Вид, неговите критериуми. Популацијата е структурна единица на еден вид и елементарна единица на еволуција. Микроеволуција. Формирање на нови видови. Методи на спецификација. Зачувување на разновидноста на видовите како основа за одржливост на биосферата
Вид, неговите критериуми
Основачот на модерната таксономија, C. Linnaeus, го сметал видот како група на организми слични по морфолошки карактеристики кои слободно се вкрстуваат. Како што се развиваше биологијата, беа добиени докази дека разликите меѓу видовите се многу подлабоки и влијаат на хемискиот состав и концентрацијата на супстанциите во ткивата, насоката и брзината на хемиските реакции, природата и интензитетот на виталните процеси, бројот и обликот на хромозомите, т.е. видот е најмалата група на организми што ја одразува нивната блиска врска. Покрај тоа, видовите не постојат вечно - тие се појавуваат, се развиваат, раѓаат нови видови и исчезнуваат.
Погледни- ова е збирка на индивидуи кои се слични по структурата и карактеристиките на животните процеси, имаат заедничко потекло, слободно се вкрстуваат меѓу себе во природата и даваат плодно потомство.
Сите единки од ист вид имаат ист кариотип и заземаат одредена географска област во природата - област
Се нарекуваат знаци на сличност меѓу поединци од ист вид критериуми за тип. Бидејќи ниту еден од критериумите не е апсолутен, за правилно да се одреди типот потребно е да се користат множество критериуми.
Главните критериуми на еден вид се морфолошки, физиолошки, биохемиски, еколошки, географски, етолошки (бихејвиорални) и генетски.
- Морфолошки- збир на надворешни и внатрешни карактеристики на организми од ист вид. Иако некои видови имаат уникатни карактери, често е многу тешко да се разликуваат тесно сродни видови користејќи само морфолошки особини. Така, неодамна беа откриени голем број видови близнаци кои живеат на иста територија, на пример, домашниот глушец и глувчето Курганчик, па затоа е неприфатливо да се користат исклучиво морфолошки критериуми за одредување на видот.
- Физиолошки- сличноста на животните процеси во организмите, првенствено репродукцијата. Исто така, не е универзално, бидејќи некои видови се вкрстуваат во природата и даваат плодни потомци.
- Биохемиски- сличност на хемискиот состав и метаболичките процеси. И покрај фактот дека овие индикатори може значително да варираат кај различни индивидуи од ист вид, тие во моментов добиваат големо внимание, бидејќи структурните карактеристики и составот на биополимерите помагаат да се идентификуваат видовите дури и на молекуларно ниво и да се утврди степенот на нивната врска.
- Еколошки- разграничување на видовите според нивната припадност на одредени екосистеми и еколошки ниши што ги заземаат. Сепак, многу неповрзани видови зафаќаат слични еколошки ниши, така што овој критериум може да се користи за да се разликува еден вид само во комбинација со други карактеристики.
- Географски- постоење на популација на секој вид во одреден дел од биосферата - област која се разликува од областите на сите други видови. Поради фактот што за многу видови границите на нивните опсези се совпаѓаат, а исто така има и голем број космополитски видови чиј опсег опфаќа огромни простори, географскиот критериум исто така не може да послужи како маркерска карактеристика „видови“.
- Генетски- постојаност на карактеристиките на хромозомскиот сет - кариотип - и нуклеотидниот состав на ДНК кај индивидуи од ист вид. Поради фактот што нехомологните хромозоми не можат да се конјугираат за време на мејозата, потомците од вкрстување на поединци од различни видови со нееднаков збир на хромозоми или воопшто не се појавуваат или не се плодни. Ова создава репродуктивна изолација на видот, го одржува неговиот интегритет и ја обезбедува реалноста на постоењето во природата. Ова правило може да биде прекршено во случај на вкрстување на видови од слично потекло со ист кариотип или појава на различни мутации, но исклучокот само го потврдува општото правило, а видовите треба да се сметаат за стабилни генетски системи. Генетскиот критериум е главен во системот на критериуми за видови, но исто така не е исцрпен.
И покрај сложеноста на системот на критериуми, еден вид не може да биде претставен како група на организми кои се апсолутно идентични во сите погледи, односно клонови. Напротив, многу видови се карактеризираат со значителна разновидност дури и во надворешните карактеристики, бидејќи, на пример, некои популации на бубамари се карактеризираат со доминација на црвена боја, додека други се карактеризираат со доминација на црна боја.
Населението е структурна единица на еден вид и елементарна единица на еволуција
Тешко е да се замисли дека во реалноста, поединците од еден вид би биле рамномерно распоредени по површината на земјата во опсегот, бидејќи, на пример, езерската жаба живее главно во прилично ретки стоечки слатки водни тела и веројатно нема да се најде во полињата и шумите. Видовите во природата најчесто спаѓаат во посебни групи, во зависност од комбинацијата на услови погодни за нивните живеалишта - популации.
Популација- група индивидуи од ист вид, кои зафаќаат дел од нејзиниот опсег, слободно се вкрстуваат едни со други и релативно изолирани од други групи поединци од истиот вид повеќе или помалку долго време.
Населението може да се одвои не само просторно, тие дури можат да живеат на иста територија, туку имаат разлики во преференциите за храна, времето на репродукција итн.
Така, вид е збир на популации на индивидуи кои имаат голем број заеднички морфолошки, физиолошки, биохемиски карактеристики и типови на односи со животната средина, кои населуваат одредена област, а исто така се способни да се вкрстуваат едни со други за да формираат плодни потомци, но речиси или воопшто не се вкрстуваат со други групи поединци од ист вид.
Во рамките на видовите со големи опсези кои покриваат територии со различни услови за живеење, тие понекогаш се разликуваат подвидови- големи популации или групи на соседни популации кои имаат постојани морфолошки разлики од другите популации.
Населението не е случајно расфрлано низ површината на земјата, тие се врзани за одредени области. Севкупноста на сите фактори од нежива природа неопходни за престој на поединци од даден вид се нарекува живеалиште. Сепак, овие фактори сами по себе можеби не се доволни за популацијата да ја заземе оваа област, бидејќи таа сепак мора да биде вклучена во блиска интеракција со популации од други видови, односно да зазема одредено место во заедницата на живи организми - еколошка ниша. Така, австралиската коала торбарска мечка, сите други работи се еднакви, не може да постои без нејзиниот главен извор на храна - еукалиптус.
Популациите од различни видови кои формираат нераскинливо единство во истите живеалишта обично обезбедуваат повеќе или помалку затворен циклус на супстанции и се елементарни еколошки системи (екосистеми) - биогеоценози.
И покрај сите нивни барања за условите на животната средина, популациите од ист вид се хетерогени по површина, број, густина и просторна дистрибуција на индивидуи, често формирајќи помали групи (семејства, стада, стада итн.), пол, возраст, генски фонд итн. Затоа, се разликуваат нивната големина, возраст, пол, просторна, генетска, етолошка и други структури, како и динамика.
Важни карактеристики на една популација се генски базен- збир на гени карактеристични за поединци од дадена популација или вид, како и зачестеноста на одредени алели и генотипови. Различни популации од ист вид првично имаат различни генски базени, бидејќи новите територии се колонизирани од поединци со случајни наместо специјално избрани гени. Под влијание на внатрешни и надворешни фактори, генскиот базен претрпува уште позначајни промени: тој се збогатува поради појавата на мутации и нова комбинација на особини и се исцрпува како резултат на губење на поединечни алели за време на смртта или миграцијата на одреден број на поединци.
Новите особини и нивните комбинации можат да бидат корисни, неутрални или штетни, затоа, само поединци приспособени на дадените услови на животната средина преживуваат и успешно се размножуваат во популацијата. Меѓутоа, на две различни точки на површината на земјата, условите на животната средина никогаш не се целосно идентични, затоа насоката на промените дури и кај две соседни популации може да биде сосема спротивна или тие ќе се појават со различна брзина. Резултатот од промените во генскиот базен е дивергенцијата на популациите според морфолошки, физиолошки, биохемиски и други карактеристики. Ако популациите се исто така изолирани едни од други, тогаш тие можат да доведат до појава на нови видови.
Така, појавата на какви било пречки при вкрстувањето на индивидуи од различни популации од ист вид, на пример, поради формирање на планински масиви, промени во речните корита, разлики во времето на репродукција итн., доведува до фактот дека популациите постепено стекнуваат се повеќе и повеќе разлики и на крајот стануваат различни видови. Некое време, на границите на овие популации, се случува вкрстување на поединци и се појавуваат хибриди, но со текот на времето, овие контакти исчезнуваат, т.е. популациите од отворените генетски системи стануваат затворени.
И покрај фактот дека поединците се првенствено изложени на фактори на животната средина, промените во генетскиот состав на еден организам се незначителни и, во најдобар случај, ќе се појават само кај неговите потомци. Подвидовите, видовите и поголемите таксони исто така не се погодни за улогата на елементарни единици на еволуцијата, бидејќи тие не се разликуваат по морфолошка, физиолошка, биохемиска, еколошка, географска и генетска единство, додека популациите се најмалите структурни единици на еден вид, кои се акумулираат различни случајни промени, од кои најлошите ќе бидат елиминирани, одговараат на оваа состојба и се елементарни единици на еволуцијата.
Микроеволуција
Промената на генетската структура на популациите не секогаш води до формирање на нов вид, туку може само да ја подобри адаптацијата на популацијата на специфичните услови на животната средина, меѓутоа, видовите не се вечни и непроменливи - тие се способни да се развиваат. Овој процес на неповратни историски промени кај живите суштества се нарекува еволуција. Примарните еволутивни трансформации се случуваат во рамките на еден вид на ниво на популација. Тие се засноваат, пред сè, на процесот на мутација и природната селекција, што доведува до промени во генскиот базен на популациите и видот како целина, па дури и до формирање на нови видови. Множеството на овие елементарни еволутивни настани се нарекува микроеволуција.
Популациите се карактеризираат со огромна генетска разновидност, која често не се изразува фенотипски. Генетската разновидност се јавува поради спонтана мутагенеза, која се јавува континуирано. Повеќето мутации се неповолни за организмот и ја намалуваат одржливоста на популацијата како целина, но доколку се рецесивни, кај хетерозиготите може да опстојуваат долго време. Некои мутации кои немаат адаптивна вредност во дадени услови на постоење се способни да добијат таква вредност во иднина или кога ќе се развијат нови еколошки ниши, со што се создава резерва на наследна варијабилност.
Микроеволутивните процеси се значително под влијание на флуктуациите во бројот на поединци во популациите, миграциите и катастрофите, како и изолацијата на популациите и видовите.
Новиот вид е среден резултат на еволуцијата, но во никој случај негов резултат, бидејќи микроеволуцијата не застанува тука - таа продолжува понатаму. Новите видови кои се појавуваат, во случај на успешна комбинација на карактеристики, населуваат нови живеалишта и, пак, создаваат нови видови. Таквите групи на тесно сродни видови се обединети во родови, семејства итн. За разлика од макроеволуција,микроеволуцијата се одвива во многу пократок временски период, додека за првата потребни се десетици и стотици илјади и милиони години, како што е човечката еволуција.
Како резултат на микроеволуцијата, се формира целата разновидност на видови живи организми кои некогаш постоеле и сега живеат на Земјата.
Во исто време, еволуцијата е неповратна, а видовите кои веќе исчезнале никогаш повеќе не се појавуваат. Видовите што се појавуваат консолидираат се што е постигнато во процесот на еволуција, но тоа не гарантира дека во иднина нема да се појават нови видови кои ќе имаат понапредни прилагодувања на условите на животната средина.
Формирање на нови видови
Во широка смисла, формирањето на нови видови се подразбира не само како отцепување на нов вид од главното стебло или дезинтеграција на родителскиот вид на неколку видови ќерки, туку и општ развој на видот како интегрален систем. , што доведува до значителни промени во неговата морфоструктурна организација. Сепак, почесто отколку не специјацијасе смета за процес на формирање на нови видови преку разгранување на „семејното стебло“ на видот.
Фундаментално решение за проблемот со спецификацијата беше предложено од Чарлс Дарвин. Според неговата теорија, ширењето на индивидуи од ист вид доведува до формирање на популации кои поради разликите во условите на околината се принудени да се прилагодат на нив. Ова, пак, повлекува интензивирање на интраспецифичната борба за егзистенција, насочена од природната селекција. Во моментов, се верува дека борбата за егзистенција воопшто не е задолжителен фактор во спецификацијата, напротив, притисокот на селекција кај одреден број популации може да се намали. Разликите во условите за живеење придонесуваат за појава на нееднакви адаптивни промени во популациите на еден вид, чија последица е дивергенција на карактеристиките и својствата на популациите - дивергенција.
Сепак, акумулацијата на разлики, дури и на генетско ниво, во никој случај не е доволна за појава на нов вид. Сè додека популациите што се разликуваат по некои карактеристики не само што се во контакт, туку се способни и за вкрстување со формирање на плодно потомство, тие припаѓаат на истиот вид. Само неможноста за проток на гени од една група на поединци во друга, дури и во случај на уништување на бариерите што ги одвојуваат, т.е. преминување, значи завршување на најкомплексниот еволутивен процес на формирање на нов вид.
Специјацијата е продолжение на микроеволутивните процеси. Постои гледна точка дека специјацијата не може да се сведе на микроеволуција таа претставува квалитативна фаза на еволуција и се спроведува благодарение на други механизми.
Методи на специјација
Постојат два главни методи на специјација: алопатриски и симпатриски.
Алопатричен, или географска спецификацијае последица на просторното одвојување на населението со физички бариери (планински венци, мориња и реки) поради нивно појавување или распрснување во нови живеалишта (географска изолација). Бидејќи во овој случај генскиот базен на одвоената популација значително се разликува од мајчината, а условите во неговото живеалиште нема да се совпаѓаат со првобитните, со текот на времето тоа ќе доведе до дивергенција и формирање на нов вид. Еклатантен пример за географска специјација е разновидноста на видовите сики откриени од Чарлс Дарвин за време на неговото патување со бродот Бигл на островите Галапагос во близина на брегот на Еквадор. Очигледно, поединечни единки од единствената сипка што го населува јужноамериканскиот континент некако завршиле на островите и, поради разликите во условите (првенствено достапноста на храна) и географската изолација, постепено еволуирале, формирајќи група сродни видови.
Во сржта симпатична, или биолошка специјацијалежи некоја форма на репродуктивна изолација, со нови видови кои се појавуваат во опсегот на оригиналниот вид. Предуслов за симпатична специјација е брзата изолација на добиените форми. Ова е побрз процес од алопатриската специјација, а новите форми се слични на првобитните предци.
Симпатичната специјација може да биде предизвикана од брзи промени во составот на хромозомите (полиплоидизација) или хромозомски преуредувања. Понекогаш се појавуваат нови видови како резултат на хибридизација на два оригинални видови, како што е, на пример, домашната слива, која е хибрид на слива и цреша. Во некои случаи, симпатичната специјација е поврзана со поделба на еколошки ниши во популации од ист вид во еден опсег или сезонска изолација - дивергенција во времето на размножување кај растенијата (различни видови борови во Калифорнија произведуваат прашина во февруари и април) и во времето на размножување кај животните.
Од целата разновидност на новопојавени видови, само неколку, најприлагодените, можат да постојат долго време и да доведат до појава на нови видови. Причините за смртта на повеќето видови сè уште се непознати, најверојатно тоа се должи на ненадејните климатски промени, геолошките процеси и нивното поместување од страна на поприлагодени организми. Во моментов, една од причините за смртта на значителен број видови е човекот, кој ги истребува најголемите животни и најубавите растенија, а ако во 17 век овој процес започнал дури со истребување на последната рунда, тогаш во 21 век повеќе од 10 видови исчезнуваат секој час.
Зачувување на разновидноста на видовите како основа за одржливост на биосферата
И покрај фактот дека, според различни проценки, планетата е дом на 5-10 милиони видови организми кои сè уште не се опишани, никогаш нема да знаеме за постоењето на повеќето од нив, бидејќи околу 50 видови исчезнуваат од лицето на Земјата секој час. Исчезнувањето на живите организми во моментов не е нужно поврзано со нивното физичко истребување, почесто се должи на уништувањето на нивните природни живеалишта како резултат на човековата активност. Смртта на поединечен вид веројатно нема да доведе до фатални последици за биосферата, но одамна е утврдено дека исчезнувањето на еден растителен вид повлекува смрт на 10-12 животински видови, а тоа веќе претставува закана и за постоењето. на поединечни биогеоценози и на глобалниот екосистем воопшто.
Тажните факти акумулирани во текот на претходните децении ја принудија Меѓународната унија за зачувување на природата и природните ресурси (IUCN) да започне со собирање информации за ретки и загрозени видови растенија и животни во 1949 година. Во 1966 година, IUCN ја објави првата Црвена книга на факти.
Црвена книгае официјален документ кој содржи редовно ажурирани податоци за статусот и распространетоста на ретки и загрозени видови растенија, животни и габи.
Со овој документ се усвои петстепена скала за статус на заштитен вид, при што првиот степен на заштита вклучува видови чиј спас е невозможен без посебни мерки, а петтиот - обновени видови, чија состојба, благодарение на преземените мерки, го прави не предизвикуваат загриженост, но тие сè уште не се предмет на индустриска употреба. Развојот на таков обем овозможува да се насочат приоритетните напори за зачувување конкретно на најретките видови, како што се тигрите Амур.
Покрај меѓународната верзија на Црвената книга, постојат и национални и регионални верзии. Во СССР, Црвената книга е основана во 1974 година, а во Руската Федерација, постапката за нејзино одржување е регулирана со Федералните закони „За заштита на животната средина“, „За дивиот свет“ и Уредбата на Владата на Руската Федерација „ За Црвената книга на Руската Федерација“. Денес, 610 видови растенија, 247 видови животни, 42 видови лишаи и 24 видови габи се наведени во Црвената книга на Руската Федерација. Популациите на некои од нив, некогаш загрозени (европски дабар, бизон), веќе се доста успешно обновени.
Следниве животински видови се заштитени во Русија: руски мускрат, тарбаган (монголски мрмот), поларна мечка, кавкаски европски визон, морска видра, манул, амур тигар, леопард, снежен леопард, морски лав, морж, фоки, делфини, китови, Пржевалски коњ, див газ, розов пеликан, обичен фламинго, црн штрк, мал лебед, степски орел, златен орел, црн кран, сибирски кран, бистар, орел був, бел галеб, медитеранска желка, јапонска змија, вајпер, џунгла жаба, касписки леле , сите видови риби есетра, езерски лосос, елен бубачки, извонреден бумбар, обичен Аполон, рак богомолка, обична бисерна школка итн.
Растенијата на Црвената книга на Руската Федерација вклучуваат 7 видови кокичиња, некои видови пелин, вистински женшен, 7 видови сини ѕвончиња, назабени дабови, шила, 11 видови ирис, руска леска тетреб, лале Шренк, орев лотос, женска влечка, тенколистен божур, пердув трева, јаглика Јулија, ливадско лумбаго (трева за спиење), беладона беладона, бор Пицунда, тис, кинеска штитна трева, езерска трева, мек сфагнум, кадрава филета, итн.
Ретките печурки се претставени со летниот тартуф, или рускиот црн тартуф, лакираната габа од габа итн.
Заштитата на ретките видови во најголем број случаи се поврзува со забрана за нивно уништување, нивно зачувување во вештачки создадени живеалишта (зоолошки градини), заштита на нивните живеалишта и создавање на нискотемпературни генетски банки.
Најефикасна мерка за заштита на ретките видови е зачувувањето на нивните живеалишта, што се постигнува со организирање мрежа на специјално заштитени подрачја кои, во согласност со Федералниот закон „За посебно заштитени природни подрачја“ (1995), имаат меѓународно , федерално, регионално или локално значење. Тие вклучуваат државни природни резервати, национални паркови, природни паркови, државни природни резервати, природни споменици, дендролошки паркови, ботанички градини итн.
Државен природен резерват- ова е специјално заштитен природен комплекс (земја, водни тела, подлога, флора и фауна) целосно повлечен од економска употреба, кој има еколошко, научно, еколошко и едукативно значење како пример за природна средина, типични или ретки пејсажи, места. каде што е зачуван генетскиот фонд на растенијата и животинскиот свет.
Резервите кои се дел од меѓународниот систем на биосферни резервати кои вршат глобален мониторинг на животната средина имаат статус државни природни биосферни резервати. Резерватот е еколошка, истражувачка и еколошка образовна институција насочена кон зачувување и проучување на природниот тек на природните процеси и појави, генетскиот фонд на флората и фауната, поединечните видови и заедници на растенија и животни, типични и единствени еколошки системи.
Во моментов, во Русија има околу 100 државни природни резервати, од кои 19 имаат статус на биосфера, вклучувајќи ги Бајкалски, Баргузински, Кавкаски, Кедроваја Пад, Кроноцки, Приоско-Терасни итн.
За разлика од природните резервати, териториите (водните области) национални парковивклучуваат природни комплекси и објекти кои имаат посебни еколошки, историски и естетски вредности, а се наменети за употреба за еколошки, образовни, научни и културни цели и за регулиран туризам. Овој статус го имаат 39 специјално заштитени природни подрачја, вклучително и националните паркови Транс-Бајкал и Сочи, како и националните паркови „Куронски спит“, „Руски север“, „Шушенски Бор“ итн.
Природни парковисе еколошки рекреативни институции под јурисдикција на конститутивните субјекти на Руската Федерација, чии територии (водни области) вклучуваат природни комплекси и објекти кои имаат значителни еколошки и естетски вредности, а се наменети за употреба за еколошки, едукативни и рекреативни цели.
Државни природни резерватисе територии (водни подрачја) кои се од особено значење за зачувување или обновување на природните комплекси или нивни компоненти и одржување на еколошката рамнотежа.
Развој на еволутивни идеи. Значењето на еволутивната теорија на Чарлс Дарвин. Меѓусебна поврзаност на движечките сили на еволуцијата. Форми на природна селекција, видови на борба за егзистенција. Синтетичка теорија на еволуција. Елементарни фактори на еволуцијата. Истражување на С.С. Четвериков. Улогата на еволутивната теорија во формирањето на модерната природонаучна слика на светот
Развој на еволутивни идеи
Сите теории за потеклото и развојот на органскиот свет можат да се сведат на три главни насоки: креационизам, трансформизам и еволуционизам. Креационизаме концептот на постојаност на видовите, земајќи ја предвид различноста на органскиот свет како резултат на неговото создавање од Бога. Оваа насока е формирана како резултат на воспоставувањето на доминацијата на христијанската црква во Европа, врз основа на библиски текстови. Истакнати претставници на креационизмот биле C. Linnaeus и J. Cuvier.
„Принцот на ботаничарите“ C. Linnaeus, кој откри и опиша стотици нови растителни видови и го создаде нивниот прв хармоничен систем, сепак тврдеше дека вкупниот број на видови организми останал непроменет од создавањето на Земјата, т.е. не само што не се појавуваат повторно, туку и не исчезнуваат. Дури кон крајот на својот живот дошол до заклучок дека родовите се Божјо дело, додека видовите можат да се развиваат поради приспособување на локалните услови.
Придонесот на извонредниот француски зоолог J. Cuvier (1769–1832) во биологијата се заснова на бројни податоци од палеонтологијата, компаративната анатомија и физиологијата доктрина за корелации- односи меѓу делови од телото. Благодарение на ова, стана возможно да се реконструира надворешниот изглед на животното во одделни делови. Сепак, во процесот на палеонтолошко истражување, J. Cuvier не можеше а да не обрне внимание и на очигледното изобилство на фосилни форми и на острите промени во животинските групи во текот на геолошката историја. Овие податоци послужија како почетна точка за формулирање теории за катастрофа, според кој сите или речиси сите организми на Земјата биле постојано убивани како резултат на периодични природни катастрофи, а потоа планетата била повторно населена со видови кои ја преживеале катастрофата. Следбениците на J. Cuvier изброиле до 27 такви катастрофи во историјата на Земјата. Размислувањата за еволуцијата се чинеше дека Џ. Кувиер е разведен од реалноста.
Противречностите во почетните премиси на креационизмот, кои стануваа се поочигледни како што се акумулираа научните факти, послужија како појдовна точка за формирање на друг систем на погледи - трансформизам, согледувајќи го вистинското постоење на видовите и нивниот историски развој. Претставниците на овој тренд - Џ. Буфон, И. Корените на трансформизмот може да се најдат во делата на античките грчки и средновековните филозофи кои ги препознале историските промени во органскиот свет. Така, Аристотел ја изразил идејата за единството на природата и постепениот премин од тела на нежива природа кон растенијата, а од нив кон животните - „скалата на природата“. Тој сметал дека главната причина за промените во живите организми е нивната внатрешна желба за совршенство.
Францускиот натуралист Ј. Буфон (1707-1788), чие главно животно дело беше 36-томната Природна историја, спротивно на ставовите на креационистите, го прошири опсегот на историјата на Земјата на 80-90 илјади години. Во исто време, тој го истакна единството на флората и фауната, како и можноста за промени во сродните организми под влијание на факторите на животната средина како резултат на припитомување и хибридизација.
Англискиот лекар, филозоф и поет Е. Дарвин (1731–1802), дедото на Чарлс Дарвин, верувал дека историјата на органскиот свет датира милиони години наназад, а различноста на животинскиот свет е резултат на мешавина од неколку „ природни“ групи, влијанието на надворешната средина, вежбањето и недостатокот на органи за вежбање и други фактори.
E. Geoffroy Saint-Hilaire (1772–1844) сметал дека единството на структурниот план на групи животни е еден од главните докази за развојот на живиот свет. Сепак, за разлика од неговите претходници, тој беше склон да верува дека промените во видовите се предизвикани од влијанието на факторите на животната средина не на возрасни поединци, туку на ембриони.
И покрај фактот што во дискусијата што се разгоре во 1831 година помеѓу Џ. Кувиер и Е. Жофрој Сен-Илер во форма на серија извештаи на Академијата на науките, јасна предност остана на страната на првиот, таа беше трансформизмот кој стана претходник на еволуционизмот. еволуционизам(теорија на еволуција, еволутивна доктрина) е систем на гледишта што го препознава развојот на природата според одредени закони. Тоа е теоретскиот врв на биологијата, кој ни овозможува да ја објасниме различноста и сложеноста на живите системи што ги набљудуваме. Меѓутоа, поради фактот што еволуциското учење опишува феномени кои се тешки за набљудување, се соочува со значителни тешкотии. Понекогаш теоријата на еволуцијата се нарекува „дарвинизам“ и се поистоветува со учењата на Чарлс Дарвин, што е фундаментално неточно, бидејќи, иако теоријата на Чарлс Дарвин даде непроценлив придонес во развојот не само на доктрината за еволуција, туку и на биологијата во општи (како и многу други науки), основите на еволутивната теорија ги поставија други научници, таа продолжува да се развива до денес, а „дарвинизмот“ во многу аспекти има само историско значење.
Креатор на првата еволутивна теорија - Ламаркизам - беше францускиот натуралист Ј.Б. Ламарк (1744–1829). Тој сметал дека движечката сила на еволуцијата е внатрешната желба на организмите за совршенство ( закон за градација), сепак, приспособувањето кон условите на животната средина ги принудува да отстапат од оваа главна линија. Во исто време, органите што животното интензивно ги користи во процесот на живот се развиваат, а оние што се непотребни за него, напротив, се ослабени, па дури и може да исчезнат ( закон за вежбање и невежба на органи). Карактеристиките стекнати во текот на животот се фиксираат и се пренесуваат на потомците. Така, тој го објасни присуството на мембрани меѓу прстите на водните птици со обидите на нивните предци да се движат во водната средина, а долгиот врат на жирафите, според Ламарк, е последица на тоа што нивните предци се обиделе да добијат лисја. од врвовите на дрвјата.
Недостатоците на Ламаркизмот беа теоретската природа на многу конструкции, како и претпоставката за интервенција на Создателот во еволуцијата. Во процесот на развој на биологијата, стана јасно дека поединечните промени што ги стекнале организмите во текот на животот, во најголем дел, спаѓаат во границите на фенотипската варијабилност, а нивното пренесување е практично невозможно. На пример, германскиот зоолог и теоретичар на еволуцијата А. Вајсман (1834–1914) ги отсекувал опашките на глувците многу генерации и секогаш добивал само опашести глодари во нивните потомци. Теоријата на Ј.Б.Ламарк не беше прифатена од неговите современици, но на крајот на векот ја формираше основата на т.н. неоламаркизам.
Значењето на еволутивната теорија на Чарлс Дарвин
Предусловите за создавање на најпознатата еволутивна теорија на Чарлс Дарвин, или Дарвинизам, беа објавувањето во 1778 година на делото на англискиот економист Т. Малтус „Трактат за населението“, делото на геологот Чарлс Лајел, формулацијата на клеточната теорија, успехот на селекцијата во Англија и сопствените набљудувања на Чарлс (1809–1882), земени за време на неговите студии во Кембриџ, за време на експедицијата како натуралист на Бигл и при нејзиното завршување.
Така, Т. Малтус тврдеше дека популацијата на Земјата експоненцијално се зголемува, што значително ја надминува способноста на планетата да и обезбеди храна и доведува до смрт на некои од потомците. Паралелите направени од Чарлс Дарвин и неговиот коавтор А. Валас (1823–1913) покажаа дека во природата, поединците се размножуваат со многу голема брзина, но големината на популацијата останува релативно константна. Истражувањето на англискиот геолог C. Lyell овозможило да се утврди дека површината на Земјата не била секогаш иста како што е сега, а нејзините промени биле предизвикани од влијанието на водата, ветерот, вулканските ерупции и активноста на живиот свет. организми. Дури и во студентските години, самиот Чарлс Дарвин бил погоден од екстремниот степен на варијабилност на бубачките, а за време на неговите патувања од сличноста на флората и фауната на континентална Јужна Америка и блиските острови Галапагос, а во исто време и од значајните разновидност на видови, како што се сибите и желките. Покрај тоа, на експедицијата тој можеше да ги набљудува скелетите на џиновските изумрени цицачи, слични на модерните армадилоси и мрзливи, што значително го разниша неговото верување во создавањето видови.
Основните принципи на теоријата на еволуцијата беа изразени од Чарлс Дарвин во 1859 година на состанокот на Кралското друштво на Лондон, а потоа беа развиени во книгите „Потекло на видовите со природна селекција или зачувување на омилените раси во борбата за Живот“ (1859), „Промени во домашните животни и култивирани растенија“ (1868), „Потеклото на човекот и сексуалната селекција“ (1871), „Изразувањето на емоциите кај човекот и животните“ (1872) итн.
Суштината развиена од Чарлс Дарвин концепти на еволуцијаможе да се сведе на низа одредби кои произлегуваат една од друга, кои имаат соодветни доказ:
- Поединците што ја сочинуваат секоја популација произведуваат многу повеќе потомци отколку што е потребно за одржување на големината на популацијата.
- Поради тоа што животните ресурси за секаков вид живи организми се ограничени, неизбежно се јавува меѓу нив борба за егзистенција. Чарлс Дарвин направи разлика помеѓу интраспецифична и меѓуспецифична борба, како и борба со фактори на животната средина. Истовремено, тој исто така посочи дека не се зборува само за борбата на одредена индивидуа за егзистенција, туку и за оставање потомство.
- Последица на борбата за егзистенција е природна селекција- доминантно преживување и размножување на организмите кои случајно се покажаа дека се најприлагодени на дадените услови на постоење. Природната селекција на многу начини е слична на вештачката селекција, која луѓето ја користеле уште од античко време за да одгледуваат нови сорти на растенија и раси на домашни животни. Со избирање на поединци кои имаат некоја посакувана особина, човекот ги зачувува овие особини преку вештачко размножување преку селективно размножување или опрашување. Посебна форма на природна селекција е сексуалната селекција за особини кои обично немаат директно адаптивно значење (долги пердуви, огромни рогови итн.), но придонесуваат за репродуктивен успех бидејќи ја прават поединецот попривлечен за спротивниот пол или пострашен за ривали од ист пол.
- Материјалот за еволуција се разликите помеѓу организмите кои се јавуваат како резултат на нивната варијабилност. Чарлс Дарвин направи разлика помеѓу неодредена и определена варијабилност. Одредени(групна) варијабилност се манифестира кај сите единки на еден вид подеднакво под влијание на одреден фактор и исчезнува кај потомците кога ќе престане дејството на овој фактор. Неизвесна(индивидуална) варијабилност се промени што се случуваат кај секој поединец, без оглед на флуктуациите во вредностите на факторите на животната средина и се пренесуваат на потомците. Таквата варијабилност нема адаптивна природа. Последователно, се покажа дека одредена варијабилност е ненаследна, а неодредената варијабилност е наследна.
- Природната селекција на крајот води до дивергенција во карактеристиките на поединечните изолирани сорти - дивергенција и, на крајот, до формирање на нови видови.
Теоријата за еволуција на Чарлс Дарвин не само што го постулираше процесот на појава и развој на видовите, туку го откри и самиот механизам на еволуција, кој се заснова на принципот на природна селекција. Дарвинизмот исто така ја негираше програмираната природа на еволуцијата и ја постулираше нејзината континуирана природа.
Во исто време, еволутивната теорија на Чарлс Дарвин не можеше да одговори на голем број прашања, на пример, за природата на генетскиот материјал и неговите својства, суштината на наследната и ненаследната варијабилност и нивната еволутивна улога. Ова доведе до криза на дарвинизмот и појава на нови теории: неоламаркизам, салтационизам, концепт на номогенеза итн. Неоламаркизамсе заснова на позицијата на теоријата на J. B. Lamarck за наследување на стекнатите карактеристики. Салтационизаме систем на гледишта за процесот на еволуција како нагли промени што доведуваат до брзо појавување на нови видови, родови и поголеми систематски групи. Концепт номогенезаја постулира програмираната насока на еволуцијата и развојот на различни карактеристики врз основа на внатрешните закони. Само синтезата на дарвинизмот и генетиката во 20-30-тите години на дваесеттиот век можеше да ги надмине противречностите што неизбежно се појавија при објаснувањето на голем број факти.
Меѓусебна поврзаност на движечките сили на еволуцијата
Еволуцијата не може да се поврзе со дејството на ниту еден фактор, бидејќи самите мутации се случајни и ненасочени промени и не можат да обезбедат адаптација на поединците на факторите на околината, додека природната селекција веќе ги сортира овие промени. Исто така, самата селекција не може да биде единствениот фактор во еволуцијата, бидејќи селекцијата бара соодветен материјал обезбеден од мутации.
Сепак, може да се забележи дека процесот на мутација и генскиот тек создаваат варијации, додека природната селекција и генетскиот нанос ја средуваат оваа варијација. Ова значи дека факторите кои создаваат варијабилност го иницираат процесот на микроеволуција, а оние кои ја сортираат варијабилноста го продолжуваат, што доведува до воспоставување на нови фреквенции на варијанти. Така, еволутивната промена во популацијата може да се гледа како резултат на спротивставените сили што создаваат и сортираат генотипски варијации.
Пример за интеракцијата помеѓу процесот на мутација и селекцијата е хемофилија кај луѓето. Хемофилија е болест предизвикана од намалено згрутчување на крвта. Претходно доведе до смрт во предрепродуктивниот период, бидејќи секое оштетување на мекото ткиво потенцијално може да доведе до значителна загуба на крв. Оваа болест е предизвикана од рецесивна мутација на половиот ген H (Xh). Жените многу ретко страдаат од хемофилија, тие се почесто хетерозиготни носители, но нивните синови можат да ја наследат болеста. Теоретски, во текот на неколку генерации, таквите мажи умираат пред пубертетот и постепено овој алел треба да исчезне од популацијата, но зачестеноста на појавата на оваа болест не се намалува поради повторените мутации во овој локус, како што се случи кај кралицата Викторија, која ја пренел болеста на три генерации од кралските куќи на Европа. Постојаната фреквенција на оваа болест укажува на рамнотежа помеѓу процесот на мутација и притисокот на селекција.
Форми на природна селекција, видови на борба за егзистенција
Природна селекцијаТие го нарекуваат селективно преживување и оставање потомство од страна на најпогодни поединци и смрт на најмалку способни.
Суштината на природната селекција во теоријата на еволуцијата лежи во диференцираното (неслучајно) зачувување на одредени генотипови во една популација и нивното селективно учество во преносот на гените на следната генерација. Покрај тоа, тоа не влијае на една особина (или ген), туку на целиот фенотип, кој се формира како резултат на интеракцијата на генотипот со факторите на животната средина. Природната селекција ќе биде од различна природа во различни услови на животната средина. Во моментов, постојат неколку форми на природна селекција: стабилизирање, возење и кинење.

Стабилизирање на селекцијае насочена кон консолидирање на тесна норма на реакција, која се покажа како најповолна во дадените услови на постоење. Тоа е типично за оние случаи кога фенотипските карактеристики се оптимални за непроменливи услови на животната средина. Впечатлив пример за дејството на стабилизирање на селекција е зачувувањето на релативно константна телесна температура на топлокрвните животни. Оваа форма на селекција беше детално проучена од извонредниот руски зоолог I. I. Shmalgauzen.
Избор на возењесе јавува како одговор на промените во условите на животната средина, како резултат на што се зачувани мутации кои отстапуваат од просечната вредност на особината, додека претходно доминантната форма е уништена бидејќи не ги исполнува доволно новите услови на постоење. На пример, во Англија, како резултат на загадувањето на воздухот од индустриски емисии, станаа широко распространети пеперутките од бреза молци, претходно невидени на многу места, со темно обоени крилја, кои беа помалку видливи за птиците на позадината на саѓи стебла од бреза. Изборот на возење не придонесува за целосно уништување на формата против која дејствува, бидејќи како резултат на мерките преземени од владата и еколошките организации, состојбата со загадувањето на воздухот е нагло подобрена, а бојата на крилата на пеперутката се врати на неговата оригинална верзија.
Кинење, или нарушувачки изборго фаворизира зачувувањето на екстремните варијанти на некоја особина и ги отстранува средните, бидејќи, на пример, како резултат на употребата на пестициди, се појавуваат групи на поединци инсекти отпорни на него. Во својот механизам, нарушувачката селекција е спротивна на стабилизирачката селекција. Преку оваа форма на селекција, кај популацијата се јавуваат неколку остро разграничени фенотипови. Овој феномен се нарекува полиморфизам. Појавата на репродуктивна изолација помеѓу различни форми може да доведе до специјација.
Понекогаш тие исто така се разгледуваат одделно дестабилизирачка селекција, кој зачувува мутации што доведуваат до широк спектар на која било карактеристика, на пример, бојата и структурата на лушпите на некои мекотели кои живеат во хетерогени микроуслови на карпестиот сурфање на морето. Оваа форма на селекција ја откри Д.К.Белјаев додека го проучуваше припитомувањето на животните.
Во природата ниту еден од облиците на природна селекција не постои во чиста форма, туку напротив, постојат различни комбинации од нив, а како што се менуваат условите на околината, прво доаѓа до израз едната или другата. Така, по завршувањето на промените во околината, селекцијата на возење се заменува со стабилизирачка селекција, која оптимизира група на поединци во нови услови на постоење.
Природната селекција се јавува на различни нивоа и затоа се разликуваат и индивидуална, групна и сексуална селекција. Индивидуаленселекцијата ги елиминира помалку прилагодените индивидуи од учество во репродукцијата, додека групната селекција е насочена кон зачувување на особина која е корисна не за поединецот, туку за групата како целина. Под притисок групаселекцијата може целосно да избрише цели популации, видови и поголеми групи организми без да остави потомство. За разлика од индивидуалната селекција, групната селекција ја намалува различноста на формите во природата.
Сексуална селекцијаспроведена во рамките на еден пол. Го промовира развојот на особини кои обезбедуваат успех во оставањето на најголемото потомство. Благодарение на оваа форма на природна селекција, се разви сексуален диморфизам, изразен во големината и бојата на опашката на паунот, роговите на елен итн.
Резултатот е природната селекција борба за егзистенцијаврз основа на наследна варијабилност. Борбата за егзистенција се подразбира како целокупен збир на односи меѓу поединците од сопствениот и другите видови, како и со абиотските фактори на животната средина. Овие односи го одредуваат успехот или неуспехот на одредена индивидуа да преживее и да произведе потомство. Причината за борбата за егзистенција е појавата на вишок на поединци во однос на расположливите ресурси. Покрај конкуренцијата, овие односи треба да вклучуваат и взаемна помош, што ги зголемува шансите за преживување на поединците.
Интеракција со фактори на животната срединаможе да доведе и до смрт на огромното мнозинство на поединци, на пример, кај инсекти, од кои само мал дел ја преживуваат зимата.
Синтетичка теорија на еволуција
Успесите на генетиката на почетокот на дваесеттиот век, на пример, откривањето на мутациите, сугерираат дека наследни промени во фенотипот на организмите се случуваат ненадејно и не се формираат во текот на долг временски период, како што претпоставува еволутивната теорија на Чарлс Дарвин. Сепак, понатамошните истражувања во областа на популациската генетика доведоа до формулација во 20-50-тите години на дваесеттиот век на нов систем на еволутивни погледи - синтетичка теорија на еволуција. Значаен придонес во неговото создавање направија научници од различни земји: советските научници С. С. Четвериков, И. Симпсон и зоологот Е. Мајр.
Основни одредби на синтетичката теорија на еволуцијата:
- Елементарниот материјал на еволуцијата е наследна варијабилност (мутациона и комбинирана) кај поединци од една популација.
- Елементарната единица на еволуцијата е населението во кое се случуваат сите еволутивни промени.
- Елементарен еволутивен феномен е промена во генетската структура на популацијата.
- Елементарните фактори на еволуцијата - генетски нанос, бранови на живот, генски тек - се ненасочени, по случаен карактер.
- Единствениот насочувачки фактор во еволуцијата е природната селекција, која е креативна по природа. Природната селекција може да биде стабилизирачка, движечка и нарушувачка.
- Еволуцијата е дивергентна по природа, односно, еден таксон може да доведе до неколку нови таксони, додека секој вид има само еден предок (вид, популација).
- Еволуцијата е постепена и континуирана. Специјацијата како фаза од еволутивниот процес е последователна замена на една популација со низа други привремени популации.
- Постојат два вида на еволутивен процес: микроеволуција и макроеволуција. Макроеволуцијата нема свои посебни механизми и се спроведува само благодарение на микроеволуционерните механизми.
- Секоја систематска група може или да процвета (биолошки напредок) или да изумре (биолошка регресија). Биолошкиот напредок се постигнува преку промени во структурата на организмите: ароморфози, идиоадаптации или општа дегенерација.
- Главните закони на еволуцијата се неговата неповратна природа, прогресивната компликација на формите на живот и развојот на приспособливоста на видовите кон нивната околина. Во исто време, еволуцијата нема крајна цел, односно процесот е ненасочен.
И покрај фактот што еволутивната теорија во изминатите децении беше збогатена со податоци од сродните науки - генетика, селекција и сл., таа сепак не зема предвид низа аспекти, на пример, насочени промени во наследниот материјал, па затоа во во иднина можно е да се создаде нов концепт на еволуција кој ќе ја замени синтетичката теорија.
Елементарни фактори на еволуцијата
Според синтетичката теорија на еволуцијата, елементарен еволутивен феномен се состои од промена на генетскиот состав на популацијата, а настаните и процесите кои водат до промени во генските базени се нарекуваат елементарни фактори на еволуцијата. Тие вклучуваат процес на мутација, популациски бранови, генетски нанос, изолација и природна селекција. Поради исклучителното значење на природната селекција во еволуцијата, ќе се разгледува посебно.
Процес на мутацијакоја е континуирана како и самата еволуција, ја одржува генетската хетерогеност на популацијата поради појавата на се повеќе нови варијанти на гени. Мутациите кои настануваат под влијание на надворешни и внатрешни фактори се класифицирани како генски, хромозомски и геномски.
Генски мутациисе јавуваат со фреквенција од 10 –4 –10 –7 по гамета, меѓутоа, поради фактот што кај луѓето и повеќето повисоки организми вкупниот број на гени може да достигне неколку десетици илјади, невозможно е да се замисли дека два организми се апсолутно идентични. Повеќето мутации што се појавуваат се рецесивни, особено затоа што доминантните мутации веднаш се предмет на природна селекција. Рецесивните мутации ја создаваат таа резерва на наследна варијабилност, но пред да се манифестираат во фенотипот, тие мора да се воспостават кај многу индивидуи во хетерозиготна состојба поради слободното преминување во популацијата.
Хромозомски мутацииповрзани со губење или пренесување на дел од хромозом (цел хромозом) на друг, исто така се доста чести кај различни организми, на пример, разликата помеѓу некои видови стаорци лежи во еден пар хромозоми, што го отежнува да ги премине.
Геномски мутации, поврзани со полиплоидизација, исто така доведуваат до репродуктивна изолација на новопојавената популација поради нарушувања во митозата на првата поделба на зиготот. Сепак, тие се доста распространети во растенијата и таквите растенија можат да растат на арктичките и алпските ливади поради нивната поголема отпорност на факторите на животната средина.
Комбинираната варијабилност, која обезбедува појава на нови варијанти на комбинирање гени во генотипот и, соодветно, ја зголемува веројатноста за појава на нови фенотипови, исто така придонесува за еволутивните процеси, бидејќи само кај луѓето бројот на варијанти на комбинации на хромозомите е 2 23, односно појавата на организам сличен на веќе постоечкиот, е речиси невозможна.
Бранови на населението.Спротивниот резултат (исцрпување на составот на гените) често е предизвикан од флуктуации во бројот на организми во природните популации, кои кај некои видови (инсекти, риби итн.) можат да се променат десетици или стотици пати - популациони бранови, или „бранови на животот“. Зголемување или намалување на бројот на поединци во популациите може да биде или периодични, така непериодични. Првите се сезонски или повеќегодишни, како што се миграциите кај птиците преселници или размножувањето кај дафнијата, кај кои има само женски единки во пролет и лето, а до есен се појавуваат мажјаци, неопходни за сексуална репродукција. Непериодични флуктуации во бројките често се предизвикани од нагло зголемување на количината на храна во поволна година, нарушување на условите на живеалиштата и зголемување на бројот на штетници или предатори.
Бидејќи реставрацијата на популацијата се јавува поради мал број индивидуи кои го немаат целиот сет на алели, новите и оригиналните популации ќе имаат различни генетски структури. Промената на фреквенцијата на гените во популацијата под влијание на случајни фактори се нарекува генетски нанос, или генетско-автоматски процеси. Тоа се јавува и при развојот на нови територии, бидејќи тие примаат исклучително ограничен број единки од даден вид, што може да доведе до нова популација. Затоа, генотиповите на овие индивидуи ( основачки ефект). Како резултат на генетскиот нанос, често се појавуваат нови хомозиготни форми (за мутантните алели), кои може да се покажат како адаптивно вредни и последователно ќе бидат прифатени од природната селекција.
Така, меѓу индиското население на американскиот континент и Лапонците, процентот на луѓе со крвна група I (0) е многу висок, додека групите III и IV се исклучително ретки. Веројатно, во првиот случај, основачите на популацијата биле поединци кои го немале алелот IB, или тој бил изгубен при процесот на селекција.
До одреден момент, се јавува размена на алели помеѓу соседните популации како резултат на вкрстување помеѓу поединци од различни популации - генски тек, што ја намалува дивергенцијата помеѓу поединечните популации, но со појавата на изолација таа престанува. Во суштина, генскиот тек е одложен процес на мутација.
Изолација.Сите промени во генетската структура на популацијата мора да се поправат, што се случува благодарение на тоа изолација- појавата на какви било бариери (географски, еколошки, бихејвиорални, репродуктивни итн.) кои го комплицираат и оневозможуваат преминувањето на поединци од различни популации. Иако самата изолација не создава нови форми, таа сепак ги зачувува генетските разлики помеѓу популациите кои се предмет на дејството на природната селекција. Постојат две форми на изолација: географска и биолошка.
Географска изолацијасе јавува како резултат на поделбата на областа со физички бариери (водни пречки за копнените организми, копнени површини за водни видови, алтернација на издигнати области и рамнини); Ова е олеснето со седентарен или приврзан (кај растенијата) начин на живот. Понекогаш географската изолација може да биде предизвикана од проширувањето на опсегот на еден вид со последователно изумирање на неговите популации во средните територии.
Биолошка изолацијае последица на одредени дивергенции на организми во рамките на истиот вид кои на некој начин го спречуваат слободното вкрстување. Постојат неколку видови на биолошка изолација: еколошка, сезонска, етолошка, морфолошка и генетска. Еколошка изолацијапостигнато преку поделбата на еколошките ниши (на пример, претпочитање за одредени живеалишта или типови на храна, како што е вкрстеното ливче од смрека и боровината). Сезонски(привремена) изолација е забележана во случај на репродукција на индивидуи од ист вид во различни времиња (различни залихи на харинга). Етолошката изолација зависи од карактеристиките на однесувањето (карактеристики на ритуалот на додворување, боење, „пеење“ на жени и мажи од различни популации). На морфолошка изолацијаПречка за вкрстување е неусогласеноста во структурата на репродуктивните органи или дури и големината на телото (Пекинез и Велика Дане). Генетска изолацијаима најголемо влијание и се манифестира во некомпатибилност на герминативните клетки (смрт на зиготот по оплодувањето), стерилитет или намалена одржливост на хибридите. Причините за ова се особеностите на бројот и обликот на хромозомите, како резултат на што целосната клеточна поделба (митоза и мејоза) станува невозможна.
Со нарушување на слободното вкрстување меѓу популациите, изолацијата на тој начин ги зајакнува кај нив оние разлики што настанале на генотипско ниво поради мутации и флуктуации во бројките. Во овој случај, секоја популација е предмет на дејство на природната селекција одделно од другата, а тоа на крајот доведува до дивергенција.
Креативната улога на природната селекција во еволуцијата
Природната селекција функционира како еден вид „сито“ што ги подредува генотиповите според нивниот степен на подготвеност. Сепак, Чарлс Дарвин нагласи дека селекцијата не е само и не толку насочена кон зачувување на исклучиво најдоброто, туку кон отстранување на најлошото, односно ви овозможува да ја зачувате мултиваријантноста. Функцијата на природната селекција не е ограничена на ова, бидејќи обезбедува репродукција на прилагодени генотипови и, на тој начин, ја одредува насоката на еволуцијата, постојано собирајќи случајни и бројни отстапувања. Природната селекција нема одредена цел: врз основа на ист материјал (наследна варијабилност) под различни услови, може да се добијат различни резултати.
Во овој поглед, факторот на еволуција што се разгледува не може да се спореди со работата на скулпторот кој копал блок од мермер, туку делува како далечен предок на човекот, правејќи алатка од фрагмент од камен, без да го замисли конечниот резултат; што зависи не само од природата на каменот и неговиот облик, туку и од силата, насоката на ударот итн. Меѓутоа, во случај на неуспех, селекцијата, како хуманоидно суштество, ја отфрла „погрешната“ форма.
Цената за избор е појава генетско оптоварување, односно акумулација на мутации во популација, која со текот на времето може да стане доминантна поради ненадејната смрт на повеќето индивидуи или миграцијата на мал број од нив.
Под притисок на природната селекција, не само што се формира разновидноста на видовите, туку се зголемува и нивното ниво на организација, вклучително и нивната компликација или специјализација. Меѓутоа, за разлика од вештачката селекција, спроведена од луѓето само заради економски вредни особини, честопати на штета на адаптивните својства, природната селекција не може да придонесе за ова, бидејќи ниту една адаптација во природата не може да ја компензира штетата од намалувањето на одржливоста на населението.
Истражување на С.С. Четвериков
Еден од важните чекори кон помирувањето на дарвинизмот и генетиката беше направен од московскиот зоолог С. С. Четвериков (1880–1959). Врз основа на резултатите од студијата за генетскиот состав на природните популации на мушичката Drosophila, тој докажа дека тие носат многу рецесивни мутации во хетерозиготна форма кои не ја нарушуваат фенотипската униформност. Повеќето од овие мутации се неповолни за телото и создаваат т.н генетско оптоварување, намалување на приспособливоста на населението како целина кон неговата околина. Некои мутации кои немаат адаптивно значење во даден момент од развојот на видот може да добијат одредена вредност подоцна, и на тој начин се резерва на наследна варијабилност.Ширењето на таквите мутации меѓу поединците од популацијата како резултат на последователни слободни вкрстувања на крајот може да доведе до нивна транзиција во хомозиготна состојба и манифестација во фенотипот. Ако оваа состојба на знакот е Фен- е адаптивен, потоа по неколку генерации целосно ќе го измести доминантниот фене, заедно со неговите носители, од популацијата што е помалку соодветна на дадените услови. Така, поради ваквите еволутивни промени, се задржува само рецесивниот мутантен алел, а неговиот доминантен алел исчезнува.
Ајде да се обидеме да го докажеме ова со конкретен пример. Кога проучувате одредена популација, можете да откриете дека не само неговата фенотипска, туку и нејзината генотипска структура може да остане непроменета долго време, поради слободното вкрстување или панмиксијадиплоидни организми.
Овој феномен е опишан со закон Харди-Вајнберг, според кој во идеална популација со неограничена големина во отсуство на мутации, миграции, популациски бранови, генетски нанос, природна селекција и предмет на слободно вкрстување, фреквенциите на алели и генотипови на диплоидни организми нема да се менуваат во текот на одреден број генерации .
На пример, кај една популација одредена карактеристика е кодирана од два алели од истиот ген - доминантна ( А) и рецесивно ( А). Фреквенцијата на доминантниот алел е означена како Ри рецесивен - q. Збирот на фреквенциите на овие алели е 1: стр + q= 1. Затоа, ако ја знаеме фреквенцијата на доминантниот алел, тогаш можеме да ја одредиме фреквенцијата на рецесивниот алел: q = 1 – стр. Всушност, фреквенциите на алелите се еднакви на веројатноста за формирање на соодветните гамети. Потоа, по формирањето на зиготите, фреквенциите на генотипот во првата генерација ќе бидат:
(pA + ка) 2 = стр 2 А.А. + 2pqAa + q 2 аа = 1,
Каде стр 2 А.А.- фреквенција на доминантни хомозиготи;
2pqAa- фреквенција на хетерозиготи;
q 2 аа- фреквенција на рецесивни хомозиготи.
Лесно е да се пресмета дека во следните генерации фреквенциите на генотиповите ќе останат исти, одржувајќи ја генетската разновидност на популацијата. Но, во природата не постојат идеални популации, и затоа, во нив, мутантните алели не само што можат да опстојуваат, туку и да се шират, па дури и да ги заменат претходно почестите алели.
Четвериков јасно сфатил дека природната селекција не ги елиминира само поединечните помалку успешни особини и, соодветно, алелите што ги кодираат, туку делува и на целиот комплекс на гени кои влијаат на манифестацијата на одреден ген во фенотипот, или генотипска средина. Како генотипска средина, целиот генотип моментално се смета како збир на гени кои можат да ја подобрат или ослабат манифестацијата на специфични алели.
Не помалку важни во развојот на еволуциското учење се студиите на С. С. Четвериков во областа на динамиката на населението, особено „брановите на животот“ или популационите бранови. Додека бил студент, во 1905 година објавил статија за можноста од појава на масовна репродукција на инсекти и подеднакво брз пад на нивниот број.
Улогата на еволутивната теорија во формирањето на модерната природонаучна слика на светот
Важноста на еволутивната теорија во развојот на биологијата и другите природни науки тешко може да се прецени, бидејќи таа прва ги објасни условите, причините, механизмите и резултатите од историскиот развој на животот на нашата планета, односно даде материјалистичко објаснување на развојот на органскиот свет. Дополнително, теоријата за природна селекција беше првата навистина научна теорија за биолошката еволуција, бидејќи кога беше создадена, Чарлс Дарвин не се потпираше на шпекулативни конструкции, туку поаѓаше од сопствените набљудувања и се потпираше на вистинските својства на живите организми. Истовремено, таа ги збогати биолошките алатки со историскиот метод.
Формулирањето на еволутивната теорија не само што предизвика жестока научна дебата, туку и даде поттик за развој на такви науки како што се општа биологија, генетика, селекција, антропологија и голем број други. Во овој поглед, не може да не се согласи со изјавата дека теоријата на еволуцијата ја круниса следната фаза во развојот на биологијата и стана почетна точка за нејзиниот напредок во дваесеттиот век.
Доказ за еволуцијата на живата природа. Резултати од еволуцијата: приспособливост на организмите кон нивната околина, разновидност на видови
Доказ за еволуцијата на дивиот свет
Во различни области на биологијата, дури и пред Чарлс Дарвин и по објавувањето на неговата теорија за еволуција, беа добиени цела низа докази за да ја поткрепат. Овој доказ се нарекува докази за еволуција. Најчесто цитирани се палеонтолошки, биогеографски, компаративни ембриолошки, компаративни анатомски и компаративни биохемиски докази за еволуцијата, иако податоците од таксономијата, како и изборот на растенија и животни, не можат да бидат намалени.
Палеонтолошки доказиврз основа на проучување на фосилни остатоци од организми. Тие вклучуваат не само добро сочувани организми замрзнати во мраз или обвиени во килибар, туку и „мумии“ откриени во кисели тресетни мочуришта, како и остатоци од организми и фосили зачувани во седиментни карпи. Присуството во античките карпи на поедноставни организми отколку во подоцнежните слоеви, и фактот дека видовите пронајдени на едно ниво исчезнуваат на друго, се смета за еден од најзначајните докази за еволуцијата и се објаснува со појавата и изумирањето на видовите во соодветните епохи поради на промените во условите на животната средина.
И покрај фактот што досега се откриени неколку фосилни остатоци и недостасуваат многу фрагменти од фосилните остатоци поради малата веројатност за зачувување на органски остатоци, сепак се пронајдени форми на организми кои имаат знаци и на еволутивно подревните и на помладите групи. на организмите. Ваквите форми на организми се нарекуваат преодни форми. Истакнати претставници на преодните форми, кои го илустрираат преминот од риби кон копнени 'рбетници, се рибите со лобуси и стегоцефалите, а Археоптериксот зазема одредено место помеѓу влекачите и птиците.
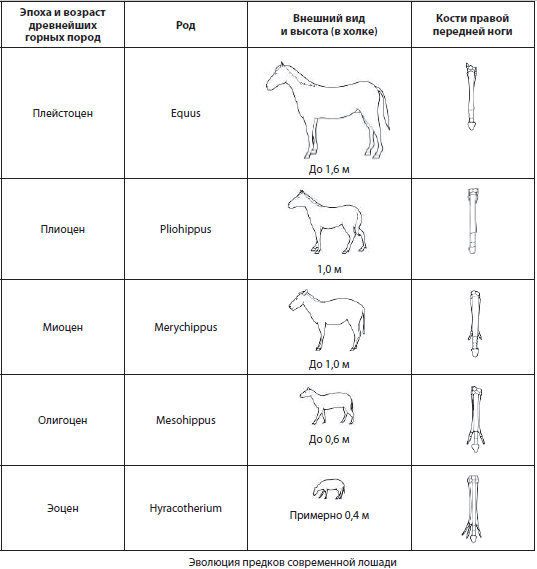
Редовите на фосилни форми кои се постојано поврзани едни со други во процесот на еволуција не само со општи, туку и со посебни структурни карактеристики се нарекуваат филогенетска серија. Тие може да бидат претставени со фосилни остатоци од различни континенти и да тврдат дека се повеќе или помалку комплетни, но нивното проучување е невозможно без споредба со живите форми со цел да се демонстрира прогресијата на еволутивниот процес. Класичен пример за филогенетска серија е еволуцијата на предците на коњот, проучувана од основачот на еволутивната палеонтологија В.О. Ковалевски.
Биогеографски докази. Биогеографијакако науката ги проучува моделите на дистрибуција и дистрибуција на видовите, родовите и другите групи на живи организми, како и нивните заедници, на површината на нашата планета.
Отсуството во кој било дел од земјината површина на видови на организми кои се приспособени на такво живеалиште и добро се вкорени кога вештачки се увезуваат, како зајаците во Австралија, како и присуството на слични форми на организми во делови од земјата лоцирани на значителни растојанија едни од други укажуваат, пред сè, дека изгледот на Земјата не бил секогаш вака, а геолошките трансформации, особено континенталниот нанос, формирањето на планини, порастот и падот на нивото на Светскиот океан влијаат еволуцијата на организмите. На пример, четири слични видови на белодробни риби живеат во тропските региони на Јужна Америка, Јужна Африка и Австралија, додека живеалиштата на камили и лами кои припаѓаат на истиот ред се наоѓаат во Северна Африка, поголемиот дел од Азија и Јужна Америка. Палеонтолошките студии покажаа дека камилите и ламите потекнуваат од заеднички предок кој некогаш живеел во Северна Америка, а потоа се прошириле во Азија преку претходно постоечкиот истмус на местото на Беринговиот теснец, а исто така и преку Панамскиот истмус до Јужна Америка. Последователно, сите претставници на ова семејство во средните региони исчезнаа, а во регионалните региони се формираа нови видови во процесот на еволуција. Поранешното одвојување на Австралија од другите копнени маси дозволило таму да се формира сосема посебна флора и фауна, во која биле зачувани такви форми на цицачи како монотреми - птицечовка и ехидна.
Од гледна точка на биогеографијата, може да се објасни и разновидноста на Дарвиновите сики на островите Галапагос, кои се оддалечени 1200 km од брегот на Јужна Америка и се со вулканско потекло. Очигледно, претставниците на единствениот вид сики во Еквадор еднаш летале или им биле запознаени, а потоа, како што се множеле, некои од поединците се населиле на преостанатите острови. На централните големи острови, борбата за егзистенција (храна, места за гнездење итн.) била најакутна, поради што се формирале видови малку различни едни од други по надворешни карактеристики, кои консумирале различна храна (семиња, овошје, нектар, инсекти, итн.) .).
Тие влијаеле на дистрибуцијата на различни групи организми и на промените во климатските услови на Земјата, што придонело за просперитет на некои групи, а за изумирање на други. Поединечни видови или групи на организми кои преживеале од претходно широко распространетите флора и фауна се нарекуваат моштите. Тука спаѓаат гинко, секвоја, лале, риба со лобус челакант, итн. Во поширока смисла, видовите растенија и животни кои живеат на ограничени области на територија или водни површини се нарекуваат ендемични, или ендемични. На пример, сите претставници на автохтоната флора и фауна на Австралија се ендемични, а во флората и фауната на Бајкалското Езеро до 75% од нив се ендемични.
Компаративни анатомски докази.Студијата за анатомијата на сродните групи на животни и растенија дава убедливи докази за сличноста во структурата на нивните органи. И покрај фактот што факторите на животната средина сигурно оставаат свој белег на структурата на органите, кај ангиоспермите, со сета своја неверојатна разновидност, цвеќињата имаат сепали, ливчиња, стомаци и пестици, а кај копнените 'рбетници, екстремитетот е изграден според пет прсти. план. Органите кои имаат слична структура, заземаат иста положба во телото и се развиваат од истите зачетоци во сродните организми, но извршуваат различни функции, се нарекуваат хомологни. Така, аудитивните жлезди (чекан, инкус и стреме) се хомологни со жабрените сводови на рибите, отровните жлезди на змиите се плунковните жлезди на другите 'рбетници, млечните жлезди на цицачите се потните жлезди, флиперите на фоките и китатите. се крилја на птици, екстремитети на коњи и кртови.

Органите кои долго време не функционираат најверојатно се претвораат во вестигијални (рудименти)- структури кои се недоволно развиени во споредба со прадедовните форми и го изгубиле своето основно значење. Тука спаѓаат фибулата кај птиците, очите кај бенките и кртовите стаорци, косата, кокцигеумот и слепото црево кај луѓето итн.
Поединечни поединци, сепак, може да покажат карактеристики кои се отсутни кај даден вид, но биле присутни кај далечните предци - атавизми, на пример, три прсти кај модерните коњи, развој на дополнителни парови на млечни жлезди, опашка и влакна на целото човечко тело.
Ако хомологните органи се доказ во корист на поврзаноста на организмите и дивергенцијата во процесот на еволуција, тогаш слични тела- слични структури во организми од различни групи кои вршат исти функции, напротив, се примери конвергенција(конвергенција е генерално независен развој на слични карактеристики кај различни групи на организми кои постојат во исти услови) и го потврдуваат фактот дека околината остава значителен печат на организмот. Аналози се крилјата на инсектите и птиците, очите на 'рбетниците и цефалоподите (лигњи, октоподи) и споените екстремитети на членконогите и копнените 'рбетници.

Компаративни ембриолошки докази.Проучувајќи го ембрионалниот развој кај претставници на различни групи 'рбетници, К. Баер го откри нивното впечатливо структурно единство, особено во раните фази на развојот ( закон за герминална сличност). Подоцна E. Haeckel формулирани биогенетски закон, според кој онтогенезата е кратко повторување на филогенезата, т.е. фазите низ кои поминува еден организам во процесот на неговиот индивидуален развој го повторуваат историскиот развој на групата на која и припаѓа.
Така, во првите фази на развој, ембрионот на 'рбетник добива структурни карактеристики карактеристични за рибите, а потоа и за водоземците и, на крајот, за групата на која припаѓа. Оваа трансформација се објаснува со фактот дека секоја од горенаведените класи има заеднички предци со модерните влекачи, птици и цицачи.

Сепак, биогенетскиот закон има голем број ограничувања, и затоа рускиот научник А.Н. Северцов значително го ограничи опсегот на неговата примена со повторување во онтогенезата исклучиво карактеристиките на ембрионските фази на развој на формите на предците.
Компаративни биохемиски докази.Развојот на попрецизни методи за биохемиска анализа им обезбеди на еволуционерните научници нова група податоци во корист на историскиот развој на органскиот свет, бидејќи присуството на исти супстанции во сите организми укажува на можна биохемиска хомологија, слична на онаа кај ниво на органи и ткива. Компаративни биохемиски студии на примарната структура на таквите широко распространети протеини како цитохром Сои хемоглобинот, како и нуклеинските киселини, особено рРНК, покажаа дека многу од нив имаат речиси иста структура и вршат исти функции кај претставници на различни видови, а колку е поблиска врската, толку е поголема сличноста во структурата на супстанциите што се испитуваат.
Така, теоријата на еволуцијата е потврдена со значителна количина на податоци од различни извори, што уште еднаш укажува на нејзината веродостојност, но таа сепак ќе се менува и рафинира, бидејќи многу аспекти од животот на организмите остануваат надвор од видното поле на истражувачите. .
Резултати од еволуцијата: приспособливост на организмите кон нивната околина, разновидност на видови
Покрај општите карактеристики карактеристични за претставниците на одредено кралство, видовите живи организми се карактеризираат со неверојатна разновидност на карактеристики на надворешна и внатрешна структура, животна активност, па дури и однесување што се појавиле и биле избрани во процесот на еволуција и обезбедуваат адаптација. на условите за живеење. Сепак, не треба да се претпоставува дека бидејќи птиците и инсектите имаат крилја, тоа се должи на директното дејство на воздухот, бидејќи има многу инсекти и птици без крилја. Горенаведените адаптации беа избрани преку процес на природна селекција од целиот спектар на достапни мутации.
Епифитичните растенија, кои живеат не на почвата, туку на дрвјата, се приспособиле да ја апсорбираат атмосферската влага со помош на корени без влакна од коренот, но со посебно хигроскопско ткиво - веламен. Некои бромелијади можат да апсорбираат водена пареа во влажната атмосфера на тропските предели користејќи ги влакната на нивните лисја.
Инсективојадните растенија (сончеви, Венера стапици за мува) кои живеат на почви каде што азот е недостапен поради една или друга причина имаат развиено механизам за привлекување и апсорпција на мали животни, најчесто инсекти, кои се извор на потребниот елемент за нив.
За да се заштитат од јадење од тревопасни животни, многу растенија кои водат приврзан начин на живот развиле пасивни средства за заштита, како што се трње (глог), трње (роза), боцкави влакна (коприва), акумулација на кристали на калциум оксалат (киселица). , биолошки активни материи во ткивата (кафе, глог) и сл. Кај некои од нив дури и семките во незрели плодови се опкружени со камени клетки кои ги спречуваат штетниците да стигнат до нив, а дури наесен се случува процесот на отсекување дрво, кој овозможува семето да влезе во почвата и да никне (круша).
Околината има формативно влијание и врз животните. Така, многу риби и водни цицачи имаат рационализирана форма на телото, што им олеснува да се движат низ нејзината дебелина. Сепак, не треба да се претпоставува дека водата директно влијае на обликот на телото, едноставно во процесот на еволуција оние животни што ја поседувале оваа особина се покажале како најприлагодени на неа.
Телото на китовите и делфините не е покриено со влакна, додека сродната група на перконози има повеќе или помалку намален слој на влакна, бидејќи, за разлика од првите, дел од своето време поминуваат на копно, каде што без волна нивната кожа веднаш би стане леден.
Телото на повеќето риби е покриено со лушпи, кои од долната страна се посветли отколку на врвот, поради што овие животни тешко се забележуваат одозгора до природните непријатели на позадината на дното, а одоздола - наспроти позадина на небото. Боење што ги прави животните невидливи за нивните непријатели или плен се нарекува покровителство. Тоа е широко распространето во природата. Впечатлив пример за такво боење е боењето на долната страна на крилјата на пеперутката калима, која, откако седна на гранка и ги преклопи крилјата заедно, излегува дека личи на сув лист. Други инсекти, како што се инсектите со стапчиња, се камуфлираат како растителни гранчиња.
Забележаното или пругастото обојување има и адаптивно значење, бидејќи на позадината на почвата, птиците како што се потполошки или бајдери не се видливи дури ни одблизу. Невидливи се и забележаните јајца на птиците кои се гнездат на земја.
Бојата на животните не е секогаш константна како онаа на зебрата, на пример, пробивањето и камелеонот можат да го променат во зависност од природата на местото каде што се наоѓаат. Кукавиците, ставајќи ги своите јајца во гнездата на разни птици, можат да ја менуваат бојата на нивните лушпи на таков начин што „сопствениците“ на гнездото не ги забележуваат разликите меѓу него и нивните јајца.
Боењето на животните не ги прави секогаш невидливи - многу од нив едноставно го привлекуваат окото, што треба да предупреди на опасност. Повеќето од овие инсекти и влекачи се отровни до еден или друг степен, како бубамара или оса, така што предаторот, откако доживеал непријатни чувства неколку пати откако јаде таков предмет, го избегнува. Сепак, предупредувачко боењене е универзален, бидејќи некои птици се приспособиле да се хранат со нив (жиче).
Зголемените шанси за преживување кај индивидуите со предупредувачка обоеност придонесоа за негово појавување кај претставници на други видови без соодветни причини. Овој феномен се нарекува мимикрија. Така, неотровните гасеници на некои видови пеперутки имитираат отровни, а бубамарите имитираат еден од видовите лебарки. Сепак, птиците можат брзо да научат да ги разликуваат отровните организми од неотровните и да ги консумираат вторите, избегнувајќи ги индивидуите кои служеле како пример.
Во некои случаи, може да се забележи и спротивен феномен - грабливите животни имитираат безопасни животни во боја, што им овозможува да и пријдат на жртвата на блиско растојание, а потоа да нападнат (блени со сабја заби).
Заштита за многу видови е обезбедена и со адаптивното однесување, кое е поврзано со складирање храна за зимата, грижа за потомството, замрзнување на место или, обратно, усвојување заканувачка поза. Така, речните дабари подготвуваат неколку кубни метри гранки, делови од стебла и друга растителна храна за зимата, поплавувајќи ја во вода во близина на „колибите“.
Грижата за потомството е карактеристична главно за цицачи и птици, меѓутоа, таа се среќава и кај претставници на други класи на хорди. На пример, познато е агресивното однесување на машките стапчиња, што ги избрка сите непријатели од гнездото во кое се наоѓаат јајцата. Машките жаби со канџи ги обвиткуваат јајцата околу нивните нозе и ги носат додека не се изведат полноглавците.
Дури и некои инсекти можат да им обезбедат на своите потомци поповолно живеалиште. На пример, пчелите ги хранат своите ларви, а младите пчели на почетокот „работат“ само во кошницата. Мравките ги движат своите кукли нагоре и надолу во мравјалникот, во зависност од температурата и влажноста, а кога постои закана од поплава, тие генерално ги носат со себе. Карабите бубачки подготвуваат специјални топчиња од животински отпад за нивните ларви.
Кога им се заканува напад, многу инсекти замрзнуваат на своето место и добиваат форма на суви стапчиња, гранчиња и лисја. Напротив, вајперите се креваат и ја надувуваат хаубата, додека змијата ѕвечарка испушта посебен звук со штракач кој се наоѓа на крајот од опашката.
Прилагодувањата на однесувањето се надополнуваат со физиолошки поврзани со карактеристиките на живеалиштето. Така, едно лице може да остане под вода без опрема за нуркање само неколку минути, по што може да изгуби свест и да умре поради недостаток на кислород, а китовите не излегуваат на површина доста долго. Нивниот волумен на белите дробови не е премногу голем, но има и други физиолошки адаптации, на пример, во мускулите има висока концентрација на респираторниот пигмент - миоглобин, кој, како што беше, складира кислород и го ослободува за време на потопувањето. Покрај тоа, китовите имаат посебна формација - „прекрасна мрежа“, која овозможува користење на кислород дури и од венска крв.
Животните во жешките живеалишта, како што се пустините, постојано се изложени на ризик од прегревање и губење на вишокот на влага. Затоа, фенек лисицата има исклучително големи уши кои и овозможуваат да зрачи топлина. Водоземците од пустинските региони, за да избегнат губење на влага преку кожата, се принудени да се префрлат на ноќен начин на живот, кога се зголемува влажноста и се појавува роса.
Птиците кои го совладале воздушното живеалиште, покрај анатомските и морфолошките прилагодувања за летот, имаат и важни физиолошки карактеристики. На пример, поради фактот што движењето во воздухот бара исклучително висока потрошувачка на енергија, оваа група на 'рбетници се карактеризира со висока стапка на метаболизам, а излачените метаболички производи веднаш се елиминираат, што помага да се намали специфичната густина на телото.
Адаптациите кон околината, и покрај сета нивна совршенство, се релативни. Така, некои видови млечка трева произведуваат алкалоиди кои се отровни за повеќето животни, но гасениците на еден вид пеперутки - данаидите - не само што се хранат со ткивата на млечната трева, туку и ги акумулираат овие алкалоиди, станувајќи нејадливи за птиците.
Покрај тоа, адаптациите се корисни само во одредена средина и се бескорисни во друга средина. На пример, реткиот и голем предатор, тигарот Усури, како и сите мачки, има меки влошки на шепите и остри канџи што се повлекуваат, остри заби, одличен вид дури и во темница, силен слух и силни мускули, што му овозможува да го открие својот плен, се прикрадне на него незабележано и направи заседа. Сепак, неговата пругаста боја го камуфлира само во пролет, лето и есен, додека на снегот станува јасно видлива и тигарот може да смета само на молскавичен напад.
Цветовите на смокви, кои даваат вредни плодови, имаат таква специфична структура што ги опрашуваат само бластофаготните оси и затоа, кога се воведуваат во културата, долго време не вродиле со плод. Само развојот на партенокарпични сорти смокви (формирање плодови без оплодување) може да ја спаси ситуацијата.
И покрај фактот што се опишани примери на специјација во многу кратки временски периоди, како што е случајот со штракаат во кавкаските ливади, кои поради редовното косење прво се поделиле на две популации - рано цветање и плодно и доцноцветно, всушност, микроеволуцијата најверојатно бара многу подолги периоди - многу векови, бидејќи човештвото, чии различни групи беа одделени едни од други со милениуми, сепак, никогаш не се делеше на различни видови. Меѓутоа, бидејќи еволуцијата има практично неограничено време, во текот на стотици милиони и милијарди години, неколку милијарди видови веќе живееле на Земјата, од кои повеќето исчезнале, а оние што дошле до нас се квалитативни фази на овој тековен процес.
Според современите податоци, на Земјата има над 2 милиони видови живи организми, од кои повеќето (приближно 1,5 милиони видови) припаѓаат на животинското царство, околу 400 илјади на растителното царство, над 100 илјади на царството на печурките и одмор - на бактерии. Ваквата неверојатна разновидност е резултат на дивергенција (дивергенција) на видовите според различни морфолошки, физиолошко-биохемиски, еколошки, генетски и репродуктивни карактеристики. На пример, еден од најголемите родови на растенија кои припаѓаат на семејството Orchidaceae, Dendrobium, вклучува над 1.400 видови, а родот на бубачки вклучува над 1.600 видови.
Класификацијата на организмите е задача на таксономијата, која веќе 2 илјади години се обидува да изгради не само хармонична хиерархија, туку „природен“ систем кој го одразува степенот на поврзаност на организмите. Сепак, сите обиди да се направи ова сè уште не се крунисани со успех, бидејќи во голем број случаи, во процесот на еволуција, не беше забележана само дивергенција на карактерите, туку и конвергенција (конвергенција), како резултат на што во многу далечни групи органите стекнале сличности, како што се очите на цефалоподите и очите на цицачите.
Макроеволуција. Насоки и патеки на еволуцијата (А. Н. Северцов, И. И. Шмалгаузен). Биолошки напредок и регресија, ароморфоза, идиоадаптација, дегенерација. Причини за биолошки напредок и регресија. Хипотези за потеклото на животот на Земјата. Основни ароморфози во еволуцијата на растенијата и животните. Компликација на живите организми во процесот на еволуција
Макроеволуција
Формирањето на еден вид означува нова рунда на еволутивниот процес, бидејќи индивидуите од овој вид, кои се поприлагодени на условите на животната средина отколку поединците од матичниот вид, постепено се населуваат на нови територии, а мутагенезата, брановите на населението, изолацијата и природната селекција играат нивната креативна улога во нејзините популации. Со текот на времето, од овие популации произлегуваат нови видови, кои поради генетската изолација имаат многу повеќе сличности меѓу себе отколку со видот од родот од кој се разгранувал родителскиот вид и на тој начин се јавува нов род, па нов семејство, ред (ред) , класа итн. Збирот на еволутивни процеси кои водат до појава на надспецифични таксони (родови, семејства, редови, класи итн.) се нарекува макроеволуција. Макроеволуционерните процеси, како што беа, ги генерализираат микроеволутивните промени кои се случуваат во долг временски период, притоа идентификувајќи ги главните трендови, насоки и модели на еволуција на органскиот свет, кои не се набљудуваат на пониско ниво. Досега не се идентификувани специфични механизми на макроеволуција, затоа се верува дека таа се спроведува само преку микроеволуциони процеси, но оваа позиција постојано е предмет на основана критика.
Појавата на сложениот хиерархиски систем на органскиот свет во голема мера е резултат на нееднаквата стапка на еволуција на различни групи на организми. Така, веќе споменатата гинко билоба беше, како да се каже, „зачувана“ со илјадници години, додека боровите кои се доста блиску до него значително се променија во ова време.
Насоки и патеки на еволуцијата (А. Н. Северцов, И. И. Шмалгаузен). Биолошки напредок и регресија, ароморфоза, идиоадаптација, дегенерација
Анализирајќи ја историјата на органскиот свет, може да се забележи дека во одредени временски периоди доминирале одредени групи на организми, кои потоа опаѓале или целосно исчезнале. Така, може да се разликуваат три главни линии насоки на еволуција: биолошки напредок, биолошка регресија и биолошка стабилизација. Значаен придонес во развојот на доктрината за насоките и патеките на еволуцијата дадоа руските еволуционисти А.Н.Северцов и И.И.Шмалгаузен.
Биолошки напредокповрзани со биолошкиот просперитет на групата како целина и го карактеризираат нејзиниот еволутивен успех. Тоа го отсликува природниот развој на живата природа од едноставна до сложена, од понизок степен на организација до повисок. Според А. род, род во семејство итн.). Во моментов, биолошки напредок е забележан кај ангиосперми, инсекти, коскени риби и цицачи.
Според А.Н. Северцов, биолошкиот напредок може да се постигне како резултат на одредени морфофизиолошки трансформации на организмите и тој идентификуваше три главни начини на постигнување: арогенеза, алогенеза и катагенеза.

Арогенеза, или морфофизиолошки напредок, е поврзан со значително проширување на опсегот на оваа група организми поради стекнување на големи структурни промени - ароморфози.
Ароморфозанаречена еволутивна трансформација на структурата и функциите на организмот, со што се зголемува неговото ниво на организација и се отвораат нови можности за адаптација на различни услови на постоење.
Примери за ароморфози се појавата на еукариотска клетка, повеќеклеточноста, појавата на срце кај рибите и нејзината поделба со целосна преграда кај птиците и цицачите, формирањето на цвет кај ангиоспермите итн.
Алогенеза, за разлика од арогенезата, не е придружена со проширување на опсегот, меѓутоа, во рамките на старата, се јавува значителна разновидност на форми кои имаат посебни прилагодувања на околината - идиоадаптации.
Идиоматска адаптација- ова е мала морфофизиолошка адаптација на посебни услови на животната средина, корисна во борбата за егзистенција, но не го менува нивото на организација. Овие промени се илустрирани со заштитната обоеност кај животните, разновидноста на устите кај инсектите, боцките на растенијата итн. Подеднакво успешен пример се сибите на Дарвин, специјализирани за разни видови храна, во кои трансформациите прво го зафатиле клунот, а потоа други делови од телото - пердуви, опашка и така натаму.
Колку и да изгледа парадоксално, поедноставувањето на организацијата може да доведе до биолошки напредок. Оваа патека се нарекува катагенеза.
Дегенерација- ова е поедноставување на организмите во процесот на еволуција, што е придружено со губење на одредени функции или органи.
Фазата на биолошки напредок се заменува со фаза биолошка стабилизација, чија суштина е да се зачуваат карактеристиките на даден вид како најповолни во дадена микросредина. Според I. I. Shmalhausen, тоа „не значи прекин на еволуцијата, напротив, тоа значи максимална конзистентност на организмот со промените во околината“. Во фаза на биолошка стабилизација се „живите фосили“ од целакант, гингко и сл.
Антиподот на биолошкиот напредок е биолошка регресија- еволутивен пад на дадена група поради неможноста да се прилагоди на промените на животната средина. Се манифестира во намалување на бројот на населението, стеснување на опсезите и намалување на бројот на пониско рангирани групи во повисок таксон. На група организми кои се во состојба на биолошка регресија и се заканува изумирање. Во историјата на органскиот свет може да се видат многу примери на овој феномен, а во моментов регресијата е карактеристична за некои папрати, водоземци и рептили. Со доаѓањето на човекот, биолошката регресија често се должи на неговите економски активности.
Насоките и патеките на еволуцијата на органскиот свет не се исклучуваат меѓусебно, односно појавата на ароморфоза не значи дека идиоадаптација или дегенерација повеќе не може да се случи. Напротив, според она што беше развиено од А. Н. Северцов и И. И. Шмалгаузен правило за промена на фаза, различните насоки на еволутивниот процес и начините за постигнување биолошки напредок природно се заменуваат едни со други. Во текот на еволуцијата, овие патишта се комбинираат: прилично ретки ароморфози пренесуваат група организми на квалитативно ново ниво на организација, а понатамошниот историски развој го следи патот на идиоадаптација или дегенерација, обезбедувајќи адаптација на специфичните услови на животната средина.
Причини за биолошки напредок и регресија
Во процесот на еволуција, барот на природната селекција е надминат и, соодветно, напредуваат само оние групи организми во кои наследната варијабилност создава доволен број комбинации кои можат да обезбедат опстанок на групата како целина.
Оние групи кои поради некоја причина немаат таков резерват, во повеќето случаи, се осудени на изумирање. Ова често се должи на нискиот притисок на селекција во претходните фази од еволутивниот процес, што доведе до тесна специјализација на групата или дури и до дегенеративни феномени. Последица на ова е неможноста да се прилагодат на новите услови на животната средина кога има нагли промени. Еклатантен пример за ова е ненадејната смрт на диносаурусите поради падот на џиновско небесно тело на Земјата пред 65 милиони години, што резултираше со земјотрес, издигнување на милиони тони прашина во воздухот, остар студен бран, и смртта на повеќето растенија и тревојади животни. Во исто време, предците на современите цицачи, немајќи тесни преференции за извори на храна и топлокрвни, можеа да ги преживеат овие услови и да заземат доминантна позиција на планетата.
Хипотези за потеклото на животот на Земјата
Од целиот опсег на хипотези за формирањето на Земјата, најголемиот број факти сведочат во корист на теоријата на „Биг Бенг“. Поради фактот што оваа научна претпоставка се заснова главно на теоретски пресметки, Големиот хадронски судирач, изграден во Европскиот центар за нуклеарни истражувања во близина на Женева (Швајцарија), е повикан експериментално да ја потврди. Според теоријата на Биг Бенг, Земјата е формирана пред повеќе од 4,5 милијарди години заедно со Сонцето и другите планети од Сончевиот систем како резултат на кондензација на облак од гас и прашина. Намалувањето на температурата на планетата и миграцијата на хемиските елементи на неа придонесоа за нејзино раслојување во јадрото, мантија и кора, а последователните геолошки процеси (движење на тектонски плочи, вулканска активност итн.) предизвикаа формирање на атмосфера и хидросфера.
Животот постои и на Земјата многу долго време, за што сведочат фосилните остатоци од различни организми во карпите, но физичките теории не можат да одговорат на прашањето за времето и причините за неговото појавување. Постојат две спротивставени гледишта за потеклото на животот на Земјата: теориите на абиогенезата и биогенезата. Теории на абиогенезатаја потврдуваат можноста за потеклото на живите од неживи суштества. Тука спаѓаат креационизмот, хипотезата за спонтано генерирање и теоријата за биохемиска еволуција од А.И.
Фундаментална позиција креационизамсоздавањето на светот било одредено натприродно битие (Создател), што се рефлектира во митовите за народите во светот и религиозните култови, но староста на планетата и животот на неа далеку ги надминуваат датумите наведени во овие извори, и има многу недоследности во нив.
Основач теории за спонтано генерирањеживотот се смета за старогрчки научник Аристотел, кој тврдел дека е можно повеќе пати да се појават нови суштества, на пример, дождовни црви од баричките, и црви и муви од расипано месо. Сепак, овие гледишта беа побиени во 17-19 век со смелите експерименти на Ф. Реди и Л. Пастер.
Италијанскиот лекар Франческо Реди во 1688 година ставал парчиња месо во саксии и цврсто ги затворал, но во нив не се појавиле црви, а тие се појавувале во отворени саксии. За да го побие преовладувачкото верување дека принципот на живот е содржан во воздухот, тој ги повторил своите експерименти, но не ги запечатил саксиите, туку ги покрил со неколку слоеви муслин и повторно животот не се појавил. И покрај убедливите податоци добиени од Ф. Реди, истражувањето на А. ван Леувенхук обезбеди нова храна за дискусии за „виталниот принцип“, кој продолжи во текот на следниот век.
Друг италијански истражувач, Лазаро Спаланзани, ги модифицирал експериментите на Ф. Реди во 1765 година со варење на лушпа од месо и зеленчук неколку часа и нивно запечатување. По неколку дена, тој исто така не нашол знаци на живот таму и заклучил дека живите суштества можат да настанат само од живи суштества.
Последниот удар на теоријата за спонтано генерирање го донел големиот француски микробиолог Луј Пастер во 1860 година, кога ставил варена супа во колба со S-врат и не добил никакви бактерии. Се чини дека ова сведочеше во корист на теориите на биогенезата, но прашањето остана отворено за тоа како се појавил самиот, првиот организам.
Советскиот биохемичар А.И. Опарин се обиде да одговори, доаѓајќи до заклучок дека составот на атмосферата на Земјата во првите фази од нејзиното постоење е сосема поинаков од она што е во нашево време. Најверојатно се состоеше од амонијак, метан, јаглерод диоксид и водена пареа, но не содржеше слободен кислород. Под влијание на електрични празнења со голема моќност и на високи температури, во него можело да се синтетизираат наједноставните органски соединенија, што е потврдено со експериментите на С. Милер и Г. Уреј во 1953 година, кои од горенаведените соединенија добиле неколку амино киселини, едноставни јаглехидрати, аденин, уреа, како и едноставни масни киселини, мравја и оцетна киселина.
Како и да е, синтезата на органски супстанции сè уште не значи појава на живот, па затоа го постави Опарин хипотеза за биохемиска еволуција, според кој разни органски материи настанале и се комбинирале во поголеми молекули во плитките води на морињата и океаните, каде што се најповолни услови за хемиска синтеза и полимеризација. Молекулите на РНК во моментов се сметаат за први носители на животот.
Некои од овие супстанции постепено формирале стабилни комплекси во водата - коацервира, или коацерват капки, што личи на капки маснотии во супа. Овие коацервати примиле различни материи од околниот раствор, кои биле подложени на хемиски трансформации кои се случуваат во капките. Како органски материи, самите коцервати не беа живи суштества, туку беа следниот чекор во нивното појавување.
Оние коацервати кои имале поволен однос на супстанции во својот состав, особено протеини и нуклеински киселини, благодарение на каталитичките својства на ензимските протеини, со текот на времето стекнале способност да го репродуцираат својот вид и да вршат метаболички реакции, додека структурата на протеините била кодирани од нуклеински киселини.
Меѓутоа, покрај репродукцијата, живите системи се карактеризираат и со зависност од снабдувањето со енергија однадвор. Овој проблем првично беше решен преку разградување без кислород на органските материи од околината (во тоа време немаше кислород во атмосферата), т.е.
хетеротрофична исхрана. Се покажа дека некои од апсорбираните органски материи можат да ја акумулираат енергијата на сончевата светлина, како што е хлорофилот, што овозможило голем број организми да се префрлат на автотрофична исхрана. Ослободувањето на кислород во атмосферата за време на фотосинтезата доведе до појава на поефикасно дишење на кислород, формирање на озонската обвивка и, на крајот, појава на организми на копно.
Така, резултатот на хемиската еволуција беше изгледот протобионти- примарни живи организми, од кои, како резултат на биолошката еволуција, настанале сите моментално постоечки видови.
Теоријата за биохемиска еволуција во нашево време е најпотврдена, но идејата за специфичните механизми за потеклото на животот е променета. На пример, се покажа дека формирањето на органски материи започнува во вселената, а органските материи играат важна улога дури и во самото формирање на планетите, обезбедувајќи адхезија на мали делови. Формирањето на органски материи се случува и во утробата на планетата: за време на една ерупција, вулканот ослободува до 15 тони органска материја. Постојат и други хипотези во однос на механизмите на концентрација на органски материи: замрзнување на растворот, апсорпција (врзување) на површината на одредени минерални соединенија, дејство на природни катализатори итн. Појавата на живот на Земјата во моментов е невозможна. бидејќи сите органски супстанци спонтано формирани во која било точка планетите веднаш би биле оксидирани од слободниот кислород на атмосферата или користени од хетеротрофни организми. Ова беше разбрано уште во 1871 година од Чарлс Дарвин.
Теории на биогенезатаго негираат спонтаното потекло на животот. Главните се хипотезата за стабилна состојба и хипотезата за панспермија. Првиот од нив се заснова на фактот дека животот постои засекогаш, меѓутоа, на нашата планета има многу антички карпи во кои нема траги од активноста на органскиот свет.
Хипотеза за панспермијатврди дека ембрионите на животот биле донесени на Земјата од вселената од некои вонземјани или од божествената промисла. Оваа хипотеза е поткрепена со два факти: потребата за сите живи суштества, што е прилично ретко на планетата, но често се среќава во метеоритите, од молибден, како и откривањето на организми слични на бактериите на метеоритите од Марс. Сепак, останува нејасно како настанал животот на други планети.
Основни ароморфози во еволуцијата на растенијата и животните
Растителните и животинските организми, кои претставуваат различни гранки на еволуцијата на органскиот свет, во процесот на историскиот развој самостојно стекнале одредени структурни карактеристики, кои ќе бидат опишани понатаму.
Кај растенијата најважни од нив се преминот од хаплоиден во диплоиден, независност од вода при процесот на оплодување, премин од надворешно во внатрешно оплодување и појава на двојно оплодување, поделба на телото на органи, развој на спроводниот систем, компликација и подобрување на ткивата, како и специјализација на опрашување со помош на инсекти и дистрибуција на семиња и плодови.
Преминот од хаплоидија во диплоидија ги направи растенијата поотпорни на факторите на околината поради намалениот ризик од рецесивни мутации. Очигледно, оваа трансформација влијаела на предците на васкуларните растенија, кои не вклучуваат бриофити, кои се карактеризираат со доминација на гаметофитот во животниот циклус.
Главните ароморфози во еволуцијата на животните се поврзани со појавата на повеќеклеточност и зголеменото распарчување на сите органски системи, појавата на силен скелет, развојот на централниот нервен систем, како и социјалното однесување во различни групи на високо организирани животни, кои му дадоа поттик на човечкиот напредок.
Компликација на живите организми во процесот на еволуција
Историјата на органскиот свет на Земјата се проучува од зачуваните остатоци, отпечатоци и други траги од виталната активност на живите организми. Таа е предмет на наука палеонтологијата. Врз основа на фактот дека остатоците од различни организми се наоѓаат во различни карпести слоеви, создадена е геохронолошка скала, според која историјата на Земјата била поделена на одредени временски периоди: еони, епохи, периоди и векови.
Еоннаречен голем временски период во геолошката историја, комбинирајќи неколку епохи. Во моментов, се разликуваат само два еони: криптозоик (скриен живот) и фанрозоичен (манифестен живот). Ера- ова е временски период во геолошката историја, кој е поделба на еден еон, кој, пак, обединува периоди. Во криптозоикот има две епохи (архејски и протерозојски), додека во фанрозојски три (палеозојски, мезозојски и кенозојски).
Важна улога во создавањето на геохронолошката скала одиграла од водечки фосили- остатоци од организми кои биле многубројни во одредени временски периоди и се добро сочувани.
Развој на животот во криптозоикот.Архејот и протерозоикот го сочинуваат најголемиот дел од историјата на животот (период пред 4,6 милијарди години - пред 0,6 милијарди години), но има малку информации за животот во тој период. Првите остатоци од органски материи од биогено потекло се стари околу 3,8 милијарди години, а прокариотските организми постоеле веќе пред 3,5 милијарди години. Првите прокариоти беа дел од специфични екосистеми - цијанобактериски душеци, благодарение на чија активност се формираа специфични седиментни карпи строматолити („камени теписи“).
Разбирањето на животот на античките прокариотски екосистеми беше помогнато со откривањето на нивните современи аналози - строматолити во заливот Ајкула во Австралија и специфични филмови на површината на почвата во заливот Сиваш во Украина. На површината на цијанобактериските душеци има фотосинтетички цијанобактерии, а под нивниот слој има исклучително разновидни бактерии од други групи и археи. Минералните материи кои се таложат на површината на подлогата и се формираат поради неговата витална активност се таложат во слоеви (приближно 0,3 mm годишно). Вакви примитивни екосистеми можат да постојат само на места непогодни за живеење за други организми, и навистина, и двете горенаведени живеалишта се карактеризираат со исклучително висока соленост.
Бројни податоци покажуваат дека на почетокот Земјата имала обновлива атмосфера, која вклучувала: јаглерод диоксид, водена пареа, сулфур оксид, како и јаглерод моноксид, водород, водород сулфид, амонијак, метан итн. Првите организми на Земјата биле анаеробите , сепак, благодарение на фотосинтезата на цијанобактериите, во околината се ослободува слободен кислород, кој на почетокот брзо се поврзува со редукционите агенси во околината, а дури по врзувањето на сите редукциони средства околината почнала да добива оксидирачки својства. За оваа транзиција сведочи таложење на оксидирани форми на железо - хематит и магнетит.
Пред околу 2 милијарди години, како резултат на геофизичките процеси, речиси целото железо неврзано во седиментните карпи се пресели во јадрото на планетата, а кислородот почна да се акумулира во атмосферата поради отсуството на овој елемент - „кислородна револуција“ се случи. Тоа беше пресвртница во историјата на Земјата, која повлекува не само промена во составот на атмосферата и формирање на озонската обвивка во атмосферата - главен предуслов за населување на земјата, туку и составот на карпите настанати на површината на Земјата.
Друг важен настан се случи во протерозоикот - појавата на еукариоти. Во последниве години, беше можно да се соберат убедливи докази за теоријата на ендосимбиогенетското потекло на еукариотската клетка - преку симбиоза на неколку прокариотски клетки. Веројатно, „главниот“ предок на еукариотите биле археите, кои се префрлиле на апсорпција на честички од храна со фагоцитоза. Наследниот апарат се пресели длабоко во клетката, сепак ја одржуваше својата врска со мембраната поради преминот на надворешната мембрана на нуклеарната мембрана што се појавува во мембраните на ендоплазматскиот ретикулум.
Геохронолошка историја на Земјата Еон Ера Период Почеток, пред милиони години Времетраење, милиони години Развој на живототФанерозоичен кенозоик антропоген 1,5 1,5 Четири ледени доба, проследени со поплави, доведоа до формирање на ладно отпорна флора и фауна (мамути, мошус волови, ирваси, леминзи). Размена на животни и растенија меѓу континентите поради појавата на копнени мостови. Доминација на плацентарните цицачи. Истребување на многу големи цицачи. Формирањето на човекот како биолошки вид и неговото населување. Припитомување на животните и одгледување растенија. Исчезнување на многу видови живи организми поради човековата економска активност Неоген 25 23.5 Распределба на житни култури. Формирање на сите модерни редови на цицачи. Појавата на мајмуните Палеоген 65 40 Доминација на цветни растенија, цицачи и птици. Појавата на копитари, месојади, перченици, примати и др. Појавата на торбари и плацентарни цицачи. Доминација на гимноспермите Тријас 225 30 Првите цицачи и птици. Рептилите се многубројни. Распределба на тревни спори Палеозојски Перм 280 55 Појавување на современи инсекти. Развој на влекачи. Истребување на голем број групи без'рбетници. Распределба на четинари Јаглерод 345 65 Први влекачи. Појавата на крилести инсекти. Доминираат папрати и конска опашка Девон 395 50 Рибите се многубројни. Првите водоземци. Појавата на главните групи на спори, првите гимносперми и габи Силуријан 430 35 Алгите се во изобилство. Првите копнени растенија и животни (пајаци). Рибите духови и раковите скорпии се вообичаени Ордовицијански 500 70 Коралите и трилобитите се во изобилство. Цветање на зелени, кафеави и црвени алги. Појавата на првите акордати Камбријан 570 70 Бројни рибини фосили. Морските ежови и трилобити се вообичаени. Појавата на повеќеклеточни алги Cryptose Proterozoic 2600 2000 Појавата на еукариоти. Вообичаени се главно едноклеточни зелени алги. Појавата на повеќеклеточност. Појава на повеќеклеточна животинска разновидност (појава на сите видови безрбетници) Archaea 3500 1500 Првите траги на живот на Земјата се бактериите и цијанобактериите. Појавата на фотосинтеза
Бактериите апсорбирани од клетката не можеле да се сварат, но останале живи и продолжиле да функционираат. Се верува дека митохондриите потекнуваат од виолетови бактерии кои ја изгубиле способноста за фотосинтеза и преминале на оксидација на органски материи. Симбиозата со други фотосинтетички клетки доведе до појава на пластиди во растителните клетки. Веројатно, флагелите на еукариотските клетки настанале како резултат на симбиоза со бактерии, кои, како и современите спирохети, биле способни да превиткуваат движења. Отпрвин, наследниот апарат на еукариотските клетки беше структуриран приближно на ист начин како оној на прокариотите, а дури подоцна, поради потребата да се контролира голема и сложена клетка, беа формирани хромозоми. Геномите на интрацелуларните симбионти (митохондрии, пластиди и флагели) генерално ја задржаа прокариотската организација, но повеќето од нивните функции беа пренесени во нуклеарниот геном.
Еукариотските клетки се појавуваа постојано и независно една од друга. На пример, црвените алги настанале како резултат на симбиогенезата со цијанобактериите, а зелените алги со прохлорофитните бактерии.
Останатите едномембрански органели и јадрото на еукариотската клетка, според ендомембранската теорија, настанале од инвагинациите на мембраната на прокариотската клетка.
Точното време на појавата на еукариотите е непознато, бидејќи веќе во седименти стари околу 3 милијарди години има отпечатоци од клетки со слични големини. Еукариотите дефинитивно се евидентирани во карпи стари околу 1,5-2 милијарди години, но само по револуцијата на кислородот (пред околу 1 милијарда години) се создале услови поволни за нив.
На крајот на протерозојската ера (пред најмалку 1,5 милијарди години), веќе постоеле повеќеклеточни еукариотски организми. Повеќеклеточната клетка, како и еукариотската клетка, постојано се појавува кај различни групи на организми.
Постојат различни гледишта за потеклото на повеќеклеточните животни. Според некои податоци, нивните предци биле повеќејадрени, слични на цилијатни клетки, кои потоа се распаднале во посебни мононуклеарни клетки.
Други хипотези го поврзуваат потеклото на повеќеклеточните животни со диференцијацијата на колонијалните едноклеточни клетки. Разликите меѓу нив се однесуваат на потеклото на клеточните слоеви кај првобитното повеќеклеточно животно. Според хипотезата за гастреа на Е. Спротивно на тоа, I. I. Mechnikov ја формулирал хипотезата за фагоцитела, сметајќи дека предците на повеќеклеточните организми се еднослојни сферични колонии како Volvox, кои ги апсорбирале честичките од храната со фагоцитоза. Клетката што ја фатила честичката го изгубила флагелумот и се преселила подлабоко во телото, каде што вршела варење, а на крајот од процесот се вратила на површината. Со текот на времето, клетките беа поделени на два слоја со специфични функции - надворешниот обезбедуваше движење, а внатрешниот обезбедуваше фагоцитоза. I. I. Mechnikov го нарече таквиот организам фагоцитела.
Долго време, повеќеклеточните еукариоти губеа во конкуренција на прокариотски организми, но на крајот на протерозоикот (пред 800-600 милиони години) поради остра промена во условите на Земјата - намалување на нивото на морето, зголемување на концентрацијата на кислород , намалување на концентрацијата на карбонати во морската вода, редовни циклуси на ладење - повеќеклеточните еукариоти добија предности во однос на прокариотите. Ако до овој момент беа пронајдени само поединечни повеќеклеточни растенија и, можеби, габи, тогаш од овој момент во историјата на Земјата беа познати и животните. Од фауните што се појавија на крајот на протерозоикот, Едијакаран и Вендијан се најдобро проучени. Животните од вендискиот период обично се вклучени во посебна група на организми или класифицирани како типови како што се колентерати, рамни црви, членконоги итн. Сепак, ниту една од овие групи нема скелети, што може да укажува на отсуство на предатори.
Развој на животот во палеозојската ера.Палеозојската ера, која траела повеќе од 300 милиони години, е поделена на шест периоди: камбриски, ордовициан, силуриски, девонски, карбон (јаглероден) и перм.
ВО Камбриски периодЗемјата се состоеше од неколку континенти, лоцирани главно на јужната хемисфера. Најзастапени фотосинтетички организми во овој период биле цијанобактериите и црвените алги. Во водениот столб живееле фораминифери и радиолари. Во Камбриј, се појавуваат огромен број скелетни животински организми, за што сведочат бројните фосилни остатоци. Овие организми припаѓале на приближно 100 видови повеќеклеточни животни, и модерни (сунѓери, колентерати, црви, членконоги, мекотели) и исчезнати, на пример: огромниот предатор Anomalocaris и колонијалните граптолити кои лебделе во водната колона или биле прикачени на дното. Земјата остана речиси ненаселена во текот на Камбриј, но процесот на формирање на почва веќе започна со бактерии, габи и, веројатно, лишаи, а на крајот на периодот, на копното се појавија олигохетни црви и милипеди.
ВО Ордовикијан периодНивото на водата на Светскиот океан се зголеми, што доведе до поплави на континенталните низини. Главните производители во овој период биле зелените, кафените и црвените алги. За разлика од Камбрискиот, во кој гребените биле изградени со сунѓери, во Ордовичанецот тие биле заменети со корални полипи. Цветаа гастроподите и цефалоподите, како и трилобитите (сега изумрени роднини на арахнидите). Во овој период, за прв пат се снимени и акордати, особено оние без вилица. На крајот на Ордовикијан, се случи голем настан на истребување, кој уништи околу 35% од семејствата и повеќе од 50% од родовите на морски животни.
Силуријасе карактеризира со зголемена планинска градба, што доведе до сушење на континенталните платформи. Водечката улога во безрбетната фауна на Силурија ја имаа цефалоподите, ехинодермите и џиновските ракови скорпии, додека меѓу 'рбетниците останаа голема разновидност на животни без вилица и се појавија риби. На крајот на периодот, првите васкуларни растенија дојдоа на копно - ринофити и ликофити, кои почнаа да ги колонизираат плитките води и плимната зона на бреговите. На копно дојдоа и првите претставници на класата на арахнидите.
ВО Девонски периодКако резултат на издигнувањето на копното, се формирале големи плитки води, кои се исушиле, па дури и замрзнале, бидејќи климата станала уште поконтинентална отколку во Силурија. Во морињата доминираат корали и ехинодерми, додека цефалоподите се претставени со спирално искривени амонити. Кај 'рбетниците на Девон, рибите процветаа, а 'рскавичните и коскените риби, како и белодробните и лобусните перки, ги заменија оклопните. На крајот на периодот се појавуваат првите водоземци, кои прво живееле во вода.
Во средниот Девон, на копно се појавија првите шуми од папрат, мов и коњско опавче, кои беа населени со црви и бројни членконоги (стоногалки, пајаци, скорпии, инсекти без крилја). На крајот на Девон, се појавија првите гимносперми. Развојот на земјиштето од страна на растенијата доведе до намалување на атмосферските влијанија и зголемено формирање на почва. Консолидацијата на почвите доведе до формирање на речни канали.
ВО Карбонски периодземјата беше претставена со два континента разделени со океан, а климата стана значително потопла и повлажна. До крајот на периодот, имаше мало издигнување на копното, а климата се промени во поконтинентална. Во морињата доминирале фораминифери, корали, ехинодерми, 'рскавици и коскени риби, а во свежите водни тела живееле двовалвни мекотели, ракови и разни водоземци. Во средината на јаглеродот, се појавија мали влекачи инсективозни, а меѓу инсектите се појавија крилести (лебарки, вилински коњчиња).
Тропските предели се карактеризираа со мочурливи шуми во кои доминираа џиновски коњски опашки, мовови и папрати, чии мртви остатоци последователно формираа наоѓалишта на јаглен. Во средината на периодот во умерената зона, благодарение на нивната независност од вода за време на процесот на оплодување и присуството на семиња, започна ширењето на гимносепермите.
Пермски периодсе одликуваше со спојување на сите континенти во еден суперконтинент Пангеа, повлекување на морињата и зајакнување на континенталната клима до таа мера што пустините се формираа во внатрешноста на Пангеа. До крајот на периодот, папратите на дрвјата, коњските опашки и мововите речиси исчезнаа на копното, а гимносепермите отпорни на суша зазедоа доминантна позиција. И покрај фактот дека големите водоземци сè уште продолжија да постојат, се појавија различни групи на влекачи, вклучително и големи тревопасни животни и предатори. На крајот на Перм, се случи најголемиот настан на истребување во историјата на животот, бидејќи исчезнаа многу групи на корали, трилобити, повеќето цефалоподи, риби (првенствено риби со 'рскавици и лобуси перки) и водоземци. Морската фауна изгуби 40-50% од семејствата и околу 70% од родовите.
Развој на животот во мезозоикот.Мезозојската ера траела околу 165 милиони години и се карактеризирала со издигнување на земјиштето, интензивна планинска градба и намалување на влажноста на климата. Поделен е на три периоди: тријаски, јура и креда.
Прво Тријасски периодКлимата била сува, но подоцна, поради покачувањето на нивото на морето, таа станала повлажна. Помеѓу растенијата преовладувале гимносперми, папрати и коњски опавчиња, но дрвенестите форми на спори речиси целосно изумреле. Некои корали, амонити, нови групи на фораминифери, бивалви и ехинодерми достигнаа висок развој, додека разновидноста на 'рскавичните риби се намали, а групите на коскени риби исто така се променија. Рептилите кои доминираа на земјата почнаа да ја совладаат водната средина, како ихтиосаурусите и плезиосаурусите. Од рептилите на Тријас, крокодилите, туатариите и желките преживеале до ден-денес. На крајот на Тријас се појавија диносауруси, цицачи и птици.
ВО Јура периодСуперконтинентот Пангеа се подели на неколку помали. Голем дел од Јура беше многу влажен, а кон крајот климата стана посува. Доминантна група на растенија биле гимносперми, од кои од тоа време преживеале црвените дрва. Во морињата цветале мекотели (амонити и белемнити, бивалви и гастроподи), сунѓери, морски ежови, рскавици и коскени риби. Големите водоземци речиси целосно изумреле во периодот Јура, но се појавиле модерни групи водоземци (опашка и без опашка) и сквамати (гуштери и змии), а разновидноста на цицачите се зголемила. До крајот на периодот се појавија и можни предци на првите птици - Археоптерикс. Сепак, во сите екосистеми доминираа рептилите - ихтиосаурусите и плезиосаурусите, диносаурусите и летечките гуштери - птеросаурусите.
Креда периодго доби своето име поради формирањето на креда во седиментните карпи од тоа време. Низ Земјата, освен поларните предели, имаше постојана топла и влажна клима. Во овој период, ангиоспермите се појавија и станаа широко распространети, поместувајќи ги гимноспермите, што доведе до нагло зголемување на разновидноста на инсектите. Во морињата, покрај мекотелите, коскените риби и плезиосаурусите, повторно се појавија огромен број фораминифери, чии лушпи ги формираа наслагите од креда, а на копното преовладуваа диносаурусите. Птиците подобро прилагодени на воздухот почнаа постепено да ги поместуваат летечките диносауруси.
На крајот на периодот, се случи глобално истребување, што резултираше со исчезнување на амонити, белемнити, диносауруси, птеросауруси и морски гуштери, древни групи птици, како и некои гимносперми. Општо земено, околу 16% од семејствата и 50% од животинските родови исчезнале од лицето на Земјата. Кризата во доцната креда се припишува на влијанието на голем метеорит во Мексиканскиот залив, но најверојатно тоа не беше единствената причина за глобалните промени. За време на последователното ладење, преживеале само мали влекачи и топлокрвни цицачи.
Развој на животот во кенозоикот.Кенозојската ера започнала пред околу 66 милиони години и продолжува до денес. Се карактеризира со доминација на инсекти, птици, цицачи и ангиосперми. Кенозоикот е поделен на три периоди - Палеоген, Неоген и Антропоцен - од кои последниот е најкраткиот во историјата на Земјата.
Во раниот и средниот палеоген, климата останала топла и влажна кон крајот на периодот станала поладна и посува. Ангиоспермите станаа доминантна група растенија, меѓутоа, ако на почетокот на периодот доминираа зимзелени шуми, тогаш на крајот се појавија многу листопадни шуми, а степите се формираа во сушните зони.
Меѓу рибите, коскените риби заземаа доминантна позиција, а бројот на 'рскавичните видови, и покрај нивната забележлива улога во солените водни тела, е незначителен. На копно, преживеале само лушпести влекачи, крокодили и желки, додека цицачите го окупирале најголемиот дел од нивните еколошки ниши. Во средината на периодот се појавиле главните редови на цицачи, вклучувајќи инсективојади, месојади, перчези, китоми, копитари и примати. Изолацијата на континентите ја направи фауната и флората географски поразновидна: Јужна Америка и Австралија станаа центри за развој на торбари, а другите континенти - за плацентарните цицачи.
Неоген период.Во неогенот, површината на земјата го добила својот модерен изглед. Климата стана поладна и посува. Во неогенот, сите редови на современи цицачи веќе биле формирани, а во африканските обвивки се појавиле семејството Хоминиди и родот Човечки. До крајот на периодот, зимзелени шуми се шират во поларните региони на континентите, се појавија тундри, а житарките ги окупираа умерените степи.
Кватернарен период(антропоцен) се карактеризира со периодични промени на глацијации и затоплувања. За време на глацијациите, високите географски широчини биле покриени со глечери, нивото на океаните нагло се намалило, а тропските и суптропските зони се стеснуваат. Во областите блиску до глечерите, беше воспоставена студена и сува клима, што придонесе за формирање на ладно отпорни групи на животни - мамути, џиновски елени, пештерски лавови итн. процесот на глацијација доведе до формирање на копнени мостови меѓу Азија и Северна Америка, Европа и Британските острови итн. реликти од вонземјани, на пример, торбари и копитари во Јужна Америка. Овие процеси, сепак, не ја погодија Австралија, која остана изолирана.
Општо земено, периодичните климатски промени доведоа до формирање на екстремно изобилна разновидност на видови, карактеристични за сегашната фаза на еволуцијата на биосферата, а исто така влијаеа и на човечката еволуција. За време на антропоценот, неколку видови од родот Човечки се прошириле од Африка до Евроазија. Пред околу 200 илјади години во Африка се појавил видот Хомо сапиенс, кој, по долг период на постоење во Африка, пред околу 70 илјади години влегол во Евроазија и пред околу 35-40 илјади години - во Америка. По период на коегзистенција со тесно сродни видови, ги помести и се рашири низ целиот свет. Пред околу 10 илјади години, човечката економска активност во умерено топлите региони на земјината топка почна да влијае и на изгледот на планетата (орање земјишта, палење шуми, прекумерно пасење на пасишта, опустинување итн.) и на животинскиот и растителниот свет поради до намалување на живеалиштата нивното живеалиште и истребување, а во игра стапи и антропогениот фактор.
Човечко потекло. Човекот како вид, неговото место во системот на органскиот свет. Хипотези за човечко потекло. Движечки сили и фази на човековата еволуција. Човечки раси, нивната генетска поврзаност. Биосоцијална природа на човекот. Социјална и природна средина, човечка адаптација кон неа
Човечко потекло
Пред само 100 години, огромното мнозинство луѓе на планетата не ни помислуваа дека луѓето можат да потекнуваат од такви „непочитувани“ животни како мајмуните. Во дискусија со еден од бранителите на Дарвиновата теорија за еволуција, професорот Томас Хаксли, неговиот жесток противник, бискупот од Оксфорд Семјуел Вилберфорс, кој се потпираше на религиозна догма, дури го праша дали се смета себеси за поврзан со предците мајмуни преку неговиот дедо или баба .
Сепак, размислувањата за еволутивното потекло ги изразиле античките филозофи, а големиот шведски таксономист C. Linnaeus во 18 век, врз основа на збир на карактеристики, му дал име на видот на човекот. Хомо сапиенс Л.(Хомо сапиенс) и го класифицираше, заедно со мајмуните, во истиот ред - Примати. Ј.Б. Ламарк го поддржувал К.Линеус и верувал дека човекот дури имал заеднички предци со модерните мајмуни, но во одреден момент од својата историја потекнува од дрвото, што било една од причините за појавата на човекот како вид.
Чарлс Дарвин, исто така, не го игнорираше ова прашање и во 70-тите години на 19 век ги објави делата „Потеклото на човекот и сексуалната селекција“ и „За изразување на емоции кај животните и човекот“, во кои даде подеднакво убедливи докази за заедничкото потекло на луѓето и мајмуните, отколку германскиот истражувач Е. . Сепак, овие студии се однесуваа само на биолошката страна на формирањето на човекот како вид, додека социјалните аспекти беа откриени од класикот на историскиот материјализам - германскиот филозоф Ф. Енгелс.
Во моментов, науката ги проучува потеклото и развојот на луѓето како биолошки вид, како и различноста на современите човечки популации и моделите на нивната интеракција. антропологијата.
Човекот како вид, неговото место во системот на органскиот свет
Хомо сапиенс ( Хомо сапиенс) како биолошки вид припаѓа на животинското царство, поткралството на повеќеклеточни организми. Присуството на нотокорд, жабрени процепи во фаринксот, неврална туба и билатерална симетрија за време на ембрионалниот развој овозможуваат да се класифицира како хордат, додека развојот на 'рбетот, присуството на два пара екстремитети и локацијата на срцето на вентралната страна на телото укажуваат на нејзината врска со другите претставници на подтипот на 'рбетници.
Хранење на младите со млеко што се излачува од млечните жлезди, топлокрвност, четирикоморно срце, присуство на влакна на површината на телото, седум пршлени во цервикалниот 'рбет, предворјето на устата, алвеоларни заби и замена. млечните заби со трајните се знаци на класата на цицачи, а интраутериниот развој на ембрионот и неговата поврзаност со телото на мајката преку плацентата - подкласа на плацентата.
Поспецифични карактеристики, како што се фаќање на екстремитетите со спротивставен палец и нокти, развој на клучната коска, очи насочени напред, зголемување на големината на черепот и мозокот, како и присуство на сите групи заби (секачи, кучешки и катници) не оставаат сомнеж дека неговото место е во редот на примати.
Значителниот развој на мозокот и мускулите на лицето, како и структурните карактеристики на забите, овозможуваат да се класифицираат луѓето како членови на подредот на повисоките примати или мајмуни.
Отсуството на опашка, присуството на искривувања на 'рбетот, развојот на церебралните хемисфери на предниот мозок, покриени со кортекс со бројни жлебови и конволуции, присуството на горната усна и ретка линија на косата даваат основа да се стави меѓу претставници на фамилијата на големите мајмуни, или големите мајмуни.
Сепак, дури и од најорганизираните мајмуни, луѓето се одликуваат со нагло зголемување на волуменот на мозокот, исправено држење, широка карлица, испакната брада, артикулиран говор и присуство на 46 хромозоми во кариотипот и ја одредуваат неговата припадност на родот Human.
Користењето на горните екстремитети за работа, производството на алатки, апстрактното размислување, колективната активност и развојот заснован на повеќе социјални отколку биолошки закони се специфичните карактеристики на хомо сапиенсот.
Сите модерни луѓе припаѓаат на еден вид - хомо сапиенс ( Хомо сапиенс), и подвидови H. sapiens sapiens. Овој вид е збир на популации кои произведуваат плодни потомци кога се вкрстуваат. И покрај прилично значајната разновидност на морфофизиолошките карактеристики, тие не се доказ за повисок или понизок степен на организација на одредени групи луѓе - сите тие се на исто ниво на развој.
Во нашево време, веќе се собрани доволен број научни факти во интерес на формирањето на човекот како вид во процесот на еволуција - антропогенеза. Специфичниот тек на антропогенезата сè уште не е целосно разбран, но благодарение на новите палеонтолошки наоди и современите методи на истражување, можеме да се надеваме дека доволно брзо ќе се појави јасна слика.
Хипотези за човечко потекло
Ако не ги земеме предвид хипотезите за божественото создавање на човекот и неговото навлегување од други планети кои не се поврзани со областа на биологијата, тогаш сите повеќе или помалку конзистентни хипотези за потеклото на човекот го трагаат до заедничките предци со модерни примати.
Значи, хипотеза за човечко потекло од античкиот тропски примати тарсиер, или тарсијална хипотеза, формулиран од англискиот биолог Ф. Вуд Џонс во 1929 година, се заснова на сличноста на пропорциите на телото на луѓето и тарсиерот, карактеристиките на линијата на косата, скратувањето на фацијалниот дел од черепот на вториот, итн. , разликите во структурата и виталната активност на овие организми се толку големи што не доби универзално препознавање.
Луѓето дури имаат премногу сличности со мајмуните. Така, покрај анатомските и морфолошките карактеристики веќе споменати погоре, треба да се обрне внимание на нивниот постембрионски развој. На пример, малите шимпанза имаат многу поретка коса, односот на волуменот на мозокот и волуменот на телото е многу поголем, а способноста за движење на задните екстремитети е нешто поширока отколку кај возрасните. Дури и пубертетот кај повисоките примати се јавува многу подоцна отколку кај претставниците на другите редови на цицачи со слични димензии на телото.
Цитогенетските студии открија дека еден од човечките хромозоми е формиран како резултат на фузија на хромозоми од два различни пара присутни во кариотипот на големите мајмуни, а тоа ја објаснува разликата во бројот на нивните хромозоми (кај луѓето 2n = 46, и кај големите мајмуни 2n = 48 ), а исто така е уште еден доказ за поврзаноста на овие организми.
Сличноста меѓу луѓето и мајмуните е исто така многу висока според молекуларните биохемиски податоци, бидејќи луѓето и шимпанзата имаат исти протеини од крвните групи AB0 и Rh, многу ензими, а низите на аминокиселините на синџирите на хемоглобин имаат само 1,6% разлики, додека со другите мајмуни ова е нешто повеќе несовпаѓање. И на генетско ниво, разликите во нуклеотидните секвенци во ДНК помеѓу овие два организми се помали од 1%. Ако ја земеме предвид просечната стапка на еволуција на таквите протеини во сродните групи на организми, можеме да утврдиме дека човечките предци се одвоиле од другите групи примати пред околу 6-8 милиони години.
Однесувањето на мајмуните на многу начини потсетува на човечкото однесување, бидејќи тие живеат во групи во кои социјалните улоги се јасно распределени. Заедничката одбрана, меѓусебната помош и ловот не се единствените цели на создавање група, бидејќи во неа мајмуните доживуваат наклонетост еден кон друг, го изразуваат на секој можен начин и реагираат емотивно на различни стимули. Дополнително, во групи се врши размена на искуства помеѓу поединци.
Така, сличностите меѓу луѓето и другите примати, особено големите мајмуни, се наоѓаат на различни нивоа на биолошка организација, а разликите меѓу луѓето како вид во голема мера се одредени од карактеристиките на оваа група цицачи.
Групата хипотези кои не го доведуваат во прашање потеклото на луѓето од заедничките предци со модерните мајмуни ги вклучуваат хипотезите за полицентризам и моноцентризам.
Почетна позиција хипотези за полицентризаме појавата и паралелната еволуција на современиот човечки вид во неколку региони на земјината топка од различни форми на антички или дури антички човек, но тоа е во спротивност со основните одредби на синтетичката теорија на еволуцијата.
Хипотезите за единственото потекло на современиот човек, напротив, го постулираат појавувањето на човекот на едно место, но се разликуваат од тоа каде се случило тоа. Значи, хипотеза за екстратропско потекло на луѓетосе заснова на фактот дека само суровите климатски услови на високите географски широчини на Евроазија би можеле да придонесат за „хуманизација“ на мајмуните. Тоа беше поддржано од откривањето на територијата на Јакутија на локалитети кои датираат од античкиот палеолит - културата Диринг, но подоцна беше утврдено дека возраста на овие наоди не е 1,8-3,2 милиони години, туку 260-370 илјади години. Така, оваа хипотеза исто така не е доволно потврдена.
Во моментов се собрани најголем број докази во корист на хипотези за африканското потекло на луѓето, но тоа не е без свои недостатоци, кои сеопфатен широка хипотеза за моноцентризам, комбинирајќи ги аргументите на хипотезите за полицентризам и моноцентризам.
Движечки сили и фази во човечката еволуција
За разлика од другите претставници на животинскиот свет, човекот во процесот на својата еволуција бил изложен не само на биолошките фактори на еволуцијата, туку и на социјалните, што придонело за појава на еден вид квалитативно нови суштества со биосоцијални својства. Социјалните фактори утврдија пробив во фундаментално нова адаптивна средина, која обезбеди огромни предности за опстанокот на човечката популација и нагло го забрза темпото на нејзината еволуција.
Биолошките фактори на еволуцијата кои играат одредена улога во антропогенезата до денес се наследната варијабилност, како и протокот на гени кои го снабдуваат примарен материјал за природна селекција. Во исто време, изолацијата, популационите бранови и генетскиот нанос речиси целосно го изгубија своето значење како резултат на научниот и технолошкиот напредок. Ова дава причина за некои научници да веруваат дека во иднина дури и минималните разлики меѓу претставниците на различни раси ќе исчезнат поради нивното мешање.
Бидејќи променливите услови на животната средина ги принудиле човечките предци да се спуштат од дрвјата во отворен простор и да се движат на два екстремитети, ослободените горни екстремитети ги користеле за носење храна и деца, како и за правење и користење алатки. Сепак, таква алатка може да се направи само ако има јасна идеја за конечниот резултат - сликата на објектот, поради што се разви и апстрактното размислување. Добро е познато дека сложените движења и процесот на размислување се неопходни за развој на одредени области на церебралниот кортекс, што се случи во процесот на еволуција. Сепак, невозможно е да се наследат такви знаења и вештини, тие можат да се пренесат само од еден на друг во текот на животот на вториот, што резултираше со создавање на посебна форма на комуникација - артикулиран говор.
Така, општествените фактори на еволуцијата вклучуваат човечка работна активност, апстрактно размислување и артикулиран говор. Не треба да се отфрлаат манифестациите на алтруизам на примитивниот човек, кој се грижел за деца, жени и стари лица.
Работната активност на човекот не само што влијаеше на неговиот изглед, туку и овозможи, во почетокот, делумно да се олеснат условите за постоење преку употреба на оган, производство на облека, изградба на станови, а подоцна активно да се менуваат преку расчистување на шумите. орање земјиште, итн. Во наше време, неконтролираната економска активност го стави човештвото под закана од глобална катастрофа како резултат на ерозија на почвата, сушење на слатководните тела и уништување на озонската обвивка, што, пак, може да се зголеми притисокот на биолошките фактори на еволуцијата.
Дриопитекус, кој живеел пред околу 24 милиони години, најверојатно бил заедничкиот предок на луѓето и мајмуните. И покрај тоа што се качуваше на дрвја и трчаше на сите четири екстремитети, можеше да се движи на две нозе и да носи храна во рацете. Целосното раздвојување на големите мајмуни и линијата што води до луѓето се случило пред околу 5-8 милиони години.

Австралопитекус. Родот очигледно потекнува од Dyopithecus Ардипитекус, кој се формирал пред повеќе од 4 милиони години во саваните во Африка како резултат на ладење и повлекување на шумите, што ги принудило овие мајмуни да се префрлат на одење на задните екстремитети. Ова мало животно очигледно родило прилично голем род Австралопитекус(„јужен мајмун“).
Австралопитекусот се појавил пред околу 4 милиони години и живеел во африкански савани и суви шуми, каде што целосно се чувствувале предностите на движењето со две нозе. Од Australopithecus дојдоа две гранки - големи тревопасни животни со моќни вилици Парантропи помали и помалку специјализирани Луѓе. Во одреден временски период, овие два рода се развиваа паралелно, што, особено, се манифестираше во зголемување на волуменот на мозокот и компликација на користените алатки. Карактеристики на нашиот род се производството на камени алатки (Paranthropus користел само коска) и релативно голем мозок.
Првите претставници на човечкиот род се појавија пред околу 2,4 милиони години. Тие припаѓале на видот вешт човек (Хомо хабилис)и беа кратки суштества (околу 1,5 m) со волумен на мозокот од приближно 670 cm 3. Тие користеле сурови алатки од камче. Очигледно, претставниците на овој вид имале добро развиени изрази на лицето и рудиментиран говор. Хомо хабилис ја напушти историската сцена пред околу 1,5 милиони години, предизвикувајќи го следниот вид - прав човек.
Човек исправен (H. еректус) како биолошки вид формиран во Африка пред околу 1,6 милиони години и постоел 1,5 милиони години, брзо се населил на огромни територии во Азија и Европа. Претставник на овој вид од островот Јава некогаш беше опишан како Питекантроп(„човек-мајмун“), откриен во Кина, беше именуван Синантропа, додека нивниот европски „колега“ е Човек од Хајделберг.
Сите овие форми се нарекуваат и архантропи(од антички луѓе). Човекот во исправена положба се одликуваше со ниско чело, големи гребени на веѓите и брадата наведната наназад неговиот волумен на мозокот беше 900-1200 cm 3. Торзото и екстремитетите на исправениот маж личат на оние на современиот човек. Без сомнение, претставниците на овој род користеле оган и правеле секири со две острици. Како што покажаа неодамнешните откритија, овој вид дури и ја совладал навигацијата, бидејќи неговите потомци биле пронајдени на оддалечените острови.
Палеоантроп.Пред околу 200 илјади години настанал човекот од Хајделберг Неандерталец (H. neandertalensis), кој се однесува палеоантропи(антички луѓе) кои живееле во Европа и Западна Азија пред помеѓу 200 и 28 илјади години, вклучително и за време на глацијационите периоди. Тие беа силни, физички прилично силни и издржливи луѓе со голем мозочен капацитет (дури и поголем од оној на современите луѓе). Тие имаа артикулиран говор, правеа сложени алатки и облека, ги закопуваа своите мртви, а можеби дури и имаа некои зачетоци на уметноста. Неандерталците не биле предци на хомо сапиенс оваа група се развивала паралелно. Нивното исчезнување е поврзано со исчезнувањето на мамутската фауна по последната глацијација, а можеби е и резултат на конкурентното поместување од нашиот вид.
Најстариот наод на претставник хомо сапиенс (Хомо сапиенс)Стара е 195 илјади години и потекнува од Африка. Најверојатно, предците на современите луѓе не се неандерталци, туку некоја форма на архантропи, како што е човекот од Хајделберг.
Неоантроп.Пред околу 60 илјади години, како резултат на непознати настани, нашиот вид речиси исчезнал, па сите следни луѓе се потомци на мала група која броела само неколку десетици единки. Откако ја надминавме оваа криза, нашиот вид почна да се шири низ Африка и Евроазија. Се разликува од другите видови по својата послаба фигура, поголема стапка на репродукција, агресивност и, се разбира, најкомплексното и најфлексибилното однесување. Се нарекуваат модерните луѓе кои ја населиле Европа пред 40 илјади години Кромањонии се однесуваат на неоантропи(на современите луѓе). Биолошки не се разликуваа од современите луѓе: висина 170-180 см, волумен на мозокот околу 1600 см 3. Кромањоните развиле уметност и религија, припитомиле многу видови диви животни и одгледувале многу видови растенија. Современите луѓе потекнуваат од Кромањоните.
Човечки раси, нивната генетска поврзаност
Како што човештвото се населило низ планетата, се појавиле одредени разлики помеѓу различни групи луѓе во однос на бојата на кожата, цртите на лицето, типот на косата, како и зачестеноста на појавата на одредени биохемиски карактеристики. Збирот на такви наследни карактеристики карактеризира група на поединци од ист вид, чии разлики се помалку значајни од подвидовите - раса.
Проучувањето и класификацијата на расите е комплицирано поради недостатокот на јасни граници меѓу нив. Целото современо човештво припаѓа на еден вид, во кој се разликуваат три големи раси: австрало-негроид (црна), кавказоидна (бела) и монголоидна (жолта). Секој од нив е поделен на мали раси. Разликите меѓу расите се сведуваат на карактеристиките на бојата на кожата, косата, обликот на носот, усните итн.
Ауси-негроид, или екваторијална тркасе карактеризира со темна боја на кожа, брановидна или кадрава коса, широк и малку испакнат нос, попречни ноздри, дебели усни и голем број кранијални црти. кавкаски, или Евроазиска расасе карактеризира со светла или темна кожа, права или брановидна мека коса, добар развој на машките влакна на лицето (брада и мустаќи), тесен испакнат нос, тенки усни и голем број кранијални црти. Монголоид(азиско-американски) расасе карактеризира со темна или светла кожа, често груба коса, просечна ширина на носот и усните, зарамнето лице, силно испакнување на јаготките, релативно голема големина на лицето, забележлив развој на „третиот очен капак“.
Овие три раси се разликуваат и по нивното населување. Пред ерата на европската колонизација, австрало-негроидната раса беше широко распространета во Стариот свет јужно од тропикот на ракот; Кавкаска раса - во Европа, Северна Африка, Западна Азија и Северна Индија; Монголоидна раса - во Југоисточна, Северна, Централна и Источна Азија, Индонезија, Северна и Јужна Америка.
Сепак, разликите меѓу расите се однесуваат само на мали карактеристики кои имаат адаптивно значење. Така, кожата на негроидите е изгорена со десеткратно поголема доза на ултравиолетово зрачење од кожата на Кавкајците, но Кавкајците страдаат помалку од рахитис на високи географски широчини, каде што може да има недостаток на ултравиолетово зрачење неопходно за формирање на витамин Д.
Претходно, некои луѓе се обидуваа да ја докажат супериорноста на една од расите за да стекнат морална супериорност над другите. Сега е јасно дека расните карактеристики ги рефлектираат само различните историски патишта на групи луѓе, но во никој случај не се поврзани со предноста или биолошка заостанатост на една или друга група. Човечките раси се помалку јасно дефинирани од подвидовите и расите на другите животни и на кој било начин не можат да се споредат, на пример, со раси на домашни животни (кои се резултат на намерна селекција). Како што покажуваат биомедицинските истражувања, последиците од меѓурасните бракови зависат од индивидуалните карактеристики на мажот и жената, а не од нивната раса. Затоа, сите забрани за меѓурасни бракови или одредени суеверија се ненаучни и нехумани.
Поконкретни од расите, групите луѓе се националности- историски формирани јазични, територијални, економски и културни заедници на луѓе. Населението на одредена земја го сочинува нејзиниот народ. Со интеракцијата на многу националности, една нација може да се појави во една нација. Сега на Земјата нема „чисти“ раси, а секоја доволно голема нација е претставена со луѓе кои припаѓаат на различни раси.
Биосоцијална природа на човекот
Несомнено, луѓето како биолошки вид мора да доживеат притисок од еволутивните фактори како што се мутагенезата, популационите бранови и изолацијата. Меѓутоа, како што се развива човечкото општество, некои од нив слабеат, додека други, напротив, се зајакнуваат, бидејќи на планетата, заробена од процесите на глобализацијата, речиси и да не останале изолирани човечки популации во кои се врши вкрстување помеѓу крвни сродници, а бројот од самите популации не подлежат на остри флуктуации. Според тоа, движечкиот фактор на еволуцијата - природната селекција - благодарение на успесите на медицината, повеќе не ја игра истата улога кај човечката популација како кај популациите на другите организми.
За жал, слабеењето на притисокот на селекција доведува до зголемување на зачестеноста на наследни болести кај популацијата. На пример, во индустријализираните земји до 5% од населението страда од далтонизам, додека во помалку развиените земји оваа бројка е до 2%. Негативните последици од овој феномен можат да се надминат благодарение на превентивните мерки и напредокот во областите на науката како што е генската терапија.
Сепак, тоа не значи дека човечката еволуција е завршена, бидејќи природната селекција продолжува да дејствува, елиминирајќи ги, на пример, гамети и поединци со неповолни комбинации на гени дури и во проембрионскиот и ембрионскиот период на онтогенезата, како и отпорноста на патогени од различни болести. Покрај тоа, материјалот за природна селекција се обезбедува не само со процесот на мутација, туку и со акумулација на знаење, способност за учење, перцепција на културата и други карактеристики што можат да се пренесат од човек на човек. За разлика од генетските информации, искуството акумулирано во процесот на индивидуален развој се пренесува и од родителите на потомството и во спротивна насока. А конкуренцијата веќе се јавува меѓу заедниците кои се разликуваат културно. Оваа форма на еволуција, единствена за луѓето, се нарекува културен, или социјална еволуција.
Сепак, културната еволуција не ја исклучува биолошката еволуција, бидејќи таа стана возможна само поради формирањето на човечкиот мозок, а самата човечка биологија моментално е одредена од културната еволуција, бидејќи во отсуство на општество и разновидност на движења, одредени зони не се формираат во мозокот.
Така, едно лице има биосоцијална природа, што остава отпечаток на манифестацијата на биолошки, вклучително и генетски, закони кои управуваат со неговиот индивидуален и еволутивен развој.
Социјална и природна средина, човечка адаптација кон неа
Под социјална срединада ги разбере, пред сè, социјалните, материјалните и духовните услови на неговото постоење и активност околу една личност. Покрај економскиот систем, општествените односи, општествената свест и култура, тој ја вклучува и непосредната околина на една личност - семејството, работните и студентските групи, како и другите групи. Околината, од една страна, има одлучувачко влијание врз формирањето и развојот на личноста, а од друга, самата се менува под влијание на личноста, што повлекува нови промени кај луѓето итн.
Приспособување на поединците или нивните групи кон социјалната средина за остварување на сопствените потреби, интереси, животни цели и вклучува адаптација на условите и природата на студирањето, работата, меѓучовечките односи, еколошката и културната средина, условите за слободно време и секојдневниот живот, како и како нивна активна промена за задоволување на вашите потреби. Голема улога во ова има и промената на себеси, своите мотиви, вредности, потреби, однесување итн.
Информациите и емоционалните искуства во современото општество често се главната причина за стрес, кој може да се надмине со помош на јасна самоорганизација, физичка обука и авто-тренинг. Во некои особено тешки случаи, потребна е посета на психотерапевт. Обидот да се најде заборав на овие проблеми преку прејадување, пушење, пиење алкохол и други лоши навики не доведува до посакуваниот резултат, туку само ја влошува состојбата на телото.
Природната средина нема помало влијание врз луѓето, и покрај фактот што луѓето се обидуваат да создадат удобно вештачко опкружување за себе околу 10 илјади години. Така, искачувањето на значителна надморска височина поради намалување на концентрацијата на кислород во воздухот доведува до зголемување на бројот на црвени крвни зрнца во крвта, зголемено дишење и отчукување на срцето, а продолжената изложеност на отворено сонце придонесува за зголемена пигментација на кожата. - тен. Сепак, наведените промени се вклопуваат во нормата на реакцијата и не се наследни. Меѓутоа, народите кои долго време живееле во такви услови може да имаат одредени прилагодувања. Така, кај северните народи, носните синуси имаат многу поголем волумен за загревање на воздухот, а големината на испакнатите делови од телото се намалува за да се намали загубата на топлина. Африканците имаат потемна боја на кожа и виткана коса бидејќи пигментот меланин ги штити органите на телото од пенетрација на штетните ултравиолетови зраци, а капачето за коса има топлинско изолациони својства. Светлите очи на Европејците се адаптација на поакутна перцепција на визуелните информации во самрак и во магла, а монголоидната форма на очите е резултат на природната селекција за дејство на ветрови и бури од прашина.
Овие промени бараат векови и милениуми, но животот во цивилизирано општество повлекува одредени промени. Така, намалувањето на физичката активност доведува до полесен скелет, намалување на неговата сила и намалување на мускулната маса. Ниската подвижност, вишокот на висококалорична храна, стресот доведуваат до зголемување на бројот на луѓе со прекумерна тежина, а соодветната протеинска исхрана и континуираните дневни часови со помош на вештачко осветлување придонесуваат за забрзување - забрзан раст и пубертет и зголемување на големината на телото .
Сади Карно, син на воениот министер за време на Наполеон Бонапарта и вујко на идниот претседател на Француската Република, анализирајќи ги последиците од освојувачките војни на Франција на почетокот на 19 век, дошол до заклучок дека една од причините бидејќи поразот на Франција беше нејзиното заостанување во користењето на енергијата.
Во последните двесте години, луѓето потрошија многу напор обидувајќи се да го разберат овој концепт. Денес стана јасно дека човековата благосостојба е поврзана со потрошувачката на енергија. На пример, обемот на бруто националниот производ на земјата е речиси пропорционален со обемот на потрошената енергија.
Термодинамиката е студија за тоа како енергијата се претвора во физичките тела. Тој одговара на прашањата: како да се добие топлина, како да се направи процесот на добивање топлина податлив, како да се заштеди и како најдобро да се користи. За да одговорат на нив, научниците воведоа дефиниции и ги формулираа основните закони или принципи на термодинамиката.
Енергија(од грчки енергија- дејство, активност) се дефинира како општа квантитативна мерка на различни форми на движење на материјата.
Во природата, секој физички процес одговара на еден или друг вид енергија: механичка, хемиска, топлинска, гравитациска, електромагнетна, нуклеарна итн. Со други зборови, тој ја изразува способноста на телото или системот да работи.
Фундаменталните истражувања во областа на теоријата на информации доведоа до концептот информациска енергија, или енергија на влијанието на информациите , како квантитативна мерка на промените во количината на информации.
Првиот закон на термодинамиката е накратко формулиран на следниов начин: „Енергијата е зачувана“. Не се појавува од ништо и не исчезнува без трага; може да се менува само од една во друга форма. Ова фундаментално откритие од 50-тите години на минатиот век им го должиме на лордот Келвин (Вилијамс Томсон) и Рудолф Клаузис. Келвин, како религиозен човек, верувал дека Создателот, во времето на создавањето на светот, го обдарил со залиха на енергија и дека овој божествен дар ќе постои засекогаш.
Вториот закон на термодинамиката го утврдува присуството во природата на основната асиметрија, односно еднонасочноста на сите спонтани процеси на дисипација на енергија што се случуваат во него: топлите тела се ладат со текот на времето, но студените не стануваат жешки сами по себе, ротирачки врв. на крајот застанува, но горниот дел од одмор нема да почне спонтано да се ротира.
Односно, дистрибуцијата на достапната енергија се менува неповратно. Ова својство на материјата се рефлектира принципот на ентропија. Ако има термин ентропијашто го карактеризира степенот на неред во природата, што значи дека мора да постои термин кој го дефинира степенот на редот. Енталпијаи е функција која го проценува степенот на редослед на системот (локална хетерогеност).
Природните процеси во природата се придружени со зголемување на ентропијата (нарушување).
На изненадувачки начин, овие идеи се рефлектираат во моделот на Универзумот. Се шири, а енергијата се троши во вселената. Континуираното и глобално природно уништување е главната сопственост на материјалниот свет околу нас. Во овој контекст, она што стои во човечкиот ум зад сликата на Мефистофеле игра важна улога во природата, обезбедувајќи процеси дисипација(од латински дисипација- расејување). Во исто време, поради локалните нехомогености во дистрибуцијата на материјата во природата, таа се акумулира во локалните зони на Универзумот. Така се формираат галаксиите, соѕвездијата, сончевите системи и планетите.
Ако свесно не одржуваме ред во куќата, тогаш, без разлика на нашата желба, во неа ќе се зголеми нередот. Спротивно на тоа, намерната креативна активност е придружена со намалување на ентропијата (нарушување) во одредена локална зона на просторот. Точно, тоа е можно само со создавање уште поголем хаос надвор од оваа зона: вака се раѓаат депониите и другиот отпад - нуспроизводи на креативната активност.
Од практиката знаеме дека креативната активност е невозможна без употреба на разум или некој алгоритам роден од природата. Во овој случај, раѓањето на кристал во аморфна карпа е пример за намерна креативна активност во природата заснована на алгоритам генериран од самата природа.
Секој жив организам е исто така пример за ограничен простор во кој ентропијата не се зголемува за време на животот. За разлика од кристалот, организмот има својство на саморегулација, односно способност да одржува постојана внатрешна средина кога се менуваат надворешните и внатрешните нарушувања. Ова својство да не се зголемува ентропијата во текот на животот е едно од главните што го разликува елементот на живата природа од неживата природа.
Така, одржувањето на редот е поврзано со алгоритам за саморегулација, свест. Расудувајќи во однос на алегориите на Гете, можеме да кажеме дека Создателот е одговорен за процесите поврзани со животот кои не ја зголемуваат ентропијата. Се чини дека тој ја користи материјата како експериментален материјал за да создаде и развие алгоритми кои поддржуваат различни животи во потрага по нивното совршенство.
Бидејќи во пракса често е тешко директно да се измери количината на енергија во телото, луѓето научија да ја оценуваат во однос на количината на работа што може да се направи со неа. Од училишниот курс по физика знаеме дека работата се оценува како производ силата, нанесена на телото, помножена со растојанието по кое се движело телото. На еден или друг начин, концептот на сила во природата е поврзан со видот на интеракцијата.
Како што веќе забележавме, сите познати сили се засноваат на четири главни типови на интеракција на физичките тела (Табела 1.1.1). Кога се разгледуваат динамичките процеси, важно е да се запамети дека секое движење на материјално тело може да се разложи на три главни: транслаторно, осцилаторно и ротационо.
Движењето и силата имаат насоки кои не секогаш се совпаѓаат. Ова несовпаѓање се зема предвид со коефициент пропорционален на аголот помеѓу насоката на силата и насоката на движење. Сега можеме да објасниме дека временскиот часовник, кој виси неподвижен на ѕидот, троши енергија поради осцилацијата на нишалото и другите делови. Оската на врвот е практично неподвижна во однос на подот. Сепак, предниот врв има ротациона енергија и ја троши на надминување на отпорот на воздухот и триењето на оската на подот.
За да се процени стапката на користење на енергија или нејзино стекнување, се користи концептот на моќност како стапка на промена на вкупната енергија на одреден систем.
Така, производот на моќта и времето дава енергија. Моќноста се мери во вати. За една секунда, уред за складирање енергија со моќност од еден вати е способен да испорача еден џул (J) енергија на системот. Еден коњски сили се дефинира како потрошувачка на енергија од 746 J во секунда.
Значи, да ги наведеме основните закони неопходни за разбирање на материјалот во книгата што лежи во основата на процесите што се случуваат во Универзумот (макросветот):
1. Закон за хиерархиска сличност.
2. Законот за ритам.
3. Закон за зачувување на енергијата.
4. Закон за ентропија.
Законите на екологијата— општи обрасци и принципи на интеракција помеѓу човечкото општество и природната средина.
Значењето на овие закони е да се регулира природата и насоката на човековата активност во рамките на екосистемите на различни нивоа. Меѓу законите на екологијата формулирани од различни автори, најпознати се четирите закони-афоризми на американскиот еколошки научник Бери Комонер (1974):
- „Сè е поврзано со сè“(законот на универзалната поврзаност на нештата и појавите во природата);
- „Сè мора да оди некаде“(закон за зачувување на масата на материјата);
- „Ништо не доаѓа бесплатно“(за цената на развојот);
- „Природата најдобро знае“(за главниот критериум на еволутивната селекција).
Од законот за универзалната поврзаност на нештата и појавите во природата(„сè е поврзано со сè“), следуваат неколку последици:
- закон за големи броеви -комбинираното дејство на голем број случајни фактори доведува до резултат кој е речиси независен од случајноста, т.е. кои имаат системска природа. Така, огромен број бактерии во почвата, водата и телата на живите организми создаваат посебна, релативно стабилна микробиолошка средина неопходна за нормално постоење на сите живи суштества. Или друг пример: случајното однесување на голем број молекули во одреден волумен на гас одредува сосема дефинитивни вредности на температура и притисок;
- Принцип Le Chatelier (Браун) -Кога надворешно влијание ќе го извади системот од состојба на стабилна рамнотежа, оваа рамнотежа се поместува во насока во која се намалува ефектот на надворешното влијание. На биолошко ниво се реализира во форма на способност на екосистемите да се саморегулираат;
- закон за оптималност— секој систем работи со најголема ефикасност во одредени просторновременски граници карактеристични за него;
- сите системски промени во природата имаат директно или индиректно влијание врз луѓето - од состојбата на поединецот до сложените општествени односи.
Од закон за зачувување на масата на материјата(„сè мора да оди некаде“), следат најмалку два постулата од практично значење:
Бери Комонер пишува: „... глобалниот екосистем е единствена целина во која ништо не може да се освои или изгуби и која не може да биде предмет на севкупно подобрување; се што е извлечено од него со човечки труд мора да се замени. Плаќањето на оваа сметка не може да се избегне; може само да се одложи. Сегашната еколошка криза значи дека доцнењето е многу долго“.
Принцип „Природата најдобро знае“одредува, пред сè, што може, а што не треба да се случува во биосферата. Сè во природата - од едноставни молекули до луѓе - помина низ жестока конкуренција за правото на постоење. Во моментов, планетата е населена со само 1/1000 видови растенија и животни тестирани со еволуција. Главниот критериум на оваа еволутивна селекција е вклучувањето во глобалниот биотски циклус, пополнување на сите еколошки ниши. Секоја супстанција произведена од организми мора да има ензим што ја разградува, а сите производи на распаѓање мора повторно да се вклучат во циклусот. Со секој биолошки вид што го прекршил овој закон, еволуцијата порано или подоцна се раздели. Човечката индустриска цивилизација грубо ја нарушува затвореноста на биотскиот циклус на глобално ниво, што не може да помине неказнето. Во оваа критична ситуација мора да се најде компромис што може да го направи само човек со интелигенција и желба за тоа.
Покрај формулациите на Бери Комонер, современите екологисти извлекоа уште еден закон за екологија - „Нема доволно за секого“ (законот за ограничени ресурси).Очигледно е дека масата на хранливи материи за сите форми на живот на Земјата е конечна и ограничена. Не е доволно за сите претставници на органскиот свет што се појавуваат во биосферата, затоа значително зголемување на бројот и масата на кој било организам на глобално ниво може да се случи само на сметка на намалување на бројот и масата на другите. Контрадикторноста помеѓу стапката на размножување и ограничените ресурси на храна во однос на популацијата на планетата прв ја забележал англискиот економист Т.Р. Малтус (1798), кој со ова се обиде да ја оправда неизбежноста на општествениот натпревар. За возврат, Чарлс Дарвин го позајмил од Малтус концептот на „борба за постоење“ за да го објасни механизмот на природна селекција во живата природа.
Закон за ограничени ресурси- изворот на сите форми на конкуренција, ривалство и антагонизам во природата и, за жал, во општеството. И колку и да се смета дека класната борба, расизмот и меѓуетничките конфликти се чисто општествени феномени, сите тие се вкоренети во интраспецифична конкуренција, која понекогаш има многу побрутални форми отколку кај животните.
Значајната разлика е во тоа што во природата, како резултат на конкуренцијата, преживуваат најдобрите, но во човечкото општество тоа воопшто не е така.
Генерализирана класификација на законите за животна средина беше претставена од познатиот советски научник Н.Ф. Рајмерс. Им се дадени следниве формулации:
- закон за социо-еколошка рамнотежа(потребата да се одржи рамнотежа помеѓу притисокот врз животната средина и обновувањето на оваа средина, природна и вештачка);
- принцип на управување со развојот на културата(наметнување ограничувања за екстензивен развој, земајќи ги предвид еколошките ограничувања);
- правило за социо-еколошка супституција(потребата да се идентификуваат начини за замена на човековите потреби);
- закон за социо-еколошка неповратност(неможност за враќање на еволутивното движење, од сложени форми во поедноставни);
- закон на ноосфератаВернадски (неизбежноста на трансформацијата на биосферата под влијание на мислата и човечкиот труд во ноосфера - геосфера во која разумот станува доминантен во развојот на системот „човек-природа“).
Усогласеноста со овие закони е можно под услов човештвото да ја разбере својата улога во механизмот за одржување на стабилноста на биосферата. Познато е дека во процесот на еволуција се зачувани само оние видови кои се способни да обезбедат одржливост на животот и животната средина. Само човекот, користејќи ја моќта на својот ум, може да го насочи понатамошниот развој на биосферата по патот на зачувување на дивата природа, зачувување на цивилизацијата и човештвото, создавање поправеден општествен систем, преминување од филозофија на војна во филозофија на мирот и партнерство, љубов и почит кон идните генерации. Сето ова се компоненти на новиот светоглед на биосферата, кој треба да стане универзален.
Закони и принципи на екологијата
Закон за минимум
Во 1840 г Ју Либиготкриле дека жетвата често е ограничена не од оние хранливи материи кои се потребни во големи количини, туку од оние кои се потребни во мали количини, но од кои има малку во почвата. Законот што тој го формулираше вели: „Супстанцијата што е во својот минимум ја контролира жетвата и ја одредува големината и стабилноста на последната со текот на времето“. Последователно, голем број други фактори, како што е температурата, беа додадени на хранливите материи. Дејството на овој закон е ограничено со два принципа. Првиот закон на Либиг строго се применува само под стационарни услови. Попрецизна формулација: „во стационарна состојба, ограничувачката супстанција ќе биде супстанцијата чии достапни количини се најблиску до потребниот минимум“. Вториот принцип се однесува на интеракцијата на факторите. Високата концентрација или достапност на одредена супстанција може да го промени внесот на минимална хранлива материја. Следниот закон е формулиран во самата екологија и го генерализира законот за минимум.
Закон за толеранција
Овој закон е формулиран на следниов начин: отсуството или неможноста за развој на екосистем се определува не само со недостаток, туку и со вишок на кој било од факторите (топлина, светлина, вода). Следствено, организмите се карактеризираат и со еколошки минимум и со максимум. Премногу добра работа е исто така лошо. Опсегот помеѓу двете вредности ги сочинува границите на толеранција во кои телото нормално реагира на влијанијата од околината. Предложен закон за толеранција В. Шелфордво 1913. Може да се формулираат голем број предлози што го надополнуваат.
- Организмите може да имаат широк опсег на толеранција за еден фактор и тесен опсег за друг.
- Организмите со широк опсег на толеранција на сите фактори обично се најраспространети.
- Ако условите за еден фактор на животната средина не се оптимални за еден вид, тогаш опсегот на толеранција на други фактори на животната средина може да се намали.
- Во природата, организмите многу често се наоѓаат во услови кои не одговараат на оптималната вредност на одреден фактор утврден во лабораторија.
- Сезоната на парење е обично критична; Во овој период, многу фактори на животната средина често излегуваат како ограничувачки.
Живите организми ги менуваат условите на животната средина за да го ослабат ограничувачкото влијание на физичките фактори. Видовите со широка географска дистрибуција формираат локално прилагодени популации наречени екотипови.Нивните оптимални и граници на толеранција одговараат на локалните услови.
Општ концепт на ограничувачки фактори
Најважни фактори на копно се светлината, температурата и водата (врнежите), додека во морето светлината, температурата и соленоста. Овие физички услови на постоење можеда биде ограничувачки и позитивно да влијае. Сите фактори на животната средина зависат еден од друг и дејствуваат заедно. Други ограничувачки фактори вклучуваат атмосферски гасови (јаглерод диоксид, кислород) и биогени соли. При формулирањето на „законот за минимум“, Либиг го имал предвид ограничувачкиот ефект на виталните хемиски елементи присутни во животната средина во мали и променливи количини. Тие се нарекуваат елементи во трагови и вклучуваат железо, бакар, цинк, бор, силициум, молибден, хлор, ванадиум, кобалт, јод, натриум. Многу микроелементи, како витамините, делуваат како катализатори. Фосфор, калиум, калциум, сулфур, магнезиум, кои се потребни од организмите во големи количини, се нарекуваат макроелементи. Важен ограничувачки фактор во современите услови е загадувањето на животната средина. Главниот ограничувачки фактор, според Ју Одуму, -големини и квалитет оикоса", или нашите" природен манастир“не само бројот на калории што може да се истиснат од земјата. Пејзажот не е само складиште за намирници, туку и куќата во која живееме. „Целта треба да биде да се зачува најмалку една третина од целото земјиште како заштитен отворен простор. Ова значи дека една третина од целото наше живеалиште треба да бидат национални или локални паркови, природни резервати, зелени површини, диви области итн“. Територијата што му е потребна на едно лице, според различни проценки, се движи од 1 до 5 хектари. Втората од овие бројки ја надминува површината на која моментално отпаѓа еден жител на Земјата.
Густината на населеност се приближува до еден човек на 2 хектари земја. Само 24% од земјиштето е погодно за земјоделство. Иако само 0,12 хектари може да обезбеди доволно калории за поддршка на едно лице, околу 0,6 хектари по лице се потребни за хранлива диета со многу месо, овошје и зеленчук. Дополнително, потребни се уште околу 0,4 хектари за производство на разни видови влакна (хартија, дрво, памук) и уште 0,2 хектари за патишта, аеродроми, згради итн. Оттука и концептот на „златната милијарда“, според кој оптималната големина на населението е 1 милијарда луѓе, и затоа, веќе има околу 5 милијарди „дополнителни луѓе“. За прв пат во својата историја, човекот се соочи со екстремни, а не локални ограничувања. Надминувањето на ограничувачките фактори бара огромни трошоци на материјата и енергијата. Удвојувањето на приносот бара десеткратно зголемување на количината на ѓубрива, пестициди и моќност (животни или машини). Големината на населението е исто така ограничувачки фактор.
Закон за конкурентско исклучување
Овој закон е формулиран на следниов начин: два вида кои заземаат иста еколошка ниша не можат да коегзистираат на едно место неодредено долго време.
Кој вид ќе победи зависи од надворешните услови. Во слични услови, секој може да победи. Важен фактор за победа е стапката на пораст на населението. Неможноста на еден вид да се натпреварува биотски доведува до негово поместување и потреба да се прилагоди на потешки услови и фактори.
Законот за конкурентно исклучување може да функционира и во човечкото општество. Особеноста на неговото дејствување во сегашно време е што цивилизациите не можат да се распрснуваат. Тие немаат каде да ја напуштат својата територија, бидејќи во биосферата нема слободен простор за населување и нема вишок ресурси, што доведува до интензивирање на борбата со сите последици што следуваат. Можеме да зборуваме за еколошки ривалство меѓу земјите, па дури и еколошки војни или војни поради еколошки причини. Едно време, Хитлер ја оправдуваше агресивната политика на нацистичка Германија со борбата за животен простор. Ресурси на нафта, јаглен итн. и тогаш тие беа важни. Тие имаат уште поголема тежина во 21 век. Покрај тоа, имаше потреба од територии за отстранување на радиоактивен и друг отпад. Војните - топли и студени - добиваат еколошка димензија. Многу настани во современата историја, како што е распадот на СССР, се перципираат на нов начин ако се гледаат од еколошка перспектива. Една цивилизација не само што може да освои друга, туку може да ја искористи за себични цели од еколошка гледна точка. Ова ќе биде еколошки колонијализам. Така се испреплетуваат политичките, социјалните и еколошките прашања.
Основен закон на екологијата
Едно од главните достигнувања на екологијата беше откритието дека не се развиваат само организмите и видовите, туку и. Се нарекува редоследот на заедници кои се заменуваат една со друга во дадена област сукцесија.Сукцесијата настанува како резултат на промените во физичката средина под влијание на заедницата, т.е. контролирани од него.
Високата продуктивност дава мала доверливост - уште една формулација на основниот закон за екологија, од која следи следното правило: „Оптималната ефикасност е секогаш помала од максималната“. Различноста, според основниот закон на екологијата, е директно поврзана со одржливоста. Сепак, сè уште не е познато до кој степен оваа врска е причинско-последична.
Некои други закони и принципи важни за екологијата.
Закон за појава: целината секогаш има посебни својства кои нејзиниот дел ги нема.
Закон за потребната разновидност: системот не може да се состои од апсолутно идентични елементи, туку може да има хиерархиска организација и интегративни нивоа.
Закон за неповратност на еволуцијата: организам (популација, вид) не може да се врати во претходната состојба постигната во низата на неговите предци.
Закон за организациска сложеност: историскиот развој на живите организми доведува до усложнување на нивната организација преку диференцијација на органите и функциите.
Биогенетски закон(Е. Хекел): онтогенезата на организмот е кратко повторување на филогенијата на даден вид, т.е. Поединецот во својот развој го повторува, во скратена форма, историскиот развој на својот вид.
Законот за нерамномерен развој на делови од системот: системите од исто ниво на хиерархија не се развиваат строго синхроно, додека некои достигнуваат повисок степен на развој, други остануваат во помалку развиена состојба. Овој закон е директно поврзан со законот за потребната различност.
Закон за зачувување на животот: животот може да постои само во процесот на движење на проток на супстанции, енергија и информации низ живо тело.
Принципот на одржување на уредност(Ј. Пригожин): во отворените системи, ентропијата не се зголемува, туку се намалува додека не се постигне минимална константна вредност, секогаш поголема од нула.
Принципот Ле Шателие-Браун: кога надворешно влијание ќе го извади системот од состојба на стабилна рамнотежа, оваа рамнотежа се поместува во насока во која ефектот на надворешното влијание е ослабен.
Принцип за заштеда на енергија(L. Onsager): со оглед на веројатноста за развој на процес во одреден сет на насоки дозволени со принципите на термодинамиката, се реализира оној кој обезбедува минимум дисипација на енергија.
Закон за енергија и максимизација на информации: најдобра шанса за самоодржување има системот кој најмногу придонесува за примање, производство и ефикасно користење на енергија и информации; максималната понуда на супстанција не гарантира успех на системот во конкуренција.
Законот за развој на системот на сметка на животната средина: секој систем може да се развие само преку употреба на материјални, енергетски и информациски способности на неговата околина; Апсолутно изолиран саморазвој е невозможен.
Шредингеровото правило„За хранење“ на организмот со негативна ентропија: уредноста на организмот е повисока од околината, а организмот дава во оваа средина повеќе неред отколку што прима. Ова правило е во корелација со принципот на Пригожин за одржување на уредност.
Правило за забрзување на еволуцијата: со зголемување на сложеноста на организацијата на биолошките системи, времетраењето на постоењето на еден вид во просек се намалува, а стапката на еволуција се зголемува. Просечниот животен век на видови птици е 2 милиони години, а видот на цицач е 800 илјади години. Бројот на изумрените видови птици и цицачи во споредба со целиот нивен број е голем.
Закон за релативна независност на адаптација: високата приспособливост на еден од факторите на животната средина не обезбедува ист степен на приспособување кон другите животни услови (напротив, може да ги ограничи овие можности поради физиолошките и морфолошките карактеристики на организмите).
Принцип на минимална големина на населението: Постои минимална големина на населението под која нејзиниот број не може да падне.
Правило за претставување на родот по еден вид: во хомогени услови и во ограничена област, таксономскиот род обично е претставен со само еден вид. Очигледно, ова се должи на близината на еколошките ниши на видовите од истиот род.
Законот за исцрпување на живата материја во нејзините островски концентрации(Г.Ф. Хилми): „Индивидуален систем кој работи во средина со ниво на организација пониско од нивото на самиот систем е осуден на пропаст: постепено губијќи ја својата структура, системот ќе се раствори во околината по некое време“. Ова води до важен заклучок за човековите еколошки активности: вештачкото зачувување на екосистемите со мала големина (во ограничена област, на пример, природен резерват) води до нивно постепено уништување и не обезбедува зачувување на видовите и заедниците.
Закон на пирамидата на енергијата(Р. Линдеман): во просек, околу 10% од енергијата добиена на претходното ниво се движи од едно трофичко ниво на еколошката пирамида на друго, повисоко ниво. Обратниот проток од повисоки кон пониски нивоа е многу послаб - не повеќе од 0,5-0,25%, и затоа нема потреба да се зборува за енергетскиот циклус во биоценозата.
Правило за задолжително пополнување на еколошки ниши: празна еколошка ниша е секогаш и секогаш природно исполнета („природата се гнаси од вакуум“).
Принципот на формирање на екосистемот: долгорочното постоење на организмите е можно само во рамките на еколошките системи, каде што нивните компоненти и елементи се надополнуваат и меѓусебно се прилагодуваат. Од овие еколошки закони и принципи произлегуваат некои заклучоци кои важат за системот „човек - природна средина“. Тие припаѓаат на типот на законот за ограничување на различноста, т.е. наметнуваат ограничувања на човековите активности за да се трансформира природата.
Законот за бумеранг: сè што е извлечено од биосферата со човечки труд мора да и се врати.
Законот за незаменливост на биосферата: биосферата не може да се замени со вештачка средина, исто како што, да речеме, не може да се создадат нови видови на живот. Човекот не може да изгради машина за постојано движење, додека биосферата е практично „машина за постојано движење“.
Закон за шагрена кожа: глобалниот првичен потенцијал на природни ресурси континуирано се исцрпува во текот на историскиот развој. Ова произлегува од фактот дека во моментов нема суштински нови ресурси кои би можеле да се појават. На секој човек за живот му се потребни 200 тони цврсти материи годишно, кои ги претвора во корисен производ со помош на 800 тони вода и просечно 1000 W енергија. Човекот сето тоа го зема од она што е веќе достапно во природата.
Принципот на оддалеченост на настанот: потомците ќе смислат нешто за да спречат можни негативни последици. Останува отворено прашањето колку законите на екологијата можат да се пренесат на односот меѓу човекот и животната средина, бидејќи човекот се разликува од сите други видови. На пример, кај повеќето видови, стапката на раст на популацијата се намалува како што се зголемува густината на населението; Кај луѓето, напротив, растот на населението во овој случај се забрзува. Некои регулаторни механизми на природата отсуствуваат кај луѓето, а тоа може да послужи како дополнителна причина за технолошки оптимизам за некои, а за еколошките песимисти, укажува на опасност од таква катастрофа, што е невозможна за кој било друг вид.
Во однос на управувањето со животната средина и активностите за заштита на животната средина, често се слуша за потребата да се земат предвид законите на природниот развој. Човекот, откако ја сфати својата улога во биосферата само како еден од многуте видови кои ја формираат нејзината разновидност, како дел од неа, мора, како и сите други, да ги почитува законите на природата. Во исто време, силата на хомо сапиенсот не лежи во преструктуирањето на природата преку покажување на нејзината моќ, туку во правилното разбирање на законите на нејзиниот развој и следење на нив. Законите на развојот на природата се закони од повисок ред за луѓето во споредба со законите на развојот на општеството. Тоа се објективни закони. По нивната акција и благодарение на нив, човекот се појави и може да постои. Законите на општеството се напишани од човекот за сопствена социјална, политичка и економска погодност, организација и обезбедување на животот во заедницата.
Познавањето и почитувањето на законите на природниот развој во активностите на човекот и општеството е од одлучувачко значење и се оценува како императив. Законите на развојот на природата, манифестирани во интеракцијата на општеството и природата, создаваат природни научни и филозофски основи за различни активности во управувањето со животната средина и заштитата на животната средина, вклучително и во областа на правото. Земањето предвид на законите на природата при планирање и спроведување на еколошки штетни активности и нивната усогласеност треба да послужи како главен критериум за еколошката валидност и допуштеност на таквите активности. Нивното знаење и внимание е особено важно при спроведувањето на таквите законски мерки за заштита на природата како што се стандардизација на максимално дозволените влијанија врз природата, проценка на влијанието на планираните активности врз животната средина, оцена на животната средина, планирање на мерки за зачувување на природата итн. на развојот на природата треба да се води сметка и при подготовката на сметките за заштита на животната средина. Обезбедувањето дека законите на природата се земаат предвид и се почитуваат при донесување економски, менаџерски и други еколошки значајни одлуки е еден од условите, методолошката основа за надминување на еколошката криза.
Ајде да разгледаме некои основни закони за развојот на природата како што ги толкува еден од водечките научници за животна средина во Русија, професорот Н.Ф. Рајмерс.
Закон за биогена миграција на атомите (В.И. Вернадски). Миграцијата на хемиските елементи на површината на земјата и во биосферата како целина се врши или со директно учество на жива материја (биогена миграција) или се јавува во средина чии геохемиски карактеристики (O 2, C0 2, H 2 итн. .) се определени од живата материја - и онаа што моментално ја населува биосферата и онаа што била на Земјата низ геолошката историја.
Според овој Закон, кој има важно теоретско и практично значење, разбирање на општите хемиски процеси кои се случувале и се случуваат на површината на земјиштето, во атмосферата и во длабочините на литосферата и водите во кои живеат организми, како и геолошките слоеви составени од минатите активности на организмите, е невозможно без да се земат предвид биотските и биогените фактори, вклучително и еволутивните. Бидејќи луѓето примарно влијаат на биосферата и нејзиното живо население, тие со тоа ги менуваат условите на биогена миграција на атомите, создавајќи предуслови за уште подлабоки хемиски промени во историската перспектива. Така, процесот може да стане саморазвивачки, независен од човечката желба и практично, на глобално ниво, неконтролиран. Оттука, една од најитните потреби е да се зачува живата површина на Земјата во релативно непроменета состојба. Истиот Закон, исто така, ја одредува потребата да се земат предвид влијанијата врз биотата во сите проекти за трансформација на природата. Во овие случаи се случуваат регионални и локални промени во хемиските процеси, што доведува до какви било големи грешки во деградацијата на животната средина - опустинување.
Закон за внатрешна динамичка рамнотежа. Материјата, енергијата, информациите и динамичките квалитети на поединечните природни системи и нивната хиерархија се толку меѓусебно поврзани што секоја промена на еден од овие индикатори предизвикува придружни функционални структурни квалитативни и квантитативни промени кои го зачувуваат вкупниот збир на материјално-енергетски, информации и динамички квалитети на системи каде што се случуваат овие промени или во нивната хиерархија.
Емпириски се утврдени низа последици од овој закон:
- а) секоја промена во животната средина (супстанции, енергија, информации, динамички квалитети на екосистемите) неизбежно води до развој на природни верижни реакции насочени кон неутрализирање на промената или формирање на нови природни системи, чиешто формирање, со значителни промени во животната средина, може да стане неповратна;
- б) интеракцијата на материјално-енергетските компоненти на животната средина (енергија, гасови, течности итн.), информациите и динамичките квалитети на природните системи не е квантитативно линеарна, т.е. слабото влијание или промена на еден од индикаторите може да предизвика силни отстапувања кај другите (и во целиот систем како целина);
- в) промените произведени во големите екосистеми се релативно неповратни. Поминувајќи низ хиерархијата од дното нагоре - од местото на влијание до биосферата како целина, тие ги менуваат глобалните процеси и со тоа ги пренесуваат на ново еволутивно ниво;
- г) секоја локална трансформација на природата предизвикува одговори во глобалната целина на биосферата и во нејзините најголеми поделби, што доведува до релативна постојаност на еколошкиот и економскиот потенцијал („Тришкиновото кафтано правило“), чие зголемување е можно само преку значително зголемување на инвестициите во енергија.
Законот за внатрешна динамичка рамнотежа е една од клучните одредби во управувањето со животната средина. Додека промените во животната средина се слаби и се прават на релативно мала област, тие се или ограничени на одредено место или „згаснуваат“ во синџирот на хиерархија на екосистемот. Но, штом промените достигнат значајни нивоа за големите екосистеми, на пример, се случуваат на обемот на големите речни сливови, тие доведуваат до значителни поместувања во овие огромни природни формации, а преку нив, во согласност со последицата б), во целата биосфера на Земјата.
Законот „се или ништо“ (Х. Булич). Слабите влијанија може да не предизвикаат одговори во природниот систем додека, откако се акумулираат, не доведат до развој на насилен динамичен процес. Законот е корисен во прогнозирањето на животната средина.
Закон на постојаност (В.И. Вернадски). Количеството на жива материја во природата (за даден геолошки период) е константа. Секоја промена во количината на жива материја во еден од регионите на биосферата неизбежно повлекува промена со иста големина во кој било регион, но со спротивен знак. Поларните промени може да се користат во процесите на управување со природата, но треба да се земе предвид дека не секогаш се случува соодветна замена. Типично, високо развиените видови и екосистеми се заменуваат со други на релативно еволутивно пониско ниво (големите организми со помали), а формите корисни за луѓето се заменуваат со помалку корисни, неутрални или дури и штетни.
Закон за минимум (Ј. Либиг). Издржливоста на еден организам ја одредува најслабата алка во синџирот на неговите еколошки потреби, т.е. можностите за живот се ограничени од фактори на животната средина, чиј квантитет и квалитет се блиску до минимумот што го бара организмот или екосистемот; нивното натамошно намалување доведува до смрт на организмот или уништување на екосистемот.
Закон за ограничени природни ресурси. Сите природни ресурси (и услови) на Земјата се конечни. Законот се заснова на фактот дека бидејќи планетата е природно ограничена целина, на неа не можат да постојат бесконечни делови. Следствено, категоријата „неисцрпни“ природни ресурси се појави поради недоразбирање.
Законот за развој на природниот систем на сметка на неговата околина. Секој природен систем може да се развие само преку употреба на материјални, енергетски и информациски способности на неговата околина. Апсолутно изолиран саморазвој е невозможен. Овој ист Закон има исклучително важно теоретско и практично значење поради неговите главни последици:
- а) апсолутно производството без отпад е невозможно;
- б) кој било повисоко организиран биотски систем (на пример, жив вид), користејќи и менувајќи ја животната средина, претставува потенцијална закана за помалку организираните системи;
- в) Земјината биосфера како систем се развива не само на сметка на ресурсите на планетата, туку индиректно на сметка и под контрола на влијанието на вселенските системи (првенствено Соларниот).
Според првата последица, можеме да сметаме само на производство со низок отпад. Затоа, првата фаза во развојот на технологиите треба да биде нивниот низок интензитет на ресурси (и на влезот и на излезот - економичност и незначителни емисии), втората фаза ќе биде создавање на циклично производство (отпадот на некои може да биде суровина за други) и третата - организација на разумно отстранување на неизбежните остатоци и неутрализирање на неотстранливиот енергетски отпад. Идејата дека биосферата работи на принципот на неотпад е погрешна, бидејќи секогаш акумулира супстанции отстранети од биолошкиот циклус кои формираат седиментни карпи.
Според втората последица од Законот што се разгледува, човечкото влијание врз природата бара мерки за неутрализирање на овие влијанија, бидејќи тие можат да бидат деструктивни за остатокот од природата и, според правилото за усогласеност на условите на животната средина со генетската предодреденост на организмот. , се закануваат на самата личност. Во овој поглед, зачувувањето на природата е една од задолжителните компоненти на социо-економскиот развој на високо развиеното општество.
Третата импликација на Законот е од особено значење за долгорочното прогнозирање. Мора да се земе предвид кога се разгледуваат сите процеси што се случуваат на Земјата.
Закон (шема) за намалување на еколошкиот интензитет на готовите производи. Специфичната содржина на природни супстанции во просечната единица на општествениот производ историски постојано опаѓа. Тоа не значи дека помалку природни супстанции се вклучени во производниот процес. Напротив, неговото количество се зголемува - до 95-98% од природната супстанција што се троши во производството се фрла. Меѓутоа, конечните слични производи на општественото производство денес веројатно содржат, во просек, помалку природна супстанција отколку во далечното минато. Тоа се објаснува со минијатуризацијата на производите, замената на природните материјали и производи со синтетички, како и со други појави. Бидејќи сè уште не се направени точни пресметки (методолошки се тешки), овој Закон има карактер на стручен заклучок.
Законот за намалување на енергетската ефикасност на управувањето со животната средина. Со напредокот на историското време, при добивање корисни производи од природни системи, во просек се троши се повеќе енергија по единица од неа.
Потрошувачката на енергија по лице (во kcal/ден) во камено доба била околу 4 илјади, во земјоделското општество - 12 илјади, во индустриската ера - 7 илјади, а во напредните развиени земји од сегашно време - 230-250 илјади , т.е. 58-62 пати повеќе отколку кај далечните предци. Од почетокот на овој век, количината на енергија потрошена за една единица земјоделски производи во развиените земји е зголемена за 8-10 пати. Целокупната енергетска ефикасност на земјоделското производство (односот на влезната енергија и енергијата добиена од готови производи) во индустријализираните земји е приближно 30 пати помала отколку во условите на примитивното земјоделство. Во некои случаи, десеткратното зголемување на трошоците за енергија за ѓубрива и одгледување на полиња доведува до многу мало (10-15%) зголемување на приносот. Ова се должи на потребата, паралелно со подобрувањето на земјоделската технологија, да се земе предвид општата еколошка состојба и ограничувањата што таа ги наметнува. Во раните 80-ти. Специфичната потрошувачка на енергија по единица бруто национален производ (БНП) е намалена за 15% како резултат на енергични мерки за зачувување на енергијата во индустријализираните земји. Во текот на последната деценија, БНП овде се зголеми за 20%, а потрошувачката на енергија - за само 2% (ова стана возможно како резултат на елиминирање на неоправданите загуби на енергија).
Закон за намалена (природна) плодност. Поради постојаното берење и нарушување на природните процеси на формирање на почвата, како и долготрајната монокултура, како резултат на акумулацијата на отровните материи што ги ослободуваат растенијата, на обработуваните површини се јавува постепено намалување на природната плодност на почвата. Овој процес делумно се неутрализира со акумулација на биомаса на подземните делови од култивирани растенија, но главно со примена на ѓубрива (создавање вештачка плодност). До денес, плодноста е изгубена до еден или друг степен во приближно 50% од сите обработливи површини во светот (од 1,5-1,6 до 2 милијарди хектари) со просечна стапка на загуба во 70-тите години. 6,8, во 80-тите - околу 7 милиони хектари годишно. Интензивирањето на земјоделството овозможува да се добијат сè поголеми приноси со помалку човечки труд и делумно да се неутрализира ефектот на Законот за намалени приноси, но во исто време, енергетската ефикасност на производството се намалува.
Законот за физичко и хемиско единство на живата материја (В.И. Вернадски). Целата жива материја на Земјата е физички и хемиски обединета. Заклучок природно произлегува од Законот: она што е штетно за еден дел од живата материја не може да биде рамнодушно кон друг дел од неа, или: она што е штетно за некои видови суштества е штетно и за другите. Оттука, сите физички и хемиски агенси кои се фатални за некои организми (на пример, средства за контрола на штетници) не можат, а да немаат штетно влијание врз други организми. Единствената разлика е во степенот на отпорност на видот на агентот. Бидејќи во секоја голема популација секогаш има индивидуи со различен квалитет, вклучувајќи ги и оние помалку или повеќе отпорни на физичко-хемиски влијанија, стапката на селекција за издржливост на популациите на штетен агенс е директно пропорционална со брзината на репродукција на организмите и брзината на алтернација на генерации. Врз основа на ова, со растечкото влијание на физичко-хемискиот фактор, на кој организам со релативно бавно менување на генерациите е отпорен, на помалку стабилен, но побрз вид кој се размножува, нивната способност да го издржат предметниот фактор станува еднаква. Затоа долгорочната употреба на хемиски методи за борба против растителните штетници и патогени на луѓето и топлокрвните животни е еколошки неприфатлива. Со изборот на резистентни единки на членконоги кои брзо се размножуваат, стапките на третман треба да се зголемат. Сепак, овие зголемени концентрации се покажаа како неефикасни, но имаат сериозно влијание врз здравјето на луѓето и 'рбетниците.
Закон за еколошка корелација. Во еден екосистем, како и во секоја друга интегрална формација на природен систем, особено во биотска заедница, сите живи видови и абиотски еколошки компоненти вклучени во него се функционално конзистентни едни со други. Губењето на еден дел од системот (на пример, уништувањето на еден вид) неизбежно води до исклучување на сите други делови од системот тесно поврзани со овој дел од системот и функционална промена во целината во рамките на Закон за внатрешна динамичка рамнотежа. Законот за еколошка корелација е особено важен за зачувување на живите видови, кои никогаш не исчезнуваат изолирано, туку секогаш во меѓусебно поврзана група. Ефектот на Законот води до нагли промени во одржливоста на животната средина: кога ќе се достигне прагот на промена на функционалниот интегритет, се случува дефект (често неочекувано) - екосистемот ја губи својата особина на доверливост. На пример, повеќекратното зголемување на концентрацијата на некој загадувач може да не доведе до катастрофални последици, но тогаш незначително зголемување ќе доведе до катастрофа.
Познатиот американски еколошки научник B. Commoner ги сведува основните закони на екологијата на следново: 1) сè е поврзано со сè; 2) сè мора да оди некаде; 3) природата „знае“ подобро; 4) ништо не се дава бесплатно.
Живеалиштето на животот се развива како резултат на постојана размена на информации врз основа на протокот на енергија во вкупното единство на животната средина и организмите што ја населуваат.
40. Закон за минимум(Либиг): Супстанцијата присутна во минимумот го контролира приносот, ја одредува нејзината големина и стабилност со текот на времето.
41. Обичните закони:
„Сè е поврзано со сè“;
„Сè мора да оди некаде“;
„Ништо не доаѓа бесплатно“;
„Природата најдобро знае“.
42. Закон за максимум(Шелфорд):Процутот на еден организам е ограничен со зони на максимум и минимум на одредени фактори на животната средина; меѓу нив постои зона на еколошки оптимум, во која телото нормално реагира на условите на околината.
43. Деградација на биосферата -Ова е уништување или значително нарушување на еколошките врски во природата, придружено со влошување на условите за живот на луѓето, предизвикани од природни катастрофи или економски активности на самиот човек, извршени без да се земе предвид знаењето за законите на природниот развој.
44. Фази на деградација на биосферата:
употреба на оган (ран палеолит);
развој на земјоделството;
индустриската револуција.
еколошка криза.
45. Извори на деградација на биосфератаможе да биде природен (природен) и вештачки (антропоген). Природно загадување на ОСпредизвикани од природни процеси (прашина, вулканизам, шумски пожари итн.). Вештачко загадувањеврски со емисиите на различни загадувачи во животната средина за време на човековите активности (земјоделство, транспорт, индустрија, итн.)
46. Последици од деградација на биосферата:
Забележливо намалување на биодиверзитетот на екосистемот, уништување и уништување на сè уште преостанатите области на дива вегетација, варварско уништување на шумите и мочуриштата, намалување на бројот на диви животни, исчезнување на многу претставници на флората и фауната. Како резултат на сите овие дејствија, до средината на дваесеттиот век, антропогеното влијание врз биосферата по своето значење влезе на исто ниво како и природното, заземајќи планетарен размер. Така, човештвото стана еден од главните гео-еколошки судбински фактори во еволуцијата на планетата.
47. Загадување– секое воведување во одреден еколошки систем (биоценоза) на живи или неживи компоненти кои не се карактеристични за него, какви било промени кои ги прекинуваат или нарушуваат процесите на циркулација и метаболизам, енергетските текови, чија последица е намалување на продуктивноста или уништување на овој систем.
48. Главни загадувачи:
јаглерод диоксид (CO 2);
јаглерод моноксид (CO);
сулфур диоксид (SO 2);
азотни оксиди (NO, NO 2, N 2 O);
тешки метали и првенствено жива, олово и кадмиум;
радионуклиди и други радиоактивни супстанции;
диоксиди (хлоројаглероди);
цврсти нечистотии (аеросоли): прашина, саѓи, чад;
нафта и нафтени деривати.
канцерогени супстанции, особено бензопирен;
пестициди;
49. Според состојбата на агрегацијаПостојат 3 типа на загадувачи: цврсти, течни и гасовити.
50. По потеклоприрода, состојба на агрегација, размер на дистрибуција, предизвикани последици, степен на токсичност
51. По природаЗагадувачите се класифицираат во следните групи: хемиски, физички, биолошки, естетски.