Type, dens kriterier. En population er en strukturel enhed af en art og en elementær evolutionsenhed. Mikroevolution. Dannelse af nye arter. Metoder til artsdannelse. Bevarelse af artsdiversitet som grundlag for biosfærens bæredygtighed
Type, dens kriterier
Grundlæggeren af moderne taksonomi, C. Linnaeus, betragtede en art som en gruppe af organismer, der ligner morfologiske egenskaber, og som frit formerer sig. Efterhånden som biologien udviklede sig, blev der opnået beviser for, at forskellene mellem arter er meget dybere og påvirker den kemiske sammensætning og koncentration af stoffer i væv, retningen og hastigheden af kemiske reaktioner, arten og intensiteten af vitale processer, antallet og formen af kromosomer, dvs. arten er den mindste gruppe af organismer, hvilket afspejler deres tætte slægtskab. Derudover eksisterer arter ikke for evigt – de opstår, udvikler sig, giver anledning til nye arter og forsvinder.
Udsigt- dette er en samling af individer, der ligner hinanden i struktur og karakteristika af livsprocesser, har en fælles oprindelse, krydser frit med hinanden i naturen og producerer frugtbart afkom.
Alle individer af samme art har den samme karyotype og indtager et bestemt geografisk område i naturen - areal
Tegn på lighed mellem individer af samme art kaldes type kriterier. Da ingen af kriterierne er absolutte, er det nødvendigt at bruge et sæt kriterier for at bestemme typen korrekt.
De vigtigste kriterier for en art er morfologiske, fysiologiske, biokemiske, økologiske, geografiske, etologiske (adfærdsmæssige) og genetiske.
- Morfologisk- et sæt ydre og indre karakteristika for organismer af samme art. Selvom nogle arter har unikke karakterer, er det ofte meget vanskeligt at skelne nært beslægtede arter alene ved hjælp af morfologiske træk. Således er der for nylig blevet opdaget en række tvillingearter, der lever i samme territorium, for eksempel husmusen og Kurganchik-musen, så det er uacceptabelt udelukkende at bruge morfologiske kriterier til at bestemme arten.
- Fysiologisk- ligheden mellem livsprocesser i organismer, primært reproduktion. Det er heller ikke universelt, da nogle arter krydser hinanden i naturen og producerer frugtbart afkom.
- Biokemisk- lighed mellem kemisk sammensætning og metaboliske processer. På trods af at disse indikatorer kan variere betydeligt blandt forskellige individer af samme art, får de i øjeblikket stor opmærksomhed, da de strukturelle træk og sammensætning af biopolymerer hjælper med at identificere arter selv på molekylært niveau og fastslå graden af deres forhold.
- Økologisk- skelnen mellem arter efter deres tilhørsforhold til visse økosystemer og økologiske nicher, som de besætter. Imidlertid optager mange ubeslægtede arter lignende økologiske nicher, så dette kriterium kan kun bruges til at skelne en art i kombination med andre egenskaber.
- Geografisk- eksistensen af en bestand af hver art i en bestemt del af biosfæren - et område, der adskiller sig fra områderne for alle andre arter. På grund af det faktum, at grænserne for deres udbredelsesområder for mange arter er sammenfaldende, og der også er en række kosmopolitiske arter, hvis udbredelse dækker store rum, kan det geografiske kriterium heller ikke tjene som et markør-"arts"-træk.
- Genetisk- konstans af kromosomsættets karakteristika - karyotype - og nukleotidsammensætningen af DNA hos individer af samme art. På grund af det faktum, at ikke-homologe kromosomer ikke kan konjugere under meiose, vises afkom fra krydsning af individer af forskellige arter med et ulige sæt kromosomer enten slet ikke eller er ikke frugtbare. Dette skaber reproduktiv isolation af arten, bevarer dens integritet og sikrer, at den eksisterer i naturen. Denne regel kan blive overtrådt i tilfælde af krydsning af arter af lignende oprindelse med samme karyotype eller forekomst af forskellige mutationer, men undtagelsen bekræfter kun den generelle regel, og arter bør betragtes som stabile genetiske systemer. Det genetiske kriterium er det vigtigste i systemet af artskriterier, men heller ikke udtømmende.
På trods af kompleksiteten af kriteriesystemet kan en art ikke repræsenteres som en gruppe af organismer, der er absolut identiske i alle henseender, det vil sige kloner. Tværtimod er mange arter kendetegnet ved en betydelig diversitet selv i ydre karakteristika, da for eksempel nogle bestande af mariehøns er kendetegnet ved en overvægt af rød farve, mens andre er kendetegnet ved en overvægt af sort.
Population er en strukturel enhed af en art og en elementær enhed af evolution
Det er svært at forestille sig, at individer af én art i virkeligheden ville være jævnt fordelt over jordens overflade inden for området, da søfrøen f.eks. hovedsageligt lever i ret sjældne stående ferskvandsforekomster, og det er usandsynligt, at den findes. i marker og skove. Arter i naturen falder oftest i separate grupper, afhængigt af kombinationen af betingelser, der er egnede til deres levesteder - populationer.
Befolkning- en gruppe individer af samme art, der optager en del af dens udbredelsesområde, frit blander sig med hinanden og relativt isoleret fra andre grupper af individer af samme art i mere eller mindre lang tid.
Populationer kan adskilles ikke kun rumligt; de kan endda leve i det samme territorium, men har forskelle i fødevarepræferencer, tidspunkt for reproduktion osv.
En art er således en samling af populationer af individer, der har en række fælles morfologiske, fysiologiske, biokemiske karakteristika og typer af forhold til miljøet, som bor i et bestemt område og også er i stand til at krydse hinanden for at danne frugtbart afkom, men næsten eller slet ikke krydsning med andre grupper individer af samme art.
Inden for arter med store udbredelsesområder, der dækker territorier med forskellige livsbetingelser, skelnes de nogle gange underarter- store populationer eller grupper af nabopopulationer, der har vedvarende morfologiske forskelle fra andre populationer.
Populationer er ikke spredt over jordens overflade tilfældigt; de er bundet til bestemte områder. Helheden af alle faktorer af livløs natur, der er nødvendige for opholdet af individer af en given art, kaldes levested. Disse faktorer alene er dog muligvis ikke nok til, at en befolkning kan besætte dette område, da den stadig skal være involveret i tæt samspil med populationer af andre arter, dvs. indtage en bestemt plads i samfundet af levende organismer - økologisk niche. Den australske koala-pungbjørn kan således alt andet lige ikke eksistere uden sin vigtigste fødekilde - eukalyptus.
Populationer af forskellige arter, der danner en uløselig enhed i de samme levesteder, giver normalt et mere eller mindre lukket kredsløb af stoffer og er elementære økologiske systemer (økosystemer) - biogeocenoser.
På trods af alle deres krav til miljøforhold er populationer af samme art heterogene i areal, antal, tæthed og rumlig fordeling af individer, og danner ofte mindre grupper (familier, flokke, besætninger osv.), køn, alder, genpulje osv. Derfor skelnes der mellem deres størrelse, alder, køn, rumlige, genetiske, etologiske og andre strukturer, såvel som dynamikker.
Vigtige egenskaber ved en befolkning er genpulje- et sæt gener, der er karakteristiske for individer af en given population eller art, samt hyppigheden af visse alleler og genotyper. Forskellige populationer af samme art har i starten forskellige genpuljer, da nye territorier koloniseres af individer med tilfældige snarere end specielt udvalgte gener. Under påvirkning af interne og eksterne faktorer gennemgår genpuljen endnu mere væsentlige ændringer: den beriges på grund af forekomsten af mutationer og en ny kombination af egenskaber og udtømmes som følge af tabet af individuelle alleler under døden eller migrationen af et vist antal individer.
Nye egenskaber og deres kombinationer kan være gavnlige, neutrale eller skadelige, derfor er det kun individer, der er tilpasset givne miljøforhold, der overlever og formerer sig med succes i befolkningen. Men på to forskellige punkter på jordens overflade er miljøforholdene aldrig helt identiske, derfor kan retningen af ændringer selv i to nabopopulationer være helt modsat, eller de vil forekomme med forskellige hastigheder. Resultatet af ændringer i genpuljen er divergensen af populationer i henhold til morfologiske, fysiologiske, biokemiske og andre karakteristika. Hvis bestande også er isoleret fra hinanden, så kan de give anledning til nye arter.
Således fører fremkomsten af eventuelle forhindringer i krydsningen af individer af forskellige populationer af samme art, for eksempel på grund af dannelsen af bjergkæder, ændringer i flodsenge, forskelle i tidspunktet for reproduktion osv., til det faktum at populationer gradvist får flere og flere forskelle og til sidst bliver til forskellige arter. I nogen tid, ved grænserne af disse populationer, sker krydsning af individer, og hybrider opstår, men over tid forsvinder disse kontakter, dvs. populationer fra åbne genetiske systemer bliver lukkede.
På trods af at individer primært er udsat for miljøfaktorer, er ændringer i den genetiske sammensætning af en enkelt organisme ubetydelige og vil i bedste fald kun optræde i dens efterkommere. Underarter, arter og større taxa er heller ikke egnede til rollen som elementære evolutionsenheder, da de ikke adskiller sig i morfologisk, fysiologisk, biokemisk, økologisk, geografisk og genetisk enhed, mens populationer er de mindste strukturelle enheder af en art, der akkumulerer en række tilfældige ændringer, hvoraf de værste vil blive elimineret, svarer til denne tilstand og er elementære evolutionsenheder.
Mikroevolution
Ændring af populationernes genetiske struktur fører ikke altid til dannelsen af en ny art, men kan kun forbedre populationens tilpasning til specifikke miljøforhold; dog er arter ikke evige og uforanderlige - de er i stand til at udvikle sig. Denne proces med irreversibel historisk forandring i levende ting kaldes evolution. Primære evolutionære transformationer forekommer inden for en art på populationsniveau. De er først og fremmest baseret på mutationsprocessen og naturlig udvælgelse, hvilket fører til ændringer i genpuljen af populationer og arten som helhed, eller endda til dannelsen af nye arter. Sættet af disse elementære evolutionære begivenheder kaldes mikroevolution.
Populationer er karakteriseret ved enorm genetisk diversitet, som ofte ikke udtrykkes fænotypisk. Genetisk diversitet opstår på grund af spontan mutagenese, som sker kontinuerligt. De fleste mutationer er ugunstige for organismen og reducerer levedygtigheden af befolkningen som helhed, men hvis de er recessive, kan de forblive i heterozygoter i lang tid. Nogle mutationer, der ikke har adaptiv værdi under givne eksistensbetingelser, er i stand til at erhverve en sådan værdi i fremtiden, eller når nye økologiske nicher udvikles, og dermed skabe en reserve af arvelig variabilitet.
Mikroevolutionære processer er væsentligt påvirket af fluktuationer i antallet af individer i populationer, migration og katastrofer, samt isolation af populationer og arter.
En ny art er et mellemresultat af evolutionen, men på ingen måde dens resultat, da mikroevolutionen ikke stopper der – den fortsætter videre. Nye arter, der opstår, i tilfælde af en vellykket kombination af egenskaber, befolker nye levesteder og giver på sin side anledning til nye arter. Sådanne grupper af nært beslægtede arter er forenet i slægter, familier osv. Evolutionære processer, der forekommer i supraspecifikke grupper, kaldes allerede makroevolution. I modsætning til makroevolution, mikroevolution finder sted i en meget kortere periode, mens den første kræver titusindvis og hundredtusinder og millioner af år, såsom menneskelig evolution.
Som et resultat af mikroevolution dannes hele mangfoldigheden af arter af levende organismer, der nogensinde har eksisteret og nu lever på Jorden.
Samtidig er evolutionen irreversibel, og arter, der allerede er forsvundet, opstår aldrig igen. Nye arter konsoliderer alt, hvad der er opnået i evolutionsprocessen, men dette garanterer ikke, at der i fremtiden ikke vil dukke nye arter op, som vil have mere avancerede tilpasninger til miljøforhold.
Dannelse af nye arter
I bred forstand forstås dannelsen af nye arter ikke kun som afspaltning af en ny art fra hovedstammen eller opløsning af forældrearten i flere datterarter, men også artens generelle udvikling som et integreret system , hvilket fører til betydelige ændringer i dens morfostrukturelle organisation. Dog oftere end ikke art betragtes som en proces til dannelse af nye arter gennem forgrening af artens "slægtstræ".
En grundlæggende løsning på problemet med artsdannelse blev foreslået af Charles Darwin. Ifølge hans teori fører spredningen af individer af samme art til dannelsen af populationer, der på grund af forskelle i miljøforhold er tvunget til at tilpasse sig dem. Dette medfører igen en intensivering af den intraspecifikke kamp for tilværelsen, styret af naturlig selektion. I øjeblikket mener man, at kampen for tilværelsen slet ikke er en obligatorisk faktor i artsdannelse, tværtimod kan selektionspresset i en række populationer falde. Forskelle i levevilkår bidrager til fremkomsten af ulige tilpasningsændringer i populationer af en art, hvis konsekvens er en divergens af populationers karakteristika og egenskaber - divergens.
Imidlertid er akkumuleringen af forskelle, selv på det genetiske niveau, på ingen måde tilstrækkelig til fremkomsten af en ny art. Så længe populationer, der adskiller sig i nogle karakteristika, ikke kun er i kontakt, men også er i stand til at krydse sig med dannelsen af frugtbart afkom, tilhører de den samme art. Kun umuligheden af strømmen af gener fra en gruppe af individer til en anden, selv i tilfælde af ødelæggelse af de barrierer, der adskiller dem, dvs. krydsning, betyder færdiggørelsen af den mest komplekse evolutionære proces med dannelsen af en ny art.
Speciation er en fortsættelse af mikroevolutionære processer. Der er et synspunkt om, at artsdannelse ikke kan reduceres til mikroevolution; det repræsenterer et kvalitativt udviklingsstadium og udføres takket være andre mekanismer.
Metoder til artsdannelse
Der er to hovedmetoder til artsdannelse: allopatrisk og sympatrisk.
Allopatrisk, eller geografisk art er en konsekvens af den rumlige adskillelse af befolkninger ved fysiske barrierer (bjergkæder, have og floder) på grund af deres opståen eller spredning til nye levesteder (geografisk isolation). Da genpuljen i den adskilte befolkning i dette tilfælde adskiller sig væsentligt fra den moderlige, og forholdene i dens habitat ikke vil falde sammen med de oprindelige, vil dette over tid føre til divergens og dannelsen af en ny art. Et slående eksempel på geografisk art er mangfoldigheden af finkearter opdaget af Charles Darwin under sin rejse på Beagle-skibet på Galapagos-øerne ud for Ecuadors kyst. Tilsyneladende endte individuelle individer af den eneste fink, der beboede det sydamerikanske kontinent, på en eller anden måde på øerne, og på grund af forskelle i forhold (primært fødetilgængelighed) og geografisk isolation udviklede de sig gradvist og dannede en gruppe af beslægtede arter.
I kernen sympatisk, eller biologisk artsdannelse ligger en form for reproduktiv isolation, hvor nye arter opstår inden for rækkevidden af den oprindelige art. En forudsætning for sympatrisk artsdannelse er den hurtige isolering af de resulterende former. Dette er en hurtigere proces end allopatrisk artsdannelse, og nye former ligner de oprindelige forfædre.
Sympatrisk artsdannelse kan være forårsaget af hurtige ændringer i kromosomsammensætning (polyploidisering) eller kromosomomlejringer. Nogle gange opstår nye arter som følge af hybridisering af to oprindelige arter, som for eksempel i tamblomme, som er en hybrid af slåen og kirsebærblomme. I nogle tilfælde er sympatrisk artsdannelse forbundet med opdelingen af økologiske nicher i populationer af samme art inden for et enkelt område eller sæsonbestemt isolation - divergens i tidspunktet for reproduktion i planter (forskellige typer fyrretræer i Californien producerer støv i februar og april) og i tidspunktet for reproduktion hos dyr.
Af hele mangfoldigheden af nyligt opståede arter kan kun få, de mest tilpassede, eksistere i lang tid og give anledning til nye arter. Årsagerne til de fleste arters død er stadig ukendte; højst sandsynligt skyldes dette pludselige klimaændringer, geologiske processer og deres forskydning af mere tilpassede organismer. I øjeblikket er en af årsagerne til et betydeligt antal arters død mennesket, der udrydder de største dyr og de smukkeste planter, og hvis denne proces i det 17. århundrede først begyndte med udryddelsen af den sidste runde, så i Det 21. århundrede forsvinder mere end 10 arter hver time.
Bevarelse af artsdiversitet som grundlag for biosfærens bæredygtighed
På trods af, at planeten ifølge forskellige skøn er hjemsted for 5-10 millioner arter af organismer, der endnu ikke er blevet beskrevet, vil vi aldrig vide om eksistensen af de fleste af dem, da omkring 50 arter forsvinder fra ansigtet jorden hver time. Forsvinden af levende organismer på nuværende tidspunkt er ikke nødvendigvis forbundet med deres fysiske udryddelse; oftere skyldes det ødelæggelsen af deres naturlige levesteder som følge af menneskelig aktivitet. Døden af en enkelt art vil næppe føre til fatale konsekvenser for biosfæren, men det har længe været fastslået, at udryddelsen af en planteart medfører døden af 10-12 dyrearter, og det udgør allerede nu en trussel mod eksistensen. af individuelle biogeocenoser og til det globale økosystem generelt.
De triste kendsgerninger akkumuleret i løbet af de foregående årtier tvang Den Internationale Union for Bevarelse af Natur og Naturressourcer (IUCN) til at begynde at indsamle oplysninger om sjældne og truede arter af planter og dyr i 1949. I 1966 udgav IUCN den første Red Book of Facts.
Rød bog er et officielt dokument, der indeholder løbende opdaterede data om status og udbredelse af sjældne og truede arter af planter, dyr og svampe.
Dette dokument vedtog en statusskala på fem niveauer for en beskyttet art, hvor det første beskyttelsesniveau inkluderer arter, hvis redning er umulig uden særlige foranstaltninger, og den femte - restaurerede art, hvis tilstand takket være de trufne foranstaltninger gør ikke give anledning til bekymring, men de er endnu ikke genstand for industriel brug. Udviklingen af en sådan skala gør det muligt at rette prioriterede bevaringsindsatser specifikt mod de mest sjældne arter, såsom Amur-tigre.
Ud over den internationale version af Røde Bog er der også nationale og regionale versioner. I USSR blev den røde bog oprettet i 1974, og i Den Russiske Føderation er proceduren for dens vedligeholdelse reguleret af de føderale love "om miljøbeskyttelse", "om dyreliv" og dekretet fra regeringen i Den Russiske Føderation " På Den Russiske Føderations Røde Bog”. I dag er 610 arter af planter, 247 dyrearter, 42 arter af lav og 24 arter af svampe opført i Den Russiske Føderations Røde Bog. Befolkningen af nogle af dem, der engang var truet (europæisk bæver, bison), er allerede blevet ret med succes genoprettet.
Følgende dyrearter er beskyttet i Rusland: russisk bisamrotte, tarbagan (mongolsk murmeldyr), isbjørn, kaukasisk europæisk mink, havodder, manul, amur tiger, leopard, sneleopard, søløve, hvalros, sæler, delfiner, hvaler, Przewalski's hest, vildrøv, pink pelikan, almindelig flamingo, sort stork, lille svane, steppeørn, kongeørn, sort trane, sibirisk trane, bustard, ørnugle, hvid måge, middelhavsskildpadde, japansk slange, hugorm, jungletudse, kaspisk lampret , alle typer størfisk, sølaks, hjortebille, ekstraordinær humlebi, almindelig Apollo, mantiskrabbe, almindelig perlemusling mv.
Planterne i Den Russiske Føderations Røde Databog omfatter 7 arter af vintergækker, nogle typer malurt, ægte ginseng, 7 typer blåklokker, takket eg, scilla, 11 arter af iris, russisk hasselryper, Schrenks tulipan, nøddebærende lotus, dametøffel, tyndbladet pæon, fjergræs, Julia's primula, eng lumbago (søvn-græs), belladonna belladonna, Pitsunda fyr, taks, kinesisk skjoldmad, søgræs, blød sphagnum, krøllet phyllophora, filamentous chara, filamentous chara, etc.
Sjældne svampe er repræsenteret af sommertrøffel eller russisk sort trøffel, lakeret tindersvamp osv.
Beskyttelsen af sjældne arter er i de fleste tilfælde forbundet med et forbud mod deres ødelæggelse, bevarelse af dem i kunstigt skabte levesteder (zoologiske haver), beskyttelse af deres levesteder og oprettelse af lavtemperatur genetiske banker.
Den mest effektive foranstaltning til beskyttelse af sjældne arter er bevarelsen af deres levesteder, hvilket opnås ved at organisere et netværk af særligt beskyttede beskyttede områder, der i overensstemmelse med den føderale lov "On Specially Protected Natural Areas" (1995) har internationale , føderal, regional eller lokal betydning. Disse omfatter statslige naturreservater, nationalparker, naturparker, statslige naturreservater, naturmonumenter, dendrologiske parker, botaniske haver mv.
Statens Naturreservat- dette er et særligt beskyttet naturkompleks (jord, vandområder, undergrund, flora og fauna) fuldstændig trukket tilbage fra økonomisk brug, som har miljømæssig, videnskabelig, miljømæssig og uddannelsesmæssig betydning som et eksempel på det naturlige miljø, typiske eller sjældne landskaber, steder hvor den genetiske fond af planter er bevaret og dyreverdenen.
Reserver, der er en del af det internationale system af biosfærereservater, der udfører global miljøovervågning, har status statens naturlige biosfærereservater. Reservatet er en miljø-, forsknings- og miljøuddannelsesinstitution, der har til formål at bevare og studere det naturlige forløb af naturlige processer og fænomener, den genetiske fond af flora og fauna, individuelle arter og samfund af planter og dyr, typiske og unikke økologiske systemer.
I øjeblikket er der omkring 100 statslige naturreservater i Rusland, hvoraf 19 har biosfærestatus, herunder Baikalsky, Barguzinsky, Kaukasisk, Kedrovaya Pad, Kronotsky, Prioksko-Terrasny osv.
I modsætning til naturreservater, territorier (vandområder) nationalparker omfatte naturkomplekser og genstande, der har særlige miljømæssige, historiske og æstetiske værdier, og som er beregnet til brug til miljømæssige, uddannelsesmæssige, videnskabelige og kulturelle formål og til reguleret turisme. 39 særligt beskyttede naturområder har denne status, herunder nationalparkerne Trans-Baikal og Sochi, samt nationalparkerne "Curonian Spit", "Russian North", "Shushensky Bor" osv.
Naturparker er miljømæssige rekreative institutioner under jurisdiktionen af de konstituerende enheder i Den Russiske Føderation, hvis territorier (vandområder) omfatter naturlige komplekser og genstande, der har betydelige miljømæssige og æstetiske værdier, og er beregnet til brug til miljømæssige, uddannelsesmæssige og rekreative formål.
Statens naturreservater er territorier (vandområder), der er af særlig betydning for bevarelse eller genopretning af naturlige komplekser eller deres komponenter og opretholdelse af den økologiske balance.
Udvikling af evolutionære ideer. Betydningen af Charles Darwins evolutionsteori. Indbyrdes sammenhæng mellem evolutionens drivkræfter. Former for naturlig udvælgelse, typer af kamp for tilværelsen. Syntetisk evolutionsteori. Elementære evolutionsfaktorer. Forskning af S. S. Chetverikov. Evolutionsteoriens rolle i dannelsen af det moderne naturvidenskabelige billede af verden
Udvikling af evolutionære ideer
Alle teorier om oprindelsen og udviklingen af den organiske verden kan reduceres til tre hovedretninger: kreationisme, transformisme og evolutionisme. Kreationisme er begrebet arternes varighed, idet man tager den organiske verdens mangfoldighed i betragtning som et resultat af dens skabelse af Gud. Denne retning blev dannet som et resultat af etableringen af den kristne kirkes dominans i Europa, baseret på bibelske tekster. Fremtrædende repræsentanter for kreationismen var C. Linnaeus og J. Cuvier.
"The Prince of Botanists" C. Linnaeus, der opdagede og beskrev hundredvis af nye plantearter og skabte deres første harmoniske system, hævdede ikke desto mindre, at det samlede antal arter af organismer er forblevet uændret siden skabelsen af Jorden, dvs. ikke blot ikke vises igen, men og forsvinder ikke. Først mod slutningen af sit liv kom han til den konklusion, at slægter er Guds værk, mens arter kan udvikle sig på grund af tilpasning til lokale forhold.
Den fremragende franske zoolog J. Cuviers (1769-1832) bidrag til biologien var baseret på talrige data fra palæontologi, komparativ anatomi og fysiologi læren om sammenhænge- forhold mellem dele af kroppen. Takket være dette blev det muligt at rekonstruere dyrets ydre udseende i individuelle dele. I processen med palæontologisk forskning kunne J. Cuvier dog ikke lade være med at være opmærksom på både den åbenlyse overflod af fossile former og de skarpe ændringer i dyregrupperne i den geologiske historie. Disse data tjente som udgangspunkt for formuleringen katastrofeteorier, ifølge hvilken alle eller næsten alle organismer på Jorden gentagne gange blev dræbt som følge af periodiske naturkatastrofer, og derefter blev planeten genbefolket af arter, der overlevede katastrofen. Tilhængerne af J. Cuvier talte op til 27 sådanne katastrofer i Jordens historie. Betragtninger om evolution syntes for J. Cuvier at være skilt fra virkeligheden.
Modsætningerne i kreationismens oprindelige præmisser, som blev mere og mere tydelige efterhånden som videnskabelige fakta akkumulerede, tjente som udgangspunkt for dannelsen af et andet system af synspunkter - transformisme, der anerkender arternes virkelige eksistens og deres historiske udvikling. Repræsentanter for denne tendens - J. Buffon, I. Goethe, E. Darwin og E. Geoffroy Saint-Hilaire, der ikke var i stand til at afsløre de sande årsager til evolution, reducerede dem til tilpasning til miljøforhold og arven af erhvervede egenskaber. Transformismens rødder kan findes i værker af antikke græske og middelalderlige filosoffer, der anerkendte historiske ændringer i den organiske verden. Således udtrykte Aristoteles ideen om naturens enhed og den gradvise overgang fra legemer af livløs natur til planter og fra dem til dyr - "naturstigen". Han anså hovedårsagen til ændringer i levende organismer for at være deres indre ønske om perfektion.
Den franske naturforsker J. Buffon (1707-1788), hvis vigtigste livsværk var Natural History i 36 bind, udvidede i modsætning til kreationisternes synspunkter omfanget af Jordens historie til 80-90 tusind år. Samtidig bemærkede han floraens og faunaens enhed samt muligheden for ændringer i beslægtede organismer under påvirkning af miljøfaktorer som følge af domesticering og hybridisering.
Den engelske læge, filosof og digter E. Darwin (1731–1802), Charles Darwins bedstefar, mente, at historien om den organiske verden går millioner af år tilbage, og dyreverdenens mangfoldighed er resultatet af en blanding af flere “ naturlige” grupper, påvirkning af det ydre miljø, træning og mangel på træningsorganer og andre faktorer.
E. Geoffroy Saint-Hilaire (1772-1844) anså enhederne i strukturplanen for grupper af dyr for at være et af de vigtigste beviser for udviklingen af den levende verden. Men i modsætning til sine forgængere var han tilbøjelig til at tro, at ændringer i arter er forårsaget af indflydelsen af miljøfaktorer ikke på voksne individer, men på embryoner.
På trods af at der i diskussionen, der blussede op i 1831 mellem J. Cuvier og E. Geoffroy Saint-Hilaire i form af en række rapporter på Videnskabsakademiet, forblev en klar fordel på førstnævntes side, var det transformisme, der blev forløberen for evolutionismen. Evolutionisme(evolutionsteori, evolutionær doktrin) er et system af synspunkter, der anerkender naturens udvikling i henhold til visse love. Det er biologiens teoretiske højdepunkt, som giver os mulighed for at forklare mangfoldigheden og kompleksiteten af levende systemer, som vi observerer. Men på grund af det faktum, at evolutionær undervisning beskriver fænomener, der er svære at observere, står den over for betydelige vanskeligheder. Nogle gange kaldes evolutionsteorien "darwinisme" og identificeres med Charles Darwins lære, hvilket er grundlæggende forkert, fordi selvom Charles Darwins teori ydede et uvurderligt bidrag til udviklingen af ikke kun evolutionslæren, men også biologien i generelt (såvel som mange andre videnskaber), grundlaget for evolutionsteori blev lagt af andre videnskabsmænd, den fortsætter med at udvikle sig til denne dag, og "darwinisme" har i mange aspekter kun historisk betydning.
Skaberen af den første evolutionsteori - Lamarckismen - var den franske naturforsker J. B. Lamarck (1744–1829). Han anså evolutionens drivkraft for at være organismers indre ønske om perfektion ( lov om graduering), dog tvinger tilpasning til miljøforhold dem til at afvige fra denne hovedlinje. Samtidig udvikles de organer, der intensivt bruges af dyret i livets proces, og de, der er unødvendige for det, tværtimod svækkes og kan endda forsvinde ( lov om motion og ikke-motion af organer). Karakteristika erhvervet i løbet af livet er fastlagt og givet videre til efterkommere. Således forklarede han tilstedeværelsen af membraner mellem tæerne på vandfugle med deres forfædres forsøg på at bevæge sig i vandmiljøet, og giraffernes lange hals er ifølge Lamarck en konsekvens af, at deres forfædre forsøgte at få blade fra toppen af træer.
Ulemperne ved Lamarckismen var den teoretiske karakter af mange konstruktioner, såvel som antagelsen om Skaberens indgriben i evolutionen. I processen med udviklingen af biologi blev det klart, at individuelle ændringer erhvervet af organismer i løbet af livet for det meste falder inden for grænserne for fænotypisk variabilitet, og deres overførsel er praktisk talt umulig. For eksempel skar den tyske zoolog og evolutionsteoretiker A. Weismann (1834-1914) haler af mus i mange generationer og fik altid kun halegnavere i deres afkom. Teorien om J. B. Lamarck blev ikke accepteret af hans samtidige, men ved århundredskiftet dannede den grundlag for den såkaldte neo-lamarckisme.
Betydningen af Charles Darwins evolutionsteori
Forudsætningerne for skabelsen af Charles Darwins mest berømte evolutionsteori, eller darwinismen, var udgivelsen i 1778 af den engelske økonom T. Malthus' arbejde "Treatise on Population", geologen Charles Lyells arbejde, formuleringen af celleteorien, selektionens succes i England og Charles egne observationer.Darwin (1809–1882), taget under sine studier i Cambridge, under ekspeditionen som naturforsker på Beagle og ved dens afslutning.
Således argumenterede T. Malthus for, at Jordens befolkning stiger eksponentielt, hvilket væsentligt overstiger planetens evne til at forsyne den med mad og fører til, at nogle af afkommet dør. Paralleller tegnet af Charles Darwin og hans medforfatter A. Wallace (1823-1913) indikerede, at individer i naturen formerer sig med meget høj hastighed, men populationsstørrelserne forbliver relativt konstante. Den engelske geolog C. Lyells forskning gjorde det muligt at fastslå, at Jordens overflade ikke altid var den samme, som den er nu, og dens ændringer var forårsaget af påvirkning af vand, vind, vulkanudbrud og levende aktivitet. organismer. Selv i sine studieår blev Charles Darwin selv ramt af billers ekstreme variation og under sine rejser af ligheden mellem floraen og faunaen på det kontinentale Sydamerika og de nærliggende Galapagos-øer, og på samme tid af den betydelige mangfoldighed af arter, såsom finker og skildpadder. Derudover var han på ekspeditionen i stand til at observere skeletterne af gigantiske uddøde pattedyr, svarende til moderne bæltedyr og dovendyr, hvilket markant rystede hans tro på skabelsen af arter.
De vigtigste bestemmelser i evolutionsteorien blev udtrykt af Charles Darwin i 1859 på et møde i Royal Society of London og udviklede efterfølgende i bøgerne "The Origin of Species by Means of Natural Selection, or the Preservation of Favored Breeds in the Kamp for livet" (1859), "Forandringer i husdyr og dyrkede planter" "(1868), "Menneskets oprindelse og seksuel udvælgelse" (1871), "Følelsernes udtryk hos mennesker og dyr" (1872) osv. .
Essensen udviklet af Charles Darwin evolutionsbegreber kan reduceres til en række af hinanden hidrørende bestemmelser, der har tilsvarende bevis:
- De individer, der udgør enhver population, producerer mange flere afkom, end det er nødvendigt for at opretholde populationsstørrelsen.
- På grund af det faktum, at livsressourcer for enhver type levende organismer er begrænsede, opstår der uundgåeligt mellem dem kamp for tilværelsen. Charles Darwin skelnede mellem intraspecifik og interspecifik kamp, såvel som kamp med miljøfaktorer. Samtidig påpegede han også, at vi ikke kun taler om et bestemt individs kamp for tilværelsen, men også for at efterlade afkom.
- Konsekvensen af kampen for tilværelsen er naturlig selektion- den overvejende overlevelse og reproduktion af organismer, der ved et uheld viste sig at være de mest tilpassede til de givne eksistensbetingelser. Naturlig selektion ligner på mange måder kunstig selektion, som mennesker har brugt siden oldtiden til at fremavle nye sorter af planter og racer af husdyr. Ved at udvælge individer, der har nogle ønskværdige egenskaber, bevarer mennesket disse egenskaber gennem kunstig avl gennem selektiv avl eller bestøvning. En særlig form for naturlig selektion er seksuel selektion for egenskaber, der normalt ikke har direkte adaptiv betydning (lange fjer, enorme horn osv.), men bidrager til reproduktiv succes, fordi de gør individet mere attraktivt for det modsatte køn eller mere formidabelt for at rivaler af samme køn.
- Materialet for evolution er forskellene mellem organismer, der opstår som følge af deres variabilitet. Charles Darwin skelnede mellem ubestemt og bestemt variabilitet. Bestemte(gruppe)variabilitet viser sig i alle individer af en art lige under indflydelse af en bestemt faktor og forsvinder i efterkommere, når virkningen af denne faktor ophører. Usikker(individuel) variabilitet er ændringer, der forekommer i hvert individ, uanset udsving i værdierne af miljøfaktorer, og som overføres til efterkommere. En sådan variabilitet har ikke en adaptiv karakter. Efterfølgende viste det sig, at en vis variabilitet er ikke-arvelig, og ubestemt variabilitet er arvelig.
- Naturlig udvælgelse fører i sidste ende til divergens i de individuelle isolerede sorters egenskaber - divergens og i sidste ende til dannelsen af nye arter.
Charles Darwins evolutionsteori postulerede ikke kun processen med arters fremkomst og udvikling, men afslørede også selve evolutionsmekanismen, som er baseret på princippet om naturlig udvælgelse. Darwinismen benægtede også evolutionens programmerede natur og postulerede dens kontinuerlige natur.
Samtidig kunne Charles Darwins evolutionsteori ikke besvare en række spørgsmål, for eksempel om arten af genetisk materiale og dets egenskaber, essensen af arvelig og ikke-arvelig variabilitet og deres evolutionære rolle. Dette førte til en darwinisme-krise og fremkomsten af nye teorier: neo-lamarckisme, saltationisme, begrebet nomogenese osv. Neo-Lamarckisme er baseret på positionen i J. B. Lamarcks teori om nedarvning af erhvervede egenskaber. Saltationisme er et system af synspunkter om evolutionsprocessen som pludselige ændringer, der fører til den hurtige fremkomst af nye arter, slægter og større systematiske grupper. Koncept nomogenese postulerer den programmerede udviklingsretning og udviklingen af forskellige karakteristika baseret på interne love. Kun syntesen af darwinisme og genetik i 20-30'erne af det tyvende århundrede var i stand til at overvinde de modsætninger, der uundgåeligt opstod, når man forklarede en række fakta.
Indbyrdes sammenhæng mellem evolutionens drivkræfter
Evolution kan ikke forbindes med virkningen af en enkelt faktor, da mutationer i sig selv er tilfældige og urettede ændringer og ikke kan sikre individers tilpasning til miljøfaktorer, mens naturlig selektion allerede sorterer disse ændringer. På samme måde kan selektion i sig selv ikke være den eneste faktor i evolutionen, da selektion kræver passende materiale leveret af mutationer.
Det kan dog bemærkes, at mutationsprocessen og genstrømmen skaber variation, mens naturlig selektion og genetisk drift sorterer denne variation fra. Dette betyder, at faktorer, der skaber variabilitet, initierer mikroevolutionsprocessen, og dem, der sorterer variabilitet, fortsætter den, hvilket fører til etablering af nye frekvenser af varianter. Således kan evolutionære ændringer inden for en befolkning ses som et resultat af modsatrettede kræfter, der skaber og sorterer genotypisk variation.
Et eksempel på samspillet mellem mutationsprocessen og selektion er hæmofili hos mennesker. Hæmofili er en sygdom forårsaget af nedsat blodpropper. Det har tidligere ført til døden i den præ-reproduktive periode, da enhver skade på det bløde væv potentielt kan føre til betydeligt blodtab. Denne sygdom er forårsaget af en recessiv mutation af det kønsbundne gen H (Xh). Kvinder lider ekstremt sjældent af hæmofili; de er oftere heterozygote bærere, men deres sønner kan arve sygdommen. Teoretisk set dør sådanne mænd i løbet af flere generationer før puberteten, og gradvist skulle denne allel forsvinde fra befolkningen, men hyppigheden af forekomsten af denne sygdom falder ikke på grund af gentagne mutationer i dette locus, som det skete i Dronning Victoria, som overførte sygdommen til tre generationer af Europas kongehuse. Den konstante hyppighed af denne sygdom indikerer en balance mellem mutationsprocessen og selektionstrykket.
Former for naturlig udvælgelse, typer af kamp for tilværelsen
Naturlig selektion De kalder selektiv overlevelse og efterladelse af afkom af de mest egnede individer og døden af de mindst egnede.
Essensen af naturlig selektion i evolutionsteorien ligger i den differentierede (ikke-tilfældige) bevarelse af visse genotyper i en population og deres selektive deltagelse i overførslen af gener til den næste generation. Desuden påvirker det ikke et enkelt træk (eller gen), men hele fænotypen, som er dannet som et resultat af genotypens interaktion med miljøfaktorer. Naturlig selektion vil være af forskellig karakter under forskellige miljøforhold. I øjeblikket er der flere former for naturlig selektion: stabilisering, kørsel og rivning.

Stabiliserende valg har til formål at konsolidere en snæver reaktionsnorm, som viste sig at være den mest gunstige under de givne eksistensbetingelser. Det er typisk for de tilfælde, hvor fænotypiske egenskaber er optimale for uændrede miljøforhold. Et slående eksempel på virkningen af stabiliserende selektion er bevarelsen af en relativt konstant kropstemperatur hos varmblodede dyr. Denne form for udvælgelse blev studeret i detaljer af den fremragende russiske zoolog I. I. Shmalgauzen.
Kørselsvalg opstår som reaktion på ændringer i miljøforhold, som følge af, at mutationer, der afviger fra egenskabens gennemsnitlige værdi, bevares, mens den tidligere dominerende form ødelægges, fordi den ikke i tilstrækkelig grad opfylder de nye eksistensbetingelser. For eksempel i England, som et resultat af luftforurening fra industrielle emissioner, blev birkemølsommerfugle, som tidligere ikke var set mange steder, med mørkefarvede vinger, som var mindre synlige for fugle på baggrund af sodede birkestammer, udbredt. Kørselsvalg bidrager ikke til den fuldstændige ødelæggelse af den form, den virker imod, da situationen med luftforurening som følge af foranstaltninger truffet af regeringen og miljøorganisationer er blevet markant forbedret, og farven på sommerfuglevingerne er vendt tilbage til dens originale version.
Rivning, eller forstyrrende valg favoriserer bevarelsen af ekstreme varianter af en egenskab og fjerner mellemliggende, da der for eksempel som følge af brugen af pesticider opstår grupper af insekter, der er resistente over for det. I sin mekanisme er disruptiv selektion det modsatte af stabiliserende selektion. Gennem denne form for selektion opstår flere skarpt afgrænsede fænotyper i en population. Dette fænomen kaldes polymorfi. Forekomsten af reproduktiv isolation mellem forskellige former kan føre til artsdannelse.
Nogle gange betragtes de også separat destabiliserende udvælgelse, som bevarer mutationer, der fører til en bred vifte af enhver karakteristik, for eksempel farven og strukturen af skallerne på nogle bløddyr, der lever under de heterogene mikrobetingelser i havets stenede brændinger. Denne form for selektion blev opdaget af D.K. Belyaev, mens han studerede domesticering af dyr.
I naturen eksisterer ingen af formerne for naturlig udvælgelse i ren form, men der er tværtimod forskellige kombinationer af dem, og efterhånden som miljøforholdene ændrer sig, kommer først den ene eller den anden af dem i forgrunden. Efter afslutning af ændringer i miljøet erstattes drivende selektion således af stabiliserende selektion, som optimerer en gruppe individer i nye eksistensbetingelser.
Naturlig selektion sker på forskellige niveauer, og derfor skelnes der også mellem individuel, gruppe og seksuel selektion. Individuel selektion fjerner mindre tilpassede individer fra at deltage i reproduktion, mens gruppeselektion har til formål at bevare en egenskab, der ikke er nyttig for et individ, men for gruppen som helhed. Under pres gruppe selektion kan fuldstændig udslette hele populationer, arter og større grupper af organismer uden at efterlade afkom. I modsætning til individuel udvælgelse reducerer gruppeudvælgelse mangfoldigheden af former i naturen.
Seksuel udvælgelse udføres inden for ét køn. Det fremmer udviklingen af egenskaber, der sikrer succes med at efterlade det største afkom. Takket være denne form for naturlig udvælgelse har seksuel dimorfisme udviklet sig, udtrykt i størrelsen og farven på påfuglens hale, hjortens gevirer osv.
Naturlig udvælgelse er resultatet kamp for tilværelsen baseret på arvelig variation. Kampen for tilværelsen forstås som hele sættet af relationer mellem individer af ens egen og andre arter, såvel som med abiotiske miljøfaktorer. Disse forhold bestemmer et bestemt individs succes eller fiasko med at overleve og producere afkom. Årsagen til kampen for tilværelsen er fremkomsten af et overskydende antal individer i forhold til de tilgængelige ressourcer. Ud over konkurrence bør disse relationer også omfatte gensidig bistand, hvilket øger chancerne for at overleve individer.
Samspil med miljøfaktorer kan også føre til døden for langt de fleste individer, for eksempel hos insekter, hvoraf kun en lille del overlever vinteren.
Syntetisk evolutionsteori
Genetikens succeser i begyndelsen af det tyvende århundrede, for eksempel opdagelsen af mutationer, antydede, at arvelige ændringer i organismers fænotype opstår pludseligt og ikke dannes over en længere periode, som postuleret af evolutionsteorien om Charles Darwin. Imidlertid førte yderligere forskning inden for befolkningsgenetik til formuleringen i 20-50'erne af det tyvende århundrede af et nyt system af evolutionære synspunkter - syntetisk evolutionsteori. Væsentlige bidrag til dets skabelse blev ydet af videnskabsmænd fra forskellige lande: sovjetiske videnskabsmænd S. S. Chetverikov, I. I. Shmalgauzen og A. N. Severtsov, engelsk biokemiker og genetiker D. Haldane, amerikanske genetikere S. Wright og F. Dobzhansky, evolutionist D. Simpson og zoolog E. Mayr.
Grundlæggende bestemmelser i den syntetiske evolutionsteori:
- Evolutionens elementære materiale er arvelig variabilitet (mutationel og kombinativ) hos individer af en population.
- Evolutionens elementære enhed er den befolkning, hvori alle evolutionære ændringer forekommer.
- Et elementært evolutionært fænomen er en ændring i den genetiske struktur af en befolkning.
- Elementære evolutionsfaktorer - genetisk drift, livsbølger, genflow - er urettede, tilfældige i naturen.
- Den eneste retningsbestemte faktor i evolutionen er naturlig udvælgelse, som er kreativ i naturen. Naturlig udvælgelse kan være stabiliserende, drivende og forstyrrende.
- Evolution er divergerende i naturen, det vil sige, at ét taxon kan give anledning til flere nye taxa, mens hver art kun har én forfader (art, population).
- Evolution er gradvis og kontinuerlig. Speciation som et trin i den evolutionære proces er den sekventielle udskiftning af en population med en række andre midlertidige populationer.
- Der er to typer evolutionære processer: mikroevolution og makroevolution. Makroevolution har ikke sine egne specielle mekanismer og udføres kun takket være mikroevolutionære mekanismer.
- Enhver systematisk gruppe kan enten blomstre (biologisk fremgang) eller dø ud (biologisk regression). Biologiske fremskridt opnås gennem ændringer i organismers struktur: aromorfoser, idioadaptationer eller generel degeneration.
- Evolutionens vigtigste love er dens irreversible natur, den progressive komplikation af livsformer og udviklingen af arternes tilpasningsevne til deres miljø. Samtidig har evolutionen ikke et endeligt mål, det vil sige, at processen er ustyret.
På trods af at evolutionsteorien gennem de seneste årtier er blevet beriget med data fra beslægtede videnskaber - genetik, selektion osv., tager den stadig ikke højde for en række aspekter, for eksempel rettede ændringer i arveligt materiale, derfor i i fremtiden er det muligt at skabe et nyt evolutionsbegreb, der vil erstatte den syntetiske teori.
Elementære evolutionsfaktorer
Ifølge den syntetiske evolutionsteori består et elementært evolutionært fænomen af en ændring i den genetiske sammensætning af en befolkning, og begivenheder og processer, der fører til ændringer i genpuljer, kaldes elementære evolutionsfaktorer. Disse omfatter mutationsprocessen, befolkningsbølger, genetisk drift, isolation og naturlig selektion. På grund af den usædvanlige betydning af naturlig udvælgelse i evolutionen, vil den blive betragtet separat.
Mutationsproces som er lige så kontinuerlig som selve evolutionen, opretholder den genetiske heterogenitet i befolkningen på grund af fremkomsten af flere og flere nye genvarianter. Mutationer, der opstår under påvirkning af eksterne og interne faktorer, klassificeres som gen, kromosomal og genomisk.
Genmutationer forekommer med en frekvens på 10 –4 –10 –7 pr. gamet, men på grund af det faktum, at det samlede antal gener hos mennesker og de fleste højere organismer kan nå op på flere titusinder, er det umuligt at forestille sig, at to organismer er absolut identisk. De fleste mutationer, der opstår, er recessive, især da dominerende mutationer umiddelbart er genstand for naturlig selektion. Recessive mutationer skaber netop den reserve af arvelig variabilitet, men før de manifesterer sig i fænotypen, skal de etablere sig hos mange individer i en heterozygot tilstand på grund af fri krydsning i befolkningen.
Kromosomale mutationer forbundet med tab eller overførsel af en del af et kromosom (et helt kromosom) til et andet, er også ret almindelige i forskellige organismer, for eksempel ligger forskellen mellem nogle arter af rotter i et enkelt kromosompar, hvilket gør det svært at krydse dem.
Genomiske mutationer, forbundet med polyploidisering, fører også til reproduktiv isolation af den nyligt opståede befolkning på grund af forstyrrelser i mitose af den første deling af zygoten. Ikke desto mindre er de ret udbredte i planter, og sådanne planter kan vokse i de arktiske og alpine enge på grund af deres større modstand mod miljøfaktorer.
Kombinativ variabilitet, som sikrer fremkomsten af nye varianter af at kombinere gener i genotypen, og følgelig øger sandsynligheden for fremkomsten af nye fænotyper, bidrager også til evolutionære processer, da antallet af varianter af kromosomkombinationer alene hos mennesker er 2 23, dvs. udseendet af en organisme, der ligner den, der allerede eksisterer, er næsten umulig.
Befolkningsbølger. Det modsatte resultat (udtømning af gensammensætning) er ofte forårsaget af udsving i antallet af organismer i naturlige populationer, som hos nogle arter (insekter, fisk osv.) kan ændre sig titusinder eller hundredvis af gange - befolkningsbølger, eller "livets bølger". En stigning eller et fald i antallet af individer i populationer kan være enten periodisk, så ikke-periodisk. De første er sæsonbestemte eller flerårige, såsom træk hos trækfugle, eller reproduktion hos dafnier, som kun har hunindivider om foråret og sommeren, og om efteråret dukker der hanner op, som er nødvendige for seksuel reproduktion. Ikke-periodiske udsving i antallet er ofte forårsaget af en kraftig stigning i mængden af mad i et gunstigt år, forstyrrelse af levesteder og spredning af skadedyr eller rovdyr.
Da populationsgenopretning sker på grund af et lille antal individer, der ikke har hele sættet af alleler, vil de nye og oprindelige populationer have forskellige genetiske strukturer. En ændring i hyppigheden af gener i en population under påvirkning af tilfældige faktorer kaldes genetisk drift, eller genetisk-automatiske processer. Det opstår også under udviklingen af nye territorier, fordi de modtager et ekstremt begrænset antal individer af en given art, som kan give anledning til en ny bestand. Derfor er genotyperne af disse individer ( grundlægger effekt). Som et resultat af genetisk drift opstår der ofte nye homozygote former (for mutante alleler), som kan vise sig at være adaptivt værdifulde og efterfølgende vil blive opfanget af naturlig selektion.
Blandt den indiske befolkning på det amerikanske kontinent og Laplanderne er andelen af personer med blodgruppe I (0) således meget høj, mens gruppe III og IV er yderst sjældne. Sandsynligvis, i det første tilfælde, var grundlæggerne af befolkningen personer, der ikke havde IB-allelen, eller den gik tabt under udvælgelsesprocessen.
Op til et vist punkt sker der en udveksling af alleler mellem nabopopulationer som følge af krydsning mellem individer af forskellige populationer - genflow, hvilket reducerer divergensen mellem individuelle populationer, men med fremkomsten af isolation stopper det. I det væsentlige er genflow en forsinket mutationsproces.
Isolering. Eventuelle ændringer i befolkningens genetiske struktur skal rettes op, hvilket er det, der sker takket være isolation- fremkomsten af eventuelle barrierer (geografiske, miljømæssige, adfærdsmæssige, reproduktive osv.), som komplicerer og umuliggør krydsning af individer af forskellige befolkningsgrupper. Selvom isolation ikke i sig selv skaber nye former, bevarer den ikke desto mindre genetiske forskelle mellem populationer, der er underlagt virkningen af naturlig selektion. Der er to former for isolation: geografisk og biologisk.
Geografisk isolation opstår som følge af opdelingen af området ved fysiske barrierer (vandhindringer for terrestriske organismer, landområder for akvatiske arter, vekslen af høje områder og sletter); Dette lettes af en stillesiddende eller vedhæftet (i planter) livsstil. Nogle gange kan geografisk isolation være forårsaget af udvidelsen af en arts rækkevidde med den efterfølgende udryddelse af dens populationer i mellemliggende territorier.
Biologisk isolation er en konsekvens af visse divergenser af organismer inden for den samme art, der på en eller anden måde forhindrer fri krydsning. Der er flere typer af biologisk isolation: miljømæssig, sæsonbestemt, etologisk, morfologisk og genetisk. Miljøisolering opnås gennem opdelingen af økologiske nicher (f.eks. præference for bestemte levesteder eller fødevaretyper, som i grankorsnæb og fyrkorsnæb). Sæson(midlertidig) isolation observeres i tilfælde af reproduktion af individer af samme art på forskellige tidspunkter (forskellige sildebestande). Etologisk isolation afhænger af adfærdens karakteristika (træk ved frieriritualet, farvning, "sang" af kvinder og mænd fra forskellige befolkningsgrupper). På morfologisk isolation En hindring for krydsning er uoverensstemmelsen i strukturen af de reproduktive organer eller endda kropsstørrelse (pekingeser og grand danois). Genetisk isolation har den største effekt og viser sig i kønscellers uforenelighed (zygotens død efter befrugtning), sterilitet eller nedsat levedygtighed af hybrider. Årsagerne til dette er ejendommelighederne ved antallet og formen af kromosomer, som et resultat af hvilken fuld celledeling (mitose og meiose) bliver umulig.
Ved at forstyrre fri krydsning mellem populationer forstærker isolation derved i dem de forskelle, der opstod på genotypisk niveau på grund af mutationer og fluktuationer i antal. I dette tilfælde er hver population underlagt virkningen af naturlig selektion adskilt fra den anden, og dette fører i sidste ende til divergens.
Den naturlige selektions kreative rolle i evolutionen
Naturlig selektion fungerer som en slags "si", der sorterer genotyper efter deres konditionsgrad. Charles Darwin understregede dog, at selektion ikke kun og ikke så meget har til formål udelukkende at bevare det bedste, men at fjerne det værste, det vil sige, at det giver dig mulighed for at bevare multivarians. Funktionen af naturlig udvælgelse er ikke begrænset til dette, da det sikrer reproduktionen af tilpassede genotyper, og dermed bestemmer udviklingsretningen, konsekvent tilføjer tilfældige og talrige afvigelser. Naturlig selektion har ikke et specifikt mål: baseret på det samme materiale (arvelig variabilitet) under forskellige forhold, kan forskellige resultater opnås.
I denne henseende kan den overvejede evolutionsfaktor ikke sammenlignes med arbejdet hos en billedhugger, der hugger en marmorblok; snarere fungerer den som en fjern forfader til mennesket, der laver et værktøj af et stenfragment uden at forestille sig det endelige resultat, hvilket ikke kun afhænger af stenens beskaffenhed og dens form, men og af styrken, retningen af slaget osv. Men i tilfælde af svigt afviser selektion, ligesom et humanoidt væsen, den "forkerte" form.
Prisen for udvælgelse er forekomsten genetisk belastning, det vil sige akkumulering af mutationer i en befolkning, som over tid kan blive dominerende på grund af de fleste individers pludselige død eller migration af et lille antal af dem.
Under presset fra naturlig selektion dannes ikke kun mangfoldigheden af arter, men deres organisationsniveau øges også, herunder deres komplikation eller specialisering. Men i modsætning til kunstig selektion, som kun udføres af mennesker for økonomisk værdifulde egenskaber, ofte på bekostning af adaptive egenskaber, kan naturlig selektion ikke bidrage til dette, da ingen tilpasning i naturen kan kompensere for skaden fra et fald i levedygtigheden af befolkningen.
Forskning af S. S. Chetverikov
Et af de vigtige skridt hen imod forsoning af darwinisme og genetik blev taget af Moskva-zoologen S. S. Chetverikov (1880-1959). Baseret på resultaterne af en undersøgelse af den genetiske sammensætning af naturlige populationer af frugtfluen Drosophila, beviste han, at de bærer mange recessive mutationer i en heterozygot form, der ikke krænker fænotypisk ensartethed. De fleste af disse mutationer er ugunstige for kroppen og skaber den såkaldte genetisk belastning, hvilket reducerer befolkningens tilpasningsevne til dets miljø. Nogle mutationer, der ikke har adaptiv betydning på et givet tidspunkt i artens udvikling, kan senere få en vis værdi og er således reserve af arvelig variabilitet. Spredningen af sådanne mutationer blandt individer af en population som et resultat af successive frie krydsninger kan i sidste ende føre til deres overgang til en homozygot tilstand og manifestation i fænotypen. Hvis denne tilstand af tegnet er hårtørrer- er adaptiv, så vil den efter et par generationer fuldstændig fortrænge den dominerende phen, sammen med dens bærere, fra den population, der er mindre passende til de givne forhold. På grund af sådanne evolutionære ændringer bibeholdes kun den recessive mutante allel, og dens dominerende allel forsvinder.
Lad os prøve at bevise dette med et specifikt eksempel. Når du studerer en bestemt population, kan du opdage, at ikke kun dens fænotypiske, men også dens genotypiske struktur kan forblive uændret i lang tid på grund af fri krydsning, eller panmixia diploide organismer.
Dette fænomen er beskrevet af loven Hardy-Weinberg, ifølge hvilken i en ideel population af ubegrænset størrelse i fravær af mutationer, migrationer, befolkningsbølger, genetisk drift, naturlig selektion og underlagt fri krydsning, vil frekvenserne af alleler og genotyper af diploide organismer ikke ændre sig over en række generationer .
For eksempel, i en population er et bestemt træk kodet af to alleler af det samme gen - dominant ( EN) og recessiv ( EN). Frekvensen af den dominante allel er betegnet som R og recessiv - q. Summen af frekvenserne af disse alleler er 1: s + q= 1. Derfor, hvis vi kender frekvensen af den dominante allel, så kan vi bestemme frekvensen af den recessive allel: q = 1 – s. Faktisk er allelernes frekvenser lig med sandsynligheden for dannelsen af de tilsvarende gameter. Derefter, efter dannelsen af zygoter, vil genotypefrekvenserne i den første generation være:
(pA + qa) 2 = s 2 A.A. + 2pqAa + q 2 aa = 1,
Hvor s 2 A.A.- hyppighed af dominerende homozygoter;
2pqAa- hyppighed af heterozygoter;
q 2 aa- hyppighed af recessive homozygoter.
Det er let at beregne, at i efterfølgende generationer vil frekvensen af genotyper forblive den samme, hvilket opretholder den genetiske diversitet i befolkningen. Men i naturen er der ingen ideelle populationer, og derfor kan mutante alleler i dem ikke kun fortsætte, men også sprede sig og endda erstatte tidligere mere almindelige alleler.
S. S. Chetverikov indså tydeligt, at naturlig selektion ikke blot eliminerer individuelle mindre succesrige egenskaber og følgelig allelerne, der koder for dem, men også virker på hele komplekset af gener, der påvirker manifestationen af et bestemt gen i fænotypen, eller genotypisk miljø. Som et genotypisk miljø betragtes hele genotypen i øjeblikket som et sæt gener, der kan forstærke eller svække manifestationen af specifikke alleler.
Ikke mindre vigtige i udviklingen af evolutionær undervisning er studierne af S. S. Chetverikov inden for befolkningsdynamik, især "livsbølger" eller befolkningsbølger. Mens han stadig var studerende, publicerede han i 1905 en artikel om muligheden for udbrud af massereproduktion af insekter og et lige så hurtigt fald i deres antal.
Evolutionsteoriens rolle i dannelsen af det moderne naturvidenskabelige billede af verden
Betydningen af evolutionsteori i udviklingen af biologi og andre naturvidenskaber kan næppe overvurderes, da den var den første til at forklare betingelserne, årsagerne, mekanismerne og resultaterne af den historiske udvikling af livet på vores planet, det vil sige, den gav en materialistisk forklaring på udviklingen af den organiske verden. Derudover var teorien om naturlig udvælgelse den første virkelig videnskabelige teori om biologisk evolution, da Charles Darwin, da den blev skabt, ikke stolede på spekulative konstruktioner, men gik ud fra sine egne observationer og stolede på levende organismers virkelige egenskaber. Samtidig berigede hun de biologiske redskaber med den historiske metode.
Formuleringen af evolutionsteorien forårsagede ikke kun en ophedet videnskabelig debat, men gav også skub til udviklingen af sådanne videnskaber som generel biologi, genetik, selektion, antropologi og en række andre. I denne henseende kan man ikke andet end at være enig i udsagnet om, at evolutionsteorien kronede det næste trin i biologiens udvikling og blev udgangspunktet for dens fremskridt i det tyvende århundrede.
Bevis på udviklingen af den levende natur. Resultater af evolution: organismers tilpasningsevne til deres miljø, mangfoldighed af arter
Bevis på udviklingen af dyrelivet
Inden for forskellige områder af biologien, selv før Charles Darwin og efter offentliggørelsen af hans evolutionsteori, blev der opnået en hel række beviser til støtte for den. Dette bevis kaldes bevis på evolution. De oftest citerede er palæontologiske, biogeografiske, komparative embryologiske, komparative anatomiske og komparative biokemiske beviser for evolution, selvom taksonomidata, såvel som udvælgelsen af planter og dyr, ikke kan udelukkes.
Palæontologiske beviser baseret på undersøgelse af fossile rester af organismer. Disse omfatter ikke kun velbevarede organismer frosset i is eller indkapslet i rav, men også "mumier" fundet i sure tørvemoser samt rester af organismer og fossiler bevaret i sedimentære bjergarter. Tilstedeværelsen i gamle klipper af simplere organismer end i senere lag, og det faktum, at arter fundet på et niveau forsvinder på et andet, betragtes som et af de mest betydningsfulde beviser på evolution og forklares ved fremkomsten og udryddelsen af arter i tilsvarende epoker pga. til ændringer i miljøforhold.
På trods af at der hidtil er blevet opdaget få fossile rester, og mange fragmenter mangler i fossilregistret på grund af den lave sandsynlighed for bevarelse af organiske rester, er der stadig fundet former for organismer, der har tegn på både evolutionært ældre og yngre grupper af organismer. Sådanne former for organismer kaldes overgangsformer. Fremtrædende repræsentanter for overgangsformer, der illustrerer overgangen fra fisk til terrestriske hvirveldyr, er lobefinnede fisk og stegocephaler, og Archaeopteryx indtager et bestemt sted mellem krybdyr og fugle.
Rækker af fossile former, der konsekvent er forbundet med hinanden i evolutionsprocessen, ikke kun af generelle, men også af særlige strukturelle træk, kaldes fylogenetiske serier. De kan være repræsenteret af fossile rester fra forskellige kontinenter og hævder at være mere eller mindre komplette, men deres undersøgelse er umulig uden sammenligning med levende former for at demonstrere udviklingen af den evolutionære proces. Et klassisk eksempel på en fylogenetisk serie er udviklingen af hestens forfædre, studeret af grundlæggeren af evolutionær palæontologi V. O. Kovalevsky.
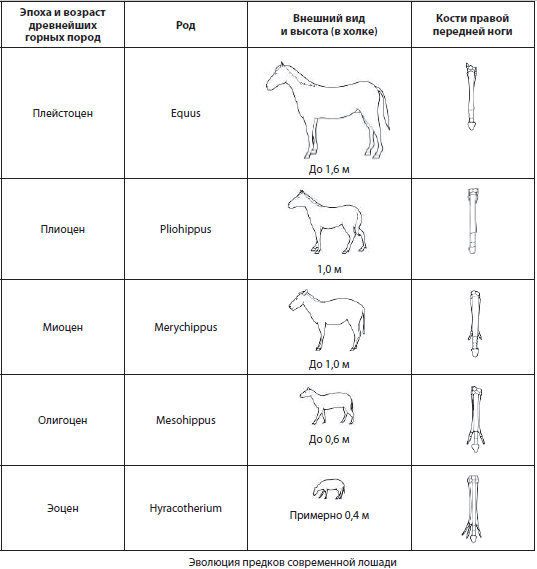
Biogeografiske beviser. Biogeografi hvordan videnskaben studerer mønstrene for distribution og udbredelse af arter, slægter og andre grupper af levende organismer, såvel som deres samfund, på overfladen af vores planet.
Fraværet i nogen del af jordens overflade af arter af organismer, der er tilpasset et sådant levested og slår godt rod, når de er kunstigt importeret, som kaniner i Australien, samt tilstedeværelsen af lignende former for organismer i dele af landet beliggende kl. betydelige afstande fra hinanden indikerer først og fremmest, at Jordens udseende ikke altid var på denne måde, og geologiske transformationer, især kontinentaldrift, dannelsen af bjerge, stigningen og faldet af verdenshavets niveau påvirker organismers udvikling. For eksempel lever fire lignende arter af lungefisk i de tropiske områder i Sydamerika, Sydafrika og Australien, mens levestederne for kameler og lamaer, der tilhører samme orden, er placeret i Nordafrika, det meste af Asien og Sydamerika. Palæontologiske undersøgelser har vist, at kameler og lamaer stammer fra en fælles forfader, der engang levede i Nordamerika, og derefter spredte sig til Asien gennem den allerede eksisterende landtange på stedet for Beringstrædet, og også gennem landtangen i Panama til Sydamerika. Efterfølgende uddøde alle repræsentanter for denne familie i de mellemliggende regioner, og i de regionale regioner blev der dannet nye arter i evolutionsprocessen. Den tidligere adskillelse af Australien fra andre landmasser tillod dannelsen af en helt speciel flora og fauna der, hvor sådanne former for pattedyr som monotremes - næbdyret og echidna - blev bevaret.
Fra et biogeografisk synspunkt kan mangfoldigheden af Darwins finker på Galapagos-øerne, som ligger 1200 km fra Sydamerikas kyst og er af vulkansk oprindelse, også forklares. Tilsyneladende fløj repræsentanter for den eneste art af finker i Ecuador engang eller blev introduceret til dem, og efterhånden som de formerede sig, slog nogle af individerne sig ned på tværs af de resterende øer. På de centrale store øer var kampen for tilværelsen (fødevarer, redepladser osv.) mest akut, hvorfor der blev dannet arter, der var lidt forskellige fra hinanden i ydre karakteristika, som indtog forskellige fødevarer (frø, frugter, nektar, insekter, etc.) .).
De påvirkede fordelingen af forskellige grupper af organismer og ændringer i klimatiske forhold på Jorden, hvilket bidrog til nogle gruppers velstand og andres udryddelse. Individuelle arter eller grupper af organismer, der har overlevet fra tidligere udbredte floraer og faunaer, kaldes relikvier. Disse omfatter ginkgo, sequoia, tulipantræ, coelacanth med fligefinnede fisk osv. I bredere forstand kaldes arter af planter og dyr, der lever i begrænsede områder af territorium eller vandområde. endemisk, eller endemisk. For eksempel er alle repræsentanter for den oprindelige flora og fauna i Australien endemiske, og i Baikal-søens flora og fauna er op til 75% af dem endemiske.
Sammenlignende anatomiske beviser. Studiet af anatomien af beslægtede grupper af dyr og planter giver overbevisende beviser for ligheden i strukturen af deres organer. På trods af at miljøfaktorer bestemt sætter deres præg på strukturen af organer, i angiospermer, med al deres fantastiske mangfoldighed, blomster har bægerblade, kronblade, støvdragere og pistiller, og i terrestriske hvirveldyr, er lemmen bygget i henhold til en femfingret plan. Organer, der har en lignende struktur, indtager samme position i kroppen og udvikler sig fra de samme rudimenter i beslægtede organismer, men udfører forskellige funktioner, kaldes homolog. Således er hørebenene (hammer, incus og stigbøjlen) homologe med fiskens gællebuer, slangernes giftige kirtler er spytkirtlerne hos andre hvirveldyr, mælkekirtlerne hos pattedyr er svedkirtlerne, svømmefødderne hos sæler og hvaler. er fuglenes vinger, lemmerne på heste og muldvarpe.

Organer, der ikke har fungeret i lang tid, bliver højst sandsynligt til rudimenter- strukturer, der er underudviklede i sammenligning med forfædres former og har mistet deres grundlæggende betydning. Disse omfatter fibula hos fugle, øjne hos muldvarpe og muldvarpe rotter, hår, haleben og blindtarm hos mennesker osv.
Individuelle individer kan dog udvise egenskaber, der er fraværende hos en given art, men som var til stede i fjerne forfædre - atavismer, for eksempel tretåethed hos moderne heste, udviklingen af yderligere par mælkekirtler, en hale og hår på hele menneskekroppen.
Hvis homologe organer er beviser til fordel for organismers slægtskab og divergens i evolutionsprocessen, så lignende organer- lignende strukturer i organismer af forskellige grupper, der udfører de samme funktioner, tværtimod er eksempler konvergens(konvergens er den generelt uafhængige udvikling af lignende egenskaber i forskellige grupper af organismer, der eksisterer under de samme forhold) og bekræfte det faktum, at miljøet efterlader et betydeligt aftryk på organismen. Analoger er vingerne på insekter og fugle, øjnene på hvirveldyr og blæksprutter (blæksprutter, blæksprutter) og ledlemmer på leddyr og landlevende hvirveldyr.

Sammenlignende embryologisk bevis. Ved at studere embryonal udvikling hos repræsentanter for forskellige grupper af hvirveldyr, opdagede K. Baer deres slående strukturelle enhed, især i de tidlige udviklingsstadier ( lov om germinal lighed). Senere formulerede E. Haeckel biogenetisk lov, ifølge hvilken ontogenese er en kort gentagelse af fylogeni, dvs. de stadier, som en organisme gennemgår i processen med sin individuelle udvikling, gentager den historiske udvikling af den gruppe, den tilhører.
Således får et hvirveldyrs embryo i de første udviklingsstadier strukturelle træk, der er karakteristiske for fisk, og derefter for padder og i sidste ende for den gruppe, det tilhører. Denne transformation forklares af det faktum, at hver af de ovennævnte klasser har fælles forfædre med moderne krybdyr, fugle og pattedyr.

Den biogenetiske lov har imidlertid en række begrænsninger, og derfor begrænsede den russiske videnskabsmand A. N. Severtsov omfanget af dens anvendelse betydeligt ved at gentage i ontogenese udelukkende funktionerne i de embryonale stadier af udvikling af forfædres former.
Sammenlignende biokemiske beviser. Udviklingen af mere nøjagtige metoder til biokemisk analyse har givet evolutionsforskere en ny gruppe af data til fordel for den historiske udvikling af den organiske verden, da tilstedeværelsen af de samme stoffer i alle organismer indikerer mulig biokemisk homologi, svarende til den ved niveau af organer og væv. Sammenlignende biokemiske undersøgelser af den primære struktur af så udbredte proteiner som cytochrom Med og hæmoglobin, såvel som nukleinsyrer, især rRNA, har vist, at mange af dem har næsten samme struktur og udfører de samme funktioner i repræsentanter for forskellige arter, og jo tættere forholdet er, jo større lighed findes i strukturen af de undersøgte stoffer.
Evolutionsteorien bekræftes således af en betydelig mængde data fra forskellige kilder, som endnu en gang indikerer dens pålidelighed, men den vil stadig ændre sig og blive forfinet, da mange aspekter af organismers liv forbliver uden for forskernes synsfelt. .
Resultater af evolution: organismers tilpasningsevne til deres miljø, mangfoldighed af arter
Ud over de generelle karakteristika, der er karakteristiske for repræsentanter for et bestemt kongerige, er arter af levende organismer kendetegnet ved en forbløffende række af funktioner i ydre og indre struktur, livsaktivitet og endda adfærd, der dukkede op og blev udvalgt i evolutionsprocessen og sikrer tilpasning til levevilkår. Man skal dog ikke gå ud fra, at da fugle og insekter har vinger, skyldes det luftens direkte påvirkning, for der er masser af vingeløse insekter og fugle. De ovennævnte tilpasninger blev udvalgt gennem en naturlig selektionsproces fra hele spektret af tilgængelige mutationer.
Epifytiske planter, som ikke lever på jorden, men på træer, har tilpasset sig at absorbere atmosfærisk fugt ved hjælp af rødder uden rodhår, men med et særligt hygroskopisk væv - velamen. Nogle bromeliaer kan absorbere vanddamp i den fugtige atmosfære i troperne ved hjælp af hårene på deres blade.
Insektædende planter (soldug, Venus-fluefælder), der lever på jord, hvor nitrogen er utilgængeligt af en eller anden grund, har udviklet en mekanisme til at tiltrække og absorbere små dyr, oftest insekter, som er en kilde til det nødvendige element for dem.
For at beskytte mod at blive spist af planteædere har mange planter, der fører en knyttet levevis, udviklet passive beskyttelsesmidler, såsom torne (tjørn), torne (rose), brændende hår (nælde), ophobning af krystaller af calciumoxalat (syre) , biologisk aktive stoffer i væv (kaffe, tjørn) osv. I nogle af dem er selv frøene i umodne frugter omgivet af stenede celler, der forhindrer skadedyr i at nå dem, og først om efteråret sker processen med aftræning, hvilket lader frøene komme ind i jorden og spire (pære).
Miljøet har også en dannende indflydelse på dyr. Mange fisk og vandpattedyr har således en strømlinet kropsform, som gør det lettere for dem at bevæge sig gennem dens tykkelse. Men man bør ikke antage, at vand direkte påvirker kroppens form; det er simpelthen, at i udviklingsprocessen viste de dyr, der havde denne egenskab, sig at være de mest tilpassede til det.
Kroppen af hvaler og delfiner er ikke dækket af hår, mens den beslægtede gruppe af pinnipeds har en mere eller mindre reduceret hårpels, da de i modsætning til førstnævnte tilbringer en del af deres tid på land, hvor deres hud uden uld umiddelbart ville blive iskold.
Kroppen af de fleste fisk er dækket af skæl, som på undersiden er lysere farvet end på toppen, som et resultat af hvilket disse dyr næppe er mærkbare ovenfra for naturlige fjender på baggrund af bunden, og nedefra - mod bunden. himlens baggrund. Farvelægning, der gør dyr usynlige for deres fjender eller bytte, kaldes nedladende. Det er udbredt i naturen. Et slående eksempel på sådan farvning er farvningen af undersiden af vingerne på callima-sommerfuglen, som efter at have siddet på en gren og foldet sine vinger sammen, viser sig at ligne et tørt blad. Andre insekter, såsom pindeinsekter, camouflerer sig selv som plantekviste.
Plettet eller stribet farve har også adaptiv betydning, da fugle som vagtler eller edderfugle på baggrund af jorden ikke er synlige selv på tæt hold. De plettede æg fra fugle, der yngler på jorden, er også usynlige.
Dyrenes farve er ikke altid lige så konstant som en zebras; for eksempel skrubber og kamæleon er i stand til at ændre det afhængigt af arten af det sted, hvor de er. Gøg kan, ved at placere deres æg i forskellige fugles reder, variere farven på deres skaller på en sådan måde, at "ejere" af reden ikke bemærker forskellene mellem den og deres egne æg.
Farvningen af dyr gør dem ikke altid usynlige - mange af dem fanger simpelthen øjet, hvilket burde advare om fare. De fleste af disse insekter og krybdyr er giftige i en eller anden grad, såsom en mariehøne eller en hveps, så et rovdyr, der har oplevet ubehagelige fornemmelser flere gange efter at have spist en sådan genstand, undgår det. Alligevel, advarselsfarve er ikke universel, da nogle fugle har tilpasset sig til at fodre på dem (musvåge).
De øgede chancer for overlevelse hos individer med advarselsfarve bidrog til dens optræden i repræsentanter for andre arter uden ordentlige grunde. Dette fænomen kaldes mimik. Således efterligner ikke-giftige larver af nogle arter af sommerfugle giftige, og mariehøns efterligner en af typerne af kakerlakker. Fugle kan dog hurtigt lære at skelne giftige organismer fra ikke-giftige og forbruge sidstnævnte og undgå de individer, der tjente som rollemodeller.
I nogle tilfælde kan det modsatte fænomen også observeres - rovdyr imiterer harmløse dyr i farven, hvilket giver dem mulighed for at nærme sig offeret på kort afstand og derefter angribe (sabeltandet blenny).
Beskyttelse for mange arter ydes også af adaptiv adfærd, som er forbundet med opbevaring af mad til vinteren, pasning af afkom, frysning på plads eller omvendt at indtage en truende stilling. Således forbereder flodbævere flere kubikmeter grene, dele af stammer og anden planteføde til vinteren og oversvømmer den i vand nær "hytterne".
Omsorg for afkom er hovedsageligt karakteristisk for pattedyr og fugle, men det findes også i repræsentanter for andre klasser af chordater. For eksempel kendes den aggressive adfærd af hanklæbende kævler, der driver alle fjender væk fra reden, hvor æggene er placeret. Hanklørede frøer vikler æggene rundt om deres ben og bærer dem, indtil haletudserne klækkes.
Selv nogle insekter er i stand til at give deres afkom et mere gunstigt levested. For eksempel fodrer bier deres larver, og unge bier "arbejder" først i stadet. Myrer flytter deres pupper op og ned i myretuen, afhængig af temperatur og luftfugtighed, og når der er fare for oversvømmelse, tager de dem generelt med. Carab biller forbereder specielle bolde fra dyreaffald til deres larver.
Når de er truet af angreb, fryser mange insekter på plads og tager form af tørre pinde, kviste og blade. Hugorme derimod rejser sig og puster deres hætte op, mens klapperslangen laver en speciel lyd med en rangle placeret for enden af dens hale.
Adfærdsmæssige tilpasninger suppleres af fysiologiske tilpasninger relateret til habitatets egenskaber. En person er således i stand til at opholde sig under vand uden dykkerudstyr i kun et par minutter, hvorefter han kan miste bevidstheden og dø på grund af iltmangel, og hvaler kommer ikke til overfladen ret længe. Deres lungevolumen er ikke for stort, men der er andre fysiologiske tilpasninger, for eksempel i musklerne er der en høj koncentration af det respiratoriske pigment - myoglobin, som så at sige lagrer ilt og frigiver det under nedsænkning. Derudover har hvaler en særlig formation - et "vidunderligt netværk", som tillader brugen af ilt selv fra venøst blod.
Dyr i varme levesteder, såsom ørkener, risikerer konstant at blive overophedet og miste overskydende fugt. Derfor har fennec-ræven ekstremt store ører, der gør, at den kan udstråle varme. Padder i ørkenområder er, for at undgå tab af fugt gennem huden, tvunget til at skifte til en natlig livsstil, når fugtigheden stiger, og duggen opstår.
Fugle, der har mestret luftens habitat, har udover anatomiske og morfologiske tilpasninger til flyvning også vigtige fysiologiske egenskaber. For eksempel på grund af det faktum, at bevægelse i luften kræver et ekstremt højt energiforbrug, er denne gruppe af hvirveldyr karakteriseret ved et højt stofskifte, og de udskilte stofskifteprodukter elimineres med det samme, hvilket er med til at reducere kroppens specifikke tæthed.
Tilpasninger til miljøet er på trods af al deres perfektion relative. Nogle arter af milkweed producerer således alkaloider, der er giftige for de fleste dyr, men larverne fra én art af sommerfugle - danaids - lever ikke kun af mælkevæv, men akkumulerer også disse alkaloider og bliver uspiselige for fugle.
Derudover er tilpasninger kun nyttige i et bestemt miljø og er ubrugelige i et andet miljø. For eksempel har et sjældent og stort rovdyr, Ussuri-tigeren, ligesom alle katte, bløde puder på poterne og udtrækkelige skarpe kløer, skarpe tænder, fremragende syn selv i mørke, skarp hørelse og stærke muskler, som gør det muligt for den at opdage sine bytte, snige sig ind på det ubemærket og baghold. Dens stribede farve camouflerer den dog kun forår, sommer og efterår, mens den i sneen bliver tydeligt synlig og tigeren kun kan regne med et lynhurtigt angreb.
Figenblomsterstande, der producerer værdifulde frugter, har en så specifik struktur, at de kun bestøves af blastophagous hvepse, og derfor, når de blev introduceret i kultur, bar de ikke frugt i lang tid. Kun udviklingen af parthenokarpiske varianter af figner (dannende frugter uden befrugtning) kunne redde situationen.
På trods af, at der er beskrevet eksempler på artsdannelse over meget korte perioder, som i tilfældet med ranglen på de kaukasiske enge, der på grund af regelmæssig slåning først delte sig i to populationer - tidlig blomstring og frugtsætning og sen blomstring, i virkeligheden kræver mikroevolution højst sandsynligt meget længere perioder - mange århundreder, fordi menneskeheden, hvis forskellige grupper var adskilt fra hinanden i årtusinder, ikke desto mindre aldrig blev opdelt i forskellige arter. Men da evolutionen har praktisk talt ubegrænset tid, over hundreder af millioner og milliarder af år, har adskillige milliarder arter allerede levet på Jorden, hvoraf de fleste er uddøde, og dem, der er kommet ned til os, er kvalitative stadier af denne igangværende proces.
Ifølge moderne data er der over 2 millioner arter af levende organismer på Jorden, hvoraf de fleste (ca. 1,5 millioner arter) tilhører dyreriget, omkring 400 tusind til planteriget, over 100 tusind til svamperiget, og hvile - til bakterier. En sådan forbløffende mangfoldighed er resultatet af divergens (divergens) af arter i henhold til forskellige morfologiske, fysiologisk-biokemiske, økologiske, genetiske og reproduktive egenskaber. For eksempel omfatter en af de største slægter af planter, der tilhører Orchidaceae-familien, dendrobium, over 1.400 arter, og slægten af biller omfatter over 1.600 arter.
Klassificering af organismer er en opgave inden for taksonomi, som i 2 tusinde år har forsøgt at bygge ikke bare et harmonisk hierarki, men et "naturligt" system, der afspejler graden af slægtskab af organismer. Alle forsøg på at gøre dette er dog endnu ikke blevet kronet med succes, da der i en række tilfælde i evolutionsprocessen ikke kun blev observeret divergens af karakterer, men også konvergens (konvergens), hvilket resulterede i meget fjerne grupper organerne erhvervede ligheder, såsom øjnene på blæksprutter og øjnene på pattedyr.
Makroevolution. Evolutionsretninger og -veje (A. N. Severtsov, I. I. Shmalgauzen). Biologiske fremskridt og regression, aromorfose, idioadaptation, degeneration. Årsager til biologisk fremskridt og regression. Hypoteser om livets oprindelse på Jorden. Grundlæggende aromorfoser i udviklingen af planter og dyr. Komplikation af levende organismer i evolutionsprocessen
Makroevolution
Dannelsen af en art markerer en ny runde af den evolutionære proces, da individer af denne art, der er mere tilpasset miljøforhold end individer af forældrearten, gradvist slår sig ned i nye territorier, og mutagenese, befolkningsbølger, isolation og naturlig selektion spiller deres kreative rolle i dens befolkninger. Disse bestande giver med tiden anledning til nye arter, som på grund af genetisk isolation har meget flere ligheder med hinanden end med arten af slægten, som forældrearten forgrenede sig fra, og dermed opstår en ny slægt, derefter en ny familie, orden (orden), klasse osv. Det sæt af evolutionære processer, der fører til fremkomsten af supraspecifikke taxa (slægter, familier, ordener, klasser osv.) kaldes makroevolution. Makroevolutionære processer generaliserer så at sige mikroevolutionære ændringer, der opstår over en lang periode, mens de identificerer de vigtigste tendenser, retninger og mønstre for udviklingen af den organiske verden, som ikke er observerbare på et lavere niveau. Indtil videre er ingen specifikke mekanismer for makroevolution blevet identificeret, derfor menes det, at det kun udføres gennem mikroevolutionære processer, men denne position er konstant underlagt velbegrundet kritik.
Fremkomsten af et komplekst hierarkisk system af den organiske verden er i vid udstrækning resultatet af den ulige udviklingshastighed af forskellige grupper af organismer. Således blev den allerede nævnte ginkgo biloba sådan set "bevaret" i tusinder af år, mens fyrretræerne, der ligger ret tæt på den, har ændret sig markant i løbet af denne tid.
Evolutionsretninger og -veje (A. N. Severtsov, I. I. Shmalgauzen). Biologiske fremskridt og regression, aromorfose, idioadaptation, degeneration
Ved at analysere den organiske verdens historie kan man bemærke, at visse grupper af organismer i visse tidsrum dominerede, som derefter faldt eller forsvandt helt. Der kan således skelnes mellem tre hovedlinjer udviklingsretninger: biologisk fremskridt, biologisk regression og biologisk stabilisering. Et væsentligt bidrag til udviklingen af doktrinen om evolutionens retninger og veje blev ydet af de russiske evolutionister A. N. Severtsov og I. I. Shmalgauzen.
Biologiske fremskridt forbundet med den biologiske velstand for gruppen som helhed og kendetegner dens evolutionære succes. Det afspejler den naturlige udvikling af den levende natur fra simpel til kompleks, fra en lavere organisationsgrad til en højere. Ifølge A.N. Severtsov er kriterierne for biologisk fremskridt en stigning i antallet af individer i en given gruppe, en udvidelse af dens rækkevidde samt fremkomsten og udviklingen af lavere rangerende grupper inden for dens sammensætning (transformation af en art til en slægt, en slægt ind i en familie osv.). I øjeblikket observeres biologiske fremskridt i angiospermer, insekter, benfisk og pattedyr.
Ifølge A. N. Severtsov kan biologiske fremskridt opnås som et resultat af visse morfofysiologiske transformationer af organismer, og han identificerede tre hovedmåder til opnåelse: arogenese, allogenese og katagenese.

Arogenese, eller morfofysiologiske fremskridt, er forbundet med en betydelig udvidelse af rækkevidden af denne gruppe af organismer på grund af erhvervelsen af store strukturelle ændringer - aromorfoser.
Aromorfose kaldet en evolutionær transformation af en organismes struktur og funktioner, som øger dens organisationsniveau og åbner op for nye muligheder for tilpasning til forskellige eksistensbetingelser.
Eksempler på aromorfoser er fremkomsten af en eukaryot celle, multicellularitet, udseendet af et hjerte i fisk og dets opdeling i en komplet septum hos fugle og pattedyr, dannelsen af en blomst i angiospermer osv.
Allogenese, i modsætning til arogenese, er ikke ledsaget af en udvidelse af sortimentet, men inden for den gamle opstår der en betydelig mangfoldighed af former, der har særlige tilpasninger til miljøet - idioadaptationer.
Idiomatisk tilpasning- dette er en mindre morfofysiologisk tilpasning til særlige miljøforhold, nyttig i kampen for tilværelsen, men ændrer ikke organisationsniveauet. Disse ændringer illustreres af den beskyttende farve hos dyr, mangfoldigheden af munddele hos insekter, planternes rygsøjler osv. Et lige så vellykket eksempel er Darwins finker, der har specialiseret sig i forskellige typer føde, hvor transformationer først påvirkede næbbet, og derefter andre dele af kroppen - fjerdragt, hale og så videre.
Hvor paradoksalt det end kan virke, kan forenkling af organisationen føre til biologiske fremskridt. Denne vej kaldes katagenese.
Degeneration- dette er forenklingen af organismer i evolutionsprocessen, som er ledsaget af tab af visse funktioner eller organer.
Fasen med biologisk fremskridt erstattes af en fase biologisk stabilisering, hvis essens er at bevare en given arts egenskaber som de mest gunstige i et givet mikromiljø. Ifølge I. I. Shmalhausen betyder det "ikke ophør af evolution; tværtimod betyder det organismens maksimale sammenhæng med ændringer i miljøet." De "levende fossiler" af coelacanth, gingko osv. er i fasen af biologisk stabilisering.
Antipoden af biologiske fremskridt er biologisk regression- evolutionær tilbagegang af en given gruppe på grund af manglende evne til at tilpasse sig miljøændringer. Det viser sig i et fald i befolkningstal, indsnævring af intervaller og et fald i antallet af lavere rangerende grupper inden for en højere taxon. En gruppe af organismer, der er i en tilstand af biologisk regression, er truet af udryddelse. I den organiske verdens historie kan man se mange eksempler på dette fænomen, og på nuværende tidspunkt er regression karakteristisk for nogle bregner, padder og krybdyr. Med menneskets fremkomst skyldes biologisk regression ofte dets økonomiske aktiviteter.
Den organiske verdens udviklingsretninger og -veje udelukker ikke hinanden, det vil sige, at udseendet af aromorfose ikke betyder, at idioadaptation eller degeneration ikke længere kan forekomme. Tværtimod, ifølge hvad der blev udviklet af A. N. Severtsov og I. I. Shmalgauzen faseskifteregel, forskellige retninger af den evolutionære proces og måder at opnå biologiske fremskridt på afløser naturligt hinanden. I løbet af evolutionen kombineres disse veje: ret sjældne aromorfoser overfører en gruppe af organismer til et kvalitativt nyt organisationsniveau, og yderligere historisk udvikling følger vejen for idioadaptation eller degeneration, hvilket sikrer tilpasning til specifikke miljøforhold.
Årsager til biologisk fremskridt og regression
I evolutionsprocessen overvindes baren for naturlig udvælgelse, og følgelig er det kun de grupper af organismer, der udvikler sig, hvor arvelig variation skaber et tilstrækkeligt antal kombinationer, der kan sikre overlevelsen af gruppen som helhed.
De grupper, der af en eller anden grund ikke har sådan en reserve, er i de fleste tilfælde dømt til at uddø. Dette skyldes ofte lavt selektionstryk på tidligere stadier af den evolutionære proces, hvilket førte til snæver specialisering af gruppen eller endda degenerative fænomener. Konsekvensen af dette er manglende evne til at tilpasse sig nye miljøforhold, når der sker pludselige ændringer. Et slående eksempel på dette er dinosaurernes pludselige død på grund af et kæmpe himmellegemes fald til Jorden for 65 millioner år siden, hvilket resulterede i et jordskælv, stigningen af millioner af tons støv i luften, en skarp kuldeknap, og de fleste planters og planteædende dyrs død. Samtidig var forfædrene til moderne pattedyr, der ikke havde snævre præferencer for fødekilder og var varmblodede, i stand til at overleve disse forhold og indtage en dominerende stilling på planeten.
Hypoteser for livets oprindelse på Jorden
Af hele rækken af hypoteser for dannelsen af Jorden vidner det største antal fakta til fordel for "Big Bang" teorien. På grund af det faktum, at denne videnskabelige antagelse hovedsageligt er baseret på teoretiske beregninger, bliver Large Hadron Collider, bygget ved European Nuclear Research Center nær Genève (Schweiz), opfordret til at bekræfte det eksperimentelt. Ifølge Big Bang-teorien blev Jorden dannet for over 4,5 milliarder år siden sammen med Solen og andre planeter i solsystemet som et resultat af kondenseringen af en gas- og støvsky. Faldet i planetens temperatur og migrationen af kemiske elementer på den bidrog til dens lagdeling i kernen, kappen og skorpen, og de efterfølgende geologiske processer (bevægelse af tektoniske plader, vulkansk aktivitet osv.) forårsagede dannelsen af atmosfære og hydrosfære.
Liv har også eksisteret på Jorden i meget lang tid, som det fremgår af fossile rester af forskellige organismer i klipper, men fysiske teorier kan ikke besvare spørgsmålet om tidspunktet og årsagerne til dets fremkomst. Der er to modsatrettede synspunkter om livets oprindelse på Jorden: teorierne om abiogenese og biogenese. Teorier om abiogenese bekræfter muligheden for oprindelsen af levende ting fra ikke-levende ting. Disse omfatter kreationisme, hypotesen om spontan generering og teorien om biokemisk evolution af A.I. Oparin.
Grundlæggende stilling kreationisme verdens skabelse var et vist overnaturligt væsen (Skaberen), hvilket afspejles i myterne om verdens folk og religiøse kulter, men planetens alder og livet på den overstiger langt de datoer, der er angivet i disse kilder, og der er masser af uoverensstemmelser i dem.
Grundlægger teorier om spontan generering livet anses for at være den antikke græske videnskabsmand Aristoteles, som hævdede, at det er muligt for nye skabninger at dukke op flere gange, for eksempel regnorme fra vandpytter og orme og fluer fra råddent kød. Disse synspunkter blev dog tilbagevist i det 17.-19. århundrede af F. Redis og L. Pasteurs dristige eksperimenter.
Den italienske læge Francesco Redi anbragte i 1688 kødstykker i gryder og forseglede dem tæt, men der kom ingen orme i dem, hvorimod de dukkede op i åbne gryder. For at tilbagevise den fremherskende tro på, at livsprincippet var indeholdt i luften, gentog han sine eksperimenter, men forseglede ikke potterne, men dækkede dem med flere lag muslin, og igen dukkede livet ikke op. På trods af de overbevisende data indhentet af F. Redi, gav A. van Leeuwenhoeks forskning nyt foder til diskussioner om det "vitale princip", som fortsatte gennem det næste århundrede.
En anden italiensk forsker, Lazzaro Spallanzani, ændrede eksperimenterne med F. Redi i 1765 ved at koge kød og grøntsagsafkog i flere timer og forsegle dem. Efter flere dage fandt han heller ingen tegn på liv der og konkluderede, at levende ting kun kan opstå fra levende ting.
Det sidste slag mod teorien om spontan generering kom fra den store franske mikrobiolog Louis Pasteur i 1860, da han anbragte kogt bouillon i en S-halset kolbe og ikke fik nogen bakterier. Det ser ud til, at dette vidnede til fordel for teorierne om biogenese, men spørgsmålet forblev åbent om, hvordan den aller, allerførste organisme opstod.
Den sovjetiske biokemiker A.I. Oparin forsøgte at svare på det og kom til den konklusion, at sammensætningen af jordens atmosfære i de første faser af dens eksistens var helt anderledes end i vores tid. Mest sandsynligt bestod den af ammoniak, metan, kuldioxid og vanddamp, men indeholdt ikke fri ilt. Under påvirkning af elektriske udladninger med høj effekt og ved høje temperaturer kunne de enkleste organiske forbindelser syntetiseres i den, hvilket blev bekræftet af S. Miller og G. Ureys eksperimenter i 1953, som opnåede flere af de ovennævnte forbindelser. aminosyrer, simple kulhydrater, adenin, urinstof, samt simple fedtsyrer, myre- og eddikesyrer.
Ikke desto mindre betyder syntesen af organiske stoffer endnu ikke livets fremkomst, derfor fremførte A.I. Oparin hypotese om biokemisk evolution, ifølge hvilke forskellige organiske stoffer opstod og forenede sig til større molekyler i det lave vand i have og oceaner, hvor betingelserne for kemisk syntese og polymerisering er mest gunstige. RNA-molekyler anses i øjeblikket for at være de første bærere af liv.
Nogle af disse stoffer dannede gradvist stabile komplekser i vand - coacervater, eller coacervate dråber, der ligner fedtdråber i bouillon. Disse koacervater modtog forskellige stoffer fra den omgivende opløsning, som undergik kemiske transformationer, der opstod i dråberne. Ligesom organiske stoffer var koacervater ikke selv levende væsener, men var det næste skridt i deres opståen.
De koacervater, der havde et gunstigt forhold mellem stoffer i deres sammensætning, især proteiner og nukleinsyrer, takket være enzymproteinernes katalytiske egenskaber, fik med tiden evnen til at reproducere deres egen art og udføre metaboliske reaktioner, mens strukturen af proteiner var kodet af nukleinsyrer.
Udover reproduktion er levende systemer dog karakteriseret ved afhængighed af tilførsel af energi udefra. Dette problem blev i første omgang løst gennem iltfri nedbrydning af organiske stoffer fra miljøet (der var ikke ilt i atmosfæren på det tidspunkt), dvs.
heterotrof ernæring. Nogle af de optagne organiske stoffer viste sig at kunne akkumulere sollysets energi, såsom klorofyl, der gjorde det muligt for en række organismer at skifte til autotrof ernæring. Frigivelsen af ilt til atmosfæren under fotosyntesen førte til fremkomsten af mere effektiv iltrespiration, dannelsen af ozonlaget og i sidste ende fremkomsten af organismer på landjorden.
Resultatet af kemisk udvikling var således udseendet protobioter- primære levende organismer, hvorfra alle eksisterende arter stammer fra som følge af den biologiske udvikling.
Teorien om biokemisk evolution i vores tid er den mest bekræftede, men ideen om de specifikke mekanismer for livets oprindelse har ændret sig. For eksempel viste det sig, at dannelsen af organiske stoffer begynder i rummet, og organiske stoffer spiller en vigtig rolle selv i selve dannelsen af planeter, hvilket sikrer vedhæftning af små dele. Dannelsen af organiske stoffer sker også i planetens tarme: under et udbrud frigiver en vulkan op til 15 tons organisk materiale. Der er andre hypoteser om mekanismerne for koncentration af organiske stoffer: frysning af opløsningen, absorption (binding) på overfladen af visse mineralforbindelser, virkningen af naturlige katalysatorer osv. Fremkomsten af liv på Jorden er i øjeblikket umulig, da alle organiske stoffer, der spontant dannes på et hvilket som helst tidspunkt, planeter straks ville blive oxideret af atmosfærens frie oxygen eller brugt af heterotrofe organismer. Dette blev forstået tilbage i 1871 af Charles Darwin.
Teorier om biogenese benægte livets spontane oprindelse. De vigtigste er steady state-hypotesen og panspermihypotesen. Den første af dem er baseret på det faktum, at liv eksisterer for evigt, men på vores planet er der meget gamle klipper, hvor der ikke er spor af den organiske verdens aktivitet.
Panspermi hypotese hævder, at livets embryoner blev bragt til Jorden fra rummet af nogle aliens eller af guddommeligt forsyn. Denne hypotese understøttes af to fakta: behovet for alle levende ting, som er ret sjældent på planeten, men ofte findes i meteoritter, af molybdæn, samt opdagelsen af organismer, der ligner bakterier på meteoritter fra Mars. Hvordan liv opstod på andre planeter er dog stadig uklart.
Grundlæggende aromorfoser i udviklingen af planter og dyr
Plante- og dyreorganismer, der repræsenterer forskellige grene af udviklingen af den organiske verden, erhvervede uafhængigt visse strukturelle træk i processen med historisk udvikling, som vil blive beskrevet yderligere.
Hos planter er de vigtigste af dem overgangen fra haploid til diploid, uafhængighed af vand under befrugtningsprocessen, overgangen fra ekstern til intern befrugtning og forekomsten af dobbeltbefrugtning, opdeling af kroppen i organer, udvikling af det ledende system, komplikation og forbedring af væv, samt specialisering af bestøvning ved hjælp af insekter og distribution af frø og frugter.
Overgangen fra haploidi til diploidi gjorde planter mere modstandsdygtige over for miljøfaktorer på grund af en reduceret risiko for recessive mutationer. Tilsyneladende påvirkede denne transformation forfædrene til karplanter, som ikke inkluderer bryophytter, som er karakteriseret ved en overvægt af gametofytten i livscyklussen.
De vigtigste aromorfoser i udviklingen af dyr er forbundet med fremkomsten af multicellularitet og den stigende opdeling af alle organsystemer, fremkomsten af et stærkt skelet, udviklingen af centralnervesystemet, såvel som social adfærd i forskellige grupper af højt organiserede dyr, hvilket satte skub i menneskets fremskridt.
Komplikation af levende organismer i evolutionsprocessen
Historien om den organiske verden på Jorden studeres ud fra de bevarede rester, aftryk og andre spor af levende organismers vitale aktivitet. Hun er et videnskabsfag palæontologi. Baseret på det faktum, at resterne af forskellige organismer er placeret i forskellige klippelag, blev der skabt en geokronologisk skala, ifølge hvilken Jordens historie blev opdelt i bestemte tidsperioder: eoner, epoker, perioder og århundreder.
Eon kaldet en stor periode i geologisk historie, der kombinerer flere epoker. I øjeblikket skelnes der kun mellem to eoner: kryptozoikum (skjult liv) og phanerozoikum (manifest liv). Æra- dette er en tidsperiode i geologisk historie, som er en opdeling af en æon, som igen forener perioder. I kryptozoikum er der to epoker (arkæisk og proterozoikum), mens der i fanerozoikum er tre (paleozoikum, mesozoikum og kenozoikum).
En vigtig rolle i skabelsen af den geokronologiske skala blev spillet af vejledende fossiler- rester af organismer, der var talrige i visse perioder, og som er velbevarede.
Udvikling af liv i kryptozoikum. Det arkæiske og proterozoikum udgør det meste af livets historie (periode for 4,6 milliarder år siden - 0,6 milliarder år siden), men der er få oplysninger om livet i den periode. De første rester af organiske stoffer af biogen oprindelse er omkring 3,8 milliarder år gamle, og prokaryote organismer eksisterede allerede for 3,5 milliarder år siden. De første prokaryoter var en del af specifikke økosystemer - cyanobakterielle måtter, takket være aktiviteten af hvilke specifikke sedimentære klipper stromatolitter ("stentæpper") blev dannet.
At forstå livet i gamle prokaryote økosystemer blev hjulpet af opdagelsen af deres moderne analoger - stromatolitter i Shark Bay i Australien og specifikke film på jordoverfladen i Syvash Bay i Ukraine. På overfladen af cyanobakterielle måtter er der fotosyntetiske cyanobakterier, og under deres lag er der ekstremt forskellige bakterier fra andre grupper og arkæer. Mineralske stoffer, der sætter sig på måttens overflade og dannes på grund af dens vitale aktivitet, aflejres i lag (ca. 0,3 mm pr. år). Sådanne primitive økosystemer kan kun eksistere på steder, der er ubeboelige for andre organismer, og begge de ovennævnte levesteder er faktisk karakteriseret ved ekstremt høj saltholdighed.
Talrige data tyder på, at Jorden til at begynde med havde en vedvarende atmosfære, som omfattede: kuldioxid, vanddamp, svovloxid, samt kulilte, brint, svovlbrinte, ammoniak, methan osv. Jordens første organismer var anaerobe , men takket være cyanobakteriers fotosyntese blev der frigivet fri ilt til miljøet, som først hurtigt associerede med reduktionsmidler i miljøet, og først efter bindingen af alle reduktionsmidler begyndte miljøet at erhverve oxiderende egenskaber. Denne overgang er bevist ved aflejring af oxiderede former af jern - hæmatit og magnetit.
For omkring 2 milliarder år siden, som et resultat af geofysiske processer, flyttede næsten alt jern ubundet i sedimentære bjergarter til planetens kerne, og ilt begyndte at akkumulere i atmosfæren på grund af fraværet af dette element - "iltrevolutionen" fandt sted. Det var et vendepunkt i Jordens historie, som ikke kun indebar en ændring i atmosfærens sammensætning og dannelsen af en ozonskærm i atmosfæren - hovedforudsætningen for landbosættelsen, men også sammensætningen af den klipper dannet på jordens overflade.
En anden vigtig begivenhed fandt sted i Proterozoic - fremkomsten af eukaryoter. I de senere år har det været muligt at indsamle overbevisende beviser for teorien om den endosymbiogenetiske oprindelse af den eukaryote celle - gennem symbiosen af flere prokaryote celler. Sandsynligvis var "hoved"-forfaderen til eukaryoter archaea, som skiftede til absorption af madpartikler ved fagocytose. Det arvelige apparat bevægede sig dybt ind i cellen, men bibeholdt ikke desto mindre sin forbindelse med membranen på grund af overgangen af den ydre membran af den fremkommende kernemembran ind i membranerne i det endoplasmatiske retikulum.
Jordens geokronologiske historie Eon Æra Periode Begyndelse for millioner år siden Varighed, millioner år Udvikling af livet Phanerozoic Cenozoic Anthropogen 1,5 1,5 Fire istider, efterfulgt af oversvømmelser, førte til dannelsen af kuldebestandig flora og fauna (mammutter, moskusokser, rensdyr, lemminger). Udveksling af dyr og planter mellem kontinenter på grund af fremkomsten af landbroer. Dominans af placenta pattedyr. Udryddelse af mange store pattedyr. Dannelsen af mennesket som biologisk art og dets bosættelse. Tamning af dyr og dyrkning af planter. Forsvinden af mange arter af levende organismer på grund af menneskelig økonomisk aktivitet Neogen 25 23.5 Fordeling af korn. Dannelse af alle moderne ordener af pattedyr. Fremkomsten af aber Paleogene 65 40 Dominans af blomstrende planter, pattedyr og fugle. Fremkomsten af hovdyr, kødædende dyr, pinnipeds, primater, etc. Mesozoikum Kridt 135 70 Fremkomsten af angiospermer, pattedyr og fugle bliver talrige Jura 195 60 Krybdyrenes og blæksprutternes æra. Fremkomsten af pungdyr og placenta pattedyr. Gymnospermers dominans Trias 225 30 De første pattedyr og fugle. Krybdyr er talrige. Udbredelse af urteagtige sporer Paleozoikum Perm 280 55 Fremkomsten af moderne insekter. Udvikling af krybdyr. Udryddelse af en række hvirvelløse grupper. Fordeling af nåletræer Kulstof 345 65 Første krybdyr. Fremkomsten af vingede insekter. Bregner og padderok dominerer Devon 395 50 Fisk er talrige. De første padder. Fremkomsten af de vigtigste grupper af sporer, de første gymnospermer og svampe Silurian 430 35 Alger er rigelige. De første landplanter og dyr (edderkopper). Spøgelsesfisk og krebsdyrskorpioner er almindelige ordovicium 500 70 Koraller og trilobitter er rigelige. Blomstring af grønne, brune og røde alger. Fremkomsten af de første akkordater Kambrium 570 70 Talrige fiskefossiler. Søpindsvin og trilobitter er almindelige. Fremkomsten af flercellede alger Cryptose Proterozoic 2600 2000 Fremkomsten af eukaryoter. For det meste encellede grønalger er almindelige. Fremkomsten af flercellethed. Udbrud af flercellet dyrediversitet (fremkomst af alle typer hvirvelløse dyr) Archaea 3500 1500 De første spor af liv på Jorden er bakterier og cyanobakterier. Fremkomsten af fotosyntese
Bakterier absorberet af cellen kunne ikke fordøjes, men forblev i live og fortsatte med at fungere. Det menes, at mitokondrier stammer fra lilla bakterier, der mistede evnen til at fotosyntese og gik over til oxidation af organiske stoffer. Symbiose med andre fotosyntetiske celler førte til fremkomsten af plastider i planteceller. Sandsynligvis opstod flagellerne af eukaryote celler som et resultat af symbiose med bakterier, der ligesom moderne spirocheter var i stand til at vride sig. Først var det arvelige apparat af eukaryote celler opbygget omtrent på samme måde som prokaryoter, og først senere, på grund af behovet for at kontrollere en stor og kompleks celle, blev der dannet kromosomer. Genomerne af intracellulære symbionter (mitokondrier, plastider og flageller) bibeholdt generelt den prokaryote organisation, men de fleste af deres funktioner blev overført til det nukleare genom.
Eukaryote celler opstod gentagne gange og uafhængigt af hinanden. For eksempel er rødalger opstået som følge af symbiogenese med cyanobakterier og grønalger med proklorfytbakterier.
De resterende enkeltmembranorganeller og kernen af den eukaryote celle opstod ifølge endomembranteorien fra invaginationer af den prokaryote celles membran.
Det nøjagtige tidspunkt for udseendet af eukaryoter er ukendt, da der allerede i sedimenter omkring 3 milliarder år gamle er aftryk af celler med lignende størrelser. Eukaryoter er bestemt registreret i bjergarter omkring 1,5-2 milliarder år gamle, men først efter iltrevolutionen (ca. 1 milliard år siden) udviklede betingelser gunstige for dem.
I slutningen af den proterozoiske æra (mindst 1,5 milliarder år siden) eksisterede allerede flercellede eukaryote organismer. Multicellularitet er ligesom den eukaryote celle opstået gentagne gange i forskellige grupper af organismer.
Der er forskellige syn på oprindelsen af flercellede dyr. Ifølge nogle data var deres forfædre multinukleære, ciliatlignende celler, som derefter brød op i separate mononukleære celler.
Andre hypoteser forbinder oprindelsen af flercellede dyr med differentieringen af koloniale encellede celler. Forskellene mellem dem vedrører oprindelsen af cellelag i det oprindelige flercellede dyr. Ifølge E. Haeckels gastrea-hypotese sker dette ved invagination af en af væggene i en enkeltlags flercellet organisme, som i coelenterater. I modsætning hertil formulerede I. I. Mechnikov phagocytella-hypotesen, idet den betragtede forfædrene til flercellede organismer som enkeltlagede sfæriske kolonier som Volvox, der absorberede madpartikler ved fagocytose. Cellen, der fangede partiklen, mistede sin flagel og bevægede sig dybere ind i kroppen, hvor den udførte fordøjelsen og ved slutningen af processen vendte tilbage til overfladen. Med tiden blev cellerne opdelt i to lag med specifikke funktioner - det ydre gav bevægelse, og det indre gav fagocytose. I. I. Mechnikov kaldte en sådan organisme en phagocytella.
I lang tid tabte flercellede eukaryoter i konkurrence med prokaryote organismer, men i slutningen af Proterozoikum (800-600 millioner år siden) på grund af en skarp ændring i forholdene på Jorden - et fald i havniveauet, en stigning i iltkoncentrationen , et fald i koncentrationen af karbonater i havvand, regelmæssige cyklusser afkøling - flercellede eukaryoter opnåede fordele i forhold til prokaryoter. Hvis der indtil dette tidspunkt kun blev fundet individuelle flercellede planter og muligvis svampe, så var dyr også kendt fra dette tidspunkt i jordens historie. Af de faunaer, der opstod i slutningen af Proterozoikum, er Ediacaran og Vendian de bedst studerede. Dyr fra den vendiske periode indgår normalt i en særlig gruppe af organismer eller klassificeres som sådanne typer som coelenterater, fladorme, leddyr osv. Ingen af disse grupper har dog skeletter, hvilket kan tyde på fravær af rovdyr.
Udvikling af liv i den palæozoiske æra. Den palæozoiske æra, som varede mere end 300 millioner år, er opdelt i seks perioder: Kambrium, Ordovicium, Silur, Devon, Carbon (Carbon) og Perm.
I Den kambriske periode Landet bestod af flere kontinenter, primært beliggende på den sydlige halvkugle. De mest udbredte fotosyntetiske organismer i denne periode var cyanobakterier og rødalger. Foraminiferer og radiolarier levede i vandsøjlen. I Kambrium optræder et enormt antal skeletdyrske organismer, som det fremgår af talrige fossile rester. Disse organismer tilhørte cirka 100 typer af flercellede dyr, både moderne (svampe, coelenterater, orme, leddyr, bløddyr) og uddøde, for eksempel: det enorme rovdyr Anomalocaris og koloniale graptolitter, der flød i vandsøjlen eller var fastgjort til bunden. Landet forblev næsten ubeboet i hele Kambrium, men jorddannelsesprocessen var allerede begyndt af bakterier, svampe og muligvis laver, og i slutningen af perioden dukkede oligochaete-orme og tusindben op på land.
I Ordovicium periode Vandstanden i Verdenshavet steg, hvilket førte til oversvømmelsen af det kontinentale lavland. De vigtigste producenter i denne periode var grønne, brune og røde alger. I modsætning til Kambrium, hvor rev blev bygget af svampe, blev de i Ordovicium erstattet af koralpolypper. Gastropoder og blæksprutter blomstrede, og det samme gjorde trilobitter (nu uddøde slægtninge til spindlere). I denne periode blev akkordater, især kæbeløse, også indspillet for første gang. I slutningen af Ordovicium skete en stor udryddelsesbegivenhed, som ødelagde omkring 35% af familierne og mere end 50% af slægterne af havdyr.
Silur præget af øget bjergbyggeri, hvilket førte til udtørring af kontinentale platforme. Den ledende rolle i den hvirvelløse fauna i Silur blev spillet af blæksprutter, pighuder og gigantiske krebsdyrskorpioner, mens der blandt hvirveldyrene forblev et stort udvalg af kæbeløse dyr, og fisk dukkede op. I slutningen af perioden kom de første karplanter til land - næsehorn og lykofytter, som begyndte at kolonisere lavt vand og kysternes tidevandszone. De første repræsentanter for arachnid-klassen kom også til land.
I Devon periode Som et resultat af landets stigning blev der dannet store lavvandede farvande, som tørrede ud og endda frøs til, da klimaet blev endnu mere kontinentalt end i Silur. Havene er domineret af koraller og pighuder, mens blæksprutter er repræsenteret af spiralformede ammonitter. Blandt devonernes hvirveldyr blomstrede fisk, og brusk- og knoglefisk, samt lungefisk og lobefinner, erstattede de pansrede. I slutningen af perioden dukker de første padder op, som først levede i vand.
I mellemdevonen dukkede de første skove af bregner, mosser og padderok op på land, som var beboet af orme og talrige leddyr (tusindbenede, edderkopper, skorpioner, vingeløse insekter). I slutningen af devonen dukkede de første gymnospermer op. Udviklingen af jord af planter førte til et fald i forvitring og øget jorddannelse. Konsolidering af jordbund førte til dannelsen af flodkanaler.
I Carbon periode landet var repræsenteret af to kontinenter adskilt af et hav, og klimaet blev mærkbart varmere og vådere. I slutningen af perioden var der en lille hævning af landet, og klimaet ændrede sig til et mere kontinentalt. Havene var domineret af foraminifer, koraller, pighuder, brusk- og benfisk, og ferskvandsområder var beboet af toskallede bløddyr, krebsdyr og forskellige padder. Midt i Carbon opstod der små insektædende krybdyr, og blandt insekter dukkede vingede (kakerlakker, guldsmede) op.
Troperne var præget af sumpede skove domineret af kæmpe padderok, køllemoser og bregner, hvis døde rester efterfølgende dannede kulforekomster. I midten af perioden i den tempererede zone, takket være deres uafhængighed af vand under befrugtningsprocessen og tilstedeværelsen af frø, begyndte spredningen af gymnospermer.
Perm periode var kendetegnet ved sammensmeltningen af alle kontinenter til et enkelt superkontinent Pangea, havenes tilbagetrækning og styrkelsen af det kontinentale klima i en sådan grad, at der dannedes ørkener i det indre af Pangea. I slutningen af perioden forsvandt træbregner, padderok og mosser næsten på land, og tørkeresistente gymnospermer indtog en dominerende stilling. På trods af at store padder stadig fortsatte med at eksistere, opstod forskellige grupper af krybdyr, herunder store planteædere og rovdyr. I slutningen af Perm skete den største udryddelsesbegivenhed i livets historie, da mange grupper af koraller, trilobitter, de fleste blæksprutter, fisk (primært brusk- og lappefinnede fisk) og padder forsvandt. Marine fauna mistede 40-50% af familierne og omkring 70% af slægterne.
Udvikling af liv i mesozoikum. Den mesozoiske æra varede omkring 165 millioner år og var præget af stigende land, intens bjergbygning og et fald i klimaets fugtighed. Den er opdelt i tre perioder: Trias, Jura og Kridt.
Først Trias periode Klimaet var tørt, men senere blev det på grund af stigende vandstand vådere. Blandt planterne var gymnospermer, bregner og padderok fremherskende, men de træagtige former af sporer døde næsten helt ud. Nogle koraller, ammonitter, nye grupper af foraminifer, toskallede og pighuder nåede høj udvikling, mens diversiteten af bruskfisk faldt, og grupper af benfisk ændrede sig også. Krybdyrene, der dominerede landet, begyndte at mestre vandmiljøet, som ichthyosaurer og plesiosaurer. Af krybdyrene i Trias har krokodiller, tuataria og skildpadder overlevet den dag i dag. I slutningen af Trias dukkede dinosaurer, pattedyr og fugle op.
I Jurassic periode Superkontinentet Pangea delte sig i flere mindre. Meget af Jura var meget vådt, og mod slutningen blev klimaet tørrere. Den dominerende gruppe af planter var gymnospermer, hvoraf redwoods overlevede fra den tid. Bløddyr (ammonitter og belemniter, toskallede og snegle), svampe, søpindsvin, brusk- og benfisk blomstrede i havene. Store padder døde næsten fuldstændigt ud i juraperioden, men moderne grupper af padder (hale- og haleløse) og squamater (firben og slanger) dukkede op, og mangfoldigheden af pattedyr steg. Ved udgangen af perioden dukkede mulige forfædre til de første fugle også op - Archaeopteryx. Alle økosystemer var dog domineret af krybdyr - ikthyosaurer og plesiosaurer, dinosaurer og flyveøgler - pterosaurer.
Kridt periode fik sit navn på grund af dannelsen af kridt i den tids sedimentære bjergarter. Over hele Jorden, bortset fra polarområderne, var der et vedvarende varmt og fugtigt klima. I denne periode opstod angiospermer og blev udbredte og fortrængte gymnospermer, hvilket førte til en kraftig stigning i mangfoldigheden af insekter. I havene dukkede udover bløddyr, benfisk og plesiosaurer et stort antal foraminifer op igen, hvis skaller dannede kridtaflejringer, og dinosaurer dominerede på land. Fugle, der var bedre tilpasset luften, begyndte gradvist at fortrænge flyvende dinosaurer.
I slutningen af perioden indtraf en global udryddelsesbegivenhed, som resulterede i forsvinden af ammonitter, belemniter, dinosaurer, pterosaurer og havøgler, gamle grupper af fugle samt nogle gymnospermer. Generelt forsvandt omkring 16 % af familierne og 50 % af dyreslægterne fra Jordens overflade. Den sene Kridt-krisen er blevet tilskrevet nedslaget af en stor meteorit i Den Mexicanske Golf, men det var højst sandsynligt ikke den eneste årsag til globale forandringer. Under den efterfølgende afkøling overlevede kun små krybdyr og varmblodede pattedyr.
Udvikling af livet i cenozoikum. Den cenozoiske æra begyndte for omkring 66 millioner år siden og fortsætter til i dag. Det er karakteriseret ved dominansen af insekter, fugle, pattedyr og angiospermer. Cenozoikum er opdelt i tre perioder - Palæogen, Neogen og Antropocæn - hvoraf sidstnævnte er den korteste i Jordens historie.
I det tidlige og mellemste palæogene miljø forblev klimaet varmt og fugtigt, mod slutningen af perioden blev det køligere og tørrere. Angiospermer blev den dominerende plantegruppe, men hvis stedsegrønne skove dominerede i begyndelsen af perioden, så dukkede der i slutningen mange løvskove op, og stepper blev dannet i tørre zoner.
Blandt fisk indtog benfisk en dominerende stilling, og antallet af bruskagtige arter er, på trods af deres mærkbare rolle i saltvandsområder, ubetydeligt. På landjorden har kun skællende krybdyr, krokodiller og skildpadder overlevet, mens pattedyr har besat de fleste af deres økologiske nicher. I midten af perioden dukkede hovedordenerne af pattedyr op, herunder insektædere, kødædere, pinnipeds, hvaler, hovdyr og primater. Isolationen af kontinenterne gjorde faunaen og floraen mere geografisk forskelligartet: Sydamerika og Australien blev centre for udviklingen af pungdyr, og andre kontinenter - for placentale pattedyr.
Neogen periode. I neogenet fik jordens overflade sit moderne udseende. Klimaet blev køligere og tørrere. I Neogenet var alle ordener af moderne pattedyr allerede dannet, og i de afrikanske ligklæder opstod Hominid-familien og Human-slægten. Ved udgangen af perioden spredte nåleskove sig i polarområderne på kontinenterne, tundraer dukkede op, og korn besatte de tempererede stepper.
Kvartær periode(antropocæn) er karakteriseret ved periodiske ændringer af istider og opvarmninger. Under istiden blev høje breddegrader dækket af gletsjere, havniveauet faldt kraftigt, og tropiske og subtropiske zoner blev indsnævret. I områderne tæt på gletsjerne blev der etableret et koldt og tørt klima, som bidrog til dannelsen af kulderesistente dyregrupper – mammutter, kæmpehjorte, huleløver osv. Faldet i verdenshavets niveau, der fulgte med glaciationsprocessen førte til dannelsen af landbroer mellem Asien og Nordamerika, Europa og de britiske øer osv. Dyrevandringer førte på den ene side til gensidig berigelse af floraer og faunaer, og på den anden side til fortrængning af relikter fra rumvæsener, for eksempel pungdyr og hovdyr i Sydamerika. Disse processer påvirkede dog ikke Australien, som forblev isoleret.
Generelt har periodiske klimaændringer ført til dannelsen af ekstremt rigelig artsdiversitet, karakteristisk for den nuværende fase af biosfærens evolution, og også påvirket menneskelig evolution. Under antropocæn spredte flere arter af menneskeslægten sig fra Afrika til Eurasien. For omkring 200 tusinde år siden i Afrika opstod arten Homo sapiens, som efter en lang periode med eksistens i Afrika for omkring 70 tusind år siden kom ind i Eurasien og for omkring 35-40 tusinde år siden - til Amerika. Efter en periode med sameksistens med nært beslægtede arter fortrængte den dem og spredte sig over hele kloden. For omkring 10 tusind år siden begyndte menneskelig økonomisk aktivitet i moderat varme områder af kloden at påvirke både planetens udseende (pløjning af jorder, afbrænding af skove, overgræsning af græsgange, ørkendannelse osv.) og dyre- og planteverdenen pga. til reduktion af levesteder deres levested og udryddelse, og den menneskeskabte faktor kom i spil.
Menneskelig oprindelse. Mennesket som art, dets plads i den organiske verdens system. Hypoteser om menneskelig oprindelse. Drivkræfter og stadier af menneskelig udvikling. Menneskeracer, deres genetiske slægtskab. Menneskets biosociale natur. Socialt og naturligt miljø, menneskelig tilpasning til det
Menneskelig oprindelse
For bare 100 år siden troede det overvældende flertal af mennesker på planeten ikke engang, at mennesker kunne nedstamme fra så "lav-respektable" dyr som aber. I en diskussion med en af forsvarerne af Darwins evolutionsteori spurgte professor Thomas Huxley, hans ivrige modstander, biskop af Oxford Samuel Wilberforce, som støttede sig på religiøse dogmer, ham endda, om han betragtede sig selv som relateret til abers forfædre gennem sin bedstefar eller bedstemor. .
Men tanker om evolutionær oprindelse blev udtrykt af oldtidens filosoffer, og den store svenske taksonom C. Linnaeus gav i det 18. århundrede, baseret på en række karakteristika, et artsnavn til mennesket Homo sapiens L.(Homo sapiens) og klassificerede ham, sammen med aberne, i samme orden - primater. J. B. Lamarck støttede C. Linnaeus og mente, at mennesket endda havde fælles forfædre med moderne aber, men på et tidspunkt i sin historie nedstammede han fra træet, hvilket var en af årsagerne til menneskets fremkomst som art.
Charles Darwin ignorerede heller ikke dette spørgsmål, og i 70'erne af det 19. århundrede udgav han værkerne "The Origin of Man and Sexual Selection" og "On the Expression of Emotions in Animals and Man", hvori han leverede lige så overbevisende beviser for menneskers og abers fælles oprindelse, end den tyske forsker E. Haeckel ("Skabelsens naturhistorie", 1868; "Anthropogenesis, or the History of the Origin of Man", 1874), der endda udarbejdede en genealogi over dyreriget . Disse undersøgelser vedrørte dog kun den biologiske side af dannelsen af mennesket som art, mens de sociale aspekter blev afsløret af den historiske materialismes klassiker - den tyske filosof F. Engels.
I øjeblikket studeres oprindelsen og udviklingen af mennesker som en biologisk art, såvel som mangfoldigheden af moderne menneskelige befolkninger og mønstrene for deres interaktion, af videnskaben antropologi.
Mennesket som art, dets plads i den organiske verdens system
Homo sapiens ( Homo sapiens) som en biologisk art tilhører dyreriget, underriget af flercellede organismer. Tilstedeværelsen af en notokord, gællespalter i svælget, et neuralrør og bilateral symmetri under embryonal udvikling gør det muligt at klassificere det som en chordat, mens udviklingen af rygsøjlen, tilstedeværelsen af to par lemmer og placeringen af hjertet på den ventrale side af kroppen angiver dets forhold til andre repræsentanter for hvirveldyrsubtypen.
Fodring af ungerne med mælk udskilt af mælkekirtlerne, varmblodighed, et firekammer hjerte, tilstedeværelsen af hår på kroppens overflade, syv hvirvler i halshvirvelsøjlen, forhallen i munden, alveolære tænder og udskiftningen af mælketænder med permanente er tegn på klassen af pattedyr, og den intrauterine udvikling af embryoet og dets forbindelse med moderens krop gennem moderkagen - en underklasse af placenta.
Mere specifikke træk, såsom at gribe om lemmer med en modsatrettet tommelfinger og fingernegle, udvikling af kravebenene, øjne rettet fremad, en stigning i størrelsen af kraniet og hjernen, samt tilstedeværelsen af alle grupper af tænder (fortænder, hjørnetænder og kindtænder) efterlader ingen tvivl om, at hans plads er i ordenen af primater.
Den betydelige udvikling af hjernen og ansigtsmusklerne samt tændernes strukturelle træk gør det muligt at klassificere mennesker som medlemmer af underordenen af højere primater eller aber.
Fraværet af en hale, tilstedeværelsen af krumninger af rygsøjlen, udviklingen af de cerebrale halvkugler i forhjernen, dækket af en cortex med talrige riller og viklinger, tilstedeværelsen af en overlæbe og sparsom hårgrænse giver grund til at placere den blandt repræsentanter for menneskeaberfamilien eller menneskeaber.
Men selv fra de mest organiserede aber er mennesker kendetegnet ved en kraftig stigning i hjernevolumen, oprejst holdning, bredt bækken, udstående hage, artikuleret tale og tilstedeværelsen af 46 kromosomer i karyotypen og bestemmer dens tilhørsforhold til slægten Human.
Brugen af de øvre lemmer til arbejde, fremstilling af redskaber, abstrakt tænkning, kollektiv aktivitet og udvikling baseret på mere sociale end biologiske love er de specifikke kendetegn ved Homo sapiens.
Alle moderne mennesker tilhører én art - Homo sapiens ( Homo sapiens), og underarter H. sapiens sapiens. Denne art er en samling af populationer, der producerer frugtbart afkom, når de krydses. På trods af den ret betydelige mangfoldighed af morfofysiologiske karakteristika er de ikke beviser for en højere eller lavere grad af organisering af visse grupper af mennesker - de er alle på samme udviklingsniveau.
I vores tid er et tilstrækkeligt antal videnskabelige fakta allerede blevet indsamlet af hensyn til dannelsen af mennesket som en art i evolutionsprocessen - antropogenese. Det specifikke forløb af antropogenese er endnu ikke fuldt ud forstået, men takket være nye palæontologiske fund og moderne forskningsmetoder kan vi håbe, at et klart billede dukker op hurtigt nok.
Hypoteser om menneskelig oprindelse
Hvis vi ikke tager hensyn til hypoteserne om menneskets guddommelige skabelse og dets indtrængen fra andre planeter, der ikke er relateret til biologiens område, så sporer alle mere eller mindre konsistente hypoteser om menneskets oprindelse ham tilbage til fælles forfædre med moderne primater.
Så, hypotese om menneskelig oprindelse fra den gamle tropiske primater tarsier, eller tarsial hypotese, formuleret af den engelske biolog F. Wood Jones i 1929, er baseret på ligheden mellem menneskets kropsproportioner og den tarsier, hårgrænsens træk, afkortningen af ansigtsdelen af kraniet på sidstnævnte osv. Men , er forskellene i disse organismers struktur og vitale aktivitet så store, at de ikke har opnået universel anerkendelse.
Mennesker har endda for mange ligheder med aber. Ud over de anatomiske og morfologiske træk, der allerede er nævnt ovenfor, bør opmærksomheden derfor rettes mod deres postembryonale udvikling. For eksempel har små chimpanser meget sparsommere hår, forholdet mellem hjernevolumen og kropsvolumen er meget større, og evnen til at bevæge sig på bagbenene er noget bredere end hos voksne. Selv puberteten hos højere primater forekommer meget senere end hos repræsentanter for andre ordener af pattedyr med lignende kropsstørrelser.
Cytogenetiske undersøgelser afslørede, at et af de menneskelige kromosomer blev dannet som et resultat af sammensmeltningen af kromosomer af to forskellige par, der er til stede i karyotypen af menneskeaber, og dette forklarer forskellen i antallet af deres kromosomer (i mennesker 2n = 46, og hos menneskeaber 2n = 48 ), og er også et andet bevis på disse organismers forhold.
Ligheden mellem mennesker og aber er også meget høj ifølge molekylær biokemiske data, da mennesker og chimpanser har de samme proteiner som AB0- og Rh-blodgrupperne, mange enzymer, og aminosyresekvenserne i hæmoglobinkæderne har kun 1,6% forskelle, hvorimod med andre aber er dette en uoverensstemmelse noget mere. Og på det genetiske niveau er forskellene i nukleotidsekvenserne i DNA mellem disse to organismer mindre end 1 %. Hvis vi tager højde for den gennemsnitlige udviklingshastighed af sådanne proteiner i beslægtede grupper af organismer, kan vi fastslå, at menneskelige forfædre adskilte sig fra andre grupper af primater for omkring 6-8 millioner år siden.
Abernes adfærd minder på mange måder om menneskelig adfærd, da de lever i grupper, hvor sociale roller er tydeligt fordelt. Fælles forsvar, gensidig bistand og jagt er ikke de eneste mål for at skabe en gruppe, da aberne inden for den oplever kærlighed til hinanden, udtrykker det på alle mulige måder og reagerer følelsesmæssigt på forskellige stimuli. Derudover er der i grupper erfaringsudveksling mellem enkeltpersoner.
Således findes lighederne mellem mennesker og andre primater, især menneskeaber, på forskellige niveauer af biologisk organisation, og forskellene mellem mennesker som art bestemmes i høj grad af denne gruppe pattedyrs egenskaber.
Gruppen af hypoteser, der ikke sætter spørgsmålstegn ved menneskers oprindelse fra fælles forfædre med moderne aber, omfatter hypoteserne om polycentrisme og monocentrisme.
Udgangsposition polycentrisme hypoteser er fremkomsten og den parallelle udvikling af den moderne menneskelige art i flere regioner på kloden fra forskellige former for oldtidens eller endda oldtidens menneske, men dette er i modstrid med de grundlæggende bestemmelser i den syntetiske evolutionsteori.
Hypoteser om det moderne menneskes enkeltoprindelse postulerer tværtimod menneskets fremkomst på ét sted, men divergerer med hensyn til, hvor dette skete. Så, hypotese om ekstratropisk oprindelse af mennesker er baseret på, at kun de barske klimatiske forhold på de høje breddegrader i Eurasien kunne bidrage til "humanisering" af aber. Det blev understøttet af opdagelsen på Yakutias territorium af steder, der går tilbage til den antikke palæolitikum - Diring-kulturen, men det blev efterfølgende fastslået, at alderen på disse fund ikke er 1,8-3,2 millioner år, men 260-370 tusind år. Denne hypotese er således heller ikke tilstrækkeligt bekræftet.
Den største mængde beviser er i øjeblikket blevet indsamlet til fordel for hypoteser om menneskers afrikanske oprindelse, men det er ikke uden sine mangler, som en omfattende bred monocentrisme-hypotese, der kombinerer argumenterne for hypoteserne om polycentrisme og monocentrisme.
Drivkræfter og stadier i menneskets evolution
I modsætning til andre repræsentanter for dyreverdenen blev mennesket i sin udvikling ikke kun udsat for biologiske evolutionsfaktorer, men også for sociale faktorer, hvilket bidrog til fremkomsten af en art af kvalitativt nye skabninger med biosociale egenskaber. Sociale faktorer bestemte et gennembrud til et fundamentalt nyt adaptivt miljø, som gav enorme fordele for overlevelse af menneskelige befolkninger og kraftigt accelererede tempoet i dets udvikling.
Biologiske evolutionsfaktorer, der spiller en vis rolle i menneskeskabt den dag i dag, er arvelig variabilitet, såvel som strømmen af gener, der leverer det primære materiale til naturlig udvælgelse. Samtidig har isolation, befolkningsbølger og genetisk drift næsten fuldstændig mistet deres betydning som følge af videnskabelige og teknologiske fremskridt. Dette giver grund for nogle videnskabsmænd til at tro, at i fremtiden vil selv minimale forskelle mellem repræsentanter for forskellige racer forsvinde på grund af deres blanding.
Da skiftende miljøforhold tvang menneskelige forfædre til at stige ned fra træerne til det åbne rum og bevæge sig på to lemmer, blev de frigjorte øvre lemmer brugt af dem til at transportere mad og børn, samt til at fremstille og bruge værktøj. Et sådant værktøj kan dog kun laves, hvis der er en klar idé om det endelige resultat - billedet af objektet, hvorfor abstrakt tænkning også udviklede sig. Det er velkendt, at komplekse bevægelser og tænkningsprocessen er nødvendige for udviklingen af visse områder af hjernebarken, hvilket skete i evolutionsprocessen. Det er imidlertid umuligt at arve sådan viden og færdigheder; de kan kun overføres fra et individ til et andet i løbet af sidstnævntes liv, hvilket resulterede i skabelsen af en særlig form for kommunikation - artikuleret tale.
Således omfatter evolutionens sociale faktorer menneskelig arbejdsaktivitet, abstrakt tænkning og artikuleret tale. Man bør ikke kassere manifestationerne af altruisme hos den primitive mand, der tog sig af børn, kvinder og ældre.
Menneskets arbejdsaktivitet påvirkede ikke kun dets udseende, men gjorde det også muligt i begyndelsen delvist at lette eksistensbetingelserne gennem brug af ild, fremstilling af tøj, opførelse af boliger og senere aktivt at ændre dem gennem skovrydning, pløjejord osv. I vor tid har ukontrolleret økonomisk aktivitet sat menneskeheden under truslen om en global katastrofe som følge af jorderosion, udtørring af ferskvandsforekomster og ødelæggelse af ozonskærmen, som igen kan øges presset af biologiske faktorer af evolution.
Dryopithecus, der levede for omkring 24 millioner år siden, var højst sandsynligt den fælles forfader til mennesker og aber. På trods af at han klatrede i træer og løb på alle fire lemmer, kunne han bevæge sig på to ben og bære mad i hænderne. Den fuldstændige adskillelse af menneskeaberne og linjen, der fører til mennesker, fandt sted for omkring 5-8 millioner år siden.

Australopithecus. Slægten stammer tilsyneladende fra Dryopithecus Ardipithecus, som blev dannet for over 4 millioner år siden i Afrikas savanner som følge af afkøling og tilbagetrækning af skove, hvilket tvang disse aber til at skifte til at gå på bagbenene. Dette lille dyr gav tilsyneladende anledning til en ret stor slægt Australopithecus("sydlig abe").
Australopithecus dukkede op for omkring 4 millioner år siden og levede i afrikanske savanner og tørre skove, hvor fordelene ved tobenet bevægelse kunne mærkes fuldt ud. Fra Australopithecus kom to grene - store planteædere med kraftige kæber Paranthropus og mindre og mindre specialiserede Mennesker. Over en periode udviklede disse to slægter sig parallelt, hvilket især viste sig i en stigning i hjernens volumen og komplikationen af de anvendte værktøjer. Funktioner i vores slægt er fremstilling af stenværktøjer (Paranthropus brugte kun knogle) og en relativt stor hjerne.
De første repræsentanter for den menneskelige slægt dukkede op for omkring 2,4 millioner år siden. De tilhørte arten af dygtige mennesker (Homo habilis) og var korte væsner (ca. 1,5 m) med et hjernevolumen på ca. 670 cm 3. De brugte rå stenværktøj. Tilsyneladende havde repræsentanter for denne art veludviklede ansigtsudtryk og rudimentær tale. Homo habilis forlod den historiske scene for omkring 1,5 millioner år siden og gav anledning til den næste art - en straight mand.
Mand oprejst (H. erectus) som en biologisk art dannet i Afrika for omkring 1,6 millioner år siden og eksisterede i 1,5 millioner år, og slog sig hurtigt ned over store territorier i Asien og Europa. En repræsentant for denne art fra øen Java blev engang beskrevet som Pithecanthropus("abemand"), opdaget i Kina, blev navngivet Sinanthropa, mens deres europæiske "kollega" er det Heidelberg mand.
Alle disse former kaldes også arkantroper(af gamle mennesker). Den oprejste mand var kendetegnet ved en lav pande, store pandekamme og en skrånende hage; hans hjernevolumen var 900-1200 cm 3. Overkroppen og lemmerne på en oprettet mand lignede det moderne menneskes. Uden tvivl brugte repræsentanter for denne slægt ild og lavede tveæggede økser. Som nyere opdagelser har vist, mestrede denne art endda navigation, for dens efterkommere blev fundet på fjerntliggende øer.
Paleoantrop. For omkring 200 tusind år siden opstod Heidelberg-mennesket Neandertalermand (H. neandertalensis), som der henvises til palæoantroper(gamle mennesker), der levede i Europa og Vestasien for mellem 200 og 28 tusinde år siden, herunder i istiderne. De var stærke, fysisk ret stærke og modstandsdygtige mennesker med en stor hjernekapacitet (endnu større end moderne menneskers). De havde artikuleret tale, lavede komplekse redskaber og tøj, begravede deres døde og havde måske endda nogle rudimenter af kunst. Neandertalerne var ikke forfædrene til Homo sapiens; denne gruppe udviklede sig parallelt. Deres udryddelse er forbundet med mammutfaunaens forsvinden efter den sidste istid, og er måske også et resultat af konkurrencedygtig fortrængning fra vores art.
Det ældste fund af en repræsentant homo sapiens (Homo sapiens) Den er 195 tusind år gammel og kommer fra Afrika. Mest sandsynligt er forfædrene til moderne mennesker ikke neandertalere, men en form for arkantroper, såsom Heidelberg-mennesket.
Neoantrop. For omkring 60 tusind år siden, som et resultat af ukendte begivenheder, uddøde vores art næsten, så alle de følgende mennesker er efterkommere af en lille gruppe, der kun talte et par dusin individer. Efter at have overvundet denne krise, begyndte vores art at sprede sig over hele Afrika og Eurasien. Den adskiller sig fra andre arter i sin slankere kropsbygning, højere reproduktionshastighed, aggressivitet og selvfølgelig den mest komplekse og mest fleksible adfærd. Moderne mennesker, der beboede Europa for 40 tusind år siden, kaldes Cro-Magnons og henvise til neoantroper(til moderne mennesker). De var biologisk ikke anderledes end moderne mennesker: højde 170-180 cm, hjernevolumen omkring 1600 cm 3. Cro-Magnonerne udviklede kunst og religion, de tæmmede mange arter af vilde dyr og dyrkede mange arter af planter. Moderne mennesker nedstammer fra Cro-Magnons.
Menneskeracer, deres genetiske slægtskab
Efterhånden som menneskeheden slog sig ned på planeten, opstod der visse forskelle mellem forskellige grupper af mennesker med hensyn til hudfarve, ansigtstræk, hårtype samt hyppigheden af forekomsten af visse biokemiske træk. Sættet af sådanne arvelige egenskaber karakteriserer en gruppe individer af samme art, hvor forskellene mellem dem er mindre signifikante end underarten - race.
Studiet og klassificeringen af racer kompliceres af manglen på klare grænser mellem dem. Hele moderne menneskehed tilhører én art, inden for hvilken der skelnes mellem tre store racer: Australo-Negroid (sort), Caucasoid (hvid) og Mongoloid (gul). Hver af dem er opdelt i små racer. Forskelle mellem racer kommer ned til træk ved hudfarve, hår, næseform, læber osv.
Aussie-negroid, eller ækvatorial race karakteriseret ved mørk hudfarve, bølget eller krøllet hår, en bred og let fremspringende næse, tværgående næsebor, tykke læber og en række kranietræk. kaukasisk, eller eurasisk race karakteriseret ved lys eller mørk hud, glat eller bølget blødt hår, god udvikling af mandligt ansigtshår (skæg og overskæg), smal udstående næse, tynde læber og en række kranietræk. Mongoloid(asiatisk-amerikansk) race karakteriseret ved mørk eller lys hud, ofte groft hår, gennemsnitlig bredde af næse og læber, fladtrykt ansigt, kraftigt fremspring af kindbenene, relativt stor ansigtsstørrelse, mærkbar udvikling af det "tredje øjenlåg".
Disse tre racer adskiller sig også i deres afvikling. Før æraen med europæisk kolonisering var den australske-negroide race udbredt i den gamle verden syd for Krebsens vendekreds; Kaukasisk race - i Europa, Nordafrika, Vestasien og det nordlige Indien; Mongoloid race - i Sydøst-, Nord-, Central- og Østasien, Indonesien, Nord- og Sydamerika.
Forskellene mellem racer vedrører dog kun mindre karakteristika, der har adaptiv betydning. Således forbrændes negroiders hud af en ti gange højere dosis ultraviolet stråling end huden på kaukasiere, men kaukasiere lider mindre af rakitis på høje breddegrader, hvor der kan være mangel på ultraviolet stråling, der er nødvendig for dannelsen af D-vitamin.
Tidligere søgte nogle mennesker at bevise en af racernes overlegenhed for at opnå moralsk overlegenhed over andre. Det er nu klart, at racekarakteristika kun afspejler de forskellige historiske veje for grupper af mennesker, men er på ingen måde forbundet med den ene eller anden gruppes fordele eller biologiske tilbagestående. Menneskeracer er mindre klart definerede end andre dyrs underarter og racer, og kan på ingen måde sammenlignes f.eks. med racer af husdyr (som er resultatet af målrettet udvælgelse). Som biomedicinsk forskning viser, afhænger konsekvenserne af ægteskab mellem racer af mandens og kvindens individuelle karakteristika og ikke af deres race. Derfor er ethvert forbud mod ægteskaber mellem racer eller visse overtro uvidenskabelige og umenneskelige.
Mere specifikke end racer er grupper af mennesker nationaliteter- historisk dannede sproglige, territoriale, økonomiske og kulturelle fællesskaber af mennesker. Befolkningen i et bestemt land udgør dets folk. Med samspillet mellem mange nationaliteter kan en nation opstå i en nation. Nu er der ingen "rene" racer på Jorden, og enhver tilstrækkelig stor nation er repræsenteret af mennesker, der tilhører forskellige racer.
Menneskets biosociale natur
Uden tvivl skal mennesker som biologisk art opleve pres fra evolutionære faktorer som mutagenese, befolkningsbølger og isolation. Men efterhånden som det menneskelige samfund udvikler sig, svækkes nogle af dem, mens andre tværtimod styrkes, da der på planeten, fanget af globaliseringsprocesserne, næsten ikke er nogen isolerede menneskelige befolkninger tilbage, hvor indavl finder sted, og antallet af befolkningerne selv er ikke udsat for skarpe udsving. Følgelig spiller den drivende faktor for evolutionen - naturlig udvælgelse - takket være medicinens succeser ikke længere den samme rolle i menneskelige populationer som i populationer af andre organismer.
Desværre fører et svækkelse af selektionspres til en stigning i hyppigheden af arvelige sygdomme i populationer. For eksempel i industrialiserede lande lider op til 5 % af befolkningen af farveblindhed, mens dette tal i mindre udviklede lande er op til 2 %. De negative konsekvenser af dette fænomen kan overvindes takket være forebyggende foranstaltninger og fremskridt inden for videnskabsområder som genterapi.
Dette betyder dog ikke, at menneskets evolution er afsluttet, da naturlig udvælgelse fortsætter med at virke og eliminerer for eksempel kønsceller og individer med ugunstige kombinationer af gener selv i de proembryonale og embryonale perioder af ontogenese, samt resistens over for patogener af forskellige sygdomme. Derudover leveres materialet til naturlig udvælgelse ikke kun af mutationsprocessen, men også af akkumulering af viden, evnen til at lære, opfattelsen af kultur og andre egenskaber, der kan overføres fra person til person. I modsætning til genetisk information overføres erfaringer akkumuleret under processen med individuel udvikling både fra forældre til afkom og i den modsatte retning. Og der opstår allerede konkurrence mellem samfund, der adskiller sig kulturelt. Denne form for evolution, unik for mennesker, kaldes kulturel, eller social udvikling.
Kulturel evolution udelukker dog ikke biologisk evolution, da det kun blev muligt på grund af dannelsen af den menneskelige hjerne, og den menneskelige biologi i sig selv er i øjeblikket bestemt af kulturel evolution, da i fravær af samfund og mangfoldighed af bevægelser, gør visse zoner ikke dannes i hjernen.
En person har således en biosocial natur, som efterlader et aftryk på manifestationen af biologiske, herunder genetiske, love, der styrer hans individuelle og evolutionære udvikling.
Socialt og naturligt miljø, menneskelig tilpasning til det
Under socialt miljø forstå først og fremmest de sociale, materielle og åndelige betingelser for hans eksistens og aktivitet omkring en person. Ud over det økonomiske system, sociale relationer, social bevidsthed og kultur, omfatter det også det umiddelbare miljø for en person - familie, arbejds- og elevgrupper samt andre grupper. Miljøet har på den ene side en afgørende indflydelse på dannelsen og udviklingen af personlighed, og på den anden side ændrer det sig selv under påvirkning af en person, hvilket medfører nye ændringer i mennesker mv.
Tilpasning af individer eller deres grupper til det sociale miljø for at realisere deres egne behov, interesser, livsmål og omfatter tilpasning til betingelserne og arten af studier, arbejde, interpersonelle relationer, økologiske og kulturelle omgivelser, fritids- og hverdagslivsforhold, samt som deres aktive forandring for at tilfredsstille dine behov. At ændre sig selv, sine motiver, værdier, behov, adfærd osv. spiller også en stor rolle heri.
Informationsmængder og følelsesmæssige oplevelser i det moderne samfund er ofte hovedårsagen til stress, som kan overvindes ved hjælp af tydelig selvorganisering, fysisk træning og autotræning. I nogle særligt alvorlige tilfælde kræves et besøg hos en psykoterapeut. Et forsøg på at finde glemsel af disse problemer gennem overspisning, rygning, indtagelse af alkohol og andre dårlige vaner fører ikke til det ønskede resultat, men forværrer kun kroppens tilstand.
Det naturlige miljø har ikke mindre indflydelse på mennesker, på trods af at mennesker har forsøgt at skabe et behageligt kunstigt miljø for sig selv i omkring 10 tusinde år. At stige til en betydelig højde på grund af et fald i iltkoncentrationen i luften fører således til en stigning i antallet af røde blodlegemer i blodet, øget vejrtrækning og puls, og langvarig eksponering for den åbne sol bidrager til øget hudpigmentering - garvning. Imidlertid passer de anførte ændringer ind i reaktionens norm og nedarves ikke. Folk, der har levet under sådanne forhold i lang tid, kan dog have nogle tilpasninger. Blandt nordlige folk har næsebihulerne således et meget større volumen til opvarmning af luften, og størrelsen af de udragende dele af kroppen falder for at reducere varmetabet. Afrikanere har mørkere hudfarve og krøllet hår, fordi pigmentet melanin beskytter kroppens organer mod indtrængen af skadelige ultraviolette stråler, og hårhætten har varmeisolerende egenskaber. Europæernes lyse øjne er en tilpasning til en mere akut opfattelse af visuel information i skumringen og i tåge, og øjnenes mongoloide form er resultatet af naturlig udvælgelse for påvirkning af vind og støvstorme.
Disse ændringer kræver århundreder og årtusinder, men livet i et civiliseret samfund medfører nogle ændringer. Et fald i fysisk aktivitet fører således til et lettere skelet, et fald i dets styrke og et fald i muskelmasse. Lav mobilitet, overskydende kaloriefattig mad, stress fører til en stigning i antallet af overvægtige mennesker, og tilstrækkelig proteinernæring og fortsatte dagslystimer ved hjælp af kunstig belysning bidrager til acceleration - accelereret vækst og pubertet, og en stigning i kropsstørrelse .
Sadi Carnot, søn af krigsministeren under Napoleon Bonaparte og onkel til den fremtidige præsident for Den Franske Republik, der analyserede konsekvenserne af Frankrigs erobringskrige i begyndelsen af det 19. århundrede, kom til den konklusion, at en af grundene for Frankrigs nederlag var dets bagud i brugen af energi.
I de sidste to hundrede år har folk brugt mange kræfter på at prøve at forstå dette koncept. I dag er det blevet klart, at menneskers velvære er forbundet med energiforbrug. For eksempel er mængden af et lands bruttonationalprodukt næsten proportional med mængden af forbrugt energi.
Termodynamik er studiet af, hvordan energi omdannes i fysiske kroppe. Den besvarer spørgsmålene: hvordan får man varme, hvordan man gør processen med at få varme overskuelig, hvordan man sparer den og hvordan man bedst bruger den. For at besvare dem introducerede videnskabsmænd definitioner og formulerede de grundlæggende love eller principper for termodynamikken.
Energi(fra græsk energi- handling, aktivitet) defineres som et generelt kvantitativt mål for forskellige former for bevægelse af stof.
I naturen svarer enhver fysisk proces til en eller anden type energi: mekanisk, kemisk, termisk, gravitationel, elektromagnetisk, nuklear og så videre. Med andre ord udtrykker det en krops eller et systems evne til at udføre arbejde.
Fundamental forskning inden for informationsteori førte til konceptet informationsenergi, eller energi af informationspåvirkning , som et kvantitativt mål for ændringer i mængden af information.
Termodynamikkens første lov er kort formuleret som følger: "Energi er bevaret." Den dukker ikke op af ingenting og forsvinder ikke sporløst; det kan kun ændre sig fra en form til en anden. Vi skylder Lord Kelvin (Williams Thomson) og Rudolf Clausis denne fundamentale opdagelse fra 50'erne i det forrige århundrede. Kelvin, som en religiøs mand, mente, at Skaberen på tidspunktet for verdens skabelse gav ham en forsyning af energi, og at denne guddommelige gave ville eksistere for evigt.
Termodynamikkens anden lov fastslår tilstedeværelsen i naturen af en fundamental asymmetri, det vil sige ensrettetheden af alle spontane energispredningsprocesser, der forekommer i den: varme kroppe afkøles over tid, men kolde bliver ikke varme af sig selv, en roterende toppen stopper til sidst, men en hviletop vil ikke begynde at rotere spontant.
Det vil sige, at fordelingen af tilgængelig energi ændres irreversibelt. Denne egenskab ved stof afspejles princippet om entropi. Hvis der er et udtryk entropi karakteriserer graden af uorden i naturen, hvilket betyder, at der skal være et udtryk, der definerer graden af orden. Entalpi og er en funktion, der evaluerer graden af orden af systemet (lokal heterogenitet).
Naturligt forekommende processer i naturen er ledsaget af en stigning i entropi (uorden).
På en overraskende måde afspejles disse ideer i universets model. Det udvider sig, og energi spredes i rummet. Kontinuerlig og global naturlig ødelæggelse er hovedegenskaben for den materielle verden omkring os. I denne sammenhæng spiller det, der står i det menneskelige sind bag billedet af Mephistopheles, en vigtig rolle i naturen, hvilket giver processer dissipation(fra latin dissipatio- spredning). På samme tid, på grund af lokale inhomogeniteter i fordelingen af stof i naturen, akkumuleres det i lokale zoner i universet. Sådan dannes galakser, stjernebilleder, solsystemer og planeter.
Hvis vi ikke bevidst opretholder orden i huset, så vil uorden, uanset vores ønske, tiltage i det. Omvendt er målrettet kreativ aktivitet ledsaget af et fald i entropi (uorden) i en bestemt lokal zone af rummet. Sandt nok er dette kun muligt ved at skabe endnu større kaos uden for denne zone: Sådan opstår lossepladser og andet affald - biprodukter af kreativ aktivitet.
Fra praksis ved vi, at kreativ aktivitet er umulig uden brug af fornuft eller en eller anden algoritme født af naturen. I dette tilfælde er fødslen af en krystal i en amorf klippe et eksempel på målrettet kreativ aktivitet i naturen baseret på en algoritme genereret af naturen selv.
Enhver levende organisme er også et eksempel på et begrænset rum, hvor entropien ikke øges i løbet af livet. I modsætning til en krystal har en organisme egenskaben af selvregulering, det vil sige evnen til at opretholde et konstant indre miljø, når ydre og indre forstyrrelser ændrer sig. Denne egenskab med ikke at øge entropien i løbet af livet er en af de vigtigste, der adskiller et element af levende natur fra livløs natur.
At opretholde orden er således forbundet med en selvreguleringsalgoritme, bevidsthed. Ud fra Goethes allegorier kan vi sige, at Skaberen er ansvarlig for processer forbundet med livet, som ikke øger entropien. Han ser ud til at bruge stof som et eksperimentelt materiale til at skabe og udvikle algoritmer, der understøtter forskellige liv på jagt efter deres perfektion.
Da det i praksis ofte er svært direkte at måle mængden af energi i en krop, har folk lært at vurdere den i forhold til mængden af arbejde, der kan udføres med den. Fra skolens fysikkursus ved vi, at arbejde vurderes som et produkt styrke, påført kroppen, ganget med den afstand, kroppen har bevæget sig over. På den ene eller anden måde er begrebet kraft i naturen forbundet med typen af interaktion.
Som vi allerede har bemærket, er alle kendte kræfter baseret på fire hovedtyper af interaktion mellem fysiske legemer (tabel 1.1.1). Når man overvejer dynamiske processer, er det vigtigt at huske, at enhver bevægelse af en materiel krop kan dekomponeres i tre hovedprocesser: translationel, oscillerende og roterende.
Bevægelse og kraft har retninger, der ikke altid er sammenfaldende. Denne uoverensstemmelse tages i betragtning af en koefficient, der er proportional med vinklen mellem kraftretningen og bevægelsesretningen. Nu kan vi forklare, at et tidsur, der hænger ubevægeligt på væggen, forbruger energi på grund af svingningen af pendulet og andre dele. Toppens akse er praktisk talt ubevægelig i forhold til gulvet. Snurretoppen har dog rotationsenergi og bruger den på at overvinde luftmodstand og friktion af akslen på gulvet.
For at estimere hastigheden af energiforbruget eller dets erhvervelse bruges begrebet strøm som ændringshastigheden i den samlede energi i et bestemt system.
Produktet af kraft og tid giver således energi. Effekten måles i watt. På et sekund er en energilagringsenhed med en effekt på én watt i stand til at levere én joule (J) energi til systemet. En hestekræfter er defineret som et energiforbrug på 746 J pr.
Så lad os liste de grundlæggende love, der er nødvendige for at forstå materialet i bogen, der ligger til grund for de processer, der forekommer i universet (makroverden):
1. Lov om hierarkisk lighed.
2. Rytmeloven.
3. Lov om bevarelse af energi.
4. Entropiens lov.
Økologiens love— generelle mønstre og principper for samspil mellem det menneskelige samfund og det naturlige miljø.
Betydningen af disse love er at regulere arten og retningen af menneskelig aktivitet i økosystemer på forskellige niveauer. Blandt økologilovene formuleret af forskellige forfattere er de mest berømte de fire love-aforismer fra den amerikanske miljøforsker Barry Commoner (1974):
- "alt er forbundet med alt"(loven om den universelle forbindelse af ting og fænomener i naturen);
- “alt skal et sted hen”(lov om bevarelse af stofmasse);
- "intet kommer gratis"(om prisen på udvikling);
- "naturen ved bedst"(om hovedkriteriet for evolutionær selektion).
Fra loven om den universelle sammenhæng mellem ting og fænomener i naturen("alt er forbundet med alt"), følger flere konsekvenser:
- lov om store tal - den kombinerede virkning af en lang række tilfældige faktorer fører til et resultat, der er næsten uafhængigt af tilfældigheder, dvs. have en systemisk karakter. Myriader af bakterier i jord, vand og levende organismers kroppe skaber således et særligt, relativt stabilt mikrobiologisk miljø, der er nødvendigt for alle levende tings normale eksistens. Eller et andet eksempel: den tilfældige opførsel af et stort antal molekyler i et bestemt volumen gas bestemmer helt bestemte værdier af temperatur og tryk;
- Le Chatelier (Brun) princip - Når en ydre påvirkning tager systemet ud af en tilstand af stabil ligevægt, skifter denne ligevægt i den retning, hvor effekten af den ydre påvirkning aftager. På det biologiske plan realiseres det i form af økosystemernes evne til selvregulering;
- optimalitetsloven— ethvert system fungerer med den største effektivitet inden for visse rumlige tidsmæssige grænser, der er karakteristiske for det;
- enhver systemisk forandring i naturen har en direkte eller indirekte indvirkning på mennesker - fra individets tilstand til komplekse sociale relationer.
Fra lov om bevarelse af stofmasse("alt skal gå et sted hen"), følger mindst to postulater af praktisk betydning:
Barry Commoner skriver: "... det globale økosystem er en enkelt helhed, hvor intet kan vindes eller tabes, og som ikke kan være genstand for en samlet forbedring; alt, hvad der er blevet udvundet af det ved menneskeligt arbejde, skal erstattes. Betaling af denne regning kan ikke undgås; det kan kun udskydes. Den nuværende miljøkrise betyder, at forsinkelsen er meget lang.”
Princip "naturen ved bedst" bestemmer først og fremmest, hvad der må og ikke må foregå i biosfæren. Alt i naturen – fra simple molekyler til mennesker – har været igennem en hård konkurrence om retten til at eksistere. I øjeblikket er planeten kun beboet af 1/1000 arter af planter og dyr, der er testet af evolution. Hovedkriteriet for denne evolutionære udvælgelse er inklusion i den globale biotiske cyklus, udfyldning af alle økologiske nicher. Ethvert stof produceret af organismer skal have et enzym, der nedbryder det, og alle henfaldsprodukter skal geninddrages i kredsløbet. Med enhver biologisk art, der overtrådte denne lov, skiltes evolutionen før eller siden. Den menneskelige industrielle civilisation krænker groft lukketheden af det biotiske kredsløb på globalt plan, som ikke kan forblive ustraffet. I denne kritiske situation skal der findes et kompromis, som kun en person med intelligens og ønske om dette kan gøre.
Ud over formuleringerne af Barry Commoner har moderne økologer udledt en anden økologilov - "der er ikke nok til alle" (loven om begrænsede ressourcer). Det er indlysende, at massen af næringsstoffer for alle former for liv på Jorden er begrænset og begrænset. Det er ikke nok for alle repræsentanter for den organiske verden, der optræder i biosfæren, derfor kan en betydelig stigning i antallet og massen af alle organismer på global skala kun ske på bekostning af et fald i antallet og massen af andre. Modsigelsen mellem reproduktionshastigheden og begrænsede føderessourcer i forhold til klodens befolkning blev først bemærket af den engelske økonom T.R. Malthus (1798), som hermed forsøgte at retfærdiggøre den sociale konkurrences uundgåelighed. Til gengæld lånte Charles Darwin fra Malthus begrebet "kamp for tilværelsen" for at forklare mekanismen for naturlig udvælgelse i den levende natur.
Lov om begrænsede ressourcer- kilden til alle former for konkurrence, rivalisering og antagonisme i naturen og desværre i samfundet. Og uanset hvor meget man anser klassekamp, racisme og interetniske konflikter for at være rene sociale fænomener, er de alle forankret i intraspecifik konkurrence, som nogle gange tager meget mere brutale former end hos dyr.
Den væsentlige forskel er, at i naturen, som følge af konkurrence, overlever de bedste, men i det menneskelige samfund er det på ingen måde tilfældet.
En generaliseret klassificering af miljølove blev præsenteret af den berømte sovjetiske videnskabsmand N.F. Reimers. De får følgende formuleringer:
- lov om socioøkologisk balance(behovet for at opretholde en balance mellem pres på miljøet og genopretning af dette miljø, både naturligt og kunstigt);
- princippet om kulturel udviklingsledelse(pålægge restriktioner for omfattende udvikling under hensyntagen til miljørestriktioner);
- reglen om socioøkologisk substitution(behovet for at identificere måder at erstatte menneskelige behov);
- lov om socioøkologisk irreversibilitet(umulighed for at vende den evolutionære bevægelse, fra komplekse former til simplere);
- noosfærens lov Vernadsky (det uundgåelige af transformationen af biosfæren under indflydelse af tanker og menneskeligt arbejde til noosfæren - geosfæren, hvor fornuften bliver dominerende i udviklingen af "menneske-natur"-systemet).
Overholdelse af disse love er mulig, forudsat at menneskeheden forstår sin rolle i mekanismen til at opretholde stabiliteten i biosfæren. Det er kendt, at i evolutionsprocessen bevares kun de arter, der er i stand til at sikre livets og miljøets bæredygtighed. Kun mennesket, ved hjælp af sindets kraft, kan lede den videre udvikling af biosfæren langs vejen til at bevare den vilde natur, bevare civilisationen og menneskeheden, skabe et mere retfærdigt socialt system, bevæge sig fra en krigsfilosofi til en filosofi om fred og fred. partnerskab, kærlighed og respekt for fremtidige generationer. Alle disse er komponenter i et nyt biosfære-verdensbillede, som burde blive universelt.
Økologiens love og principper
Minimumsloven
I 1840 Yu Liebig konstateret, at høsten ofte ikke er begrænset af de næringsstoffer, der er nødvendige i store mængder, men af dem, der er nødvendige i små mængder, men som der er lidt af i jorden. Den lov, han formulerede, sagde: "Det stof, der er i sit minimum, kontrollerer høsten og bestemmer størrelsen og stabiliteten af sidstnævnte over tid." Efterfølgende blev en række andre faktorer, såsom temperatur, tilført næringsstofferne. Virkningen af denne lov er begrænset af to principper. Liebigs første lov gælder strengt taget kun under stationære forhold. En mere præcis formulering: "i stationær tilstand vil det begrænsende stof være det stof, hvis tilgængelige mængder er tættest på det krævede minimum." Det andet princip vedrører samspillet mellem faktorer. Høj koncentration eller tilgængelighed af et bestemt stof kan ændre indtaget af et minimumsnæringsstof. Følgende lov er formuleret i selve økologien og generaliserer minimumsloven.
Toleranceloven
Denne lov er formuleret som følger: fraværet eller umuligheden af udvikling af et økosystem bestemmes ikke kun af en mangel, men også af et overskud af nogen af faktorerne (varme, lys, vand). Derfor er organismer karakteriseret ved både et økologisk minimum og et maksimum. For meget af det gode er også dårligt. Området mellem de to værdier udgør de tolerancegrænser, inden for hvilke kroppen reagerer normalt på miljøpåvirkninger. Toleranceloven foreslået W. Shelford i 1913. Der kan formuleres en række forslag, der supplerer den.
- Organismer kan have en bred vifte af tolerance for én faktor og et snævert interval for en anden.
- Organismer med en bred vifte af tolerance over for alle faktorer er normalt de mest udbredte.
- Hvis betingelserne for én miljøfaktor ikke er optimale for en art, kan toleranceområdet for andre miljøfaktorer indsnævres.
- I naturen befinder organismer sig meget ofte i forhold, der ikke svarer til den optimale værdi af en bestemt faktor bestemt i laboratoriet.
- Ynglesæsonen er normalt kritisk; I denne periode viser mange miljøfaktorer sig ofte at være begrænsende.
Levende organismer ændrer miljøforhold for at svække fysiske faktorers begrænsende indflydelse. Arter med en bred geografisk udbredelse danner lokalt tilpassede populationer kaldet økotyper. Deres optimale og tolerancegrænser svarer til lokale forhold.
Generelt koncept for begrænsende faktorer
De vigtigste faktorer på land er lys, temperatur og vand (nedbør), mens de i havet er lys, temperatur og saltholdighed. Disse fysiske eksistensbetingelser kan at være begrænsende og gunstigt påvirkende. Alle miljøfaktorer afhænger af hinanden og virker i samspil. Andre begrænsende faktorer omfatter atmosfæriske gasser (kuldioxid, oxygen) og biogene salte. Ved formuleringen af "minimumsloven" havde Liebig den begrænsende virkning af vitale kemiske grundstoffer, der er til stede i miljøet i små og varierende mængder, i tankerne. De kaldes sporstoffer og omfatter jern, kobber, zink, bor, silicium, molybdæn, klor, vanadium, kobolt, jod, natrium. Mange mikroelementer, som vitaminer, fungerer som katalysatorer. Fosfor, kalium, calcium, svovl, magnesium, som kræves af organismer i store mængder, kaldes makroelementer. En vigtig begrænsende faktor i moderne forhold er miljøforurening. Den væsentligste begrænsende faktor, iflg Yu. Odumu, - størrelser og kvalitet oikosa", eller vores" naturligt kloster" ikke kun antallet af kalorier, der kan presses ud af jorden. Landskabet er ikke kun et lager for forsyninger, men også det hus, vi bor i. ”Målet bør være at bevare mindst en tredjedel af al jord som beskyttet friareal. Det betyder, at en tredjedel af alle vores levesteder bør være nationale eller lokale parker, naturreservater, grønne områder, vildmarksområder osv. Det område, som en person har brug for, varierer ifølge forskellige skøn fra 1 til 5 hektar. Det andet af disse tal overstiger det område, der i øjeblikket tegner sig for én indbygger på Jorden.
Befolkningstætheden nærmer sig én person pr. 2 hektar jord. Kun 24 % af jorden er egnet til landbrug. Selvom kun 0,12 hektar kan give nok kalorier til at støtte én person, er der brug for omkring 0,6 hektar per person til en nærende kost med masser af kød, frugt og grønt. Derudover kræves der ca. 0,4 hektar mere til produktion af forskellige typer fiber (papir, træ, bomuld) og yderligere 0,2 hektar til veje, lufthavne, bygninger mv. Deraf begrebet "den gyldne milliard", ifølge hvilken den optimale befolkningsstørrelse er 1 milliard mennesker, og derfor er der allerede omkring 5 milliarder "ekstra mennesker". For første gang i sin historie blev mennesket konfronteret med ekstreme snarere end lokale restriktioner. At overvinde begrænsende faktorer kræver enorme udgifter til stof og energi. Fordobling af udbyttet kræver en tidobling af mængden af gødning, pesticider og kraft (dyr eller maskiner). Befolkningsstørrelsen er også en begrænsende faktor.
Lov om konkurrencemæssig udelukkelse
Denne lov er formuleret som følger: to arter, der besætter den samme økologiske niche, kan ikke eksistere sammen på ét sted i uendeligt lang tid.
Hvilken art der vinder afhænger af ydre forhold. Under lignende forhold kan enhver vinde. En vigtig faktor for sejren er befolkningstilvæksten. En arts manglende evne til at konkurrere biotisk fører til dens fortrængning og behovet for at tilpasse sig vanskeligere forhold og faktorer.
Loven om konkurrencemæssig udelukkelse kan også fungere i det menneskelige samfund. Det særlige ved dens handling på nuværende tidspunkt er, at civilisationer ikke kan spredes. De har ingen steder at forlade deres territorium, for i biosfæren er der intet frirum til bosættelse, og der er intet overskud af ressourcer, hvilket fører til en intensivering af kampen med alle de deraf følgende konsekvenser. Vi kan tale om miljørivalisering mellem lande og endda miljøkrige eller krige på grund af miljømæssige årsager. På et tidspunkt retfærdiggjorde Hitler Nazitysklands aggressive politik med kampen om boligareal. Ressourcer af olie, kul mv. og så var de vigtige. De har endnu større vægt i det 21. århundrede. Derudover var der behov for territorier til bortskaffelse af radioaktivt og andet affald. Krige – varme og kolde – får en miljødimension. Mange begivenheder i moderne historie, såsom USSR's sammenbrud, opfattes på en ny måde, hvis de ses fra et miljømæssigt perspektiv. En civilisation kan ikke kun erobre en anden, men bruge den til selviske formål fra et miljømæssigt synspunkt. Dette vil være økologisk kolonialisme. Sådan hænger politiske, sociale og miljømæssige spørgsmål sammen.
Grundlæggende lov om økologi
En af økologiens vigtigste resultater var opdagelsen af, at ikke kun organismer og arter udvikler sig, men også. Rækkefølgen af fællesskaber, der afløser hinanden i et givet område, kaldes arvefølge. Succession sker som følge af ændringer i det fysiske miljø under påvirkning af fællesskabet, dvs. kontrolleret af ham.
Høj produktivitet giver lav pålidelighed - en anden formulering af den grundlæggende lov om økologi, hvorfra følgende regel følger: "Optimal effektivitet er altid mindre end maksimum." Diversitet er ifølge økologiens grundlov direkte relateret til bæredygtighed. Det vides dog endnu ikke, i hvilket omfang denne sammenhæng er kausal.
Nogle andre love og principper, der er vigtige for økologi.
lov om emergens: helheden har altid særlige egenskaber, som dens del ikke har.
Lov om nødvendig variation: et system kan ikke bestå af absolut identiske elementer, men kan have en hierarkisk organisation og integrerende niveauer.
Lov om evolutionens irreversibilitet: en organisme (population, art) kan ikke vende tilbage til den tidligere tilstand opnået i rækken af sine forfædre.
Lov om organisatorisk kompleksitet: den historiske udvikling af levende organismer fører til komplikation af deres organisation gennem differentiering af organer og funktioner.
Biogenetisk lov(E. Haeckel): en organismes ontogeni er en kort gentagelse af en given arts fylogeni, dvs. Individet i sin udvikling gentager i forkortet form sin arts historiske udvikling.
Loven om ujævn udvikling af dele af systemet: systemer på samme hierarkiniveau udvikler sig ikke strengt synkront, mens nogle når et højere udviklingstrin, andre forbliver i en mindre udviklet tilstand. Denne lov er direkte relateret til loven om nødvendig mangfoldighed.
Loven om bevarelse af liv: liv kan kun eksistere i bevægelsesprocessen af en strøm af stoffer, energi og information gennem en levende krop.
Princippet om at opretholde orden(J. Prigogine): i åbne systemer stiger entropien ikke, men falder indtil en minimum konstant værdi er nået, altid større end nul.
Le Chatelier-Brown princippet: når en ydre påvirkning tager systemet ud af en tilstand af stabil ligevægt, forskydes denne ligevægt i den retning, hvori effekten af den ydre påvirkning svækkes.
Energispareprincip(L. Onsager): givet sandsynligheden for, at en proces udvikler sig i et bestemt sæt af retninger tilladt af termodynamikkens principper, realiseres den, der giver et minimum af energispredning.
Loven om energi og informationsmaksimering: den bedste chance for selvopretholdelse har det system, der mest bidrager til modtagelse, produktion og effektiv brug af energi og information; det maksimale udbud af et stof garanterer ikke systemets succes i konkurrencen.
Loven om systemudvikling på bekostning af miljøet: Ethvert system kan kun udvikles gennem brug af materiale, energi og informationskapacitet i dets miljø; Helt isoleret selvudvikling er umulig.
Schrödingers styre"om at fodre" organismen med negativ entropi: Organismens orden er højere end miljøet, og organismen giver dette miljø mere uorden, end den modtager. Denne regel korrelerer med Prigogines princip om at opretholde orden.
Regel for at accelerere evolution: Med stigende kompleksitet af organiseringen af biologiske systemer falder varigheden af eksistensen af en art i gennemsnit, og udviklingshastigheden stiger. Den gennemsnitlige levetid for en fugleart er 2 millioner år, og en pattedyrart er 800 tusind år. Antallet af uddøde arter af fugle og pattedyr i forhold til hele deres antal er stort.
Loven om relativ uafhængighed af tilpasning: høj tilpasningsevne til en af miljøfaktorerne giver ikke samme grad af tilpasning til andre livsbetingelser (tværtimod kan det begrænse disse muligheder på grund af organismers fysiologiske og morfologiske egenskaber).
Princippet om mindste befolkningsstørrelse: Der er en minimumspopulationsstørrelse, under hvilken dens antal ikke kan falde.
Reglen for repræsentation af en slægt ved én art: under homogene forhold og i et begrænset område er en taksonomisk slægt normalt kun repræsenteret af én art. Tilsyneladende skyldes dette nærheden af de økologiske nicher af arter af samme slægt.
Loven om udtømning af levende stof i dets økoncentrationer(G.F. Hilmi): "Et individuelt system, der opererer i et miljø med et organisationsniveau, der er lavere end selve systemets niveau, er dødsdømt: gradvist at miste sin struktur, vil systemet efter nogen tid opløses i miljøet." Dette fører til en vigtig konklusion for menneskelige miljøaktiviteter: den kunstige bevarelse af små økosystemer (i et begrænset område, for eksempel et naturreservat) fører til deres gradvise ødelæggelse og sikrer ikke bevarelsen af arter og samfund.
Loven om energipyramiden(R. Lindeman): i gennemsnit bevæger omkring 10% af den energi, der modtages på det tidligere niveau, sig fra et trofisk niveau i den økologiske pyramide til et andet, højere niveau. Det omvendte flow fra højere til lavere niveauer er meget svagere – ikke mere end 0,5-0,25 %, og derfor er der ingen grund til at tale om energikredsløbet i biocenosen.
Reglen om obligatorisk påfyldning af økologiske nicher: en tom økologisk niche er altid og altid naturligt fyldt ("naturen afskyr et vakuum").
Princippet om økosystemdannelse: organismers langsigtede eksistens er kun mulig inden for rammerne af økologiske systemer, hvor deres komponenter og elementer komplementerer hinanden og er gensidigt tilpasset. Fra disse miljølove og -principper følger nogle konklusioner, der er gyldige for "menneske - naturligt miljø"-systemet. De hører til typen af lov om mangfoldighedsbegrænsning, dvs. pålægge begrænsninger på menneskelige aktiviteter for at transformere naturen.
Boomerang lov: alt, der udvindes fra biosfæren ved menneskeligt arbejde, skal returneres til den.
Loven om biosfærens uundværlighed: Biosfæren kan ikke erstattes af et kunstigt miljø, ligesom der ikke kan skabes nye typer liv. Mennesket kan ikke bygge en evighedsmaskine, mens biosfæren praktisk talt er en "perpetual motion-maskine".
Lov om shagreen læder: Det globale oprindelige naturressourcepotentiale udtømmes løbende i løbet af den historiske udvikling. Dette følger af, at der i øjeblikket ikke er nogen grundlæggende nye ressourcer, der kunne dukke op. Hver person har brug for 200 tons faste stoffer om året for at leve, som han omdanner til et nyttigt produkt ved hjælp af 800 tons vand og et gennemsnit på 1000 W energi. Mennesket tager alt dette fra det, der allerede er tilgængeligt i naturen.
Princippet om begivenhedens fjernhed: efterkommere vil finde på noget for at forhindre mulige negative konsekvenser. Spørgsmålet om, hvor langt økologiens love kan overføres til forholdet mellem mennesket og miljøet, står åbent, da mennesket adskiller sig fra alle andre arter. For eksempel, i de fleste arter, falder hastigheden af befolkningstilvækst, når befolkningstætheden stiger; Hos mennesker accelererer befolkningstilvæksten i dette tilfælde tværtimod. Nogle reguleringsmekanismer i naturen er fraværende hos mennesker, og dette kan tjene som en yderligere årsag til teknologisk optimisme for nogle, og for miljøpessimister, indikerer faren for en sådan katastrofe, som er umulig for nogen anden art.
I forhold til miljøledelse og miljøbeskyttelsesaktiviteter hører man ofte om behovet for at tage hensyn til den naturlige udviklings love. Mennesket, der kun har realiseret sin rolle i biosfæren som en af de mange arter, der udgør dens mangfoldighed, som en del af den, må ligesom alle andre adlyde naturens love. Samtidig ligger homo sapiens styrke ikke i at omstrukturere naturen ved at vise dens magt, men i korrekt at forstå lovene for dens udvikling og følge dem. Lovene for naturens udvikling er love af en højere orden for mennesker i sammenligning med lovene for samfundets udvikling. Det er objektive love. I kraft af deres handling og takket være dem dukkede mennesket op og kan eksistere. Samfundets love er skrevet af mennesket for dets egen sociale, politiske og økonomiske bekvemmelighed, organisering og tilvejebringelse af samfundslivet.
Kendskab til og overholdelse af den naturlige udviklings love i menneskets og samfundets aktiviteter er af afgørende betydning og vurderes som et imperativ. Lovene om naturens udvikling, manifesteret i samspillet mellem samfund og natur, skaber det naturvidenskabelige og filosofiske grundlag for forskellige aktiviteter inden for miljøforvaltning og miljøbeskyttelse, herunder på retsområdet. At tage hensyn til naturlovene ved planlægning og gennemførelse af miljøskadelige aktiviteter og deres overholdelse bør tjene som hovedkriteriet for sådanne aktiviteters miljømæssige gyldighed og tilladelighed. Deres viden og hensyn er særlig vigtig ved gennemførelsen af sådanne lovmæssige tiltag til naturbeskyttelse som standardisering af maksimalt tilladte påvirkninger af naturen, vurdering af planlagte aktiviteters indvirkning på miljøet, miljøvurdering, planlægning af nosv. Lovene af naturudvikling bør også tages i betragtning ved udarbejdelse af lovforslag om miljøbeskyttelse. At sikre, at naturlovene tages i betragtning og overholdes, når der træffes økonomiske, ledelsesmæssige og andre miljømæssigt væsentlige beslutninger, er en af betingelserne, det metodiske grundlag for at overvinde miljøkrisen.
Lad os overveje nogle grundlæggende love for udviklingen af naturen som fortolket af en af de førende miljøforskere i Rusland, professor N.F. Reimers.
Loven om biogen migration af atomer (V.I. Vernadsky). Migrationen af kemiske grundstoffer på jordens overflade og i biosfæren som helhed udføres enten med direkte deltagelse af levende stof (biogen migration) eller sker i et miljø, hvis geokemiske træk (O 2, C0 2, H 2 osv.) .) er bestemt af levende stof - både den, der i øjeblikket bebor biosfæren, og den, der har været på Jorden gennem geologisk historie.
I henhold til denne lov, som har vigtig teoretisk og praktisk betydning, forstå de generelle kemiske processer, der har fundet sted og finder sted på jordens overflade, i atmosfæren og i dybet af litosfæren og vandet, der er beboet af organismer, samt geologiske lag sammensat af organismers tidligere aktiviteter, er umuligt uden at tage biotiske og biogene faktorer i betragtning, herunder evolutionære. Da mennesker primært påvirker biosfæren og dens levende befolkning, ændrer de dermed betingelserne for biogen migration af atomer, hvilket skaber forudsætningerne for endnu dybere kemiske ændringer i det historiske perspektiv. Processen kan således blive selvudviklende, uafhængig af menneskelig lyst og praktisk talt, på globalt plan, ukontrollerbar. Derfor er et af de mest presserende behov at bevare jordens levende overflade i en relativt uændret tilstand. Den samme lov bestemmer også behovet for at tage hensyn til indvirkningen på biota i alle naturomdannelsesprojekter. I disse tilfælde sker der regionale og lokale ændringer i kemiske processer, hvilket fører til større fejl i miljøforringelsen - ørkendannelse.
Lov om intern dynamisk ligevægt. Stof, energi, information og dynamiske kvaliteter af individuelle naturlige systemer og deres hierarki er så indbyrdes forbundne, at enhver ændring i en af disse indikatorer forårsager ledsagende funktionelle strukturelle kvalitative og kvantitative ændringer, der bevarer den samlede sum af materiale-energi, information og dynamiske kvaliteter af systemer, hvor disse ændringer forekommer, eller i deres hierarki.
En række konsekvenser af denne lov er empirisk fastlagt:
- a) enhver ændring i miljøet (stoffer, energi, information, økosystemers dynamiske kvaliteter) fører uundgåeligt til udviklingen af naturlige kædereaktioner med det formål at neutralisere ændringen eller dannelsen af nye naturlige systemer, hvis dannelse, med væsentlige ændringer i miljøet, kan blive irreversibelt;
- b) samspillet mellem materiale-energi miljøkomponenter (energi, gasser, væsker osv.), information og dynamiske kvaliteter af naturlige systemer er ikke kvantitativt lineært, dvs. en svag effekt eller ændring i en af indikatorerne kan forårsage stærke afvigelser i andre (og i hele systemet som helhed);
- c) ændringerne i store økosystemer er relativt irreversible. Når de passerer gennem hierarkiet fra bunden og op - fra indvirkningens sted til biosfæren som helhed, ændrer de globale processer og overfører dem derved til et nyt evolutionært niveau;
- d) enhver lokal transformation af naturen forårsager reaktioner i den globale helhed af biosfæren og i dens største divisioner, hvilket fører til den relative konstanthed af det økologiske og økonomiske potentiale ("Trishkins kaftanregel"), hvis forøgelse kun er mulig gennem en markant stigning i energiinvesteringer.
Loven om intern dynamisk ligevægt er en af nøglebestemmelserne i miljøledelse. Mens miljøændringer er svage og foretages over et relativt lille område, er de enten begrænset til et bestemt sted eller "fader ud" i kæden af økosystemhierarki. Men så snart ændringer når betydelige niveauer for store økosystemer, for eksempel, sker de på skalaen af store vandløbsoplande, de fører til betydelige forskydninger i disse enorme naturlige formationer, og gennem dem, i overensstemmelse med følge b), i hele jordens biosfære.
"Alt eller intet"-loven (X. Boulich). Svage påvirkninger forårsager muligvis ikke reaktioner i det naturlige system, før de efter at have akkumuleret fører til udviklingen af en voldsom dynamisk proces. Loven er nyttig i miljøprognoser.
Lov om konstanthed (V.I. Vernadsky). Mængden af levende stof i naturen (for en given geologisk periode) er en konstant. Enhver ændring i mængden af levende stof i en af regionerne i biosfæren medfører uundgåeligt en ændring af samme størrelse i enhver region, men med det modsatte fortegn. Polære ændringer kan bruges i naturforvaltningsprocesser, men det skal tages i betragtning, at der ikke altid sker tilstrækkelig udskiftning. Typisk erstattes højtudviklede arter og økosystemer af andre på et relativt evolutionært lavere niveau (store organismer med mindre), og former, der er gavnlige for mennesker, erstattes af mindre nyttige, neutrale eller endda skadelige.
Minimumsloven (J. Liebig). En organismes udholdenhed bestemmes af det svageste led i kæden af dens miljøbehov, dvs. livsmuligheder begrænses af miljøfaktorer, hvis mængde og kvalitet er tæt på det minimum, som organismen eller økosystemet kræver; deres yderligere reduktion fører til organismens død eller ødelæggelse af økosystemet.
Loven om begrænsede naturressourcer. Alle jordens naturressourcer (og betingelser) er begrænsede. Loven er baseret på det faktum, at da planeten er en naturligt begrænset helhed, kan uendelige dele ikke eksistere på den. Som følge heraf opstod kategorien "uudtømmelige" naturressourcer på grund af en misforståelse.
Loven om udvikling af et naturligt system på bekostning af dets miljø. Ethvert naturligt system kan kun udvikle sig gennem brug af materialer, energi og informationskapaciteter i sit miljø. Helt isoleret selvudvikling er umulig. Denne samme lov har ekstremt vigtig teoretisk og praktisk betydning på grund af dens vigtigste konsekvenser:
- a) absolut affaldsfri produktion er umulig;
- b) ethvert mere højt organiseret biotisk system (f.eks. en levende art), der bruger og modificerer det levende miljø, udgør en potentiel trussel mod mindre organiserede systemer;
- c) Jordens biosfære som et system udvikler sig ikke kun på bekostning af planetens ressourcer, men indirekte på bekostning og under kontrolpåvirkning af rumsystemer (primært det Solar).
Ifølge den første konsekvens kan vi kun regne med lav-spild produktion. Derfor bør den første fase i udviklingen af teknologier være deres lave ressourceintensitet (både ved input og output - økonomi og ubetydelige emissioner), anden fase vil være skabelsen af cyklisk produktion (spild af nogle kan være råmaterialer for andre ) og den tredje - organisering af rimelig bortskaffelse af uundgåelige rester og neutralisering af uaftageligt energiaffald. Ideen om, at biosfæren fungerer efter princippet om ikke-affald, er fejlagtig, da den altid akkumulerer stoffer fjernet fra det biologiske kredsløb, som danner sedimentære bjergarter.
Ifølge lovens anden konsekvens kræver menneskelig påvirkning af naturen foranstaltninger til at neutralisere disse påvirkninger, da de kan være ødelæggende for resten af naturen og ifølge reglen om overholdelse af miljøforhold med organismens genetiske forudbestemmelse , true personen selv. I denne henseende er naturbeskyttelse en af de obligatoriske komponenter i den socioøkonomiske udvikling af et højt udviklet samfund.
Den tredje implikation af loven er af særlig betydning for langsigtede prognoser. Det skal tages i betragtning, når man overvejer alle processer, der finder sted på Jorden.
Lov (mønster) for at reducere miljøintensiteten af færdige produkter. Det specifikke indhold af naturlige stoffer i den gennemsnitlige enhed af socialt produkt har historisk set været støt faldende. Det betyder ikke, at der indgår færre naturlige stoffer i produktionsprocessen. Tværtimod stiger dens mængde - op til 95-98% af det naturlige stof, der forbruges i produktionen, smides væk. Imidlertid indeholder de endelige lignende produkter af social produktion i dag sandsynligvis i gennemsnit mindre naturligt stof end i en fjern fortid. Dette forklares ved miniaturisering af produkter, udskiftning af naturlige materialer og produkter med syntetiske såvel som andre fænomener. Da der endnu ikke er foretaget nøjagtige beregninger (de er metodologisk vanskelige), har denne lov karakter af en ekspertkonklusion.
Loven om at reducere energieffektiviteten af miljøledelse. Med fremskridt i historisk tid, når man opnår nyttige produkter fra naturlige systemer, bliver der i gennemsnit brugt mere og mere energi per enhed af det.
Energiforbrug pr. person (i kcal/dag) i stenalderen var omkring 4 tusinde, i et landbrugssamfund - 12 tusind, i den industrielle æra - 7 tusind, og i de avancerede udviklede lande i nutiden - 230-250 tusinde ., dvs. 58-62 gange mere end hos fjerne forfædre. Siden begyndelsen af dette århundrede er mængden af energi brugt på en enhed landbrugsprodukter i udviklede lande steget 8-10 gange. Den samlede energieffektivitet i landbrugsproduktionen (forholdet mellem energiinput og energi modtaget fra færdige produkter) i industrialiserede lande er ca. 30 gange lavere end under forhold med primitivt landbrug. I nogle tilfælde fører en tidobling af energiomkostningerne til gødning og markbearbejdning kun til en meget lille (10-15 %) stigning i udbyttet. Dette skyldes behovet for, sideløbende med forbedring af landbrugsteknologien, at tage hensyn til den generelle miljøsituation og de begrænsninger, den pålægger. I begyndelsen af 80'erne. Det specifikke energiforbrug pr. enhed af bruttonationalproduktet (BNP) er blevet reduceret med 15 % som følge af kraftige ei industrialiserede lande. I løbet af det sidste årti er BNI steget her med 20%, og energiforbruget - med kun 2% (dette blev muligt som følge af eliminering af uberettigede energitab).
Lov om aftagende (naturlig) frugtbarhed. På grund af den konstante høst og afbrydelse af naturlige jorddannelsesprocesser såvel som langvarig monokultur, som et resultat af ophobning af giftige stoffer frigivet af planter, sker der et gradvist fald i den naturlige jordfrugtbarhed på dyrkede arealer. Denne proces neutraliseres delvist af akkumulering af biomasse i de underjordiske dele af dyrkede planter, men hovedsageligt ved påføring af gødning (skabelse af kunstig frugtbarhed). Til dato er frugtbarheden gået tabt i en eller anden grad i cirka 50 % af al agerjord i verden (fra 1,5-1,6 til 2 milliarder hektar) med en gennemsnitlig tabsrate i 70'erne. 6,8, i 80'erne - omkring 7 millioner hektar om året. Intensivering af landbruget gør det muligt at opnå stadig større udbytter med mindre menneskelig arbejdskraft og delvist neutralisere effekten af loven om faldende afkast, men samtidig falder produktionens energieffektivitet.
Loven om fysisk og kemisk enhed af levende stof (V.I. Vernadsky). Alt levende stof på Jorden er fysisk og kemisk forenet. En konsekvens følger naturligvis af loven: hvad der er skadeligt for en del af et levende stof kan ikke være ligegyldigt for en anden del af det, eller: hvad der er skadeligt for nogle typer skabninger er skadeligt for andre. Derfor kan fysiske og kemiske agenser, der er dødelige for nogle organismer (f.eks. skadedyrsbekæmpelsesmidler), ikke andet end have en skadelig virkning på andre organismer. Den eneste forskel er artens resistensgrad over for midlet. Da der i enhver stor population altid er individer af forskellig kvalitet, inklusive dem, der er mindre eller mere modstandsdygtige over for fysisk-kemiske påvirkninger, er udvælgelseshastigheden for populationers udholdenhed over for et skadeligt middel direkte proportional med organismernes reproduktionshastighed og hastigheden af generationsskifte. Baseret på dette, med den voksende påvirkning af en fysisk-kemisk faktor, som en organisme med et relativt langsomt generationsskifte er modstandsdygtig overfor, på en mindre stabil, men hurtigere reproducerende art, bliver deres evne til at modstå den pågældende faktor lig. Derfor er langvarig brug af kemiske metoder til bekæmpelse af planteskadedyr og patogener hos mennesker og varmblodede dyr miljømæssigt uacceptabel. Med udvælgelsen af resistente individer af hurtigt reproducerende leddyr skal behandlingsraterne øges. Disse øgede koncentrationer viser sig dog at være ineffektive, men har en alvorlig indvirkning på sundheden for mennesker og hvirveldyr.
Lov om økologisk korrelation. I et økosystem, som i enhver anden integreret naturlig systemdannelse, især i et biotisk samfund, er alle levende arter og abiotiske økologiske komponenter inkluderet i det funktionelt konsistente med hinanden. Tabet af en del af systemet (for eksempel ødelæggelsen af en art) fører uundgåeligt til udelukkelse af alle andre dele af systemet, der er tæt knyttet til denne del af systemet og en funktionel ændring i helheden inden for rammerne af Lov om intern dynamisk ligevægt. Loven om økologisk korrelation er især vigtig for bevarelsen af levende arter, som aldrig forsvinder isoleret, men altid i en sammenkoblet gruppe. Lovens virkning fører til bratte ændringer i miljømæssig bæredygtighed: når tærsklen for ændring i funktionel integritet er nået, sker der et sammenbrud (ofte uventet) - økosystemet mister sin egenskab af pålidelighed. For eksempel kan en multipel stigning i koncentrationen af et forurenende stof ikke føre til katastrofale konsekvenser, men så vil en ubetydelig stigning føre til en katastrofe.
Den berømte amerikanske miljøforsker B. Commoner reducerer økologiens grundlæggende love til følgende: 1) alt er forbundet med alt; 2) alt skal et sted hen; 3) naturen "ved" bedre; 4) intet gives gratis.
Livets habitat udvikler sig som et resultat af konstant udveksling af information baseret på strømmen af energi i den samlede enhed af miljøet og de organismer, der beboer det.
40. Minimumsloven(Liebig): Stoffet til stede i minimum kontrollerer udbyttet, bestemmer dets størrelse og stabilitet over tid.
41. Almindelige love:
"Alt er forbundet med alt";
"Alt skal et sted hen";
"Intet kommer gratis";
"Naturen ved bedst."
42. Loven om det maksimale(Shelford): En organismes opblomstring er begrænset af zoner med maksimum og minimum af visse miljøfaktorer; mellem dem er der en zone med økologisk optimum, inden for hvilken kroppen reagerer normalt på miljøforhold.
43. Biosfærens nedbrydning - Dette er ødelæggelse eller væsentlig forstyrrelse af økologiske forbindelser i naturen, ledsaget af en forringelse af menneskets levevilkår, forårsaget af naturkatastrofer eller menneskets økonomiske aktiviteter, udført uden at tage hensyn til kendskabet til den naturlige udviklings love.
44. Stadier af nedbrydning af biosfæren:
brug af ild (tidlig palæolitikum);
landbrugsudvikling;
industrielle revolution.
økologisk krise.
45. Kilder til biosfærens nedbrydning kan være naturlig (naturlig) og kunstig (antropogen). Naturlig OS-forurening forårsaget af naturlige processer (støvstorme, vulkanisme, skovbrande osv.). Kunstig forurening sammenhænge med emissioner af forskellige forurenende stoffer til miljøet under menneskelige aktiviteter (landbrug, transport, industri osv.)
46. Konsekvenser af biosfærens nedbrydning:
Et mærkbart fald i økosystemets biodiversitet, ødelæggelsen og ødelæggelsen af stadig tilbageværende områder med vild vegetation, den barbariske ødelæggelse af skove og sumpe, en reduktion i antallet af vilde dyr, forsvinden af mange repræsentanter for flora og fauna. Som et resultat af alle disse handlinger, ved midten af det tyvende århundrede, indtrådte den menneskeskabte påvirkning af biosfæren i dens betydning på samme niveau som den naturlige, idet den antog en planetarisk skala. Således er menneskeheden blevet en af de vigtigste geo-økologiske skæbnesvangre faktorer i planetens udvikling.
47. Forurening- enhver introduktion i et bestemt økologisk system (biocenose) af levende eller ikke-levende komponenter, som ikke er karakteristiske for det, enhver ændring, der afbryder eller forstyrrer cirkulations- og metabolismeprocesserne, energistrømme, hvis konsekvens er et fald i produktiviteten eller ødelæggelse af dette system.
48. Vigtigste forurenende stoffer:
kuldioxid (CO 2);
carbonmonoxid (CO);
svovldioxid (SO 2);
nitrogenoxider (NO, NO 2, N 2 O);
tungmetaller og primært kviksølv, bly og cadmium;
radionuklider og andre radioaktive stoffer;
dioxider (chlorcarboner);
faste urenheder (aerosoler): støv, sod, røg;
olie og olieprodukter.
kræftfremkaldende stoffer, især benzopyren;
pesticider;
49. Ifølge aggregeringstilstand Der er 3 typer forurenende stoffer: fast, flydende og gasformig.
50. Efter oprindelse natur, aggregeringstilstand, distributionsskala, forårsagede konsekvenser, grad af toksicitet
51. Af natur Forurenende stoffer er klassificeret i følgende grupper: kemiske, fysiske, biologiske, æstetiske.