ចងចាំពីសៀវភៅសិក្សា "រុក្ខជាតិ។ បាក់តេរី។ ផ្សិត និង lichens” ដែលដំណើរការកំណត់លក្ខណៈជីវិតរបស់កោសិកា។ តើអ្វីជារចនាសម្ព័ន្ធនៃស្នូលកោសិកា? តើក្រូម៉ូសូមជាអ្វី? តើអ្វីជារចនាសម្ព័ន្ធនៃម៉ូលេគុល DNA? តើការចម្លង DNA គឺជាអ្វី?
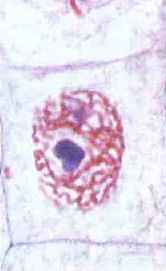
រយៈពេលនៃសកម្មភាពកោសិកាចាប់ពីពេលដើមរហូតដល់ស្លាប់ត្រូវបានគេហៅថា វដ្តជីវិតកោសិកា ឬវដ្តកោសិកា។ ក្នុងអំឡុងពេលនេះ ការលូតលាស់កោសិកា ការអភិវឌ្ឍន៍ និងការបន្តពូជកើតឡើង។ រយៈពេលនៃវដ្តកោសិកានៅក្នុងកោសិកាផ្សេងៗគ្នា សូម្បីតែនៅក្នុងសារពាង្គកាយដូចគ្នាក៏ខុសគ្នាដែរ។ ឧទាហរណ៍រយៈពេលនៃវដ្តនេះនៅក្នុងកោសិកាជាលិកា epithelial របស់មនុស្សគឺប្រហែល 10-15 ម៉ោងហើយនៅក្នុងកោសិកាថ្លើមពេញមួយឆ្នាំ។ វដ្តកោសិកាមានចន្លោះពេលពីរនៃរយៈពេលខុសៗគ្នា៖ អន្តរដំណាក់កាល និងការបែងចែកកោសិកា (រូបភាព 66) ។
អង្ករ។ 66. វដ្តជីវិតកោសិកា (វដ្តកោសិកា): 1 - interphase; 2 - ជំងឺ mitosis
អន្តរដំណាក់កាល។ផ្នែកនៃវដ្តជីវិតរបស់កោសិការវាងការបែងចែកបន្តបន្ទាប់គ្នាពីរត្រូវបានគេហៅថា interphase (ពីឡាតាំងអន្តរ - រវាងនិងដំណាក់កាលក្រិក - រូបរាង) ។ វាត្រូវបានកំណត់លក្ខណៈដោយដំណើរការមេតាបូលីសសកម្ម ជីវសំយោគនៃប្រូតេអ៊ីន អាស៊ីត nucleic កាបូអ៊ីដ្រាត និង lipid ។ នៅក្នុង interphase ដំណើរការដែលទាក់ទងនឹងជីវិតរបស់កោសិកាកើតឡើង - dissimilation និង assimilation ។ ការផ្គត់ផ្គង់ថាមពលនៅក្នុងកោសិកាកើនឡើងដោយសារតែការសំយោគ ATP ។ គ្រប់ប្រភេទនៃ RNA ត្រូវបានសំយោគយ៉ាងសកម្មនៅក្នុងស្នូល ហើយ ribosomes ត្រូវបានបង្កើតឡើង និងប្រមូលផ្តុំនៅក្នុង nucleolus ។ កោសិកាឆ្លងកាត់ការលូតលាស់ដែលពឹងផ្អែកខ្លាំង ហើយចំនួននៃសរីរាង្គទាំងអស់របស់វាកើនឡើង។
ព្រឹត្តិការណ៍សំខាន់នៃ interphase គឺការចម្លង DNA - ការចម្លងដោយខ្លួនឯង។ នេះជារបៀបដែលកោសិការៀបចំដើម្បីបែងចែក។
រយៈពេលនៃ interphase អាស្រ័យលើប្រភេទកោសិកា ហើយជាមធ្យមគឺយ៉ាងហោចណាស់ 90% នៃពេលវេលាសរុបនៃវដ្តកោសិកា។ បន្ទាប់ពីការបញ្ចប់នៃ interphase កោសិកាចូលទៅក្នុងផ្នែកបន្ទាប់នៃវដ្ត - ការបែងចែក។
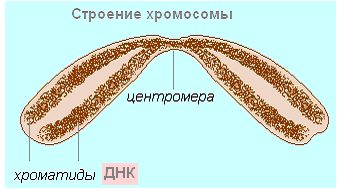
រចនាសម្ព័ន្ធនៃក្រូម៉ូសូម។ក្រូម៉ូសូមដើរតួយ៉ាងសំខាន់ក្នុងវដ្តកោសិកា។ ក្រូម៉ូសូមគឺជាស្មុគ្រស្មាញនៃម៉ូលេគុល DNA និងប្រូតេអ៊ីនដែលមានរាងជារង្វង់ (មកពីក្រូម៉ូសូមក្រិក - ពណ៌និងសូម៉ូ - រាងកាយ) ។ ពួកគេមិនត្រឹមតែគ្រប់គ្រងដំណើរការមេតាបូលីសទាំងអស់នៅក្នុងកោសិកាប៉ុណ្ណោះទេ ប៉ុន្តែថែមទាំងធានានូវការផ្ទេរព័ត៌មានតំណពូជពីកោសិកា និងសារពាង្គកាយមួយជំនាន់ទៅអ្នកដទៃទៀតផង។ កោសិកា prokaryotic មានម៉ូលេគុល DNA រាងជារង្វង់តែមួយគត់ដែលមិនមានទំនាក់ទំនងជាមួយប្រូតេអ៊ីន។ ដូច្នេះវាមិនអាចត្រូវបានគេហៅថាក្រូម៉ូសូមទេ។
អង្ករ។ 67. ខ្សែស្រឡាយ Chromatin នៅក្នុង interphase នៃវដ្តជីវិតកោសិកា
ក្រូម៉ូសូមភាគច្រើននៅក្នុង interphase គឺនៅក្នុងទម្រង់នៃ chromatin threads ដែលធ្វើឱ្យពួកវាមើលមិនឃើញជាក់ស្តែង (រូបភាព 67) ។ បន្ទាប់ពីការចម្លងឡើងវិញ ក្រូម៉ូសូមនីមួយៗមានម៉ូលេគុល DNA ពីរដែលតំរៀបស្លឹក ភ្ជាប់ជាមួយប្រូតេអ៊ីន និងទទួលបានរាងខុសៗគ្នា។ ម៉ូលេគុល DNA របស់កូនស្រីទាំងពីរត្រូវបានខ្ចប់ដោយឡែកពីគ្នា ហើយបង្កើតជា chromatids បងប្អូនស្រី (មកពីភាសាក្រិក chromium - ពណ៌ និង eidos - រូបរាង)។ Sister chromatids ត្រូវបាននៅជាមួយគ្នានិងបង្កើតជាក្រូម៉ូសូមមួយ (រូបភាព 68) ។ ទីតាំងនៃភាពស្អិតរមួតរវាងក្រូម៉ាទីតបងប្អូនស្រីពីរនាក់ត្រូវបានគេហៅថា centromere (ពីកណ្តាលឡាតាំង - កណ្តាលនិង meros - ផ្នែក) ។
អង្ករ។ 68. រចនាសម្ព័ន្ធក្រូម៉ូសូមបន្ទាប់ពីការចម្លង DNA: 1 - centromere: 2 - chromosome arms; 3 - បងស្រី chromatids; 4 - ម៉ូលេគុល DNA: 5 - ប្រូតេអ៊ីន
វាអាចទៅរួចដើម្បីសិក្សារូបរាង និងទំហំនៃក្រូម៉ូសូម និងកំណត់ចំនួនរបស់វានៅក្នុងកោសិកាតែក្នុងអំឡុងពេលបែងចែក នៅពេលដែលពួកវាត្រូវបានខ្ចប់ជាអតិបរមា ខ្ចប់យ៉ាងតឹង មានស្នាមប្រឡាក់ល្អ និងអាចមើលឃើញដោយមីក្រូទស្សន៍ពន្លឺ។
សំណុំក្រូម៉ូសូមនៃកោសិកា។កោសិកានៃសារពាង្គកាយនីមួយៗមានសំណុំជាក់លាក់នៃក្រូម៉ូសូមដែលត្រូវបានគេហៅថា karyotype (ពីភាសាក្រិច karyon - nucleus និង typos - គំរូរូបរាង) ។ ប្រភេទនៃសារពាង្គកាយនីមួយៗមាន karyotype របស់វា។ ក្រូម៉ូសូមនៃ karyotypes ខុសគ្នានៅក្នុងរូបរាង ទំហំ និងសំណុំនៃព័ត៌មានហ្សែន។ សំណុំក្រូម៉ូសូមគឺមានលក្ខណៈបុគ្គលយ៉ាងតឹងរ៉ឹងសម្រាប់ប្រភេទនីមួយៗនៃសារពាង្គកាយ។ ដូច្នេះ karyotype របស់មនុស្សមានក្រូម៉ូសូម 23 គូ (រូបភាព 69) រុយផ្លែឈើ Drosophila មានក្រូម៉ូសូម 4 គូ ហើយប្រភេទស្រូវសាលីមួយគូមាន 14 គូ។
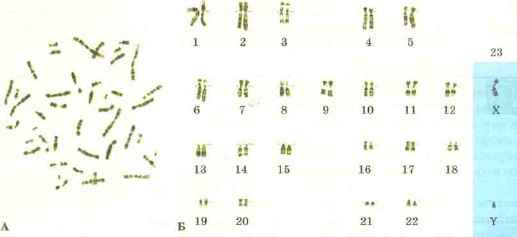
អង្ករ។ 69. សំណុំក្រូម៉ូសូមនៃកោសិកាមនុស្ស: A - រូបថតទូទៅ; ខ - ក្រូម៉ូសូមចំនួន ២៣ គូ
ការសិក្សាអំពី karyotypes នៃសារពាង្គកាយជាច្រើនបានបង្ហាញថាកោសិការបស់ពួកគេអាចមានក្រូម៉ូសូមទ្វេ និងតែមួយ។
សំណុំក្រូម៉ូសូមទ្វេតែងតែមានក្រូម៉ូសូមគូដែលដូចគ្នាបេះបិទក្នុងទំហំ រូបរាង និងធម្មជាតិនៃព័ត៌មានតំណពូជ។ ក្រូម៉ូសូមគូត្រូវបានគេហៅថា homologous (ពីភាសាក្រិក homos - ដូចគ្នា) ។ ដូច្នេះ កោសិកាមនុស្សមិនបន្តពូជទាំងអស់មានក្រូម៉ូសូម 23 គូ ពោលគឺក្រូម៉ូសូមចំនួន 46 ត្រូវបានបង្ហាញក្នុងទម្រង់ 23 គូ។ នៅក្នុង Drosophila ក្រូម៉ូសូមចំនួន 8 បង្កើតបាន 4 គូ។ ក្រូម៉ូសូមដែលបានផ្គូផ្គងមានរូបរាងស្រដៀងគ្នាខ្លាំងណាស់។ centromeres របស់ពួកគេស្ថិតនៅកន្លែងដូចគ្នា ហើយហ្សែនរបស់ពួកគេស្ថិតនៅក្នុងលំដាប់ដូចគ្នា។
កោសិកាខ្លះអាចមានសំណុំក្រូម៉ូសូមតែមួយ។ ឧទាហរណ៍នៅក្នុងកោសិកានៃរុក្ខជាតិទាប - សារាយពណ៌បៃតង unicellular - សំណុំនៃក្រូម៉ូសូមគឺនៅលីវខណៈពេលដែលនៅក្នុងរុក្ខជាតិនិងសត្វខ្ពស់ជាងវាមានទ្វេដង។ កោសិកាផ្លូវភេទសត្វក៏មានសំណុំក្រូម៉ូសូមតែមួយផងដែរ។ ក្នុងករណីនេះ មិនមានក្រូម៉ូសូមផ្គូផ្គង គ្មានក្រូម៉ូសូមដូចគ្នា ប៉ុន្តែមានក្រូម៉ូសូមដែលមិនមានលក្ខណៈដូចគ្នា។ ដូច្នេះកោសិកាមេរោគរបស់មនុស្សមាន 23 ក្រូម៉ូសូម។ លើសពីនេះទៅទៀត សំណុំក្រូម៉ូសូមនៃកោសិកាមេជីវិតឈ្មោល និងស្ត្រីមានភាពខុសគ្នានៅក្នុងក្រូម៉ូសូមទី 23 ។ វាប្រហាក់ប្រហែលនឹងរាងអក្សរឡាតាំង X ឬ Y ។ មេជីវិតឈ្មោលអាចមានក្រូម៉ូសូម X ឬ Y ។ ស៊ុតតែងតែផ្ទុកក្រូម៉ូសូម X ។
សំណុំក្រូម៉ូសូមជាធម្មតាត្រូវបានតំណាងដោយអក្សរឡាតាំង p សំណុំទ្វេត្រូវបានតំណាងឱ្យស្របតាម 2p និងសំណុំតែមួយ - ទំ។
លំហាត់ផ្អែកលើសម្ភារៈគ្របដណ្តប់
- កំណត់វដ្តជីវិតនៃកោសិកា (វដ្តកោសិកា) ។
- តើអ្វីជា inferphase? តើព្រឹត្តិការណ៍សំខាន់មួយណាក្នុងដំណាក់កាលអន្តរកាល? បញ្ជាក់ចម្លើយរបស់អ្នក។
- តើម៉ូលេគុល DNA ប៉ុន្មានដែលក្រូម៉ូសូមមាននៅដើមនៃ interphase និងមុនការបែងចែកកោសិកា?
- តើចំនួន និងរូបរាងរបស់ក្រូម៉ូសូមត្រូវបានកំណត់ក្នុងប្រភេទផ្សេងៗនៃសារពាង្គកាយយ៉ាងដូចម្តេច?
- តើសំណុំក្រូម៉ូសូមពីរខុសគ្នាពីសំណុំតែមួយយ៉ាងដូចម្តេច?
- មានក្រូម៉ូសូមចំនួន 44 នៅក្នុង karyotype របស់ទន្សាយ។ តើមានក្រូម៉ូសូមប៉ុន្មាននៅក្នុងកោសិកាមិនបន្តពូជរបស់ទន្សាយ ហើយតើមានប៉ុន្មាននៅក្នុងកោសិកាមេជីវិត?
វាត្រូវបានរៀបរាប់ខាងលើរួចហើយថានៅក្នុងស្នូលកោសិកា ម៉ូលេគុល DNA មានទីតាំងនៅក្នុងរចនាសម្ព័ន្ធពិសេសហៅថា ក្រូម៉ូសូម. ការស្រាវជ្រាវរបស់ពួកគេបានចាប់ផ្តើមជាង 100 ឆ្នាំមុនដោយប្រើមីក្រូទស្សន៍ពន្លឺធម្មតា។ នៅចុងបញ្ចប់នៃសតវត្សទី 19 អ្វីមួយបានក្លាយទៅជាច្បាស់លាស់អំពីអាកប្បកិរិយារបស់ក្រូម៉ូសូមក្នុងអំឡុងពេលការបែងចែកកោសិកាហើយគំនិតនេះត្រូវបានបង្ហាញអំពីការចូលរួមរបស់ពួកគេក្នុងការបញ្ជូនតំណពូជ។
ក្រូម៉ូសូមអាចមើលឃើញនៅក្នុងមីក្រូទស្សន៍ នៅពេលដែលកោសិកាមួយបែងចែកនៅដំណាក់កាលជាក់លាក់មួយនៃវដ្តកោសិកាដែលហៅថា មីតូស៊ីស. ក្រូម៉ូសូមនៅក្នុងរដ្ឋនេះគឺជារចនាសម្ព័ន្ធរាងជាដំបងដែលមានប្រវែងខុសៗគ្នាជាមួយនឹងកម្រាស់មិនទៀងទាត់ ក្រូម៉ូសូមភាគច្រើនមានការរឹតបន្តឹងដែលបែងចែកក្រូម៉ូសូមជាពីរដៃ។ នៅក្នុងតំបន់នៃ constriction មានរចនាសម្ព័ន្ធសំខាន់សម្រាប់ការកើនឡើងទ្វេដងនៃក្រូម៉ូសូមដែលហៅថា កណ្តាល. នៅពេលដែលកោសិកាមួយបែងចែកកំឡុងពេល mitosis ចំនួននៃក្រូម៉ូសូមកើនឡើងទ្វេដង ដែលជាលទ្ធផលនៃកោសិកាដែលទើបបង្កើតថ្មីទាំងពីរត្រូវបានផ្តល់ឱ្យនូវសំណុំក្រូម៉ូសូមស្តង់ដារដូចគ្នា។
មានតែនៅក្នុងឆ្នាំ 1956 ជាលើកដំបូង Y. Tio និង A. Levan បានពិពណ៌នាអំពីសំណុំក្រូម៉ូសូមរបស់មនុស្ស កំណត់សមាសភាពបរិមាណនៃក្រូម៉ូសូម និងផ្តល់នូវលក្ខណៈទូទៅនៃរូបវិទ្យារបស់ពួកគេ។ តាមពិត ការងារទាំងនេះបានសម្គាល់ការចាប់ផ្តើមនៃការសិក្សាអំពីរចនាសម្ព័ន្ធនៃហ្សែនរបស់មនុស្ស។ នៅក្នុងមនុស្ស កោសិកានីមួយៗនៃរាងកាយមានក្រូម៉ូសូមចំនួន 46 ដែលប្រវែងរាងកាយមានចាប់ពី 1.5 ដល់ 10 microns (រូបភាព 7) ។
អង្ករ។ ៧. ទិដ្ឋភាពមីក្រូទស្សន៍នៃសំណុំក្រូម៉ូសូមពេញលេញដែលមាននៅក្នុងស្នូលនៃកោសិកាមនុស្សនីមួយៗ
ចូរយើងរំលឹកអ្នកអានថាសំណុំនៃក្រូម៉ូសូមនៅក្នុងកោសិកាមនុស្សទាំងអស់ (លើកលែងតែកោសិកាផ្លូវភេទ) ត្រូវបានគេហៅថា diploid (ទ្វេ) ចាប់តាំងពីក្រូម៉ូសូមនីមួយៗត្រូវបានតំណាងដោយពីរច្បាប់ចម្លង (សរុប 23 គូ) ។ កោសិកា somatic របស់មនុស្សនីមួយៗ (លើកលែងតែកោសិកាឈាមក្រហម) មាន 2 សំណុំក្រូម៉ូសូមពេញលេញ។ សំណុំនីមួយៗ (haploid) មានក្រូម៉ូសូម 23 - ក្រូម៉ូសូមធម្មតា 22 (អូតូសូម) និងក្រូម៉ូសូមភេទមួយ - X ឬ Y. ដូច្នេះ ហ្សែនរបស់មនុស្សម្នាក់ៗមាន 23 គូនៃម៉ូលេគុល DNA យក្សដែលចែកចាយក្នុងក្រូម៉ូសូមផ្សេងៗគ្នា ហើយប្រសិនបើយើងនិយាយ អំពីហ្សែនរបស់មនុស្សជាទូទៅ (បុរស និងស្ត្រី) បន្ទាប់មកចំនួនសរុបនៃម៉ូលេគុលបែបនេះគឺ 24 ។ នេះគឺជាព័ត៌មានមូលដ្ឋានដំបូងគេដែលត្រូវបានទទួលអំពីហ្សែនរបស់មនុស្សដោយការវិភាគក្រូម៉ូសូម។
ការសិក្សាអំពីរចនាសម្ព័ន្ធ (ទំហំ និងរូបរាង) នៃក្រូម៉ូសូមរបស់មនុស្ស បានបង្ហាញថា ភាគច្រើននៃពួកវាស្រដៀងទៅនឹង skittles នៅក្នុងរូបរាង ដែលមានផ្នែកក្រាស់ពីរ (chromatids) និងការរឹតបន្តឹងស្តើង (centromeres) រវាងពួកវា។ ភាពស្រដៀងគ្នាជាមួយ skittles និងមិនមែនជាមួយ dumbbells គឺថា centromere ភាគច្រើនមិនមានទីតាំងនៅកណ្តាលនៃក្រូម៉ូសូមនោះទេប៉ុន្តែត្រូវបានផ្លាស់ប្តូរទៅចុងម្ខាងរបស់វា។ ទំហំក្រូម៉ូសូមប្រែប្រួលយ៉ាងខ្លាំង ដោយក្រូម៉ូសូមខ្លីបំផុតមានទំហំតូចជាងប្រហែលដប់ដង។ នេះគឺជាព័ត៌មានសំខាន់ទីពីរអំពីរចនាសម្ព័ន្ធនៃហ្សែនរបស់មនុស្ស - ម៉ូលេគុល DNA 24 ដែលបង្កើតវាមានទំហំខុសៗគ្នា។
ប្រសិនបើអ្នកប្រៀបធៀបចំនួន និងទំហំនៃក្រូម៉ូសូមនៅក្នុងមនុស្ស និងនៅក្នុងប្រភេទសារពាង្គកាយផ្សេងទៀត អ្នកអាចមើលឃើញភាពខុសគ្នាយ៉ាងខ្លាំង។ ឧទាហរណ៍ គោមួយក្បាលដែលទំហំហ្សែនប្រហែលស្មើនឹងហ្សែនមនុស្ស មានក្រូម៉ូសូម ៦០ គូ។ កង្កែបក្រញ៉ាំមានក្រូម៉ូសូមតែ ១៨ ប៉ុណ្ណោះ ប៉ុន្តែសូម្បីតែក្រូម៉ូសូមតូចបំផុតក៏ធំជាងក្រូម៉ូសូមធំបំផុតរបស់មនុស្សដែរ។ នៅក្នុងបក្សី ផ្ទុយទៅវិញចំនួនក្រូម៉ូសូមឈានដល់ 40 ឬច្រើនជាងនេះ ហើយពួកវាទាំងអស់មានទំហំតូចណាស់។ ដូច្នេះភាពចម្រុះនៃក្រូម៉ូសូមនៅក្នុងធម្មជាតិមានទំហំធំណាស់។
ដោយប្រើមីក្រូទស្សន៍ពន្លឺ ទំហំនៃក្រូម៉ូសូមរបស់មនុស្សទាំងអស់ត្រូវបានកំណត់។ បន្ទាប់មក ក្រូម៉ូសូមមិនមែនភេទទាំងអស់ត្រូវបានដាក់លេខតាមលំដាប់លំដោយនៃទំហំថយចុះពី 1 ដល់ 22។ ក្រូម៉ូសូមភេទមិនត្រូវបានកំណត់ជាលេខទេ ប៉ុន្តែត្រូវបានគេដាក់ឈ្មោះថា X និង Y។ ដូចដែលការសិក្សាជាបន្តបន្ទាប់បានបង្ហាញឱ្យឃើញកាន់តែត្រឹមត្រូវ ក្រូម៉ូសូម 21 ពិតជាប្រែទៅជាបន្តិច។ តូចជាង 22 ប៉ុន្តែលេខក្រូម៉ូសូមមិនត្រូវបានផ្លាស់ប្តូរទេ (ដើម្បីកុំឱ្យមានការភ័ន្តច្រឡំ)។ ភាពខុសគ្នានៃសំណុំក្រូម៉ូសូមរវាងបុរសនិងស្ត្រីគឺថាស្ត្រីមានក្រូម៉ូសូម X ភេទពីរ (ឧទាហរណ៍ក្រូម៉ូសូមក្នុង 23 គូគឺដូចគ្នា) ហើយចំពោះបុរសគូដែលមានក្រូម៉ូសូម X ត្រូវបានបង្កើតឡើងដោយក្រូម៉ូសូមភេទបុរស - Y ក្រូម៉ូសូមនីមួយៗអាចត្រូវបានចាត់ទុកថាជាបរិមាណដាច់ដោយឡែកនៃការប្រមូលស្នាដៃចំនួនម្ភៃបួនដែលហៅថា Encyclopedia of Man។
កោសិកាមេរោគរបស់មនុស្ស មិនដូចកោសិកានៃរាងកាយរបស់សារពាង្គកាយមនុស្សពេញវ័យ (កោសិកា somatic) មិនមាន 2 សំណុំនៃបរិមាណនៃអត្ថបទ DNA ទេ ប៉ុន្តែមានតែមួយប៉ុណ្ណោះ។ មុនពេលមានគភ៌ ក្រូម៉ូសូមនីមួយៗ (បរិមាណដាច់ដោយឡែកនៅក្នុងសព្វវចនាធិប្បាយមនុស្ស) នៃមេជីវិតឈ្មោលរបស់ឪពុក និងស៊ុតរបស់ម្តាយ មានជំពូកផ្សេងៗគ្នានៃអត្ថបទ DNA របស់ឪពុកម្តាយរបស់ពួកគេ លាយបញ្ចូលគ្នាក្នុងបន្សំផ្សេងៗគ្នា។ ក្រូម៉ូសូមណាមួយដែលយើងបានទទួលពីឪពុករបស់យើងត្រូវបានបង្កើតឡើងនៅក្នុងពងស្វាសរបស់គាត់មិនយូរប៉ុន្មានមុនពេលយើងមានផ្ទៃពោះ។ ពីមុននៅក្នុងប្រវត្តិសាស្រ្តទាំងមូលរបស់មនុស្សជាតិ ក្រូម៉ូសូមបែបនេះមិនដែលមានទេ។ វាត្រូវបានបង្កើតឡើងតាមរយៈដំណើរការនៃការលាយចំរុះដោយចៃដន្យដែលកើតឡើងកំឡុងពេលបែងចែក ដែលបណ្តើរៗត្រូវបានបង្កើតឡើងពីផ្នែកនៃក្រូម៉ូសូមរបស់បុព្វបុរសនៅខាងឪពុកដែលរួបរួមគ្នាជាមួយគ្នា។ ស្ថានភាពគឺដូចគ្នាជាមួយនឹងក្រូម៉ូសូមនៃស៊ុត លើកលែងតែវាត្រូវបានបង្កើតឡើងនៅក្នុងរាងកាយរបស់ម្តាយយើងមុនពេលយើងកើត (ស្ទើរតែភ្លាមៗបន្ទាប់ពីកំណើតរបស់ម្តាយខ្លួនឯង)។
នៅក្នុង zygote ដែលត្រូវបានបង្កើតឡើងជាលទ្ធផលនៃការលាយបញ្ចូលគ្នានៃមេជីវិតឈ្មោល និងស៊ុតមួយ ហ្សែនរបស់មាតា និងបិតាត្រូវបានលាយបញ្ចូលគ្នា និងសាប់នៅក្នុងបន្សំផ្សេងៗគ្នា។ នេះកើតឡើងជាលទ្ធផលនៃការពិតដែលថាក្រូម៉ូសូមមិននៅតែមិនផ្លាស់ប្តូរពីមួយជំនាន់ទៅមួយជំនាន់ - ពួកគេមានទំនាក់ទំនងជាមួយគូដែលជួបប្រទះដោយចៃដន្យរបស់ពួកគេដោយផ្លាស់ប្តូរសម្ភារៈជាមួយវា។ ដំណើរការដែលកំពុងដំណើរការនេះត្រូវបានគេហៅថា ការផ្សំឡើងវិញ. ហើយមនុស្សជំនាន់ក្រោយច្រើនតែទទួលបានក្រូម៉ូសូមកូនកាត់ - មួយផ្នែកពីជីតា និងផ្នែកពីជីដូន។ បន្ថែមទៀតនៅក្នុងស៊េរីនៃជំនាន់មួយ ផ្លូវហ្សែនប្រសព្វគ្នាឥតឈប់ឈរ និងខុសគ្នា។ ជាលទ្ធផលនៃការលាយបញ្ចូលគ្នានៃស៊ុតតែមួយគត់ជាមួយនឹងមេជីវិតឈ្មោលតែមួយគត់ ហ្សែនដែលមានលក្ខណៈប្លែកពីគេក្នុងគ្រប់ទិដ្ឋភាពទាំងអស់កើតឡើង។ ហើយក្នុងន័យនេះ យើងទាំងអស់គ្នាគឺប្លែក បុគ្គលម្នាក់ៗរក្សាទុកព័ត៌មានហ្សែនតែមួយគត់ ដែលរួមមានការរួមផ្សំដោយចៃដន្យនៃហ្សែនផ្សេងៗគ្នា។
ហ្សែនតែមួយអាចត្រូវបានគេមើលថាជាឯកតាដែលបន្តមានក្នុងជំនាន់ជាច្រើន។ ហើយក្នុងន័យនេះហ្សែនគឺអមតៈ! មានសូម្បីតែទស្សនៈដើមដែលថាវាមិនមែនជាមនុស្សខ្លួនឯងទេ ប៉ុន្តែហ្សែនរបស់ពួកគេដែលគ្រប់គ្រងពិភពលោក ហើយសារពាង្គកាយមានជីវិតជាក់លាក់នីមួយៗបម្រើជាជម្រកបណ្តោះអាសន្នសម្រាប់ពួកគេ។ គំនិតចម្រូងចម្រាសនេះបានមកពី Richard Dawkins អ្នកនិពន្ធសៀវភៅ The Selfish Gene ។ តាមគំនិតរបស់គាត់ ហ្សែនគឺពិតជាអមតៈ មិនដូចសារពាង្គកាយមានជីវិតដែលពួកគេមាននោះទេ។ ហ្សែនខ្លះមានអាយុរាប់សិប ឬរាប់រយលានឆ្នាំ។ ហ្សែន ដើម្បីប្រើវាក្យស័ព្ទរបស់ Dawkins ធ្វើអ្វីគ្រប់យ៉ាងដែលពួកគេអាចធ្វើបានដើម្បីរស់។ ពួកគេសម្របខ្លួនទៅនឹងកំដៅ និងត្រជាក់ ជ្រើសរើសកន្លែងល្អសម្រាប់ខ្លួនគេ ធ្វើចំណាកស្រុកដោយមានជំនួយពីមនុស្ស និងចូលទៅក្នុងបន្សំថ្មី។ បុរសនោះបានក្លាយទៅជាម្ចាស់ដែលមិនស្រួលខ្លួន។ អស់រយៈពេលរាប់ពាន់ឆ្នាំគាត់បានធ្វើដំណើរយ៉ាងទូលំទូលាយនៅជុំវិញពិភពលោកដោយផ្សព្វផ្សាយវត្តមានរបស់គាត់ឥទ្ធិពលនិងវត្ថុរបស់គាត់ - ហ្សែន។ (អ្នកអានដែលចង់ដឹងចង់ឃើញអាចស្វែងយល់បន្ថែមអំពីគំនិត និងការវែកញែករបស់ R. Dawkins នៅក្នុង ឧបសម្ព័ន្ធ ១) ទស្សនៈនេះគឺនៅឆ្ងាយពីមិនអាចប្រកែកបាន ហើយពីការបង្ហាញបន្ថែមទៀត វានឹងកាន់តែច្បាស់សម្រាប់ពួកយើងថា ហ្សែនគឺជាមនុស្សដំបូងគេ មិនមែនជាមនុស្សអាត្មានិយមទេ ប៉ុន្តែជាអ្នកធ្វើការ។ មានហ្សែនដែលជា "អាណាព្យាបាល" នៃហ្សែន ហ្សែនដែលជា "អ្នកថែទាំ" ហ្សែនដែលជា "អ្នកចំអិន" និងហ្សែនដែលជា "អ្នកថែរក្សាផ្ទះ" ។ តាមរយៈការធានាអត្ថិភាពរបស់ពួកគេ ពួកគេធានានូវអត្ថិភាពរបស់យើង។
ភ្លាមៗបន្ទាប់ពីការមានគភ៌ មនុស្សអនាគតគឺគ្រាន់តែជាកោសិកាមួយ (ហ្សីហ្គោត) ដែលផ្តល់ដោយបណ្ណាល័យ DNA ដំបូងមួយដែលមាន 46 ភាគ។ ក្នុងចំណោម 46 ភាគ 23 តែងតែទទួលបានពីឪពុកនិង 23 ផ្សេងទៀតពីម្តាយ។ អត្ថបទនៃភាគ 23 ប៉ា និង 23 របស់មាតា ថ្វីត្បិតតែជាទូទៅស្រដៀងគ្នាខ្លាំងណាស់ យ៉ាងណាក៏ដោយ ខុសគ្នានៅក្នុងព័ត៌មានលម្អិត។ ជាឧទាហរណ៍ នៅក្នុងសៀវភៅប៉ាក់ លេខ 18 នៅទំព័រ 253 មានប្រយោគបញ្ជាមួយ (ក្នុងទម្រង់ហ្សែន) ដែលនិយាយថា ភ្នែករបស់កុមារគួរតែមានពណ៌ត្នោត ហើយក្នុងបរិមាណមាតាដូចគ្នានៅលើទំព័រដូចគ្នា វាក៏ត្រូវបានសរសេរផងដែរ។ អំពីពណ៌ភ្នែក ប៉ុន្តែយោងទៅតាមពណ៌អត្ថបទនេះគួរតែជាពណ៌ខៀវ។ ការចង្អុលបង្ហាញដំបូងគឺតឹងរ៉ឹងជាង (លេចធ្លោ) ជាងទីពីរហើយជាលទ្ធផលភ្នែករបស់កុមារនឹងមានពណ៌ត្នោត។ ហ្សែនដែលកំណត់សិទ្ធិរបស់វាត្រូវបានគេហៅថា លេចធ្លោហើយអ្នកដែលលះបង់សិទ្ធិរបស់គាត់ - ធ្លាក់ចុះ. មានតែមនុស្សទាំងនោះដែលមានអត្ថបទទាំងមាតា និងបិតាដែលមានហ្សែនដែលបង្ហាញអំពីភ្នែកពណ៌ខៀវមានពណ៌ភ្នែកពណ៌ខៀវ។ បន្ទាប់មក zygote បែងចែកជាពីរកោសិកា ពួកវានីមួយៗបែងចែកម្តងទៀត ហើយបន្តរហូតដល់កោសិការាប់ពាន់លានលេចឡើង។ ដំណើរការនៃការបែងចែកកោសិកាត្រូវបានបង្ហាញជាគ្រោងការណ៍នៅក្នុងរូបភព។ ៨.
ជាមួយនឹងការបែងចែកកោសិកានីមួយៗ បរិមាណនៃអត្ថបទ DNA ដែលមាននៅក្នុងបណ្ណាល័យត្រូវបានចម្លងយ៉ាងពិតប្រាកដ ដោយស្ទើរតែគ្មានកំហុស។ រាងកាយរបស់មនុស្សពេញវ័យមានជាមធ្យម 10 14 កោសិកា។ ជាឧទាហរណ៍ មានកោសិកាប្រហែល 10 ពាន់លាននៅក្នុងខួរក្បាល និងថ្លើម និង 300 ពាន់លានកោសិកានៅក្នុងប្រព័ន្ធភាពស៊ាំ។ ក្នុងអំឡុងពេលពេញមួយជីវិតរបស់មនុស្ស ការបែងចែកកោសិកាប្រហែល 10 16 កើតឡើងនៅក្នុងខ្លួនរបស់គាត់។ សមាសភាពកោសិកានៃសរីរាង្គជាច្រើនត្រូវបានបន្តឡើងវិញជាច្រើនដងក្នុងរយៈពេល 70 ឆ្នាំនៃជីវិត។ ហើយកោសិកាទាំងនេះនីមួយៗមាន 46 បរិមាណដូចគ្នានៃអត្ថបទ DNA ។
នៅចុងបញ្ចប់នៃទសវត្សរ៍ទី 60 នៃសតវត្សទី 20 របកគំហើញដ៏សំខាន់មួយត្រូវបានធ្វើឡើងក្នុងការសិក្សាអំពីក្រូម៉ូសូម។ វាគឺដោយសារតែការពិតដែលថាពួកគេបានចាប់ផ្តើមប្រើភ្នាក់ងារកម្រិតពណ៌ពិសេសសម្រាប់ការដាក់ពណ៌របស់ពួកគេ - mustard akrichine ហើយបន្ទាប់មកសមាសធាតុផ្សេងទៀតដែលស្រដៀងនឹងវា។ ស្នាមប្រឡាក់នេះបានធ្វើឱ្យវាអាចកំណត់អត្តសញ្ញាណមួយចំនួនធំនៃរចនាសម្ព័ន្ធផ្សេងគ្នានៅក្នុងក្រូម៉ូសូមដែលមិនអាចមើលឃើញនៅក្រោមមីក្រូទស្សន៍ដោយគ្មានស្នាមប្រឡាក់។ បន្ទាប់ពីលាបពណ៌ក្រូម៉ូសូមជាមួយនឹងការជ្រលក់ពណ៌ Giemsa-Romanovsky ជាក់លាក់ ពួកវាមើលទៅដូចជាសេះបង្កង់៖ ឆ្នូតពន្លឺឆ្លងកាត់ និងងងឹតនៃអាំងតង់ស៊ីតេពណ៌ខុសៗគ្នាអាចមើលឃើញនៅតាមបណ្តោយប្រវែងទាំងមូល។
អង្ករ។ ៨. ដំណាក់កាលសំខាន់នៃវដ្តកោសិកាដែលនាំទៅដល់ការបែងចែកកោសិកា
ក្រុមតន្រ្តីទាំងនេះត្រូវបានគេហៅថា chromosomal G segments ឬ bands (រូបភាព 9) ។ គំរូនៃការបែងចែកមានភាពខុសប្លែកគ្នាយ៉ាងខ្លាំងក្នុងចំណោមក្រូម៉ូសូមផ្សេងៗគ្នា ប៉ុន្តែការរៀបចំនៃផ្នែកក្រូម៉ូសូមគឺថេរនៅលើក្រូម៉ូសូមនីមួយៗនៅក្នុងគ្រប់ប្រភេទនៃកោសិកាមនុស្ស។
លក្ខណៈនៃស្នាមឆ្នូតដែលបង្ហាញដោយការប្រឡាក់មិនទាន់ច្បាស់លាស់នៅឡើយទេ។ ឥឡូវនេះវាត្រូវបានបង្កើតឡើងតែប៉ុណ្ណោះដែលថាតំបន់នៃក្រូម៉ូសូមដែលត្រូវគ្នានឹងក្រុមងងឹត (ហៅថា R-bands) ចម្លងមុនជាងតំបន់ពន្លឺ (ហៅថា G-bands)។ ដូច្នេះ ការភ្ជាប់ក្រូម៉ូសូមភាគច្រើននៅតែមានអត្ថន័យដែលមិនទាន់យល់ច្បាស់នៅឡើយ។
ស្នាមប្រឡាក់ក្រូម៉ូសូមជួយសម្រួលការកំណត់អត្តសញ្ញាណរបស់ពួកគេយ៉ាងខ្លាំង ហើយក្រោយមកបានរួមចំណែកក្នុងការកំណត់ទីតាំងនៃហ្សែននៅលើពួកវា (ការធ្វើផែនទីហ្សែន)។
អង្ករ។ ៩. ក្រូម៉ូសូមជាក់លាក់ G-segments កំណត់ដោយស្នាមប្រឡាក់ក្រូម៉ូសូមរបស់មនុស្ស និងប្រព័ន្ធនៃការកំណត់របស់វាយោងទៅតាមការសម្រេចចិត្តរបស់សន្និសីទអន្តរជាតិនៅទីក្រុងប៉ារីសក្នុងឆ្នាំ 1971។ លេខខាងក្រោមក្រូម៉ូសូមបង្ហាញពីលេខរបស់វា។ X និង Y - ក្រូម៉ូសូមភេទ, ទំ - ដៃខ្លី, q - ដៃវែងនៃក្រូម៉ូសូម
ទោះបីជាដំណើរការលម្អិតដែលកើតឡើងកំឡុងពេលស្នាមប្រឡាក់មិនទាន់ច្បាស់ទាំងស្រុងក៏ដោយ វាច្បាស់ណាស់ថាលំនាំពណ៌អាស្រ័យលើប៉ារ៉ាម៉ែត្រដូចជាមាតិកាកើនឡើង ឬថយចុះនៃគូ AT ឬ GC នៅក្នុងក្រុមក្រូម៉ូសូមនីមួយៗ។ ហើយនេះគឺជាព័ត៌មានទូទៅមួយទៀតអំពីហ្សែន - វាមិនដូចគ្នាទេ វាមានតំបន់ដែលសំបូរទៅដោយគូនុយក្លេអូទីតជាក់លាក់។
ជាពិសេស នេះអាចបណ្តាលមកពីការធ្វើឡើងវិញនៃប្រភេទមួយចំនួននៃ DNA nucleotide បន្តបន្ទាប់គ្នានៅក្នុងតំបន់មួយចំនួន។
ភាពខុសគ្នានៃពណ៌នៃក្រូម៉ូសូមបានរកឃើញកម្មវិធីធំទូលាយសម្រាប់ការរកឃើញ និងការកំណត់អត្តសញ្ញាណនៃការផ្លាស់ប្តូរបុគ្គលតូចៗនៅក្នុងហ្សែនរបស់មនុស្សជាក់លាក់មួយ ( polymorphismជាពិសេសនាំទៅរករោគសាស្ត្រផ្សេងៗ។ ឧទាហរណ៏នៃការនេះគឺជាការរកឃើញនៃក្រូម៉ូសូម Philadelphia ដែលត្រូវបានគេរកឃើញនៅក្នុងអ្នកជំងឺដែលមានជំងឺមហារីកឈាម myeloid រ៉ាំរ៉ៃ។ ដោយប្រើស្នាមប្រឡាក់ក្រូម៉ូសូម វាត្រូវបានបង្កើតឡើងថា ចំពោះអ្នកជំងឺដែលមានជំងឺនេះ បំណែកជាក់លាក់មួយបាត់នៅលើក្រូម៉ូសូម 21 ហើយលេចឡើងនៅចុងបញ្ចប់នៃដៃវែងនៃក្រូម៉ូសូម 9 (ការផ្ទេរបំណែក ឬ ការផ្លាស់ប្តូរទីតាំង, អក្សរកាត់ t) ។ អ្នកសេនេទិចកំណត់ព្រឹត្តិការណ៍ដូចជា t (9; 21) ។ ដូច្នេះ ការវិភាគក្រូម៉ូសូមបង្ហាញថា ម៉ូលេគុល DNA ផ្សេងៗគ្នាអាចផ្លាស់ប្តូរផ្នែកដាច់ដោយឡែកពីគ្នាទៅវិញទៅមក ដែលបណ្តាលឱ្យមានការបង្កើត "កូនកាត់" នៅក្នុងហ្សែនដែលមានម៉ូលេគុល DNA នៃក្រូម៉ូសូមផ្សេងៗគ្នា។ ការវិភាគលើលក្ខណៈសម្បត្តិដែលបានសិក្សារួចហើយនៃក្រូម៉ូសូមបានធ្វើឱ្យវាអាចបង្កើតជាគំនិតនៃពហុវចនៈនៃហ្សែនរបស់មនុស្ស។
ដើម្បីកំណត់ការធ្វើមូលដ្ឋានីយកម្មនៃហ្សែនបុគ្គលនៅលើក្រូម៉ូសូម (នោះគឺការគូសផែនទីហ្សែន) ឃ្លាំងអាវុធទាំងមូលនៃវិធីសាស្រ្តពិសេសដែលជាញឹកញាប់ស្មុគស្មាញក្នុងការរចនា និងការអនុវត្តត្រូវបានប្រើ។ កត្តាសំខាន់មួយគឺការបង្កាត់ម៉ូលេគុល (ការបង្កើតកូនកាត់) នៃហ្សែនមួយ ឬបំណែករបស់វាជាមួយនឹងការត្រៀមក្រូម៉ូសូមដែលបានជួសជុលនៅលើការគាំទ្រដ៏រឹងមាំ ដាច់ដោយឡែកពីកោសិកាក្នុងទម្រង់សុទ្ធ (នេះហៅថាការបង្កាត់ នៅកន្លែង) ខ្លឹមសារនៃវិធីសាស្ត្របង្កាត់ នៅកន្លែងមាននៅក្នុងអន្តរកម្ម (hybridization) រវាង denatured (unbraided) DNA strands នៅក្នុង chromosomes និង nucleotide sequences នៃ chromosomes ដែលត្រូវបានបន្ថែមទៅក្នុងការរៀបចំ, DNA single-stranded បុគ្គល ឬ RNA (ពួកវាត្រូវបានគេហៅថា ការស៊ើបអង្កេត) ប្រសិនបើមានការបំពេញបន្ថែមរវាងខ្សែ DNA ក្រូម៉ូសូមមួយ និងការស៊ើបអង្កេត នោះកូនកាត់ម៉ូលេគុលដែលមានស្ថេរភាពត្រឹមត្រូវត្រូវបានបង្កើតឡើងរវាងពួកវា។ ការស៊ើបអង្កេតត្រូវបានដាក់ស្លាកជាមុនដោយប្រើស្លាកផ្សេងៗគ្នា (វិទ្យុសកម្ម fluorescent ។ល។)។ ទីកន្លែងនៃការបង្កើតកូនកាត់នៅលើក្រូម៉ូសូមត្រូវបានកំណត់អត្តសញ្ញាណដោយទីតាំងនៃសញ្ញាទាំងនេះនៅលើការត្រៀមលក្ខណៈក្រូម៉ូសូម។ ដូច្នេះ សូម្បីតែមុនពេលការមកដល់នៃវិធីសាស្រ្តវិស្វកម្មហ្សែន និងការរៀបចំលំដាប់ DNA ពួកគេបានរកឃើញឧទាហរណ៍ ទីតាំងនៅក្នុងហ្សែនរបស់មនុស្សដែលអ៊ិនកូដ RNAs ribosomal ធំ និងតូច (rRNAs)។ ហ្សែននៃអតីតបានប្រែទៅជាត្រូវបានធ្វើមូលដ្ឋានីយកម្មនៅក្នុងក្រូម៉ូសូមរបស់មនុស្សចំនួនប្រាំផ្សេងគ្នា (13, 14, 15, 21 និង 22) ខណៈពេលដែលភាគច្រើននៃហ្សែន rRNA តូច ( 5 ស RNA) ត្រូវបានប្រមូលផ្តុំនៅកន្លែងមួយនៅលើដៃវែងនៃក្រូម៉ូសូម 1 ។
ឧទាហរណ៍នៃរូបភាពដែលទទួលបានដោយការបង្កាត់នៃការស៊ើបអង្កេតហ្សែនដែលមានស្លាកសញ្ញា fluorescent ត្រូវបានបង្ហាញនៅក្នុងរូបភព។ 10 នៅលើការបញ្ចូលពណ៌។
អង្ករ។ ១០. ការធ្វើកូនកាត់នៃក្រូម៉ូសូមរបស់មនុស្សជាមួយនឹងការស៊ើបអង្កេតហ្សែនដែលមានស្លាកពណ៌ fluorescent ក្រហម និងបៃតង។ ព្រួញបង្ហាញពីទីតាំងនៃហ្សែនដែលត្រូវគ្នានៅចុងនៃក្រូម៉ូសូមពីរផ្សេងគ្នា (ខាងស្តាំខាងលើបង្ហាញពីការពង្រីករូបភាពនៃក្រូម៉ូសូមកូនកាត់)។
ហ្សែនដែលមានទីតាំងនៅលើក្រូម៉ូសូមដូចគ្នាត្រូវបានកំណត់ថាជាហ្សែនដែលភ្ជាប់គ្នា។ ប្រសិនបើហ្សែនមានទីតាំងនៅលើក្រូម៉ូសូមផ្សេងៗគ្នា ពួកគេត្រូវបានទទួលមរតកដោយឯករាជ្យ (ការបែងចែកដោយឯករាជ្យ)។ នៅពេលដែលហ្សែនស្ថិតនៅលើក្រូម៉ូសូមដូចគ្នា (ដែលភ្ជាប់គ្នា) ពួកគេមិនអាចបែងចែកដោយឯករាជ្យបានទេ។ ម្តងម្កាល ការផ្លាស់ប្តូរជាច្រើននៅក្នុងក្រូម៉ូសូមអាចកើតឡើងនៅក្នុងកោសិកាមេរោគដែលជាលទ្ធផលនៃដំណើរការផ្សំឡើងវិញរវាងក្រូម៉ូសូមដូចគ្នា។ ដំណើរការមួយក្នុងចំណោមដំណើរការទាំងនេះត្រូវបានគេហៅថា ឆ្លងកាត់. ដោយសារតែការឆ្លងកាត់ ការផ្សារភ្ជាប់គ្នារវាងហ្សែននៃក្រុមដូចគ្នាគឺមិនពេញលេញទេ។ ហ្សែនដែលជាប់ទាក់ទងគ្នាកាន់តែជិតគ្នា ទំនងជាតិចថាទីតាំងនៃហ្សែនបែបនេះនឹងផ្លាស់ប្តូរចំពោះកុមារបើធៀបនឹងឪពុកម្តាយរបស់ពួកគេ។ ការវាស់ស្ទង់ភាពញឹកញាប់នៃការផ្សំឡើងវិញ (ឆ្លងកាត់) ត្រូវបានប្រើដើម្បីបង្កើតលំដាប់លីនេអ៊ែរនៃហ្សែននៅលើក្រូម៉ូសូមនៅក្នុងក្រុមតំណភ្ជាប់មួយ។ ដូច្នេះ នៅពេលគូសផែនទីក្រូម៉ូសូម វាត្រូវបានកំណត់ដំបូងថាតើហ្សែនទាំងនេះស្ថិតនៅលើក្រូម៉ូសូមដូចគ្នា ដោយមិនបញ្ជាក់ថាមួយណា។ បន្ទាប់ពីយ៉ាងហោចណាស់មួយនៃហ្សែននៃក្រុមតំណភ្ជាប់ដែលបានផ្តល់ឲ្យត្រូវបានធ្វើមូលដ្ឋានីយកម្មលើក្រូម៉ូសូមជាក់លាក់មួយ (ឧទាហរណ៍ការប្រើការបង្កាត់ នៅកន្លែង) វាច្បាស់ណាស់ថាហ្សែនផ្សេងទៀតទាំងអស់នៃក្រុមតំណភ្ជាប់នេះមានទីតាំងនៅលើក្រូម៉ូសូមដូចគ្នា។
ឧទាហរណ៍ដំបូងនៃការតភ្ជាប់នៃហ្សែនជាមួយក្រូម៉ូសូមជាក់លាក់អាចជាការរកឃើញនៃការភ្ជាប់នៃលក្ខណៈតំណពូជមួយចំនួនជាមួយនឹងក្រូម៉ូសូមភេទ។ ដើម្បីបញ្ជាក់ពីការធ្វើមូលដ្ឋានីយកម្មនៃហ្សែននៅលើក្រូម៉ូសូម Y ភេទរបស់បុរស វាគ្រប់គ្រាន់ដើម្បីបង្ហាញថាលក្ខណៈនេះតែងតែត្រូវបានរកឃើញតែចំពោះបុរស ហើយមិនត្រូវបានរកឃើញចំពោះស្ត្រីឡើយ។ ក្រុមតំណភ្ជាប់នៃក្រូម៉ូសូម X របស់ស្ត្រីត្រូវបានកំណត់លក្ខណៈដោយអវត្តមាននៃលក្ខណៈដែលអាចចម្លងពីឪពុកទៅកូនប្រុសនិងមរតកនៃលក្ខណៈមាតា។
សារៈសំខាន់ជាពិសេសសម្រាប់ការសិក្សាហ្សែនរបស់មនុស្សនៅក្នុងដំណាក់កាលដំបូងនៃការស្រាវជ្រាវរបស់វាគឺវិធីសាស្រ្តមួយហៅថា ការបង្កាត់កោសិកា somatic. នៅពេលដែលកោសិកា somatic (មិនបន្តពូជ) របស់មនុស្សត្រូវបានលាយជាមួយកោសិកានៃប្រភេទសត្វដទៃទៀត (ជាញឹកញាប់បំផុត កណ្តុរ ឬកោសិកា hamster ចិនត្រូវបានប្រើសម្រាប់គោលបំណងនេះ) ការលាយនៃស្នូលរបស់ពួកគេ (hybridization) អាចកើតឡើងនៅក្នុងវត្តមាននៃភ្នាក់ងារជាក់លាក់។ នៅពេលដែលកោសិកាកូនកាត់បែបនេះបង្កើតឡើងវិញ ក្រូម៉ូសូមមួយចំនួនត្រូវបានបាត់បង់។ ដោយឧបទ្ទវហេតុដ៏រីករាយមួយសម្រាប់អ្នកពិសោធន៍ នៅក្នុងកោសិកាកូនកាត់កណ្តុរមនុស្ស ក្រូម៉ូសូមរបស់មនុស្សភាគច្រើនត្រូវបានបាត់បង់។ បន្ទាប់មក កូនកាត់ត្រូវបានជ្រើសរើស ដែលក្រូម៉ូសូមរបស់មនុស្សនៅសល់តែមួយ។ ការសិក្សាអំពីកូនកាត់បែបនេះបានធ្វើឱ្យវាអាចធ្វើទៅបានដើម្បីភ្ជាប់លក្ខណៈជីវគីមីមួយចំនួននៃកោសិកាមនុស្សជាមួយនឹងក្រូម៉ូសូមរបស់មនុស្សជាក់លាក់។ បន្តិចម្ដងៗ តាមរយៈការប្រើប្រាស់ប្រព័ន្ធផ្សព្វផ្សាយជ្រើសរើស ពួកគេរៀនដើម្បីសម្រេចបាននូវការអភិរក្ស ឬការបាត់បង់ក្រូម៉ូសូមរបស់មនុស្សម្នាក់ៗដែលផ្ទុកហ្សែនជាក់លាក់។ គ្រោងការណ៍នៃការជ្រើសរើស ទោះបីជាមិនសាមញ្ញបំផុតនៅ glance ដំបូងក៏ដោយ បានបង្ហាញខ្លួនឯងយ៉ាងល្អនៅក្នុងការពិសោធន៍។ ដូច្នេះហើយ ពួកគេបានបង្កើតឧបករណ៍ជ្រើសរើសពិសេសមួយ ដែលមានតែកោសិកាទាំងនោះដែលអង់ស៊ីម thymidine kinase ត្រូវបានសំយោគអាចរស់បាន។ ប្រសិនបើសម្រាប់ការបង្កាត់ជាមួយកោសិកាមនុស្ស យើងយកកោសិកាកណ្ដុរដែលផ្លាស់ប្តូរជាដៃគូដែលមិនសំយោគ thymidine kinase នោះមានតែកូនកាត់ទាំងនោះដែលមានក្រូម៉ូសូមរបស់មនុស្សជាមួយនឹងហ្សែន thymidine kinase ប៉ុណ្ណោះដែលនឹងនៅរស់។ តាមរបៀបនេះ ជាលើកដំបូង គេអាចបង្កើតការធ្វើមូលដ្ឋានីយកម្មនៃហ្សែន thymidine kinase នៅលើក្រូម៉ូសូម 17 របស់មនុស្ស។
ទោះបីជាការពិតដែលថាការសិក្សាអំពីហ្សែនរបស់មនុស្សនៅកម្រិតក្រូម៉ូសូមផ្តល់នូវលក្ខណៈសំខាន់ៗមួយចំនួន ពួកវាមានលក្ខណៈទូទៅបំផុត និងផ្តល់តិចតួចសម្រាប់ការយល់ដឹងពេញលេញអំពីរចនាសម្ព័ន្ធ និងមុខងារនៃបរិធានហ្សែននៃកោសិកាមនុស្ស។
| |
យើងបាននិយាយថាកោសិកា somatic មានសំណុំក្រូម៉ូសូមទ្វេ diploid ហើយកោសិកាផ្លូវភេទចាស់ទុំមានសំណុំ haploid តែមួយ។ សំណុំក្រូម៉ូសូម diploid ក៏មានវត្តមាននៅក្នុងកោសិកាដំណុះដែលមិនទាន់ទុំផងដែរ។ ពាក់កណ្តាលនៃចំនួនក្រូម៉ូសូម ហើយតាមនោះ DNA ដែលហៅថាការកាត់បន្ថយរបស់វាកើតឡើងនៅក្នុងដំណើរការនៃ gametogenesis ពោលគឺការវិវត្តនៃកោសិកាមេរោគ - gametes ។ ការកាត់បន្ថយចំនួនពាក់កណ្តាលនៃក្រូម៉ូសូមនៅក្នុង gametes គឺជាការរៀបចំសម្រាប់ការបង្កកំណើតនាពេលអនាគត ដែលសំណុំ diploid របស់ពួកគេត្រូវបានស្ដារឡើងវិញដោយសារតែការបញ្ចូលគ្នានៃសំណុំក្រូម៉ូសូម haploid នៃមេជីវិតឈ្មោល និងស៊ុត។
Gametogenesis កើតឡើងនៅក្នុង gonads: នៅក្នុងពងស្វាសនៅក្នុងរាងកាយបុរសនិងនៅក្នុងអូវែរនៅក្នុងស្ត្រី។ ដូច្នោះហើយវាត្រូវបានគេហៅថា spermatogenesis និង oogenesis ។ ធម្មតាចំពោះមេជីវិតឈ្មោល- និង oogenesis គឺជារយៈពេល 3 ដំបូងនៃ gametogenesis: ការបន្តពូជ ការលូតលាស់ និងការពេញវ័យ។ ការអភិវឌ្ឍកោសិកាមេជីវិតឈ្មោលឆ្លងកាត់ដំណាក់កាលទី 4 បន្ថែមទៀត - ការបង្កើត (រូបភាព 26) ។
នៅក្នុងអំឡុងពេលដំបូងនៃ gametogenesis កោសិកាមេរោគបានបែងចែកយ៉ាងខ្លាំងក្លា ហើយចំនួនរបស់វាកើនឡើង។ នៅក្នុងអំឡុងពេលនេះកោសិកាមេរោគត្រូវបានគេហៅថា spermato- និង oogonia រៀងគ្នា។ សម្រាប់ភាពសាមញ្ញ ដ្យាក្រាមបង្ហាញករណីនៅពេលដែលសំណុំ haploid រួមបញ្ចូលតែក្រូម៉ូសូមចំនួន 3 ដែលមួយក្នុងចំនោមនោះគឺជាភេទ - gonosome ឬ heterochromosome (ជាភាសាក្រិច "heteros" មានន័យថាខុសគ្នា) និងពីរមិនមែនផ្លូវភេទ - autosomes ។ នៅក្នុងកោសិកា សំណុំ haploid មួយត្រូវបានធ្វើឱ្យខ្មៅ ទីពីរត្រូវបានចង្អុលបង្ហាញដោយបន្ទាត់វណ្ឌវង្ក។ ស្វ័យភាពដូចគ្នា និងគ្មានភាពច្បាស់លាស់នៃឈុតទាំងពីរត្រូវបានគូរក្នុងរូបរាង និងទំហំដូចគ្នា (នេះគឺជាបន្ទាត់ និងរង្វង់វែងបំផុត)។ Heterochromosomes ត្រូវបានបង្ហាញខុសគ្នា - ជាមួយនឹងបន្ទាត់ត្រង់ (ក្រូម៉ូសូម X) និងបន្ទាត់កោងនៃប្រវែងដូចគ្នា (ក្រូម៉ូសូម Y) ។ ក្រូម៉ូសូមស្ថិតនៅដាច់ដោយឡែកពីគ្នានៅក្នុងកោសិកា។
កោសិកាផ្លូវភេទដែលបានចូលទៅក្នុងដំណាក់កាលលូតលាស់ត្រូវបានកំណត់ថាជាមេជីវិតឈ្មោល- និង oocytes នៃលំដាប់ទីមួយ។ ពួកវាកើនឡើងក្នុងទំហំ ជាពិសេស oocytes ហើយបរិធាននុយក្លេអ៊ែររបស់ពួកគេកំពុងដំណើរការរៀបចំរចនាសម្ព័ន្ធឡើងវិញ។ ក្រូម៉ូសូម homologous ស្ថិតនៅស្របគ្នាទៅវិញទៅមក បង្កើតជា bivalents ចំនួនដែលស្មើនឹងចំនួនក្រូម៉ូសូមនៅក្នុងសំណុំ haploid ។ ក្រូម៉ូសូមទាំងពីរនៃ bivalent នីមួយៗគឺជារចនាសម្ព័ន្ធគូ - dyad ព្រោះវាមានក្រូម៉ាទីតប្អូនស្រីពីរ។ នៅពេលដែលគម្លាតរវាង chromatids ទាំងនេះត្រូវបានកំណត់យ៉ាងល្អនោះ bivalents មើលទៅដូចជា tetrads រួចទៅហើយ។ ចំនួននៃ tetrads ត្រូវគ្នាទៅនឹងចំនួន haploid នៃក្រូម៉ូសូម។ ចំនួនសរុបនៃក្រូម៉ាទីតដែលអាចរកឃើញ - ក្រូម៉ូសូមនាពេលអនាគតនៃកោសិកាមេជីវិតចាស់ - គឺ tetraploid ។ បរិមាណ DNA នៅក្នុង spermato- និង oocytes មុនពេលកាលកំណត់ក៏ជា tetraploid ផងដែរ។
បន្ទាប់មកមករយៈពេលនៃភាពចាស់ទុំដែលត្រូវបានកំណត់ដោយ meiosis ("meiosis" មានន័យថាការកាត់បន្ថយ) - ការបែងចែកកោសិកាបន្តបន្ទាប់គ្នាយ៉ាងឆាប់រហ័សចំនួនពីរដែលក្នុងអំឡុងពេលនោះការកាត់បន្ថយក្រូម៉ូសូមកើតឡើង។ កោសិកាមេជីវិតឈ្មោលដែលបានបញ្ចប់ការបែងចែកដំបូងនៃភាពចាស់ទុំត្រូវបានគេហៅថា spermatocytes នៃលំដាប់ទីពីរ ឬ prespermatids ហើយកោសិកាមេជីវិតឈ្មោលដែលត្រូវគ្នាត្រូវបានគេហៅថា oocytes នៃលំដាប់ទីពីរ បន្ទាប់ពីការបែងចែកទីពីរនៃភាពចាស់ទុំ prespermatids ក្លាយជា operamatids ដែលជា oocytes ។ លំដាប់ទីពីរក្លាយជាស៊ុតចាស់ទុំ ការបែងចែកដំបូងនៃភាពចាស់ទុំគឺកាត់បន្ថយក្នុងអំឡុងពេលនៃការបែងចែកនេះ ក្រូម៉ូសូមទាំងមូល - dyads - ត្រូវបានចែកចាយរវាងកោសិកាកូនស្រី ហើយសំណុំក្រូម៉ូសូមក្លាយជា haploid ការបែងចែកទីពីរនៃភាពចាស់ទុំត្រូវបានគេហៅថាស្មើភាពស្មើគ្នា នៃក្រូម៉ូសូម (dyads) សំខាន់ក្រូម៉ាទីតរបស់ពួកគេ ខុសគ្នារវាងកោសិកាកូនស្រី។
interphase រវាងការបែងចែកទីមួយ និងទីពីរនៃភាពចាស់ទុំអាចខ្លីណាស់ ឬសូម្បីតែអវត្តមានទាំងអស់គ្នា ដោយសារនៅពេលនេះ ការចម្លង DNA ឬការកើនឡើងទ្វេដងនៃចំនួនក្រូម៉ូសូមកើតឡើងនៅក្នុងកោសិកា។ ការបែងចែកចាប់ផ្តើមម្តងទៀត ឥឡូវនេះមានកោសិកាពីរ ហើយកោសិកាចៅនីមួយៗទទួលបានក្រូម៉ូសូមមួយពីឌីយ៉ាត។ ដូច្នេះ កោសិកានីមួយៗនៃកោសិកាទាំងបួននេះ ដែលកើតចេញពីការបែងចែក 2 នៃភាពចាស់ទុំ ទទួលបានធាតុមួយនៃ tetrad ។ ចំនួន haploid នៃ tetrads ត្រូវគ្នាទៅនឹងចំនួន haploid នៅក្នុងសំណុំក្រូម៉ូសូមនៃកោសិកាដែលបានឆ្លងកាត់ meiosis ។ ដូចគ្នានេះដែរបរិមាណ DNA ដែលជា tetraploid នៅដើមដំបូងនៃ meiosis (នៅក្នុង oocyte និង spermatocyte នៃលំដាប់ទីមួយ) បន្ទាប់ពីការបែងចែកជា 4 ផ្នែកក្លាយជា haploid នៅចុងបញ្ចប់របស់វា។
ក្នុងអំឡុងពេលនៃការបង្កើត spermatogenesis ពី spermatogonia នីមួយៗដែលបានចូលដល់ដំណាក់កាលលូតលាស់ មេជីវិតឈ្មោលពេញលក្ខណៈចំនួន 4 ត្រូវបានទទួលជាលទ្ធផលនៃការបែងចែកនៃភាពចាស់ទុំ។ ក្នុងអំឡុងពេល oogenesis, oocyte លំដាប់ទីពីរដែលកើតចេញពី oocyte លំដាប់ទីមួយរក្សាទំហំស្ទើរតែទាំងមូលនៃកោសិកាម្តាយ។ កោសិកាកូនស្រីទីពីរទទួលបានពាក់កណ្តាលនៃសារធាតុក្រូម៉ូសូមរបស់ម្តាយ ហើយមានតែផ្នែកមួយមិនសំខាន់នៃ cytoplasm របស់វា។ កោសិកាតូចនេះត្រូវបានគេហៅថារាងកាយកាត់បន្ថយ។ រូបភាពស្រដៀងគ្នានេះត្រូវបានធ្វើម្តងទៀតក្នុងអំឡុងពេលនៃការបែងចែកទីពីរនៃភាពចាស់ទុំ - oocyte លំដាប់ទីពីរផ្តល់ការកើនឡើងដល់ស៊ុតដែលមានទំហំប្រហាក់ប្រហែលនិងរាងកាយកាត់បន្ថយទីពីរ។ ក្នុងពេលជាមួយគ្នានេះរាងកាយកាត់បន្ថយដំបូងក៏បែងចែកជាពីរ។ ជាលទ្ធផលពី oogonia មួយដែលបានឆ្លងផុតពីរយៈពេលនៃការបន្តពូជទៅអំឡុងពេលនៃការលូតលាស់ហើយបន្ទាប់មកចូលទៅក្នុងរយៈពេលនៃភាពចាស់ទុំមានតែពងចាស់មួយប៉ុណ្ណោះដែលត្រូវបានបង្កើតឡើង។ នេះគឺជាការសម្របខ្លួនដ៏ល្អសម្រាប់ការបង្កើត - ការផ្គត់ផ្គង់ទាំងមូលនៃសារធាតុចិញ្ចឹមដែលប្រមូលផ្តុំដោយ oocyte លំដាប់ទីមួយក្នុងអំឡុងពេលលូតលាស់ ដែលចាំបាច់ដើម្បីធានាបាននូវដំណាក់កាលដំបូងនៃការអភិវឌ្ឍន៍នៃអំប្រ៊ីយ៉ុងនាពេលអនាគតនៅតែមាននៅក្នុងស៊ុត។
ក្នុងអំឡុងពេលនៃការបង្កើត មេជីវិតឈ្មោលនៃកោសិកាធម្មតាមួយត្រូវបានផ្លាស់ប្តូរទៅជាមេជីវិតឈ្មោលដែលមានរចនាសម្ព័ន្ធស្មុគ្រស្មាញខ្លាំង ដែលធានានូវតួនាទីរបស់វាជាដៃគូចល័តសកម្មក្នុងសកម្មភាពបង្កកំណើត។ សមាសធាតុទាំងអស់នៃ spermatid ចូលរួមក្នុងការរៀបចំឡើងវិញនេះ។
ដំបូងបង្អស់ centrioles របស់វាមានទីតាំងនៅមួយទៅមួយ ដូច្នេះកំណត់អ័ក្សវែងនៃមេជីវិតឈ្មោលនាពេលអនាគត (រូបភាព 27A) ។ ក្នុងចំណោមគ្រាប់ proacrosomal តូចៗជាច្រើនដែលបង្កើតឡើងនៅក្នុង spermatocyte - នៅចំកណ្តាលនៃបរិធាន Golgi របស់វា ដែលឥឡូវនេះផ្លាស់ទីទៅផ្នែកខាងចុងនៃកោសិកា (រូបភាព 27B-3) គ្រាប់ acrosomal ដ៏ធំមួយ ("acron" មានន័យថាចុង) លេចឡើង។ ដែលបន្ទាប់មកស្ថិតនៅលើស្នូលនៅកន្លែងនៃបង្គោលក្បាលនាពេលអនាគតរបស់វា (រូបភាព 27B-9) ។ ស្មុគ្រស្មាញ lamellar ត្រូវបានកាត់បន្ថយ ដែលបណ្តាលឱ្យមាន vesicle acroblast រុំព័ទ្ធ granule acrosomal ។ រាងកាយរបស់មេជីវិតឈ្មោលចាប់ផ្តើមពង្រីកបន្តិចម្តងៗ ហើយស្នូលកោសិកាកាន់តែក្រាស់។ វាមានទីតាំងនៅខាងចុងនៃមេជីវិតឈ្មោលដែលកំពុងលូតលាស់។ ចំនុចកណ្តាលជិតស្ថិតនៅខាងក្រោយស្នូល ហើយផ្នែកខាងចុងបង្កើតបានជា flagellum ដូចជា kintosome ។ បន្ទាប់មកវាត្រូវបានបែងចែកទៅជាពីរផ្នែកហើយផ្នែកខាងក្រោយនៃពួកវាមានរាងជារង្វង់ (រូបភាព 27B-8) ហើយផ្លាស់ទីឆ្ងាយពីផ្នែកខាងមុខដោយរអិលតាមបណ្តោយទង់ជាតិដែលកំពុងលូតលាស់ - សរសៃអ័ក្សនាពេលអនាគតនៃកន្ទុយមេជីវិតឈ្មោល។ ចិញ្ចៀនស្ថិតនៅគែមខាងក្រោយនៃក្រឡា។ នៅពេលនេះ mitochondria ភាគច្រើនត្រូវបានប្រមូលផ្តុំនៅជិត filament axial ។ Acroblast, រីកលូតលាស់, ផ្លាស់ទីក្នុងទម្រង់ជាមួកមួយនៅលើផ្នែកខាងមុខនៃស្នូល។
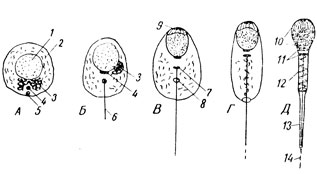
នៅចុងបញ្ចប់នៃកំឡុងពេលនៃការបង្កើត ភាពខុសគ្នាទៅជាផ្នែកត្រូវបានសម្តែងយ៉ាងល្អនៅក្នុងមេជីវិតឈ្មោល: ក្បាលដែលជាស្នូលដែលមានរាងសំប៉ែត និងបង្រួមខ្លាំង គ្របដណ្តប់នៅខាងមុខដោយស្រោបមួយ ក្រោមដែល acrosome ស្ថិតនៅគែមបំផុត; កដែលបង្កើតឡើងដោយ centrioles; កម្រិតមធ្យម ផ្នែកតភ្ជាប់ និងកន្ទុយ។ មីតូខនឌ្រីទាំងអស់ត្រូវបានប្រមូលផ្តុំនៅក្នុងផ្នែកមធ្យម ដែលវិលជុំវិញសរសៃអំបោះ។ ព្រំដែន distal នៃផ្នែកមធ្យមគឺ centriole ring បិទ។ នៅក្នុង cytoplasm មានសារធាតុជាច្រើន (glycogen, lipids) ដោយសារតែការបែកខ្ញែកដែលមេជីវិតឈ្មោលទទួលបានថាមពលសម្រាប់ចលនា។ នៅកន្ទុយនៅពេលពិនិត្យនៅក្រោមមីក្រូទស្សន៍អុបទិកផ្នែកពីរត្រូវបានសម្គាល់ - ផ្នែកសំខាន់គ្របដណ្តប់ដោយស៊ីតូប្លាសនិងផ្នែកស្ថានីយ "ទទេ" ដែលមានតែសរសៃកន្ទុយប៉ុណ្ណោះ។
ដោយប្រើមីក្រូទស្សន៍អេឡិចត្រុង វាត្រូវបានបង្កើតឡើងថាមួកស្តើងដែលគ្របលើស្នូលមេជីវិតឈ្មោល គឺជាធុងដែលមានរាងសំប៉ែត។ ភ្នាសខាងក្នុងរបស់វានៅជាប់នឹងស្នូល ហើយភ្នាសខាងក្រៅរបស់វានៅជាប់នឹងប្លាស្មា។ នៅផ្នែកខាងក្រោយនៃក្បាលមេជីវិតឈ្មោល ប្លាស្មាម៉ាឡាមម៉ាដោយផ្ទាល់គ្របដណ្តប់ស្នូល។ ស្នូលខ្លួនវាត្រូវបានបំពេញយ៉ាងក្រាស់ជាមួយនឹងខ្សែស្រឡាយជាប់គ្នាដែលមានកម្រាស់ 40 Å ដែលជាម៉ូលេគុល DNP (nucleohistone) ។ ការវិភាគគីមីនៃ chromatin នុយក្លេអ៊ែរ condensed បង្ហាញថាវាមានប្រហែលពាក់កណ្តាល DNA និងពាក់កណ្តាលប្រូតេអ៊ីន។ centrioles ដែលមានទីតាំងនៅតំបន់កមានរចនាសម្ព័ន្ធធម្មតានៃសរីរាង្គទាំងនេះ។ ពួកវាតូចជាងដែលនៅជាប់នឹងផ្នែកខាងក្រោយនៃស្នូលនៅផ្នែកកណ្តាលរបស់វាមានរូបរាងនៃស៊ីឡាំងដែលបង្កើតឡើងដោយបំពង់ 9 គូ។ មជ្ឈមណ្ឌល distal ត្រូវបានអភិវឌ្ឍជាង។ វាដូចជា kinetochore គឺទាក់ទងនឹងគែមសរសៃចំនួន 9 គូនៃ flagellum ដែលមានកណ្តាល 2 បន្ថែមទៀតដូចជា cilia និង flagella ទូទៅ។
មីតូខនឌ្រី ដែលស្ថិតនៅយ៉ាងក្រាស់នៅក្នុងផ្នែកមធ្យមជុំវិញសរសៃអ័ក្ស បង្កើតបានប្រហែល 14 វេននៃវង់។ ជាក់ស្តែង ជាដំបូងពួកគេផ្តល់ថាមពលដល់ធាតុ contractile នៃមេជីវិតឈ្មោល។ នៅក្នុងផ្នែកសំខាន់នៃកន្ទុយ សរសៃអ័ក្សចំនួន 9 គូត្រូវបានហ៊ុំព័ទ្ធដោយសរសៃក្រវ៉ាត់ដែលនៅជាប់គ្នាដោយខ្សែក្រាស់វែងពីរ។ នៅក្នុងផ្នែកស្ថានីយនៃកន្ទុយ មិនមានដុំសាច់ក្រវ៉ាត់កទេ បណ្តុំនៃសរសៃអ័ក្សដែលបង្កើតជាសរសៃអ័ក្សត្រូវបានរុំព័ទ្ធក្នុងម៉ាស់ដូចគ្នា និងគ្របដណ្តប់លើផ្ទៃដោយប្លាស្មា។ ដូច្នេះនៅទីនេះយើងជួបប្រទះលក្ខណៈរចនាសម្ព័ន្ធនៃ cilia និង flagella ទាំងអស់។
ប្រវែងសរុបនៃមេជីវិតឈ្មោលរបស់មនុស្សឈានដល់ប្រហែល 60 mk. វាផ្លាស់ទីយ៉ាងសកម្មក្នុងល្បឿន 3.5 មីលីម៉ែត្រក្នុងមួយនាទី។ ក្នុងពេលជាមួយគ្នានេះ វាបង្វិលជុំវិញអ័ក្សរបស់វាតាមទ្រនិចនាឡិកា ធ្វើឱ្យមានបដិវត្តន៍ពេញលេញមួយក្នុងរយៈពេល 15 នាទី។ សមត្ថភាពផ្លាស់ទីរបស់វាអាស្រ័យលើ pH និងលក្ខណៈសម្បត្តិផ្សេងទៀតនៃបរិស្ថាន។ អាយុកាលរបស់មេជីវិតឈ្មោលនៅក្នុងទ្វាមាសគឺត្រឹមតែ 1 ម៉ោងប៉ុណ្ណោះ។ នៅតាមបណ្តោយប្រដាប់បន្តពូជស្ត្រី មេជីវិតឈ្មោលធ្វើចលនាដោយអសកម្ម (ជាលទ្ធផលនៃការកន្ត្រាក់សាច់ដុំនៃជញ្ជាំងស្បូន និងបំពង់អូវែ) ឈានដល់ផ្នែកខាងលើទីបីនៃបំពង់អូវុល ដែលជាកន្លែងបង្កកំណើតកើតឡើង។ ដូច្នេះហើយ គាត់មិនចាំបាច់ខ្ជះខ្ជាយថាមពលពីទុនបម្រុងដែលមិនសូវសំខាន់របស់គាត់ ដើម្បីយកឈ្នះលើផ្លូវដ៏វែងឆ្ងាយបែបនេះសម្រាប់គាត់នោះទេ។
កោសិកាស៊ុតចាស់ទុំមានរាងស្វ៊ែរ។ អង្កត់ផ្ចិតរបស់វានៅក្នុងមនុស្សគឺ 135 mk. វាតែងតែត្រូវបានគ្របដណ្ដប់ដោយភ្នាសដែលអាចមើលឃើញដោយមីក្រូទស្សន៍ ហើយរចនាសម្ព័ន្ធរបស់វាខុសពីកោសិកា somatic ជាចម្បងតាមពីរវិធី។ ទីមួយនៅក្នុងវា សមាមាត្រនុយក្លេអ៊ែរ - ស៊ីតូប្លាមិកត្រូវបានផ្លាស់ប្តូរយ៉ាងខ្លាំងនៅក្នុងការពេញចិត្តនៃ cytoplasm ដែលត្រូវបានពន្យល់ដោយការប្រមូលផ្តុំនៅក្នុងខ្លួនរបស់ oocyte ក្នុងអំឡុងពេលនៃការលូតលាស់នៃសារធាតុចិញ្ចឹមសម្រាប់តម្រូវការនៃអំប្រ៊ីយ៉ុងនាពេលអនាគត។ ទីពីរ វាខ្វះមជ្ឈមណ្ឌលកោសិកា ដែលបាត់ក្នុងអំឡុងពេលដូចគ្នានៃការលូតលាស់។ មជ្ឈមណ្ឌលកោសិកាណែនាំមេជីវិតឈ្មោលចូលទៅក្នុងស៊ុតអំឡុងពេលបង្កកំណើត ហើយបន្ទាប់ពីនេះការបែងចែក mitotic នៃ zygote ចាប់ផ្តើម - សារពាង្គកាយដែលមានទីតាំងនៅដំណាក់កាល unicellular នៃ ontogenesis ។
ចូរយើងនិយាយដោយសង្ខេបអំពីបញ្ហានៃការកំណត់ភេទ ដែលកើតឡើងបន្ទាប់ពីការបង្កកំណើតនៃសារពាង្គកាយ។ ដូចដែលវាបានប្រែក្លាយ វាកើតឡើងនៅពេលនៃការមានគភ៌ ពោលគឺការបង្កកំណើត និងបណ្តាលមកពីការរួមផ្សំនៃ heterochromosomes នៅក្នុង zygote ។ ចូរយើងងាកទៅរកដ្យាក្រាម (រូបភាពទី 26) ហើយពិចារណាអំពីសំណុំក្រូម៉ូសូមក្នុងការអភិវឌ្ឍកោសិកាមេជីវិតឈ្មោល និងស្ត្រី។ oogonia ទាំងអស់មានក្រូម៉ូសូម X ពីរ។ វាធ្វើតាមថាសំណុំក្រូម៉ូសូម haploid នៃស៊ុតពេញវ័យនីមួយៗនឹងចាំបាច់រួមបញ្ចូលក្រូម៉ូសូម X មួយ។ នៅក្នុង spermatogonia ក្នុងចំណោម heterochromosomes ពីរមានក្រូម៉ូសូម X និងក្រូម៉ូសូម Y មួយ។ ដូច្នេះមេជីវិតឈ្មោលត្រូវតែខុសគ្នា - ពាក់កណ្តាលជាមួយក្រូម៉ូសូម X និងពាក់កណ្តាលជាមួយក្រូម៉ូសូម Y ។
នៅក្នុងថនិកសត្វ និងមនុស្ស នៅពេលដែលក្រូម៉ូសូម X ពីរត្រូវបានបញ្ចូលគ្នានៅក្នុង zygote មួយ សារពាង្គកាយស្ត្រីមានការវិវឌ្ឍន៍នៅក្នុងវត្តមាននៃក្រូម៉ូសូម XY សារពាង្គកាយបុរសមានការរីកចម្រើន។ ដូច្នេះ ភេទរបស់កូនដែលមិនទាន់កើតនឹងអាស្រ័យទៅលើមេជីវិតឈ្មោលរបស់ឪពុកដែលស៊ុតរបស់ម្តាយភ្ជាប់ជាមួយ (រូបភាពទី 28)។ សំណុំ haploid របស់មនុស្សមានមួយឬមួយផ្សេងទៀតនៃ heterochromosomes (ក្រូម៉ូសូមផ្លូវភេទ - gonosomes) និង 22 autosomes (ក្រូម៉ូសូមមិនមែនភេទ) ។ វាគួរតែមានក្រូម៉ូសូមចំនួន 46 នៅក្នុងកោសិកាហ្សីហ្គោត អំប្រ៊ីយ៉ុង សូម៉ាត និងកោសិកាមេជីវិតមិនទាន់ពេញវ័យរបស់មនុស្ស - 44+XY នៅក្នុងរាងកាយបុរស និង 44+XX ក្នុងស្ត្រី។ មានតែបន្ទាប់ពីការបែងចែក meiotic លើកដំបូងនៅក្នុងកោសិកាដំណុះដែលចាស់ទុំចំនួនក្រូម៉ូសូមត្រូវបានកាត់បន្ថយមកត្រឹម 23។ ដើម្បីឱ្យការអភិវឌ្ឍន៍ធម្មតាកើតឡើង ចាំបាច់ត្រូវមានសំណុំក្រូម៉ូសូម diploid ដោយគ្មានគម្លាតណាមួយទាំងបរិមាណ និងរចនាសម្ព័ន្ធ។
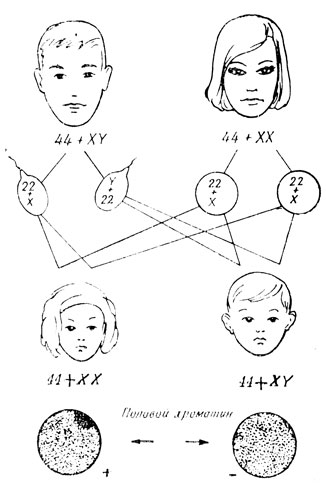
នៅឆ្នាំ 1949 លោក Barr បានបង្កើតថានៅក្នុងថនិកសត្វញី និងស្ត្រី ក្រូម៉ូសូម X នៅក្នុងសំណុំ diploid មានឥរិយាបទខុសគ្នា - មួយក្នុងចំនោមពួកគេបន្ទាប់ពីការបែងចែកកោសិកា despirals ដូចជា autosomes ហើយក្លាយទៅជាមិនអាចបែងចែកបាននៅក្នុងស្នូល interphase មួយទៀតនៅតែវិលជុំវិញខ្ពស់ដូចជា ក្រូម៉ូសូម heterochromatic ។ ក្រូម៉ូសូម X heterochromatic នេះដែលអាចមើលឃើញយ៉ាងច្បាស់នៅក្នុងស្នូលក្នុងទម្រង់ជារាងកាយងងឹតនៅក្នុងស្នាមប្រឡាក់ធម្មតានៃការត្រៀមលក្ខណៈត្រូវបានគេហៅថា chromatin ផ្លូវភេទ។ នៅក្នុងការពិសោធន៍ហ្សែនដើម្បីកំណត់អត្តសញ្ញាណ Barr សាកសព (សាកសព chromatin ផ្លូវភេទ) វាងាយស្រួលបំផុតដើម្បីពិនិត្យមើល lymphocytes ឈាមឬកោសិកា epithelial desquamated ។ វាត្រូវបានគេរកឃើញថាក្រូម៉ូសូម X ទាំងពីរអាចក្លាយទៅជាអសកម្ម។
ក្នុងករណីដ៏កម្រក្នុងអំឡុងពេល oogenesis ក្រូម៉ូសូម X មិនបំបែកចេញពីគ្នាក្នុងអំឡុងពេល meiosis ។ ជាលទ្ធផល ស៊ុតដែលខុសពីបទដ្ឋានអាចត្រូវបានបង្កើតឡើង៖ ជំនួសឱ្យក្រូម៉ូសូម X មួយអាចមានពីរ ឬប្រហែលជាមិនមានក្រូម៉ូសូម X ទាល់តែសោះ។ ក្នុងករណីចុងក្រោយ ក្នុងអំឡុងពេលបង្កកំណើត ហ្សីហ្គោតពីរប្រភេទអាចកើតឡើង ដែលមាននៅក្នុងសំណុំ diploid ទាំងក្រូម៉ូសូម Y មួយ ឬក្រូម៉ូសូម X មួយដែលត្រូវបានបញ្ចូលទៅក្នុងស៊ុតអំឡុងពេលបង្កកំណើតដោយមេជីវិតឈ្មោល។ ហ្សីកូតដែលមានក្រូម៉ូសូម Y មួយ ជាទូទៅមិនអាចដំណើរការបាន ហើយស្លាប់។ ហ្សីហ្គោតដែលមានក្រូម៉ូសូម X មួយនៅក្នុងមនុស្សមាន 45 ក្រូម៉ូសូម: 44+XO ។ ជាមួយនឹងការរួមបញ្ចូលគ្នានៃក្រូម៉ូសូមនេះ ហ្សីហ្គោតនឹងវិវឌ្ឍន៍ទៅជាស្ត្រីទាបដែលមានកម្ពស់តូច ជាមួយនឹងអូវែរមូល ហើយជាលទ្ធផល អវត្ដមាននៃលក្ខណៈផ្លូវភេទបន្ទាប់បន្សំ។ រោគសាស្ត្រនេះត្រូវបានគេស្គាល់ថាជារោគសញ្ញា Turner ។ ក្រូម៉ូសូម X តែមួយគត់នៅក្នុងករណីនេះត្រូវបាន despiralized ហើយដូច្នេះ chromatin ផ្លូវភេទមិនត្រូវបានរកឃើញនៅក្នុងកោសិកា somatic នៅក្នុងក្មេងស្រីបែបនេះ។
លទ្ធផលនៃការមិនបំបែកក្រូម៉ូសូម X ក្នុងអំឡុងពេល meiosis ក៏នឹងមានគម្លាតនៅក្នុងសំណុំនៃធម្មជាតិផ្ទុយគ្នាផងដែរ ពោលគឺវត្តមាននៃក្រូម៉ូសូម X បី ឬក្រូម៉ូសូម X ពីរ និងក្រូម៉ូសូម Y មួយនៅក្នុងហ្សីហ្គោត។ សារពាង្គកាយស្ត្រីដែលមានក្រូម៉ូសូម X ចំនួនបីត្រូវបានគេហៅថា "ស្ត្រីកំពូល" ឬ "ស្ត្រីកំពូល" (សម្រាប់មនុស្ស) ។ ទោះជាយ៉ាងណាក៏ដោយ បុគ្គលបែបនេះត្រូវបានគេហៅថា "ស្ត្រីកំពូល" តែប៉ុណ្ណោះតាមលក្ខខណ្ឌ ដោយផ្អែកលើចំនួនក្រូម៉ូសូម X លើសលប់។ តាមពិតជាមួយនឹងសំណុំនៃក្រូម៉ូសូម 44 + XXX មានការវិវឌ្ឍន៍នៃអូវែរ ហើយជារឿយៗការបាត់បង់ការមានកូន។ គួរឱ្យចាប់អារម្មណ៍ "ស្ត្រីកំពូល" ដែលមានក្រូម៉ូសូម X ចំនួនបួន (សំណុំក្រូម៉ូសូម 44 + XXXX) មានកូន ប៉ុន្តែបានកាត់បន្ថយការអភិវឌ្ឍន៍ផ្លូវចិត្ត។ នៅពេលវិភាគក្រូម៉ូសូមភេទនៃកោសិកា somatic ចំពោះស្ត្រីដែលមានក្រូម៉ូសូម XXX ឬ XXXX សាកសព Barr 2 ឬ 3 ត្រូវបានរកឃើញនៅក្នុងស្នូលរៀងគ្នា។ ដូច្នេះ នៅក្នុងករណីទាំងនេះ មានតែក្រូម៉ូសូម X មួយប៉ុណ្ណោះដែលត្រូវបានបំផ្លិចបំផ្លាញ និងសកម្មនៅក្នុងស្នូលអន្តរដំណាក់កាល។
Zygotes ដែលមានសំណុំក្រូម៉ូសូមនៃប្រភេទ 44+XXY វិវឌ្ឍន៍លើបុរសដែលទទួលរងពីរោគសញ្ញា Klinefelter - វិកលចរិត និងការអភិវឌ្ឍន៍នៃពងស្វាសដែលនាំទៅដល់ភាពគ្មានកូន។ កោសិកា somatic របស់ពួកគេមាន chromatin ផ្លូវភេទ និងមានរាងកាយ 1 Barr ។ X ទីពីរដូចបុរសធម្មតាដែរគឺត្រូវបានរំសាយក្នុងដំណាក់កាលអន្តរកាល។ ពិការភាពនៃការអភិវឌ្ឍន៍ស្រដៀងគ្នានេះត្រូវបានគេសង្កេតឃើញជាមួយនឹងចំនួនក្រូម៉ូសូម X ធំជាងដែលរួមបញ្ចូលគ្នាជាមួយក្រូម៉ូសូម Y ដែលមានប្រភេទ XXXY, XXXXY និង XXXXXY ។ វត្តមានរបស់ក្រូម៉ូសូម Y នៅក្នុងសំណុំកំណត់ការវិវឌ្ឍន៍របស់បុគ្គលបុរស ប៉ុន្តែមានកម្រិតទាបជាង។ ចំនួនសាកសព Barr នៅក្នុងស្នូលអន្តរដំណាក់កាលនៃកោសិកា somatic គឺតិចជាងចំនួនក្រូម៉ូសូម X នៅក្នុងសំណុំ។
ថ្មីៗនេះ ភាពមិនធម្មតានៃក្រូម៉ូសូមមួយទៀតត្រូវបានរកឃើញ - ប្រភេទ XYY ។ សត្វ "បុរសកំពូល" ដែលមានក្រូម៉ូសូម Y បន្ថែមត្រូវបានសម្គាល់ដោយភាពខ្លាំងនិងភាពឆេវឆាវ។ បុរសដែលមានក្រូម៉ូសូម Y ពីរមានកម្ពស់ (លើសពី 180 សង់ទីម៉ែត្រ) មានកម្លាំងរាងកាយដ៏អស្ចារ្យ ប៉ុន្តែមានការថយចុះសមត្ថភាពផ្លូវចិត្ត។ ដូចទៅនឹងបុរសធម្មតាដែរ ស្នូល interphase នៃកោសិកា somatic របស់ពួកគេមិនមាន chromatin ភេទទេ។
អ្នកវិទ្យាសាស្ត្របានរកឃើញថាមរតកនៃជំងឺមួយចំនួនត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងហ្សែនដែលមានទីតាំងនៅលើក្រូម៉ូសូមភេទ។ ឧទាហរណ៍ ជំងឺភ្នែកពណ៌ពីកំណើត (ជំងឺដែលពីមុនគេហៅថា ពិការភ្នែកពណ៌) ជាទូទៅ ភាពងងឹតភ្នែកដែលបណ្តាលមកពីការដាច់នៃសរសៃប្រសាទអុបទិក ជំងឺ hemophilia (ការលំបាកខាងរោគសាស្ត្រក្នុងការបញ្ឈប់ការហូរឈាម) ត្រូវបានបញ្ជូនតាមរយៈក្រូម៉ូសូម X ។
បាតុភូតនៅពេលដែលមានក្រូម៉ូសូមបន្ថែមនៅក្នុងសំណុំ ឬក្រូម៉ូសូមមួយចំនួនត្រូវបានបាត់ត្រូវបានគេហៅថា aneuploidy ។ វត្តមាននៃក្រូម៉ូសូមបន្ថែមមួយត្រូវបានគេហៅថា trisomy; ប្រសិនបើក្រូម៉ូសូមមួយបាត់ វាត្រូវបានគេហៅថា monosomy ។
ឧទាហរណ៏នៃជំងឺដែលត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការកើនឡើងនៃចំនួន autosomes គឺជា trisomy ដ៏ល្បីល្បាញបំផុត - មួយនៃក្រូម៉ូសូមតូចបំផុត - ទី 21 ។ វត្តមាននៃក្រូម៉ូសូមចំនួន 21 ជំនួសឱ្យពីរនៅក្នុងសំណុំក្រូម៉ូសូមកើតឡើងនៅក្នុងជម្ងឺ Down - ទម្រង់មួយនៃទម្រង់នៃការវិកលចរិតរួមជាមួយនឹងការពន្យាពេលនិងការអភិវឌ្ឍរាងកាយចុះខ្សោយហើយជួនកាលជាមួយនឹងវត្តមាននៃការខូចទ្រង់ទ្រាយមួយចំនួន (រូបរាងរបស់អ្នកជំងឺគឺស្រដៀងគ្នាខ្លាំងណាស់។ ត្រូវបានកំណត់លក្ខណៈដោយលលាដ៍ក្បាលតូចមួយ ខ្នងរាបស្មើនៃក្បាល ភ្នែកស្លេក ស្ពានលិចនៃច្រមុះធំទូលាយ មាត់បើកចំហពាក់កណ្តាល ត្រចៀកខូច)។ ប្រដាប់ភេទមានការអភិវឌ្ឍន៍តិចតួច ហើយលក្ខណៈផ្លូវភេទបន្ទាប់បន្សំត្រូវបានបង្ហាញមិនល្អ។ ពាក់កណ្តាលនៃកុមារដែលមានជម្ងឺ Down មិនរស់នៅលើសពី 2 ឆ្នាំទេ។
ជំងឺជាច្រើនត្រូវបានគេរកឃើញដែលបណ្តាលមកពីហ្សែនដែលមានទីតាំងនៅលើអូតូសូមផ្សេងៗគ្នា។ ក្នុងចំនោមពួកគេអាចត្រូវបានគេដាក់ឈ្មោះថាជំងឺផ្លូវចិត្តដែលទទួលមរតកដូចជាជំងឺវិកលចរិកនិងជំងឺឆ្កួតជ្រូក។
គ្រប់ប្រភេទនៃជំងឺតំណពូជរបស់មនុស្សដែលបណ្តាលមកពីជំងឺហ្សែនផ្សេងៗគ្នាគឺជាប្រធានបទនៃការសិក្សានៃសាខា cytogenetics ដែលត្រូវបានអភិវឌ្ឍយ៉ាងខ្លាំងនាពេលបច្ចុប្បន្ន - ពន្ធុវិទ្យាវេជ្ជសាស្រ្ត។
ក្រូម៉ាទីត- (ពីក្រូម៉ាក្រិក - ពណ៌ថ្នាំលាបនិងអ៊ីដូស - រូបរាង) - ធាតុរចនាសម្ព័ន្ធនៃក្រូម៉ូសូមដែលបានបង្កើតឡើងនៅចន្លោះស្នូលកោសិកាដែលជាលទ្ធផលនៃក្រូម៉ូសូមទ្វេដង (ការចម្លង) ។
ក្រូម៉ាទីត- អំបោះ nucleoprotein ពាក់កណ្តាលនៃក្រូម៉ូសូមទ្វេ។ ក្រូម៉ូសូមអាចមានតែមួយ (ពីក្រូម៉ាទីតមួយ) ឬទ្វេ (ពីក្រូម៉ាទីតពីរ) ។
មុខងាររបស់ក្រូម៉ូសូមគឺរក្សាទុកព័ត៌មានតំណពូជ។ ក្រូម៉ូសូមស្ថិតនៅក្នុងស្នូលនៃកោសិកា និងជាសមាសធាតុសំខាន់នៃស្នូល។
ក្រូម៉ាទីតគឺជាច្បាប់ចម្លងណាមួយនៃម៉ូលេគុល DNA ពីរដែលរួមគ្នាបង្កើតជាក្រូម៉ូសូមចម្លង ហើយត្រូវបានតភ្ជាប់ដោយ centromeres របស់ពួកគេ។ ពាក្យនេះត្រូវបានប្រើដរាបណា centromeres នៅតែមានទំនាក់ទំនង។ បន្ទាប់ពីក្រូម៉ូសូមដាច់ដោយឡែកក្នុងអំឡុងពេល anaphase នៃ mitosis ឬ meiosis, strands ត្រូវបានគេហៅថាក្រូម៉ូសូមកូនស្រី។ ក្នុងន័យផ្សេងទៀត, chromatids គឺជាផ្នែកពាក់កណ្តាលនៃក្រូម៉ូសូមចម្លង. សមាសធាតុគីមីនៃក្រូម៉ូសូមគឺ 50% DNA និង 50% ប្រូតេអ៊ីន។
ជាធម្មតាមនុស្សមានក្រូម៉ូសូមដូចគ្នាចំនួន 23 គូនៅក្នុងកោសិកានីមួយៗ (N = 23) ។ ទោះជាយ៉ាងណាក៏ដោយ ចំនួនក្រូម៉ាទីតនឹងជាពហុគុណនៃ 23 ហើយអាចជា 4N, 2N ឬ 1N។ លេខនេះមិនសំដៅទៅលើសំណុំ haploid ឬ diploid ទេ វាសំដៅទៅលើចំនួនក្រូម៉ាទីតនៅក្នុងកោសិកានីមួយៗ ដែលជាពហុគុណនៃសំណុំក្រូម៉ូសូម haploid នៅក្នុងខ្លួន។
- 4N. នៅក្នុងកោសិកាមួយដែលមានក្រូម៉ូសូម 4N មានក្រូម៉ូសូមចំនួន 23 គូ (46 ក្រូម៉ូសូម) ហើយក្រូម៉ូសូមនីមួយៗមានក្រូម៉ូសូមពីរ។ ដូច្នេះមាន 92 chromatids នៅក្នុងកោសិកានីមួយៗ (4N) ។
- 2N. ភ្លាមៗបន្ទាប់ពី mitosis ក្នុងកំឡុងពេលដែលកោសិកាបែងចែកជាពាក់កណ្តាល ក្រូម៉ូសូម 23 គូ (46 chromatids) លេចឡើង។ ទោះយ៉ាងណាក៏ដោយ ក្រូម៉ូសូមមានក្រូម៉ូសូមតែមួយប៉ុណ្ណោះ។ ដូច្នេះមាន 46 chromatids សរុប (2xN) ។ ម្យ៉ាងវិញទៀត កោសិកា haploid ដែលមានក្រូម៉ាទីពីរក្នុងមួយក្រូម៉ូសូមក៏មាន 46 chromatids ផងដែរ។ ទោះជាយ៉ាងណាក៏ដោយនេះមិនកើតឡើងចំពោះមនុស្សទេ។
- 1N. ភ្លាមៗបន្ទាប់ពី meiosis កោសិកានីមួយៗត្រូវបានគេហៅថា gameteមានក្រូម៉ូសូមពាក់កណ្តាល (២៣ ក្រូម៉ូសូម) ។ លើសពីនេះ ក្រូម៉ូសូមនីមួយៗមានក្រូម៉ូសូមតែមួយប៉ុណ្ណោះ។ ដូច្នេះមាន 23 chromatids សរុប (1xN) ។
ក្រូម៉ូសូមតែមួយប្រែទៅជាក្រូម៉ូសូមទ្វេកំឡុងពេលដំណើរការចម្លង (ទ្វេដងនៃ DNA) ។ ក្រូម៉ូសូមទ្វេប្រែទៅជាក្រូម៉ូសូមតែមួយពីរ (ក្រូម៉ូសូមក្លាយជាក្រូម៉ូសូមកូនស្រី) បន្ទាប់ពីការបំបែកនៃ centromere ភ្ជាប់ពួកវា (នៅក្នុង anaphase នៃ mitosis និង anaphase II នៃ meiosis) ។ នោះគឺជា chromatids ក្លាយជាក្រូម៉ូសូមបន្ទាប់ពី centromere ភ្ជាប់ពួកវាបែងចែក។
ការរឹតបន្តឹងក្រូម៉ូសូម៖
ក្រូម៉ូសូម- ក្រុមហ៊ុនដឹកជញ្ជូនសំខាន់នៃសម្ភារៈហ្សែនធានាការចម្លងរបស់វាពីជំនាន់មួយទៅជំនាន់មួយ។
ក្នុងអំឡុងពេលរវាងការបែងចែកកោសិកា (នៅក្នុងដំណាក់កាលនៃ mitosis) ក្រូម៉ូសូមគឺមើលមិនឃើញនៅក្នុងមីក្រូទស្សន៍ពន្លឺ ហើយត្រូវបានតំណាងដោយខ្សែស្រឡាយ chromatin ដែលមិនមានចលនា។ ក្នុងអំឡុងពេលនេះព្រឹត្តិការណ៍ហ្សែនដ៏សំខាន់មួយកើតឡើង - ការចម្លង DNA និងការកើនឡើងទ្វេដងនៃក្រូម៉ូសូមដោយផ្អែកលើវា។ ដរាបណាច្បាប់ចម្លងលទ្ធផលទាំងពីរត្រូវបានរក្សារួមគ្នាដោយ centromere ពួកគេត្រូវបានគេហៅថា chromatids បងស្រី។ ជាមួយនឹងការចាប់ផ្តើមនៃការបែងចែកកោសិកា ក្រូម៉ូសូមវិល និងកាន់តែក្រាស់។ នៅក្រោមមីក្រូទស្សន៍ពន្លឺ វាច្បាស់ណាស់ថាពួកវាមានក្រូម៉ាទីតពីរ។ ក្នុងអំឡុងពេល mitosis, chromatids បំបែក និងក្លាយជាក្រូម៉ូសូមឯករាជ្យ។ដូច្នេះក្នុងអំឡុងពេលវដ្តកោសិការចនាសម្ព័ន្ធនៃក្រូម៉ូសូមឆ្លងកាត់ការផ្លាស់ប្តូរ។
ក្រូម៉ូសូមនីមួយៗមានលក្ខណៈបុគ្គល i.e. កំណត់លក្ខណៈដោយទំហំ រូបរាង និងទីតាំងរបស់ centromere ប៉ុណ្ណោះ។ នៅក្នុងកោសិការាងកាយរបស់សារពាង្គកាយដែលបង្កើតឡើងវិញតាមការរួមភេទ ក្រូម៉ូសូមណាមួយត្រូវបានតំណាងដោយច្បាប់ចម្លងពីរ ឬដូចគ្នា ។ នៅពេលដែលកោសិកាមេរោគត្រូវបានបង្កើតឡើងនៅក្នុង meiosis ពួកវានីមួយៗទទួលបានក្រូម៉ូសូមដូចគ្នាពីរ។ កំឡុងពេលបង្កកំណើត ការផ្គូផ្គងក្រូម៉ូសូមដូចគ្នាត្រូវបានស្ដារឡើងវិញ៖ ក្រូម៉ូសូមមួយនៃគូនីមួយៗគឺជាឪពុក មួយទៀតជាមាតា។
សំណុំនៃលក្ខណៈនៃសំណុំក្រូម៉ូសូម (ចំនួនក្រូម៉ូសូម ទំហំ និងរូបរាងរបស់វា) គឺថេរសម្រាប់កោសិកានៃប្រភេទនីមួយៗ ហើយត្រូវបានគេហៅថា karyotype របស់វា។ karyotype បែងចែករវាងក្រូម៉ូសូមភេទមួយគូ ដែលកំណត់ភេទរបស់សារពាង្គកាយមួយ ហើយក្រូម៉ូសូមផ្សេងទៀតទាំងអស់គឺជា autosomes ។ ការសិក្សាអំពីឥរិយាបថរបស់ក្រូម៉ូសូមក្នុង mitosis និង meiosis ក៏ដូចជាតួនាទីរបស់ក្រូម៉ូសូម ជាពិសេសក្រូម៉ូសូមភេទ ក្នុងការបញ្ជូនលក្ខណៈពីជំនាន់មួយទៅជំនាន់មួយទៀត បាននាំឱ្យមានការបង្កើតទ្រឹស្តីក្រូម៉ូសូមនៃតំណពូជនៅដើមសតវត្សទី 20 ។ សតវត្ស។ ក្រូម៉ូសូមជារឿយៗត្រូវបានគេហៅថាជាសម្ភារៈហ្សែននៃបាក់តេរី និងមេរោគ។ទោះបីជារចនាសម្ព័ន្ធរបស់វាខុសពីក្រូម៉ូសូមនៃសារពាង្គកាយ eukaryotic ក៏ដោយ។
សំណុំនៃក្រូម៉ូសូម
សំណុំក្រូម៉ូសូមតែមួយ (haploid) គឺជាលក្ខណៈនៃកោសិកាមេរោគ ( gametes), ទ្វេ (diploid) - សម្រាប់កោសិកា somatic ។
- សំណុំតែមួយប្រែទៅជាសំណុំពីរដងនៅពេលនៃការបង្កកំណើត (ការបញ្ចូលគ្នានៃមេជីវិតឈ្មោលនិងស៊ុត) ។
- សំណុំទ្វេដងប្រែទៅជាសំណុំតែមួយនៅពេលនៃការបង្វែរឯករាជ្យនៃក្រូម៉ូសូមទ្វេនៅក្នុង anaphase I នៃ meiosis ។
នៅក្នុងសំណុំទ្វេរដង មនុស្សមានក្រូម៉ូសូមចំនួន 46 ។
បញ្ហាលើចំនួនក្រូម៉ូសូម៖
- ប្រសិនបើមានក្រូម៉ូសូមចំនួន 12 នៅក្នុងកោសិកាបន្តពូជ បន្ទាប់មកនៅក្នុងស៊ុតបង្កកំណើត ( ហ្សីហ្គោត) និងនៅក្នុង កោសិកា somatic(ថ្លើម ស្បែក សាច់ដុំ។ល។) - 24 ក្រូម៉ូសូមនីមួយៗ។
- ហើយផ្ទុយទៅវិញប្រសិនបើមានក្រូម៉ូសូមចំនួន 36 នៅក្នុងស្នូលនៃកោសិកា somatic (ពោះវៀន ខួរក្បាល។ ស្នូលហ្សីហ្គោត។
តើអ្នកអាចស្វែងរកបញ្ជីមេធាវីនៅប្រទេសរុស្ស៊ីនៅឯណានៅលើអ៊ីនធឺណិត?
ព័ត៌មានអំពីអង្គជំនុំជម្រះនៃអង្គភាពដែលមានធាតុផ្សំនៃសហព័ន្ធរុស្ស៊ី (ចំនួនសមាជិក ប្រធានសភា អាស័យដ្ឋាន លេខទូរស័ព្ទ គេហទំព័រ) បញ្ជីការិយាល័យច្បាប់ ការិយាល័យ ក្រុមប្រឹក្សា បញ្ជីមេធាវីដែលរួមបញ្ចូលក្នុងបញ្ជីឈ្មោះមេធាវី។ ស្ថាប័នដែលមានធាតុផ្សំនៃសហព័ន្ធរុស្ស៊ី ក៏ដូចជាព័ត៌មានអំពីមេធាវីជាក់លាក់ ឬពិនិត្យមើលថាតើបុគ្គលនោះជាមេធាវី ឬអត់ ថាតើអំណាចរបស់គាត់ត្រូវបានបញ្ចប់ អាចរកបាននៅលើគេហទំព័រ៖ ចុះឈ្មោះមេធាវី
តើឈ្មោះដំបូង Galina មានន័យយ៉ាងម៉េច?
Galina - មកពីពាក្យក្រិក galene ដែលមានន័យថាស្ងប់ស្ងាត់ភាពស្ងប់ស្ងាត់។ ទម្រង់ដែលទទួលបាន៖ Alya, Gala, Galinka, Galyunya, Galyusya, Galyukha, Galyusha, Galya, Pebble, Galenka, Galonka, Galka, Galochka, Galchonok, Galchonochek ។ ឈ្មោះថ្ងៃ៖ ថ្ងៃទី ២៣ ខែកុម្ភៈ ថ្ងៃទី ២៣ ខែមីនា ថ្ងៃទី ២៩ ខែមេសា។ (យកចិត្តទុកដាក់! បន្ថែមទៀត
តើ Prada បញ្ចេញក្លិនដំបូងនៅឆ្នាំណា?
Prada គឺជាក្រុមហ៊ុនឯកជនដ៏ល្បីរបស់ប្រទេសអ៊ីតាលីដែលមានឯកទេសក្នុងការផលិតសំលៀកបំពាក់ ស្បែកជើង និងគ្រឿងប្រើប្រាស់ទាន់សម័យ ដែលជាម្ចាស់ហាងម៉ូដ និងយីហោដែលមានឈ្មោះដូចគ្នា។ រចនាបថរបស់ Prada គឺអាចស្គាល់បាន វាមាន laconic តឹងរឹង និងឆើតឆាយ។ ប្រវត្តិសាស្រ្តនៃម៉ាកនេះបានចាប់ផ្តើមនៅទីក្រុង Milan ក្នុងឆ្នាំ 1913 នៅពេលដែល Mario Prada -
តើ botulism ត្រូវបានព្យាបាលយ៉ាងដូចម្តេច?
botulism គឺជាអ្វី? Botulism គឺជាជំងឺឆ្លងស្រួចស្រាវដែលកើតឡើងជាលទ្ធផលនៃការទទួលទានអាហារ ទឹក ឬសារធាតុ aerosols ដែលត្រូវបានបំពុលដោយ bacillus Clostridium botulinum ដែលបង្កើតជាស្ពែរ។ បង្ហាញថាខ្លួនវាជាការរំលោភលើការកន្ត្រាក់នៃសាច់ដុំដែលមានសរសៃនិងរលោងត្រូវបានស្រូបយកពីបំពង់អាហារឬសួត
តើអ្វីទៅជាគុណសម្បត្តិនិងគុណវិបត្តិនៃចង្កៀង LED
ចង្កៀង LED ឬអំពូល LED ប្រើ LEDs ជាប្រភពពន្លឺ ហើយប្រើសម្រាប់ភ្លើងក្នុងផ្ទះ ឧស្សាហកម្ម និងតាមដងផ្លូវ។ អំពូល LED គឺជាប្រភពពន្លឺដែលមិនប៉ះពាល់ដល់បរិស្ថានបំផុត។ គោលការណ៍ពន្លឺ LED អនុញ្ញាតឱ្យប្រើសមាសធាតុសុវត្ថិភាពក្នុងការផលិត និងប្រតិបត្តិការនៃចង្កៀងខ្លួនឯង។ ខ្ញុំមិនប្រើអំពូល LED ទេ។
អ្វីដែលត្រូវផ្តល់ឱ្យគ្រូនៅថ្ងៃគ្រូ
ទិវាគ្រូបង្រៀនគឺជាថ្ងៃឈប់សម្រាកវិជ្ជាជីវៈសម្រាប់បុគ្គលិកអប់រំ។ ប្រវត្តិនៃថ្ងៃឈប់សម្រាក លក្ខខណ្ឌជាប្រវត្តិសាស្ត្រសម្រាប់ការបង្កើតទិវាគ្រូបង្រៀន គឺជាសន្និសីទអន្តររដ្ឋាភិបាលពិសេសស្តីពីស្ថានភាពគ្រូបង្រៀនដែលបានធ្វើឡើងនៅថ្ងៃទី 5 ខែតុលា ឆ្នាំ 1966 នៅទីក្រុងប៉ារីស។
យកចិត្តទុកដាក់! ការព្យាបាលទាំងអស់ត្រូវតែអនុវត្តបន្ទាប់ពីការពិគ្រោះយោបល់ជាមួយវេជ្ជបណ្ឌិតជំនាញ និងស្ថិតក្រោមការត្រួតពិនិត្យជាប្រចាំរបស់គាត់។ អ្នកជំងឺខ្លួនឯងទទួលខុសត្រូវចំពោះផលវិបាកនៃការប្រើប្រាស់ព័ត៌មានដែលបានផ្តល់។ ជំងឺរលាកក្រពះគឺជាការរលាកនៃភ្នាសរំអិល (ខាងក្នុង) នៃជញ្ជាំងក្រពះ។ នៅពេលដែលការរលាករាលដាលដល់ duodenum, gastroduodenitis ត្រូវបានបង្កើតឡើង។ មានពីរប្រភេទ
តើស្មៀនពន្លឺជាកម្មសិទ្ធិរបស់លំដាប់អ្វី?
ស្រមោចស្រាល (lat. Accipiter novaehollandiae) គឺជាសត្វស្លាបព្រៃនៃគ្រួសារ hawk ដែលជាទូទៅនៅក្នុងព្រៃនៃភាគខាងជើង និងភាគខាងកើតនៃប្រទេសអូស្ត្រាលី។ រាជាណាចក្រ៖ ប្រភេទសត្វ៖ អង្កត់ធ្នូ ថ្នាក់៖ បក្សី លំដាប់៖ Falconiformes គ្រួសារ៖ Accipitridae គ្រួសាររង៖ Accipites ពូជ៖ ស្ទាំងពិត ប្រភេទសត្វ៖ ស្ទាំងពន្លឺ ស្ទាំងពន្លឺ &md
តើទិវាតន្ត្រីអន្តរជាតិត្រូវបានបង្កើតឡើងនៅពេលណា?
ទិវាតន្ត្រីអន្តរជាតិត្រូវបានប្រារព្ធនៅថ្ងៃទី 1 ខែតុលា។ ទិវាតន្ត្រីអន្តរជាតិត្រូវបានបង្កើតឡើងនៅថ្ងៃទី 1 ខែតុលា ឆ្នាំ 1975 ដោយសេចក្តីសម្រេចរបស់អង្គការយូណេស្កូ។ អ្នកផ្តួចផ្តើមគំនិតបង្កើតទិវាតន្ត្រីអន្តរជាតិគឺ Dmitry Shostakovich ដែលជាអ្នកនិពន្ធបុរាណនៃសតវត្សទី 20 ។ វាត្រូវបានប្រារព្ធជារៀងរាល់ឆ្នាំនៅទូទាំងពិភពលោកជាមួយនឹងកម្មវិធីប្រគុំតន្ត្រីធំ ៗ ដោយមានការចូលរួមពីសិល្បករនិងក្រុមសិល្បៈល្អបំផុត។ នៅក្នុងអ៊ី
របៀបដែលបុណ្យណូអែលត្រូវបានប្រារព្ធនៅប្រទេសដាណឺម៉ាក
បុណ្យណូអែលត្រូវបានប្រារព្ធនៅទូទាំងពិភពលោក ហើយប្រទេសជាច្រើនមានទំនៀមទម្លាប់ និងទំនៀមទម្លាប់រៀងៗខ្លួនដែលទាក់ទងនឹងការប្រារព្ធពិធីរបស់ខ្លួន។ តាំងពីបុរាណកាល ទិវាកំណើតរបស់ព្រះគ្រីស្ទត្រូវបានចាត់ថ្នាក់ក្នុងចំណោមថ្ងៃឈប់សម្រាកដ៏អស្ចារ្យដោយសាសនាចក្រ នេះបើយោងតាមដំណឹងល្អ ដែលពណ៌នាព្រឹត្តិការណ៍នេះថាអស្ចារ្យ និងអស្ចារ្យ៖ «ខ្ញុំប្រកាសប្រាប់អ្នក» ទេវតាប្រាប់អ្នកគង្វាលបេថ្លេហិម , "អស្ចារ្យ
តើមនុស្សល្បីណាខ្លះបានស្លាប់នៅថ្ងៃទី ២៧ ខែធ្នូ
ថ្ងៃទី 27 ខែធ្នូគឺជាថ្ងៃទី 361 នៃឆ្នាំ (362 ឆ្នាំបង្គ្រប់) នៅក្នុងប្រតិទិនហ្គ្រេហ្គោរៀន។ នៅសល់ 4 ថ្ងៃទៀតរហូតដល់ដំណាច់ឆ្នាំ ថ្ងៃឈប់សម្រាករបស់ប្រទេសរុស្ស៊ី ទិវាអ្នកជួយសង្គ្រោះ។ ថ្ងៃនៃសាវក និងជាអ្នកផ្សាយដំណឹងល្អ ចន (ថ្ងៃឈប់សម្រាកនៅក្នុងនិកាយកាតូលិក និងប្រូតេស្តង់)។ ឈ្មោះថ្ងៃ Firs, Leonid, Gennady, Hilarion, Nikolai ។ ព្រឹត្តិការណ៍នៃថ្ងៃទី 27 ខែធ្នូសតវត្សទី 6
1. កោសិកា Somatic ផ្ទុយទៅនឹងកោសិកាមេរោគ
- អសមត្ថភាពក្នុងការបែងចែក
- មាន n ក្រូម៉ូសូម
- មានក្រូម៉ូសូម 2n
កោសិកា Somatic- កោសិកានៃរាងកាយរបស់សត្វឬរុក្ខជាតិ (ឧទាហរណ៍កោសិកាមិនបន្តពូជ) ។ នៅក្នុងកោសិកា somatic (កោសិការាងកាយ) ចំនួននៃក្រូម៉ូសូមជាធម្មតាមានទំហំធំជាងពីរដងនៃកោសិកាដំណុះចាស់ទុំ។ នេះត្រូវបានពន្យល់ដោយការពិតដែលថាក្នុងអំឡុងពេលបង្កកំណើត ពាក់កណ្តាលនៃក្រូម៉ូសូមបានមកពីរាងកាយរបស់ម្តាយ (នៅក្នុងស៊ុត) និងពាក់កណ្តាលពីឪពុក (នៅក្នុងមេជីវិតឈ្មោល) ពោលគឺឧ។ នៅក្នុងស្នូលនៃកោសិកា somatic ក្រូម៉ូសូមទាំងអស់ត្រូវបានផ្គូផ្គង។ ជាងនេះទៅទៀត ក្រូម៉ូសូមនៃគូនីមួយៗមានភាពខុសប្លែកពីក្រូម៉ូសូមផ្សេងទៀត។ ក្រូម៉ូសូមបែបនេះ មានរូបរាង និងទំហំដូចគ្នាបេះបិទ ផ្ទុកហ្សែនដូចគ្នា ត្រូវបានគេហៅថា homologous ។ មួយក្នុងចំនោមក្រូម៉ូសូមដូចគ្នាគឺជាច្បាប់ចម្លងនៃក្រូម៉ូសូមមាតា ហើយមួយទៀតគឺចម្លងនៃក្រូម៉ូសូមប៉ា។ សំណុំក្រូម៉ូសូម តំណាងដោយក្រូម៉ូសូមផ្គូផ្គងត្រូវបានគេហៅថា double ឬ diploid ហើយត្រូវបានកំណត់ 2 ន.
2. កោសិកាមនុស្សខុសពីកោសិកាសត្វស្វា
- វត្តមាននៃ ribosomes
- ចំនួនក្រូម៉ូសូម
- កង្វះ DNA
កោសិការបស់មនុស្សខុសគ្នាពីកោសិកាស្វានហ្សីក្នុងចំនួនក្រូម៉ូសូម: មនុស្សមានក្រូម៉ូសូមចំនួន 23 គូ សត្វស្វាដ៏អស្ចារ្យមាន 24 ។ Chimpanzees គឺជាសាច់ញាតិជិតបំផុតរបស់យើងដែលមានស្ទើរតែដូចគ្នាទៅនឹងប្រភេទ karyotype របស់យើង (សត្វស្វា Chimpanzee គឺនៅជិតយើងជាពិសេសទាក់ទងនឹងក្រូម៉ូសូម) ។
សំណុំក្រូម៉ូសូមនៃកោសិកា somatic ដែលក្រូម៉ូសូមនីមួយៗមានគូ - ទ្វេ ឬ diploid ( 2 ន) បរិមាណ DNA ដែលត្រូវគ្នានឹងសំណុំក្រូម៉ូសូម diploid គឺ − 2 វិ. ពីគូនៃក្រូម៉ូសូមដូចគ្នា មានតែមួយប៉ុណ្ណោះចូលទៅក្នុងកោសិកាមេរោគ ដូច្នេះសំណុំក្រូម៉ូសូមនៃ gametes គឺទោល ឬ haploid ( 1 ន).
3. Sphagnum spores គឺជាកោសិកាពិសេសដែលអនុវត្ត
- ការបន្តពូជផ្លូវភេទ
- ការបន្តពូជផ្លូវភេទ
- ការបែកខ្ញែក
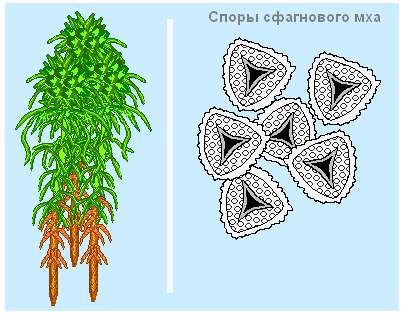 Sphagnumអាចបន្តពូជដោយ spores និងលូតលាស់។
Sphagnumអាចបន្តពូជដោយ spores និងលូតលាស់។
ការបន្តពូជដោយ spores គឺជាវិធីសាស្រ្តចម្បងសម្រាប់ការបែកខ្ចាត់ខ្ចាយនៃ sphagnum ក្នុងចម្ងាយឆ្ងាយ - តំបន់ថ្មីឬតំបន់ដែលរងការខូចខាតដោយភ្លើងឬសកម្មភាពសេដ្ឋកិច្ច។ នៅផ្នែកខាងលើនៃរុក្ខជាតិប្រអប់នៅលើ stalks ទុំដែលក្នុងនោះ spores ត្រូវបានបង្កើតឡើង។
ដើម្បីបង្កើតរុក្ខជាតិពី spore មួយវាចាំបាច់ដែលវាធ្លាក់នៅលើដីសមរម្យ - peat មានសំណើម។ ស្ព័រធំជាងមានការផ្គត់ផ្គង់សារធាតុចិញ្ចឹមកាន់តែច្រើន ដូច្នេះហើយមានឱកាសល្អក្នុងការរង់ចាំលក្ខខណ្ឌត្រឹមត្រូវ។
4. កោសិកាពិសេសដែលមានជំនួយដែលការបន្តពូជផ្លូវភេទត្រូវបានអនុវត្តត្រូវបានគេហៅថា
- ជម្លោះ
- blastomeres
- oocytes
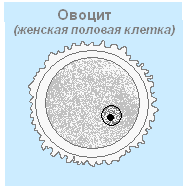
ដំណើរការនៃការអភិវឌ្ឍកោសិកាមេជីវិតញី - ស៊ុតពី primordia នៃ epithelium (ជាលិកា oogenic) ។ Ontogenesis កើតឡើងនៅក្នុងអូវែរជាបីដំណាក់កាល៖ ការបន្តពូជ ការលូតលាស់ និងការពេញវ័យ។ ដំណាក់កាលដំបូង - ការបន្តពូជ - កោសិកានៃជាលិកា diploid នៃ epithelium rudimentary បែងចែកម្តងហើយម្តងទៀតតាមរយៈ mitosis បង្កើតកោសិកា diploid ។ អូតូស៊ីត (oocytes)លំដាប់ដំបូង។
5. រូបភាពបង្ហាញ 
- ក្រូម៉ូសូម
- chromatid
- កណ្តាល
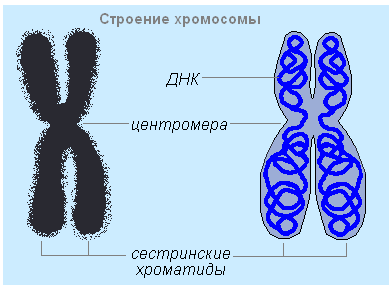 ក្រូម៉ូសូម- រចនាសម្ព័ន្ធសំខាន់បំផុតនៃស្នូល។ ក្នុងអំឡុងពេលរវាងការបែងចែកកោសិកា (អន្តរដំណាក់កាល) ក្រូម៉ូសូមមិនអាចមើលឃើញទេ។ ពួកវាអាចស្ថិតនៅក្នុងរដ្ឋពីរ: spiralized - ខ្លីនិងក្រាស់អាចមើលឃើញយ៉ាងច្បាស់នៅក្នុងមីក្រូទស្សន៍ពន្លឺ; despiralized (untwisted) - វែងនិងស្តើងហៅថា chromatin (សារធាតុពណ៌) ។ មិនមានភាពខុសគ្នានៅក្នុងសមាសធាតុគីមីនៃក្រូម៉ូសូមនិងក្រូម៉ាទីន; ទាំងនេះគឺជាម៉ូលេគុល DNA និងប្រូតេអ៊ីន (ប្រូតេអ៊ីន) ដែលរួមគ្នាបង្កើតជានុយក្លេអូទីត។ នៅក្នុង metaphase នៃកោសិកាបែងចែក វាអាចមើលឃើញយ៉ាងច្បាស់ថា ក្រូម៉ូសូមនីមួយៗមានពាក់កណ្តាលស៊ីមេទ្រីបណ្តោយពីរ - chromatids ។ នៅក្នុងកោសិកាឯកទេស ក្រូម៉ូសូមជាធម្មតាមានក្រូម៉ាទីតតែមួយ។ ក្រូម៉ូសូមមានការរឹតបន្តឹងចម្បងដែល centromere ស្ថិតនៅ; constriction បែងចែកក្រូម៉ូសូមទៅជាដៃពីរដែលមានប្រវែងស្មើគ្នា ឬខុសគ្នា។ centromere ភ្ជាប់ទាំង chromatids និងបម្រើជាចំណុចភ្ជាប់សម្រាប់ spindle filaments នៃការបែងចែកកោសិកា។ មុខងារចម្បងនៃក្រូម៉ូសូមគឺការផ្ទុក និងការបញ្ជូនព័ត៌មានតំណពូជ ដែលជាក្រុមហ៊ុនដឹកជញ្ជូនដែលជាម៉ូលេគុល DNA - ហ្សែន - ដែលទទួលខុសត្រូវចំពោះលក្ខណៈជាក់លាក់នីមួយៗ ឬទ្រព្យសម្បត្តិនៃសារពាង្គកាយ។ ព័ត៌មានតំណពូជត្រូវបានបញ្ជូនពីកោសិកាមួយទៅកោសិកាដោយការបង្កើនទ្វេដងនៃម៉ូលេគុល DNA (ការចម្លង) ការចម្លង និងការបកប្រែ។
ក្រូម៉ូសូម- រចនាសម្ព័ន្ធសំខាន់បំផុតនៃស្នូល។ ក្នុងអំឡុងពេលរវាងការបែងចែកកោសិកា (អន្តរដំណាក់កាល) ក្រូម៉ូសូមមិនអាចមើលឃើញទេ។ ពួកវាអាចស្ថិតនៅក្នុងរដ្ឋពីរ: spiralized - ខ្លីនិងក្រាស់អាចមើលឃើញយ៉ាងច្បាស់នៅក្នុងមីក្រូទស្សន៍ពន្លឺ; despiralized (untwisted) - វែងនិងស្តើងហៅថា chromatin (សារធាតុពណ៌) ។ មិនមានភាពខុសគ្នានៅក្នុងសមាសធាតុគីមីនៃក្រូម៉ូសូមនិងក្រូម៉ាទីន; ទាំងនេះគឺជាម៉ូលេគុល DNA និងប្រូតេអ៊ីន (ប្រូតេអ៊ីន) ដែលរួមគ្នាបង្កើតជានុយក្លេអូទីត។ នៅក្នុង metaphase នៃកោសិកាបែងចែក វាអាចមើលឃើញយ៉ាងច្បាស់ថា ក្រូម៉ូសូមនីមួយៗមានពាក់កណ្តាលស៊ីមេទ្រីបណ្តោយពីរ - chromatids ។ នៅក្នុងកោសិកាឯកទេស ក្រូម៉ូសូមជាធម្មតាមានក្រូម៉ាទីតតែមួយ។ ក្រូម៉ូសូមមានការរឹតបន្តឹងចម្បងដែល centromere ស្ថិតនៅ; constriction បែងចែកក្រូម៉ូសូមទៅជាដៃពីរដែលមានប្រវែងស្មើគ្នា ឬខុសគ្នា។ centromere ភ្ជាប់ទាំង chromatids និងបម្រើជាចំណុចភ្ជាប់សម្រាប់ spindle filaments នៃការបែងចែកកោសិកា។ មុខងារចម្បងនៃក្រូម៉ូសូមគឺការផ្ទុក និងការបញ្ជូនព័ត៌មានតំណពូជ ដែលជាក្រុមហ៊ុនដឹកជញ្ជូនដែលជាម៉ូលេគុល DNA - ហ្សែន - ដែលទទួលខុសត្រូវចំពោះលក្ខណៈជាក់លាក់នីមួយៗ ឬទ្រព្យសម្បត្តិនៃសារពាង្គកាយ។ ព័ត៌មានតំណពូជត្រូវបានបញ្ជូនពីកោសិកាមួយទៅកោសិកាដោយការបង្កើនទ្វេដងនៃម៉ូលេគុល DNA (ការចម្លង) ការចម្លង និងការបកប្រែ។
6. ក្រូម៉ូសូមមានម៉ូលេគុលមួយ។
- គ្លុយកូស
ក្រូម៉ូសូមមួយមាន ឌីអិនអេនិងកំប្រុក។ ស្មុគ្រស្មាញនៃប្រូតេអ៊ីនដែលភ្ជាប់ទៅនឹង DNA បង្កើតជាក្រូម៉ាទីន។ ប្រូតេអ៊ីនដើរតួនាទីយ៉ាងសំខាន់ក្នុងការវេចខ្ចប់ម៉ូលេគុល DNA នៅក្នុងស្នូល។ មុនពេលការបែងចែកកោសិកា DNA ត្រូវបានចងយ៉ាងតឹងរ៉ឹងដើម្បីបង្កើតជាក្រូម៉ូសូម ហើយប្រូតេអ៊ីននុយក្លេអ៊ែរ - អ៊ីស្តូន - គឺចាំបាច់សម្រាប់ការបត់ត្រឹមត្រូវនៃ DNA ដែលជាលទ្ធផលដែលបរិមាណរបស់វាថយចុះច្រើនដង។ ក្រូម៉ូសូមនីមួយៗត្រូវបានបង្កើតឡើងដោយម៉ូលេគុលមួយ។ ឌីអិនអេ.
7. ក្នុងអំឡុងពេលបន្តពូជផ្លូវភេទ ប្រភេទនីមួយៗរក្សាចំនួនក្រូម៉ូសូមថេរពីជំនាន់មួយទៅជំនាន់មួយ ដោយសារ
- មីតូស៊ីស
- meiosis
- gametogenesis
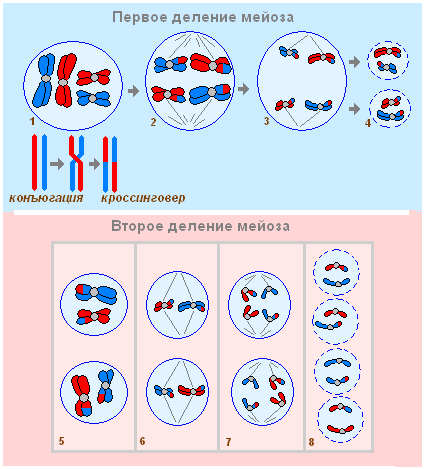
Meiosis គឺជាប្រភេទពិសេសនៃការបែងចែកកោសិកា ដែលជាលទ្ធផលដែលកោសិកាកូនស្រីក្លាយទៅជា haploid ។ Meiosis មានការបែងចែកពីរដែលកើតឡើងពីមួយទៅមួយ; ការចម្លង DNA កើតឡើងតែម្តងប៉ុណ្ណោះ។ សារធាតុ និងថាមពលចាំបាច់ទាំងអស់ (ATP) កកកុញក្នុងដំណាក់កាល I, អន្តរដំណាក់កាល II គឺអវត្តមានជាក់ស្តែង។
ការបែងចែកដំបូងនៃ meiosis ។
1. Prophase I - spiralization នៃក្រូម៉ូសូមជាមួយនឹងការបង្កើត bivalents (រចនាសម្ព័ន្ធដែលមានក្រូម៉ូសូមពីរ និង chromatids បួន) chromosome conjugation - ប្រមូលផ្តុំក្រូម៉ូសូម homologous ពីរតាមបណ្តោយប្រវែងទាំងមូល ហើយឆ្លងកាត់ - ការផ្លាស់ប្តូរផ្នែកនៃ chromosomes ដូចគ្នា ការបាត់ខ្លួន។ nucleolus ការបង្កើត spindle ការបំផ្លាញភ្នាសនុយក្លេអ៊ែរ។ (2n 2chr 4c) - 1n bivalent
2. Metaphase I – ការរៀបចំគូ (bivalents) នៃក្រូម៉ូសូមដូចគ្នានៅអេក្វាទ័រនៃកោសិកា។ (2n 2chr 4c) - 1n bivalent
3. Anaphase I – ការបង្វែរក្រូម៉ូសូម homologous ដែលមានក្រូម៉ាទីពីរទៅប៉ូលទល់មុខនៃកោសិកា (ចំនួនក្រូម៉ូសូមត្រូវបានកាត់បន្ថយ)។ (1n 2chr 2c)- នៅបង្គោលនីមួយៗនៃកោសិកា។
4. Telophase I - ការបង្កើតស្នូល ការបែងចែក cytoplasm - ការបង្កើតកោសិកាកូនស្រីពីរ។ (1n 2chr 2c)
ការបែងចែកទីពីរនៃ meiosis ។
5. Prophase II – spiralization បន្តិចនៃក្រូម៉ូសូម, ការបង្កើត spindle, ការបំផ្លិចបំផ្លាញនៃភ្នាសនុយក្លេអ៊ែរ។ (1n 2chr 2c)
6. Metaphase II - ការរៀបចំតាមលំដាប់នៃក្រូម៉ូសូមដែលមានក្រូម៉ាទីតពីរនៅអេក្វាទ័រនៃកោសិកា។ (1n 2chr 2c)
7. Anaphase II - ការបង្វែរក្រូម៉ាទីតកូនស្រីទៅប៉ូលទល់មុខនៃកោសិកា។ (1n 1chr 1c)- នៅបង្គោលនីមួយៗ។
8. Telophase II - ការបាត់ខ្លួននៃ spindle, despiralization នៃក្រូម៉ូសូម, ការបង្កើតភ្នាសនុយក្លេអ៊ែរ, ការបែងចែកនៃ cytoplasm, ការបង្កើតភ្នាសកោសិកាថ្មី, ការបង្កើតកោសិកាកូនស្រីបួន។ (1n 1chr 1c)- នៅក្នុងស្នូលនីមួយៗដែលបានបង្កើតឡើង។
Meiosis បម្រើជាមូលដ្ឋានសម្រាប់ការបន្តពូជផ្លូវភេទ និងភាពប្រែប្រួលចម្រុះ។
8. ក្រូម៉ូសូមរួមបញ្ចូល
- ATP និង mRNA
- RNA និងប្រូតេអ៊ីន
- DNA និងប្រូតេអ៊ីន
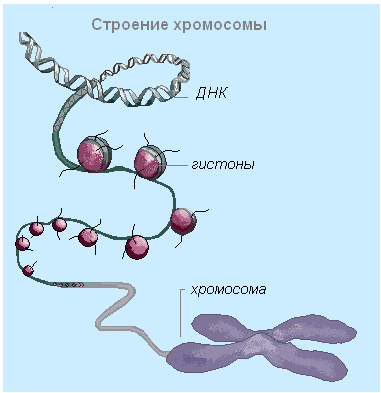
នៅក្នុងកោសិកា interphase, chromatin មានទម្រង់នៃរចនាសម្ព័ន្ធដូចខ្សែស្រឡាយល្អិតល្អន់ដែលមានម៉ូលេគុល DNA និងប្រូតេអ៊ីន(នុយក្លេអូប្រូតេអ៊ីន) សំបក។ ក្នុងការបែងចែកកោសិកា ក្រូម៉ាទីនបង្កើតជាវង់ និងបង្កើតជាក្រូម៉ូសូម។ ក្រូម៉ូសូមមួយមានក្រូម៉ាទីតពីរ ហើយបន្ទាប់ពីការបែងចែកនុយក្លេអ៊ែរ វាក្លាយជាក្រូម៉ាទីតតែមួយ។
ម៉ូលេគុល DNA នៅក្នុងក្រូម៉ូសូមត្រូវបានផ្សារភ្ជាប់យ៉ាងជិតស្និទ្ធជាមួយនឹងប្រូតេអ៊ីនពីរប្រភេទ - អ៊ីស្តូន (ប្រូតេអ៊ីនមូលដ្ឋាន) និង non-histones (ប្រូតេអ៊ីនអាស៊ីត) ។ Histones គឺជាប្រូតេអ៊ីនតូចៗដែលមានមាតិកាខ្ពស់នៃអាស៊ីតអាមីណូដែលត្រូវបានចោទប្រកាន់ (lysine និង arginine) ដែលវាចាំបាច់សម្រាប់ការប្រមូលផ្តុំ និងការវេចខ្ចប់ខ្សែស្រឡាយ។ ឌីអិនអេចូលទៅក្នុងក្រូម៉ូសូម។
9. ក្រូម៉ូសូមទ្វេដងកើតឡើង
- អាណាហ្វាស
- អន្តរដំណាក់កាល
- ព្យាករណ៍
Interphase - ការរៀបចំកោសិកាសម្រាប់ការបែងចែកមានបីដំណាក់កាល
ការរចនានៃសម្ភារៈហ្សែន។
1 ន- សំណុំក្រូម៉ូសូម haploid
2 ន- សំណុំក្រូម៉ូសូម diploid
chr- បរិមាណ
chromatids នៅលើក្រូម៉ូសូមមួយ។
គ- ចំនួនក្រូម៉ូសូមនៅក្នុងសំណុំនៃក្រូម៉ូសូម
2 គ- បរិមាណ DNA ដែលត្រូវនឹងសំណុំក្រូម៉ូសូម diploid
10. កោសិកា Somatic ត្រូវបានបង្កើតឡើងជាលទ្ធផល
- មីតូស៊ីស
- ការបង្កកំណើត
- oogenesis
11. leukocytes របស់មនុស្សមាន... chromosome(s)
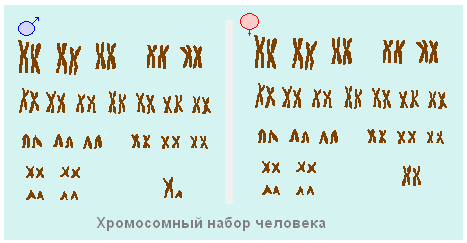
ក្រូម៉ូសូមរបស់មនុស្សមានហ្សែន (៤៦ គ្រឿង),បង្កើត 23 គូ។ មួយគូនៃឈុតនេះកំណត់ភេទរបស់មនុស្សម្នាក់។ សំណុំក្រូម៉ូសូមរបស់ស្ត្រីមានក្រូម៉ូសូម X ចំនួនពីរដែលជារបស់បុរស - មួយ X និងមួយ Y ។ កោសិកាផ្សេងទៀតទាំងអស់នៃរាងកាយមនុស្សមានផ្ទុកមេជីវិតឈ្មោល និងស៊ុតពីរដង។
12. ការបំបែករបស់បងស្រី chromatids កើតឡើងនៅក្នុង ... mitosis
- អាណាហ្វាស
- metaphase
- ព្យាករណ៍
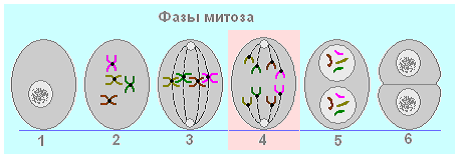
IN អាណាហ្វាស(4) Sister chromatids ត្រូវបានបំបែកនៅក្រោមសកម្មភាពរបស់ spindle: ដំបូងនៅក្នុងតំបន់ centromere ហើយបន្ទាប់មកតាមបណ្តោយប្រវែងទាំងមូល។ ចាប់ពីពេលនេះតទៅ ពួកគេក្លាយជាក្រូម៉ូសូមឯករាជ្យ។ ខ្សែស្រឡាយ spindle លាតសន្ធឹងពួកវាទៅបង្គោលផ្សេងគ្នា។ ដូច្នេះដោយសារតែអត្តសញ្ញាណរបស់កូនស្រី chromatids ប៉ូលទាំងពីរនៃកោសិកាមានសម្ភារៈហ្សែនដូចគ្នា: ដូចគ្នាទៅនឹងអ្វីដែលនៅក្នុងកោសិកាមុនពេលចាប់ផ្តើមនៃ mitosis ។
បរិមាណនៃសម្ភារៈហ្សែន 2n 1chr 2cនៅបង្គោលនីមួយៗនៃកោសិកា។
13. Zygote ផ្ទុយទៅនឹង gamete
- បង្កើតឡើងក្នុងអំឡុងពេល mitosis
- មានក្រូម៉ូសូម 2n
- បង្កើតឡើងក្នុងអំឡុងពេលនៃដំណើរការ meiosis
ហ្សីហ្គោត គឺជាកោសិកាដំបូងនៃសារពាង្គកាយថ្មីមួយ ដែលមាននៅក្នុងស្នូលនៃសារធាតុហ្សែននៃមេជីវិតឈ្មោល និងស៊ុត ពោលគឺឧ។ សំណុំក្រូម៉ូសូម diploid ត្រូវបានស្ដារឡើងវិញ៖ =n + n = 2n.
14. កោសិកា somatic ថ្មីនៅក្នុងរាងកាយរបស់ដង្កូវនាងមួយត្រូវបានបង្កើតឡើងជាលទ្ធផល
- មីតូស៊ីស
- gametogenesis
- meiosis
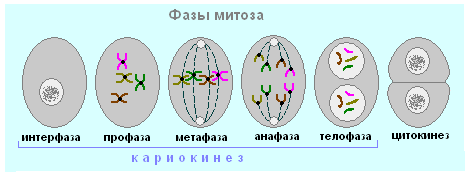
មីតូស៊ីស- ការបែងចែកដោយប្រយោលនៃកោសិកា eukaryotic somatic រួមមានបួនដំណាក់កាល។ ជាលទ្ធផលនៃ mitosis សម្ភារៈតំណពូជនៃកោសិកាម្តាយត្រូវបានកើនឡើងទ្វេដងហើយបន្ទាប់មកចែកចាយស្មើៗគ្នារវាងកោសិកាកូនស្រី។ រយៈពេលនៃ mitosis នៅក្នុងកោសិកាសត្វគឺ 30-60 នាទីហើយនៅក្នុងកោសិការុក្ខជាតិ - 2-3 ម៉ោង។
ដង្កូវដី។

| ដំណាក់កាល mitosis | គ្រោងការណ៍ | បរិមាណនៃសម្ភារៈហ្សែន | ដំណើរការ |
| ប្រូហ្វាស | 2n 2chr 4c |
|
|
| មេតាហ្វាស | 2n 2chr 4c |
|
|
| អាណាផាស | 2n 1chr 2c នៅបង្គោលនីមួយៗនៃកោសិកា |
|
|
| តេឡូហ្វាស | 2n 1chr 2c |
|
|
| 2n 1chr 2c |
|
15. ជាលទ្ធផលនៃ meiosis,
- កោសិកាកូនស្រី 2 ដែលមានសំណុំក្រូម៉ូសូម haploid
- កោសិកាកូនស្រី 2 ដែលមានសំណុំក្រូម៉ូសូម diploid
- កោសិកាកូនស្រី 4 ដែលមានសំណុំក្រូម៉ូសូម haploid
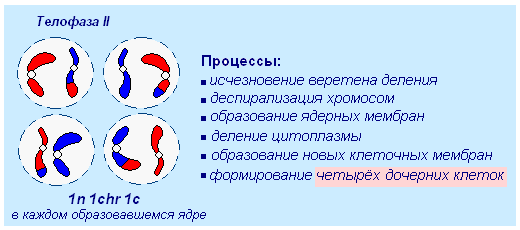
Telophase II ។ បន្ទាប់ពីការបញ្ចប់នៃការបែងចែក meiotic ទី 1 ដំណាក់កាលខ្លីនៃផ្នែក meiotic ទីពីរកើតឡើង។ ជាងនេះទៅទៀត នៅដំណាក់កាលនេះ ការចម្លង DNA (ទ្វេដង) មិនកើតឡើងទេ ហើយដូច្នេះ ភាពច្របូកច្របល់មិនត្រូវបានស្ដារឡើងវិញទេ។