ក្រណាត់អប់រំ ជាលិកាអប់រំ
(meristems) ជាលិការុក្ខជាតិដែលមានកោសិកាដែលរក្សាសមត្ថភាពក្នុងការបែងចែកបានយូរ។ សូមអរគុណចំពោះការបែងចែកកោសិកាទាំងនេះម្តងហើយម្តងទៀត រុក្ខជាតិលូតលាស់ពេញមួយជីវិតរបស់ពួកគេ (សម្រាប់ដើមឈើខ្លះវាមានរយៈពេលរាប់រយ ឬរាប់ពាន់ឆ្នាំ)។ ទ្រព្យសម្បត្តិសំខាន់ទីពីរនៃកោសិកា meristem គឺថាពួកគេបង្កើតកោសិកាពិសេសដែលបង្កើតជាជាលិកាអចិន្រ្តៃយ៍ - integumentary, basal, conductive, mechanical, excretory ។ អាស្រ័យលើការចែកចាយនៅលើរាងកាយរបស់រុក្ខជាតិដែលកំពុងអភិវឌ្ឍ 4 ប្រភេទ meristems ត្រូវបានសម្គាល់។ Apical ឬ apical meristems ធានាដល់ការលូតលាស់នៃពន្លក និងឫសតាមប្រវែង។ meristems ចំហៀង ឬ lateral កំណត់ការលូតលាស់នៃដើម និងឫសក្នុងក្រាស់ ហើយត្រូវបានគេហៅថា ខេមមៀម. meristems intercalary ឬ intercalary ត្រូវបានរក្សាទុកជាបណ្ដោះអាសន្ននៅក្នុង internodes នៃដើម និងនៅមូលដ្ឋាននៃស្លឹកខ្ចី ធានានូវការលូតលាស់នៃតំបន់ទាំងនេះ ប៉ុន្តែបន្ទាប់មកប្រែទៅជាជាលិកាអចិន្រ្តៃយ៍។ meristems របួស ឬរបួសកើតឡើងនៅកន្លែងដែលរុក្ខជាតិត្រូវបានខូចខាត ដែលពួកគេបង្កើតជាការការពារ callus. សំណួរគឺថាតើជាលិកាផ្សេងគ្នាត្រូវបានបង្កើតឡើងពីកោសិកា meristematic ដូចគ្នា i.e. របៀបដែលភាពខុសគ្នានៃកោសិកាកើតឡើងនៅតែមិនទាន់ដោះស្រាយបាន។
.(ប្រភព៖ “ជីវវិទ្យា។ សព្វវចនាធិប្បាយរូបភាពទំនើប។” និពន្ធនាយក A.P. Gorkin; M.: Rosman, 2006។)
សូមមើលអ្វីដែល "ក្រណាត់អប់រំ" មាននៅក្នុងវចនានុក្រមផ្សេងទៀត៖
ក្រណាត់អប់រំ- មើលជាលិកា meristematic... វចនានុក្រមនៃពាក្យរុក្ខសាស្ត្រ
ជាលិកាអប់រំបន្ទាប់បន្សំ- មានន័យដូច៖ meristems អនុវិទ្យាល័យ គឺជាជាលិកាអប់រំដែលកើតចេញពី meristems បឋម ឬពីជាលិកាអចិន្ត្រៃយ៍។ តំណាងដោយ cambium, phellogen និងរបួស meristems ...
ជាលិកាអប់រំបឋម- មានន័យដូច: meristems បឋម, promeristems, ជាលិកាអប់រំបានមកដោយផ្ទាល់ពី meristems នៃអំប្រ៊ីយ៉ុង; កោសិការបស់ពួកគេដំបូងមានសមត្ថភាពក្នុងការបែងចែក។ ទៅ P. o ។ t. រួមបញ្ចូល meristems apical នៃពន្លកនិង root, procambium, ... ... កាយវិភាគសាស្ត្រ និងសរីរវិទ្យានៃរុក្ខជាតិ
ដាំជាលិកា- ក្រុម ឬស្មុគ្រស្មាញនៃកោសិកាដែលទាក់ទងនឹងរចនាសម្ព័ន្ធទូទៅ ប្រភពដើម មុខងារ និងទីតាំង។ ដោយអនុលោមតាមនេះការអប់រំ, បញ្ចូល, មូលដ្ឋាន, មេកានិច, ចរន្តនិង excretory T. r ត្រូវបានសម្គាល់។ ក្រណាត់អប់រំ ឬ ...... វចនានុក្រមសព្វវចនាធិប្បាយកសិកម្ម
ជាលិការុក្ខជាតិ- ជាលិការុក្ខជាតិ ក្រុម ឬស្មុគ្រស្មាញនៃកោសិកាដែលទាក់ទងនឹងរចនាសម្ព័ន្ធទូទៅ ប្រភពដើម មុខងារ និងទីតាំង។ អនុលោមតាមនេះ ការអប់រំ អាំងតេក្រាល មូលដ្ឋាន មេកានិច ចរន្ត និងការបញ្ចេញទឹកកាម T. r. កសិកម្ម។ វចនានុក្រមសព្វវចនាធិប្បាយធំ
ប្រព័ន្ធកោសិកា ស្រដៀងគ្នានៅក្នុងរចនាសម្ព័ន្ធ ប្រភពដើម និងមុខងារ ខុសគ្នាក្នុងទំហំ រូបរាង និងទីតាំង។ សមាសភាពនៃជាលិការួមមានសារធាតុរាវជាលិកា (បំពេញចន្លោះរវាងកោសិកា) និងសារធាតុដែលស្ថិតនៅចន្លោះកោសិកា។ អំបិលកាល់ស្យូម ...... វចនានុក្រមសព្វវចនាធិប្បាយជីវសាស្រ្ត
គំរូនៃ meristem apical "tunica corpus" ។ Meristems ឬជាលិកាអប់រំ ឬជាលិកា meristematic (ក្រិកបុរាណ ... វិគីភីឌា
Tunica body Model Meristems, or Educational tissues, or Meristematic tissues (ភាសាក្រិច meristos divisible) គឺជាឈ្មោះទូទៅសម្រាប់ជាលិការុក្ខជាតិដែលមានការបែងចែកយ៉ាងខ្លាំង និងរក្សាសកម្មភាពសរីរវិទ្យានៅទូទាំង ... ... Wikipedia
Tunica body Model Meristems, or Educational tissues, or Meristematic tissues (ភាសាក្រិច meristos divisible) គឺជាឈ្មោះទូទៅសម្រាប់ជាលិការុក្ខជាតិដែលមានការបែងចែកយ៉ាងខ្លាំង និងរក្សាសកម្មភាពសរីរវិទ្យានៅទូទាំង ... ... Wikipedia
សរីរាង្គរុក្ខជាតិត្រូវបានបង្កើតឡើងដោយជាលិកា។ ជាលិការុក្ខជាតិគឺជាបណ្តុំនៃកោសិកាដែលមានរចនាសម្ព័ន្ធស្រដៀងគ្នា មានប្រភពដើមដូចគ្នា និងអនុវត្តមុខងារដូចគ្នា។ជាលិកានៃរុក្ខជាតិដីខ្ពស់បានឈានដល់ការអភិវឌ្ឍន៍ពិសេស។ នៅក្នុងរុក្ខជាតិ ជាលិកាអប់រំត្រូវបានសម្គាល់: integumentary, មេកានិច, conductive, មូលដ្ឋាន និង excretory ។
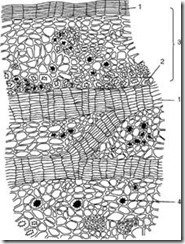
ជាលិកាអប់រំ ឬ meristems បម្រើនៅក្នុងរុក្ខជាតិសម្រាប់ការលូតលាស់ ពួកវាមានដើមកំណើតបឋម និងអនុវិទ្យាល័យ។ ជាលិកាអប់រំបឋមមានទីតាំងនៅកោណលូតលាស់នៃឫសនិងដើម។ សូមក្រឡេកមើលជាមុនសិន កោណលូតលាស់ជា root(រូបទី 1, ខ្ញុំ) ។
រូប ១. ជាលិកាអប់រំបឋមនៅក្នុងកោណលូតលាស់ឫស:
ខ្ញុំ - កោណលូតលាស់ជា root: 1 - កោសិកានៃជាលិកាអប់រំ; 2 - កោសិកានៃមួកឫស។ II - ដំណាក់កាលបន្តបន្ទាប់នៃការបែងចែកនុយក្លេអ៊ែរ និងកោសិកាស្មុគស្មាញ(karyokinesis, ឬ mitosis): 1 - interphase; 2 - prophase; 3 - metaphase; 4-5 - អាណាហ្វាស; 6-7 - telophase; 8 - cytokinesis ។
ចុងឫសមានរូបរាងរាងកោណ គ្របដណ្ដប់ដោយមួកឫស ក្រោមដែលជាលិកាអប់រំបឋមស្ថិតនៅ។ កោសិកានៃជាលិកាអប់រំគឺ tetrahedral ឬ multifaceted មាន protoplasm ក្រាស់ ស្នូលធំ និងភ្នាស cellulose ស្តើង។ vacuoles របស់ពួកគេមិនត្រូវបានអភិវឌ្ឍឬអភិវឌ្ឍតិចតួច។ កោសិកាទាំងនេះមានសមត្ថភាពបែងចែកយ៉ាងខ្លាំងក្លា។ ការបែងចែកកោសិកាត្រូវបាននាំមុខដោយការបែងចែកស្មុគស្មាញនៃស្នូលដែលក្នុងនោះការផ្លាស់ប្តូរពិសេសកើតឡើង។ នៅលើការត្រៀមលក្ខណៈដែលត្រៀមរួចជាស្រេចដែលបានរៀបចំយ៉ាងល្អ និងមានពណ៌ ស្នូលអាចមើលឃើញដែលស្ថិតនៅក្នុងដំណាក់កាលផ្សេងគ្នានៃការបែងចែកស្មុគស្មាញ បើមិនដូច្នេះទេគេហៅថា karyokinesis ("karyon" - nucleus "kinesis" - ចលនា) (រូបភាពទី 1, II) ។ ការសង្កេតជាច្រើនបានបង្កើតឡើងថានៅក្នុងស្នូល សរសៃក្រូម៉ាទីនស្តើងដំបូងកើតចេញពីសារធាតុក្រូម៉ាទីន ដែលខ្លី ក្រាស់ និងប្រែទៅជាភាគល្អិតនីមួយៗ - ក្រូម៉ូសូម ("ក្រូម៉ូសូម" - ពណ៌ "សូម៉ា" - រាងកាយ) ជាញឹកញាប់កោងនៅក្នុងរូបរាង។ horseshoe (ការព្យាករណ៍នៃការបែងចែក) ។ ក្រូម៉ូសូមនីមួយៗនៅក្នុង prophase ហាក់ដូចជាត្រូវបានបំបែកតាមប្រវែងជាពីរពាក់កណ្តាល ប៉ុន្តែការពិតក្រូម៉ូសូមទ្វេដង (ការចម្លងឡើងវិញ) កើតឡើង។ ស្រោមសំបុត្រនុយក្លេអ៊ែរ និងនុយក្លេអូលបាត់បន្តិចម្តងៗ។ ក្រូម៉ូសូមមានទីតាំងនៅផ្នែកកណ្តាលនៃស្នូលក្នុងយន្តហោះតែមួយក្នុងទម្រង់ជាចាន។ ពាក់កណ្តាលនៃក្រូម៉ូសូមនីមួយៗ (ក្រូម៉ូសូម "កូនស្រី") សមនឹងគ្នាទៅវិញទៅមក ហើយក្នុងដំណាក់កាលនេះ (មេតាហ្វាស) នៅលើផ្នែកឆ្លងកាត់នៃឫស អ្នកអាចរាប់ចំនួនក្រូម៉ូសូមយ៉ាងងាយស្រួល និងពិនិត្យមើលរូបរាងរបស់វា។ នៅបង្គោលនៃស្នូល (សូម្បីតែមុនពេលការបាត់ខ្លួននៃភ្នាសនុយក្លេអ៊ែរ) ប្រព័ន្ធនៃសរសៃអំបោះស្តើង (មិនមានស្នាមប្រឡាក់) លេចឡើងដែលត្រូវបានរៀបចំជាទម្រង់មួកពីរ។ ខ្សែស្រឡាយដុះហើយយកទម្រង់ជា spindle ។ បន្ទាប់ពីនេះ ភាពខុសគ្នានៃក្រូម៉ូសូមពាក់កណ្តាល (ក្រូម៉ូសូមកូនស្រី) ចាប់ផ្តើមឆ្ពោះទៅរកប៉ូលទល់មុខពីរនៃកោសិកា (អាណាហ្វាស)។ ការបង្វែរក្រូម៉ូសូមបញ្ចប់ដោយក្រូម៉ូសូមកូនស្រីប្រមូលផ្តុំនៅប៉ូលនៃកោសិកាដោយភ្ជាប់គ្នាទៅវិញទៅមកប្រែទៅជាបាល់ពីរ (តេឡូហ្វាស) ។ ក្នុងពេលជាមួយគ្នានេះ pectin thickenings លេចឡើងនៅលើខ្សែស្រឡាយ achromatin នៅផ្នែកកណ្តាលនៃកោសិកា - phragmoplast មួយត្រូវបានបង្កើតឡើង នោះគឺ rudiment នៃភ្នាសឆ្លងកាត់។ ទីបំផុតស្នូលត្រូវបានហ៊ុំព័ទ្ធដោយភ្នាសមួយ nucleoli (មួយឬច្រើន) ត្រូវបានបង្កើតឡើងនៅក្នុងពួកវា ហើយ protoplasm ត្រូវបានបែងចែកដោយភ្នាសឆ្លងកាត់ (cytokinesis) ដែលបណ្តាលឱ្យមានរូបរាងនៃកោសិកាថ្មីពីរ។
ដំណើរការនៃការបែងចែកនុយក្លេអ៊ែរ និងកោសិកាកើតឡើងញឹកញាប់ជាងនៅពេលយប់ ល្បឿនរបស់វាអាស្រ័យលើសីតុណ្ហភាព និងសំណើម។ ដំណើរការបែងចែកមានរយៈពេលជាមធ្យម 75-120 នាទី ប៉ុន្តែពេលខ្លះមានរយៈពេលជាច្រើនម៉ោង ដោយដំណាក់កាលដំបូង (prophase) ចំណាយពេលច្រើនបំផុត។ ក្នុងអំឡុងពេលនៃការបែងចែកកោសិកាភ្នាសកោសិកាកើតឡើងជាផលិតផលនៃសកម្មភាពសំខាន់នៃ protoplasm និង nucleus ។ ដំបូងចានមធ្យមលេចឡើង។ វាមានសារធាតុអន្តរកោសិកាដែលស្អិតជាប់គ្នានូវភ្នាសកោសិកាបឋមដែលជាកម្មសិទ្ធិរបស់កោសិកាពីរ។
ជាលើកដំបូង ការបង្វែរក្រូម៉ូសូមទៅកាន់ប៉ូលនៃកោសិកា ក៏ដូចជាព័ត៌មានលម្អិតផ្សេងទៀតនៃការបែងចែកស្នូល និងកោសិកាត្រូវបានបង្កើតឡើងក្នុងឆ្នាំ 1875 ដោយសាស្រ្តាចារ្យសាកលវិទ្យាល័យម៉ូស្គូ I. D. Chistyakov ។
ខ្លឹមសារនៃការបែងចែកស្មុគ្រស្មាញនៃស្នូល និងកោសិកា ដែលត្រូវបានគេហៅថា mitosis ជាក់ស្តែងគឺស្ថិតនៅក្នុងការពិតដែលថា ដោយការបំបែកក្រូម៉ូសូមឯកសណ្ឋានជាពីរផ្នែក (ឬច្បាស់ជាងនេះទៅទៀត ក្រូម៉ូសូមទ្វេដង) កោសិកាកូនស្រីទទួលបានពីម្តាយច្រើន ឬតិច។ បរិមាណស្មើគ្នានៃសារធាតុ chromatin (អាស៊ីត deoxyribonucleic) - DNA) ដែលបញ្ជូនតំណពូជពីកោសិកាម្តាយទៅកោសិកាកូនស្រី។ ("Mitos" គឺជាខ្សែស្រឡាយ។ វាត្រូវបានទទួលយកជាធម្មតាថា mitosis = karyokinesis + cytokinesis) ។ រូបរាង និងចំនួនក្រូម៉ូសូមគឺជាលក្ខណៈជាក់លាក់សម្រាប់ប្រភេទរុក្ខជាតិនីមួយៗ (ស្រូវសាលីទន់ 2n = 42 ក្រូម៉ូសូម durum wheat 2n = 28, barley 2n = 14 ។ល។)។ ទោះបីជាយ៉ាងណាក៏ដោយ ពេលខ្លះមានភាពប្រែប្រួលនៃចំនួនក្រូម៉ូសូម ហើយកោសិកាដែលមានក្រូម៉ូសូមផ្សេងគ្នាអាចត្រូវបានគេសង្កេតឃើញនៅក្នុងសារពាង្គកាយមួយ។
បន្ថែមពីលើ mitosis (ឬ karyokinesis) ការបែងចែកដោយផ្ទាល់នៃស្នូលកោសិកា (amitosis) កើតឡើងនៅក្នុងរុក្ខជាតិ - ការទាញស្នូលសាមញ្ញហើយជួនកាលកោសិកាទាំងមូលទៅជាពីរពាក់កណ្តាល។
រួមជាមួយ mitosis ការកាត់បន្ថយការបែងចែកនុយក្លេអ៊ែរគឺជារឿងធម្មតានៅក្នុងរុក្ខជាតិ - meiosis ដែលក្នុងនោះចំនួនក្រូម៉ូសូមនៅក្នុងកោសិកាថ្មីត្រូវបានកាត់បន្ថយពាក់កណ្តាលបើប្រៀបធៀបទៅនឹងកោសិកាម្តាយ។ ការបែងចែកនេះកើតឡើងនៅក្នុងរុក្ខជាតិក្នុងអំឡុងពេល sporulation បន្ទាប់ពីនោះកោសិកាផ្លូវភេទបុរសនិងស្ត្រី (gametes) កើតឡើងនិងអភិវឌ្ឍ។ ក្រោយមកទៀតមានចំនួនពាក់កណ្តាលនៃក្រូម៉ូសូមនៅក្នុងស្នូលរបស់ពួកគេ ហើយស្នូលបែបនេះត្រូវបានគេហៅថា haploid ផ្ទុយទៅនឹង nuclei diploid នៃកោសិកាលូតលាស់ដែលបង្កើតជាសរីរាង្គលូតលាស់របស់រុក្ខជាតិ។ ដំណើរការផ្លូវភេទរបស់រុក្ខជាតិមានការលាយបញ្ចូលគ្នានៃ gametes ពីរ (ប្រុសនិងស្រី) ។ ប្រសិនបើមិនមានការថយចុះនៃសារធាតុក្រូម៉ាទីនទេ នោះគឺពាក់កណ្តាលនៃម៉ាស់របស់វា នោះនៅក្នុងជំនាន់ថ្មីនៃការរួមភេទនីមួយៗ បរិមាណនៃសារធាតុនេះ ហើយដូច្នេះចំនួនក្រូម៉ូសូមនឹងកើនឡើងដោយជៀសមិនរួច។ នៅក្នុងកោសិកាមេរោគ (gametes) ស្នូលគឺ haploid ហើយមានតែបន្ទាប់ពីដំណើរការបង្កកំណើតប៉ុណ្ណោះ ទើបស្នូលកោសិកាក្លាយជា diploid ម្តងទៀត។
ការបែងចែកកាត់បន្ថយត្រូវបានរកឃើញដោយអ្នកវិទ្យាសាស្ត្រ V.I. Belyaev ហើយអ្នកវិទ្យាសាស្ត្រ S.G. Navashin បានរកឃើញការបង្កកំណើតទ្វេរដងនៅក្នុងរុក្ខជាតិផ្កា និងជាស្ថាបនិកនៃសាលាអ្នកវិទ្យាសាស្ត្រទាំងមូលដែលសិក្សាលម្អិតអំពីការបែងចែកនុយក្លេអ៊ែរ និងកោសិកា។
ជាលិកាអប់រំបឋមផ្តល់នូវការកើនឡើងដល់ជាលិកាផ្សេងទៀតចាប់តាំងពីកោសិការបស់វាផ្លាស់ប្តូរភាពខុសគ្នា។ សមត្ថភាពនៃកោសិកាក្នុងការបែងចែកយ៉ាងឆាប់រហ័សនៅក្នុងដើម និងឫសនៃរុក្ខជាតិ dicotyledonous ត្រូវបានរក្សាសម្រាប់ជីវិតនៅក្នុងកោសិកា meristematic ហៅថា cambium ដោយសារតែដើម និងឫសក្រាស់។ ជាលិកាអប់រំបន្ទាប់បន្សំរួមមានជាលិកាដែលកើតចេញពីជាលិកាផ្សេងទៀត។ ឧទាហរណ៍មួយគឺ phellogen ដែលជារឿយៗកើតឡើងពីជាលិកាដី។
ក្រណាត់។ ការចាត់ថ្នាក់នៃក្រណាត់។
ការរៀបចំនៃរុក្ខជាតិខ្ពស់គឺផ្អែកលើគោលការណ៍នៃឯកទេសកោសិកាដែលស្ថិតនៅក្នុងការពិតដែលថាកោសិកានីមួយៗនៃរាងកាយមិនអនុវត្តមុខងារទាំងអស់របស់វាទេប៉ុន្តែមានតែមួយចំនួនប៉ុណ្ណោះប៉ុន្តែកាន់តែពេញលេញនិងល្អឥតខ្ចោះ។
ក្រណាត់- ស្មុគ្រស្មាញដែលកើតឡើងដដែលៗដោយធម្មជាតិនៃកោសិកា ដែលមានលក្ខណៈស្រដៀងនឹងប្រភពដើម រចនាសម្ព័ន្ធ និងប្រែប្រួលដើម្បីអនុវត្តមុខងារមួយ ឬច្រើន។
មានការចាត់ថ្នាក់ផ្សេងៗគ្នានៃក្រណាត់ ប៉ុន្តែពួកគេទាំងអស់សុទ្ធតែបំពាន។
អាស្រ័យលើមុខងារចម្បងក្រុមជាច្រើននៃជាលិការុក្ខជាតិត្រូវបានសម្គាល់។
1. ក្រណាត់អប់រំ,ឬ meristems,- មានសមត្ថភាពក្នុងការបែងចែក និងបង្កើតជាលិកាផ្សេងៗទាំងអស់។
2. ក្រណាត់គ្របដណ្តប់៖
បឋមសិក្សា;
អនុវិទ្យាល័យ;
ឧត្តមសិក្សា។
3. ក្រណាត់សំខាន់ៗ- បង្កើតជារាងកាយរុក្ខជាតិភាគច្រើន។ ក្រណាត់សំខាន់ៗដូចខាងក្រោមត្រូវបានសម្គាល់:
assimilation (chlorophyll-bearing);
ស្តុក;
ខ្យល់ (aerenchyma);
Aquifers ។
4. ក្រណាត់មេកានិច(គាំទ្រ, គ្រោងឆ្អឹង):
Collenchyma;
Sclerenchyma ។
5. ក្រណាត់ដែលមានចរន្តអគ្គិសនី៖
Xylem (ឈើ) គឺជាជាលិការឡើង;
phloem (phloem) គឺជាជាលិកានៃលំហូរចុះ។
6. ជាលិកាបញ្ចេញចោល៖
ខាងក្រៅ៖
សក់ក្រពេញ;
Hydathodes គឺជា stomata ទឹក;
សត្វស្លាបព្រះអាទិត្យ;
ខាងក្នុង:
កោសិកា excretory ជាមួយប្រេងសំខាន់, ជ័រ, តានីន;
បង្កាន់ដៃពហុកោសិកាសម្រាប់ការសម្ងាត់, lacticifers ។
ដោយផ្អែកលើសមត្ថភាពនៃការបែងចែកកោសិកា ជាលិកាពីរប្រភេទត្រូវបានសម្គាល់៖ អប់រំ,ឬ meristems,និង អចិន្ត្រៃយ៍- integumentary, excretory, មូលដ្ឋាន, មេកានិច, conductive ។
ក្រណាត់ត្រូវបានគេហៅថា សាមញ្ញ,ប្រសិនបើកោសិកាទាំងអស់របស់វាមានលក្ខណៈដូចគ្នាបេះបិទនៅក្នុងរូបរាង និងមុខងារ (parenchyma, sclerenchyma, collenchyma)។ ស្មុគស្មាញជាលិកាមានកោសិកាដែលមានរូបរាង រចនាសម្ព័ន្ធ និងមុខងារខុសៗគ្នា ប៉ុន្តែមានទំនាក់ទំនងដោយប្រភពដើមទូទៅ (ឧទាហរណ៍ xylem, phloem)។
វាក៏មានការចាត់ថ្នាក់នៃជាលិកាដោយផ្អែកលើប្រភពដើមរបស់វា (ontogenetic) ។ យោងតាមចំណាត់ថ្នាក់នេះ ជាលិកាបឋម និងបន្ទាប់បន្សំត្រូវបានសម្គាល់។ ពី meristem បឋមដែលមានទីតាំងនៅផ្នែកខាងលើនៃពន្លកនិងចុងឫសក៏ដូចជាពីអំប្រ៊ីយ៉ុងគ្រាប់ពូជ។ អថេរបឋមជាលិកា (epidermis, collenchyma, sclerenchyma, ជាលិកា assimilation, epiblema) ។ កោសិកានៃជាលិកាអចិន្រ្តៃយ៍មិនអាចបែងចែកបន្ថែមទៀតបានទេ។ ពីកោសិកានៃ meristem ឯកទេស - ប្រូខេមប៊ី - ត្រូវបានបង្កើតឡើង អ្នកដឹកនាំបឋមជាលិកា ( xylem បឋម, phloem បឋម) ។
ពី meristem ទីពីរ - ខេមមៀម - ត្រូវបានបង្កើតឡើង អនុវិទ្យាល័យជាលិកា៖ xylem ទីពីរ phloem ទីពីរ; ពី ផូហ្សែន ដោត, phelloderm, និង lentils ត្រូវបានបង្កើតឡើងដែលកើតឡើងនៅពេលដែលដើមនិងឫសក្រាស់។ ជាលិកាបន្ទាប់បន្សំត្រូវបានរកឃើញជាធម្មតានៅក្នុង gymnosperms និង dicotyledonous angiosperms ។ ការអភិវឌ្ឍដ៏មានឥទ្ធិពលនៃជាលិកាបន្ទាប់បន្សំ - ឈើនិងបាស - គឺជាលក្ខណៈនៃរុក្ខជាតិឈើ។
ក្រណាត់អប់រំ
ក្រណាត់អប់រំសូមអរគុណដល់ការបែងចែក mitotic ថេរនៃកោសិការបស់ពួកគេពួកគេធានានូវការបង្កើតជាលិការុក្ខជាតិទាំងអស់ពោលគឺឧ។ តាមពិតរូបរាងរាងកាយរបស់គាត់។ កោសិកាណាមួយនៅក្នុងការអភិវឌ្ឍន៍របស់វាឆ្លងកាត់បីដំណាក់កាល៖ អំប្រ៊ីយ៉ុង ដំណាក់កាលលូតលាស់ និងភាពខុសគ្នា (នោះគឺកោសិកាទទួលបានមុខងារជាក់លាក់មួយ)។ ដោយសារអំប្រ៊ីយ៉ុងមានភាពខុសគ្នា មេរីស្តមបឋមត្រូវបានរក្សាទុកតែនៅចុងពន្លកនាពេលអនាគត (នៅក្នុងកោណលូតលាស់) និងនៅចុងឫស - apical (apical) meristems ។អំប្រ៊ីយ៉ុងនៃរុក្ខជាតិណាមួយមានកោសិកា meristem ។
លក្ខណៈ Cytological នៃ meristems ។លក្ខណៈធម្មតាត្រូវបានបង្ហាញយ៉ាងច្បាស់បំផុតនៅក្នុង meristems apical ។ meristems ទាំងនេះត្រូវបានផ្សំឡើងដោយកោសិកា isodiametric polyhedral ដែលមិនត្រូវបានបំបែកដោយចន្លោះ intercellular ។ សំបករបស់វាស្តើង មានសែលុយឡូសតិចតួច និងអាចលាតសន្ធឹងបាន។
បែហោងធ្មែញនៃកោសិកានីមួយៗត្រូវបានបំពេញដោយ cytoplasm ក្រាស់ជាមួយនឹងស្នូលដ៏ធំមួយដែលកាន់កាប់ទីតាំងកណ្តាលនិងបែងចែកយ៉ាងខ្លាំងដោយ mitosis ។ hyaloplasm មាន ribosomes រាយប៉ាយជាច្រើន, proplastids, mitochondria និង dictyosomes ។ មាន vacuoles តិចតួចហើយវាតូច។ ជាលិកាដំណើរការត្រូវបានបង្កើតឡើងពី meristem ដែលមានរាង prosenchymal និង vacuoles ធំ - procambium និង cambium ។ កោសិកា Procambium មានពហុកោណនៅក្នុងផ្នែកឆ្លងកាត់ កោសិកា cambium មានរាងចតុកោណ។
កោសិកាដែលរក្សាលក្ខណៈសម្បត្តិ meristematic របស់ពួកគេបន្តបែងចែក បង្កើតកោសិកាថ្មីកាន់តែច្រើនឡើងហៅថា អក្សរកាត់។កោសិកាកូនស្រីមួយចំនួនមានភាពខុសគ្នាប្រែក្លាយទៅជាកោសិកានៃជាលិកាផ្សេងៗគេហៅថា ដេរីវេនៃអក្សរកាត់។កោសិកាដំបូងអាចបែងចែកមិនកំណត់បានច្រើនដង ហើយដេរីវេនៃសញ្ញាដំបូងបែងចែកមួយដង ឬច្រើនដង ហើយវិវត្តទៅជាជាលិកាអចិន្ត្រៃយ៍។
ដោយផ្អែកលើប្រភពដើមរបស់ពួកគេ meristems បឋមនិងមធ្យមត្រូវបានសម្គាល់។
meristems បឋម
meristems បឋមកើតឡើងដោយផ្ទាល់ពី meristem នៃអំប្រ៊ីយ៉ុងហើយមានសមត្ថភាពបែងចែក។ យោងតាមទីតាំងរបស់ពួកគេនៅក្នុងរោងចក្រ meristems បឋមអាចជា apical (apical), intercalary (intercalary) និង lateral (lateral) ។
Apical (apical) meristems- meristems បែបនេះដែលមានទីតាំងនៅក្នុងរុក្ខជាតិពេញវ័យនៅកំពូលនៃដើមនិងចុងនៃឫសនិងធានាបាននូវកំណើនរាងកាយនៅក្នុងប្រវែង។ នៅក្នុងដើម នៅក្នុងកោណលូតលាស់ ស្រទាប់ meristematic ពីរត្រូវបានសម្គាល់៖ tunica ដែលជាលិកាភ្ជាប់ និងផ្នែកខាងចុងនៃ Cortex បឋមត្រូវបានបង្កើតឡើង និង corpus ដែលផ្នែកខាងក្នុងនៃ Cortex បឋម និងផ្នែកកណ្តាល។ ស៊ីឡាំងអ័ក្សត្រូវបានបង្កើតឡើង (រូបភាព 2.3) ។
អង្ករ។ ២.៣. meristems apical នៃដើម: ក- ផ្នែកបណ្តោយ៖ ១ - កោណលូតលាស់; 2 - ស្លឹក primordium; 3 - tubercle នៃ axillary bud;
មានបីស្រទាប់នៅចុងឫស៖
1) dermatogen ដែលជាលិកាស្រូបយកបឋម - rhizoderm - ត្រូវបានបង្កើតឡើង;
2) periblema ដែលជាលិកានៃ Cortex បឋមអភិវឌ្ឍ;
3) plerome បង្កើតជាលិកានៃស៊ីឡាំងអ័ក្សកណ្តាល។
ពួកវាអាចជាបឋម ឬអនុវិទ្យាល័យ នៅក្នុងផ្នែកឆ្លងកាត់នៃសរីរាង្គអ័ក្សដែលពួកគេមើលទៅដូចជាចិញ្ចៀន។ ឧទាហរណ៏នៃ meristem ខាងក្រោយចម្បងគឺ procambium និង pericycle ។ ពី ប្រូខេមប៊ី cambium និងធាតុចម្បងនៃបណ្តុំសរសៃឈាម-សរសៃ (primary phloem និង primary xylem) ត្រូវបានបង្កើតឡើង ខណៈពេលដែលកោសិកា procambium ខុសគ្នាដោយផ្ទាល់ទៅក្នុងកោសិកានៃជាលិកាដែលដឹកនាំបឋម។
meristems ពេលក្រោយមានទីតាំងស្ថិតនៅស្របទៅនឹងផ្ទៃនៃសរីរាង្គនិងធានាបាននូវការរីកលូតលាស់នៃសរីរាង្គអ័ក្សនៅក្នុងកម្រាស់។
meristems អន្តរកាលជាញឹកញាប់ពួកវាជាចម្បងហើយត្រូវបានរក្សាទុកក្នុងទម្រង់ជាតំបន់ដាច់ដោយឡែកនៅក្នុងតំបន់នៃការលូតលាស់យ៉ាងសកម្មនៅក្នុងផ្នែកផ្សេងៗនៃរុក្ខជាតិ (ឧទាហរណ៍នៅមូលដ្ឋាននៃស្លឹក petioles នៅមូលដ្ឋាននៃ internodes) ។ នៅមូលដ្ឋាននៃ internodes នៅក្នុងធញ្ញជាតិ សកម្មភាពនៃ meristem នេះនាំឱ្យមានការពន្លូតនៃ internodes ដែលធានាដល់ការលូតលាស់នៃដើមនៅក្នុងប្រវែង។
meristems អនុវិទ្យាល័យ
meristems ទីពីររួមមាន meristems ចំហៀងនិងរបួស។
meristems ចំហៀង (ចំហៀង)បានបង្ហាញ ខេមមៀមនិង ផូហ្សែន។ពួកវាត្រូវបានបង្កើតឡើងពី promeristems (procambium) ឬជាលិកាអចិន្រ្តៃយ៍ដោយ dedifferentiation របស់ពួកគេ។ កោសិកា Cambium ត្រូវបានបែងចែកដោយ Septa ស្របទៅនឹងផ្ទៃនៃសរីរាង្គ (periclinally) ។ ធាតុនៃ phloem ទីពីរវិវត្តចេញពីកោសិកាដែលដាក់នៅខាងក្រៅដោយ cambium ហើយធាតុនៃ xylem បន្ទាប់បន្សំកើតឡើងពីកោសិកាដែលដាក់នៅខាងក្នុង។ cambium ដែលកើតចេញពីជាលិកាអចិន្រ្តៃយ៍តាមរយៈ dedifferentiation ត្រូវបានគេហៅថា បន្ថែមនៅក្នុងរចនាសម្ព័ន្ធនិងមុខងារវាមិនខុសគ្នាពី cambium ដែលកើតឡើងពី promeristems ។ Phellogen ត្រូវបានបង្កើតឡើងពីជាលិកាអចិន្រ្តៃយ៍ដែលស្ថិតនៅក្នុងស្រទាប់ subepidermal (នៅក្រោម epidermis) ។ ការបែងចែក periclinally, phellogen បំបែកកោសិកាដោតនាពេលអនាគត (phelleme) ខាងក្រៅ និងកោសិកា phelloderm ខាងក្នុង។ ដូច្នេះ phellogen បង្កើតជាជាលិកាភ្ជាប់បន្ទាប់បន្សំ - periderm ។ meristems ពេលក្រោយមានទីតាំងស្ថិតនៅស្របទៅនឹងផ្ទៃនៃសរីរាង្គនិងធានាបាននូវការរីកលូតលាស់នៃសរីរាង្គអ័ក្សនៅក្នុងកម្រាស់។
របួសស្នាមរបួសត្រូវបានបង្កើតឡើងនៅពេលដែលជាលិកា និងសរីរាង្គត្រូវបានខូចខាត។ ជុំវិញការខូចខាត កោសិកាមានជីវិតមិនខុសគ្នា ចាប់ផ្តើមបែងចែក ហើយដោយហេតុនេះបំលែងទៅជា meristem ទីពីរ។ ភារកិច្ចរបស់ពួកគេគឺបង្កើតជាលិកាការពារក្រាស់ដែលមានកោសិកា parenchyma - callus ។ជាលិកានេះមានពណ៌ស ឬលឿង កោសិការបស់វាមានស្នូលធំ និងជញ្ជាំងកោសិកាក្រាស់ល្មម។ Callus កើតឡើងកំឡុងពេលផ្សាំ ធានាការលាយបញ្ចូលគ្នានៃ Scion ជាមួយឫសគល់ និងនៅមូលដ្ឋាននៃការកាត់។ វាអាចបង្កើតជាឫស និងពន្លក ដូច្នេះវាត្រូវបានប្រើដើម្បីទទួលបានវប្បធម៌ជាលិកាដាច់ស្រយាល។
គម្របក្រណាត់
ជាលិកាភ្ជាប់បឋម
TO ការរួមបញ្ចូលបឋមជាលិការួមមាន epidermis, epidermal ខ្លួនវា, កោសិកា parastomatal, កោសិកាការពារនៃ stomata និង trichomes ។
សារធាតុ Pectic និងសែលុយឡូសដែលរួមបញ្ចូលនៅក្នុងជញ្ជាំងកោសិកាអាចជាកម្មវត្ថុនៃការបង្កើតស្លសជាមួយនឹងការបង្កើត ទឹករំអិលនិង អញ្ចាញធ្មេញ។ពួកវាជាកាបូអ៊ីដ្រាតវត្ថុធាតុ polymeric ដែលទាក់ទងទៅនឹងសារធាតុ pectin ហើយត្រូវបានកំណត់លក្ខណៈដោយសមត្ថភាពរបស់ពួកគេក្នុងការហើមយ៉ាងខ្លាំងនៅពេលមានទំនាក់ទំនងជាមួយទឹក។ អញ្ចាញធ្មេញក្នុងស្ថានភាពហើមមានភាពស្អិត ហើយអាចទាញចេញជាខ្សែស្រឡាយ ខណៈពេលដែលទឹករំអិលមានភាពស្រពិចស្រពិល និងមិនអាចទាញចេញជាខ្សែស្រឡាយបានទេ។ ទឹករំអិល Pectic ត្រូវបានរកឃើញនៅក្នុងអ្នកតំណាងនៃគ្រួសារ Liliaceae, Cruciferae, Malvaceae, Linden និង Rosaceae ផ្ទុយទៅនឹងភ្នាសកោសិកាដែលមិនសូវមានច្រើនទេ (ឧទាហរណ៍នៅក្នុងផ្កាអ័រគីដេ)។
 ស្តូម៉ាតា
ពួកវាជាទម្រង់ពិសេសខ្ពស់នៃអេពីដេមី ដែលមានកោសិកាយាមរាងសណ្តែកពីរ និងការប្រេះស្រាំ (ប្រភេទនៃចន្លោះរវាងពួកវា)។ ពួកវាត្រូវបានរកឃើញជាចម្បងនៅក្នុងស្លឹក ប៉ុន្តែក៏ត្រូវបានរកឃើញនៅក្នុងដើមផងដែរ (រូបភាព 2.6) ។
ស្តូម៉ាតា
ពួកវាជាទម្រង់ពិសេសខ្ពស់នៃអេពីដេមី ដែលមានកោសិកាយាមរាងសណ្តែកពីរ និងការប្រេះស្រាំ (ប្រភេទនៃចន្លោះរវាងពួកវា)។ ពួកវាត្រូវបានរកឃើញជាចម្បងនៅក្នុងស្លឹក ប៉ុន្តែក៏ត្រូវបានរកឃើញនៅក្នុងដើមផងដែរ (រូបភាព 2.6) ។
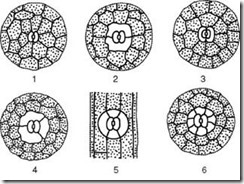
អង្ករ។ ២.៦.រចនាសម្ព័ន្ធក្រពះ៖ ក, ខ- ស្បែកនៃស្លឹក thyme (ទិដ្ឋភាពកំពូលនិងផ្នែកឆ្លងកាត់); វ- បកចេញពីដើមរបស់ Cereus (គ្រួសារ cactus); 1 - កោសិកាស្បែកពិតប្រាកដ; 2 - កោសិកាការពារនៃ stomata; 3 - ការប្រេះស្រាំនៃក្រពះ; 4 - រន្ធខ្យល់; 5 - កោសិកានៃ chlorophyll-bearing parenchyma; A - cuticle; ខ - ស្រទាប់ cuticular - សែលជាមួយ suberin និង wax; ខ - ស្រទាប់សែលុយឡូសនៃជញ្ជាំង; G - ស្នូលជាមួយ nucleolus; ឃ - chloroplasts
ជញ្ជាំងនៃកោសិកាឆ្មាំត្រូវបានក្រាស់មិនស្មើគ្នា: ជញ្ជាំងដែលតម្រង់ឆ្ពោះទៅរកគម្លាត (ពោះ) គឺក្រាស់ជាងបើប្រៀបធៀបទៅនឹងជញ្ជាំងដែលដឹកនាំឆ្ងាយពីគម្លាត (ខាងក្រោយ) ។ គម្លាតអាចពង្រីក និងចុះកិច្ចសន្យា គ្រប់គ្រងការហូរចេញ និងការផ្លាស់ប្តូរឧស្ម័ន។ នៅក្រោមគម្លាតមានប្រហោងផ្លូវដង្ហើមធំមួយ (ចន្លោះរវាងកោសិកា) ហ៊ុំព័ទ្ធដោយកោសិកា mesophyll នៃស្លឹក។
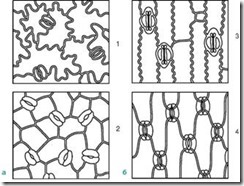
 កោសិកាការពារត្រូវបានហ៊ុំព័ទ្ធដោយកោសិកាប៉ារ៉ាស្តូម៉ាត ដែលរួមគ្នាបង្កើត ស្មុគ្រស្មាញ stomal(រូបភាព 2.7) ។ ប្រភេទសំខាន់ៗនៃស្មុគ្រស្មាញខាងក្រោមត្រូវបានសម្គាល់៖
កោសិកាការពារត្រូវបានហ៊ុំព័ទ្ធដោយកោសិកាប៉ារ៉ាស្តូម៉ាត ដែលរួមគ្នាបង្កើត ស្មុគ្រស្មាញ stomal(រូបភាព 2.7) ។ ប្រភេទសំខាន់ៗនៃស្មុគ្រស្មាញខាងក្រោមត្រូវបានសម្គាល់៖
អង្ករ។ ២.៧.ប្រភេទឧបករណ៍ stomatal សំខាន់ៗ: 1 - anomocytic (នៅក្នុងរុក្ខជាតិខ្ពស់ទាំងអស់លើកលែងតែ horsetails); 2 - diacytic (នៅក្នុង ferns និងរុក្ខជាតិផ្កា); 3 - paracytic (នៅក្នុង ferns, horsetails, ការចេញផ្កានិងការគៀបសង្កត់); 4 - anisocytic (តែនៅក្នុងរុក្ខជាតិផ្កា); 5 - តេត្រាស៊ីទិក (ជាចម្បងនៅក្នុង monocots); 6 - ancyclocytic (នៅក្នុង ferns, gymnosperms និងរុក្ខជាតិផ្កា)
1) អាម៉ូស៊ីទិច(មិនមានសណ្តាប់ធ្នាប់) - កោសិកាការពារមិនមានកោសិកាប៉ារ៉ាស្តូម៉ាតដែលបានកំណត់ច្បាស់លាស់។ លក្ខណៈពិសេសនៃរុក្ខជាតិខ្ពស់ទាំងអស់ដោយមិនរាប់បញ្ចូល conifers;
2) anisocytic(កោសិកាមិនស្មើគ្នា) - កោសិកាការពារនៃ stomata ត្រូវបានហ៊ុំព័ទ្ធដោយកោសិកា parastomatal បីដែលមួយក្នុងចំណោមនោះមានទំហំធំ (ឬតូចជាង) ជាងផ្សេងទៀត។
3) ប៉ារ៉ាស៊ីត(កោសិកាប៉ារ៉ាឡែល) - កោសិកា peristomatal មួយ (ឬច្រើន) មានទីតាំងនៅស្របទៅនឹងកោសិកាយាម។
4) diacytic(កោសិកាឆ្លង) - កោសិកាប៉ារ៉ាស្តូម៉ាតពីរមានទីតាំងនៅកាត់កែងទៅនឹងកោសិកាយាម។
5) តេត្រាស៊ីត(មកពីភាសាក្រិក តេត្រា- បួន) - ជាចម្បងនៅក្នុង monocots;
 Stomata មានទីតាំងនៅផ្នែកខាងក្រោមនៃស្លឹក ប៉ុន្តែនៅក្នុងរុក្ខជាតិទឹកដែលមានស្លឹកអណ្តែត គេឃើញតែនៅផ្នែកខាងលើនៃស្លឹកប៉ុណ្ណោះ។ ដោយផ្អែកលើរូបរាងនៃកោសិកា epidermal ស្លឹក និងទីតាំងនៃ stomata រុក្ខជាតិ monocotyledonous អាចត្រូវបានសម្គាល់ពី dicotyledonous មួយ (រូបភាព 2.8) ។ កោសិកា epidermal ពិតប្រាកដនៃស្លឹករបស់រុក្ខជាតិ dicotyledonous មានរលកនៅក្នុងគ្រោង (រូបភាព 2.9) ខណៈពេលដែលនៅក្នុងរុក្ខជាតិ monocotyledonous ពួកគេត្រូវបានពន្លូត, rhombic នៅក្នុងរូបរាង។
Stomata មានទីតាំងនៅផ្នែកខាងក្រោមនៃស្លឹក ប៉ុន្តែនៅក្នុងរុក្ខជាតិទឹកដែលមានស្លឹកអណ្តែត គេឃើញតែនៅផ្នែកខាងលើនៃស្លឹកប៉ុណ្ណោះ។ ដោយផ្អែកលើរូបរាងនៃកោសិកា epidermal ស្លឹក និងទីតាំងនៃ stomata រុក្ខជាតិ monocotyledonous អាចត្រូវបានសម្គាល់ពី dicotyledonous មួយ (រូបភាព 2.8) ។ កោសិកា epidermal ពិតប្រាកដនៃស្លឹករបស់រុក្ខជាតិ dicotyledonous មានរលកនៅក្នុងគ្រោង (រូបភាព 2.9) ខណៈពេលដែលនៅក្នុងរុក្ខជាតិ monocotyledonous ពួកគេត្រូវបានពន្លូត, rhombic នៅក្នុងរូបរាង។
អង្ករ។ ២.៨.ទីតាំងនៃ stomata នៅលើ epidermis (មើលពីផ្ទៃ): ក- រុក្ខជាតិ dicotyledonous៖ 1 - អក្សរដើម; 2 - ឪឡឹក; ខ- monocots៖ 3 - ពោត; ៤- អាយរីស
ប្រភេទនៃ stomata អាចត្រូវបានបែងចែកទៅតាមកម្រិតនៃទីតាំងរបស់វាទាក់ទងទៅនឹងផ្ទៃនៃ epidermis ដូចខាងក្រោម។
1.7.1. Stomata ស្ថិតនៅក្នុងយន្តហោះដូចគ្នាទៅនឹងអេពីដេមី។ ប្រភេទទូទៅបំផុតនិងជាធម្មតាមិនត្រូវបានចង្អុលបង្ហាញនៅក្នុងការពិពណ៌នានៃមីក្រូទស្សន៍នៃសម្ភារៈរុក្ខជាតិឱសថ, i.e. កថាខណ្ឌនេះត្រូវបានលុបចោល។ សញ្ញារោគវិនិច្ឆ័យនឹងមានស្នាមលេចចេញ ឬលិចទឹក។
1.7.2. ពងបែក - stomata ដែលមានទីតាំងនៅខាងលើ epidermis ។ ជាធម្មតានៅពេលដែលមីក្រូទស្សន៍មីក្រូទស្សន៍ត្រូវបានបង្វិល (នៅពេលដែលកញ្ចក់ត្រូវបានបន្ទាប) ស្នាមប្រេះបែបនេះត្រូវបានរកឃើញជាលើកដំបូង ហើយមានតែកោសិកាអេពីដេមឺមប៉ុណ្ណោះលេចឡើង ដូច្នេះវាស្ទើរតែមិនអាចទៅរួចទេក្នុងការចាប់យកពួកវានៅក្នុងរូបថតពីផ្ទៃស្លឹក ក៏ដូចជា ដើម្បីពណ៌នាពួកគេនៅក្នុងគំនូរ។ នៅក្នុងយន្តហោះដូចគ្នាទៅនឹង epidermis នេះ stomata អាចត្រូវបានគេមើលឃើញនៅក្នុងផ្នែកឆ្លងកាត់ ប៉ុន្តែសម្រាប់ផ្នែកនេះ ត្រូវតែឆ្លងកាត់ stomata ដែលពិបាកក្នុងការទទួលបានទីតាំងដ៏កម្ររបស់ពួកគេនៅលើស្លឹក។ stomata បែបនេះគឺជាលក្ខណៈឧទាហរណ៍នៃស្លឹក bearberry ។
1.7.3. stomata លិចទឹក។ - stomata ជ្រមុជនៅក្នុង epidermis ។ នៅពេលសង្កេតឃើញនៅក្រោមមីក្រូទស្សន៍ដោយការបង្វិលមីក្រូទស្សន៍ (ខណៈពេលកំពុងបន្ទាបកែវ) កោសិកាអេពីដេមឺមត្រូវបានគេរកឃើញយ៉ាងច្បាស់ជាមុនសិន បន្ទាប់មកវាអាចមើលឃើញយ៉ាងច្បាស់នូវវណ្ឌវង្កនៃស្តូម៉ាតា។ វាក៏ពិបាកក្នុងការបង្ហាញពួកវានៅក្នុងរូបថតនិងគំនូរនៃការរៀបចំពីផ្ទៃ។ រកឃើញនៅក្នុង Lily នៃស្លឹកជ្រលងភ្នំ, ស្លឹកមើល, ស្លឹក eucalyptus ។ ជួនកាល រន្ធដែល stomata ស្ថិតនៅត្រូវបានតម្រង់ជួរ ឬគ្របដោយរោម ហើយត្រូវបានគេហៅថា គ្រីបស្តូត។
១.៨. ប្រភេទនៃកោសិកាលំពែង
មាន 19 ប្រភេទដែលបានពិពណ៌នានៅក្នុងអក្សរសិល្ប៍; យើងបានជ្រើសរើសតែប្រភេទដែលត្រូវបានប្រើក្នុងការវិភាគសម្ភារៈរុក្ខជាតិឱសថ ** ។
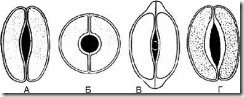
អង្ករ។ ៦៣.ប្រភេទនៃកោសិកាលំពែង។ ក - ទម្រង់ខ្ចី; ខ - ស្វ៊ែរ; ខ - រាងមួក; G - scaphoid
1.8.1. Lenticular - កោសិកាដែលមានរាងដូចអឌ្ឍចន្ទដូចគ្នាចំនួន 2 ត្រូវបានរៀបចំដោយស៊ីមេទ្រី។ នៅលើយន្តហោះផ្នែកខាងមុខការឡើងក្រាស់នៃសែលគឺស្ទើរតែឯកសណ្ឋាន។ ការប្រេះស្រាំគឺ fusiform (រូបភាព 63, ក) ។ ប្រភេទនៃកោសិកា stomatal គឺជាលក្ខណៈរបស់រុក្ខជាតិភាគច្រើន។
1.8.2. ស្វ៊ែរ - កោសិការាងជារង្វង់រាងមូលដូចគ្នាគ្នាខ្លាំងពីរមានទីតាំងស៊ីមេទ្រី។ នៅលើយន្តហោះផ្នែកខាងមុខការឡើងក្រាស់នៃសែលគឺស្ទើរតែឯកសណ្ឋាន។ រន្ធដោតមានរាងមូល (រូបភាព 63, ខ) ។
1.8.3. រាងមួក - កោសិកាដែលមានរាងដូចអឌ្ឍចន្ទដូចគ្នានៅក្នុងផ្នែកប៉ូលមានភាពក្រាស់ក្នុងទម្រង់ជាមួក។ ការប្រេះស្រាំគឺ fusiform (រូបភាព 63, ខ) ។ រកឃើញនៅក្នុងស្រោមដៃ។
1.8.4. Scaphoid - ជញ្ជាំងខាងក្នុងនៃកោសិកា stomatal ត្រូវបានក្រាស់។ ការប្រេះស្រាំគឺ fusiform (រូបភាព 63, ឃ) ។ សង្កេតឃើញនៅស្មៅសតវត្ស និងមើលស្លឹកឈើ។
យន្តការនៃប្រតិបត្តិការ stomata ត្រូវបានកំណត់ដោយលក្ខណៈសម្បត្តិ osmotic នៃកោសិកា។ នៅពេលដែលផ្ទៃស្លឹកត្រូវបានបំភ្លឺដោយព្រះអាទិត្យ ដំណើរការសកម្មនៃការធ្វើរស្មីសំយោគកើតឡើងនៅក្នុង chloroplasts នៃកោសិកាឆ្មាំ។ ការតិត្ថិភាពនៃកោសិកាជាមួយនឹងផលិតផលសំយោគរស្មីសំយោគ និងជាតិស្ករធ្វើឱ្យមានធាតុសកម្មនៃអ៊ីយ៉ុងប៉ូតាស្យូមទៅក្នុងកោសិកា ដែលជាលទ្ធផលដែលកំហាប់នៃកោសិកាបឹងទន្លេសាបក្នុងកោសិកាការពារកើនឡើង។ មានភាពខុសប្លែកគ្នានៅក្នុងកំហាប់នៃកោសិកាបឹងប៉ារ៉ាសិត និងកោសិកាការពារ។ ដោយសារតែលក្ខណៈសម្បត្តិ osmotic នៃកោសិកាទឹកពីកោសិកា parastomatal ចូលទៅក្នុងកោសិកាយាមដែលនាំឱ្យមានការកើនឡើងនៃបរិមាណនៃក្រោយនិងការកើនឡើងយ៉ាងខ្លាំងនៃ turgor ។ ការឡើងក្រាស់នៃជញ្ជាំង "ពោះ" នៃកោសិកាយាមដែលប្រឈមមុខនឹងការប្រេះស្រាំ ធានាបាននូវការលាតសន្ធឹងមិនស្មើគ្នានៃជញ្ជាំងកោសិកា។ កោសិកាឆ្មាំទទួលបានរូបរាងសណ្ដែកដាច់ដោយឡែក ហើយប្រេះស្រាំនឹងបើក។ នៅពេលដែលអាំងតង់ស៊ីតេនៃការធ្វើរស្មីសំយោគមានការថយចុះ (ឧទាហរណ៍នៅពេលល្ងាច) ការបង្កើតជាតិស្ករនៅក្នុងកោសិកាយាមថយចុះ។ ការហូរចូលនៃអ៊ីយ៉ុងប៉ូតាស្យូមឈប់។ កំហាប់នៃកោសិកាបឹងទន្លេសាបនៅក្នុងកោសិកាយាមត្រូវបានកាត់បន្ថយបើប្រៀបធៀបទៅនឹងកោសិកាប៉ារ៉ាស្តូម៉ាត។ ទឹកទុកកោសិកាការពារដោយ osmosis, បន្ថយ turgor របស់ពួកគេជាលទ្ធផល, ការប្រេះស្រាំនៃ stomatal បិទនៅពេលយប់។
កោសិកានៃអេពីដេមីសត្រូវបានបិទយ៉ាងស្អិតជាប់គ្នា ដោយសារអេពីដេមីបំពេញមុខងារមួយចំនួន៖
ការពារការជ្រៀតចូលនៃសារធាតុបង្កជំងឺចូលទៅក្នុងរុក្ខជាតិ;
ការពារជាលិកាខាងក្នុងពីការខូចខាតមេកានិច;
គ្រប់គ្រងការផ្លាស់ប្តូរឧស្ម័ន និងការហូរចេញ;
ទឹកនិងអំបិលត្រូវបានបញ្ចេញតាមរយៈវា;
អាចដំណើរការជាជាលិកាស្រូប;
ចូលរួមក្នុងការសំយោគសារធាតុផ្សេងៗ ការយល់ឃើញនៃការរលាក និងចលនាស្លឹក។
Trichomes - ការរីកដុះដាលនៃកោសិកាស្បែកដែលមានរាង រចនាសម្ព័ន្ធ និងមុខងារផ្សេងៗគ្នា៖ រោម ជញ្ជីង សរសែ។ល។ ពួកវាត្រូវបានបែងចែកទៅជាគម្របនិងក្រពេញ។ trichomes ក្រពេញ,មិនដូចការលាក់បាំងទេ ពួកវាមានកោសិកាដែលលាក់កំបាំង។ គ្របដណ្តប់សក់បង្កើតជារោមចៀម មានអារម្មណ៍ ឬគម្របផ្សេងទៀតនៅលើរុក្ខជាតិ ពួកវាឆ្លុះបញ្ចាំងពីផ្នែកមួយនៃកាំរស្មីព្រះអាទិត្យ ហើយដោយហេតុនេះកាត់បន្ថយការសាយភាយ។ ជួនកាលរោមត្រូវបានគេរកឃើញតែកន្លែងដែល stomata មានទីតាំងនៅឧទាហរណ៍នៅផ្នែកខាងក្រោមនៃស្លឹក coltsfoot ។ នៅក្នុងរុក្ខជាតិខ្លះ រោមដែលនៅរស់បង្កើនផ្ទៃហួតសរុប ដែលជួយពន្លឿនការហូរចេញ។
ទំហំ trichome ប្រែប្រួលយ៉ាងខ្លាំង។ trichomes វែងបំផុត (រហូតដល់ 5-6 សង់ទីម៉ែត្រ) គ្របដណ្តប់គ្រាប់ពូជកប្បាស។ គម្រប trichomes មានទម្រង់នៃរោមទោល ឬច្រើនកោសិកា សាខា ឬផ្កាយ។ គម្រប trichomes អាចនៅរស់បានយូរឬឆាប់ស្លាប់ដោយបំពេញដោយខ្យល់។
ពួកវាខុសគ្នាពី trichomes ដែលកើតឡើងតែជាមួយនឹងការចូលរួមនៃកោសិកាអេពីដេមី។ ការកើតឡើង, នៅក្នុងការបង្កើតដែលជាលិកាដែលមានទីតាំងកាន់តែជ្រៅនៃស្រទាប់ subepidermal ក៏ចូលរួមផងដែរ។
លក្ខណៈកាយវិភាគសាស្ត្រ និងរោគវិនិច្ឆ័យ ដែលមានសារៈសំខាន់បំផុត និងភាពប្រែប្រួលខ្ពស់ក្នុងការកំណត់វត្ថុធាតុដើមឱសថ។ សក់អាចមានលក្ខណៈសាមញ្ញ ឬរាងមូល ដែលនៅក្នុងវេនអាចជា unicellular ឬ multicellular ។ សក់ពហុកោសិកាអាចជាជួរតែមួយ ជួរដេកពីរ ឬមែក។
1.9.1. សក់សាមញ្ញ។
ក. សក់ឯកតាសាមញ្ញ
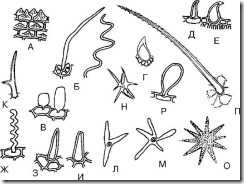
អង្ករ។ ៦៨.សក់ឯកតាសាមញ្ញ។ ក - papillary; ខ - សរសៃ; ខ - សរសៃឈាមវ៉ែន; G - ឆ្អឹងខ្នង; ឃ - រាងជាទំពក់; អ៊ី - រាងកោង; F - កំណាត់ដូចខ្សែស្រឡាយ; Z - រាងកោណមុតស្រួច; ខ្ញុំ - រាងពងក្រពើ; K - ចុងពីរ; L - បីចង្អុល; M, N, O - ពហុចំណុច; P - ដុំពក; P - រាងជាក្លឹប
1. Papillary (រូបភព 68, A; រូបភព 69-74) - ការរីកដុះដាលទាបប៉ុន្តែធំទូលាយនៃកោសិការអេពីដេមី។ ភាគច្រើនត្រូវបានបង្កើតឡើងនៅលើ petals ។ ពួកវាអាចជា apical blunt-apical ឬ acute-apical ។ ពួកវាត្រូវបានរកឃើញនៅលើស្លឹក thyme, Lily នៃផ្កាជ្រលងភ្នំ, ផ្កា violet tricolor, នៅតាមគែមស្លឹក centaury, នៅក្នុងស្មៅពុកចង្ការ gentian ជាដើម។
2. រាងកោណ (រូបភព 68, H, I; រូបភព 75-79) - សក់ត្រង់មានទំនោរនៅមុំមួយទៅផ្ទៃ ដោយបញ្ចប់ដោយចុង (រាងកោណ) ឬមុតស្រួច (រាងកោណ)។ សក់ទូទៅបំផុត។ ជាធម្មតាពួកគេត្រង់។ សក់ទំនោរទៅផ្ទៃត្រូវបានគេសង្កេតឃើញនៅលើស្លឹក thyme ។ រោមរាងកោណចង្អុលត្រូវបានរកឃើញនៅលើស្លឹកសេណា ស្លឹកលីងហ្គនបឺរី ក្នុងស្មៅពណ៌ស្វាយបីពណ៌ (នៅលើស្លឹក) ក្នុងស្មៅ thyme ជាដើម .
3. Filiform (រូបភាព 68, B, G; រូបភព 80, 81) – សក់ស្តើង និងវែង។ អាចត្រូវបានត្រង់ឬ corrugated ។ រោមដូចសរសៃអំបោះត្រង់ត្រូវបានគេរកឃើញនៅលើស្បែកនៃផ្លែ hawthorn និងផ្លែ Raspberry។ រាងដូចខ្សែស្រឡាយ រោមត្រូវបានគេសង្កេតឃើញនៅលើ epidermis នៃផ្កា violet (រូបភាព 82) ។
4. រាងទ្រេត (រូបភាព 68, អ៊ី; រូបភព 83) - សក់ដែលមានមូលដ្ឋានពង្រីក និងចុងកោងតូចចង្អៀត ឬត្រង់។ រកឃើញនៅក្នុងផ្លែឈើហប និងស្មៅ។
5. រាងប្លោកនោម (រូបភាព 68,B; រូបភព 84) – រោមនៅក្នុងទម្រង់ជាពពុះតូចៗ។ ឧទាហរណ៍ពួកគេអាចត្រូវបានរកឃើញនៅក្នុងផ្កា immortelle ។
6. រាងទំពក់ (រូបភព 68, អ៊ី; រូបភព 85-87) - សក់ចង្អុលទៅកំពូល និងកោងក្នុងទម្រង់ជាទំពក់។ រោមទាំងនេះអាចត្រូវបានរកឃើញនៅមូលដ្ឋាននៃស្លឹក bearberry នៅលើផ្ទៃស្លឹក lingonberry ស្លឹក senna និងផ្លែឈើ anise ។
7. គែម (រូបភព 88, 89) - ការលូតលាស់ដ៏វែងនៃកោសិកាអេពីដេមមី ដែលមានទីតាំងនៅតាមគែមស្លឹក ផ្កា ឬ sepal ។ សក់បែបនេះត្រូវបានរកឃើញឧទាហរណ៍នៅតាមគែមនៃផ្កាលីលីនៃជ្រលងភ្នំនិងពណ៌ស្វាយ។
8. រាងឆ្អឹងខ្នង (រូបភព 68, D; រូបភព 90) – រោមគឺស្ទើរតែមានរាងមូលជាមួយនឹងចុងចង្អុល។ ពណ៌ស្វាយបីពណ៌ត្រូវបានគេសង្កេតឃើញនៅលើស្មៅ។
9. រាងក្លឹប (រូបភព 68, P; រូបភព 91) - សក់ជាមួយនឹងផ្នែកចុងពង្រីក, នឹកឃើញដល់ក្លឹបមួយ។ ត្រូវបានរកឃើញនៅលើ thyme petals និង petals violet tricolor ។
10. ទ្វេរដង (រូបភាព 68, K) - សក់បែកជាពីរចុង។ ត្រូវបានគេសង្កេតឃើញនៅក្នុងស្មៅនៃកាបូបរបស់អ្នកគង្វាល។
11. បីចង្អុល (ពហុចំណុច) (រូបភព 68, L-O) - សក់បែកជាបី (ឬច្រើន) ចុង។ ត្រូវបានគេសង្កេតឃើញនៅក្នុងស្មៅនៃកាបូបរបស់អ្នកគង្វាល។
12. ដុំពក (រូបភព 68, ទំ) - រោមរាងកោណជាមួយ protrusions (រលាក់) នៅលើផ្ទៃរបស់ពួកគេ។
ខ. សក់ជួរតែមួយពហុកោសិកាសាមញ្ញ
វណ្ឌវង្កនៃសក់ពហុកោសិកាអាចស្របគ្នាជាមួយនឹងសក់ឯកកោ ប៉ុន្តែមានកោសិកាពីរ ឬច្រើន ហើយតាមនោះ ត្រូវបានគេហៅថាជាសក់ unicellular ស្រដៀងគ្នាដែលបង្ហាញពីចំនួនកោសិកា ឧទាហរណ៍ សក់រាងពងក្រពើ 2 កោសិកា។ 11- សក់ filamentous កោសិកា 10-15 កោសិកា-សក់រាងកោណ។

អង្ករ។ ៩២.សក់ពហុកោសិកាសាមញ្ញ។ ក - រាងកោណ; ខ - រាងពពុះ; ខ - សន្លាក់; G - រាងពងក្រពើ; ឃ - រោម; អ៊ី - រាងអក្សរ T; F - ជួរពីរ; Z - ស្នាមប្រឡាក់; ខ្ញុំ - ធ្នឹម; L - ប៉ារ៉ាឡែល; ម - កាច់ជ្រុង; K - យ៉ាងតឹងរ៉ឹង
1. រាងកោណ (រូបភព ៩២, ក; ៩៣-៩៦)។ សក់ដែលកើតឡើងយ៉ាងទូលំទូលាយបំផុត។ មានត្រង់និងទំនោរទៅនឹងផ្ទៃ, ក្រោយមកទៀតគឺកម្រ (ឧទាហរណ៍នៅក្នុងឱសថ thyme) ។ សក់រាងកោណអាចស្រួច ឬរាងកោណ។ សក់រាងកោណមុតស្រួចគឺជារឿងធម្មតាជាង (នៅក្នុងឱសថនៃ oregano, នៅក្នុងស្លឹកនៃ mint, sage ជាដើម) ។ កោណ Blunt ត្រូវបានរកឃើញនៅក្នុងផ្កា marigold ។
2. រាងទ្រេត - សក់ដែលមានមូលដ្ឋានពង្រីក និងចុងកោងតូចចង្អៀត ឬត្រង់។
3. រាងប្លោកនោម (រូបភាព 92, B; រូបភព 97) - រោមនៅក្នុងទម្រង់ជាពពុះតូចៗ។ មានអូវែ immortelle នៅលើផ្ទៃ។
4. Filiform - សក់ស្តើងនិងវែងពហុកោសិកា។
5. រាងទំពក់ - សក់ចង្អុលនៅកំពូល និងកោងក្នុងទម្រង់ជាទំពក់។
6. Caterpillars (រូបភព 98, 99) - រោមដែលមានកម្រាស់ស្ទើរតែដូចគ្នានៅទូទាំងប្រវែងរបស់វា មានកោសិកាខ្លីស្ទើរតែដូចគ្នា ជាមួយនឹងចុងមិនច្បាស់ និងនឹកឃើញដល់ដង្កូវនាង។ ពួកវាត្រូវបានគេសង្កេតឃើញនៅក្នុងស្មៅនៃ violets tricolor និងនៅក្នុងស្មៅនៃការបន្តពូជ។
7. គែម (រូបភព 100) - ការលូតលាស់ពហុកោសិកាវែងនៃកោសិកាអេពីដេមី ដែលមានទីតាំងនៅតាមគែមស្លឹក ផ្កា ឬ sepal ។ លំដាប់ត្រូវបានគេរកឃើញនៅក្នុងស្មៅ។
8. សាហាវ (រូបភព 92, ឃ; រូបភព 101-104) - រោមដែលមានមូលដ្ឋានពហុកោសិកា រួមមានខ្សែសង្វាក់នៃកោសិកាខ្លី និងកោសិកាស្ថានីយដែលមានរាងជាសរសៃវែង។ នៅក្នុងអក្សរសិល្ប៍ សក់បែបនេះត្រូវបានពិពណ៌នាថាដូចជារំពាត់ ខ្សែដូចខ្សែ អំបោះ ឬមានអារម្មណ៍ដូចរោម។ ប្រភេទសក់គឺជាលក្ខណៈនៃគ្រួសារ Asteraceae ។ ត្រូវបានរកឃើញនៅក្នុងស្មៅ yarrow ផ្កា immortelle ស្លឹក coltsfoot ផ្កា tansy ជាដើម។
9. សន្លាក់ (រូបភព 92, B; រូបភព 105, 106) - សក់ដែលបានពង្រីកមូលដ្ឋានកោសិកានៅកន្លែងនៃសន្លាក់របស់ពួកគេ (ស្រដៀងនឹងសន្លាក់) ។ រោមស្រដៀងគ្នានេះត្រូវបានរកឃើញនៅក្នុងស្មៅ motherwort នៅលើ petals oregano និងកម្រនៅក្នុងស្មៅ thyme ។
10. ដុំពក - រោមដែលមានរោមលើផ្ទៃ។ សរសៃមើមកោសិកាពីរត្រូវបានគេសង្កេតឃើញឧទាហរណ៍នៅក្នុងស្លឹកនៃ thermopsis ។
ខ. សក់មែកច្រើនកោសិកាសាមញ្ញ
1. ប៉ារ៉ាឡែល (រូបភាព 92, អិល; រូបភព 107, 108) - រោមដែលមានកោសិកាត្រង់វែងពីរ (រោម) បញ្ចូលគ្នានៅមូលដ្ឋាន។ ត្រូវបានរកឃើញនៅក្នុងផ្កា linden កម្រនៅក្នុង rosehips និង hawthorns ។
2. សម (រូបភព 92, M; រូបភព 109, 110) - រោមដែលមានកោសិកាខ្យល់វែងពីរ (រោម) បញ្ចូលគ្នានៅមូលដ្ឋាន។ ត្រូវបានគេសង្កេតឃើញនៅក្នុងផ្កា linden កម្រណាស់នៅក្នុងផ្កា hawthorn និងផ្លែឈើ។
3. រាងផ្កាយ (រូបភព 111) - រោមដែលមានកោសិការវែងៗចំនួន 3 ឬច្រើនជាងនេះ (រោម) បញ្ចូលគ្នានៅមូលដ្ឋាន។ ត្រូវបានរកឃើញនៅក្នុងផ្កាលីនដិន។
4. ស្កាលី (រូបភាព 92, H) - រោមដែលមានចានពហុកោសិកា (ក្នុងទម្រង់ជាផ្កាកុលាប) និងដើមខ្លី (ដើមអាចអវត្តមាន) ។ សក់បែបនេះត្រូវបានរកឃើញនៅក្នុងសមុទ្រ buckthorn ។ (ឈ្មោះនេះសម្រាប់សក់ត្រូវបានផ្តល់ឱ្យស្របតាមវាក្យស័ព្ទរុក្ខសាស្ត្រទំនើប។ យោងតាមប្រភពផ្សេងទៀត សក់ទាំងនេះត្រូវបានគេហៅថា stellate ឬ corymbose scaly) ។
5. រាងអក្សរ T (រូបភាព 92, អ៊ី) គឺជាសក់ពីរចង្អុលដែលមានដើមតែមួយ ឬច្រើនកោសិកា។ នៅក្នុងអក្សរសិល្ប៍គេក៏ត្រូវបានគេពិពណ៌នាថាជាស្នែង រាងដូចថ្ម។
6. សៀក (រូបភព 92, ឃ) - រោមពហុកោសិកា នឹកឃើញដល់មែកធាងមែកធាង។ បានរកឃើញឧទាហរណ៍នៅក្នុង mullein ។
ឃ. សក់ពហុកោសិកាសាមញ្ញ
1. ជួរដេកទ្វេ (រូបភព 92, G; រូបភព 112, 113) – រោម កោសិកាដែលត្រូវបានរៀបចំជាពីរជួរ។ ពួកគេអាចត្រូវបានគេសង្កេតឃើញនៅក្នុងផ្កា calendula និងស្មៅស្ងួត។
2. ធ្នឹម (រូបទី 92, I) - រោមដែលមានកោសិកាពីរឬច្រើនត្រូវបានបញ្ចូលគ្នាយ៉ាងតឹងជាមួយគ្នាទៅវិញទៅមកបង្កើតជាបាច់។
3. យ៉ាងតឹងរ៉ឹង (រូបភព 92 K, រូបភព 114) - សក់ពហុកោសិកា មានរោមដែលមានប្រវែងខុសៗគ្នា ប្រសព្វគ្នាទៅវិញទៅមក និងមានចុងចង្អុលដោយឥតគិតថ្លៃ។ សក់បែបនេះច្រើនតែជាសក់រុយ (គ្រួសារ Asteraceae) ។នៅក្នុងអក្សរសិល្ប៍ពួកគេត្រូវបានគេហៅផងដែរថា jagged complex, awl-shaped ។
1.9.2. កោរសក់។ សក់ Capitate ត្រូវបានគេហៅផងដែរថាសក់ក្រពេញ។ ពួកវាអាចជា unicellular ឬ multicellular ។

អង្ករ។ ១១៥.កោរសក់។ ក - មានដើមកោសិកាតែមួយ និងក្បាលកោសិកាតែមួយ។ ខ - ជាមួយនឹងជើងពីរជួរនិងក្បាលកោសិកាតែមួយ; ខ - ជាមួយជើងពីរជួរនិងក្បាលពីរជួរ; G - មានដើម unicellular និងក្បាល multicellular; ឃ - មានដើមពហុកោសិកា និងក្បាល unicellular; F - មានដើមពហុកោសិកា និងក្បាលពហុកោសិកា; Z - unicellular (ដុត); អ៊ី - ទីរ៉ូអ៊ីត
ក. រោម Unicellular capitate
អ្នកតំណាងនៃសក់ប្រភេទនេះអាចមានរាងកោណ ឬរាងមូល ប៉ុន្តែតែងតែមានក្បាលនៅចុងបញ្ចប់។ សក់បែបនេះត្រូវបានរកឃើញឧទាហរណ៍នៅលើស្លឹក nettle ។ ខ. រោមពហុកោសិកា (ក្រពេញ)
1. សក់មានក្បាលច្រើនកោសិកា និងមានទងជាកោសិកា (រូបភាព 115, ឃ; រូបភព 116-120) ។ ត្រូវបានរកឃើញនៅក្នុងស្មៅ motherwort និងស្លឹក bearberry ។
2. សក់ដែលមានក្បាលកោសិកាតែមួយ និងដើមកោសិកាតែមួយ (រូបភព 115, A; រូបភព 121-123)។ ត្រូវបានរកឃើញនៅក្នុងស្មៅ motherwort, ស្លឹក sage, ផ្កា elderberry, ស្លឹក mint ។
3. សក់មានក្បាល unicellular និង stalk ពហុកោសិកា (រូបភាព 115, D; រូបភព 124) ។ ត្រូវបានរកឃើញនៅក្នុងស្មៅ motherwort ស្លឹក sage ផ្កា calendula និង pedicels chamomile ។
4. សក់មានក្បាលពហុកោសិកា និងដើមពហុកោសិកា (រូបភាព 115, G; រូបភព 125) ។ ត្រូវបានរកឃើញនៅក្នុងស្មៅ motherwort, ស្លឹក bearberry, ផ្កា linden, ផ្កា calendula, ស្មៅ violet ។
5. សក់មានក្បាលក្រឡាតែមួយ និងដើមពីរជួរ (រូបភាព 115, ខ; រូបភព 126) ។ ត្រូវបានរកឃើញនៅក្នុងផ្កានៃ calendula និង immortelle ។
6. សក់មានក្បាលពីរជួរ និងដើមពីរជួរ (រូបភព 115, B; រូបភព 127-130)។ ពួកគេត្រូវបានរកឃើញនៅក្នុងផ្កានៃ calendula និង immortelle និងនៅក្នុងស្មៅនៃ marsh ស្មៅ។
7. រាងក្លឹប សក់ក្រពេញពហុកោសិកា (រូបភាព 131-133; សូមមើលរូបទី 155, អ៊ី) - សក់ដែលមានវណ្ឌវង្កជាមួយនឹងផ្នែកចុងដែលពង្រីក ដែលនឹកឃើញដល់ក្លឹប។ ពួកគេអាចត្រូវបានគេសង្កេតឃើញឧទាហរណ៍នៅក្នុងស្លឹក lingonberry និងស្មៅ violet ។
8. ក្រពេញទីរ៉ូអ៊ីត រោមក្រពេញពហុកោសិកា (រូបទី 115, I; រូបភព 134) គឺជាខែលនៃកោសិកាដែលមានជញ្ជាំងស្តើងពហុកោណ ដែលអង្គុយនៅលើដើមខ្លីមួយ ឬពីរកោសិកា។ cuticle នៃ scutellum ផ្លាស់ទីឆ្ងាយពីកោសិកាជាមួយនឹងប្រេងសំខាន់ៗដែលលាក់នៅក្រោមវា។ ពួកវាត្រូវបានរកឃើញនៅក្នុងផ្លែឈើហប។
១.១០. ធម្មជាតិនៃការឡើងក្រាស់នៃជញ្ជាំងកោសិកា និងរោមដែលគ្របដណ្តប់លើ cuticle
A. លក្ខណៈនៃការឡើងក្រាស់នៃជញ្ជាំងកោសិកា
1.10.1. ជញ្ជាំងស្តើង (រូបភព ១៣៥-១៣៧)។ សក់ភាគច្រើនមានជញ្ជាំងស្តើង។ ជញ្ជាំងកោសិកានៃសក់ស្តើង ជញ្ជាំងវែង ជួនកាលដួលរលំ ដែលរំខានដល់វណ្ឌវង្កនៃសក់។ ជាឧទាហរណ៍ សក់បែបនេះត្រូវបានរកឃើញនៅលើស្រទាប់ខាងក្រោមនៃស្លឹក kakali ។
1.10.2. ជញ្ជាំងក្រាស់ (រូបភាព 138, 139) ។ ត្រូវបានគេរកឃើញនៅក្នុង rosehips, string grass, Dahurian lollipop grass ជាដើម។
1.10.3. ក្រាស់មិនស្មើគ្នា។ អ្នកអាចមើលឃើញ motherwort នៅក្នុងស្មៅ (រូបភាព 140) ។ សក់ Rosehip ជាមួយនឹងរន្ធញើសឆ្លងកាត់ oblique ក៏ជាកម្មសិទ្ធិរបស់ប្រភេទនេះផងដែរ (រូបភាព 141, 142) ។
B. ធម្មជាតិនៃរោមដែលគ្របដណ្តប់លើ cuticle
1.10.4. ផ្ទៃរលោង (រូបភាព 143) ។ សក់ដែលមានផ្ទៃបែបនេះត្រូវបានគេសង្កេតឃើញឧទាហរណ៍នៅក្នុងស្លឹក plantain ស្មៅ celandine ជាដើម។
1.10.5. ផ្ទៃ warty (សូមមើលរូបទី 1, G; រូបភព 144, 145) - epidermis បង្កើតជាទម្រង់នៃ tubercles (ឬស) ។ មានរោម ជាឧទាហរណ៍ នៅក្នុងស្មៅពណ៌ស្វាយ ស្មៅ thyme ក្នុងស្មៅ motherwort ក្នុងស្លឹក mint ស្លឹក senna ជាដើម ក្នុងករណីនេះ ផ្ទៃនៃសក់អាចប្រេះស្រាំបន្តិច នៅពេលដែលមានស្នាមតូចៗនៃ cuticle ត្រូវបានបង្កើតឡើង។ និងមានភាពស្វិតស្វាញនៅពេលដែលមានស្នាមប្រេះសំខាន់ៗត្រូវបានបង្កើតឡើង។ ក្នុងករណីទី 1 រោមនៅក្នុងស្មៅពណ៌ស្វាយអាចត្រូវបានលើកឡើងជាឧទាហរណ៍នៅក្នុងទីពីរ - រោមនៅក្នុងស្លឹក senna និងស្មៅ thyme ។
1.10.6. ផ្ទៃរដុប (សូមមើលរូបទី 68, P) - cuticle បង្កើតជាប្រហោងធំៗ ឧទាហរណ៍រោមរបស់ស្មៅ Thermopsis lanceolata ។
10.7. ផ្ទៃដែលមានស្នាមជ្រួញ - បត់ឬរលកនៃ cuticle សក់ពីមូលដ្ឋានទៅចុងរបស់វា (រូបភាព 146) ។ វាត្រូវបានគេសង្កេតឃើញជាញឹកញាប់នៅក្នុង papillae ឧទាហរណ៍នៅក្នុងស្មៅ centaury និងផ្កា elderberry ។ ទោះជាយ៉ាងណាក៏ដោយ សក់ធម្មតាក៏អាចមានផ្ទៃជ្រីវជ្រួញផងដែរ ឧទាហរណ៍នៅក្នុងស្មៅខ្សែអក្សរ។ នៅមូលដ្ឋានរោមរបស់ស្លឹក mint មានស្នាមជ្រីវជ្រួញ (រូបភាព 147) ។
1.10.8. ផ្ទៃញាស់ - អេពីដេមីបង្កើតជាប្រហោងខ្លីក្នុងទម្រង់ជាស្នាមប្រេះ (មើម ឬស)។ វាកម្រ និងជាជម្រើសកម្រិតមធ្យមរវាងផ្ទៃដែលមានស្នាមជ្រីវជ្រួញ។ ជាឧទាហរណ៍ វាត្រូវបានគេសង្កេតឃើញនៅលើរោមរបស់ផ្កាចាស់ (រូបភាព 148)។
១.១១. លក្ខណៈពិសេសនៃកន្លែងភ្ជាប់សក់
1.11.1. គេហទំព័រឯកសារភ្ជាប់ទូទៅ (រូបភាព 149, 150) - រោមត្រូវបានភ្ជាប់ទៅនឹងកោសិកាឬរវាងកោសិកានៃ epidermis ។ ប្រភេទទូទៅបំផុតត្រូវបានគេរកឃើញឧទាហរណ៍នៅក្នុងស្លឹក coltsfoot ស្លឹក mint ស្លឹក sage ឱសថ motherwort ឱសថ violet ជាដើម។
1.11.2. ការរីកដុះដាលចេញពីកោសិកាអេពីដេមី (សូមមើលរូប 68, A; 69-74)។ ត្រូវបានគេសង្កេតឃើញនៅក្នុងផ្កា elderberry, ស្មៅ centaury, petals motherwort, ស្មៅពុកចង្ការ gentian, epidermis នៃផ្លែឈើ anise ជាដើម។
1.11.3. នៅលើមូលដ្ឋាននៃសក់ rosette នៃកោសិកា epidermal ត្រូវបានបង្កើតឡើង (រូបភាព 151, 152) ។ មាននៅក្នុងស្លឹក senna, ស្លឹក plantain, ឱសថ oregano ជាដើម។
1.11.4. មូលដ្ឋានសក់ពង្រីក (រូបភាព 153) ។ ត្រូវបានរកឃើញនៅក្នុងស្លឹក plantain ។
1.11.5. ផ្នែកដែលពង្រីកនៃមូលដ្ឋានសក់ត្រូវបានជ្រមុជនៅក្នុងមូលដ្ឋាននៃជាលិកា subepidermal - ការលេចឡើង (សូមមើលរូប 115, 3)។ វាអាចត្រូវបានគេសង្កេតឃើញឧទាហរណ៍នៅក្នុងស្លឹក nettle ។
1.11.6. មូលដ្ឋានសក់ពហុកោសិកា (រូបភាព 154) ។ ឧទាហរណ៍អ្នកអាចសង្កេតមើលខ្សែនៅក្នុងស្មៅ។
ជាញឹកញាប់ សក់បែកចេញដោយបន្សល់ទុកចំណុចភ្ជាប់របស់វានៅលើអេពីដេមី ដែលចាំបាច់ត្រូវកត់សំគាល់ថាជាសញ្ញាកាយវិភាគសាស្ត្រ និងរោគវិនិច្ឆ័យនៃវត្ថុធាតុដើមរុក្ខជាតិឱសថ។
ក្រដាសបិទបាំងបន្ទាប់បន្សំ
ជាលិកាភ្ជាប់បន្ទាប់បន្សំត្រូវបានគេហៅថា periderma ។នេះគឺជាជាលិកាគ្របដណ្តប់ស្មុគស្មាញនៃដើម, ឫសនិង rhizomes នៃរុក្ខជាតិដែលមានអាយុច្រើនឆ្នាំ។ វាជំនួសអេពីដេមីនៃសរីរាង្គអ័ក្ស ដែលងាប់បន្តិចម្តងៗ ហើយរលត់ទៅវិញ។ periderm ត្រូវបានបង្កើតឡើងពី phellogen ( meristem ទីពីរ) ។ Phellogen ត្រូវបានបង្កើតឡើងនៅក្នុង epidermis ស្រទាប់ subepidermal និងសូម្បីតែនៅក្នុងស្រទាប់ជ្រៅនៃសរីរាង្គអ័ក្ស។ កោសិកា Phellogen បែងចែកដូចខាងក្រោមៈ កោសិកាដោតត្រូវបានដាក់នៅខាងក្រៅ ហើយកោសិកា parenchyma ដែលមានជីវិតរបស់ phelloderm ត្រូវបានដាក់នៅខាងក្នុង។ កោសិកា phelloderm នៃដើមមានផ្ទុក chloroplasts ។
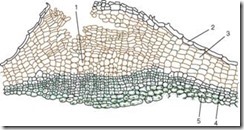 ដោតត្រូវបានបង្កើតឡើងដោយកោសិកាងាប់ដែលជញ្ជាំងកោសិកាត្រូវបាន impregnated ជាមួយសារធាតុដូចជាជាតិខ្លាញ់ដែលគេហៅថា suberin ។ ក្រឡាត្រូវបានរៀបចំជាជួរស្មើគ្នា មានរាងចតុកោណកែង (នៅលើផ្នែកឈើឆ្កាង) ហើយត្រូវគ្នាយ៉ាងតឹងណែន បង្កើតជាស្រោមពហុស្រទាប់។ Cork ការពារជាលិការស់ខាងក្នុងពីការបាត់បង់ជាតិសំណើម ការប្រែប្រួលសីតុណ្ហភាពភ្លាមៗ និងការជ្រៀតចូលនៃអតិសុខុមប្រាណ។ ជាលិការស់ដែលដេកនៅក្រោមដោតត្រូវការការផ្លាស់ប្តូរឧស្ម័ន និងការដកសំណើមលើស។ នោះហើយជាមូលហេតុដែលនៅក្រោម stomata ដែលជាលទ្ធផលនៃការបែងចែកនៃស្រទាប់ subepidermal (សូម្បីតែមុនពេលរូបរាងនៃ periderm នេះ) និងជាបន្តបន្ទាប់ phellogen, រស់នៅ, កោសិកា parenchyma ទីតាំងរលុងជាមួយនឹងចន្លោះ intercellular ជាច្រើន, ហៅថា សម្តែងការត្បាញថ្មី។ដែលបំបែកអេពីដេមី និងបង្កើតលទ្ធភាពនៃការផ្លាស់ប្តូរឧស្ម័ន និងការឆ្លងជាមួយបរិយាកាសខាងក្រៅ។ ការបង្កើតរចនាសម្ព័ន្ធនេះត្រូវបានគេហៅថា សណ្តែកសៀង(រូបភាព 2.12) ។
ដោតត្រូវបានបង្កើតឡើងដោយកោសិកាងាប់ដែលជញ្ជាំងកោសិកាត្រូវបាន impregnated ជាមួយសារធាតុដូចជាជាតិខ្លាញ់ដែលគេហៅថា suberin ។ ក្រឡាត្រូវបានរៀបចំជាជួរស្មើគ្នា មានរាងចតុកោណកែង (នៅលើផ្នែកឈើឆ្កាង) ហើយត្រូវគ្នាយ៉ាងតឹងណែន បង្កើតជាស្រោមពហុស្រទាប់។ Cork ការពារជាលិការស់ខាងក្នុងពីការបាត់បង់ជាតិសំណើម ការប្រែប្រួលសីតុណ្ហភាពភ្លាមៗ និងការជ្រៀតចូលនៃអតិសុខុមប្រាណ។ ជាលិការស់ដែលដេកនៅក្រោមដោតត្រូវការការផ្លាស់ប្តូរឧស្ម័ន និងការដកសំណើមលើស។ នោះហើយជាមូលហេតុដែលនៅក្រោម stomata ដែលជាលទ្ធផលនៃការបែងចែកនៃស្រទាប់ subepidermal (សូម្បីតែមុនពេលរូបរាងនៃ periderm នេះ) និងជាបន្តបន្ទាប់ phellogen, រស់នៅ, កោសិកា parenchyma ទីតាំងរលុងជាមួយនឹងចន្លោះ intercellular ជាច្រើន, ហៅថា សម្តែងការត្បាញថ្មី។ដែលបំបែកអេពីដេមី និងបង្កើតលទ្ធភាពនៃការផ្លាស់ប្តូរឧស្ម័ន និងការឆ្លងជាមួយបរិយាកាសខាងក្រៅ។ ការបង្កើតរចនាសម្ព័ន្ធនេះត្រូវបានគេហៅថា សណ្តែកសៀង(រូបភាព 2.12) ។
អង្ករ។ ២.១២.រចនាសម្ព័ន្ធនៃ periderm ជាមួយ lentil: 1 - ជាលិកាគាំទ្រនៃ lentil; 2 - នៅសល់នៃ epidermis; 3 - ឆ្នុក (phellema); 4 - phellogen; 5 - phelloderm
Lentils ដែលមើលទៅដូចជា tubercles តូចៗអាចមើលឃើញយ៉ាងច្បាស់នៅលើផ្ទៃនៃពន្លកនៃដើមឈើនិង shrubs (រូបភាព 2.13) ។
នៅលើដើម birch នៅសល់របស់ពួកគេត្រូវបានគេសង្កេតឃើញនៅក្នុងទម្រង់នៃឆ្នូតខ្មៅឆ្លងកាត់លក្ខណៈនៅលើ aspen ពួកគេយកទម្រង់ពេជ្រ។
ជាលិកាភ្ជាប់ទីបី
 សំបក (rhytide)គឺជាជាលិកាភ្ជាប់ថ្នាក់ទីបី ដែលត្រូវបានបង្កើតឡើងនៅក្នុងរុក្ខជាតិដែលមានអាយុច្រើនឆ្នាំនៅក្នុងឫស ដើម និង rhizome ។ ជារៀងរាល់ឆ្នាំនៅក្នុងស្រទាប់កាន់តែជ្រៅ ស្រទាប់ phellogen ថ្មីមួយត្រូវបានដាក់ចុះ ហើយ periderm ត្រូវបានបង្កើតឡើង។ ស្រទាប់ខាងក្រៅនៃ periderm - ឆ្នុក - ដាច់ដោយឡែកពីជាលិកាទាំងអស់ដែលជាលទ្ធផលដែលពួកគេស្លាប់។ ដូច្នេះការប្រមូលផ្តុំនៃ periderms ជាច្រើនដែលមានជាលិកាស្លាប់រវាងពួកវាគឺសំបក (រូបភាព 2.14) ។
សំបក (rhytide)គឺជាជាលិកាភ្ជាប់ថ្នាក់ទីបី ដែលត្រូវបានបង្កើតឡើងនៅក្នុងរុក្ខជាតិដែលមានអាយុច្រើនឆ្នាំនៅក្នុងឫស ដើម និង rhizome ។ ជារៀងរាល់ឆ្នាំនៅក្នុងស្រទាប់កាន់តែជ្រៅ ស្រទាប់ phellogen ថ្មីមួយត្រូវបានដាក់ចុះ ហើយ periderm ត្រូវបានបង្កើតឡើង។ ស្រទាប់ខាងក្រៅនៃ periderm - ឆ្នុក - ដាច់ដោយឡែកពីជាលិកាទាំងអស់ដែលជាលទ្ធផលដែលពួកគេស្លាប់។ ដូច្នេះការប្រមូលផ្តុំនៃ periderms ជាច្រើនដែលមានជាលិកាស្លាប់រវាងពួកវាគឺសំបក (រូបភាព 2.14) ។
អង្ករ។ ២.១៤.សំបកអុក៖ ១ - ស្រទាប់ឆ្នុក; 2 - សរសៃ; 3 - នៅសល់នៃសំបកដើម; ៤- ដ្រូសេននៃកាល់ស្យូម oxalate
(កោសិកា meristematic) ហើយត្រូវបានរកឃើញនៅក្នុងផ្នែកនៃរុក្ខជាតិដែលការលូតលាស់កើតឡើង។
1. លក្ខណៈ Cytological នៃ meristems
កោសិការុក្ខជាតិដែលបែងចែកខុសគ្នាមិនអាចបែងចែក ឬបំប្លែងទៅជាប្រភេទកោសិកាផ្សេងទៀតបានទេ។ ដូច្នេះការបែងចែកកោសិកានៅក្នុង meristems គឺចាំបាច់ដើម្បីផ្តល់កោសិកាថ្មីសម្រាប់ការលូតលាស់នៃជាលិកាផ្សេងទៀត ការបង្កើតសរីរាង្គថ្មី និងរចនាសម្ព័ន្ធនៃរាងកាយរបស់រុក្ខជាតិ។ មុខងារនៃកោសិកា meristematic គឺស្រដៀងទៅនឹងកោសិកាដើមរបស់សត្វ ដែលមិនមានភាពខុសប្លែកគ្នា ឬខុសគ្នាបន្តិចបន្តួច និងមានសមត្ថភាពបែងចែកកោសិកាជាបន្តបន្ទាប់។ កោសិកា Meristematic មានទំហំតូច ហើយ cytoplasm និង nucleus បំពេញកោសិកាទាំងស្រុង។ vacuoles មានទំហំតូចខ្លាំង ហើយ cytoplasm មិនមានផ្ទុកផ្លាស្ទីតខុសគ្នា (chloroplasts ឬ chromoplasts) ទោះបីជាពួកវាមានវត្តមាននៅក្នុងទម្រង់ vestigial (protoplastids) ក៏ដោយ។ កោសិកា Meristematic ត្រូវបានប្រមូលផ្តុំយ៉ាងតឹងរ៉ឹង ដោយស្ទើរតែគ្មានចន្លោះរវាងកោសិកា។ ជញ្ជាំងកោសិកាគឺជាជញ្ជាំងកោសិកាបឋមស្តើងណាស់។
សកម្មភាពសរីរវិទ្យានៃ meristems តម្រូវឱ្យមានតុល្យភាពរវាងដំណើរការប្រឆាំងពីរ: ការបង្កើតជាលិកាថ្មីនិងការគាំទ្រសម្រាប់ការបន្តនៃចំនួនប្រជាជននៃកោសិកា meristematic ។
Meristem- បង្កើតជាលិការុក្ខជាតិ ដែលជាលិកាផ្សេងទៀតទាំងអស់អាចត្រូវបានបង្កើតឡើង កោសិកា meristematic សម្រាប់រយៈពេលដ៏យូររក្សាសមត្ថភាពក្នុងការបែងចែកជាមួយនឹងការបង្កើតកោសិកាថ្មីដែលមិនពិសេស។
កោសិកានៃ meristem ត្រូវបានរុញភ្ជាប់, តូច, ក្រាស់ដែលមានទីតាំងនៅជាប់គ្នាដោយគ្មាន intercliniforms, ស្នូលមានទីតាំងនៅកណ្តាលកោសិកា, vacuoles តូច, ជាច្រើន, ភ្នាសកោសិកាគឺបឋម, មានតែផ្លាស្ទីតមុនគេ។
លក្ខណៈ cytological នៃ meristems croup ត្រូវបានបង្ហាញជាធម្មតានៅក្នុង meristems apical ។ កោសិកាគឺជាពហុកោណ isodiametric មិនត្រូវបានបំបែកដោយចន្លោះរវាងកោសិកាទេ។ ជញ្ជាំងកោសិកាគឺស្តើង ដោយមានបរិមាណសែលុយឡូសតិចតួច។ cytoplasm គឺក្រាស់ ស្នូលមានទំហំធំ ដែលមានទីតាំងនៅកណ្តាល។ នៅក្នុង cytoplasm មាន ribosomes និង mitochondria មួយចំនួនធំ (ការសំយោគប្រូតេអ៊ីន និងសារធាតុផ្សេងៗទៀតកើតឡើង)។ vacuoles ជាច្រើនគឺតូចណាស់។
កោសិកានៃ meristems ខាងក្រោយមានទំហំ និងរូបរាងមិនស្មើគ្នា។ នេះគឺដោយសារតែភាពខុសគ្នានៃកោសិកានៃជាលិកាអចិន្រ្តៃយ៍ដែលពួកគេត្រូវបានបង្កើតឡើង។ ឧទាហរណ៍នៅក្នុង cambium មានកោសិកា parenchymal និង prosenchymal ។ parenchyma នៃស្មុគស្មាញឈានមុខគេត្រូវបានបង្កើតឡើងពីការចាប់ផ្តើម parenchymal ហើយធាតុ conductive ខ្លួនវាត្រូវបានបង្កើតឡើងពី prosenchymal បឋម។
Meristems ត្រូវបានចាត់ថ្នាក់តាមទីតាំងរបស់វានៅក្នុងរុក្ខជាតិ៖
និងប្រភពដើម៖
- បឋម
- អនុវិទ្យាល័យ។
meristems បឋម មានទំនាក់ទំនងជិតស្និទ្ធជាមួយ meristem apical ហើយដោយផ្ទាល់ពីពួកវាត្រូវបានបង្កើតឡើងជាពិសេស protoderm មេ meristem និង procambium ។ Protoderm គឺជាស្រទាប់តែមួយនៃកោសិកាដែលមានភាពខុសប្លែកគ្នាតិចតួចដែលគ្របដណ្ដប់លើកំពូលពីខាងក្រៅ អេពីដេមីត្រូវបានបង្កើតឡើងជាបន្តបន្ទាប់ពីវា។ Cortex ត្រូវបានបង្កើតឡើងពី meristem មេ ហើយស៊ីឡាំងកណ្តាលត្រូវបានបង្កើតឡើងពី procambium ដែលជាកោសិកាដែលត្រូវបានពន្លូតតាមអ័ក្សនៃដើម។
meristems អនុវិទ្យាល័យ ជ្រាបចូលពីកោសិកាឯកទេសនៃសរីរាង្គរុក្ខជាតិចាស់ទុំ និងធានាបាននូវការលូតលាស់នៅពេលក្រោយ (phelogen, pericycle, cambium)។ ជាលិកាដែលបង្កើតឡើងពី meristem បឋមត្រូវបានគេហៅថាបឋម ហើយជាលិកាដែលបង្កើតឡើងពី meristem ទីពីរត្រូវបានគេហៅថាអនុវិទ្យាល័យ។ meristems ទីពីរក៏គួររួមបញ្ចូលផងដែរនូវអ្វីដែលគេហៅថា traumatic meristems ដែលត្រូវបានបង្កើតឡើងនៅកន្លែងដែលរាងកាយរបស់រុក្ខជាតិត្រូវបានខូចខាត និងផ្តល់នូវការបង្កើតឡើងវិញ។
ក្នុងចំណោមប្រភេទនៃ meristems ដែលបានរាយបញ្ជី ទីមួយដែលលេចឡើងក្នុង ontogenesis គឺជា meristem apical ដែលភាពខុសគ្នានៃជាលិកាផ្សេងទៀតទាំងអស់កើតឡើង។ វាមានទីតាំងនៅផ្នែកខាងលើ (APEX) នៃដើម និងឫស និងសាខាក្រោយរបស់វា។ លក្ខណៈពិសេសនៃសកម្មភាពជីវិតរបស់សារពាង្គកាយរុក្ខជាតិគឺជាពិសេសការរក្សាសកម្មភាពទ្រង់ទ្រាយនៅក្នុង meristems apical នៅទូទាំង ontogenesis ទាំងមូល ដូច្នេះពួកគេនិយាយថារុក្ខជាតិមានសមត្ថភាពលូតលាស់គ្មានដែនកំណត់។
រចនាសម្ព័ន្ធនៃ meristem apical (ចំណុចលូតលាស់)
1 - ផ្នែកកណ្តាល
2 - គ្រឿងកុំព្យូទ័រ
2. meristem apical
4. meristem អន្តរកាល
intercalary meristem (មានន័យដូច - intercalary meristem) - កោសិកាមានទីតាំងស្ថិតនៅរវាងតំបន់នៃជាលិកាផ្សេងគ្នាដែលផ្តល់នូវការលូតលាស់ intercalary ។
ឧទាហរណ៍ meristem intercalary មានទីតាំងស្ថិតនៅក្នុងថ្នាំងនៃធញ្ញជាតិនៅមូលដ្ឋាននៃ petioles និង stamen filaments ។ ទាំងនេះគឺជា meristems បឋមដែលនៅសល់។ ពួកវាកើតឡើងពី meristems ខ្សែពួរ ប៉ុន្តែការបំប្លែងរបស់ពួកគេទៅជាជាលិកាអចិន្ត្រៃយ៍ត្រូវបានពន្យារពេលបើប្រៀបធៀបទៅនឹងជាលិកាដើមផ្សេងទៀត។ meristems ឆ្ងាញ់ទាំងនេះគឺគួរឱ្យកត់សម្គាល់ជាពិសេសនៅក្នុងធញ្ញជាតិ។ ក្នុងករណីនំប៉័ងពួកគេធានានូវការចិញ្ចឹមដើមដោយសារតែការបែងចែកកោសិកាមិនស្មើគ្នាពីផ្នែកខាងក្រោមនិងខាងលើនៃចំបើង។
5. របួស meristem
បង្កើតឡើងនៅពេលដែលជាលិកា និងសរីរាង្គត្រូវបានរងរបួស។ កោសិការស់នៅជុំវិញតំបន់ដែលរងផលប៉ះពាល់បានបែងចែក ហើយចាប់ផ្តើមបែងចែក ពោលគឺពួកវាប្រែទៅជា meristem ទីពីរ។ ទម្រង់មុខរបួស កាឡូស- ជាលិកាក្រាស់នៃពណ៌ខៀវ ឬពណ៌លឿង ដែលមានកោសិកា parenchyma ដែលមានទំហំខុសៗគ្នា រៀបចំតាមរបៀបមិនសមរម្យ។ កោសិកា Callus មានស្នូលធំ និងជញ្ជាំងកោសិកាក្រាស់។ callus អាចបង្កើតជាលិការុក្ខជាតិ ឬសរីរាង្គណាមួយ។ ដោតត្រូវបានបង្កើតឡើងនៅតាមបរិវេណ; ឫសនិងពន្លកបន្ថែមអាចត្រូវបានបង្កើតឡើងនៅក្នុង Kalus ។ Callus កើតឡើងនៅពេលដែលរុក្ខជាតិត្រូវបាន grafted និងធានាការរីកលូតលាស់នៃ sion និង rootstock; នៅលើមូលដ្ឋាននៃ cuttings នេះ។ Calus ក៏ត្រូវបានគេប្រើដើម្បីទទួលបានវប្បធម៌ជាលិកាដាច់ស្រយាលផងដែរ។