysaccharid bao gồm các monome của nhiều loại đường khác nhau, đặc biệt phổ biến ở chi Penicillium. Ở dạng giống nấm men, gây bệnh cho động vật, như Coccidioides và Cryptococcus, các polysaccharide tương tự nhưng có tính axit là một phần của các viên nang bao quanh tế bào của chúng. Ví dụ về glucans và các sản phẩm trùng hợp khác của monosacarit và axit đường tạo nên thành tế bào và chất dự trữ của nấm được đưa ra trong Bảng. 1.5.

Mannans là các polysacarit bao gồm các monome mannose. được tìm thấy đặc biệt nhiều trong nấm men và chúng thường thấy ở các loài nấm men sống trên bề mặt tầng sinh gỗ dưới vỏ cây. Những dạng này bao gồm Hansenula, sống dưới vỏ cây lá kim; Trong viên nang bao quanh tế bào của các loại nấm men này, polysaccharide hiện diện ở dạng phosphomannan. Polysacarit ưa nước và nhầy nhụa này dính cùng với nấm men vào các sợi lông bao phủ cơ thể bọ cánh cứng, và theo cách này, nấm men được chuyển từ cây này sang cây khác với sự trợ giúp của chúng (Wickerham và Barton, 1961). Mannans chưa được tìm thấy ở nấm hình thành sợi nấm, nhưng mannose được tìm thấy trong dịch thủy phân của thành tế bào của chúng.
Nấm cũng chứa các polysaccharide chứa galactose, 6-deoxypentose, methylpentose và thường gặp nhất là fructose, đặc biệt là trong họ mucoraceae. Trong số các pentose được tìm thấy trong thành tế bào của Penicillium chrysogenum có 6-deoxypentose, rhamnose và xyloza, cũng được tìm thấy trong nấm bùi nhùi Polysiictus sanguineus.
Phức hợp polysaccharide-protein đã được phát hiện trong nấm men, ví dụ như ở Candida albicans, tác nhân gây bệnh tưa miệng ở trẻ sơ sinh. Phức hợp mannan-protein được tìm thấy ở Saccharomyces.
Lipid trong nấm rất khác nhau về số lượng tùy thuộc vào điều kiện môi trường và độ tuổi nuôi cấy. Đôi khi số lượng của chúng đạt tới 35-36% khối lượng chất khô của tế bào. Trong nấm men, hơn 3% lipid được tìm thấy trong màng của chúng. Chúng cũng được tìm thấy trong vỏ của nấm nhầy, ví dụ như Mycor rouxii và Phycomyces, trong bào tử chứa khoảng 25% lipid từ khối lượng khô của chúng. Chúng có thể được chứa ở đó dưới dạng dạng biểu bì (Aronson, 1965).
Các sắc tố cũng có mặt với số lượng đáng kể trong thành tế bào của nấm. Các sắc tố, đặc biệt là sắc tố đen melanin, thường tập trung ở một lớp đặc biệt, thường được tìm thấy trong thành tế bào của sợi nấm hoặc trong vỏ bào tử của nhiều loại nấm. Lớp melanin như vậy hiện diện trong màng ascospore của Neurospora tetrasperma (Aronson, 1965).
Melanin không có ở những loại nấm có hoạt tính polyphenol oxidase thấp, tham gia vào quá trình sinh tổng hợp và có ưu thế là dehydrogenase hoạt động trong quá trình trao đổi chất. Những loại nấm như vậy, bao gồm đại diện của các chi Fusarium, Trichothecium, Arthrobotris, Cephalosporium và nhiều loại khác, thường có đặc điểm là cấu trúc bào tử có màu hồng hoặc cam, tùy thuộc vào lượng carotenoid dồi dào, chúng đảm nhận vai trò bảo vệ ánh sáng. sàng lọc và chống oxy hóa, thuộc về melanin trong nấm có màu sẫm. Mối liên hệ giữa sự hiện diện của carotenoid và mức độ hoạt động dehydrogenase cao được giải thích bởi thực tế là các điều kiện tối ưu của chế độ oxy hóa khử cho hoạt động của dehydrogenase trùng với các điều kiện tối ưu cho quá trình sinh tổng hợp carotenoid và các sản phẩm shunt terpenoid khác. Các sắc tố carotenoid thường không còn là một phần của thành tế bào nấm mà nằm ở màng tế bào hoặc trong các giọt lipid phân tán trong tế bào chất. Trong một số trường hợp, các sắc tố hoàn toàn đặc biệt được tìm thấy ở nấm, chẳng hạn như ommochromes, sắc tố mắt của côn trùng, ở polypore Pycnoporus (Polyporus) cinnabarinus (Shivrina, 1965) hoặc phycobilin ở loài russula Russula emetica và R. paludosa (Efimenko, 1972), là một phần của hệ thống quang hợp ở tảo xanh lam và tảo đỏ.
Actinomycetes (Actinomycetales, từ tiếng Hy Lạp aktis - ray, mykes - nấm) là vi khuẩn phân nhánh thuộc ngành Actinobacteria. Chúng là một phần của hệ vi sinh vật bình thường trong hệ tiêu hóa của động vật có xương sống và động vật không xương sống trên cạn, đồng thời hiện diện rất nhiều trong đất và đóng một vai trò quan trọng trong hệ sinh thái và chu trình của các chất trong đất.
Những vi sinh vật này là tác nhân gây ra nhiều bệnh lý cơ hội - những bệnh phát sinh do chức năng hệ thống miễn dịch của cơ thể bị suy giảm. Actinomycetes được sử dụng rộng rãi trong công nghệ sinh học vì chúng là nguồn cung cấp một số chất kháng khuẩn và chống ung thư.
Cơm. 1. Streptomycetes tổng hợp một lượng lớn thuốc kháng khuẩn và chống ung thư.
Cấu trúc của xạ khuẩn: tại sao chúng là vi khuẩn mà không phải nấm?
1. Tổ chức vật chất di truyền
Vật liệu di truyền của Actinomycetes được chứa trong một phân tử axit deoxyribonucleic, có hình vòng và nằm tự do trong tế bào chất - cùng một dạng tổ chức vật liệu di truyền, được gọi là nucleoid, đặc trưng của các vi khuẩn khác. Ở nấm, vật chất di truyền được tổ chức và là một phần của nhân tế bào.
DNA của xạ khuẩn chứa một số lượng lớn các cặp GC (65-75% tổng số nucleotide). Đặc tính này không đổi, không phụ thuộc vào đột biến nên được sử dụng trong phân loại vi sinh vật. Hàm lượng các cặp GC này làm cho DNA của Actinomycetes rất bền; do đó, việc phân tích DNA của Actinomycetes mất nhiều thời gian hơn so với các vi khuẩn khác.

Cơm. 2. Sơ đồ cấu trúc thành tế bào của vi khuẩn Gr+.

Cơm. 3. Actinomycetes, nhuộm Gram.
Actinomycetes có thành tế bào vi khuẩn dày đặc, nằm bên ngoài màng tế bào chất và gây ra hiện tượng nhuộm Gram dương tính. Giống như các vi khuẩn gram dương khác, nó bao gồm vài chục lớp polymer murein (peptidoglycan), được thấm axit teichoic và lipoteichoic. Axit lipoteichoic được neo trong màng tế bào chất của vi khuẩn và kết nối nó với thành tế bào. Axit Teichoic mang điện tích âm cho thành tế bào. Thành tế bào của nấm bao gồm các polyme khác - chitin và glucan.
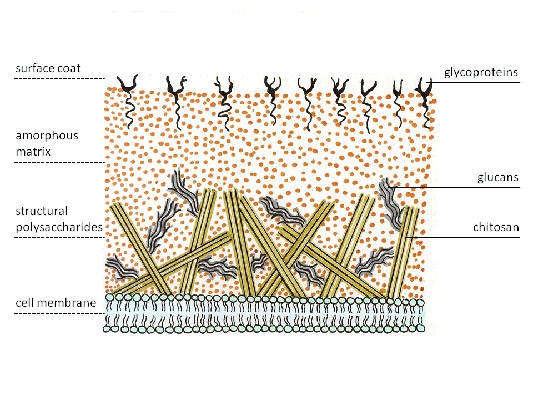
Cơm. 4. Vách tế bào nấm.
3. Bào quan tế bào
Actinomycetes, giống như các vi khuẩn khác, không có bào quan có màng. Actinomycetes có ribosome 70S, trong khi nấm có ribosome 80S, cũng như các sinh vật nhân chuẩn khác.
4. Sự phát triển của khuẩn lạc
Sự hình thành sợi nấm trong quá trình sinh trưởng là điều mà xạ khuẩn giống với nấm nhất. Sợi nấm trong trường hợp xạ khuẩn là tập hợp các sợi nấm phân nhánh. Các sợi nấm được phân chia bằng vách ngăn thành các tế bào vi khuẩn dài chứa một số nucleoid. Các vách ngăn ở một số loài có thể chạy theo hướng vuông góc với nhau. Sợi nấm phân nhánh bằng cách nảy chồi.
Sợi nấm phát triển thành chất nền (đất, bùn hoặc môi trường dinh dưỡng) được gọi là sợi nấm chất nền. Nó cung cấp chất dinh dưỡng cho thuộc địa. Sợi nấm trên không nổi lên trên bề mặt, tạo cho khuẩn lạc một "độ mịn" - nó hình thành các bào tử, cũng như cái gọi là "chất chuyển hóa thứ cấp" (ngược lại với "chất chuyển hóa sơ cấp" của sợi nấm cơ chất), trong đó có rất nhiều chất kháng khuẩn.
Vòng đời và sinh lý của Actinomycetes
Trong vòng đời, hầu hết xạ khuẩn đều hình thành bào tử. Một số xạ khuẩn sinh sản bằng cách phân mảnh sợi nấm.
1. Bào tử
Bào tử Actinomycete có nguồn gốc từ sợi nấm trên không. Đây là những ngoại bào tử - chúng phát triển bên ngoài tế bào mẹ. Các sợi nấm trên không nơi bào tử phát triển được gọi là vật mang bào tử. Các bào tử có thể được chứa trong một lớp dày ở phần cuối của bào tử - bào tử (ví dụ, ở streptomycetes, Actinoplanes và plymelia), hoặc có thể nằm trong một chuỗi dọc theo bào tử (ví dụ, ở nocardia và Actinomadura).
Theo số lượng bào tử được hình thành, xạ khuẩn được chia thành:
- Đơn bào tử (ví dụ, Saccaromonospora, Micromonospora, Thermomonospora) - hình thành các bào tử đơn lẻ, thường bằng cách nảy chồi và sau đó tách khỏi sợi nấm mẹ bằng một vách ngăn;
- Oligosporous (ví dụ, Actinomadura) - tạo thành chuỗi bào tử ngắn dọc theo chất mang bào tử;
- Đa bào tử (hầu hết các loại xạ khuẩn khác, ví dụ Streptomyces, Frankia, Geodermatophilus) - tạo thành nhiều bào tử được bao bọc trong túi bào tử.

Cơm. 5. Bào tử xạ khuẩn thuộc chi Frankia.
Các bào tử Actinomycete có thể di động - trong trường hợp này, bào tử có một lá cờ và có thể di chuyển (bào tử của Actinoplanes, geodermatophiles và dermatophiles). Trong hầu hết các trường hợp, bào tử không di động và lây lan nhờ gió, nước hoặc động vật.

Cơm. 6. Dermatophils, kính hiển vi ánh sáng.
Sự hình thành bào tử ở xạ khuẩn đặc biệt hoạt động trong điều kiện không thuận lợi. Khả năng chịu nhiệt của bào tử thấp hơn so với bào tử của các vi khuẩn khác, nhưng chúng có thể chịu được khô không kém các loại khác và do đó có ý nghĩa thích nghi rất lớn. Actinomycetes thống trị các vi sinh vật khác trong đất sa mạc khô.
Sự nảy mầm của bầy đòi hỏi độ ẩm nhất định ở môi trường bên ngoài. Khi có nước, bào tử phồng lên, các enzyme được kích hoạt và các quá trình trao đổi chất được bắt đầu, kèm theo sự giải phóng các ống tăng trưởng (cơ thể vi khuẩn trong tương lai) và quá trình tổng hợp axit nucleic.
2. Kiểu thở
Hầu hết các xạ khuẩn đều là vi khuẩn hiếu khí (chúng cần oxy để duy trì sự sống). Vi khuẩn kỵ khí tùy ý (vi khuẩn có thể sống trong cả điều kiện có và không có oxy) được tìm thấy ở những loài có giai đoạn sợi nấm ngắn, sinh sản bằng cách phân mảnh sợi nấm.
3. Kháng axit
Actinomycetes có khả năng chịu axit - khả năng kháng trong môi trường axit, cho phép chúng sống trong đất rừng bão hòa axit. Khả năng kháng axit trong phòng thí nghiệm có thể được xác định bằng cách nhuộm chế phẩm có chứa xạ khuẩn theo Ziehl-Neelsen (fuchsin sau đó ăn mòn bằng axit sulfuric và nhuộm bằng xanh methylene). Hầu hết xạ khuẩn có màu này không bị mất màu sau khi ăn mòn bằng axit và giữ được màu đỏ tươi. Môi trường kiềm không thuận lợi cho những vi khuẩn này: ở độ pH cao, chúng dễ hình thành bào tử.
4. Đặc điểm trao đổi chất

Cơm. 7. Actinomycetes hiếu khí hình thành sắc tố trên mặt thạch nghiêng. Từ trái sang phải: Actinomadura madurae, Nocardia tiểu hành tinh, Micromonospora.
Sự hình thành các “chất chuyển hóa thứ cấp” bằng sợi nấm trên không đã được đề cập ở trên. Trong số đó:
- sắc tố tạo ra các màu sắc khác nhau của sợi nấm trên không khi phát triển trên môi trường;
- các chất dễ bay hơi tạo mùi đặc trưng cho đất sau mưa, nước đọng, mùi da của một số loài động vật;
- thuốc kháng sinh:
Một. thuốc chống nấm – polyene;
b. kháng khuẩn - ví dụ streptomycin, erythromycin, tetracycline, vancomycin;
c. thuốc chống ung thư - anthracycline, bleomycin.
Actinomycetes sống ở đâu?
Actinomycetes được tìm thấy với số lượng lớn nhất trong đất và có số lượng sợi nấm ít hơn đáng kể so với bào tử. Chúng đóng vai trò quan trọng trong việc hình thành mùn, phân hủy các chất hữu cơ mà các vi khuẩn khác khó sử dụng. Về vấn đề này, xạ khuẩn được sử dụng làm vi sinh vật chỉ thị vệ sinh trong các vấn đề vệ sinh và dịch tễ học: việc phát hiện chúng với số lượng lớn trong đất hoặc nước cho thấy sự hiện diện của phân trộn trong chất nền tương ứng.

Cơm. 8. Actinomycetes trong phân trộn.
Actinomycetes là cộng sinh của nhiều loại thực vật, giúp chúng cố định đạm. Đồng thời, nhiều vi sinh vật thuộc lớp này là tác nhân gây bệnh cho cây trồng.

Cơm. 9. Bệnh liên cầu khuẩn khoai tây.
Chúng cũng được tìm thấy trong hệ vi sinh bình thường của hệ tiêu hóa của một số động vật, từ giun đốt đất (như giun đất) đến vật nuôi lớn.
Những vi sinh vật này giúp phân hủy cellulose, chất có nhiều trong thực phẩm thực vật. Ở người, xạ khuẩn được tìm thấy trong khoang miệng (nướu và mảng bám), ruột (phần xa của ruột già), trên da (mặt, cánh mũi, sau tai, giữa các ngón tay) và trong hệ hô hấp. (chủ yếu ở đường hô hấp trên).
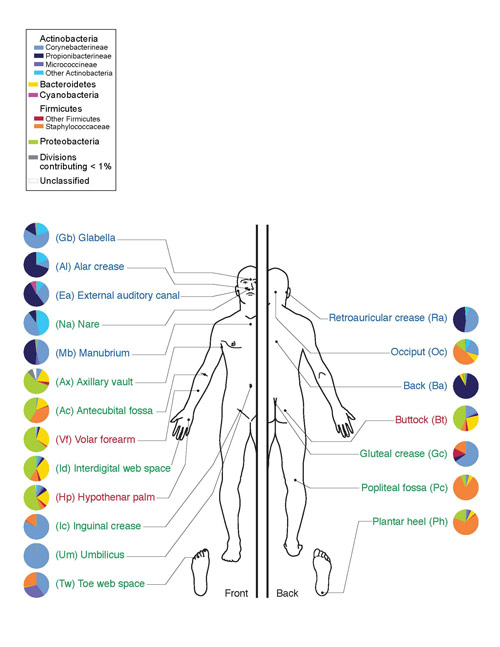
Cơm. 10. Hệ vi sinh vật trên da người. Ngành Actinobacteria được biểu thị bằng màu xanh lam, lớp Actinomycetes được biểu thị bằng màu xanh lam sáng.
Actinomycetes, nếu khả năng phản ứng miễn dịch của cơ thể bị giảm, có thể gây ra bệnh Actinomycosis - bệnh cơ hội bao gồm sự hình thành u hạt Actinomycosis - sự tích tụ của cơ thể vi khuẩn giống như hạt lưu huỳnh màu vàng ("drusen"), được bao quanh bởi các tế bào có thẩm quyền miễn dịch.

Phản ứng viêm dẫn đến sự tan chảy của u hạt, hình thành các lỗ rò, dẫn đến thủng các cơ quan và lây lan vi khuẩn qua đường máu.

Cơm. 11. Actinomycosis drusen, nhuộm Gram.

Cơm. 12. Bệnh Actinomycosis ở hàm trên ở bò.
Cơm. 13. Bệnh Actinomycosis hàm trên ở người.