Chương 1
CƠ SỞ VỀ SINH LÝ TẾ BÀO
I. anh bạn
Màng huyết tương . Tế bào động vật được bao bọc bởi một màng sinh chất (Hình 1.1). Chúng ta sẽ tập trung vào cấu trúc của nó, rất giống với cấu trúc của nhiều màng nội bào, chi tiết hơn một chút. Chất nền chính của màng bao gồm lipid, chủ yếu là phosphatidylcholin. Những lipid này bao gồm một nhóm đầu ưa nước mà các chuỗi hydrocarbon kỵ nước dài được gắn vào. Trong nước, các lipid như vậy tự hình thành một màng hai lớp dày 4–5 nm, trong đó các nhóm ưa nước hướng vào môi trường nước và các chuỗi hydrocarbon kỵ nước được sắp xếp thành hai hàng, tạo thành pha lipid khan. Màng tế bào là lớp lipid kép thuộc loại này và chứa glycolipids, cholesterol và phospholipids (Hình 1.2). Phần ưa nước của glycolipid được hình thành bởi oligosacarit. Glycolipids luôn nằm ở bề mặt ngoài của màng sinh chất, với phần oligosaccharide của phân tử được định hướng giống như một sợi tóc ngâm trong môi trường. Nằm rải rác giữa các phospholipid với số lượng gần như bằng nhau, các phân tử cholesterol ổn định màng. Sự phân bố của các loại lipid khác nhau ở lớp bên trong và bên ngoài của màng là không giống nhau, và ngay cả trong một lớp cũng có những khu vực tập trung một số loại lipid nhất định. Sự phân bố không đồng đều này
Cơm. 1.1. Sơ đồ tế bào thể hiện các bào quan quan trọng nhất
có lẽ có một số ý nghĩa chức năng, nhưng vẫn chưa rõ ràng.
Các yếu tố chức năng chính được nhúng trong ma trận lipid tương đối trơ của màng là sóc(Hình 1.2). Protein theo trọng lượng chiếm từ 25 đến 75% ở các màng khác nhau, nhưng do phân tử protein lớn hơn nhiều so với phân tử lipid nên 50% trọng lượng tương đương với tỷ lệ 1 phân tử protein trên 50 phân tử lipid. Một số protein xuyên qua màng từ bề mặt ngoài đến bề mặt bên trong, trong khi một số protein khác được cố định trong một lớp. Các phân tử protein thường được định hướng sao cho các nhóm kỵ nước của chúng được gắn vào màng lipid và các nhóm ưa nước phân cực trên bề mặt màng được gắn vào pha nước. Nhiều protein ở bề mặt ngoài của màng là glycoprotein; các nhóm sacarit ưa nước của chúng phải đối mặt với môi trường ngoại bào.
Hệ thống màng của các bào quan nội bào .
Khoảng một nửa thể tích tế bào được chiếm giữ bởi các bào quan được phân lập từ bào tương bằng màng. Tổng diện tích bề mặt của màng tế bào nội bào lớn hơn ít nhất 10 lần so với bề mặt của màng sinh chất. Hệ thống màng được sử dụng rộng rãi nhất là mạng lưới nội chất,đại diện cho một mạng

Cơm. 1.2.Sơ đồ biểu diễn màng sinh chất. Protein được nhúng vào lớp kép phospholipid, một số protein xuyên qua lớp kép, trong khi một số khác chỉ bám vào lớp ngoài hoặc lớp trong.
các ống rất phức tạp hoặc các cấu trúc kéo dài giống như túi; khu vực rộng lớn của mạng lưới nội chất được đính đầy ribosome; một mạng lưới như vậy được gọi là dạng hạt hoặc thô (Hình 1.1). bộ máy Golgi cũng bao gồm các phiến có màng bao bọc, từ đó các mụn nước hoặc mụn nước vỡ ra (Hình 1.1). Lysosome và peroxisome- Đây là những mụn nước nhỏ chuyên biệt. Trong tất cả các bào quan khác nhau này, màng và không gian bao quanh nó chứa các bộ enzyme cụ thể; Bên trong các bào quan, các sản phẩm trao đổi chất đặc biệt được tích lũy, được sử dụng để thực hiện các chức năng khác nhau của các bào quan.
Cốt lõiVà ty thể khác nhau ở chỗ mỗi bào quan này được bao quanh bởi hai màng. Nhân chịu trách nhiệm điều khiển động học của quá trình trao đổi chất; màng gấp bên trong của ty thể là nơi diễn ra quá trình trao đổi chất oxy hóa; ở đây, do quá trình oxy hóa pyruvate hoặc axit béo, hợp chất năng lượng cao adenosine triphosphate (ATP, hoặc ATP) được tổng hợp.
Bộ xương tế bào . Tế bào chất bao quanh các bào quan không thể được coi là vô định hình; nó được thấm vào bởi một mạng lưới tế bào. Bộ khung tế bào bao gồm các vi ống, sợi Actin và sợi trung gian (Hình 1.1). Vi ống có đường kính ngoài khoảng 25 nm; chúng được hình thành, giống như một loại polyme thông thường, là kết quả của sự lắp ráp các phân tử protein tubulin. Sợi actin- các sợi co bóp, nằm ở lớp gần màng và khắp tế bào, chủ yếu tham gia vào các quá trình liên quan đến chuyển động. Sợi trung gian bao gồm các khối có thành phần hóa học khác nhau trong các loại tế bào khác nhau; chúng tạo thành nhiều mối liên kết khác nhau giữa hai thành phần khung tế bào khác đã đề cập ở trên. Các bào quan và màng sinh chất cũng liên kết với bộ xương tế bào, bộ xương này không chỉ duy trì hình dạng của tế bào và vị trí của các bào quan trong đó mà còn quyết định những thay đổi về hình dạng của tế bào và khả năng di chuyển của nó.
Cytosol . Khoảng một nửa thể tích tế bào bị chiếm giữ bởi bào tương. Vì trọng lượng của nó là khoảng 20% protein nên nó giống gel hơn là dung dịch nước. Các phân tử nhỏ, bao gồm cả hữu cơ và vô cơ ion, hòa tan trong pha nước. Sự trao đổi ion xảy ra giữa tế bào và môi trường (không gian ngoại bào); những quá trình trao đổi chất này sẽ được thảo luận trong phần tiếp theo. Nồng độ ion trong không gian ngoại bào được duy trì với độ chính xác đáng kể ở mức không đổi; nồng độ nội bào của mỗi ion cũng có nồng độ riêng, khác với nồng độ bên ngoài tế bào (Bảng 1.1). Cation có nhiều nhất trong môi trường ngoại bào là Na+ trong tế bào nồng độ của nó thấp hơn 10 lần. Ngược lại, nồng độ K+ bên trong tế bào là cao nhất; bên ngoài tế bào thì nồng độ này thấp hơn nhiều. Sự chênh lệch lớn nhất giữa nồng độ ngoại bào và nội bào tồn tại đối với Ca 2+, nồng độ ion tự do bên trong tế bào thấp hơn bên ngoài ít nhất 10.000 lần. Không phải tất cả các ion đều được hòa tan trong bào tương; một số ion được hấp phụ trên protein hoặc lắng đọng trong các bào quan. Ví dụ, trong trường hợp Ca 2+, số ion liên kết nhiều hơn số ion tự do. Hầu hết các protein tế bào là enzyme, với sự tham gia của nhiều quá trình chuyển hóa trung gian được thực hiện: glycolysis và gluconeogen, tổng hợp hoặc phá hủy axit amin, tổng hợp protein trên ribosome (Hình 1.1). Cytosol cũng chứa các giọt lipid và hạt glycogen, đóng vai trò dự trữ các phân tử quan trọng.
Bảng 1.1.Nồng độ ion trong và ngoại bào trong tế bào cơ của động vật đẳng nhiệt. MỘT – – “anion tế bào trọng lượng phân tử cao”
|
Nồng độ nội bào |
Nồng độ ngoại bào |
||
|
Na+ |
12 mmol/l |
Na+ |
145 mmol/l |
|
155 mmol/l |
K+ |
4 mmol/l |
|
|
Ca 2+ |
10 –7 10 –8 mmol/l |
Ca 2+ |
2 mmol/l |
|
C l – |
4 mmol/l |
C1 – |
120 mmol/l |
|
HCO3 – |
8 mmol/l |
HCO3 – |
27 mmol/l |
|
MỘT - |
155 mmol/l |
Người khác cation |
5 mmol/l |
|
Điện thế nghỉ –90 mV |
|||
1.2. Trao đổi chất giữa tế bào và môi trường
Chúng tôi đã mô tả ngắn gọn cấu trúc của tế bào để có thể sử dụng mô tả này để xem xét những điều cơ bản về sinh lý tế bào. Trong mọi trường hợp, tế bào không thể được coi là một thực thể tĩnh, vì có sự trao đổi chất liên tục giữa các ngăn nội bào khác nhau, cũng như giữa các ngăn và môi trường. Cấu trúc tế bào ở trạng thái cân bằng động và sự tương tác của các tế bào với nhau và với môi trường bên ngoài là điều kiện cần thiết để duy trì sự sống của một sinh vật hoạt động. Trong chương này chúng ta sẽ xem xét các cơ chế cơ bản của sự trao đổi đó. Trong các chương tiếp theo, các cơ chế này sẽ được xem xét khi áp dụng cho tế bào thần kinh và các chức năng của nó;
tuy nhiên, các cơ chế tương tự làm nền tảng cho hoạt động của tất cả các cơ quan khác.
Khuếch tán.Quá trình chuyển động đơn giản nhất của vật chất là khuếch tán. Trong dung dịch (hoặc chất khí), các nguyên tử và phân tử chuyển động tự do và sự khác biệt về nồng độ được cân bằng do sự khuếch tán. Hãy xem xét hai thể tích chứa đầy chất lỏng hoặc khí (Hình 1.3), trong đó các chất có nồng độ c 1 và c 2 và được ngăn cách bởi một lớp có diện tích bề mặt A và độ dày d. Dòng vật chất m trong thời gian t mô tả Định luật khuếch tán đầu tiên của Fick:
dm/ dt= DMỘT/ d ( C 1 –С 2)=DMỘT/ dD C(1)
Trong đó D là hệ số khuếch tán, không đổi đối với một chất, dung môi và nhiệt độ nhất định. Ở dạng tổng quát hơn cho sự chênh lệch nồng độ dc ở khoảng cách dx
dm/dt= –D A dc/dx,(2)
dòng chảy qua phần A tỷ lệ thuận với gradient nồng độ dc/dx . Dấu trừ xuất hiện trong phương trình vì sự thay đổi nồng độ theo hướng x là âm.
Khuếch tán là quá trình quan trọng nhất do hầu hết các phân tử trong dung dịch nước di chuyển trên một khoảng cách ngắn. Điều này cũng áp dụng cho sự di chuyển của chúng trong tế bào khi sự khuếch tán không bị cản trở bởi màng. Nhiều chất có thể khuếch tán tự do qua màng lipid, đặc biệt là nước và các khí hòa tan như O 2 và CO 2 . tan trong chất béo

Cơm. 1.3.Sơ đồ khuếch tán định lượng. Hai không gian được ngăn cách bởi một lớp dàydvà diện tích MỘT. C; – nồng độ hạt cao ở phần bên trái của thể tích, C: – nồng độ hạt thấp ở phần bên phải các bộ phận, bề mặt màu hồng- gradient nồng độ trong lớp khuếch tán. Dòng khuếch tán dm/dt – xem phương trình (1)
các chất cũng khuếch tán tốt qua màng; Điều này cũng áp dụng cho các phân tử phân cực có kích thước khá nhỏ, chẳng hạn như ethanol và urê, trong khi đường khó đi qua lớp lipid. Đồng thời, các lớp lipid thực tế không thể thấm qua các phân tử tích điện, kể cả các ion vô cơ. Đối với chất không điện phân, phương trình khuếch tán (1) thường được biến đổi bằng cách kết hợp các đặc tính của màng và chất khuếch tán thành một độ thấm tham số (P):
dm/dt=P AD c.(3)
Trong bộ lễ phục. 1.4 được so sánh tính thấm (P) của màng lipid đối với các phân tử khác nhau.
Khuếch tán qua các lỗ màng . Màng sinh chất (và các màng tế bào khác) không chỉ có khả năng thẩm thấu với các chất khuếch tán qua lớp lipid mà còn với nhiều ion, đường, axit amin và nucleotide. Các chất này xuyên qua màng thông qua các lỗ hình thành protein vận chuyển, ngâm trong màng. Bên trong những protein như vậy có một kênh chứa đầy nước có đường kính nhỏ hơn 1 nm, qua đó các phân tử nhỏ có thể khuếch tán. Chúng di chuyển dọc theo gradient nồng độ và nếu chúng mang điện tích thì chuyển động của chúng qua các kênh cũng được điều chỉnh bởi điện thế màng. Các kênh màng tương đối chọn lọc

Cơm. 1.4.Tính thấm của lớp lipid kép nhân tạo với các chất khác nhau
Liên quan đến loại phân tử có thể đi qua chúng, chẳng hạn, có các kênh kali, natri và canxi, mỗi kênh đều không thấm đối với hầu hết mọi ion ngoại trừ một ion cụ thể. Như là tính chọn lọcđược xác định bởi điện tích hoặc cấu trúc của các vị trí liên kết trong thành kênh, tạo điều kiện thuận lợi cho việc vận chuyển một phân tử cụ thể và ngăn chặn sự xâm nhập của các chất khác qua kênh (Hình 2). 1,5, A) .
Đằng sau hành vi kênh ion màng dễ quan sát vì có thể đo được dòng điện phát sinh từ chuyển động của các ion, ngay cả đối với một kênh đơn lẻ. Nó cho thấy rằng các kênh một cách tự nhiên và với tần số cao thay đổi trạng thái của chúng từ mở sang đóng. Kênh kali được đặc trưng bởi các xung dòng điện có biên độ khoảng 2 pA (2 10 –12 A) và thời lượng vài mili giây (xem Hình 2.12, trang 37) [3]. Trong thời gian này, hàng chục nghìn ion đi qua nó. Sự chuyển đổi của protein từ cấu hình này sang cấu hình khác được nghiên cứu bằng phương pháp nhiễu xạ tia X, quang phổ Mössbauer và cộng hưởng từ hạt nhân (NMR). Do đó, protein có cấu trúc rất năng động, di động và kênh đi qua protein không chỉ là một ống cứng, chứa đầy nước (Hình 1.5, A), mà là một mê cung gồm các nhóm phân tử và điện tích chuyển động nhanh. Đặc tính động này của kênh được phản ánh trong hồ sơ năng lượng của kênh, thể hiện trong hình. 1.5, B. Ở đây, trục hoành biểu thị chiều dài kênh từ dung dịch bên ngoài có nồng độ ion C 0 và điện thế 0 đến dung dịch bên trong có nồng độ C 1 và điện thế E. Trục tọa độ

Cơm. 1.5.A. Sơ đồ protein tạo thành kênh kali gắn trong lớp lipid kép của màng sinh chất. Bốn điện tích âm được cố định trên “bức tường” của kênh. B. Sơ đồ năng lượng của kênh được hiển thị trong Hình. A. Trục hoành biểu thị các giá trị động năng cần thiết để kênh đi qua; dọc theo trục hoành - khoảng cách giữa bề mặt bên trong và bên ngoài của màng. Năng lượng cực tiểu tương ứng với vị trí liên kết của các ion tích điện dương với điện tích âm cố định trên thành kênh. Cực đại năng lượng tương ứng với các trở ngại khuếch tán trong kênh. Cấu trúc của protein kênh được cho là dao động tự phát; các tùy chọn hồ sơ năng lượng được hiển thị bằng các đường nét liền và nét đứt; những dao động này tạo điều kiện thuận lợi rất nhiều cho sự liên kết của các ion khi vượt qua hàng rào năng lượng (đã được sửa đổi)
mức năng lượng ion tại các vị trí liên kết kênh được trình bày; đỉnh trong biểu đồ biểu thị hàng rào thấm mà năng lượng ion phải vượt qua để xuyên qua kênh và điểm “nhúng” trong biểu đồ biểu thị trạng thái tương đối ổn định (liên kết). Bất chấp trở ngại của đỉnh năng lượng, ion có thể xuyên qua kênh nếu cấu hình năng lượng thay đổi một cách tự phát; do đó, ion có thể đột nhiên thấy mình “ở phía bên kia” của đỉnh năng lượng và có thể tiếp tục di chuyển vào trong tế bào. Tùy thuộc vào điện tích, kích thước và mức độ hydrat hóa của ion cũng như khả năng liên kết với cấu trúc thành kênh của nó, cấu hình năng lượng của kênh thay đổi đối với các ion khác nhau, điều này có thể giải thích tính chọn lọc của từng loại kênh riêng lẻ.
Cân bằng khuếch tán của các ion . Sự khuếch tán của các ion khác nhau qua các kênh màng sẽ dẫn đến việc loại bỏ sự khác biệt về nồng độ giữa môi trường ngoại bào và nội bào. Tuy nhiên, có thể thấy từ bảng. 1.1, những khác biệt như vậy vẫn còn tồn tại, do đó phải có một số trạng thái cân bằng giữa quá trình khuếch tán và các quá trình vận chuyển khác qua màng. Hai phần tiếp theo đề cập đến những cách thức đạt được trạng thái cân bằng này. Trong trường hợp các ion, trạng thái cân bằng khuếch tán bị ảnh hưởng bởi điện tích của chúng. Sự khuếch tán của các phân tử không tích điện được đảm bảo bằng sự chênh lệch nồng độ dc và khi nồng độ được cân bằng thì quá trình vận chuyển sẽ dừng lại. Các hạt mang điện còn bị ảnh hưởng bởi điện trường. Ví dụ, khi ion kali rời khỏi tế bào dọc theo gradient nồng độ, nó mang một điện tích dương. Do đó, môi trường nội bào trở nên tích điện âm hơn, dẫn đến sự chênh lệch điện thế trên màng. Điện tích âm nội bào ngăn cản các ion kali mới rời khỏi tế bào và những ion rời khỏi tế bào sẽ làm tăng thêm điện tích trên màng. Dòng ion kali dừng lại khi tác động của điện trường bù lại áp suất khuếch tán do chênh lệch nồng độ. Các ion tiếp tục đi qua màng nhưng với số lượng bằng nhau theo cả hai hướng. Do đó, đối với một sự khác biệt nhất định về nồng độ ion trên màng, sẽ có thế năng cân bằng E ion tại đó dòng ion qua màng dừng lại. Thế cân bằng có thể được xác định dễ dàng bằng cách sử dụng Phương trình Nernst:
Eion= RT/ zF* lnC ra/ C trong(4)
ở đâu R – hằng số khí, T – nhiệt độ tuyệt đối, z – hóa trị ion (âm đối với anion) C ra - nồng độ ion ngoại bào, C trong – nồng độ ion nội bào, F Số Faraday. Nếu chúng ta thay các hằng số vào phương trình thì ở nhiệt độ cơ thể (T = 310 K) thế cân bằng của ion kali E K bằng:
ek= –61 tôiNhật ký B /(5)
Nếu [ K + ngoài ]/[ K + trong ] = 39, như sau từ bảng. 1.1 thì
Ek= –61 m B log 39= –97 mV.
Thật vậy, người ta thấy rằng tất cả các tế bào đều có điện thế màng; trong tế bào cơ của động vật có vú mức độ của nó là khoảng -90 mV. Tùy thuộc vào điều kiện và nồng độ ion tương đối, tế bào có thể có điện thế màng từ -40 đến -120 mV. Đối với ô trong ví dụ trên (Bảng 1.1) tiềm năng nghỉ ngơi, bằng khoảng -90 mV, cho thấy dòng ion kali qua các kênh màng gần như ở trạng thái cân bằng. Điều này không có gì đáng ngạc nhiên, vì trong màng nghỉ, trạng thái mở của các kênh kali rất có thể xảy ra, tức là. màng có tính thấm tốt nhất đối với ion kali. Tuy nhiên, điện thế màng được xác định bởi dòng chảy của các ion khác.
Mức độ dễ dàng mà các hạt không tích điện có thể khuếch tán qua màng được định lượng theo phương trình (3). Tính thấm của các hạt tích điện được mô tả bằng một phương trình phức tạp hơn một chút:
P= tôi RT/ dF(6)
Ở đâu tôi- Khả năng di chuyển của ion trong màng, d - độ dày màng, một R, T và F – các hằng số nhiệt động đã biết. Giá trị độ thấm của các ion khác nhau được xác định theo cách này có thể được sử dụng để tính toán điện thế màng Em , khi các ion kali, natri và clo đồng thời đi qua màng (có độ thấm P K, P Na và PCl tương ứng). Người ta giả định rằng điện thế giảm đồng đều trong màng, do đó cường độ trường không đổi. Trong trường hợp này nó được áp dụng Phương trình Goldman, hay phương trình trường hằng số :
Em= R T/ F * ln(P K + P Na + P Cl )/ (P K + P Na + P Cl )(7)
Đối với hầu hết màng tế bào P K cao hơn khoảng 30 lần so với P Na (xem thêm phần 1.3). Giá trị tương đối PCl thay đỗi lớn; cho nhiều màng PCl nhỏ so với R K , tuy nhiên đối với những người khác (ví dụ: trong cơ xương) PCl , cao hơn đáng kể so với P K.
Vận chuyển tích cực, bơm natri . Phần trước mô tả sự khuếch tán thụ động của các ion và điện thế màng thu được ở nồng độ ion trong và ngoài tế bào nhất định. Tuy nhiên, kết quả của quá trình này là nồng độ ion bên trong tế bào không được tự động ổn định vì màng
điện thế có độ âm điện lớn hơn một chút so với EK, và nhiều so với E Na (khoảng +60 mV). Do sự khuếch tán, nồng độ ion nội bào, ít nhất là kali và natri, phải được cân bằng với nồng độ ngoại bào. Sự ổn định của gradient ion đạt được thông qua vận chuyển tích cực: protein màng vận chuyển các ion qua màng chống lại gradient điện và/hoặc nồng độ, tiêu thụ năng lượng trao đổi chất cho việc này. Quá trình quan trọng nhất của vận chuyển tích cực là hoạt động Na/K – một máy bơm tồn tại ở hầu hết các tế bào;
máy bơm bơm ion natri ra khỏi tế bào đồng thời bơm ion kali vào tế bào. Điều này đảm bảo nồng độ ion natri nội bào thấp và nồng độ ion kali cao (Bảng 1.1). Độ dốc nồng độ của các ion natri trên màng có chức năng cụ thể liên quan đến việc truyền thông tin dưới dạng xung điện (xem Phần 2.2), cũng như duy trì các cơ chế vận chuyển tích cực khác và điều chỉnh thể tích tế bào (xem bên dưới). Do đó, không có gì đáng ngạc nhiên khi hơn 1/3 năng lượng mà tế bào tiêu thụ được dành cho bơm Na/K và ở một số tế bào hoạt động mạnh nhất, tới 70% năng lượng được sử dụng cho hoạt động của nó.
Protein vận chuyển Na/K là ATPase. Ở bề mặt bên trong của màng, nó phân hủy ATP thành ADP và photphat (Hình 1.6). Năng lượng của một phân tử ATP được sử dụng để vận chuyển ba ion natri từ tế bào và đồng thời hai ion kali vào tế bào, tức là tổng cộng một điện tích dương được lấy ra khỏi tế bào trong một chu kỳ. Vì vậy, bơm Na/K là mang điện(tạo ra dòng điện qua màng), dẫn đến độ âm điện của điện thế màng tăng khoảng 10 mV. Protein vận chuyển thực hiện hoạt động này ở tốc độ cao: từ 150 đến 600 ion natri mỗi giây. Trình tự axit amin của protein vận chuyển đã được biết nhưng cơ chế vận chuyển trao đổi chất phức tạp này vẫn chưa rõ ràng. Quá trình này được mô tả bằng cách sử dụng các biểu đồ năng lượng của quá trình chuyển ion natri hoặc kali bằng protein (Hình 1.5,5). Theo bản chất của những thay đổi trong các cấu hình này, liên quan đến những thay đổi liên tục về cấu hình của protein vận chuyển (một quá trình cần năng lượng), người ta có thể đánh giá phép cân bằng hóa học của quá trình trao đổi: hai ion kali được trao đổi cho ba ion natri.
Bơm Na/K, giống như bơm cách ly Na+ /K + - ATPase màng phụ thuộc, bị ức chế đặc biệt bởi ouabain glycoside tim (strophanthin). Vì hoạt động của bơm Na/K là một phản ứng hóa học nhiều giai đoạn nên nó cũng giống như tất cả các phản ứng hóa học khác, phần lớn phụ thuộc vào nhiệt độ.

Cơm. 1.6.Sơ đồ bơm Na/K-ATPase (ngâm trong lớp lipid kép của màng plasma), trong một chu kỳ sẽ loại bỏ ba ion Na + khỏi tế bào theo chiều chênh lệch điện thế và nồng độ và đưa hai ion K vào tế bào + . Trong quá trình này, một phân tử ATP được phân hủy thành ADP và photphat. Sơ đồ cho thấy ATPase dưới dạng dimer bao gồm các tiểu đơn vị lớn (chức năng) và nhỏ; trong màng nó tồn tại dưới dạng tetramer được hình thành bởi hai tiểu đơn vị lớn và hai tiểu đơn vị nhỏ
thể hiện trong hình. 1.7. Ở đây, dòng ion natri từ tế bào cơ được thể hiện tương đối theo thời gian; điều này thực tế tương đương với dòng ion natri qua trung gian hoạt động của bơm Na/K, bởi vì dòng thụ động của ion natri chống lại nồng độ và gradient điện thế là cực kỳ nhỏ. Nếu thuốc được làm lạnh xuống khoảng 18°C, dòng ion natri từ tế bào sẽ nhanh chóng giảm đi 15 lần và ngay sau khi đun nóng sẽ được phục hồi về mức ban đầu. Sự giảm dòng ion natri từ tế bào này lớn hơn nhiều lần so với mức giảm tương ứng với sự phụ thuộc vào nhiệt độ của quá trình khuếch tán hoặc một phản ứng hóa học đơn giản. Một hiệu ứng tương tự cũng được quan sát thấy khi năng lượng trao đổi chất bị cạn kiệt do ngộ độc dinitrophenol (DNP) (Hình 1.7,5). Do đó, dòng ion natri từ tế bào được đảm bảo bằng phản ứng phụ thuộc vào năng lượng - một máy bơm hoạt động. Một đặc điểm khác của máy bơm, cùng với sự phụ thuộc đáng kể vào nhiệt độ và năng lượng, là sự hiện diện của mức bão hòa (giống như tất cả các phản ứng hóa học khác); điều này có nghĩa là tốc độ của máy bơm không thể tăng vô thời hạn khi nồng độ các ion vận chuyển tăng lên (Hình 1.8). Ngược lại, dòng chất khuếch tán thụ động tăng tỷ lệ thuận với chênh lệch nồng độ theo định luật khuếch tán (phương trình 1 và 2).

Cơm. 1.7. A, B. Vận chuyển Na tích cực + . Trục Y: dòng phóng xạ 24 Na + từ tế bào (imp./min). Trục hoành: thời gian kể từ khi bắt đầu thí nghiệm. MỘT. Tế bào được làm lạnh từ 18,3 °C xuống 0,5 °C; chảy Na+ ra khỏi tế bào trong thời gian này bị ức chế. B.Ức chế dòng Na + từ tế bào bằng dinitrophenol (DNP) ở nồng độ 0,2 mmol/l (đã biến đổi)
Ngoài bơm Na/K, màng plasma còn chứa ít nhất một bơm nữa - canxi; máy bơm này bơm các ion canxi (Ca 2+) ra khỏi tế bào và tham gia vào việc duy trì nồng độ nội bào của chúng ở mức cực thấp (Bảng 1.1). Bơm canxi hiện diện với mật độ rất cao trong mạng lưới cơ tương của tế bào cơ, nơi tích tụ các ion canxi do sự phân hủy các phân tử ATP (xem Chương 4).
Ảnh hưởng của bơm Na/K lên điện thế màng và thể tích tế bào . Trong bộ lễ phục. Hình 1.9 cho thấy các thành phần khác nhau của dòng điện qua màng và cho thấy nồng độ nội bào của các ion

Cơm. 1.8.Mối quan hệ giữa tốc độ vận chuyển của các phân tử và nồng độ của chúng (tại lối vào kênh hoặc tại vị trí liên kết của bơm) trong quá trình khuếch tán qua kênh hoặc trong quá trình vận chuyển bơm. Sau này trở nên bão hòa ở nồng độ cao (tốc độ tối đa, Vmax ) trên trục x tương ứng với một nửa tốc độ bơm tối đa ( Vmax /2), là nồng độ cân bằng ĐẾN tôi

Cơm. 1.9.Sơ đồ thể hiện nồng độ Na+ , K+ và Cl – bên trong và bên ngoài tế bào và con đường xâm nhập của các ion này qua màng tế bào (thông qua các kênh ion cụ thể hoặc sử dụng bơm Na/K. Tại gradient nồng độ nhất định, điện thế cân bằng E Na, E K và E С l – bằng với giá trị được chỉ định, điện thế màng Em = – 90 mV
đảm bảo sự tồn tại của chúng. Dòng ion kali đi ra ngoài được quan sát qua các kênh kali, vì điện thế màng có độ dương điện cao hơn một chút so với điện thế cân bằng của ion kali. Độ dẫn tổng thể của kênh natri thấp hơn nhiều so với kênh kali, tức là kênh natri mở ít thường xuyên hơn so với kênh kali ở điện thế nghỉ; tuy nhiên, số lượng ion natri đi vào tế bào cũng gần bằng số lượng ion kali thoát ra khỏi tế bào, vì nồng độ và gradient điện thế cần lớn để ion natri khuếch tán vào tế bào. Bơm Na/K cung cấp khả năng bù lý tưởng cho dòng khuếch tán thụ động, vì nó vận chuyển các ion natri ra khỏi tế bào và các ion kali vào trong tế bào. Do đó, máy bơm có khả năng sinh điện do sự khác biệt về số lượng điện tích truyền vào và ra khỏi tế bào, ở tốc độ hoạt động bình thường của nó sẽ tạo ra điện thế màng xấp xỉ 10 mV có độ âm điện lớn hơn nếu nó được tạo ra chỉ bởi dòng ion thụ động (xem Công thức 7). Kết quả là điện thế màng tiến tới điện thế cân bằng kali, làm giảm sự rò rỉ ion kali. hoạt động Na Bơm /K được điều chỉnh bởi nồng độ ion natri nội bào. Tốc độ của bơm chậm lại khi nồng độ ion natri cần loại bỏ khỏi tế bào giảm (Hình 1.8), do đó hoạt động của bơm và dòng ion natri vào tế bào cân bằng lẫn nhau, duy trì nồng độ natri nội bào ion ở mức xấp xỉ 10 mmol/L.
Để duy trì trạng thái cân bằng giữa dòng bơm và dòng màng thụ động, cần nhiều phân tử bơm Na/K hơn các protein kênh cho ion kali và natri. Khi kênh mở, hàng chục nghìn ion đi qua nó trong vài mili giây (xem ở trên) và vì kênh thường mở vài lần trong một giây nên tổng cộng có hơn 10 ion 5 đi qua kênh trong thời gian này. Một protein bơm duy nhất di chuyển hàng trăm ion natri mỗi giây, do đó màng plasma phải chứa số lượng phân tử bơm nhiều hơn khoảng 1000 lần so với các phân tử kênh. Các phép đo dòng điện trong kênh ở trạng thái nghỉ cho thấy sự hiện diện trung bình của một kênh mở kali và một natri trên mỗi màng 1 μm; Từ đó suy ra rằng khoảng 1000 phân tử của bơm Na/K sẽ có mặt trong cùng một không gian, tức là. khoảng cách giữa chúng trung bình là 34 nm; Đường kính của protein bơm, với tư cách là protein kênh, là 8–10 nm. Do đó, màng được bão hòa khá dày đặc với các phân tử bơm
Thực tế là dòng ion natri vào tế bào và dòng ion kali ra khỏi tế bào được bù đắp bằng hoạt động của máy bơm, còn có một hệ quả khác, bao gồm việc duy trì áp suất thẩm thấu ổn định và khối lượng không đổi. Bên trong tế bào có nồng độ cao các anion lớn, chủ yếu là protein (A - trong Bảng 1.1), không có khả năng xuyên qua màng (hoặc xuyên qua màng rất chậm) và do đó là thành phần cố định bên trong tế bào. Để cân bằng điện tích của các anion này, cần có số lượng cation bằng nhau. Do hoạt động của bơm Na/K, các cation này chủ yếu là ion kali. Sự gia tăng đáng kể nồng độ ion nội bào chỉ có thể xảy ra khi tăng nồng độ anion do dòng C1 - dọc theo gradient nồng độ vào trong tế bào (Bảng 1.1), nhưng điện thế màng lại chống lại điều này. Dòng điện đến Cl – chỉ quan sát cho đến khi đạt đến thế cân bằng của các ion clo; điều này được quan sát thấy khi gradient ion clo gần như đối lập với gradient ion kali, vì ion clo tích điện âm (Công thức 4). Do đó, nồng độ ion clo nội bào thấp được thiết lập, tương ứng với nồng độ ion kali ngoại bào thấp. Kết quả là tổng số ion trong tế bào bị giới hạn. Nếu điện thế màng giảm khi bơm Na/K bị chặn, ví dụ như trong quá trình thiếu oxy, thì điện thế cân bằng của các ion clo sẽ giảm và nồng độ ion clorua nội bào sẽ tăng theo. Khôi phục lại sự cân bằng điện tích, ion kali cũng đi vào tế bào; tổng nồng độ ion trong tế bào tăng lên, làm tăng áp suất thẩm thấu; điều này buộc nước vào tế bào. Tế bào sưng lên. Sự sưng tấy này được quan sát thấy trong cơ thể sống trong điều kiện thiếu năng lượng.
Độ dốc nồng độ Na + Là động lực cho sự vận chuyển qua màng . Tầm quan trọng của bơm Na/K đối với tế bào không chỉ giới hạn ở việc ổn định gradient K+ và Na+ bình thường trên màng. Năng lượng dự trữ trong gradient màng Na+ , thường được sử dụng để tạo điều kiện cho màng vận chuyển các chất khác. Ví dụ, trong hình. 1.10 hiển thị “simport” Na+ và các phân tử đường vào trong tế bào. Protein vận chuyển màng vận chuyển phân tử đường vào trong tế bào ngay cả khi đi ngược lại gradient nồng độ, đồng thời Na + di chuyển dọc theo gradient nồng độ và gradient thế năng, cung cấp năng lượng cho vận chuyển đường. Việc vận chuyển đường này hoàn toàn phụ thuộc vào sự tồn tại của gradient cao Na+ ; nếu nồng độ nội bào Na+ tăng lên đáng kể, quá trình vận chuyển đường dừng lại. Cho nhiều c akharov, có nhiều hệ thống simport khác nhau. Vận chuyển axit amin vào một tế bào tương tự như vận chuyển c akharov được thể hiện trong hình. 1,10; nó cũng được cung cấp bởi gradient Na+ , Có ít nhất năm hệ thống vận chuyển khác nhau, mỗi hệ thống chuyên biệt cho một nhóm axit amin liên quan.
Ngoài hệ thống simport, còn có "chống người khuân vác". Ví dụ, một trong số chúng chuyển một ion canxi ra khỏi tế bào trong một chu kỳ để đổi lấy ba ion natri đến (Hình 1.10). Năng lượng cho Vận chuyển Ca 2+được hình thành do sự xâm nhập của ba ion natri dọc theo nồng độ và gradient thế năng. Năng lượng này đủ (ở điện thế nghỉ) để duy trì gradient ion canxi cao (từ dưới 10–7 mol/L bên trong tế bào đến khoảng 2 mmol/L bên ngoài tế bào).
Endo- và exocytosis . Đối với một số chất đi vào tế bào hoặc phải thải ra ngoài

Cơm. 1.10.Các protein gắn trong lớp lipid kép của màng làm trung gian cho sự vận chuyển glucose và Na+ vào trong tế bào, cũng như Ca2+/Na+ –phản cổng, trong đó động lực là gradient Na+ trên màng tế bào
từ đó không còn kênh vận chuyển; Những chất này bao gồm, ví dụ, protein và cholesterol. Chúng có thể đi qua màng tế bào vào mụn nước, hoặc bong bóng, thông qua nội bào và ngoại bào. Trong bộ lễ phục. Hình 1.11 cho thấy cơ chế chính của các quá trình này. Trong quá trình xuất bào, một số bào quan nhất định (xem bên dưới) hình thành các túi chứa đầy chất cần được loại bỏ khỏi tế bào, chẳng hạn như hormone hoặc enzyme ngoại bào. Khi các túi này đến màng sinh chất, màng lipid của chúng sẽ kết hợp với nó, do đó cho phép các chất bên trong thoát ra môi trường bên ngoài. Trong quá trình ngược lại, nhập bào, màng sinh chất xâm lấn, tạo thành một hố, sau đó sâu hơn và đóng lại, tạo thành một túi nội bào chứa đầy dịch ngoại bào và một số đại phân tử. Để đảm bảo sự hợp nhất màng này và đóng túi, các yếu tố co bóp của khung tế bào hoạt động kết hợp với chính màng (xem bên dưới). Endocytosis không phải lúc nào cũng chỉ đơn giản liên quan đến việc thu giữ môi trường ngoại bào vào tế bào. Màng tế bào thường chứa các thụ thể đặc hiệu cho các đại phân tử, chẳng hạn như insulin hoặc kháng nguyên, thường được tổ chức thành các nhóm chuyên biệt. Sau khi các đại phân tử này liên kết với các thụ thể của chúng, quá trình nội bào xảy ra ở vùng màng xung quanh thụ thể và đại phân tử được vận chuyển có chọn lọc vào tế bào (Hình 1.12, B).
Endo- và exocytosis xảy ra liên tục trong tế bào. Lượng luân chuyển vật liệu màng là đáng kể; trong vòng 1 giờ, đại thực bào hấp thụ gấp đôi diện tích bề mặt màng tế bào chất của nó dưới dạng túi. Ở hầu hết các tế bào, sự luân chuyển vật liệu màng không diễn ra mạnh mẽ nhưng vẫn đáng kể.

Cơm. 1.11.Xuất bào và nhập bào. Hướng lên: túi nội bào kết hợp với lớp lipid kép của màng sinh chất và mở ra khoảng ngoại bào. Quá trình này được gọi là exocytosis. Ở phía dưới: màng sinh chất xâm lấn vào một khu vực nhỏ và giải phóng một túi chứa đầy chất ngoại bào. Quá trình này được gọi là nhập bào
1.3. Vận chuyển các chất trong tế bào
Endo- và exocytosis không chỉ là quá trình vận chuyển các chất qua màng tế bào mà còn là quá trình trao đổi màng - thành phần cấu trúc của chính tế bào. Chủ đề được xem xét trong phần này là các quá trình vận chuyển tương tự khác trong tế bào và các bào quan của nó.

Cơm. 1.12. A–B. Sơ đồ các quá trình bao gồm ngoại bào và nội bào. MỘT. Protein được tổng hợp trong mạng lưới nội chất dạng hạt được vận chuyển qua bộ máy Golgi đến màng sinh chất, nơi nó được tiết ra bằng quá trình ngoại bào. B. Cholesterol liên kết với các hạt LDL (lipoprotein mật độ thấp) gắn vào màng huyết tương, tạo ra sự hình thành túi nội tiết ở vùng màng này và được vận chuyển đến lysosome, nơi nó được giải phóng. TRONG. Vật liệu ngoại bào được thu giữ trong quá trình nhập bào (trong hình ở bên phải),được vận chuyển qua tế bào dưới dạng túi hoặc túi và được giải phóng thông qua quá trình xuất bào (trong hình bên trái)
Khuếch tán . Đương nhiên, trong bào tương, sự khác biệt về nồng độ bị loại bỏ do sự khuếch tán; điều tương tự cũng đúng với chất lỏng có trong các bào quan. Do nồng độ protein hòa tan cao nên quá trình khuếch tán ở đây chậm hơn nhiều so với trong nước. Màng lipid—xung quanh tế bào và bên trong các bào quan—là chất lỏng hai chiều trong đó xảy ra sự khuếch tán. Lipid trong lớp màng kép khuếch tán trong lớp riêng của chúng, hiếm khi di chuyển từ lớp này sang lớp khác. Các protein được nhúng trong chúng cũng khá cơ động; chúng quay quanh một trục vuông góc với màng hoặc khuếch tán sang một bên với hằng số khuếch tán rất khác nhau, chậm hơn 2–10.000 lần so với phospholipid. Vì vậy, nếu một số protein di chuyển tự do trong lớp lipid và có cùng tốc độ với chính các phân tử lipid, thì những protein khác sẽ được neo giữ, tức là. được kết nối khá chắc chắn với khung tế bào. Có những tập hợp “vĩnh viễn” của các protein cụ thể trong màng, chẳng hạn như cấu trúc trước và sau khớp thần kinh của các tế bào thần kinh. Các protein di chuyển tự do có thể được chứng minh bằng cách liên kết chúng với thuốc nhuộm huỳnh quang, sự phát quang của chúng được tạo ra bằng cách chiếu sáng nhanh một vùng nhỏ của màng. Những thí nghiệm như vậy cho thấy trong vòng chưa đầy 1 phút, các protein liên kết với thuốc nhuộm được phân bố đồng đều trên màng trong khoảng cách lên tới 10 μm.
Vận chuyển tích cực ở màng tế bào .
Các quá trình vận chuyển tích cực, đóng vai trò quan trọng trong hoạt động của màng sinh chất, cũng xảy ra bên trong tế bào, trong màng của các bào quan. Nội dung cụ thể của các bào quan khác nhau được tạo ra một phần do quá trình tổng hợp bên trong và một phần do sự vận chuyển tích cực từ tế bào chất. Một ví dụ sau đó là bơm Ca 2+ nêu trên trong mạng lưới cơ tương của tế bào cơ. Điều đặc biệt thú vị là trong trường hợp tổng hợp ATP ở ty thể, nguyên tắc ngược lại áp dụng cho những gì xảy ra trong bơm ATPase của màng tế bào (Hình 1.6). Trong quá trình tổng hợp ATP, quá trình trao đổi chất oxy hóa dẫn đến sự hình thành gradient dốc H+ trên màng bên trong. Độ dốc này là động lực cho quá trình ngược lại của chu trình bơm vận chuyển tích cực các phân tử: các ion H + di chuyển qua màng dọc theo độ dốc và năng lượng được giải phóng do đó cung cấp sự tổng hợp ATP từ ADP và photphat. Ngược lại, ATP thu được sẽ cung cấp năng lượng cho tế bào, bao gồm cả năng lượng cho quá trình vận chuyển tích cực.
Vận chuyển trong túi nước . Tế bào có một số lượng lớn các bào quan và các túi liên quan (Hình 1.1). Các bào quan này, và đặc biệt là các túi, chuyển động liên tục, vận chuyển nội dung của chúng đến các bào quan khác hoặc tới màng sinh chất. Các mụn nước cũng có thể di chuyển từ màng tế bào đến các bào quan, như trong quá trình nhập bào.
Quá trình bài tiết protein thể hiện trong hình. 1.12, MỘT. Protein được tổng hợp gần nhân tế bào trên các ribosome liên kết với mạng lưới nội chất (còn gọi là mạng lưới nội chất dạng hạt hoặc thô); Khi ở trong mạng lưới nội chất, protein được đóng gói thành các túi vận chuyển, tách ra khỏi bào quan và di chuyển đến bộ máy Golgi. Ở đây, chúng hợp nhất với các bể chứa của bộ máy Golgi, nơi protein được biến đổi (tức là chuyển đổi thành glycoprotein). Ở cuối các bể chứa nước, các túi lại tách ra. Các túi tiết mang protein biến đổi di chuyển đến màng sinh chất và giải phóng nội dung của chúng bằng cách xuất bào.
Một ví dụ khác về con đường vận chuyển trong tế bào được thể hiện trong hình. 1.12, B; Đây là sự hấp thụ cholesterol của tế bào. Cholesterol vận chuyển trong máu chủ yếu liên kết với protein, chẳng hạn như các hạt "mật độ lipoprotein thấp"(LDL). Các hạt như vậy gắn vào các vùng màng cụ thể có chứa thụ thể LDL, nơi xảy ra hiện tượng nhập bào và LDL được vận chuyển vào tế bào trong các túi “có viền”. Những mụn nước này hợp nhất để tạo thành nội nhũ và mất đi “viền” trong quá trình này. Ngược lại, các nội thể hợp nhất với các lysosome sơ cấp, chứa chủ yếu các enzyme thủy phân và tạo thành các lysosome thứ cấp, lớn hơn. Ở đó, cholesterol được giải phóng khỏi các hạt LDL và khuếch tán vào bào tương, nơi nó có sẵn, chẳng hạn như để tổng hợp màng lipid. Các túi không chứa LDL cũng được tách ra khỏi nội bào, chúng di chuyển theo một cách đặc biệt đến màng sinh chất và hợp nhất với nó, trả lại vật liệu màng và có thể cả các thụ thể cho LDL. Từ thời điểm hạt LDL liên kết với màng cho đến khi cholesterol được giải phóng khỏi lysosome thứ cấp, mất 10-15 phút. Sự suy giảm khả năng liên kết và hấp thu LDL, tức là trong việc cung cấp cholesterol cho tế bào, đóng vai trò quyết định trong sự phát triển của một căn bệnh nghiêm trọng và lan rộng, chứng xơ vữa động mạch (“xơ cứng” động mạch).
Có nhiều tuyến đường vận chuyển khác tương tự như trong Hình 2. 1.11 và 1.12, A, với sự trợ giúp của các túi cụ thể di chuyển trong tế bào. Người ta không biết chính xác chúng di chuyển như thế nào, nhưng các yếu tố khung tế bào có thể tham gia vào quá trình này. Các túi có thể trượt dọc theo các vi ống, trong trường hợp đó, năng lượng cho sự di chuyển rõ ràng được cung cấp bởi protein liên kết với các túi, ATPase (xem bên dưới). Hiện vẫn chưa rõ có bao nhiêu túi khác nhau, lần lượt di chuyển theo mọi hướng, đến đích. Rõ ràng là chúng cần được “đánh dấu” theo cách được hệ thống giao thông công nhận và chuyển thành chuyển động có mục tiêu.
Vận chuyển bằng cách hình thành và phá hủy các bào quan . Cho đến nay, chúng ta đã coi quá trình nội bào và ngoại bào là các quá trình vận chuyển nội dung của túi. Có một khía cạnh khác của các quá trình này, đó là việc loại bỏ mục tiêu màng plasma trên một phần của bề mặt tế bào bằng quá trình nhập bào và ngược lại, việc bổ sung nó vào một phần khác bằng quá trình xuất bào sẽ di chuyển các vùng rộng lớn của màng (Hình .12.E), tạo cơ hội cho tế bào, ví dụ, hình thành sự phát triển hoặc di chuyển.
Sự sắp xếp lại tương tự cũng điển hình cho bộ khung tế bào, đặc biệt là đối với các vi sợi và vi ống (Hình 1.1). Vi sợi bao gồm chủ yếu là protein F-actin, có khả năng tập hợp thành bó sợi do quá trình trùng hợp monome từ cytosol. Các bó có tính phân cực, tức là chúng thường chỉ phát triển ở một đầu, tích lũy các phân tử Actin mới, trong khi đầu kia trơ hoặc xảy ra hiện tượng phân tách ở đây. Do sự tăng trưởng phân cực này, các vi chất di chuyển hiệu quả và cấu trúc mạng lưới của chúng có thể thay đổi. Sự chuyển đổi Actin từ trạng thái khử polyme (sol) sang trạng thái có tổ chức (gel) có thể xảy ra rất nhanh dưới tác động của các protein khác hoặc sự thay đổi nồng độ ion (xem bên dưới). Ngoài ra còn có các protein khiến các sợi Actin bị phân hủy tạo thành các đoạn ngắn. Các phần nhô ra mỏng của nhiều tế bào - filmaia - chứa bó Actin trung tâm (Hình 1.1), và các chuyển động khác nhau của filmaia có thể là do sự chuyển tiếp Actin: trùng hợp - khử polyme.
Vi ốngcũng thường xuyên trải qua những chuyển động tương tự. Cơ chế của những chuyển động này tương tự nhau - sự trùng hợp của tubulin từ bào tương theo cách mà một trong hai đầu của vi ống phát triển, trong khi đầu kia không thay đổi hoặc xảy ra quá trình phân tách ở đó. Do đó, một vi ống, bằng cách bổ sung hoặc loại bỏ vật liệu thích hợp, có thể di chuyển khắp bào tương.
Chuyển động tích cực của tế bào . Những thay đổi trong cấu trúc khung tế bào có thể xảy ra do cả sự chuyển động tích cực và sự sắp xếp lại được mô tả ở trên. Trong nhiều trường hợp, sự chuyển động của vi ống và sợi Actin được điều khiển bởi các protein co bóp liên kết các sợi hoặc ống và có thể di chuyển chúng tương đối với nhau. Sóc myosin và dynein hiện diện trong bào tương của tất cả các tế bào với nồng độ tương đối cao; chúng là những yếu tố chuyển hóa năng lượng thành chuyển động trong các tế bào chuyên biệt (cơ) và bào quan (lông mao). Trong tế bào cơ, myosin tạo thành các sợi dày hướng song song với các sợi Actin. Phân tử myosin với “đầu” của nó gắn vào sợi Actin và sử dụng năng lượng của ATP, đẩy myosin dọc theo phân tử Actin. Myosin sau đó tách ra khỏi Actin. Sự kết hợp của nhiều chu kỳ kết nối-ngắt kết nối như vậy dẫn đến những ảnh hưởng vĩ mô sự co rút của các sợi cơ(chương 4). Dynein đóng vai trò tương tự trong sự chuyển động của các vi ống trong lông mao (Hình 1.1). Trong tế bào chất của các tế bào không chuyên biệt, myosin và dynein không tạo thành các sợi thông thường mà trong hầu hết các trường hợp là các nhóm phân tử nhỏ. Ngay cả trong những tập hợp nhỏ như vậy, chúng vẫn có khả năng di chuyển các sợi Actin hoặc vi ống. Cơm. Hình 1.13 minh họa quá trình này khi các phân tử myosin phân cực trái dấu cũng được gắn vào hai sợi Actin, phân cực theo các hướng khác nhau. Các nhóm đầu myosin uốn cong về phía đuôi của phân tử, tiêu thụ ATP trong quá trình này và hai sợi Actin di chuyển theo hướng ngược lại, sau đó myosin tách ra khỏi chúng. Các chuyển động thuộc loại này, trong đó năng lượng ATP được chuyển đổi thành công cơ học, có thể thay đổi hình dạng của khung tế bào và do đó là hình dạng tế bào, đồng thời cung cấp sự vận chuyển các bào quan liên quan đến khung tế bào.
Các quá trình vận chuyển nội bào có thể được thể hiện rõ ràng nhất trên sợi trục của tế bào thần kinh. Vận chuyển sợi trụcđược thảo luận chi tiết ở đây để minh họa các sự kiện có khả năng xảy ra theo những cách tương tự trong hầu hết các ô. Một sợi trục có đường kính chỉ vài micron có thể đạt chiều dài từ một mét trở lên và quá trình di chuyển của protein bằng cách khuếch tán từ nhân đến đầu xa của sợi trục sẽ mất nhiều năm. Từ lâu, người ta đã biết rằng khi bất kỳ phần nào của sợi trục bị co lại, phần sợi trục ở gần sẽ giãn ra nhiều hơn. Có vẻ như dòng ly tâm bị chặn trong sợi trục. Như là khả năng vận chuyển sợi trục nhanhđược chứng minh bằng sự chuyển động của các chất đánh dấu phóng xạ, như trong thí nghiệm ở Hình 2. 1.14. Leucine được dán nhãn phóng xạ được tiêm vào hạch rễ lưng, sau đó đo hoạt độ phóng xạ ở dây thần kinh tọa ở khoảng cách 166 mm tính từ thân tế bào thần kinh từ giờ thứ 2 đến giờ thứ 10. Sau hơn 10 giờ, đỉnh điểm phóng xạ tại vị trí tiêm có thay đổi nhẹ. Nhưng sóng phóng xạ lan truyền dọc theo sợi trục với tốc độ không đổi khoảng 34 mm trong 2 giờ, hay 410 mm/ngày. Người ta đã chứng minh rằng trong tất cả các tế bào thần kinh của động vật đẳng nhiệt, sự vận chuyển sợi trục nhanh xảy ra ở cùng một tốc độ và không có sự khác biệt đáng chú ý nào được quan sát thấy giữa các sợi mỏng, không có myelin và các sợi trục dày nhất, cũng như giữa sợi vận động và sợi cảm giác. Loại chất đánh dấu phóng xạ cũng không ảnh hưởng đến tốc độ vận chuyển nhanh của sợi trục; chất đánh dấu có thể phục vụ như một loạt các chất phóng xạ

Cơm. 1.13.Phức hợp myosin không cơ, với một hướng nhất định, có thể liên kết với các sợi Actin có cực tính khác nhau và sử dụng năng lượng của ATP, dịch chuyển chúng so với nhau.
các phân tử, chẳng hạn như các axit amin khác nhau, có trong protein của thân tế bào thần kinh. Nếu chúng ta phân tích phần ngoại vi của dây thần kinh để xác định bản chất của các chất mang chất phóng xạ được vận chuyển đến đây, thì những chất mang như vậy chủ yếu được tìm thấy trong phần protein, mà còn trong thành phần của các chất trung gian và axit amin tự do. Biết rằng tính chất của các chất này là khác nhau và kích thước phân tử của chúng đặc biệt khác nhau, chúng ta có thể giải thích tốc độ vận chuyển không đổi chỉ bằng một cơ chế vận chuyển chung cho tất cả chúng.
Miêu tả trên vận chuyển sợi trục nhanh là trước, tức là hướng ra khỏi cơ thể tế bào. Người ta đã chứng minh rằng một số chất di chuyển từ ngoại vi đến thân tế bào bằng cách sử dụng vận chuyển ngược dòng. Ví dụ, acetylcholinesterase được vận chuyển theo hướng này với tốc độ chậm hơn 2 lần so với tốc độ vận chuyển sợi trục nhanh. Một chất đánh dấu thường được sử dụng trong giải phẫu thần kinh, peroxidase cải ngựa, cũng di chuyển bằng cách vận chuyển ngược. Vận chuyển ngược có lẽ đóng một vai trò quan trọng trong việc điều hòa tổng hợp protein trong cơ thể tế bào. Một vài ngày sau khi sợi trục bị cắt, sự phân giải sắc tố được quan sát thấy trong cơ thể tế bào, điều này cho thấy sự gián đoạn trong quá trình tổng hợp protein. Thời gian cần thiết cho quá trình sắc ký tương quan với thời gian vận chuyển ngược từ vị trí chuyển tiếp sợi trục đến thân tế bào. Kết quả này cũng gợi ý lời giải thích cho tình trạng rối loạn này - việc truyền từ ngoại vi “chất tín hiệu” điều chỉnh quá trình tổng hợp protein bị gián đoạn. Rõ ràng là các "phương tiện" chính được sử dụng để truyền sợi trục nhanh

Cơm. 1.14.Thí nghiệm chứng minh sự vận chuyển sợi trục nhanh chóng trong các sợi cảm giác của dây thần kinh tọa ở mèo. Leucine gắn nhãn tritium được tiêm vào hạch rễ lưng và hoạt tính phóng xạ trong hạch và các sợi cảm giác được đo vào lúc 2, 4, 6, 8 và 10 giờ sau khi tiêm. (phía dưới ảnh). Qua trục x Khoảng cách từ hạch đến các vùng của dây thần kinh tọa nơi thực hiện phép đo được vẽ trên đồ thị. Trên trục tọa độ chỉ dành cho các đường cong trên và dưới, hoạt độ phóng xạ (xung/phút) được vẽ theo thang logarit. “Làn sóng” phóng xạ gia tăng (mũi tên) di chuyển với tốc độ 410 mm/ngày (dọc theo)
vận chuyển đang túi (túi) và bào quan, chẳng hạn như ty thể, nơi chứa các chất cần được vận chuyển. Sự chuyển động của các túi lớn nhất hoặc ty thể có thể được quan sát bằng kính hiển vi trong cơ thể sống . Các hạt như vậy thực hiện các chuyển động ngắn, nhanh theo một hướng, dừng lại, thường di chuyển nhẹ về phía sau hoặc sang một bên, dừng lại và sau đó giật theo hướng chính. 410 mm/ngày tương ứng với tốc độ di chuyển xuôi dòng trung bình khoảng 5 μm/s; Do đó, tốc độ của từng chuyển động riêng lẻ phải cao hơn nhiều và nếu chúng ta tính đến kích thước của các bào quan, sợi và vi ống thì những chuyển động này thực sự rất nhanh. Vận chuyển sợi trục nhanh đòi hỏi nồng độ ATP đáng kể. Các chất độc như colchicine, phá vỡ các vi ống, cũng ngăn chặn sự vận chuyển nhanh của sợi trục. Từ đó, trong quá trình vận chuyển mà chúng ta đang xem xét, các túi và bào quan di chuyển dọc theo các vi ống và sợi Actin; chuyển động này được cung cấp bởi các tập hợp nhỏ các phân tử dynein và myosin, hoạt động như trong Hình 2. 1.13, sử dụng năng lượng ATP.
Vận chuyển sợi trục nhanh cũng có thể liên quan đến các quá trình bệnh lý. Một số virus hướng thần kinh (ví dụ, virus herpes hoặc virus bại liệt) xâm nhập vào sợi trục ở ngoại vi và di chuyển theo phương thức vận chuyển ngược đến cơ thể tế bào thần kinh, nơi chúng nhân lên và phát huy tác dụng độc hại. Độc tố uốn ván, một loại protein do vi khuẩn sản xuất, xâm nhập vào cơ thể khi da bị tổn thương, bị các đầu dây thần kinh bắt giữ và vận chuyển đến thân tế bào thần kinh, nơi nó gây ra các cơn co thắt cơ đặc trưng. Có những trường hợp đã biết về tác dụng độc hại đối với chính quá trình vận chuyển sợi trục, ví dụ như tiếp xúc với dung môi công nghiệp acrylamide. Ngoài ra, người ta tin rằng cơ chế bệnh sinh của bệnh beriberi do thiếu vitamin và bệnh đa dây thần kinh do rượu liên quan đến sự vi phạm vận chuyển sợi trục nhanh.
Ngoài sự vận chuyển sợi trục nhanh chóng trong tế bào, còn có sự vận chuyển khá mạnh mẽ. vận chuyển sợi trục chậm. Tubulin di chuyển dọc theo sợi trục với tốc độ khoảng 1 mm/ngày và Actin di chuyển nhanh hơn—lên tới 5 mm/ngày. Các protein khác di chuyển cùng với các thành phần khung tế bào này; ví dụ, các enzyme dường như có liên quan đến Actin hoặc tubulin. Tốc độ di chuyển của tubulin và actin gần như phù hợp với tốc độ tăng trưởng được tìm thấy đối với cơ chế được mô tả trước đây, trong đó các phân tử được tích hợp vào đầu hoạt động của vi ống hoặc vi sợi. Do đó, cơ chế này có thể làm chậm quá trình vận chuyển sợi trục. Tốc độ vận chuyển sợi trục chậm cũng xấp xỉ tương ứng với tốc độ tăng trưởng sợi trục, điều này rõ ràng cho thấy những hạn chế do cấu trúc của khung tế bào áp đặt lên quá trình thứ hai.
Kết thúc phần này, cần nhấn mạnh rằng các tế bào hoàn toàn không phải là cấu trúc tĩnh, như chúng xuất hiện, ví dụ, trong các bức ảnh kính hiển vi điện tử. Màng huyết tương và đặc biệt các bào quan liên tục chuyển động nhanh và tái cơ cấu liên tục;đó là lý do tại sao họ có thể hoạt động. Hơn nữa, đây không phải là những căn phòng đơn giản trong đó diễn ra các phản ứng hóa học, mà là các tập đoàn có tổ chức cao của màng và sợi, trong đó các phản ứng xảy ra theo trình tự được tổ chức tối ưu.
1.4. Điều hòa chức năng tế bào
Việc duy trì từng tế bào như một đơn vị chức năng chủ yếu được điều hòa bởi nhân; việc nghiên cứu các cơ chế điều hòa như vậy là chủ đề của sinh học tế bào và hóa sinh. Đồng thời, các tế bào phải sửa đổi chức năng của chúng để phù hợp với điều kiện môi trường và nhu cầu của các tế bào khác trong cơ thể, tức là chúng đóng vai trò là đối tượng điều chỉnh chức năng. Dưới đây chúng ta sẽ xem xét ngắn gọn cách thức những ảnh hưởng điều tiết này tác động lên màng sinh chất và cách chúng tiếp cận các bào quan nội bào.
Tác dụng điều hòa trên màng tế bào
Điện thế màng . Trong nhiều trường hợp, việc điều chỉnh chức năng tế bào được thực hiện bằng cách thay đổi điện thế màng. Những thay đổi cục bộ về điện thế có thể xảy ra khi: 1) dòng điện từ khu vực lân cận của tế bào hoặc do một tế bào khác tạo ra chảy qua màng; 2) nồng độ ion ngoại bào thay đổi (thường là [K + ] ngoài ); 3) kênh ion màng mở. Những thay đổi về điện thế màng có thể ảnh hưởng đến cấu trúc của protein màng, đặc biệt là khiến các kênh mở hoặc đóng. Như đã mô tả ở trên, hoạt động của một số máy bơm màng phụ thuộc vào điện thế màng. Tế bào thần kinh có chức năng chuyên biệt để nhận biết những thay đổi về điện thế màng dưới dạng thông tin cần được xử lý và truyền đi (xem Chương 2).
Chất điều hòa ngoại bào . Cơ chế điều hòa quan trọng nhất liên quan đến các chất ngoại bào là sự tương tác của chúng với các thụ thể đặc hiệu trên màng sinh chất hoặc bên trong tế bào. Những chất này bao gồm các chất dẫn truyền qua khớp thần kinh, truyền thông tin giữa các tế bào thần kinh, tác nhân tại chỗ và các chất lưu thông trong máu và đến tất cả các tế bào của cơ thể, chẳng hạn như hormone và kháng nguyên. Bộ truyền dẫn synap là những phân tử nhỏ được giải phóng từ các đầu dây thần kinh ở khớp thần kinh;
khi chúng đến màng sinh chất của một tế bào sau khớp thần kinh lân cận, chúng sẽ kích hoạt các tín hiệu điện hoặc các cơ chế điều hòa khác. Vấn đề này được thảo luận chi tiết ở Chap. 3.
Tác nhân hóa học địa phương thường được tiết ra bởi các tế bào chuyên biệt. Chúng khuếch tán tự do vào không gian ngoại bào, nhưng hoạt động của chúng bị giới hạn ở một nhóm nhỏ tế bào do sự phá hủy nhanh chóng của các chất này, một cách tự nhiên hoặc dưới tác dụng của enzyme. Một ví dụ về việc giải phóng các tác nhân như vậy là việc giải phóng histamin tế bào mast do tổn thương hoặc phản ứng miễn dịch. Histamine gây giãn các tế bào cơ trơn mạch máu, làm tăng tính thấm của nội mô mạch máu và kích thích các đầu dây thần kinh cảm giác dẫn truyền cảm giác ngứa. Các tác nhân hóa học cục bộ khác được giải phóng bởi nhiều tế bào khác. Các đại lý địa phương điển hình là tuyến tiền liệt, tạo thành một nhóm gồm khoảng 20 dẫn xuất của axit béo. Chúng được giải phóng liên tục từ các tế bào phân bố rộng rãi, nhưng chỉ hoạt động cục bộ vì chúng nhanh chóng bị phá hủy bởi các phospholipase màng. Các loại prostaglandin khác nhau có phổ tác dụng rộng: chúng có thể kích hoạt sự co bóp của các tế bào cơ trơn, gây ra sự kết tụ các tiểu cầu trong máu (tiểu cầu) hoặc ngăn chặn sự phát triển của hoàng thể trong buồng trứng.
Các đại lý địa phương khác phục vụ yếu tố tăng trưởng.Được biết đến nhiều nhất là yếu tố tăng trưởng thần kinh (NGF) dành cho tế bào thần kinh giao cảm, cần thiết cho sự tăng trưởng và tồn tại của các tế bào thần kinh này trong quá trình phát triển. trong cơ thể sống hoặc trong nuôi cấy tế bào. Rõ ràng, các tế bào đích của lớp tế bào thần kinh này tiết ra NGF và do đó đảm bảo sự phân bố thích hợp. Khi hình thành cơ quan, tế bào thường phải “tìm đường” đến các tế bào đích, có thể định vị ở khoảng cách khá xa. Theo đó, phải có nhiều yếu tố tăng trưởng chuyên biệt tương tự như NGF.
Hormon và kháng nguyên được máu đưa tới mọi tế bào. Kháng nguyên kích hoạt phản ứng miễn dịch từ các tế bào mang kháng thể cụ thể. Tuy nhiên, kháng nguyên, theo nguyên tắc, là những chất lạ không được tạo ra trong cơ thể phản ứng (để biết thêm chi tiết, xem Chương 18). Một số hormone, chẳng hạn như insulin hoặc thyroxine, ảnh hưởng đến nhiều loại tế bào, trong khi những loại khác, chẳng hạn như hormone giới tính, chỉ ảnh hưởng đến một loại tế bào cụ thể. Hormon là các peptide, hoạt động của chúng được kích hoạt bằng cách liên kết với một thụ thể trên màng tế bào, hoặc steroid và thyroxine, khuếch tán qua màng lipid và liên kết với các thụ thể nội bào. Các hormone steroid liên kết với nhiễm sắc thể hạt nhân, dẫn đến sự phiên mã của một số gen nhất định. Các protein tạo thành gây ra những thay đổi trong chức năng tế bào, đó là tác dụng cụ thể của hormone. Các vấn đề liên quan đến việc giải phóng và hoạt động của hormone sẽ được thảo luận chi tiết trong Chương. 17.
Truyền thông nội bào liên quan đến chất truyền tin thứ hai
Các chức năng điều tiết được mô tả ở trên bao gồm các tác động lên màng tế bào. Thông tin mà màng tế bào nhận được thường phải khiến các bào quan phản ứng và được chuyển đến chúng bằng nhiều chất khác nhau được gọi là chất truyền tin thứ hai (trái ngược với chất truyền tin thứ nhất đến tế bào từ các nguồn bên ngoài). Nghiên cứu về chất hòa giải thứ hai đang phát triển nhanh chóng và không có gì đảm bảo rằng mức độ hiểu biết hiện tại về vấn đề sẽ đầy đủ. Ở đây chúng ta sẽ đề cập đến ba chất trung gian được nghiên cứu kỹ lưỡng: Ca 2+, cAMP và inositol triphosphate.
Canxi.Chất trung gian nội bào đơn giản nhất là ion Ca 2+. Nồng độ tự do của nó trong tế bào nghỉ rất thấp và lên tới 10_–8–10–7 mol/l. Nó có thể xâm nhập vào tế bào thông qua các kênh màng cụ thể khi chúng ở trạng thái mở, ví dụ như khi điện thế màng thay đổi (xem Chương 2). Kết quả là nồng độ Ca 2+ tăng lên sẽ kích hoạt các phản ứng quan trọng trong tế bào, chẳng hạn như sự co của các sợi cơ, là cơ sở của sự co cơ (xem Chương 4), hoặc giải phóng các túi chứa chất dẫn truyền thần kinh từ các đầu dây thần kinh (xem Chương 3) . Cả hai phản ứng đều yêu cầu nồng độ Ca 2+ khoảng 10 –5 mol/l. Ca 2+, có tác dụng điều hòa, cũng có thể được giải phóng từ các kho dự trữ nội bào, chẳng hạn như mạng lưới nội chất. Việc giải phóng Ca 2+ từ kho cần có sự tham gia của các trung gian khác (ví dụ, xem Hình 1.16).
Adenosine monophosphate tuần hoàn, cAMP. Gần đây, người ta đã chứng minh rằng adenosine monophosphate vòng (cAMP), một dẫn xuất của nguồn năng lượng chính của cơ thể, ATP, là chất truyền tin thứ hai quan trọng. Chuỗi phản ứng phức tạp được thể hiện trong hình. 1.15, bắt đầu bằng thụ thể R s trên bề mặt bên ngoài của màng sinh chất, có thể đóng vai trò là vị trí liên kết cụ thể cho các chất trung gian và hormone khác nhau. Sau khi liên kết với một phân tử “kích thích” cụ thể R s thay đổi hình dạng của nó; những thay đổi này ảnh hưởng đến protein Gs trên bề mặt bên trong của màng theo cách có thể kích hoạt chất sau bằng guanosine triphosphate (GTP) nội bào. Protein hoạt hóa Gs , lần lượt, kích thích enzyme trên bề mặt bên trong của màng, adenylate cyclase (AC), xúc tác cho sự hình thành cAMP từ ATP. cAMP tan trong nước là chất trung gian truyền tác dụng

Cơm. 1.15.Một chuỗi phản ứng liên quan đến chất truyền tin nội bào cAMP (cycladenosine monophosphate). Các tín hiệu bên ngoài kích thích hoặc ức chế kích hoạt thụ thể màng R s hoặc Ri . Những thụ thể này điều chỉnh quá trình liên kết G – protein có GTP nội bào (guanosine triphosphate), do đó kích thích hoặc ức chế adenylate cyclase nội bào (AC). Enzim khuếch đại AC chuyển đổi adenosine triphosphate (ATP) thành cAMP, sau đó được phân hủy thành AMP bởi phosphodiesterase. CAMP tự do khuếch tán vào tế bào và kích hoạt adenylate kinase (A-kinase), giải phóng tiểu đơn vị xúc tác C của nó, xúc tác quá trình phosphoryl hóa các protein nội bào, tức là. tạo thành tác dụng cuối cùng của một kích thích ngoại bào. Sơ đồ cũng cho thấy các loại thuốc dược lý và độc tố kích hoạt (+) hoặc ức chế (-) một số phản ứng nhất định (đã được sửa đổi)
kích thích thụ thể ngoại bào R s đến cấu trúc bên trong của tế bào.
Song song với chuỗi phản ứng kích thích liên quan đến R s Có thể liên kết các chất trung gian ức chế và hormone với thụ thể tương ứng tôi một lần nữa thông qua protein được kích hoạt GTP G , ức chế AC và do đó tạo ra cAMP. Khuếch tán vào tế bào, cAMP phản ứng với adenylate kinase (A-kinase); điều này giải phóng tiểu đơn vị C, xúc tác quá trình phosphoryl hóa protein P. Quá trình phosphoryl hóa này chuyển đổi protein thành dạng hoạt động và giờ đây chúng có thể phát huy tác dụng điều chỉnh cụ thể của mình (ví dụ: gây thoái hóa glycogen). Hệ thống điều hòa phức tạp này cực kỳ hiệu quả vì kết quả cuối cùng là sự phosphoryl hóa nhiều protein, tức là tín hiệu điều hòa đi qua mạch với mức tăng cao. Các chất trung gian bên ngoài liên kết với các thụ thể R s và R tôi dành riêng cho từng người trong số họ, vô cùng đa dạng. Adrenaline, kết nối với R s hoặc R i tham gia vào quá trình điều hòa chuyển hóa lipid và glycogen, cũng như tăng cường sự co bóp của cơ tim và các phản ứng khác (xem Chương 19). Hormon kích thích tuyến giáp, kích hoạt R s , kích thích tuyến giáp giải phóng hormone thyroxine và prostaglandin I ức chế sự kết tập tiểu cầu trong máu. Tác dụng ức chế, bao gồm adrenaline, qua trung gian tôi được thể hiện ở việc làm chậm quá trình phân giải lipid. Như vậy, Hệ thống cAMP là một hệ thống điều hòa nội bào đa chức năng, có thể được kiểm soát chính xác bởi các chất truyền tín hiệu kích thích và ức chế ngoại bào.
Inositol photphat "NẾU" h " Hệ thống nội bào của chất truyền tin thứ hai, inositol phosphate, chỉ được phát hiện gần đây (Hình 1.16). Trong trường hợp này, không có con đường ức chế, nhưng có điểm tương đồng với hệ thống cAMP, trong đó tác dụng kích thích thụ thể R được truyền đến protein G được kích hoạt GTP trên bề mặt bên trong của màng. Ở giai đoạn tiếp theo, phosphatidylinositol lipid màng thông thường (PI), trước đó đã nhận được hai nhóm photphat bổ sung, được chuyển thành PI-diphosphate (PIP 2), được phân cắt bởi phosphodiesterase hoạt hóa (PDE) thành inositol triphosphate(IFz) và lipid diacylglycerol(DAG). Inositol triphosphate là chất truyền tin thứ hai hòa tan trong nước khuếch tán vào bào tương. Nó hoạt động chủ yếu bằng cách giải phóng Ca 2+ từ mạng lưới nội chất. Ca 2+ lần lượt đóng vai trò là chất trung gian, như đã mô tả ở trên; ví dụ, nó kích hoạt phosphokinase phụ thuộc Ca 2+, giúp phosphoryl hóa các enzyme. Tiểu đơn vị lipid của DAG (Hình 1.16) cũng mang tín hiệu, khuếch tán trong pha lipid của màng sinh chất tới C-kinase nằm ở bề mặt bên trong của nó, được kích hoạt với sự tham gia của phosphatidylserine làm đồng yếu tố. Sau đó, C-kinase kích hoạt quá trình phosphoryl hóa protein, chuyển chúng thành dạng hoạt động.
Hệ thống truyền tin thứ hai nội bào IF 3 cũng có thể được kiểm soát bởi nhiều chất trung gian và hormone bên ngoài, bao gồm acetylcholine, serotonin, vasopressin và hormone kích thích tuyến giáp; giống như hệ thống cAMP, nó được đặc trưng bởi nhiều hiệu ứng nội bào. Có thể hệ thống này cũng được kích hoạt bởi ánh sáng trong cơ quan thụ cảm thị giác của mắt và đóng vai trò trung tâm trong quá trình dẫn truyền ánh sáng (xem Chương 11). Lần đầu tiên trong quá trình phát triển cá thể của sinh vật, thụ thể của hệ thống IG được kích hoạt bởi tinh trùng, do đó IG tham gia vào các phản ứng điều hòa đi kèm với quá trình thụ tinh của trứng.
Hệ thống cAMP và IFz-DAG có hiệu quả cao chất tăng cường sinh học. Họ

Cơm. 1.16.Một chuỗi phản ứng liên quan đến chất trung gian nội bào IPG (inositol triphosphate). Giống như trong hệ thống cAMP, tín hiệu ngoại bào được trung gian thông qua proteinG, trong trường hợp này kích hoạt phosphodiesterase (PDE). Enzym này phân hủy phosphatidylinosine diphosphate (PIP 2 ) trong màng tế bào trước IF h và diacylglycerol (DAG); NẾU NHƯ h khuếch tán vào tế bào chất. Tại đây nó gây ra sự giải phóng Ca 2+ từ mạng lưới nội chất; tăng nồng độ Ca 2+ trong tế bào chất ([Ca 2+] tôi ) kích hoạt protein kinase, chất này phosphoryl hóa và do đó kích hoạt các enzyme. Một sản phẩm khác, DAG, vẫn còn trong màng và kích hoạt protein kinase C (đồng yếu tố phosphatidylserine, PS). Protein kinase C cũng phosphoryl hóa các enzyme điều hòa các hoạt động cụ thể liên quan đến kích thích thụ thể bên ngoài R . Các nhánh của chuỗi phản ứng liên quan đến IF h và DAG có thể được kích hoạt độc lập bởi ionomycin và phorbol ester tương ứng (đã được sửa đổi)
chuyển đổi phản ứng giữa chất dẫn truyền và thụ thể màng ngoài thành quá trình phosphoryl hóa nhiều loại protein nội bào, sau đó có thể ảnh hưởng đến các chức năng khác nhau của tế bào. Một trong những khía cạnh quan trọng của vấn đề là, theo như những gì được biết ngày nay, chỉ có hai hệ thống điều hòa liên quan chặt chẽ thuộc loại này, được sử dụng bởi nhiều chất trung gian bên ngoài để điều chỉnh nhiều quá trình nội bào. Đồng thời, các hệ thống điều hòa này, bao gồm Ca 2+, tương tác chặt chẽ với nhau, cho phép chúng thực hiện tốt việc điều chỉnh các chức năng của tế bào.
Hướng dẫn và hướng dẫn
1. Alberts TRONG., Bray D., Lewis J., Raff M., Roberts ĐẾN., Watson J.D.Sinh học phân tử của tế bào, New York và London, Nhà xuất bản Garland, 1983.
2. Czihak G., Longer H., Ziegler H.(eds.). Sinh vật học. Berlin, Heidelberg, New York, Springer, 1983.
3. Hille TRONG. Các kênh ion của màng dễ bị kích thích. Sunderland, Mass., Sinauer PGS., 1984.
4. Hoppe W., Lohmann W.. Marki H., Ziegler H.(eds.). Sinh lý học. Berlin, Heidelberg, New York, Springer, 1984.
5. Jungermann ĐẾN., Mahler H.Hóa sinh. Berlin, Heidelberg, New York, Springer, 1980.
6. Kandel E. R., Schwartz– J. H.,(eds.). Nguyên tắc khoa học thần kinh, New York, Amsterdam, Oxford, Elsevier, 1985.
7. Schiebler T. H., Schmidt W.Giải phẫu des Menschen. Berlin, Heidelberg, New York, Tokyo, Springer, 1983.
Bài viết gốc và đánh giá
8. Berridge M.J. Cơ sở phân tử của giao tiếp trong tế bào, Sci. Amer, 253, 124-134 (1985).
9. Berridge M. J., Irvine R. F. Inositol triphosphate, chất truyền tin thứ hai mới trong truyền tín hiệu tế bào. Thiên nhiên, 312, 315 321 (1984).
10. Bretscher MS Các phân tử của màng tế bào, Sci. Amer., 253, 124–134 (1985).
11. Đậu J. Tế bào sống như một cỗ máy chuyển hóa năng lượng. Một mô hình tối thiểu về chuyển hóa cơ tim, Biochem. và Sinh lý học. Acta, 895, 41–62 (1987).
12. Hodgkin AL, Katz TRONG. Ảnh hưởng của ion natri đến hoạt động điện của sợi trục khổng lồ của mực. J. Physiol. (Lond.), 108, 37–77 (1949).
13. Hodgkin AL, Keynes R.D. Vận chuyển tích cực các cation trong sợi trục khổng lồ từ màu nâu đỏ Và Loligo, J. Physiol. (Lond.), 128, 28–42 (1955).
14. P dài hơn Các kênh ion với các trạng thái hình thành, Sinh lý học. J., 47, 581–590 (1985).
15. Ochs S., Worth P.M. Vận chuyển ngoại chất trong hệ thống bình thường và bệnh lý. Trong: Sinh lý học và bệnh lý của sợi trục, S.G. Waxmann, Ed. New York, Nhà xuất bản Raven, 1978.
Lý thuyết nhiệm vụ 5 trong kỳ thi Thống nhất môn sinh học
Cấu trúc tế bào. Mối quan hệ giữa cấu trúc và chức năng của các bộ phận, bào quan của tế bào là cơ sở cho tính toàn vẹn của nó
Cấu trúc tế bào
Cấu trúc của tế bào nhân sơ và nhân chuẩn
Các thành phần cấu trúc chính của tế bào là màng sinh chất, tế bào chất và bộ máy di truyền. Tùy thuộc vào đặc điểm của tổ chức, hai loại tế bào chính được phân biệt: tế bào nhân sơ và tế bào nhân chuẩn. Sự khác biệt chính giữa tế bào nhân sơ và tế bào nhân chuẩn là cách tổ chức bộ máy di truyền của chúng: ở sinh vật nhân sơ, nó nằm trực tiếp trong tế bào chất (khu vực này của tế bào chất được gọi là hạt nhân) và không bị phân tách khỏi nó bởi các cấu trúc màng, trong khi ở sinh vật nhân chuẩn, hầu hết DNA tập trung ở nhân, được bao quanh bởi màng kép. Ngoài ra, thông tin di truyền của tế bào nhân sơ, nằm trong nhân, được ghi trong phân tử DNA hình tròn, trong khi ở sinh vật nhân chuẩn, phân tử DNA mở.
Không giống như sinh vật nhân chuẩn, tế bào chất của tế bào nhân sơ cũng chứa một số lượng nhỏ các bào quan, trong khi tế bào nhân chuẩn được đặc trưng bởi sự đa dạng đáng kể của các cấu trúc này.
Cấu trúc và chức năng của màng sinh học
Cấu trúc của màng sinh học. Các màng bao quanh tế bào và các bào quan màng của tế bào nhân chuẩn có thành phần và cấu trúc hóa học chung. Chúng bao gồm lipid, protein và carbohydrate. Lipid màng chủ yếu được đại diện bởi phospholipid và cholesterol. Hầu hết các protein màng là các protein phức tạp, chẳng hạn như glycoprotein. Carbohydrate không xuất hiện độc lập trong màng; chúng liên kết với protein và lipid. Độ dày của màng là 7-10 nm.
Theo mô hình cấu trúc màng tế bào khảm chất lỏng được chấp nhận rộng rãi hiện nay, lipid tạo thành một lớp kép, hoặc lớp lipid kép, trong đó các “đầu” ưa nước của các phân tử lipid hướng ra ngoài và các “đuôi” kỵ nước được giấu bên trong màng. Những “đuôi” này, do tính kỵ nước của chúng, đảm bảo sự phân tách các pha nước của môi trường bên trong tế bào và môi trường của nó. Protein được liên kết với lipid thông qua nhiều loại tương tác khác nhau. Một số protein nằm trên bề mặt của màng. Những protein như vậy được gọi là ngoại vi, hoặc hời hợt. Các protein khác được ngâm một phần hoặc hoàn toàn trong màng - đây là tích phân, hoặc protein ngập nước. Protein màng thực hiện các chức năng cấu trúc, vận chuyển, xúc tác, thụ thể và các chức năng khác.
Màng không giống như tinh thể; các thành phần của chúng chuyển động liên tục, do đó xuất hiện các khoảng trống giữa các phân tử lipid - lỗ chân lông mà qua đó các chất khác nhau có thể đi vào hoặc rời khỏi tế bào.
Các màng sinh học khác nhau về vị trí của chúng trong tế bào, thành phần hóa học và chức năng. Các loại màng chính là huyết tương và nội bộ. Màng huyết tương chứa khoảng 45% lipid (bao gồm glycolipids), 50% protein và 5% carbohydrate. Các chuỗi carbohydrate, là một phần của protein-glycoprotein phức tạp và lipid-glycolipid phức tạp, nhô ra trên bề mặt màng. Các glycoprotein Plasmalemma cực kỳ đặc hiệu. Ví dụ, chúng được sử dụng để nhận dạng lẫn nhau các tế bào, bao gồm cả tinh trùng và trứng.
Trên bề mặt tế bào động vật, các chuỗi cacbohydrat tạo thành một lớp bề mặt mỏng - glycocalyx. Nó được phát hiện ở hầu hết các tế bào động vật, nhưng mức độ biểu hiện của nó khác nhau (10-50 µm). Glycocalyx cung cấp sự liên lạc trực tiếp giữa tế bào và môi trường bên ngoài, nơi xảy ra quá trình tiêu hóa ngoại bào; Các thụ thể nằm trong glycocalyx. Ngoài plasmalemma, các tế bào của vi khuẩn, thực vật và nấm cũng được bao quanh bởi màng tế bào.
Màng trong tế bào nhân chuẩn phân định các phần khác nhau của tế bào, tạo thành các “ngăn” đặc biệt - ngăn, thúc đẩy sự phân tách các quá trình trao đổi chất và năng lượng khác nhau. Chúng có thể khác nhau về thành phần hóa học và chức năng, nhưng sơ đồ cấu trúc chung của chúng vẫn giống nhau.
Chức năng màng:
- Hạn chế.Ý tưởng là chúng tách không gian bên trong tế bào khỏi môi trường bên ngoài. Màng có tính chất bán thấm, tức là chỉ những chất mà tế bào cần mới có thể tự do đi qua nó và có cơ chế vận chuyển các chất cần thiết.
- Tiếp nhận. Nó chủ yếu liên quan đến việc nhận thức các tín hiệu môi trường và truyền thông tin này vào tế bào. Các protein thụ thể đặc biệt chịu trách nhiệm cho chức năng này. Các protein màng cũng chịu trách nhiệm nhận dạng tế bào theo nguyên tắc “bạn hay thù”, cũng như hình thành các kết nối giữa các tế bào, trong đó được nghiên cứu nhiều nhất là các khớp thần kinh của tế bào thần kinh.
- Xúc tác. Nhiều phức hợp enzyme nằm trên màng, do đó các quá trình tổng hợp chuyên sâu xảy ra trên chúng.
- Chuyển hóa năng lượng. Liên quan đến sự hình thành năng lượng, lưu trữ năng lượng dưới dạng ATP và tiêu thụ.
- Sự phân chia. Màng còn phân định không gian bên trong tế bào, từ đó ngăn cách các nguyên liệu ban đầu của phản ứng và các enzyme có thể thực hiện các phản ứng tương ứng.
- Sự hình thành các liên hệ giữa các tế bào. Mặc dù độ dày của màng nhỏ đến mức không thể phân biệt được bằng mắt thường nhưng một mặt nó đóng vai trò là rào cản khá đáng tin cậy đối với các ion và phân tử, đặc biệt là các chất hòa tan trong nước, mặt khác. , đảm bảo sự vận chuyển chúng vào và ra khỏi tế bào.
- Chuyên chở.

Vận chuyển màng. Do tế bào, với tư cách là hệ thống sinh học cơ bản, là hệ thống mở, để đảm bảo quá trình trao đổi chất và năng lượng, duy trì cân bằng nội môi, tăng trưởng, kích thích và các quá trình khác, nên cần phải vận chuyển các chất qua màng - vận chuyển qua màng. Hiện nay, việc vận chuyển các chất qua màng tế bào được chia thành chủ động, thụ động, nội bào và ngoại bào.
Vận chuyển thụ động- Đây là loại hình vận chuyển diễn ra không tiêu hao năng lượng từ nơi có nồng độ cao đến nơi có nồng độ thấp. Các phân tử nhỏ không phân cực hòa tan trong lipid (O 2, CO 2) dễ dàng xâm nhập vào tế bào bằng cách khuếch tán đơn giản. Những chất không hòa tan trong lipid, kể cả các hạt nhỏ mang điện, được protein vận chuyển thu giữ hoặc đi qua các kênh đặc biệt (glucose, axit amin, K+, PO 4 3-). Kiểu vận chuyển thụ động này được gọi là khuếch tán thuận lợi. Nước xâm nhập vào tế bào qua các lỗ chân lông trong pha lipid, cũng như thông qua các kênh đặc biệt được lót bằng protein. Sự vận chuyển nước qua màng gọi là bằng thẩm thấu.

Sự thẩm thấu cực kỳ quan trọng đối với sự sống của tế bào, vì nếu nó được đặt trong dung dịch có nồng độ muối cao hơn trong dung dịch tế bào thì nước sẽ bắt đầu rời khỏi tế bào và thể tích của các chất sống sẽ bắt đầu giảm. Ở tế bào động vật, toàn bộ tế bào co lại và ở tế bào thực vật, tế bào chất nằm sau thành tế bào, được gọi là quá trình plasmolysis. Khi tế bào được đặt trong dung dịch ít đậm đặc hơn tế bào chất, quá trình vận chuyển nước diễn ra theo hướng ngược lại - vào trong tế bào. Tuy nhiên, khả năng mở rộng của màng tế bào chất có những giới hạn và tế bào động vật cuối cùng sẽ bị vỡ, trong khi tế bào thực vật không cho phép điều này xảy ra do thành tế bào chắc chắn của nó. Hiện tượng lấp đầy toàn bộ không gian bên trong của tế bào bằng nội dung tế bào được gọi là sự phân hủy chất plasmo. Cần tính đến nồng độ muối nội bào khi chuẩn bị thuốc, đặc biệt là tiêm tĩnh mạch, vì điều này có thể dẫn đến tổn thương tế bào máu (đối với điều này, người ta sử dụng dung dịch muối có nồng độ 0,9% natri clorua). Điều này không kém phần quan trọng khi nuôi cấy tế bào và mô cũng như các cơ quan động vật và thực vật.

Vận chuyển chủ động tiến hành tiêu hao năng lượng ATP từ nồng độ chất thấp hơn đến nồng độ cao hơn. Nó được thực hiện bằng cách sử dụng các protein bơm đặc biệt. Protein bơm K + , Na + , Ca 2+ và các ion khác qua màng, giúp thúc đẩy quá trình vận chuyển các chất hữu cơ thiết yếu, cũng như xuất hiện các xung thần kinh, v.v.
nhập bào- đây là một quá trình tích cực hấp thụ các chất của tế bào, trong đó màng hình thành các vết lõm và sau đó hình thành các túi màng - thể thực bào, chứa các vật thể bị hấp thụ. Sau đó lysosome chính kết hợp với phagosome và hình thành lysosome thứ cấp, hoặc phagolysosome, hoặc không bào tiêu hóa. Nội dung của túi được tiêu hóa bởi enzyme lysosome và các sản phẩm phân hủy được tế bào hấp thụ và đồng hóa. Dư lượng không tiêu hóa được loại bỏ khỏi tế bào bằng cách xuất bào. Có hai loại nhập bào chính: thực bào và pinocytosis.
thực bào là quá trình bề mặt tế bào bắt giữ và hấp thụ các hạt rắn của tế bào, và chứng pinocytosis- chất lỏng. Quá trình thực bào xảy ra chủ yếu ở tế bào động vật (động vật đơn bào, bạch cầu ở người), nó cung cấp dinh dưỡng cho chúng và thường bảo vệ cơ thể. Bằng cách pinocytosis, protein, phức hợp kháng nguyên-kháng thể được hấp thụ trong các phản ứng miễn dịch, v.v. Tuy nhiên, nhiều loại virus cũng xâm nhập vào tế bào bằng cách pinocytosis hoặc thực bào. Ở tế bào thực vật và nấm, thực tế là không thể thực hiện được vì chúng được bao quanh bởi màng tế bào bền.

Xuất bào- một quá trình đảo ngược với quá trình nhập bào. Bằng cách này, thức ăn chưa tiêu hóa còn sót lại sẽ được giải phóng khỏi không bào tiêu hóa và các chất cần thiết cho sự sống của tế bào và toàn bộ cơ thể sẽ bị loại bỏ. Ví dụ, việc truyền xung thần kinh xảy ra do tế bào thần kinh gửi xung thần kinh giải phóng các chất truyền tin hóa học - người trung gian, và trong tế bào thực vật, đây là cách các carbohydrate phụ của màng tế bào được tiết ra.
Thành tế bào của tế bào thực vật, nấm và vi khuẩn. Bên ngoài màng, tế bào có thể tiết ra một khung vững chắc - màng tế bào, hoặc vách tế bào.
Ở thực vật, cơ sở của thành tế bào là xenluloza, được đóng gói thành bó 50-100 phân tử. Khoảng trống giữa chúng chứa đầy nước và các carbohydrate khác. Màng tế bào thực vật được thấm qua các ống - plasmodesmata, qua đó các màng của lưới nội chất đi qua. Plasmodesmata vận chuyển các chất giữa các tế bào. Tuy nhiên, việc vận chuyển các chất, chẳng hạn như nước, cũng có thể xảy ra dọc theo thành tế bào. Theo thời gian, các chất khác nhau, bao gồm cả tannin hoặc các chất giống như chất béo, tích tụ trong thành tế bào của thực vật, dẫn đến sự hóa gỗ hoặc suberization của chính thành tế bào, sự dịch chuyển nước và làm chết các chất trong tế bào. Giữa các thành tế bào của các tế bào thực vật lân cận có các miếng đệm giống như thạch - các tấm ở giữa giữ chúng lại với nhau và gắn kết toàn bộ cơ thể thực vật. Chúng chỉ bị phá hủy trong quá trình quả chín và khi lá rụng.

Thành tế bào của tế bào nấm được hình thành chitin- một loại carbohydrate có chứa nitơ. Chúng khá chắc chắn và là bộ xương bên ngoài của tế bào, nhưng cũng giống như ở thực vật, chúng ngăn chặn quá trình thực bào.
Ở vi khuẩn, thành tế bào chứa cacbohydrat và các đoạn peptit - murein tuy nhiên, hàm lượng của nó thay đổi đáng kể giữa các nhóm vi khuẩn khác nhau. Các polysaccharide khác cũng có thể được tiết ra phía trên thành tế bào, tạo thành một lớp màng nhầy bảo vệ vi khuẩn khỏi những tác động từ bên ngoài.
Màng xác định hình dạng của tế bào, đóng vai trò hỗ trợ cơ học, thực hiện chức năng bảo vệ, cung cấp các đặc tính thẩm thấu cho tế bào, hạn chế sự kéo dài của các chất sống và ngăn ngừa sự vỡ của tế bào, tăng lên do sự xâm nhập của nước. . Ngoài ra, nước và các chất hòa tan trong nó sẽ vượt qua thành tế bào trước khi đi vào tế bào chất hoặc ngược lại, khi rời khỏi nó, trong khi nước được vận chuyển qua thành tế bào nhanh hơn qua tế bào chất.
Tế bào chất
Tế bào chất- Đây là nội dung bên trong của ô. Tất cả các bào quan của tế bào, nhân và các chất thải khác nhau đều được ngâm trong đó.
Tế bào chất kết nối tất cả các phần của tế bào với nhau và nhiều phản ứng trao đổi chất diễn ra trong đó. Tế bào chất được tách ra khỏi môi trường và được chia thành các ngăn bằng màng, nghĩa là tế bào có cấu trúc màng. Nó có thể ở hai trạng thái - sol và gel. Sol- đây là trạng thái bán lỏng, giống như thạch của tế bào chất, trong đó các quá trình quan trọng diễn ra mạnh mẽ nhất, và gel- trạng thái đặc hơn, sền sệt cản trở sự xuất hiện của các phản ứng hóa học và sự vận chuyển các chất.
Phần chất lỏng của tế bào chất không có bào quan được gọi là hyaloplasm. Hyaloplasm, hay cytosol, là một dung dịch keo trong đó có một loại huyền phù gồm các hạt khá lớn, ví dụ như protein, được bao quanh bởi các lưỡng cực của các phân tử nước. Sự kết tủa của huyền phù này không xảy ra do chúng có cùng điện tích và đẩy nhau.
Chất hữu cơ
Chất hữu cơ- Đây là những thành phần cố định của tế bào thực hiện các chức năng cụ thể.
Tùy thuộc vào đặc điểm cấu trúc của chúng, chúng được chia thành màng và không màng. Màng lần lượt, các bào quan được phân loại thành màng đơn (lưới nội chất, phức hợp Golgi và lysosome) hoặc màng kép (ty thể, lạp thể và nhân). Không màng Các bào quan là ribosome, vi ống, vi sợi và trung tâm tế bào. Trong số các bào quan được liệt kê, chỉ có ribosome là có ở sinh vật nhân sơ.
Cấu trúc và chức năng của hạt nhân. Cốt lõi- một cơ quan lớn có màng kép nằm ở trung tâm tế bào hoặc ở ngoại vi. Kích thước của hạt nhân có thể dao động từ 3-35 micron. Hình dạng của hạt nhân thường là hình cầu hoặc hình elip, nhưng cũng có những hạt nhân hình que, hình thoi, hình hạt đậu, thùy và thậm chí là phân đoạn. Một số nhà nghiên cứu tin rằng hình dạng của nhân tương ứng với hình dạng của tế bào.
Hầu hết các tế bào đều có một nhân, nhưng, ví dụ, trong tế bào gan và tim có thể có hai nhân, và ở một số tế bào thần kinh - lên tới 15. Các sợi cơ xương thường chứa nhiều nhân, nhưng chúng không phải là tế bào theo nghĩa đầy đủ của từ này, vì chúng được hình thành do sự hợp nhất của một số tế bào.
Cốt lõi được bao quanh Màng nhân, và không gian bên trong của nó được lấp đầy nước ép hạt nhân, hoặc nucleoplasm (karyoplasm), trong đó họ đắm chìm chất nhiễm sắc Và hạch nhỏ. Nhân thực hiện các chức năng quan trọng như lưu trữ và truyền thông tin di truyền, cũng như kiểm soát sự sống của tế bào.

Vai trò của nhân trong việc truyền thông tin di truyền đã được chứng minh một cách thuyết phục trong các thí nghiệm với tảo xanh Acetabularia. Trong một tế bào khổng lồ duy nhất, đạt chiều dài 5 cm, có mũ, thân và thân rễ. Hơn nữa, nó chỉ chứa một hạt nhân nằm trong rhizoid. Vào những năm 1930, I. Hemmerling đã cấy nhân của một loài acetabularia có màu xanh lục vào thân rễ của một loài khác, có màu nâu, từ đó nhân đã bị loại bỏ. Sau một thời gian, cây được cấy nhân đã mọc ra một mũ mới, giống như tảo cho nhân. Đồng thời, mũ hoặc cuống tách khỏi thân rễ và không chứa nhân sẽ chết sau một thời gian.

Màng nhânđược hình thành bởi hai màng - bên ngoài và bên trong, giữa đó có không gian. Khoảng gian màng giao tiếp với khoang của mạng lưới nội chất thô và màng ngoài của nhân có thể mang ribosome. Lớp vỏ hạt nhân được thấm vào nhiều lỗ chân lông được lót bằng các protein đặc biệt. Sự vận chuyển các chất xảy ra qua các lỗ chân lông: các protein cần thiết (bao gồm enzyme), ion, nucleotide và các chất khác đi vào nhân và các phân tử RNA, protein đã qua sử dụng và các tiểu đơn vị của ribosome rời khỏi nhân. Do đó, chức năng của vỏ hạt nhân là tách các chất trong nhân ra khỏi tế bào chất, cũng như điều hòa quá trình trao đổi chất giữa nhân và tế bào chất.
Nucleoplasm gọi là phần bên trong nhân, trong đó chất nhiễm sắc và nhân được nhúng vào. Nó là một dung dịch keo, về mặt hóa học gợi nhớ đến tế bào chất. Các enzyme của nucleoplasm xúc tác cho quá trình trao đổi axit amin, nucleotide, protein, v.v. Nhân tế bào được kết nối với hyaloplasm thông qua các lỗ nhân. Chức năng của nucleoplasm, giống như hyaloplasm, là đảm bảo sự liên kết của tất cả các thành phần cấu trúc của nhân và thực hiện một số phản ứng enzyme.
Chất nhiễm sắc gọi là tập hợp các sợi và hạt mỏng nằm trong nhân tế bào. Nó chỉ có thể được phát hiện bằng cách nhuộm màu, vì chỉ số khúc xạ của nhiễm sắc và nucleoplasm gần như nhau. Thành phần sợi của chất nhiễm sắc được gọi là chất nhiễm sắc trung tính và dạng hạt - dị nhiễm sắc. Euchromatin được cô đặc yếu vì thông tin di truyền được đọc từ nó, trong khi các chất dị nhiễm sắc xoắn ốc hơn thì không hoạt động về mặt di truyền.
Chromatin là sự biến đổi cấu trúc của nhiễm sắc thể trong nhân không phân chia. Do đó, nhiễm sắc thể liên tục hiện diện trong nhân; chỉ có trạng thái của chúng thay đổi tùy thuộc vào chức năng mà nhân thực hiện vào thời điểm đó.
Thành phần của chất nhiễm sắc chủ yếu bao gồm các protein nucleoprotein (deoxyribonucleoprotein và ribonucleoprotein), cũng như các enzyme, trong đó quan trọng nhất có liên quan đến quá trình tổng hợp axit nucleic và một số chất khác.
Các chức năng của chất nhiễm sắc bao gồm, trước hết là tổng hợp các axit nucleic đặc trưng cho một sinh vật nhất định, chỉ đạo quá trình tổng hợp các protein cụ thể, và thứ hai là chuyển các đặc tính di truyền từ tế bào mẹ sang tế bào con, nhằm mục đích đó các sợi nhiễm sắc được đóng gói thành nhiễm sắc thể trong quá trình phân chia.
nhân- thân hình cầu, nhìn rõ dưới kính hiển vi, có đường kính 1-3 micron. Nó được hình thành trên các phần của chất nhiễm sắc, trong đó thông tin về cấu trúc của rRNA và protein ribosome được mã hóa. Thường chỉ có một nhân trong nhân, nhưng trong những tế bào diễn ra các quá trình quan trọng chuyên sâu, có thể có hai hoặc nhiều nhân. Chức năng của nucleoli là tổng hợp rRNA và lắp ráp các tiểu đơn vị ribosome bằng cách kết hợp rRNA với các protein đến từ tế bào chất.
ty thể- các bào quan màng kép có dạng tròn, hình bầu dục hoặc hình que, mặc dù cũng có những bào quan có hình xoắn ốc (trong tinh trùng). Đường kính của ty thể lên tới 1 µm và chiều dài lên tới 7 µm. Không gian bên trong ty thể chứa đầy ma trận. Ma trận- Đây là chất chính của ty thể. Một phân tử DNA tròn và ribosome được nhúng vào đó. Màng ngoài của ty thể nhẵn và không thấm được nhiều chất. Màng bên trong có các hình chiếu - Crista, làm tăng diện tích bề mặt của màng để các phản ứng hóa học xảy ra. Trên bề mặt màng có rất nhiều phức hợp protein tạo nên cái gọi là chuỗi hô hấp, cũng như các enzyme ATP synthetase hình nấm. Giai đoạn hô hấp hiếu khí xảy ra ở ty thể, trong đó ATP được tổng hợp.

Plastid- Bào quan lớn có màng kép, đặc trưng chỉ có ở tế bào thực vật. Không gian bên trong của plastid được lấp đầy chất nền, hoặc ma trận. Chất nền chứa một hệ thống các túi màng ít nhiều phát triển - thylakoid, được thu thập thành từng đống - hạt, cũng như phân tử DNA vòng và ribosome của chính nó. Có bốn loại lạp thể chính: lục lạp, sắc lạp, bạch cầu và tiền lạp thể.
lục lạp- đây là những plastid màu xanh lá cây có đường kính 3-10 micron, có thể nhìn thấy rõ dưới kính hiển vi. Chúng chỉ được tìm thấy ở những phần còn xanh của cây - lá, thân non, hoa và quả. Lục lạp thường có hình bầu dục hoặc hình elip, nhưng cũng có thể có hình cốc, hình xoắn ốc hoặc thậm chí là thùy. Số lượng lục lạp trong một tế bào trung bình từ 10 đến 100 mảnh. Tuy nhiên, ví dụ, ở một số loài tảo, nó có thể là một, có kích thước đáng kể và hình dạng phức tạp - khi đó nó được gọi là tế bào sắc tố. Trong các trường hợp khác, số lượng lục lạp có thể lên tới vài trăm, trong khi kích thước của chúng nhỏ. Màu sắc của lục lạp là do sắc tố chính của quá trình quang hợp - chất diệp lục, mặc dù chúng cũng chứa các sắc tố bổ sung - carotenoit. Carotenoid chỉ trở nên đáng chú ý vào mùa thu, khi chất diệp lục trong lá già bị phá vỡ. Chức năng chính của lục lạp là quang hợp. Phản ứng sáng của quá trình quang hợp xảy ra trên màng thylakoid, trên đó các phân tử diệp lục được gắn vào, và phản ứng tối diễn ra trong chất nền, nơi chứa nhiều enzyme.

sắc lạp- Đây là các plastid màu vàng, cam và đỏ chứa sắc tố caroten. Hình dạng của sắc lạp cũng có thể thay đổi đáng kể: chúng có thể ở dạng hình ống, hình cầu, tinh thể, v.v. Sắc lạp tạo màu cho hoa và quả của cây, thu hút các loài thụ phấn và phân phối hạt và quả.
Bạch cầu- Đây là những plastid màu trắng hoặc không màu, chủ yếu có hình tròn hoặc hình bầu dục. Chúng phổ biến ở các bộ phận không quang hợp của thực vật, ví dụ như ở vỏ lá, củ khoai tây, v.v. Chúng dự trữ chất dinh dưỡng, thường là tinh bột, nhưng ở một số thực vật, nó có thể là protein hoặc dầu.
Plastid được hình thành trong tế bào thực vật từ proplastid, chúng đã có sẵn trong các tế bào của mô giáo dục và là những cơ thể màng đôi nhỏ. Ở giai đoạn đầu phát triển, các loại lạp thể khác nhau có khả năng biến đổi lẫn nhau: khi tiếp xúc với ánh sáng, bạch cầu của củ khoai tây và sắc lạp của rễ cà rốt chuyển sang màu xanh lục.
Plastid và ty thể được gọi là các bào quan bán tự trị của tế bào, vì chúng có các phân tử DNA và ribosome riêng, thực hiện quá trình tổng hợp protein và phân chia độc lập với sự phân chia tế bào. Những đặc điểm này được giải thích là do chúng có nguồn gốc từ các sinh vật nhân sơ đơn bào. Tuy nhiên, tính “độc lập” của ty thể và lục lạp bị hạn chế, vì DNA của chúng chứa quá ít gen để tồn tại tự do, trong khi phần thông tin còn lại được mã hóa trong nhiễm sắc thể của nhân, cho phép nó kiểm soát các bào quan này.
Mạng lưới nội chất (ER), hoặc lưới nội chất (ER), là một cơ quan màng đơn, là một mạng lưới các khoang màng và ống chiếm tới 30% nội dung của tế bào chất. Đường kính của ống EPS khoảng 25-30 nm. Có hai loại EPS - thô và mịn. XPS thô mang ribosome, nơi xảy ra quá trình tổng hợp protein. XPS mượt mà thiếu ribosome. Chức năng của nó là tổng hợp lipid và carbohydrate, cũng như vận chuyển, lưu trữ và trung hòa các chất độc hại. Nó đặc biệt phát triển ở những tế bào nơi diễn ra các quá trình trao đổi chất chuyên sâu, ví dụ như trong tế bào gan - tế bào gan - và các sợi cơ xương. Các chất được tổng hợp trong ER được vận chuyển đến bộ máy Golgi. Sự lắp ráp màng tế bào cũng xảy ra trong ER, nhưng sự hình thành của chúng được hoàn thành trong bộ máy Golgi.
Bộ máy Golgi, hoặc phức hợp Golgi, là một cơ quan đơn màng được hình thành bởi một hệ thống các bể chứa phẳng, các ống và túi tách ra khỏi chúng. Đơn vị cấu trúc của bộ máy Golgi là dictyosome- một chồng bể, ở một cực chứa các chất từ EPS đến, và từ cực đối diện, sau khi trải qua những biến đổi nhất định, chúng được đóng gói thành các túi và gửi đến các phần khác của tế bào. Đường kính của bể khoảng 2 micron, đường kính của bong bóng nhỏ khoảng 20-30 micron. Các chức năng chính của phức hợp Golgi là tổng hợp một số chất và biến đổi (thay đổi) protein, lipid và carbohydrate đến từ ER, sự hình thành cuối cùng của màng, cũng như vận chuyển các chất khắp tế bào, đổi mới cấu trúc của nó. và sự hình thành lysosome. Bộ máy Golgi được đặt tên để vinh danh nhà khoa học người Ý Camillo Golgi, người đầu tiên phát hiện ra cơ quan này (1898).

Lysosome- các bào quan đơn màng nhỏ có đường kính lên tới 1 μm, chứa các enzym thủy phân tham gia vào quá trình tiêu hóa nội bào. Màng lysosome có khả năng thấm kém đối với các enzyme này, vì vậy lysosome thực hiện chức năng của chúng rất chính xác và có mục tiêu. Do đó, chúng tham gia tích cực vào quá trình thực bào, hình thành không bào tiêu hóa và trong trường hợp một số bộ phận của tế bào bị đói hoặc bị tổn thương, chúng sẽ tiêu hóa chúng mà không ảnh hưởng đến những bộ phận khác. Vai trò của lysosome trong quá trình chết tế bào gần đây đã được phát hiện.
không bào là một khoang trong tế bào chất của tế bào thực vật và động vật, được bao bọc bởi một màng và chứa đầy chất lỏng. Không bào tiêu hóa và co bóp được tìm thấy trong các tế bào đơn bào. Loại thứ nhất tham gia vào quá trình thực bào, vì chúng phá vỡ các chất dinh dưỡng. Loại thứ hai đảm bảo duy trì cân bằng nước-muối do điều hòa thẩm thấu. Ở động vật đa bào, không bào tiêu hóa chủ yếu được tìm thấy.
Trong tế bào thực vật luôn có không bào; chúng được bao quanh bởi một lớp màng đặc biệt và chứa đầy nhựa tế bào. Màng bao quanh không bào có thành phần hóa học, cấu trúc và chức năng tương tự như màng sinh chất. Tế bào nhựa cây là dung dịch nước của nhiều chất vô cơ và hữu cơ khác nhau, bao gồm muối khoáng, axit hữu cơ, carbohydrate, protein, glycoside, alkaloid, v.v. Không bào có thể chiếm tới 90% thể tích tế bào và đẩy nhân ra ngoại vi. Phần này của tế bào thực hiện các chức năng lưu trữ, bài tiết, thẩm thấu, bảo vệ, lysosomal và các chức năng khác, vì nó tích lũy chất dinh dưỡng và chất thải, đảm bảo cung cấp nước và duy trì hình dạng và thể tích của tế bào, đồng thời cũng chứa các enzyme để phân hủy tế bào. nhiều thành phần tế bào. Ngoài ra, các hoạt chất sinh học của không bào có thể ngăn cản nhiều động vật ăn những thực vật này. Ở một số thực vật, do sự phồng lên của không bào, sự phát triển của tế bào xảy ra do sự kéo dài.
Không bào cũng có mặt trong tế bào của một số loại nấm và vi khuẩn, nhưng ở nấm, chúng chỉ thực hiện chức năng điều hòa thẩm thấu, trong khi ở vi khuẩn lam, chúng duy trì độ nổi và tham gia vào quá trình đồng hóa nitơ từ không khí.
Ribosome- các bào quan nhỏ không màng có đường kính 15-20 micron, bao gồm hai tiểu đơn vị - lớn và nhỏ. Các tiểu đơn vị ribosome của sinh vật nhân chuẩn được tập hợp trong nhân và sau đó được vận chuyển vào tế bào chất. Ribosome ở sinh vật nhân sơ, ty thể và lục lạp có kích thước nhỏ hơn ribosome ở sinh vật nhân chuẩn. Các tiểu đơn vị ribosome bao gồm rRNA và protein.

Số lượng ribosome trong tế bào có thể lên tới vài chục triệu: trong tế bào chất, ty thể và lục lạp, chúng ở trạng thái tự do và trên ER thô - ở trạng thái liên kết. Chúng tham gia vào quá trình tổng hợp protein, đặc biệt là thực hiện quá trình dịch mã - sinh tổng hợp chuỗi polypeptide trên phân tử mRNA. Ribosome tự do tổng hợp protein của hyaloplasm, ty thể, plastid và protein ribosome của chính chúng, trong khi ribosome gắn vào ER thô thực hiện quá trình dịch mã protein để loại bỏ khỏi tế bào, lắp ráp màng và hình thành lysosome và không bào.
Ribosome có thể được tìm thấy đơn lẻ trong hyaloplasm hoặc được tập hợp thành nhóm trong quá trình tổng hợp đồng thời một số chuỗi polypeptide trên một mRNA. Những nhóm ribosome như vậy được gọi là polyribosome, hoặc polysome.

Vi ống- Đây là những bào quan rỗng, không có màng, hình trụ, xâm nhập vào toàn bộ tế bào chất của tế bào. Đường kính của chúng khoảng 25 nm, độ dày thành 6-8 nm. Chúng được hình thành bởi nhiều phân tử protein ống,đầu tiên tạo thành 13 sợi giống như hạt và sau đó tập hợp lại thành một vi ống. Các vi ống tạo thành mạng lưới tế bào chất, tạo nên hình dạng và thể tích của tế bào, kết nối màng sinh chất với các phần khác của tế bào, đảm bảo vận chuyển các chất đi khắp tế bào, tham gia vào sự chuyển động của tế bào và các thành phần nội bào, cũng như trong sự phân chia vật chất di truyền. Chúng là một phần của trung tâm tế bào và các cơ quan vận động - roi và lông mao.
vi sợi, hoặc microthread, cũng là những bào quan không có màng, tuy nhiên, chúng có dạng sợi và được hình thành không phải bởi tubulin mà là Actin. Chúng tham gia vào các quá trình vận chuyển màng, nhận biết giữa các tế bào, phân chia tế bào chất và chuyển động của nó. Trong tế bào cơ, sự tương tác của các vi sợi Actin với các sợi myosin làm trung gian cho sự co bóp.
Các vi ống và vi sợi tạo thành bộ xương bên trong của tế bào - bộ xương tế bào. Đó là một mạng lưới các sợi phức tạp cung cấp hỗ trợ cơ học cho màng sinh chất, xác định hình dạng của tế bào, vị trí của các bào quan trong tế bào và sự chuyển động của chúng trong quá trình phân chia tế bào.

Trung tâm tế bào- một cơ quan không có màng nằm trong tế bào động vật gần nhân; nó không có trong tế bào thực vật. Chiều dài của nó khoảng 0,2-0,3 micron và đường kính của nó là 0,1-0,15 micron. Trung tâm tế bào được hình thành bởi hai trung thể, nằm trong các mặt phẳng vuông góc với nhau và quả cầu rạng rỡ từ các vi ống. Mỗi trung thể được hình thành bởi chín nhóm vi ống, được tập hợp thành nhóm ba, tức là bộ ba. Trung tâm tế bào tham gia vào quá trình lắp ráp vi ống, phân chia vật liệu di truyền của tế bào, cũng như hình thành tiên mao và lông mao.

Các bào quan vận động. Roi Và lông mi Chúng là sự phát triển của tế bào được bao phủ bởi plasmalemma. Cơ sở của các bào quan này được tạo thành từ chín cặp vi ống nằm dọc theo ngoại vi và hai vi ống tự do ở trung tâm. Các vi ống được liên kết với nhau bởi nhiều protein khác nhau, đảm bảo độ lệch phối hợp của chúng khỏi trục - dao động. Dao động phụ thuộc vào năng lượng, nghĩa là năng lượng của các liên kết ATP năng lượng cao được sử dụng cho quá trình này. Phục hồi các roi và lông mao bị mất là một chức năng cơ thể cơ bản, hoặc kinetosome nằm ở căn cứ của họ.
Chiều dài của lông mao khoảng 10-15 nm, và chiều dài của roi là 20-50 µm. Do sự di chuyển được chỉ đạo chặt chẽ của roi và lông mao, không chỉ xảy ra sự di chuyển của động vật đơn bào, tinh trùng, v.v. mà còn xảy ra quá trình làm sạch đường hô hấp và sự di chuyển của trứng qua ống dẫn trứng, vì tất cả các bộ phận này của cơ thể con người được lót bằng biểu mô có lông.

Bao gồm
Bao gồm- Đây là những thành phần không cố định của tế bào, được hình thành và biến mất trong suốt cuộc đời của nó. Chúng bao gồm cả các chất dự trữ, ví dụ như hạt tinh bột hoặc protein trong tế bào thực vật, hạt glycogen trong tế bào của động vật và nấm, volutin ở vi khuẩn, giọt chất béo trong tất cả các loại tế bào và các chất thải, đặc biệt là dư lượng thực phẩm. không được tiêu hóa do quá trình thực bào, tạo thành cái gọi là thể sót lại.
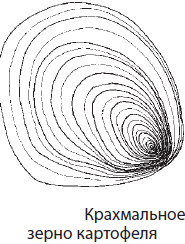
Mối quan hệ giữa cấu trúc và chức năng của các bộ phận, bào quan của tế bào là cơ sở cho tính toàn vẹn của nó
Mỗi bộ phận của tế bào, một mặt, là một cấu trúc riêng biệt với cấu trúc và chức năng cụ thể, mặt khác, là thành phần của một hệ thống phức tạp hơn gọi là tế bào. Hầu hết thông tin di truyền của tế bào nhân chuẩn tập trung ở nhân, nhưng bản thân nhân không thể đảm bảo việc thực hiện nó, vì điều này đòi hỏi ít nhất tế bào chất, đóng vai trò là chất chính và ribosome, nơi diễn ra quá trình tổng hợp này. . Hầu hết các ribosome đều nằm trên mạng lưới nội chất dạng hạt, từ đó protein thường được vận chuyển đến phức hợp Golgi, và sau đó, sau khi sửa đổi, đến các phần của tế bào mà chúng dự định hoặc được bài tiết. Màng bao bì protein và carbohydrate có thể được nhúng vào màng của các bào quan và màng tế bào chất, đảm bảo chúng được đổi mới liên tục. Lysosome và không bào, thực hiện các chức năng quan trọng, cũng tách ra khỏi phức hợp Golgi. Ví dụ, nếu không có lysosome, tế bào sẽ nhanh chóng biến thành nơi chứa các phân tử và cấu trúc chất thải.
Sự xuất hiện của tất cả các quá trình này đòi hỏi năng lượng được tạo ra bởi ty thể và ở thực vật là bởi lục lạp. Và mặc dù các bào quan này tương đối tự chủ, vì chúng có các phân tử DNA riêng nên một số protein của chúng vẫn được bộ gen hạt nhân mã hóa và tổng hợp trong tế bào chất.
Vì vậy, tế bào là một thể thống nhất không thể tách rời của các thành phần cấu thành của nó, mỗi thành phần thực hiện chức năng riêng của mình.
Trao đổi chất và chuyển đổi năng lượng là đặc tính của sinh vật sống. Chuyển hóa năng lượng và nhựa, mối quan hệ của chúng. Các giai đoạn chuyển hóa năng lượng. Lên men và hô hấp. Quang hợp, ý nghĩa của nó, vai trò vũ trụ. Các giai đoạn của quá trình quang hợp Phản ứng sáng và tối của quang hợp, mối quan hệ của chúng. Hóa tổng hợp. Vai trò của vi khuẩn hóa tổng hợp trên Trái đất
Trao đổi chất và chuyển hóa năng lượng - đặc điểm của cơ thể sống
Một tế bào có thể được ví như một nhà máy hóa chất thu nhỏ, trong đó có hàng trăm, hàng nghìn phản ứng hóa học xảy ra.
Sự trao đổi chất- một tập hợp các biến đổi hóa học nhằm bảo tồn và tự tái tạo các hệ thống sinh học.
Nó bao gồm việc đưa các chất vào cơ thể trong quá trình dinh dưỡng và hô hấp, trao đổi chất nội bào hoặc sự trao đổi chất, cũng như việc phân lập các sản phẩm trao đổi chất cuối cùng.
Trao đổi chất gắn bó chặt chẽ với các quá trình chuyển đổi loại năng lượng này sang loại năng lượng khác. Ví dụ, trong quá trình quang hợp, năng lượng ánh sáng được lưu trữ dưới dạng năng lượng liên kết hóa học của các phân tử hữu cơ phức tạp, và trong quá trình hô hấp, nó được giải phóng và dành cho quá trình tổng hợp các phân tử mới, công việc cơ học và thẩm thấu, tiêu tan dưới dạng nhiệt, v.v.
Sự xuất hiện của các phản ứng hóa học trong cơ thể sống được đảm bảo nhờ các chất xúc tác sinh học có tính chất protein - enzim, hoặc enzim. Giống như các chất xúc tác khác, enzym làm tăng tốc độ xảy ra các phản ứng hóa học trong tế bào lên hàng chục, hàng trăm nghìn lần, thậm chí đôi khi còn có thể thực hiện được nhưng không làm thay đổi bản chất hoặc tính chất của (các) sản phẩm cuối cùng của phản ứng và làm không thay đổi chính mình. Enzyme có thể là protein đơn giản và phức tạp, ngoài phần protein còn bao gồm phần phi protein - đồng yếu tố (coenzym). Ví dụ về các enzyme là amylase nước bọt, giúp phân hủy các polysacarit trong quá trình nhai kéo dài và pepsin, đảm bảo quá trình tiêu hóa protein trong dạ dày.
Enzyme khác với các chất xúc tác không phải protein ở tính đặc hiệu cao của hoạt động, tốc độ phản ứng tăng đáng kể với sự trợ giúp của chúng, cũng như khả năng điều chỉnh hoạt động bằng cách thay đổi điều kiện phản ứng hoặc tương tác của các chất khác nhau với chúng. Ngoài ra, các điều kiện xảy ra xúc tác enzyme khác biệt đáng kể so với các điều kiện xảy ra xúc tác không enzyme: nhiệt độ tối ưu cho hoạt động của enzyme trong cơ thể con người là $37°C$, áp suất phải gần với khí quyển và $pH$ của môi trường có thể gây cản trở đáng kể. Vì vậy, amylase cần môi trường kiềm và pepsin cần môi trường axit.
Cơ chế hoạt động của enzyme là làm giảm năng lượng hoạt hóa của các chất (cơ chất) tham gia phản ứng do hình thành phức hợp enzyme-cơ chất trung gian.

Chuyển hóa năng lượng và nhựa, mối quan hệ của chúng
Trao đổi chất bao gồm hai quá trình xảy ra đồng thời trong tế bào: chuyển hóa nhựa và chuyển hóa năng lượng.
Chuyển hóa nhựa (đồng hóa, đồng hóa) là một tập hợp các phản ứng tổng hợp liên quan đến việc tiêu hao năng lượng ATP. Trong quá trình chuyển hóa nhựa, các chất hữu cơ cần thiết cho tế bào được tổng hợp. Ví dụ về các phản ứng trao đổi nhựa là quang hợp, sinh tổng hợp protein và sao chép DNA (tự nhân đôi).
Chuyển hóa năng lượng (dị hóa, hòa tan) là tập hợp các phản ứng phân hủy các chất phức tạp thành chất đơn giản hơn. Là kết quả của quá trình chuyển hóa năng lượng, năng lượng được giải phóng và lưu trữ dưới dạng ATP. Các quá trình chuyển hóa năng lượng quan trọng nhất là hô hấp và lên men.
Trao đổi nhựa và năng lượng có mối liên hệ chặt chẽ với nhau, vì trong quá trình trao đổi nhựa, các chất hữu cơ được tổng hợp và điều này đòi hỏi năng lượng ATP, và trong quá trình trao đổi năng lượng, các chất hữu cơ bị phân hủy và năng lượng được giải phóng, sau đó sẽ được sử dụng cho quá trình tổng hợp .
Các sinh vật nhận năng lượng trong quá trình dinh dưỡng, giải phóng nó và chuyển đổi nó thành dạng dễ tiếp cận chủ yếu trong quá trình hô hấp. Theo phương pháp dinh dưỡng, tất cả các sinh vật được chia thành tự dưỡng và dị dưỡng. Sinh vật tự dưỡng có khả năng tổng hợp độc lập các chất hữu cơ từ các chất vô cơ và dị dưỡng sử dụng các chất hữu cơ được chuẩn bị độc quyền.
Các giai đoạn chuyển hóa năng lượng
Bất chấp sự phức tạp của các phản ứng chuyển hóa năng lượng, nó thường được chia thành ba giai đoạn: chuẩn bị, kỵ khí (không có oxy) và hiếu khí (có oxy).
TRÊN giai đoạn chuẩn bị các phân tử polysaccharide, lipid, protein, axit nucleic phân hủy thành các phân tử đơn giản hơn như glucose, glycerol và các axit béo, axit amin, nucleotide, v.v. Giai đoạn này có thể xảy ra trực tiếp trong tế bào hoặc trong ruột, từ đó cơ thể bị phân hủy. các chất xuống được vận chuyển qua máu.
Giai đoạn kỵ khí Quá trình chuyển hóa năng lượng đi kèm với sự phân hủy sâu hơn các monome của các hợp chất hữu cơ thành các sản phẩm trung gian đơn giản hơn, ví dụ như axit pyruvic hoặc pyruvate. Nó không cần sự hiện diện của oxy và đối với nhiều sinh vật sống trong bùn đầm lầy hoặc trong ruột con người, đó là cách duy nhất để có được năng lượng. Giai đoạn kỵ khí của quá trình chuyển hóa năng lượng xảy ra trong tế bào chất.
Nhiều chất khác nhau có thể bị phân cắt không cần oxy, nhưng chất nền của các phản ứng thường là glucose. Quá trình phân tách không có oxy được gọi là đường phân. Trong quá trình đường phân, một phân tử glucose mất đi bốn nguyên tử hydro, tức là nó bị oxy hóa và hai phân tử axit pyruvic, hai phân tử ATP và hai phân tử chất mang hydro khử $NADH + H^(+)$ được hình thành:
$C_6H_(12)O_6 + 2H_3PO_4 + 2ADP + 2NAD → 2C_3H_4O_3 + 2ATP + 2NADH + H^(+) + 2H_2O$.
Sự hình thành ATP từ ADP xảy ra do sự vận chuyển trực tiếp anion photphat từ đường tiền phosphoryl hóa và được gọi là sự phosphoryl hóa cơ chất.
Giai đoạn hiếu khí trao đổi năng lượng chỉ có thể xảy ra khi có mặt oxy, trong khi các hợp chất trung gian hình thành trong quá trình phân tách không có oxy bị oxy hóa thành sản phẩm cuối cùng (carbon dioxide và nước) và phần lớn năng lượng dự trữ trong liên kết hóa học của các hợp chất hữu cơ được giải phóng. Nó biến thành năng lượng của các liên kết năng lượng cao của 36 phân tử ATP. Giai đoạn này còn được gọi là hô hấp mô. Khi không có oxy, các hợp chất trung gian sẽ được chuyển hóa thành các chất hữu cơ khác, một quá trình gọi là quá trình lên men.
Hơi thở
Cơ chế hô hấp tế bào được mô tả sơ đồ trong hình 2.

Quá trình hô hấp hiếu khí xảy ra ở ty thể, với axit pyruvic đầu tiên mất đi một nguyên tử cacbon, sau đó là sự tổng hợp một chất khử tương đương $NADH + H^(+)$ và một phân tử acetyl coenzym A (acetyl-CoA):
$C_3H_4O_3 + NAD + H~CoA → CH_3CO~CoA + NADH + H^(+) + CO_2$.
Acetyl-CoA trong ma trận ty thể tham gia vào một chuỗi các phản ứng hóa học, toàn bộ phản ứng đó được gọi là chu trình Krebs (chu trình axit tricarboxylic, chu trình axit citric). Trong quá trình biến đổi này, hai phân tử ATP được hình thành, acetyl-CoA bị oxy hóa hoàn toàn thành carbon dioxide, đồng thời các ion và electron hydro của nó được thêm vào các chất mang hydro $NADH + H^(+)$ và $FADH_2$. Các chất mang vận chuyển proton và electron hydro đến màng trong của ty thể, tạo thành các cristae. Với sự trợ giúp của protein vận chuyển, các proton hydro được bơm vào không gian giữa các màng và các electron được truyền qua cái gọi là chuỗi hô hấp của các enzyme nằm trên màng trong của ty thể và thải ra các nguyên tử oxy:
$O_2+2e^(-)→O_2^-$.
Cần lưu ý rằng một số protein chuỗi hô hấp có chứa sắt và lưu huỳnh.
Từ không gian giữa các màng, các proton hydro được vận chuyển trở lại ma trận ty thể với sự trợ giúp của các enzyme đặc biệt - ATP synthase và năng lượng giải phóng trong trường hợp này được dùng để tổng hợp 34 phân tử ATP từ mỗi phân tử glucose. Quá trình này được gọi là oxy hóa phosphoryl. Trong ma trận ty thể, các proton hydro phản ứng với các gốc oxy để tạo thành nước:
$4H^(+)+O_2^-→2H_2O$.
Tập hợp các phản ứng hô hấp oxy có thể được biểu diễn như sau:
$2C_3H_4O_3 + 6O_2 + 36H_3PO_4 + 36ADP → 6CO_2 + 38H_2O + 36ATP.$
Phương trình thở tổng thể trông như thế này:
$C_6H_(12)O_6 + 6O_2 + 38H_3PO_4 + 38ADP → 6CO_2 + 40H_2O + 38ATP.$
Lên men
Trong trường hợp không có oxy hoặc thiếu oxy, quá trình lên men xảy ra. Lên men là một phương pháp tiến hóa sớm hơn để thu được năng lượng so với hô hấp, nhưng nó ít có lợi hơn về mặt năng lượng vì quá trình lên men tạo ra các chất hữu cơ vẫn giàu năng lượng. Có một số loại lên men chính: axit lactic, rượu, axit axetic, v.v. Do đó, trong cơ xương khi không có oxy trong quá trình lên men, axit pyruvic bị khử thành axit lactic, trong khi chất khử tương đương được hình thành trước đó được tiêu thụ và chỉ còn lại hai phân tử ATP:
$2C_3H_4O_3 + 2NADH + H^(+) → 2C_3H_6O_3 + 2NAD$.
Trong quá trình lên men với sự trợ giúp của nấm men, axit pyruvic với sự có mặt của oxy sẽ được chuyển hóa thành rượu etylic và carbon monoxide (IV):
$C_3H_4O_3 + NADH + H^(+) → C_2H_5OH + CO_2 + NAD^(+)$.
Trong quá trình lên men với sự trợ giúp của vi sinh vật, axit axetic, butyric, formic, v.v. cũng có thể được hình thành từ axit pyruvic.
ATP, thu được từ quá trình chuyển hóa năng lượng, được sử dụng trong tế bào cho nhiều loại công việc khác nhau: hóa học, thẩm thấu, điện, cơ học và điều tiết. Công việc hóa học liên quan đến quá trình sinh tổng hợp protein, lipid, carbohydrate, axit nucleic và các hợp chất quan trọng khác. Công việc thẩm thấu bao gồm các quá trình tế bào hấp thụ và loại bỏ khỏi nó các chất có trong không gian ngoại bào với nồng độ lớn hơn trong chính tế bào. Công điện có mối quan hệ chặt chẽ với công thẩm thấu, vì đó là kết quả của sự chuyển động của các hạt tích điện qua màng mà điện tích màng được hình thành và thu được các tính chất dễ bị kích thích và dẫn điện. Công việc cơ học liên quan đến sự chuyển động của các chất và cấu trúc bên trong tế bào, cũng như toàn bộ tế bào. Công việc điều tiết bao gồm tất cả các quy trình nhằm phối hợp các quy trình trong tế bào.
Quang hợp, ý nghĩa của nó, vai trò vũ trụ
quang hợp là quá trình chuyển hóa năng lượng ánh sáng thành năng lượng của các liên kết hóa học của các hợp chất hữu cơ với sự tham gia của diệp lục.
Kết quả của quá trình quang hợp là khoảng 150 tỷ tấn chất hữu cơ và khoảng 200 tỷ tấn oxy được tạo ra hàng năm. Quá trình này đảm bảo chu trình carbon trong sinh quyển, ngăn chặn carbon dioxide tích tụ và từ đó ngăn ngừa hiệu ứng nhà kính và tình trạng quá nóng của Trái đất. Các chất hữu cơ được hình thành do quá trình quang hợp không được các sinh vật khác tiêu thụ hoàn toàn; một phần đáng kể trong số chúng trong suốt hàng triệu năm đã hình thành các mỏ khoáng sản (than cứng và nâu, dầu). Gần đây, dầu hạt cải (“diesel sinh học”) và rượu thu được từ tàn dư thực vật cũng đã bắt đầu được sử dụng làm nhiên liệu. Ozone được hình thành từ oxy dưới tác động của sự phóng điện, tạo thành màn chắn ozone bảo vệ mọi sự sống trên Trái đất khỏi tác động hủy diệt của tia cực tím.
Đồng hương của chúng ta, nhà sinh lý học thực vật xuất sắc K. A. Timiryazev (1843-1920), gọi vai trò của quang hợp là “vũ trụ”, vì nó kết nối Trái đất với Mặt trời (không gian), cung cấp dòng năng lượng cho hành tinh.
Các giai đoạn của quá trình quang hợp Phản ứng sáng và tối của quang hợp, mối quan hệ giữa chúng
Năm 1905, nhà sinh lý học thực vật người Anh F. Blackman đã phát hiện ra rằng tốc độ quang hợp không thể tăng vô thời hạn; Dựa trên điều này, ông đưa ra giả thuyết rằng quá trình quang hợp có hai giai đoạn: ánh sáng Và tối tăm. Ở cường độ ánh sáng yếu, tốc độ phản ứng ánh sáng tăng tỷ lệ thuận với sự tăng cường độ ánh sáng, ngoài ra, những phản ứng này không phụ thuộc vào nhiệt độ vì chúng không cần enzyme để xảy ra. Phản ứng ánh sáng xảy ra trên màng thylakoid.
Ngược lại, tốc độ phản ứng tối tăng khi nhiệt độ tăng, tuy nhiên, khi đạt đến ngưỡng nhiệt độ $30°C$, mức tăng này sẽ dừng lại, điều này cho thấy bản chất enzyme của những biến đổi này xảy ra trong chất nền. Cần lưu ý rằng ánh sáng cũng có ảnh hưởng nhất định đến phản ứng tối, mặc dù thực tế chúng được gọi là phản ứng tối.

Pha sáng của quá trình quang hợp xảy ra trên màng thylakoid mang một số loại phức hợp protein, trong đó chính là hệ thống quang I và II, cũng như ATP synthase. Hệ thống quang học bao gồm các phức hợp sắc tố, ngoài diệp lục còn chứa carotenoid. Carotenoid thu giữ ánh sáng ở những vùng quang phổ mà diệp lục không có, đồng thời bảo vệ diệp lục khỏi bị phá hủy bởi ánh sáng cường độ cao.
Ngoài các phức hợp sắc tố, các hệ thống ảnh còn bao gồm một số protein nhận điện tử, chúng chuyển các điện tử từ các phân tử diệp lục sang nhau một cách tuần tự. Trình tự của các protein này được gọi là chuỗi vận chuyển điện tử của lục lạp.
Một phức hợp protein đặc biệt cũng liên kết với hệ thống quang học II, đảm bảo giải phóng oxy trong quá trình quang hợp. Phức hợp giải phóng oxy này chứa các ion mangan và clo.
TRONG pha sáng lượng tử ánh sáng, hay photon, rơi vào các phân tử diệp lục nằm trên màng thylakoid, chuyển chúng sang trạng thái kích thích, đặc trưng bởi năng lượng điện tử cao hơn. Trong trường hợp này, các electron bị kích thích từ diệp lục của hệ thống ảnh I được chuyển qua một chuỗi trung gian tới chất mang hydro NADP, chất này gắn các proton hydro, luôn có mặt trong dung dịch nước:
$NADP + 2e^(-) + 2H^(+) → NADPH + H^(+)$.
$NADPH + H^(+)$ đã giảm sau đó sẽ được sử dụng trong giai đoạn tối. Các electron từ diệp lục của quang hệ II cũng được chuyển dọc theo chuỗi vận chuyển điện tử, nhưng chúng lấp đầy các “lỗ điện tử” của diệp lục của quang hệ I. Sự thiếu hụt electron trong diệp lục của quang hệ II được lấp đầy bằng cách lấy đi các phân tử nước, điều này xảy ra với sự tham gia của phức hợp giải phóng oxy đã đề cập ở trên. Là kết quả của sự phân hủy các phân tử nước, được gọi là quang phân, các proton hydro được hình thành và oxy phân tử được giải phóng, là sản phẩm phụ của quá trình quang hợp:
$H_2O → 2H^(+) + 2e^(-) + (1)/(2)O_2$.
Thông tin di truyền trong tế bào. Gen, mã di truyền và các đặc tính của nó. Bản chất ma trận của các phản ứng sinh tổng hợp. Sinh tổng hợp protein và axit nucleic
Thông tin di truyền trong tế bào
Sinh sản của chính mình là một trong những đặc tính cơ bản của sinh vật sống. Nhờ hiện tượng này, có sự giống nhau không chỉ giữa các sinh vật mà còn giữa các tế bào riêng lẻ, cũng như các bào quan của chúng (ty thể và lục lạp). Cơ sở vật chất của sự giống nhau này là việc truyền thông tin di truyền được mã hóa theo trình tự nucleotide DNA, được thực hiện thông qua các quá trình sao chép DNA (tự nhân đôi). Tất cả các đặc điểm và tính chất của tế bào và sinh vật được thực hiện nhờ protein, cấu trúc của chúng chủ yếu được xác định bởi trình tự các nucleotide DNA. Do đó, quá trình sinh tổng hợp axit nucleic và protein đóng vai trò hết sức quan trọng trong quá trình trao đổi chất. Đơn vị cấu trúc của thông tin di truyền là gen.
Gen, mã di truyền và các đặc tính của nó
Thông tin di truyền trong tế bào không phải là nguyên khối; nó được chia thành các “từ” riêng biệt - gen.
gen là đơn vị cơ bản của thông tin di truyền.
Công việc trong chương trình “Bộ gen người”, được thực hiện đồng thời ở một số quốc gia và hoàn thành vào đầu thế kỷ này, đã cho chúng tôi hiểu rằng một người chỉ có khoảng 25-30 nghìn gen, nhưng thông tin từ hầu hết DNA của chúng tôi không bao giờ được đọc, vì nó chứa một số lượng lớn các phần vô nghĩa, các phần lặp lại và các gen mã hóa các đặc điểm đã mất ý nghĩa đối với con người (đuôi, lông trên cơ thể, v.v.). Ngoài ra, một số gen chịu trách nhiệm phát triển các bệnh di truyền cũng như gen mục tiêu của thuốc đã được giải mã. Tuy nhiên, việc áp dụng thực tế các kết quả thu được trong quá trình thực hiện chương trình này bị hoãn lại cho đến khi bộ gen của nhiều người hơn được giải mã và người ta thấy rõ chúng khác nhau như thế nào.
Các gen mã hóa cấu trúc bậc một của protein, ribosome hoặc RNA vận chuyển được gọi là cấu trúc và các gen cung cấp sự kích hoạt hoặc ngăn chặn việc đọc thông tin từ các gen cấu trúc - quy định. Tuy nhiên, ngay cả các gen cấu trúc cũng chứa các vùng điều hòa.
Thông tin di truyền của sinh vật được mã hóa trong DNA dưới dạng sự kết hợp nhất định của các nucleotide và trình tự của chúng - mã di truyền. Tính chất của nó là: tính bộ ba, tính đặc hiệu, tính phổ quát, tính dư thừa và không chồng chéo. Ngoài ra, không có dấu chấm câu trong mã di truyền.
Mỗi axit amin được mã hóa trong DNA bởi ba nucleotide - sinh ba, ví dụ, methionine được mã hóa bởi bộ ba TAC, nghĩa là mã là bộ ba. Mặt khác, mỗi bộ ba chỉ mã hóa một axit amin, đó là tính đặc hiệu hoặc rõ ràng của nó. Mã di truyền có tính phổ biến đối với mọi sinh vật sống, tức là vi khuẩn có thể đọc được thông tin di truyền về protein của con người và ngược lại. Điều này cho thấy sự thống nhất về nguồn gốc của thế giới hữu cơ. Tuy nhiên, 64 tổ hợp ba nucleotide chỉ tương ứng với 20 axit amin, do đó một axit amin có thể được mã hóa bởi 2-6 bộ ba, nghĩa là mã di truyền là dư thừa hoặc thoái hóa. Ba bộ ba không có axit amin tương ứng gọi là dừng codon, vì chúng biểu thị sự kết thúc quá trình tổng hợp chuỗi polypeptide.
Trình tự các bazơ trong bộ ba ADN và các axit amin mà chúng mã hóa

*Codon dừng, biểu thị sự kết thúc quá trình tổng hợp chuỗi polypeptide.
Các chữ viết tắt của tên axit amin:
Ala - alanine
Arg - arginine
Asn-măng tây
Asp - axit aspartic
Val - valine
Của anh ấy - histidine
Gly - glycine
Gln - glutamine
Glu - axit glutamic
Ile - isoleucine
Leu - leucine
Liz - lysine
Meth - methionin
Pro-proline
Ser - serine
Tyr - tyrosine
Tre - threonin
Ba - tryptophan
Fen-phenylalanin
Cys - cystein
Nếu bạn bắt đầu đọc thông tin di truyền không phải từ nucleotide đầu tiên trong bộ ba mà từ nucleotide thứ hai, thì không chỉ khung đọc sẽ dịch chuyển mà protein được tổng hợp theo cách này sẽ hoàn toàn khác không chỉ ở trình tự nucleotide mà còn ở cấu trúc và tính chất. Không có dấu chấm câu giữa các bộ ba nên không có trở ngại nào trong việc dịch chuyển khung đọc, điều này mở ra không gian cho sự xuất hiện và duy trì các đột biến.
Bản chất ma trận của các phản ứng sinh tổng hợp
Tế bào vi khuẩn có khả năng nhân đôi sau mỗi 20-30 phút và tế bào nhân chuẩn - hàng ngày và thậm chí thường xuyên hơn, đòi hỏi quá trình sao chép DNA tốc độ cao và chính xác. Ngoài ra, mỗi tế bào còn chứa hàng trăm, hàng nghìn bản sao của nhiều protein, đặc biệt là enzyme, do đó, phương pháp sản xuất “từng phần” là không thể chấp nhận được đối với quá trình sinh sản của chúng. Một phương pháp tiến bộ hơn là dán tem, cho phép bạn có được nhiều bản sao chính xác của sản phẩm và cũng giảm giá thành. Để dập, cần có một ma trận để tạo ấn tượng.
Trong tế bào, nguyên tắc tổng hợp mẫu là các phân tử protein và axit nucleic mới được tổng hợp theo chương trình được nhúng trong cấu trúc của các phân tử có sẵn của cùng một axit nucleic (DNA hoặc RNA).
Sinh tổng hợp protein và axit nucleic
Sao chép DNA. DNA là một polyme sinh học sợi đôi, các monome của nó là các nucleotide. Nếu quá trình sinh tổng hợp DNA diễn ra theo nguyên tắc sao chép thì chắc chắn sẽ phát sinh nhiều biến dạng và sai sót trong thông tin di truyền, cuối cùng dẫn đến cái chết của các sinh vật mới. Vì vậy, quá trình nhân đôi DNA diễn ra khác nhau, một cách nửa bảo thủ: phân tử DNA tháo ra và một chuỗi mới được tổng hợp trên mỗi chuỗi theo nguyên tắc bổ sung. Quá trình tự sinh sản của một phân tử DNA, đảm bảo sao chép chính xác thông tin di truyền và truyền nó từ thế hệ này sang thế hệ khác, được gọi là nhân rộng(từ lat. bản sao- sự lặp lại). Kết quả của sự sao chép là hai bản sao hoàn toàn chính xác của phân tử DNA mẹ được hình thành, mỗi bản sao mang một bản sao của phân tử DNA mẹ.

Quá trình sao chép thực sự cực kỳ phức tạp vì có một số protein tham gia vào quá trình này. Một số trong chúng làm đứt chuỗi xoắn kép của DNA, một số khác phá vỡ liên kết hydro giữa các nucleotide của chuỗi bổ sung, một số khác (ví dụ, enzyme DNA polymerase) chọn lọc các nucleotide mới dựa trên nguyên tắc bổ sung, v.v. Hai phân tử DNA hình thành như một kết quả của sự sao chép phân thành hai trong quá trình phân chia tế bào con mới hình thành.
Lỗi trong quá trình sao chép cực kỳ hiếm khi xảy ra, nhưng nếu chúng xảy ra, chúng sẽ bị loại bỏ rất nhanh bởi cả DNA polymerase và các enzyme sửa chữa đặc biệt, vì bất kỳ lỗi nào trong trình tự nucleotide đều có thể dẫn đến sự thay đổi không thể đảo ngược trong cấu trúc và chức năng của protein. và cuối cùng, ảnh hưởng xấu đến khả năng tồn tại của một tế bào mới hoặc thậm chí là một cá thể.
Sinh tổng hợp protein. Như nhà triết học xuất sắc của thế kỷ 19 F. Engels đã nói một cách hình tượng: “Sự sống là một dạng tồn tại của các cơ thể protein”. Cấu trúc và tính chất của các phân tử protein được xác định bởi cấu trúc bậc một của chúng, tức là trình tự các axit amin được mã hóa trong DNA. Không chỉ sự tồn tại của chính polypeptide mà cả hoạt động của toàn bộ tế bào cũng phụ thuộc vào độ chính xác của việc tái tạo thông tin này, vì vậy quá trình tổng hợp protein có tầm quan trọng rất lớn. Nó dường như là quá trình tổng hợp phức tạp nhất trong tế bào, vì nó liên quan đến tới ba trăm enzym khác nhau và các đại phân tử khác. Ngoài ra, nó chảy ở tốc độ cao, đòi hỏi độ chính xác cao hơn nữa.
Có hai giai đoạn chính trong quá trình sinh tổng hợp protein: phiên mã và dịch mã.
Phiên mã(từ lat. phiên mã- viết lại) là quá trình sinh tổng hợp các phân tử mRNA trên ma trận DNA.

Vì phân tử DNA chứa hai chuỗi đối song song, việc đọc thông tin từ cả hai chuỗi sẽ dẫn đến sự hình thành các mRNA hoàn toàn khác nhau, do đó quá trình sinh tổng hợp của chúng chỉ có thể thực hiện được trên một trong các chuỗi, được gọi là mã hóa, hoặc codogen, trái ngược với chuỗi thứ hai, không mã hóa hoặc không mã hóa. Quá trình viết lại được đảm bảo bởi một enzyme đặc biệt, RNA polymerase, chọn lọc các nucleotide RNA theo nguyên tắc bổ sung. Quá trình này có thể xảy ra cả trong nhân và các bào quan có DNA riêng - ty thể và lục lạp.
Các phân tử mRNA được tổng hợp trong quá trình phiên mã trải qua một quá trình chuẩn bị dịch mã phức tạp (các mRNA ty thể và plastid có thể tồn tại bên trong các bào quan, nơi xảy ra giai đoạn thứ hai của quá trình sinh tổng hợp protein). Trong quá trình trưởng thành của mRNA, ba nucleotide đầu tiên (AUG) và một đuôi nucleotide adenyl được gắn vào nó, độ dài của chúng quyết định số lượng bản sao của protein có thể được tổng hợp trên một phân tử nhất định. Chỉ khi đó các mRNA trưởng thành mới rời khỏi nhân qua các lỗ nhân.
Song song đó, quá trình kích hoạt axit amin xảy ra trong tế bào chất, trong đó axit amin tham gia vào tRNA tự do tương ứng. Quá trình này được xúc tác bởi một enzyme đặc biệt và cần ATP.
Phát tin(từ lat. phát tin- chuyển giao) là quá trình sinh tổng hợp chuỗi polypeptide trên ma trận mRNA, trong đó thông tin di truyền được dịch thành chuỗi axit amin của chuỗi polypeptide.
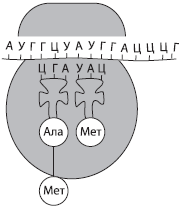
Giai đoạn thứ hai của quá trình tổng hợp protein thường xảy ra nhất trong tế bào chất, ví dụ như ở ER thô. Để nó xuất hiện, cần phải có sự hiện diện của ribosome, kích hoạt tRNA, trong đó chúng gắn các axit amin tương ứng, sự hiện diện của ion Mg2+, cũng như các điều kiện môi trường tối ưu (nhiệt độ, pH, áp suất, v.v.).
Để bắt đầu phát sóng ( bắt đầu) một tiểu đơn vị ribosome nhỏ được gắn vào phân tử mRNA sẵn sàng để tổng hợp, và sau đó, theo nguyên tắc bổ sung với codon đầu tiên (AUG), một tRNA mang axit amin methionine được chọn. Chỉ sau đó tiểu đơn vị ribosome lớn mới gắn vào. Trong ribosome được lắp ráp có hai codon mRNA, codon đầu tiên đã được sử dụng. Một tRNA thứ hai, cũng mang một axit amin, được thêm vào codon liền kề với nó, sau đó liên kết peptide được hình thành giữa các gốc axit amin với sự trợ giúp của các enzym. Ribosome di chuyển một codon của mRNA; tRNA đầu tiên được giải phóng khỏi một axit amin sẽ quay trở lại tế bào chất sau axit amin tiếp theo và một đoạn của chuỗi polypeptide trong tương lai sẽ treo trên tRNA còn lại. TRNA tiếp theo được gắn vào codon mới nằm trong ribosome, quá trình này được lặp lại và chuỗi polypeptide dần dài ra, tức là. sự kéo dài.
Kết thúc quá trình tổng hợp protein ( chấm dứt) xảy ra ngay khi gặp một trình tự nucleotide cụ thể trong phân tử mRNA không mã hóa cho một axit amin (codon dừng). Sau đó, chuỗi ribosome, mRNA và polypeptide được tách ra và protein mới được tổng hợp sẽ có cấu trúc thích hợp và được vận chuyển đến phần tế bào nơi nó sẽ thực hiện các chức năng của mình.
Dịch mã là một quá trình tiêu tốn rất nhiều năng lượng, vì năng lượng của một phân tử ATP được tiêu thụ để gắn một axit amin vào tRNA và một số axit amin khác được sử dụng để di chuyển ribosome dọc theo phân tử mRNA.
Để tăng tốc độ tổng hợp các phân tử protein nhất định, một số ribosome có thể được gắn liên tiếp vào một phân tử mRNA, tạo thành một cấu trúc duy nhất - polysome.
Tế bào là đơn vị di truyền của một sinh vật sống. Nhiễm sắc thể, cấu trúc (hình dạng và kích thước) và chức năng của chúng. Số lượng nhiễm sắc thể và tính hằng định loài của chúng. Tế bào soma và tế bào mầm. Chu kỳ sống của tế bào: xen kẽ và nguyên phân. Nguyên phân là sự phân chia của tế bào soma. Giảm phân. Các giai đoạn của nguyên phân và giảm phân. Sự phát triển của tế bào mầm ở thực vật và động vật. Sự phân chia tế bào là cơ sở cho sự sinh trưởng, phát triển và sinh sản của sinh vật. Vai trò của giảm phân và nguyên phân
Tế bào là đơn vị di truyền của sinh vật.
Mặc dù thực tế là axit nucleic là chất mang thông tin di truyền, nhưng việc thực hiện thông tin này là không thể bên ngoài tế bào, điều này dễ dàng được chứng minh bằng ví dụ về virus. Những sinh vật này, thường chỉ chứa DNA hoặc RNA, không thể sinh sản độc lập; để làm được điều này, chúng phải sử dụng bộ máy di truyền của tế bào. Chúng thậm chí không thể xâm nhập vào tế bào nếu không có sự trợ giúp của chính tế bào đó, ngoại trừ thông qua việc sử dụng các cơ chế vận chuyển màng hoặc do tế bào bị tổn thương. Hầu hết các loại virus đều không ổn định; chúng chết chỉ sau vài giờ tiếp xúc với không khí ngoài trời. Do đó, tế bào là một đơn vị di truyền của một sinh vật sống, có một bộ thành phần tối thiểu để bảo tồn, thay đổi và thực hiện thông tin di truyền cũng như truyền nó cho con cháu.
Hầu hết thông tin di truyền của tế bào nhân chuẩn nằm trong nhân. Điểm đặc biệt trong tổ chức của nó là, không giống như DNA của tế bào nhân sơ, các phân tử DNA của sinh vật nhân chuẩn không khép kín và tạo thành các phức hợp phức tạp với protein - nhiễm sắc thể.
Nhiễm sắc thể, cấu trúc (hình dạng và kích thước) và chức năng của chúng
Nhiễm sắc thể(từ tiếng Hy Lạp crom- màu sắc, màu sắc và soma- cơ thể) là cấu trúc của nhân tế bào, chứa gen và mang thông tin di truyền nhất định về các đặc điểm và tính chất của sinh vật.
Đôi khi các phân tử DNA vòng của sinh vật nhân sơ còn được gọi là nhiễm sắc thể. Nhiễm sắc thể có khả năng tự nhân đôi; chúng có cấu trúc và chức năng riêng biệt và duy trì nó qua nhiều thế hệ. Mỗi tế bào mang tất cả thông tin di truyền của cơ thể, nhưng chỉ một phần nhỏ hoạt động trong đó.
Cơ sở của nhiễm sắc thể là một phân tử DNA sợi đôi chứa đầy protein. Ở sinh vật nhân chuẩn, protein histone và không phải histone tương tác với DNA, trong khi ở sinh vật nhân sơ, protein histone không có.
Nhiễm sắc thể được nhìn thấy rõ nhất dưới kính hiển vi ánh sáng trong quá trình phân chia tế bào, khi do quá trình nén, chúng mang hình dáng của các cơ thể hình que được ngăn cách bởi một cơ thắt sơ cấp - tâm động — trên vai. Trên nhiễm sắc thể cũng có thể có sự co thắt thứ cấp, trong một số trường hợp tách biệt cái gọi là vệ tinh. Các đầu của nhiễm sắc thể được gọi là telomere. Telomere ngăn chặn các đầu của nhiễm sắc thể dính vào nhau và đảm bảo chúng gắn vào màng nhân trong tế bào không phân chia. Khi bắt đầu phân chia, nhiễm sắc thể được nhân đôi và bao gồm hai nhiễm sắc thể con - chất nhiễm sắc, gắn chặt ở tâm động.

Theo hình dạng của chúng, nhiễm sắc thể được chia thành các nhiễm sắc thể có nhánh bằng nhau, nhánh không bằng nhau và hình que. Kích thước của nhiễm sắc thể thay đổi đáng kể, nhưng nhiễm sắc thể trung bình có kích thước 5 $×$ 1,4 micron.
Trong một số trường hợp, nhiễm sắc thể, là kết quả của nhiều lần nhân đôi DNA, chứa hàng trăm, hàng nghìn nhiễm sắc thể: những nhiễm sắc thể khổng lồ như vậy được gọi là polyten. Chúng được tìm thấy trong tuyến nước bọt của ấu trùng Drosophila, cũng như trong tuyến tiêu hóa của giun tròn.
Số lượng nhiễm sắc thể và tính hằng định loài của chúng. Tế bào soma và tế bào mầm
Theo lý thuyết tế bào, tế bào là đơn vị cấu trúc, hoạt động sống và phát triển của cơ thể. Do đó, các chức năng quan trọng của sinh vật như sinh trưởng, sinh sản và phát triển của sinh vật đều được cung cấp ở cấp độ tế bào. Tế bào của sinh vật đa bào có thể được chia thành tế bào soma và tế bào sinh sản.
Tế bào soma- đây là tất cả các tế bào của cơ thể được hình thành do sự phân chia nguyên phân.
Nghiên cứu về nhiễm sắc thể đã cho phép xác định rằng các tế bào soma trong cơ thể của mỗi loài sinh học được đặc trưng bởi số lượng nhiễm sắc thể không đổi. Ví dụ, một người có 46 nhiễm sắc thể của tế bào soma được gọi là. lưỡng bội(2n), hoặc gấp đôi.

Tế bào sinh dục, hoặc giao tử, là những tế bào chuyên biệt được sử dụng để sinh sản hữu tính.
Giao tử luôn chứa số lượng nhiễm sắc thể bằng một nửa so với tế bào soma (ở người - 23), do đó bộ nhiễm sắc thể của tế bào mầm được gọi là đơn bội(n), hoặc đơn. Sự hình thành của nó gắn liền với sự phân chia tế bào giảm phân.
Lượng DNA trong tế bào soma được chỉ định là 2c và trong tế bào sinh dục - 1c. Công thức di truyền của tế bào soma được viết là 2n2c và tế bào sinh dục - 1n1c.
Trong nhân của một số tế bào soma, số lượng nhiễm sắc thể có thể khác với số lượng của chúng trong tế bào soma. Nếu sự khác biệt này lớn hơn một, hai, ba, v.v. các bộ đơn bội thì các ô như vậy được gọi là đa bội(tương ứng là tri-, tetra-, pentaploid). Trong những tế bào như vậy, quá trình trao đổi chất thường diễn ra rất mạnh mẽ.
Bản thân số lượng nhiễm sắc thể không phải là một đặc điểm riêng của loài, vì các sinh vật khác nhau có thể có số lượng nhiễm sắc thể bằng nhau, nhưng những sinh vật có liên quan có thể có số lượng khác nhau. Ví dụ, ký sinh trùng sốt rét và giun đũa ngựa đều có hai nhiễm sắc thể, trong khi con người và tinh tinh có lần lượt là 46 và 48.
Nhiễm sắc thể ở người được chia thành hai nhóm: nhiễm sắc thể thường và nhiễm sắc thể giới tính (dị nhiễm sắc thể). nhiễm sắc thể trong tế bào soma của con người có 22 cặp, chúng giống nhau ở nam và nữ, và nhiễm sắc thể giới tính chỉ có một cặp, nhưng chính điều này quyết định giới tính của cá thể. Có hai loại nhiễm sắc thể giới tính - X và Y. Tế bào cơ thể của phụ nữ mang hai nhiễm sắc thể X và của nam giới - X và Y.
kiểu nhân- đây là tập hợp các đặc điểm của bộ nhiễm sắc thể của một sinh vật (số lượng nhiễm sắc thể, hình dạng và kích thước của chúng).
Bản ghi có điều kiện của kiểu nhân bao gồm tổng số nhiễm sắc thể, nhiễm sắc thể giới tính và các sai lệch có thể có trong bộ nhiễm sắc thể. Ví dụ, kiểu nhân của một người đàn ông bình thường được viết là 46, XY và kiểu nhân của một phụ nữ bình thường là 46, XX.
Chu kỳ sống của tế bào: xen kẽ và nguyên phân
Các tế bào không phải lúc nào cũng sinh ra mới, chúng chỉ được hình thành do sự phân chia của tế bào mẹ. Sau khi phân chia, các tế bào con cần một thời gian để hình thành các bào quan và có được cấu trúc thích hợp để đảm bảo thực hiện một chức năng cụ thể. Khoảng thời gian này được gọi là sự trưởng thành.
Khoảng thời gian từ khi tế bào xuất hiện do sự phân chia cho đến khi tế bào phân chia hoặc chết đi được gọi là vòng đời của một tế bào.
Ở tế bào nhân chuẩn, vòng đời được chia thành hai giai đoạn chính: xen kẽ và nguyên phân.
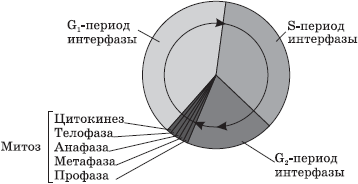
Kỳ trung gian- đây là khoảng thời gian trong vòng đời mà tế bào không phân chia và hoạt động bình thường. Kỳ trung gian được chia thành 3 kỳ: G 1 -, S- và G 2 -.
G 1 -thời gian(tiền tổng hợp, hậu phân bào) là giai đoạn tăng trưởng và phát triển của tế bào, trong đó diễn ra quá trình tổng hợp tích cực RNA, protein và các chất khác cần thiết để hỗ trợ sự sống hoàn chỉnh cho tế bào mới hình thành. Đến cuối giai đoạn này, tế bào có thể bắt đầu chuẩn bị nhân đôi DNA của mình.
TRONG chu kỳ S(tổng hợp) quá trình sao chép DNA tự xảy ra. Phần duy nhất của nhiễm sắc thể không trải qua quá trình sao chép là tâm động, do đó các phân tử DNA thu được không phân kỳ hoàn toàn mà vẫn được giữ lại với nhau trong đó và khi bắt đầu phân chia, nhiễm sắc thể có hình chữ X. Công thức di truyền của tế bào sau khi nhân đôi DNA là 2n4c. Cũng trong thời kỳ S, số trung thể của trung tâm tế bào tăng gấp đôi.
G 2 -thời kỳ(hậu tổng hợp, tiền nguyên phân) được đặc trưng bởi sự tổng hợp mạnh mẽ RNA, protein và ATP cần thiết cho quá trình phân chia tế bào, cũng như sự phân tách các trung thể, ty thể và lục lạp. Cho đến khi kết thúc kỳ trung gian, chất nhiễm sắc và nhân vẫn có thể phân biệt rõ ràng, tính toàn vẹn của lớp vỏ hạt nhân không bị phá vỡ và các bào quan không thay đổi.
Một số tế bào của cơ thể có thể thực hiện các chức năng của chúng trong suốt cuộc đời của cơ thể (tế bào thần kinh của não, tế bào cơ tim), trong khi những tế bào khác tồn tại trong một thời gian ngắn, sau đó chúng chết (tế bào biểu mô ruột, tế bào biểu bì của ruột). làn da). Do đó, cơ thể phải liên tục trải qua quá trình phân chia tế bào và hình thành những tế bào mới thay thế những tế bào đã chết. Tế bào có khả năng phân chia được gọi là thân cây. Trong cơ thể con người, chúng được tìm thấy trong tủy xương đỏ, trong các lớp sâu của biểu bì da và những nơi khác. Sử dụng những tế bào này, bạn có thể phát triển một cơ quan mới, trẻ hóa và nhân bản cơ thể. Triển vọng của việc sử dụng tế bào gốc là hoàn toàn rõ ràng, nhưng các khía cạnh luân lý và đạo đức của vấn đề này vẫn đang được thảo luận, vì trong hầu hết các trường hợp, tế bào gốc phôi thu được từ phôi người bị giết trong quá trình phá thai vẫn được sử dụng.
Thời gian xen kẽ ở tế bào thực vật và động vật trung bình là 10-20 giờ, trong khi quá trình nguyên phân mất khoảng 1-2 giờ.
Trong quá trình phân chia liên tiếp ở các sinh vật đa bào, các tế bào con ngày càng trở nên đa dạng hơn khi chúng đọc thông tin từ số lượng gen ngày càng tăng.
Một số tế bào ngừng phân chia theo thời gian và chết, điều này có thể là do một số chức năng nhất định đã hoàn thành, như trong trường hợp tế bào da biểu bì và tế bào máu, hoặc do các tế bào này bị tổn thương bởi các yếu tố môi trường, đặc biệt là mầm bệnh. Sự chết tế bào theo chương trình di truyền được gọi là sự chết tế bào, trong khi cái chết do tai nạn - hoại tử.
Nguyên phân là sự phân chia của tế bào soma. Các giai đoạn của quá trình nguyên phân
nguyên phân- một phương pháp phân chia gián tiếp các tế bào soma.
Trong quá trình nguyên phân, tế bào trải qua một loạt các giai đoạn liên tiếp, kết quả là mỗi tế bào con nhận được bộ nhiễm sắc thể giống như tế bào mẹ.
Nguyên phân được chia thành bốn giai đoạn chính: tiên tri, metaphase, anaphase và telophase. tiên tri- giai đoạn nguyên phân dài nhất, trong đó chất nhiễm sắc ngưng tụ, dẫn đến nhiễm sắc thể hình X bao gồm hai nhiễm sắc thể (nhiễm sắc thể con) trở nên rõ ràng. Trong trường hợp này, các nhân biến mất, các trung thể phân kỳ về các cực của tế bào và một trục sắc thể (trục phân chia) từ các vi ống bắt đầu hình thành. Vào cuối kỳ tiên tri, màng nhân phân hủy thành các túi riêng biệt.

TRONG siêu hình Các nhiễm sắc thể được xếp dọc theo đường xích đạo của tế bào với tâm động của chúng, nơi gắn các vi ống của trục chính hình thành đầy đủ. Ở giai đoạn phân chia này, các nhiễm sắc thể được nén chặt nhất và có hình dạng đặc trưng, giúp nghiên cứu kiểu nhân.
TRONG phản vệ Sự sao chép DNA nhanh chóng xảy ra ở tâm động, do đó nhiễm sắc thể bị tách ra và các nhiễm sắc thể phân kỳ về các cực của tế bào, được kéo dài bởi các vi ống. Sự phân bố của các nhiễm sắc thể phải hoàn toàn bằng nhau, vì chính quá trình này đảm bảo duy trì số lượng nhiễm sắc thể không đổi trong các tế bào của cơ thể.
Trên sân khấu kỳ cuối các nhiễm sắc thể con tập trung ở hai cực, các màng nhân hình thành xung quanh chúng từ các túi và các hạt nhân xuất hiện trong các nhân mới hình thành.
Sau khi phân chia hạt nhân, sự phân chia tế bào chất xảy ra - sự phân bào, trong đó xảy ra sự phân bố ít nhiều đồng đều của tất cả các bào quan của tế bào mẹ.
Do đó, do nguyên phân, hai tế bào con được hình thành từ một tế bào mẹ, mỗi tế bào là bản sao di truyền của tế bào mẹ (2n2c).
Ở các tế bào bị bệnh, bị tổn thương, lão hóa và các mô chuyên biệt của cơ thể, một quá trình phân chia hơi khác có thể xảy ra - amitosis. amitosisđược gọi là sự phân chia trực tiếp của tế bào nhân chuẩn, trong đó sự hình thành các tế bào tương đương về mặt di truyền không xảy ra do các thành phần tế bào phân bố không đều. Nó được tìm thấy ở thực vật ở nội nhũ và ở động vật - ở gan, sụn và giác mạc của mắt.
Giảm phân. Các giai đoạn của giảm phân
Giảm phân là phương pháp phân chia gián tiếp các tế bào mầm sơ cấp (2n2c), dẫn đến hình thành các tế bào đơn bội (1n1c), thường là tế bào mầm.
Không giống như nguyên phân, giảm phân bao gồm hai lần phân chia tế bào liên tiếp, mỗi lần phân chia đều xảy ra trước kỳ trung gian. Sự phân chia đầu tiên của bệnh teo cơ (meiosis I) được gọi là người theo chủ nghĩa giản lược, vì trong trường hợp này số lượng nhiễm sắc thể giảm đi một nửa và lần phân chia thứ hai (meiosis II) - phương trình, vì trong quá trình của nó, số lượng nhiễm sắc thể được bảo tồn.

Kỳ trung gian I diễn ra giống như kỳ giữa của quá trình nguyên phân. Giảm phân Iđược chia thành bốn giai đoạn: tiên tri I, metaphase I, anaphase I và telophase I. B tiên tri tôi Hai quá trình quan trọng xảy ra: liên hợp và trao đổi chéo. sự chia động từ- Đây là quá trình hợp nhất các nhiễm sắc thể tương đồng (theo cặp) dọc theo toàn bộ chiều dài. Các cặp nhiễm sắc thể hình thành trong quá trình tiếp hợp được bảo tồn cho đến hết kỳ giữa I.
Băng qua- Trao đổi lẫn nhau các vùng tương đồng của nhiễm sắc thể tương đồng. Kết quả của việc trao đổi chéo, các nhiễm sắc thể mà cơ thể nhận được từ cả bố và mẹ sẽ thu được các tổ hợp gen mới, gây ra sự xuất hiện của con cái đa dạng về mặt di truyền. Vào cuối kỳ đầu I, giống như trong kỳ đầu của nguyên phân, nhân biến mất, các trung thể phân kỳ về hai cực của tế bào và màng nhân tan rã.

TRONG siêu hình tôi các cặp nhiễm sắc thể được sắp xếp dọc theo đường xích đạo của tế bào và các vi ống trục chính được gắn vào tâm động của chúng.
TRONG phản vệ tôi Toàn bộ nhiễm sắc thể tương đồng, bao gồm hai nhiễm sắc thể, phân kỳ về hai cực.
TRONG kỳ cuối tôi Màng nhân được hình thành xung quanh các cụm nhiễm sắc thể ở hai cực của tế bào và các nucleoli được hình thành.
Vận động tế bào tôiđảm bảo sự phân tách tế bào chất của tế bào con.
Các tế bào con (1n2c) được hình thành do kết quả của bệnh teo cơ I không đồng nhất về mặt di truyền, vì nhiễm sắc thể của chúng, phân tán ngẫu nhiên đến các cực của tế bào, chứa các gen khác nhau.
Đặc điểm so sánh của nguyên phân và giảm phân
| Dấu hiệu | nguyên phân | Giảm phân | |
| Những tế bào nào bắt đầu phân chia? | Cơ thể (2n) | Tế bào mầm sơ cấp (2n) | |
| Số lượng phân chia | 1 | 2 | |
| Có bao nhiêu và loại tế bào nào được hình thành trong quá trình phân chia? | 2 soma (2n) | 4 tình dục (n) | |
| Kỳ trung gian | Chuẩn bị tế bào phân chia, nhân đôi DNA | Rất ngắn, DNA nhân đôi không xảy ra | |
| Giai đoạn | Giảm phân I | Giảm phân II | |
| tiên tri | Có thể xảy ra hiện tượng ngưng tụ nhiễm sắc thể, biến mất nhân, phân rã màng nhân, liên hợp và trao đổi chéo | Sự ngưng tụ nhiễm sắc thể, sự biến mất của nucleolus, sự phân hủy của màng nhân | |
| Siêu hình | Các cặp nhiễm sắc thể nằm dọc theo đường xích đạo, hình thành trục chính | Nhiễm sắc thể xếp dọc theo xích đạo, hình thành trục chính | |
| Kỳ sau | Các nhiễm sắc thể tương đồng của hai nhiễm sắc thể di chuyển về hai cực | Các nhiễm sắc thể di chuyển về phía cực | |
| Kỳ cuối | Nhiễm sắc thể xoắn, màng nhân và nucleoli mới được hình thành | Nhiễm sắc thể xoắn, màng nhân và nucleoli mới được hình thành | |
Kỳ trung gian II rất ngắn, vì quá trình nhân đôi DNA không xảy ra trong đó, nghĩa là không có chu kỳ S.
Giảm phân II cũng được chia thành bốn giai đoạn: tiên tri II, metaphase II, anaphase II và telophase II. TRONG tiên tri II các quá trình tương tự xảy ra như ở kì đầu I, ngoại trừ sự chia động từ và chuyển giao chéo.
TRONG kỳ giữa II nhiễm sắc thể nằm dọc theo đường xích đạo của tế bào.
TRONG phản vệ II nhiễm sắc thể được phân chia ở tâm động và nhiễm sắc thể được kéo dài về phía cực.
TRONG kì cuối II Màng nhân và nucleoli được hình thành xung quanh các cụm nhiễm sắc thể con gái.
Sau đó tế bào học II Công thức di truyền của cả bốn tế bào con là 1n1c, nhưng chúng đều có bộ gen khác nhau, là kết quả của quá trình trao đổi chéo và sự kết hợp ngẫu nhiên các nhiễm sắc thể của sinh vật mẹ và sinh vật cha trong tế bào con.
Sự phát triển của tế bào mầm ở thực vật và động vật
Phát sinh giao tử(từ tiếng Hy Lạp giao tử- vợ, giao tử- chồng và nguồn gốc- nguồn gốc, sự xuất hiện) là quá trình hình thành tế bào mầm trưởng thành.
Vì sinh sản hữu tính thường đòi hỏi hai cá thể - một con cái và một con đực, tạo ra các tế bào sinh dục khác nhau - trứng và tinh trùng, nên quá trình hình thành các giao tử này phải khác nhau.
Bản chất của quá trình này phụ thuộc rất nhiều vào việc nó xảy ra ở tế bào thực vật hay động vật, vì ở thực vật chỉ có quá trình nguyên phân xảy ra trong quá trình hình thành giao tử, còn ở động vật thì cả quá trình nguyên phân và giảm phân đều xảy ra.
Sự phát triển của tế bào mầm ở thực vật.Ở thực vật hạt kín, sự hình thành tế bào sinh sản đực và cái xảy ra ở các phần khác nhau của hoa - tương ứng là nhị hoa và nhụy hoa.
Trước khi hình thành tế bào sinh sản nam - sự phát sinh vi giao tử(từ tiếng Hy Lạp vi mô- nhỏ) - xảy ra sự hình thành vi bào tử, tức là sự hình thành các vi bào tử trong bao phấn của nhị hoa. Quá trình này gắn liền với sự phân chia vi bào tử của tế bào mẹ, tạo ra 4 vi bào tử đơn bội. Quá trình tạo giao tử có liên quan đến sự phân chia phân bào của vi bào tử, tạo ra giao tử đực từ hai tế bào - một thể giao tử lớn thực vật(siphonogen) và nông có tính sinh sản. Sau khi phân chia, giao tử đực được bao phủ bởi màng dày đặc và tạo thành hạt phấn hoa. Trong một số trường hợp, ngay cả trong quá trình trưởng thành của phấn hoa và đôi khi chỉ sau khi chuyển đến đầu nhụy của nhụy hoa, tế bào thế hệ mới phân chia theo kiểu phân bào để tạo thành hai tế bào mầm đực bất động - tinh trùng. Sau khi thụ phấn, một ống phấn được hình thành từ tế bào sinh dưỡng, qua đó tinh trùng xâm nhập vào buồng trứng của nhụy hoa để thụ tinh.

Sự phát triển của tế bào mầm cái ở thực vật được gọi là siêu giao tử(từ tiếng Hy Lạp mega- to lớn). Nó xảy ra ở buồng trứng của nhụy hoa, trước đó là sự phát sinh bào tử, kết quả là bốn đại bào tử được hình thành từ tế bào mẹ của đại bào tử nằm trong nhân tế bào thông qua quá trình phân chia giảm nhiễm. Một trong những đại bào tử phân chia theo phương pháp phân bào ba lần, tạo ra giao tử cái - một túi phôi có tám nhân. Với sự phân tách tiếp theo của tế bào chất của các tế bào con, một trong những tế bào thu được sẽ trở thành một quả trứng, ở hai bên của nó có cái gọi là chất tổng hợp, ở đầu đối diện của túi phôi, ba phản cực được hình thành và ở trung tâm. , là kết quả của sự hợp nhất của hai nhân đơn bội, một tế bào trung tâm lưỡng bội được hình thành.
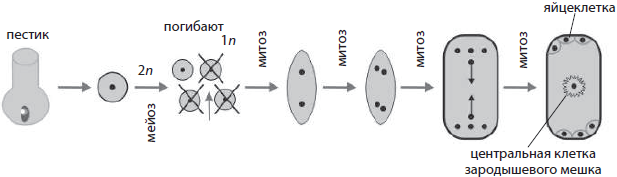
Sự phát triển của tế bào mầm ở động vật.Ở động vật, có hai quá trình hình thành tế bào mầm - sinh tinh và sinh trứng.
sinh tinh(từ tiếng Hy Lạp tinh trùng, tinh trùng- hạt giống và nguồn gốc- nguồn gốc, sự xuất hiện) là quá trình hình thành tế bào mầm đực trưởng thành - tinh trùng. Ở người, nó xảy ra ở tinh hoàn và được chia thành bốn giai đoạn: sinh sản, tăng trưởng, trưởng thành và hình thành.

TRONG mùa sinh sản Tế bào mầm nguyên thủy phân chia theo nguyên phân, tạo thành thể lưỡng bội nguyên bào sinh tinh. TRONG thời kỳ tăng trưởng tinh trùng tích tụ chất dinh dưỡng trong tế bào chất, tăng kích thước và biến thành tế bào sinh tinh sơ cấp, hoặc Tế bào sinh tinh bậc 1. Chỉ sau đó chúng mới bước vào quá trình giảm phân ( thời kỳ trưởng thành), kết quả là hai cái đầu tiên được hình thành tinh bào thứ cấp, hoặc tinh trùng bậc 2, và sau đó - bốn tế bào đơn bội với số lượng tế bào chất khá lớn - tinh trùng. TRONG thời kỳ hình thành chúng mất gần như toàn bộ tế bào chất và hình thành tiên mao, biến thành tinh trùng.
tinh trùng, hoặc sinh động, - tế bào sinh sản đực di động rất nhỏ có đầu, cổ và đuôi.

TRONG cái đầu, ngoài cốt lõi, là đầu cực- một phức hợp Golgi được sửa đổi đảm bảo sự hòa tan của màng trứng trong quá trình thụ tinh. TRONG cổ tử cung là trung tâm của trung tâm tế bào và là cơ sở tóc đuôi gà hình thành các vi ống hỗ trợ trực tiếp cho sự di chuyển của tinh trùng. Nó cũng chứa ty thể, cung cấp năng lượng ATP cho tinh trùng để di chuyển.
quá trình tạo trứng(từ tiếng Hy Lạp LHQ- trứng và nguồn gốc- nguồn gốc, sự xuất hiện) là quá trình hình thành tế bào mầm cái trưởng thành - trứng. Ở người, nó xảy ra ở buồng trứng và bao gồm ba giai đoạn: sinh sản, sinh trưởng và trưởng thành. Các giai đoạn sinh sản và tăng trưởng, tương tự như giai đoạn sinh tinh, xảy ra trong quá trình phát triển trong tử cung. Trong trường hợp này, các tế bào lưỡng bội được hình thành từ các tế bào mầm sơ cấp do quá trình nguyên phân. oogonia, sau đó chuyển thành lưỡng bội sơ cấp tế bào trứng, hoặc Tế bào trứng bậc 1. Giảm phân và quá trình phân bào tiếp theo xảy ra ở thời kỳ trưởng thành, được đặc trưng bởi sự phân chia tế bào chất của tế bào mẹ không đồng đều, do đó, kết quả là lúc đầu thu được một tế bào chất tế bào trứng thứ cấp, hoặc tế bào trứng bậc 2, Và Cơ cực phân cực đầu tiên, và sau đó từ tế bào trứng thứ cấp - trứng, nơi giữ lại toàn bộ nguồn cung cấp chất dinh dưỡng, và thể cực thứ hai, trong khi thể cực thứ nhất được chia thành hai. Cơ thể cực chiếm vật liệu di truyền dư thừa.
Ở người, trứng được sản xuất trong khoảng thời gian 28-29 ngày. Chu kỳ liên quan đến sự trưởng thành và giải phóng trứng được gọi là kinh nguyệt.
Trứng- một tế bào sinh sản cái lớn không chỉ mang bộ nhiễm sắc thể đơn bội mà còn là nguồn cung cấp chất dinh dưỡng đáng kể cho sự phát triển tiếp theo của phôi.

Trứng ở động vật có vú được bao phủ bởi bốn màng, giúp giảm khả năng bị hư hại bởi các yếu tố khác nhau. Đường kính của trứng ở người đạt 150-200 micron, trong khi ở đà điểu có thể lên tới vài cm.
Sự phân chia tế bào là cơ sở cho sự sinh trưởng, phát triển và sinh sản của sinh vật. Vai trò của nguyên phân và giảm phân
Nếu ở sinh vật đơn bào, sự phân chia tế bào dẫn đến sự gia tăng số lượng cá thể, tức là sinh sản, thì ở sinh vật đa bào, quá trình này có thể có những ý nghĩa khác nhau. Như vậy, sự phân chia tế bào phôi, bắt đầu từ hợp tử, là cơ sở sinh học của các quá trình sinh trưởng và phát triển liên kết với nhau. Những thay đổi tương tự cũng được quan sát thấy ở con người trong thời niên thiếu, khi số lượng tế bào không chỉ tăng lên mà còn xảy ra những thay đổi về chất trong cơ thể. Sự sinh sản của sinh vật đa bào cũng dựa trên sự phân chia tế bào, ví dụ, trong sinh sản vô tính, nhờ quá trình này, toàn bộ cơ thể được phục hồi, và trong sinh sản hữu tính, trong quá trình phát sinh giao tử, các tế bào giới tính được hình thành, sau đó hình thành cơ thể mới. Cần lưu ý rằng các phương pháp phân chia chính của tế bào nhân chuẩn - nguyên phân và giảm phân - có những ý nghĩa khác nhau trong vòng đời của sinh vật.
Kết quả của quá trình nguyên phân là có sự phân bố đồng đều vật chất di truyền giữa các tế bào con - bản sao chính xác của mẹ. Nếu không có nguyên phân, sự tồn tại và phát triển của các sinh vật đa bào phát triển từ một tế bào duy nhất, hợp tử, sẽ không thể xảy ra, vì tất cả các tế bào của những sinh vật đó phải chứa cùng một thông tin di truyền.
Trong quá trình phân chia, các tế bào con ngày càng đa dạng về cấu trúc và chức năng, điều này gắn liền với việc kích hoạt ngày càng nhiều nhóm gen mới trong chúng do sự tương tác giữa các tế bào. Vì vậy, nguyên phân là cần thiết cho sự phát triển của sinh vật.
Phương pháp phân chia tế bào này là cần thiết cho quá trình sinh sản vô tính và tái tạo (phục hồi) các mô cũng như các cơ quan bị tổn thương.
Ngược lại, bệnh teo cơ đảm bảo sự ổn định của kiểu nhiễm sắc thể trong quá trình sinh sản hữu tính, vì nó làm giảm một nửa bộ nhiễm sắc thể trước khi sinh sản hữu tính, sau đó được phục hồi nhờ quá trình thụ tinh. Ngoài ra, bệnh teo cơ dẫn đến sự xuất hiện của các tổ hợp gen mới của bố mẹ do sự trao đổi chéo và sự kết hợp ngẫu nhiên của các nhiễm sắc thể trong tế bào con. Nhờ đó, con cái trở nên đa dạng về mặt di truyền, cung cấp nguyên liệu cho chọn lọc tự nhiên và là cơ sở vật chất cho quá trình tiến hóa. Một mặt, sự thay đổi về số lượng, hình dạng và kích thước của nhiễm sắc thể có thể dẫn đến sự xuất hiện của nhiều sai lệch khác nhau trong quá trình phát triển của sinh vật và thậm chí là cái chết của sinh vật, mặt khác, nó có thể dẫn đến sự xuất hiện của các cá thể. thích nghi hơn với môi trường.
Như vậy, tế bào là đơn vị sinh trưởng, phát triển và sinh sản của sinh vật.
Chúng tôi mời bạn làm quen với các tài liệu và.
: màng xenlulo, màng, tế bào chất với bào quan, nhân, không bào với nhựa tế bào.Sự hiện diện của plastid là đặc điểm chính của tế bào thực vật.
Chức năng của màng tế bào- quyết định hình dạng của tế bào, bảo vệ chống lại các yếu tố môi trường.
Màng huyết tương- một màng mỏng, bao gồm các phân tử tương tác giữa lipid và protein, ngăn cách các chất bên trong với môi trường bên ngoài, đảm bảo vận chuyển nước, khoáng chất và các chất hữu cơ vào tế bào bằng thẩm thấu và vận chuyển tích cực, đồng thời loại bỏ các chất thải.
Tế bào chất- môi trường bán lỏng bên trong tế bào, trong đó có nhân và các bào quan, cung cấp các kết nối giữa chúng và tham gia vào các quá trình sống cơ bản.
Lưới nội chất- mạng lưới các kênh phân nhánh trong tế bào chất. Nó tham gia vào quá trình tổng hợp protein, lipid và carbohydrate và vận chuyển các chất. Ribosome là các cơ thể nằm trên ER hoặc trong tế bào chất, bao gồm RNA và protein và tham gia vào quá trình tổng hợp protein. EPS và ribosome là một bộ máy duy nhất để tổng hợp và vận chuyển protein.
ty thể- Các bào quan được phân cách với tế bào chất bằng hai màng. Các chất hữu cơ bị oxy hóa trong đó và các phân tử ATP được tổng hợp với sự tham gia của các enzyme. Tăng bề mặt của màng bên trong nơi chứa các enzyme do cristae. ATP là một chất hữu cơ giàu năng lượng.
Plastid(lục lạp, bạch cầu, sắc lạp), hàm lượng của chúng trong tế bào là đặc điểm chính của cơ thể thực vật. Lục lạp là các plastid chứa chất diệp lục sắc tố màu xanh lá cây, hấp thụ năng lượng ánh sáng và sử dụng nó để tổng hợp các chất hữu cơ từ carbon dioxide và nước. Lục lạp được tách ra khỏi tế bào chất bằng hai màng, nhiều phần phát triển - grana trên màng bên trong, trong đó có các phân tử diệp lục và enzyme.
phức hợp Golgi- một hệ thống các khoang được phân cách với tế bào chất bằng một màng. Sự tích tụ protein, chất béo và carbohydrate trong đó. Thực hiện quá trình tổng hợp chất béo và carbohydrate trên màng.
Lysosome- Các cơ thể được phân cách với tế bào chất bằng một màng duy nhất. Các enzym chứa trong chúng đẩy nhanh quá trình phân hủy các phân tử phức tạp thành các phân tử đơn giản: protein thành axit amin, carbohydrate phức tạp thành đơn giản, lipid thành glycerol và axit béo, đồng thời phá hủy các phần chết của tế bào và toàn bộ tế bào.
không bào- các khoang trong tế bào chất chứa đầy nhựa tế bào, nơi tích tụ các chất dinh dưỡng dự trữ và các chất có hại; chúng điều chỉnh hàm lượng nước trong tế bào.
Cốt lõi- phần chính của tế bào, được bao phủ bên ngoài bằng một lớp vỏ hạt nhân hai màng, có lỗ rỗng. Các chất đi vào lõi và được loại bỏ khỏi nó qua các lỗ chân lông. Nhiễm sắc thể là vật mang thông tin di truyền về các đặc điểm của cơ thể, cấu trúc chính của nhân, mỗi hạt bao gồm một phân tử DNA kết hợp với protein. Nhân là nơi tổng hợp DNA, mRNA và r-RNA.

Sự hiện diện của màng ngoài, tế bào chất với các bào quan và nhân có nhiễm sắc thể.
Màng ngoài hoặc màng sinh chất- phân định các chất trong tế bào với môi trường (tế bào khác, chất nội bào), bao gồm các phân tử lipid và protein, đảm bảo sự liên lạc giữa các tế bào, vận chuyển các chất vào tế bào (pinocytosis, phagocytosis) và ra khỏi tế bào.
Tế bào chất- môi trường bán lỏng bên trong tế bào, cung cấp sự liên lạc giữa nhân và các bào quan nằm trong đó. Các quá trình sống chính diễn ra trong tế bào chất.
Các bào quan của tế bào:
1) lưới nội chất (ER)- một hệ thống ống phân nhánh, tham gia vào quá trình tổng hợp protein, lipid và carbohydrate, tham gia vận chuyển các chất trong tế bào;
2) ribosome- các cơ thể chứa rRNA nằm trên ER và trong tế bào chất và tham gia tổng hợp protein. EPS và ribosome là một bộ máy duy nhất để tổng hợp và vận chuyển protein;
3) ty thể- “Trạm năng lượng” của tế bào, được phân cách với tế bào chất bằng hai màng. Phần bên trong tạo thành cristae (nếp gấp), làm tăng bề mặt của nó. Enzyme trên cristae đẩy nhanh quá trình oxy hóa các chất hữu cơ và tổng hợp các phân tử ATP giàu năng lượng;
4) phức hợp Golgi- một nhóm các khoang được giới hạn bởi một màng tế bào chất, chứa đầy protein, chất béo và carbohydrate, được sử dụng trong các quá trình quan trọng hoặc được loại bỏ khỏi tế bào. Các màng của phức hợp thực hiện quá trình tổng hợp chất béo và carbohydrate;
5) lysosome- Cơ thể chứa đầy enzyme đẩy nhanh quá trình phân hủy protein thành axit amin, lipid thành glycerol và axit béo, polysacarit thành monosacarit. Trong lysosome, các phần chết của tế bào, toàn bộ tế bào, bị phá hủy.
Bao gồm tế bào- tích lũy các chất dinh dưỡng dự trữ: protein, chất béo và carbohydrate.
Cốt lõi- phần quan trọng nhất của tế bào. Nó được bao phủ bởi một lớp vỏ màng đôi có lỗ chân lông, qua đó một số chất xâm nhập vào nhân và một số khác đi vào tế bào chất. Nhiễm sắc thể là cấu trúc chính của nhân, mang thông tin di truyền về các đặc điểm của cơ thể. Nó được truyền trong quá trình phân chia tế bào mẹ sang tế bào con và từ tế bào mầm sang cơ thể con. Nhân là nơi tổng hợp DNA, mRNA và rRNA.
Bài tập:
Giải thích tại sao bào quan được gọi là cấu trúc tế bào chuyên biệt?
Trả lời: các bào quan được gọi là cấu trúc tế bào chuyên biệt, vì chúng thực hiện các chức năng được xác định nghiêm ngặt, thông tin di truyền được lưu trữ trong nhân, ATP được tổng hợp trong ty thể, quá trình quang hợp xảy ra ở lục lạp, v.v.
Nếu có thắc mắc về tế bào học, bạn có thể liên hệ
tóm tắt các bài thuyết trình khác“Phương pháp dạy học sinh học” - Động vật học trường học. Giới thiệu cho học sinh cách sử dụng dữ liệu động vật học khoa học. Giáo dục đạo đức. Phước lành bổ sung của chuồng gà. Lựa chọn các phương pháp. Quy trình sống. Cá cảnh. Dinh dưỡng. Giáo dục môi trường. Tính vật chất của các quá trình sống. Kết quả tiêu cực. Các bạn sinh viên chú ý nhé. Mẫu bắt buộc. Nhìn vào động vật nhỏ. Mục đích và mục tiêu của sinh học. Câu chuyện.
“Học dựa trên vấn đề trong bài học sinh học” - Kiến thức. Sách giáo khoa mới. Con đường dẫn đến giải pháp. Vấn đề. Hội thảo. Một nhiệm vụ là gì? Albrecht Durer. Dạy học dựa trên vấn đề trong bài học sinh học. Bài học không chuẩn. Học tập dựa trên vấn đề có nghĩa là gì? Chất lượng cuộc sống. Sinh học như một môn học. Câu hỏi. Bài học giải quyết vấn đề. Giảm sự quan tâm đến chủ đề này. Các lớp học thí nghiệm dựa trên vấn đề.
“Tư duy phê phán trong bài học sinh học” - Công nghệ “tư duy phê phán”. Sử dụng công nghệ “phát triển tư duy phản biện”. Bảng cho bài học. Động lực học tập. Hệ sinh thái. Ý nghĩa của việc “phát triển tư duy phản biện”. Dấu hiệu của công nghệ. Công nghệ RKM. Cấu trúc bài học. Các hướng chính. Lịch sử công nghệ. Công nghệ sư phạm. Quy luật công nghệ. Bài tập sinh học. Quang hợp. Các kỹ thuật được sử dụng ở các giai đoạn khác nhau của bài học.
“Bài học Sinh học với bảng tương tác” - Sách giáo khoa điện tử. Lợi ích cho sinh viên. Bảng trắng tương tác giúp truyền tải thông tin đến từng học sinh. Nhiệm vụ giáo khoa. Giải quyết các vấn đề sinh học. Ưu điểm khi làm việc với bảng trắng tương tác. Làm việc với bài thuyết trình. Làm việc để so sánh các đối tượng. Đồ vật chuyển động. Sử dụng bảng tính. Sử dụng bảng tương tác trong quá trình dạy học cho học sinh. Lợi ích cho giáo viên.
“Phương pháp tiếp cận hoạt động hệ thống trong sinh học” - Câu hỏi hội thảo. Phương pháp hoạt động. Dryopithecus. Con đường ngoài trái đất có nguồn gốc từ con người. Lysosome. Tổ chức hóa học Thực vật hạt trần. Sự trao đổi chất. Máy phân tích. Phương pháp hoạt động hệ thống trong dạy học sinh học. Nhiễm sắc thể. Tế bào chất. Mù quáng. Chiều dài tai. Phân loại một người. Bộ xương của động vật có vú. Con đường tiến hóa của loài người. Nguyên phân. Phức hợp bề ngoài. Câu hỏi có vấn đề. Hạt nhân. Màng nhân.
“Máy tính trong Sinh học” - Hoạt động hợp tác của học sinh. Họ thực vật hạt kín. Đào tạo tương tác. Các mô hình học tập. Ví dụ về hệ thống đánh giá Câu hỏi về thẻ hướng dẫn. Ví dụ về thẻ hướng dẫn Các nhà nghiên cứu. Nhóm vi mô. Công nghệ học tập tương tác. Băng chuyền. Công nghệ học tập tương tác. Phương pháp tương tác trong bài học sinh học. Hình thức làm việc nhóm. Nhiệm vụ của nhóm “nhà nghiên cứu”.
Mối liên hệ của sinh vật với môi trường, theo quan điểm hóa lý, là một hệ thống mở, tức là một hệ thống trong đó các quá trình sinh hóa đang diễn ra. Các chất ban đầu đến từ môi trường, còn các chất cũng liên tục được hình thành sẽ được đưa ra bên ngoài. Sự cân bằng giữa tốc độ và nồng độ sản phẩm của các phản ứng đa chiều trong cơ thể là có điều kiện, tưởng tượng, vì việc hấp thụ và loại bỏ các chất không dừng lại. Sự kết nối liên tục với môi trường cho phép chúng ta coi sinh vật sống như một hệ thống mở.
Đối với mọi tế bào sống, nguồn năng lượng là Mặt trời. Tế bào thực vật thu năng lượng từ ánh sáng mặt trời với sự trợ giúp của chất diệp lục, sử dụng nó cho các phản ứng đồng hóa trong quá trình quang hợp. Tế bào của động vật, nấm và vi khuẩn sử dụng năng lượng mặt trời một cách gián tiếp, trong quá trình phân hủy các chất hữu cơ được tổng hợp bởi thực vật trên trái đất.
Một số chất dinh dưỡng của tế bào bị phân hủy trong quá trình hô hấp tế bào, do đó cung cấp năng lượng cần thiết cho nhiều loại hoạt động của tế bào. Quá trình này diễn ra trong các bào quan gọi là ty thể. Ty thể bao gồm hai màng: màng ngoài, ngăn cách cơ quan với tế bào chất và màng trong, tạo thành nhiều nếp gấp. Sản phẩm chính của hô hấp là ATP. Nó rời khỏi ty thể và được sử dụng làm nguồn năng lượng cho nhiều phản ứng hóa học trong tế bào chất và màng tế bào. Nếu oxy cần thiết cho quá trình hô hấp của tế bào thì hô hấp được gọi là hiếu khí, nhưng nếu các phản ứng xảy ra trong điều kiện không có oxy thì chúng ta gọi là hô hấp kỵ khí.
Đối với bất kỳ loại công việc nào được thực hiện trong tế bào, năng lượng được sử dụng ở một dạng duy nhất - dưới dạng năng lượng từ các liên kết photphat của ATP. ATP là một hợp chất dễ di chuyển. Sự hình thành ATP xảy ra ở màng trong của ty thể. ATP được tổng hợp ở tất cả các tế bào trong quá trình hô hấp nhờ năng lượng oxy hóa carbohydrate, chất béo và các chất hữu cơ khác. Ở tế bào thực vật xanh, lượng ATP chủ yếu được tổng hợp ở lục lạp nhờ năng lượng mặt trời. Trong quá trình quang hợp, chúng tạo ra lượng ATP nhiều gấp nhiều lần so với ty thể. ATP bị phân hủy làm đứt liên kết phốt pho-oxy và giải phóng năng lượng. Điều này xảy ra dưới tác dụng của enzyme ATPase trong quá trình thủy phân ATP - bổ sung nước và loại bỏ phân tử axit photphoric. Kết quả là ATP được chuyển đổi thành ADP và nếu hai phân tử axit photphoric bị tách ra thì thành AMP. Phản ứng loại bỏ từng gram phân tử axit đi kèm với việc giải phóng 40 kJ. Đây là sản lượng năng lượng rất lớn, đó là lý do tại sao liên kết phốt pho-oxy của ATP thường được gọi là vĩ mô (năng lượng cao).
Việc sử dụng ATP trong các phản ứng trao đổi nhựa được thực hiện bằng cách ghép chúng với quá trình thủy phân ATP. Các phân tử của nhiều chất khác nhau được tích năng lượng bằng cách gắn nhóm phốt pho được giải phóng trong quá trình thủy phân từ phân tử ATP, tức là bằng quá trình phosphoryl hóa.
Điểm đặc biệt của các dẫn xuất photphat là chúng không thể rời khỏi tế bào, mặc dù dạng “thải” của chúng tự do đi qua màng. Nhờ đó, các phân tử được phosphoryl hóa vẫn tồn tại trong tế bào cho đến khi chúng được sử dụng trong các phản ứng thích hợp.
Quá trình ngược lại chuyển đổi ADP thành ATP xảy ra bằng cách thêm một phân tử axit photphoric vào ADP, giải phóng nước và hấp thụ một lượng lớn năng lượng.
Như vậy, ATP là nguồn năng lượng phổ biến và trực tiếp cho hoạt động của tế bào. Điều này tạo ra một nguồn năng lượng duy nhất cho tế bào và giúp phân phối lại và vận chuyển nó từ vùng này sang vùng khác của tế bào.
Việc chuyển nhóm photphat đóng vai trò quan trọng trong các phản ứng hóa học như sự lắp ráp các đại phân tử từ các đơn phân. Ví dụ, các axit amin chỉ có thể được kết hợp thành peptide sau khi được phosphoryl hóa trước đó. Các quá trình cơ học co lại hoặc chuyển động, vận chuyển chất hòa tan ngược với gradient nồng độ và các quá trình khác liên quan đến việc tiêu thụ năng lượng dự trữ trong ATP.
Quá trình chuyển hóa năng lượng có thể được biểu diễn như sau. Các chất hữu cơ phân tử cao trong tế bào chất được enzyme, thủy phân, chuyển đổi thành các chất đơn giản hơn, từ đó chúng bao gồm: protein - thành axit amin, poly- và disacarit - thành monosacarit (+ glucose), chất béo thành glycerol và axit béo. Không có quá trình oxy hóa, ít năng lượng được giải phóng, không được sử dụng và chuyển thành dạng nhiệt. Hầu hết các tế bào sử dụng carbohydrate đầu tiên. Polysaccharides (tinh bột trong thực vật và glycogen ở động vật) bị thủy phân thành glucose. Quá trình oxy hóa glucose xảy ra theo ba giai đoạn: glycolysis, quá trình khử carboxyl oxy hóa (chu trình Krebs - chu trình axit citric) và quá trình phosphoryl hóa oxy hóa (chuỗi hô hấp). Quá trình đường phân, do đó một phân tử glucose được chia thành hai phân tử axit pyruvic với sự giải phóng hai phân tử ATP, xảy ra trong tế bào chất. Khi không có oxy, axit pyruvic được chuyển thành ethanol (lên men) hoặc axit lactic (hô hấp kỵ khí).
Khi quá trình đường phân xảy ra ở tế bào động vật, phân tử glucose sáu carbon sẽ phân hủy thành hai phân tử axit lactic. Quá trình này là nhiều giai đoạn. Nó được thực hiện tuần tự bởi 13 enzyme. Trong quá trình lên men rượu, hai phân tử ethanol và hai phân tử CO2 được hình thành từ một phân tử glucose.
Glycolysis là một giai đoạn phổ biến của hô hấp kỵ khí và hiếu khí; hai giai đoạn còn lại chỉ xảy ra trong điều kiện hiếu khí. Quá trình oxy hóa không có oxy, trong đó chỉ một phần năng lượng của các chất chuyển hóa được giải phóng và sử dụng, là quá trình cuối cùng đối với các sinh vật kỵ khí. Với sự hiện diện của oxy, axit pyruvic đi vào ty thể, tại đây, do một số phản ứng tuần tự, nó bị oxy hóa hoàn toàn trong điều kiện hiếu khí thành H2O và CO2 với quá trình phosphoryl hóa đồng thời ADP thành ATP. Trong trường hợp này, hai phân tử ATP được tạo ra bởi quá trình đường phân, hai phân tử được tạo ra bởi chu trình Krebs và 34 phân tử được tạo ra bởi chuỗi hô hấp. Hiệu suất thực của quá trình oxy hóa hoàn toàn một phân tử glucose thành H2O và CO2 là 38 phân tử.
Do đó, ở sinh vật hiếu khí, quá trình phân hủy cuối cùng của các chất hữu cơ được thực hiện bằng cách oxy hóa chúng bằng oxy trong khí quyển thành các chất vô cơ đơn giản: CO2 và H2O. Quá trình này diễn ra trên các cristae của ty thể. Trong trường hợp này, lượng năng lượng tự do tối đa được giải phóng, một phần đáng kể được dự trữ trong các phân tử ATP. Dễ dàng nhận thấy rằng quá trình oxy hóa hiếu khí cung cấp cho tế bào năng lượng tự do ở mức độ lớn nhất.
Kết quả của quá trình dị hóa là các phân tử ATP giàu năng lượng tích tụ trong tế bào, CO2 và nước dư thừa được thải ra môi trường bên ngoài.
Các phân tử đường không cần thiết cho quá trình hô hấp có thể được lưu trữ trong tế bào. Lipid dư thừa sẽ bị phân hủy, sau đó các sản phẩm phân hủy của chúng sẽ đi vào ty thể làm chất nền cho quá trình hô hấp hoặc được lưu trữ dưới dạng dự trữ trong tế bào chất dưới dạng các giọt chất béo. Protein được xây dựng từ các axit amin đi vào tế bào. Sự tổng hợp protein xảy ra ở các bào quan gọi là ribosome. Mỗi ribosome bao gồm hai hạt con - lớn và nhỏ: cả hai hạt con đều bao gồm các phân tử protein và phân tử RNA.
Ribosome thường được gắn vào một hệ thống màng đặc biệt bao gồm các bể chứa và túi - cái gọi là mạng lưới nội chất (ER); ở những tế bào sản xuất nhiều protein, mạng lưới nội chất thường phát triển rất tốt và được bao phủ bởi ribosome. Một số enzyme chỉ có hiệu quả nếu chúng được gắn vào màng. Hầu hết các enzyme tham gia vào quá trình tổng hợp lipid đều nằm ở đây. Vì vậy, mạng lưới nội chất giống như một loại bàn làm việc của tế bào.
Ngoài ra, ER chia tế bào chất thành các ngăn riêng biệt, tức là nó phân tách các quá trình hóa học khác nhau xảy ra đồng thời trong tế bào chất và do đó làm giảm khả năng các quá trình này ảnh hưởng lẫn nhau.
Sản phẩm được sản xuất bởi một tế bào nhất định thường được sử dụng bên ngoài tế bào. Trong những trường hợp như vậy, các protein được tổng hợp trên ribosome sẽ đi qua màng của lưới nội chất và được đóng gói thành các túi màng hình thành xung quanh chúng, sau đó được tách ra khỏi ER. Những mụn nước này dẹt và xếp chồng lên nhau, giống như những chiếc bánh kếp xếp chồng lên nhau, tạo thành một cấu trúc đặc trưng gọi là phức hợp Golgi hay bộ máy Golgi. Trong thời gian ở trong bộ máy Golgi, protein trải qua những thay đổi nhất định. Khi đến thời điểm chúng rời khỏi tế bào, các túi màng hợp nhất với màng tế bào và được làm trống, đổ nội dung của chúng ra ngoài, tức là sự bài tiết xảy ra bằng quá trình ngoại bào.
Bộ máy Golgi còn sản xuất lysosome - túi màng chứa enzym tiêu hóa. Tìm hiểu cách tế bào tạo ra, đóng gói và xuất khẩu một số protein nhất định cũng như cách tế bào “biết” loại protein nào nên giữ lại cho riêng mình, là một trong những nhánh hấp dẫn nhất của tế bào học hiện đại.
Màng của bất kỳ tế bào nào cũng liên tục chuyển động và thay đổi. Màng ER di chuyển chậm khắp tế bào. Các phần riêng lẻ của các màng này tách ra và tạo thành các túi, chúng tạm thời trở thành một phần của bộ máy Golgi, và sau đó, thông qua quá trình xuất bào, hợp nhất với màng tế bào.
Sau đó, vật liệu màng được đưa trở lại tế bào chất, nơi nó được sử dụng lại.