See artikkel tutvustab lugejat kõige lihtsamate organismide ehitusega, nimelt keskendub see kokkutõmbuva vakuooli struktuurile, mis täidab eritus- (ja muud) funktsiooni, räägib kõige lihtsamate organismide tähendusest ja kirjeldab nende tekkeviise. nende olemasolu keskkonnas.
Kontraktiivne vakuool. Kontseptsioon
Vacuool (prantsuse vacuole, ladina sõnast vacuus - tühi), sfäärilised väikesed õõnsused taime- ja loomarakkudes või üherakulistes organismides. Kokkutõmbuvad vakuoolid on levinud eeskätt magevees elavate lihtorganismide seas, nagu protistid nagu amoeba proteus ja ripsmeline suss, mis sai oma algse nimetuse tänu kehakujule, mis sarnaneb jalatsi talla kujuga. Lisaks loetletud algloomadele leiti identseid struktuure ka erinevate Badyagovi perekonda kuuluvate mageveekäsnade rakkudest.

Kokkutõmbuva vakuooli struktuur. Selle omadused
Kokkutõmbuv vakuool on membraani organell, mis vabastab tsütoplasmast liigse vedeliku. Selle seadme lokaliseerimine ja struktuur on erinevate mikroorganismide lõikes erinev. Vesikulaarsete või torukujuliste vakuoolide kompleksist, mida nimetatakse spongiaks, siseneb vedelik kontraktiilsesse vakuooli. Tänu selle süsteemi pidevale tööle säilitatakse stabiilne rakumaht. Algloomadel on kokkutõmbuvad vakuoolid, mis on aparaat, mis reguleerib osmootset rõhku ja aitab ka jääkaineid organismist väljutada. Algloomade keha koosneb ainult ühest rakust, mis omakorda täidab kõiki vajalikke elutähtsaid funktsioone. Selle alamkuningriigi esindajatel, nagu sussripslane, harilik amööb ja teised üherakulised organismid, on kõik iseseisva organismi omadused.
Lihtorganismide roll
Rakk täidab kõiki elutähtsaid funktsioone: väljutamine, hingamine, ärrituvus, liikumine, paljunemine, ainevahetus. Algloomad on kõikjal. Suurim arv liike elab mere- ja magevetes, paljud elavad niiskes pinnases, võivad nakatada taimi ning elavad mitmerakuliste loomade ja inimeste kehas. Looduses täidavad algloomad sanitaarset rolli, nad osalevad ka aineringes ja on toiduks paljudele loomadele.

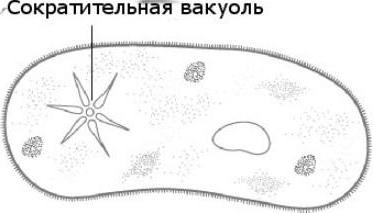
Amoeba vulgaris'e kontraktiilne vakuool
Harilik amööb on risopoodide klassi esindaja, erinevalt teistest esindajatest pole tal püsivat kehakuju. Liikumine toimub pseudopoodide abil. Nüüd selgitame välja, millist funktsiooni täidab kontraktiilne vakuool amööbis. See on osmootse rõhu taseme reguleerimine selle rakus. See võib moodustuda raku mis tahes osas. Välismembraani kaudu siseneb vesi keskkonnast osmootselt. Lahustunud ainete kontsentratsioon amööbarakus on suurem kui keskkonnas. Seega tekib rõhuerinevus algloomarakus ja väljaspool seda. Kokkutõmbuva vakuooli funktsioonid amööbis on omamoodi pumpamisaparaat, mis eemaldab lihtsa organismi rakust liigse vee. Amoeba Proteus võib vabastada kogunenud vedeliku keskkonda kõikjal kehapinnal.
Lisaks osmoregulatsioonile täidab see elus hingamisfunktsiooni, kuna osmoosi tulemusena tarnib sissetulev vesi selles lahustunud hapnikku. Millist funktsiooni kontraktiilne vakuool veel täidab? Samuti täidab see eritusfunktsiooni, nimelt eemaldatakse ainevahetusproduktid koos veega oma keskkonda.

Hingamine, eritumine, osmoregulatsioon ripsisussis
Algloomade keha on kaetud tiheda kestaga, millel on püsiv kuju. ja vetikad, sealhulgas mõned algloomad. Ripslooma keha on keerulisema ehitusega kui amööbil. Sussikambris on kaks kontraktiilset vakuooli, mis asuvad ees ja taga. Selles seadmes on eristatav reservuaar ja mitu väikest torukest. Kokkutõmbuvad vakuoolid paiknevad tänu sellele struktuurile (mikrotuubulitest) pidevalt rakus püsivas kohas.
Selle algloomade esindaja elutegevuses on kontraktiilse vakuooli põhifunktsiooniks osmoregulatsioon, samuti eemaldab see rakust liigse vee, mis osmoosi toimel rakku tungib. Esiteks paisuvad aferentsed kanalid, seejärel pumbatakse neist vesi spetsiaalsesse reservuaari. Mahuti tõmbub kokku, eraldub toitekanalitest ja vesi paiskub pooride kaudu välja. Ripsirakus on kaks kontraktiilset vakuooli, mis omakorda toimivad antifaasis. Kahe sellise seadme töö tõttu on tagatud pidev protsess. Lisaks ringleb vesi pidevalt kontraktiilsete vakuoolide tegevuse tõttu. Need suruvad vaheldumisi kokku ja kontraktsioonide sagedus sõltub ümbritsevast temperatuurist.
Seega on toatemperatuuril (+18 - +20 kraadi Celsiuse järgi) vakuoolide kontraktsioonide sagedus mõningatel andmetel 10-15 sekundit. Ja arvestades, et sussi looduslik elupaik on igasugune seisva veega mageveekogu ja selles laguneva orgaanilise aine olemasolu, muutub selle keskkonna temperatuur sõltuvalt aastaajast mitu kraadi ja seega ka kokkutõmbumise sagedust. võib ulatuda 20-25 sekundini. Tunni jooksul on lihtsa organismi kontraktiilne vakuool võimeline vabastama rakust suures koguses vett. vastavuses selle suurusega. Neisse koguneb toitaineid, seedimata toidujääke, ainevahetuse lõppprodukte ning leidub ka hapnikku ja lämmastikku.
Reovee puhastamine algloomadega
Algloomade mõju aineringele looduses on väga oluline. Reovee ärajuhtimise tõttu reservuaarides paljunevad bakterid suurel hulgal. Selle tulemusena tekivad erinevad lihtsad organismid, kes kasutavad neid baktereid toiduna ja aitavad seega kaasa looduslikule

Järeldus
Vaatamata nende üherakuliste organismide lihtsale ehitusele, mille keha koosneb ühest rakust, kuid täidab terve organismi funktsioone, mis on keskkonnaga hämmastavalt kohanenud. Seda võib täheldada isegi kontraktiilse vakuooli struktuuri näitel. Tänaseks on looduses kõige lihtsamate tohutu tähtsus ja nende osalemine aineringes juba tõestatud.
KONTRAKTIVNE VACUOL, vt VACUOL... Teaduslik ja tehniline entsüklopeediline sõnastik
Kokkutõmbuv vakuool kontraktiilne vakuool. Teatud tüüpi vakuool mõnes protistide rühmas, mis on seotud vee (lahuste) eemaldamisega rakust kokkutõmbumise ajal ja vee imendumisega raku poolt paisumise ajal, mis aitab reguleerida osmootset rõhku.... Molekulaarbioloogia ja geneetika. Sõnastik.
Eukarüootse raku struktuur. Vakuool on loetletud numbriga 10. Vakuool on ühemembraaniline organell, mida leidub mõnes eukarüootses rakus ja ... Wikipedia
Või Heliozoa klassi Sarcodica (q.v.) tüüpi algloomade (q.v.) loomad. Morfoloogilised omadused. Neid eristab sfääriline protoplasmaatiline keha, millest õhukesed, niitjad, mitte... ... ulatuvad igas suunas, nagu kiired. Entsüklopeediline sõnaraamat F.A. Brockhaus ja I.A. Efron
Või Ciliata, ripslaste klassi üksus ehk ripslased (q.v.), algloomade tüüp (q.v.). RIIMELISED SILAADID. I (Aspirotricha). Tähtede tähendus: pulber; ektoplasma alveolaarne kiht; ad.Z ripsmete jumalik rida; b kombatav seta; cl ripsmed; ... Entsüklopeediline sõnaraamat F.A. Brockhaus ja I.A. Efron
- (Lobosea), kõige lihtsamalt organiseeritud algloomade klass, mis ületab risoomide klassi. Ilma sisemisest skelett ja välimine kestad. Keha kuju on muutuv, suurused jäävad tavaliselt vahemikku 20-700 mikronit, harva veidi rohkem. Pseudopoodide kuju ja suurus on iseloomulikud... ... Bioloogia entsüklopeediline sõnastik
- (Flagellata s. Mastigophora, vt tabel. Flagellates, Flagellata) algloomade (Protozoa) klass. Nagu kõigil teistel seda tüüpi esindajatel, on ka neil keha, mis koosneb ainult ühest rakust, mis esindab protoplasmat ja tuuma koos tuumaga.... Entsüklopeediline sõnaraamat F.A. Brockhaus ja I.A. Efron
Või algloomad. Artikli sisu: Tunnused ja klassifikatsioon. Ajalooline sketš. Morfoloogia; protoplasma koos lisanditega (trihhotsüstid, tuum, kontraktiilsed vakuoolid, kromatofoorid jne). Kaaned ja luustik. Liikumine P.; pseudopoodiad, flagellad ja... Entsüklopeediline sõnaraamat F.A. Brockhaus ja I.A. Efron
A. KONTRAKTIIVSED VAKUULID
1. Kokkutõmbuvate vakuoolide morfoloogia
Enamik algloomi (v.a Sporozoa) Neil on spetsiaalne, enam-vähem rangelt lokaliseeritud, mõnikord väga keeruline eritusaparaat, millest enamikul on nn kontraktiilsete ehk pulseerivate vakuoolide iseloom.
Üldreeglina on kõigil mageveeloomadel vakuoolid. Mastigophora Ja Sarcodina ja kõigis ripslastes.
U Mastigophora enamasti on ainult 1 kontraktiilne vakuool, harva 2 (fütomastigina).
Vacuolaaraparaadi mõõtmed on üldiselt otseselt proportsionaalsed looma keha suurusega ja pöördvõrdelised vakuoolide arvuga looma kehas. Niisiis, suurtes ripsloomades Heterotricha (Stentor, Spirostomum) seal on ainult 1 vakuool, kuid see (koos kanalitega) ulatub väga suurte mõõtmeteni. U Trahheeli munarakk kuni 30 vakuooli, kuid need on väga väikesed.
Mõnikord täheldatakse looma kasvades vakuoolide arvu suurenemist, nagu näiteks aastal Collinia branchiarum ja mõned teised Astomata. See vakuoolide arvu korrutis, mis Collinia (anoplophrya) toimub indiviidi elu jooksul, ilmselt toimus kogu rühma evolutsiooni käigus Astomata(joonis 124). Tõepoolest, nende hulgas võib tuvastada mitmeid vorme, millel on järk-järgult keerukam vakuolaarseade. Jah, y Dogielella perekonnas on ainult 1 subterminaalne vakuool Anoplofria 1 pikisuunaline rida väikese arvu vakuoolidega; sünnil Butschliella Ja Mopodontofria vakuoolide arv selles reas suureneb oluliselt ja sisse Radiofria vakuoolid moodustavad 2 pikisuunalist rida. Lõpuks selline Haptofria hulk arvukaid vakuoole ühinevad ühiseks pikisuunaliseks vakuoolikanaliks, mille tekkemeetod ilmneb mitmete eraldi väljalaskeavade olemasolust piki seda.
U Sarcodina Ja Mastigophora kogu eritusaparaat on piiratud vakuooli endaga. Mõnes flagellaadis (nt. Euglena) kontraktiilne vakuool on ümbritsetud väikeste harivate vakuoolide servaga, mis on nendega ühendatud õhukeste torukestega.
Pappas ja Brandt (1958) arvavad, et vesi vakuoolis Amööb siseneb sinna ümberringi lebavate väikeste vesiikulite väljavalamise tulemusena nende vesiikulite membraanide sulandumise teel keskvakuooliga. Ripslastel on kontraktiilsed vakuoolid tavaliselt varustatud ühe või mitme pulseeriva aferentse kanaliga. U Spirostomum - 1 selline kanal, Stentor- 2, a Frontonia- IO pikad kanalid jne Kanalid lahknevad ilmselt tsütoplasmas palju suuremale kaugusele, kui mikroskoobi all näha. Vähemalt osmoosi ajal Parametsium märgatavaks muutub kanalite perifeersete otste jätkumine pikkadeks õhukesteks otsteks. Haridusvakuoolid ja aferentsed kanalid koguvad tsütoplasmast vedelikku (st peamiselt vett) ja toimetavad selle kontraktiilsesse vakuooli. Kogu ripslaste vakuolaaraparaat asub sügaval endoplasmas ega muuda oma asendit kehas.
Hiljuti avaldas Schneider (1960) huvitavaid andmeid kontraktiilsete vakuoolide ja adduktorkanalite ultramikroskoopilise struktuuri kohta. Paramecium caudatum. D. N. Nasonovi (1924) järgi on adduktorkanalid ümbritsetud osmiofiilse sidemega ja vastavalt
Gelei (Gelei, 1925a, 1925b), käsnjas plasma (Nierenplasma või Nephridialplasma) vaatlused.
Schneideri uuringute kohaselt koosneb see adduktorkanaleid ümbritsev plasmapiirkond paljudest õhukestest ja keerdunud.

Riis. 125. Adduktorkanaliga kontraktiilse vakuooli ja seda ümbritseva tsütoplasma ehituse skeem Parametsiumis elektronmikroskoopia järgi. (Schneideri järgi, 1960). A - radiaalne kanal süstolis, kontraktiilne vakuool diastolis; B - radiaalne kanal diastolis, kontraktiilne vakuool süstolis. AK - vakuooli väljumiskanal; Amp - perifeerse kanali ampulla; EK - kanal, mis ühendab ampulli kontraktiilse vakuooliga; EPR - endoplasmaatilise retikulumi kanalid; FB - vakuooli kontraktiilsed fibrillid; KV - kontraktiilne vakuool; NK - kontraktiilne vakuooli adduktorkanal; NT - endoplasmaatiliste retikulumi kanalitega seotud "nefriidikanalite" võrk; RS - ristlõikes "nefridiaalsete kanalite" klaster.
osmiofiilsete seintega torud (joon. 125). Nende torude sisemine luumen süstooli ajal on 100 Å ja diastoli ajal 150 Å. Kogu nefridiaalse plasma kihi paksus on 1-1,5 μ. Selle perifeerias lähevad torud otse endoplasmaatilise retikulumi tuubulitesse, mis tungivad läbi kogu parametsiumi keha. Lisaks on nefriidiplasma ümbritsetud spetsiaalsete osmiofiilsete seintega torudega. Mõnikord on need paigutatud kimpudesse. Nende läbimõõt ulatub 500 Å-ni. Nende torude pikkus on umbes 1 μ.
Adduktorkanalil on osmiofiilne membraan. Süstooli ajal muutub see kanal pilulaadseks ja selle luumeniga on umbes 300 Å, diastoli korral laieneb see 4000 Å-ni. Adduktorkanalit ümbritsevad tuubulid avanevad sellesse ainult diastooli ajal, kui nende läbimõõt suureneb, samas kui süstooli ajal on need torukesed suletud ega ole kanaliga ühendatud. Proksimaalses otsas olev aferentne kanal läheb otse ampulli, mis avaneb terminaalse kanali kaudu vakuooli reservuaari. Ampulli ja terminali kanali seinas on torukujulised kontraktiilsed fibrillid, mis on paigutatud mitmesse kimpu. Terminali kanali seintelt lähevad need otse kontraktiilse vakuooli reservuaari seinale, jaotudes seal ka eraldi kimpudena.
Rudzinska (1957) järgi vakuooli ümbritsevas tsütoplasmas Tokophrya infusioon Väikesed ergastoplasma mullid kontsentreeritakse ja mitokondrid kogunevad. Lisaks leidub vakuooli ümbert struktuure, mida saab võrrelda diktüosoomidega. Tundub, et nad osalevad tsütoplasmast tuleva vedeliku sekretsioonis ja kogunemises.

A. A. Strelkov (1939) leitud väljaheidete kanali ümbert kl Tsüklopostium selgelt nähtav rõngakujuline kiud, mis on suure tõenäosusega müoneem, mis reguleerib vedeliku väljumist kanali kaudu väljapoole. Kõigil ripsloomadel on kontraktiilse vakuooli eritusava eelnevalt moodustunud poorid. Tavaliselt on see ümmargune või ovaalne ja sellel on iga liigi ripsmete ridade seas täiesti konstantne asukoht. Perekonnas Conchophthirus auk näeb välja nagu kitsas paksendatud servaga pilu. Võib-olla on see pooride kuju Conchophthirus on selle sulgemisel teadaolevalt funktsionaalne tähendus. Nimelt on väga tõenäoline, et siin ei toimu sulgumine mitte väljalaskekanali rõngakujulise ahenemise, vaid pilu servade kokkuvarisemise kaudu nagu kaks huulet. Poor avaneb erituskanalisse, tavaliselt väga lühike, kuid mõnikord (kell Paramecium trichium) moodustades mitu silmusetaolist paindet.
Kokkutõmbuva vakuooli elektronmikroskoopiline uurimine Tokophrya infusioon kinnitas püsivalt eksisteeriva poori ja erituskanali olemasolu, mille ümber paiknevad fibrillid (180 A paksus), mis seda kokku suruvad (Rudzinska, 1957). Sarnased fibrillid leiti aastal Parametsium(Schneider, 1960).
2. Kokkutõmbuvate vakuoolide funktsionaalne tähtsus
1) Pulseerivate vakuoolide kokkutõmbumise protsess
Vakuoolid töötavad rütmiliselt, vaheldumisi laienedes (diastooli staadium) ja täitudes aeglaselt vedelikuga, seejärel järsult kokku tõmbudes (süstooli staadium) ja surudes oma sisu läbi erituskanali välja. Seal, kus on spetsiaalsed adduktorkanalid, muutub kogu aparaadi kokkutõmbumistsükkel keerulisemaks; nimelt algab see kanalite laienemisega, mis tekib siis, kui vakuool on süstoli seisundis (kanalidiastool); sellele järgneb kanalite kokkutõmbumine (süstool), nende sisu valamine kontraktiilsesse vakuooli, mis põhjustab selle süstoli; samal ajal hakkavad kanalid uuesti laienema. Tegelikult on vastavad staadiumid olemas ka vakuoolides, millel puudub kanalisüsteem, kuid seal on neid vähe või mitte näha, kuna vakuooli diastooli ajal valatakse sellesse ümbritsevas plasmas paiknevad väga väikesed ja ebakorrapärase asukohaga vakuoolid (vt . Ophryoscolecidae, riis. 126).
Pulsatsiooni kiirus on erinevate algloomade lõikes erinev ja muutub ka sama liigi vahel keskkonnategurite muutuste mõjul.
Niisiis, erinevate magevee jaoks Algloomad pulsatsioonide vahelised intervallid erinevatel temperatuuridel on näidatud tabelis. 6.
TABEL 6
Muutused kontraktiilse vakuooli pulsatsiooni kiiruses ripsloomadel(sekundis) erinevatel temperatuuridel

Oligochaeta. Selgus, et kontraktiilse vakuooli pulsatsiooni kiirus ei ole kõigil ühesugune Astomata ja on seotud omaniku elukeskkonna osmootsete tingimustega. U Mesnilella clavataw. Hoplitophrya secans mageveeoligoheetide seas on pulseerimine sama sage kui vabalt elavatel mageveeripslastel: pulsatsiooni sagedus ei ületa 1 minutit. (20-30 sekundit).
U Radiofria mereoligoheedest Enchytraeus sp., kes elab Barentsi mere rannikuvööndis, toimub pulsatsioon iga 6-8 minuti järel.
Lõpuks kl Metaradiophrya lumbrici Ja Anoplophrya lumbrici mulla oligohaete soolestikust Eisenia foetida ja kell Mesnilella fastigata Ja Radiophrya prolijera mullast Enchytraeus sp. pulsatsioonisagedus on aeglasem (1,5-4 min) kui mageveeripslastel ja mõnevõrra kiirem kui Astomata mere oligoheetidest. Seega lähisugulasliikidel ripsloomadel Astomata ilmneb oluline füsioloogiline erinevus osmootse keskkonnaga kohanemise tõttu. Mageveeussidest pärit ripslaste puhul on isotooniliseks keskkonnaks 0,4% rõngastaja, mullausside soolestiku ripslaste puhul vastab isotoonilisus 0,75–0,8% ringeri lahusele ja merevormide puhul 3% lahusele.
Keskkonna soolade hulga muutuste mõju vakuoolide pulsatsiooni kiirusele näitas Zuelzer (1910). Ta näitas seda Amoeba verrucosa merevees eluga harjudes näitab see, alustades soolakontsentratsioonist 0,3%, vakuooli pulsatsiooni järkjärgulist aeglustumist ja kui soola kontsentratsioon jõuab 1,5% -ni, vakuool kaob täielikult, alles siis, kui kultuur ilmub uuesti järk-järgult lahjendatakse värske veega. Samamoodi sai Herfs (1922) eest parametsium, NaGl lahustes ja magevees eluga harjunud, järgmised andmed:
| Soolasisaldus vees (%) | 0 | 0.25 | 0.5 | 0.75 | 1 |
| Pulsatsioonide vaheline intervall (sek.) | 6.2 | 9.3 | 18.4 | 24.8 | 163 |
| 1 tunni jooksul eritunud vedeliku kogus, väljendatuna kehamahtudes | 4.8 | 2.82 | 1.38 | 1.08 | 0.16 |
Üldiselt on vaakumite kaudu teatud aja jooksul eritunud vedeliku kogus, kui muud näitajad on võrdsed, võrdeline pulsatsiooni kiirusega ja erinevatel juhtudel. Algloomad võib olla üsna erinev. Näiteks kontraktiilne vakuool Uronema nigricans temperatuuril 28° 2 minutit. vabastab ripslooma keha mahuga võrdse veemahu ja Euplotes patella selleks kulub 14 minutit, Paramecium aurelia- 46 min.
2) Kokkutõmbuvate vakuoolide füsioloogiline tähtsus
Vakuoolide funktsioonid on ilmselt mitmekesised. Esiteks on põhjust arvata, et vakuoolid vastutavad sellesse kogunevate lõpp-ainevahetusproduktide organismist väljaviimise eest.
Ettepanekuid selle funktsiooni kohta tegid eelmisel sajandil Stein (1878) ja teised teadlased, kuid Howland (1924a, 1924b) oli esimene, kes näitas kaudselt, et kusihapet eritavad algloomad. Tema parametsiumi ja amööbide kultuurid sisaldasid kusihapet, mille kontsentratsioon suurenes ligikaudu proportsionaalselt kultuuri hoidmise ajaga. M. Novikov (1908), Shumway (1917) ja Flather (1919) näitasid, et kilpnäärme ripsmete toitmisel epinefriini ja käbinäärme ekstrakti toimel vakuoolide pulseerimine sageneb ja vakuoolid ise suurenevad, st stimulatsiooni saadakse eritumine samade ravimite mõjul, mis põhjustavad selgroogsetel diureesi.
Weatherby (1927) avastas kontraktiilse vakuooli vedelikust uurea Spirostomum, ekstraheeritakse mikropipeti abil. Kuid selle autori arvutuste põhjal eemaldab vakuool ainult 1% massikultuuride poolt eritatavast koguhulgast. Spirostomum uurea.
Üldiselt on andmed lämmastiku eritumise produktide olemuse kohta väga vastuolulised. Lisaks kusihappele ja uureale märkisid mõned autorid nende ainete asemel eritumisproduktidena ammoniaaki (näiteks Glaukoom Ja Spirostomum). Ainus asjaolu, mis kontraktiilsete vakuoolide eritusfunktsiooniga täielikult ei nõustu, on see, et nad ei osale kunagi plasmasse viidud intravitaalsete värvainete eritumises, mis erituvad väga sageli kõrgematel loomadel neerude ja teiste eritusorganite kaudu.
Korduvalt on avaldatud arvamust, et kokkutõmbuvad vakuoolid võtavad teatud osa algloomade hingamisteede ainevahetuses, hõlbustades süsihappe eemaldamist tsütoplasmast; siiski puuduvad tegelikud tõendid vakuooli spetsiifilise aktiivsuse kohta selles suunas. Ainult Ludwigi (Ludwig, 1928) kaudseid andmeid vakuooli kaudu eritunud süsihappe tegeliku koguse ja loomade poolt eritatava hinnangulise süsihappe koguse vahelise vastavuse kohta tõlgendab see autor vakuooli hingamisfunktsiooni kasuks.
Enamik autoreid rõhutab praegu kontraktiilsete vakuoolide olulist rolli osmoregulatoorse aparaadina. Isegi Degen (1905) tõi välja, et magevee algloomade plasmal on selles sisalduvate soolade tõttu kõrgem osmootne rõhk kui keskkonnas; ja see asjaolu toob kaasa vee pideva imendumise tsütoplasmas. Kokkutõmbuva vakuooli eesmärk on eemaldada plasmasse pidevalt sisenev liigne vesi. Selle seisukoha õigsust kinnitavad nii kontraktiilsete vakuoolide jaotumise olemus algloomade vahel kui ka katsed magevee algloomade aklimatiseerimisel kasvava soolakontsentratsiooniga veega (Firiley, 1930; Loefer, 1939 jt).
Hiljuti tehti eksperimentaalne uuring Jl. H. Seravini (1958) vakuooli pulsatsiooni üle Paramecium caidatum, mis annab andmeid, mis näitavad, et kui ripsloomad viiakse üle kõrgendatud kontsentratsiooniga soolalahustesse, siis esimese 15-30 minuti jooksul. Vakuooli pulsatsioonikiirus väheneb mitu korda, seejärel pulsatsiooni kiirus järk-järgult kiireneb ja läheneb kontrollripsmetele iseloomulikule kiirusele, kuid tavaliselt ei saavuta selle taset. Näiteks 0,2% NaCl-s 30 minuti pärast. Pulsatsiooni sagedus oli keskmiselt 40,5 sekundit ja 48 tunni pärast. saavutas 13,1 sek., samas kui kontrollrühmas kulges pulsatsioon kiirusega 10,3 sek. Sama täheldati 0,1% LiCi, 0,5% CaCl2, 1% sahharoosi ja uurea lahuse lahustes. Aeglustumine toimus 7-15 korda, kuid 2 päeva pärast
pulsatsioonikiirus nendes lahustes aeglustus vaid 1,5-3 korda. Madalamate soolakontsentratsioonide korral täheldati vakuooli pulsatsioonikiiruse täielikku taastumist. Sellised ained nagu HGl, NaOH, GaGl 2, formaliin põhjustasid tühistes kontsentratsioonides pulsatsiooni kerget aeglustumist, mis seejärel kiiresti normaliseerus. Adrenaliin kontsentratsioonis 0,0005% isegi suurendas veevahetust, kuid aja jooksul normaliseerus ka pulsatsioon. Seega põhjustavad osmootselt aktiivsed ja mitteaktiivsed ained kontraktiilse vakuooli aktiivsuses sarnaseid muutusi. Sel juhul on üldiselt kalduvus reguleerida ripsmeke tsütoplasmat läbivat veevoolu, muutes vakuooli pulsatsiooni kiirust. Lisaks kohanevad ripsloomad väliskeskkonna osmootse rõhu muutuvate tingimustega. Seda märkis ka Kitching (1952) seoses Archesium Ja Podophrya. Nendel ripsloomadel lakkab etüleenglükooli ja sahharoosi kõrge kontsentratsiooni korral vakuooli aktiivsus täielikult, kuid mõne aja pärast vakuooli pulseerimine ja veevahetus taastuvad.
Kõik see viitab kontraktiilse vakuooli keerulisele toimemehhanismile, millel on ripslaste veevahetuses kahtlemata roll. Seravin usub aga, et lähtudes tõsiasjast, et osmootselt inaktiivsed ained põhjustavad vakuoolide pulsatsiooni viivitusi, võib eeldada, et veevahetus ripsloomadel ei ole osmootsete mustritega määratud ja seetõttu ei saa kontraktiilse vakuooli rolli osmoregulatsioonis pidada tõestatuks. .
Vakuooli funktsiooni mõistmiseks on loomulikult vaja mõista ka vee tsirkulatsiooni mehhanismi tsütoplasmast vakuooli reservuaari ja vakuooli enda kokkutõmbumise mehhanismi. I. Gelei ja G. Gelei (Gelei, 1928; Gelei, 1939), nagu ka mitmed teised uurijad, usuvad, et ainult tsütoplasmas tekkiv turgori pinge ja sellest tulenevalt ka osmootsed tingimused määravad vakuoolide reservuaari vähenemise. Võimalik, et on olemas ka aktiivne mehhanism vee eritumiseks vakuooli (Kitching, 1956). Schneideri (Schneider, 1950) järgi põhjustab vakuooli pulseerimist vakuooli seinas paiknevate kontraktiilsete fibrillide tegevus ning aferentsete kanalite täitumine veega toimub järgmiselt. Vedelik koguneb ergastoplasmaatilistesse tuubulitesse ja seejärel, kuna need torukesed on suurema läbimõõduga kui nefriidi plasmatorukesed, tekib ergastoplasma tuubulitest veevool nefriidi plasmatuubulitesse ja neist kanali luumenisse. Loomulikult on nende protsesside mehhanismi täpsemaks mõistmiseks vaja täiendavaid uuringuid. Kokkutõmbuva vakuooli eritus- ja hingamisaktiivsus näib olevat väga võimalik, kuid mitte lõplikult tõestatud.
3) kontraktiilne vakuool ja Golgi aparaat
Küsimus kontraktiilse vakuooli morfoloogilisest tähendusest jõudis uude faasi pärast D. N. Nasonovi (1924, 1925) tööd, kes väljendas ideed, et algloomade kontraktiilne vakuool on homoloogne rakkudes oleva Golgi aparaadiga. Metazoa. Nasonov (1924) ja peaaegu samaaegselt temaga I. Gelei (J. Gelei, 1925a, 1925b) avastasid ripsloomadele osmatsioonitehnikat rakendades olulisi, uusi jooni kontraktiilsete vakuoolide struktuuris (joon. 127). Nasonov leidis, et paljude ripsloomade kontraktiilse vakuooli ümber (Campanella, Lionotus, Parameciuin, Nassula), sama hästi kui Chilomonas poolt tugevalt mustaks
vähendatud osmiumiga kest, sileda pinnaga või ümbritsetud väikeste osmiofiilsete terade kihiga. Kest kas ümbritseb tervet vakuooli (ülaltoodud liikidel) või on võrkrõnga kujul, mis ümbritseb vakuooli piki selle ekvaatorit. (Chilodon, Dogielella). Adduktorkanalite süsteemiga vormides (parametsium), kest jätkub viimasele; sel juhul on kanalid endoplasmas palju kaugemal kui ühegi teise uurimismeetodiga jälgitavad ning nende distaalsed osad on ümbritsetud halli võrkhülsiga. Nasonov peab seda kanalite lõiku aparaadi tegelikuks ekskretoorseks osaks, samas kui kanalite proksimaalne osa ja vakuool ise on vaid vakuolaarsüsteemi eritusosa. U Dogielella Nasonov võiks väita, et vakuooli süstooli ajal ei järgne seda ümbritsev must rõngas selle kokkutõmbumise ajal viimasele, vaid jääb oma algsele kohale. Gelei (1925a, 1925b) püüdis neid võrrelda nefriidisüsteemiga metazoa, võrdsustada kontraktiilne vakuool ise põiega ja aparaadi muud osad nefriidi teiste osadega. aastal keskendus Nasonov vakuoolide osmootse membraani sarnasusele Golgi aparaadiga. Metazoa. Tõepoolest, tema esitatud joonised erinevat tüüpi kontraktiilsetest vakuoolidest ja Golgi aparaatidest, mis on võetud erinevat tüüpi rakkudest metazoa,äärmiselt veenvad oma sarnasuses.

Sellest lähtuvalt peab Nasonov võimalikuks otsida osmiofiilsest käsnjas membraanist adduktorkanaleid Parameciurn ja võrkrõngas Dogitlella mitte ainult morfoloogiline, vaid ka funktsionaalne paralleelsus nende moodustiste ja Golgi aparaadi vahel. Rõngasaine võrgustruktuur sõltub väikeste hariduslike vakuoolide olemasolust selles - seotud sekretsiooni staadiumis. Need vakuoolid ühinevad seejärel tsentraalse vakuooliga, mis asub rõnga keskel - väljapoole sekreteeritud vaba sekretsiooni staadiumis. Nasonovi kontseptsiooni kohaselt kogub Golgi aparaat plasmast teadaolevaid aineid, mis valmistatakse selles ette nende eemaldamiseks kehast vaakumi kaudu. Nasonovi leidude kogu faktilist külge kinnitasid täielikult mitmete autorite hilisemad uuringud: A. A. Strelkov (1939) Cycloposlhiidae Fauré-Fremiet, 1925) Vorlicella, Kuningas (1928) Parametsium trihium Ja Euplotes jne. Tõsi, mõne alglooma puhul ei olnud võimalik tuvastada vakuooli ümbert erilist osmiofiilset membraani, mis võis sõltuda metoodilistest küsimustest.
igaühe rakkudes Metazoa võrgusilma moodustumise kujul või koosnedes üksikutest diktüosoomidest. Seetõttu võime arvata, et seda leidub ka kõigis ainuraksetes organismides. Tõepoolest, Hirschleri (1927), Hovasse (1937), Joyet-Lavererie (1926) jt uuringud näitasid paljude algloomade esinemist plasmas. (Gregarina, Coccidia, Flagellata, Sarcodina) väikesed vardad, rõngad, tugevalt osmiofiilse servaga kuulid ja diktüosoomidega väga sarnased rakud Metazoa.
Golgi aparaadi märgid Metazoa Esiteks on see seade võimeline spetsiifiliselt redutseerima teatud raskmetallide ühendeid - hõbedat ja osmiumi, mis väljendub seadme struktuuride mustaks muutumises. Neid struktuure ei värvita intravitaalselt neutraalse punase ja metüleensinisega. Sageli on Golgi aparaadi iseloomulikud struktuurid diktüosoomid, mis koosnevad kahest osast – kromofiilsest ja kromofoobsest. Tavaliselt sukeldatakse sekretoorsed graanulid kromofoobiosasse. Reeglina paikneb Golgi aparaat täpselt raku sekretoorses tsoonis.
Elektronmikroskoobi uuringud on näidanud Golgi aparaadi struktuuride ühtsust erinevates rakkudes Metazoa. Kõigis uuritud rakkudes tuvastati topeltmembraanid (ehk γ-tsütomembraanid), mis paiknesid üksteisega paralleelselt ja moodustasid 3-7 või enamast paarist koosneva süsteemi. Iga paar ümbritseb 50–200 Å laiust kergemat ruumi ja moodustab omamoodi lameda koti. Mõnikord laieneb see suureks vakuooliks. Membraanid on sukeldatud peeneteralise alusmaterjali sisse. Mõnel juhul võivad membraanid ümbritseda erineva suurusega vakuoole. Kõik need struktuurid on üsna stabiilsed.
On märkimisväärne, et Nasonovi ja teiste rakkude uurijate poolt valgusmikroskoobiga kirjeldatud Golgi aparaadi piltide vahel on täielik vastavus. metazoa, ja elektronmikroskoobiga saadud kaasaegsed kujutised.
Dalton ja Felix (1957) märgivad samuti, et kontraktiilne vakuool Chlamidomonas mida piiravad Golgi rakuaparaadi membraanidega sarnased membraanid metazoa, ja seega toetavad autorid Nasonovi hüpoteesi nende organellide homoloogia kohta. Kokkutõmbuva vakuooli ümber ripsloomadel Campanella Ja Ophridium on arvukalt tihedate osmiofiilsete seintega torusid, mis võivad olla mingil määral võrreldavad Golgi aparaadi ultrastruktuuridega (Fauré-Fremiet et Rouiller, 1959). Ülaltoodud Schneideri andmed nefridiaalse plasma struktuuri kohta kontraktiilse vakuooli adduktorkanalite ümber Parametsiiim raskendab selle struktuuri homologiseerimist diktüosoomide ultrastruktuuriga.
Veel varem lükkasid prantsuse autorid (Duboscq et Grassé, 1933; Grassé et Hollande, 1941) tagasi ka kontraktiilsete vakuoolide osmiofiilsete membraanide homoloogia Golgi aparaadiga. Nad usuvad, et vakuoolide osmiofiilsed seinad, nagu ka kontraktiilsed vakuoolid ise, on Golgi aparaadist täiesti sõltumatud moodustised.
Tuleb märkida, et mõnedel ripsloomadel leiti sirbi- või rõngakujulisi diktüosoome, näiteks Balantidium elongatum,ühendusest väljas, vakuoolidega (Villeneuve-Brachori, 1940). Noirot-Timothee (1957) tuvastas elektronmikroskoobiga Golgi aparaadi tüüpilised struktuurid mõne inimese endoplasmas. Ophryoscolecidae, ei ole seotud kontraktiilse vakuooliga.
Prantsuse autorid homologiseerivad parabasaalkeha Golgi aparaadiga (vt lk 115), kuna see moodustis, eriti elektronmikroskoobiga uurides, osutus sarnaseks rakkude diktüosoomidega. Metazoa(joonis 80).
Kahtlemata on vaja täiendavaid algloomade võrdlevaid uuringuid, et lahendada teatud struktuuride homoloogia küsimus rakkude Golgi aparaadiga. Metazoa.