Тази статия ще запознае читателя със структурата на най-простите организми, а именно ще се съсредоточи върху структурата на контрактилната вакуола, която изпълнява отделителна (и не само) функция, ще говори за значението на най-простите организми и ще опише начините, по които на тяхното съществуване в околната среда.
Контрактилна вакуола. Концепция
Вакуола (от френски vacuole, от латинската дума vacuus - празен), малки кухини със сферична форма в растителни и животински клетки или едноклетъчни организми. Контрактилните вакуоли са предимно често срещани сред прости организми, които живеят в сладка вода, като протисти като амеба протей и ресничести чехли, които са получили първоначалното си име поради формата на тялото, подобно на формата на подметката на обувка. В допълнение към изброените протозои, идентични структури са открити и в клетките на различни сладководни гъби, които принадлежат към семейство Бадягов.

Структурата на контрактилната вакуола. Характеристиките му
Контрактилната вакуола е мембранна органела, която освобождава излишната течност от цитоплазмата. Локализацията и структурата на този апарат варират при различните микроорганизми. От комплекс от везикуларни или тубулни вакуоли, наречени спонгии, течността навлиза в контрактилната вакуола. Благодарение на постоянната работа на тази система се поддържа стабилен клетъчен обем. Протозоите имат контрактилни вакуоли, които са апарат, който регулира осмотичното налягане и също така служи за отделяне на отпадъчни продукти от тялото. Тялото на протозоите се състои само от една клетка, която от своя страна изпълнява всички необходими жизнени функции. Представители на това подцарство, като ресничести чехли, обикновена амеба и други едноклетъчни организми, притежават всички свойства на независим организъм.
Ролята на простите организми
Клетката изпълнява всички жизненоважни функции: отделяне, дишане, раздразнителност, движение, размножаване, метаболизъм. Протозоите са повсеместни. Най-голям брой видове живеят в морски и сладки води, много обитават влажна почва, могат да заразяват растенията и живеят в телата на многоклетъчни животни и хора. В природата протозоите изпълняват и санитарна роля, участват в кръговрата на веществата и са храна за много животни.

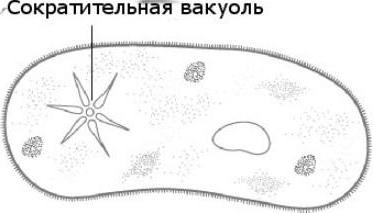
Контрактилна вакуола при амеба вулгарис
Обикновената амеба е представител на класа на коренопогите, за разлика от другите представители, тя няма постоянна форма на тялото. Движението се извършва с помощта на псевдоподи. Сега нека разберем каква функция изпълнява контрактилната вакуола в амеба. Това е регулирането на нивото на осмотичното налягане вътре в клетката. Може да се образува във всяка част на клетката. През външната мембрана водата от околната среда постъпва осмотично. Концентрацията на разтворени вещества в клетката на амебата е по-висока, отколкото в околната среда. Така се създава разлика в налягането вътре в протозойната клетка и извън нея. Функциите на контрактилната вакуола в амеба са вид помпен апарат, който премахва излишната вода от клетката на прост организъм. Amoeba Proteus може да освободи натрупаната течност в околната среда навсякъде по повърхността на тялото.
В допълнение към осморегулаторната, тя изпълнява функцията на дишане в живота, тъй като в резултат на осмозата входящата вода доставя разтворен в нея кислород. Каква друга функция изпълнява контрактилната вакуола? Той също така изпълнява екскреторна функция, а именно метаболитните продукти се отстраняват заедно с водата в околната среда.

Дишане, отделяне, осморегулация в ресничеста чехълка
Тялото на протозоите е покрито с плътна черупка, която има постоянна форма. и водорасли, включително някои протозои. Тялото на ресничките има по-сложна структура от тази на амеба. В чехлевата клетка има две контрактилни вакуоли, разположени отпред и отзад. В това устройство се различават резервоар и няколко малки тубули. Контрактилните вакуоли са постоянно разположени, благодарение на тази структура (от микротубули), на постоянно място в клетката.
Основната функция на контрактилната вакуола в жизнената дейност на този представител на протозоите е осморегулацията; тя също така премахва излишната вода от клетката, която прониква в клетката поради осмоза. Първо, адукторните канали набъбват, след което водата от тях се изпомпва в специален резервоар. Резервоарът се свива, отделя се от захранващите канали и водата се изхвърля през порите. В ресничеста клетка има две контрактилни вакуоли, които от своя страна действат в антифаза. Благодарение на работата на две такива устройства се осигурява непрекъснат процес. В допълнение, водата непрекъснато циркулира поради активността на контрактилните вакуоли. Те се компресират последователно, а честотата на контракциите зависи от температурата на околната среда.
Така при стайна температура (+18 - +20 градуса по Целзий) честотата на контракциите на вакуолите е според някои данни 10-15 секунди. И като се има предвид, че естественото местообитание на чехъла е всяко сладководно тяло със застояла вода и наличието на разлагащи се органични вещества в него, температурата на тази среда се променя с няколко градуса в зависимост от времето на годината и следователно от честотата на контракциите може да достигне 20-25 секунди. За един час контрактилната вакуола на прост организъм е в състояние да освободи големи количества вода от клетката. съизмерими с размера си. В тях се натрупват хранителни вещества, несмлени остатъци от храна, крайни продукти от обмяната на веществата, а също могат да се намерят кислород и азот.
Пречистване на отпадни води с протозои
Влиянието на протозоите върху кръговрата на веществата в природата е от голямо значение. В резервоарите, поради изхвърлянето на отпадъчни води, бактериите се размножават в големи количества. В резултат на това се появяват различни прости организми, които използват тези бактерии като храна и така допринасят за естественото

Заключение
Въпреки простата структура на тези едноклетъчни организми, тялото на които се състои от една клетка, но изпълнява функциите на цял организъм, удивително адаптиран към околната среда. Това може да се наблюдава дори в примера за структурата на контрактилната вакуола. Днес вече е доказано огромното значение на най-простите в природата и тяхното участие в кръговрата на веществата.
КОНТРАКТИВЕН ВАКУОЛ, вижте ВАКУОЛ... Научно-технически енциклопедичен речник
Контрактилна вакуола контрактилна вакуола. Вид вакуола в някои групи протисти, участваща в отстраняването на вода (разтвори) от клетката по време на свиване и в абсорбцията на вода от клетката по време на разширяване, което служи за регулиране на осмотичното налягане.... ... Молекулярна биология и генетика. Тълковен речник.
Устройство на еукариотна клетка. Вакуолата е посочена под номер 10. Вакуолата е едномембранен органел, открит в някои еукариотни клетки и ... Уикипедия
Или разред Heliozoa от клас Sarcodica (q.v.) тип протозои (q.v.) животни. Морфологични свойства. Отличават се със сферично протоплазмено тяло, от което като лъчи излизат във всички посоки тънки, нишковидни, не... .... Енциклопедичен речник F.A. Брокхаус и И.А. Ефрон
Или Ciliata, отряд от клас Ciliates, или ресничести (q.v.), вид протозои (q.v.). ресничести цилати. I (Aspirotricha). Значение на буквите: прах; алвеоларен слой на ектоплазмата; ad.Z адорален ред реснички; b тактилна сета; cl реснички; ... Енциклопедичен речник F.A. Брокхаус и И.А. Ефрон
- (Lobosea), клас от най-просто организираните протозои над класа на коренищата. Лишен от вътрешен скелет и външни черупки. Формата на тялото е променлива, размерите обикновено варират от 20 до 700 микрона, рядко малко повече. Формата и размерът на псевдоподиите са характерни за... ... Биологичен енциклопедичен речник
- (Flagellata s. Mastigophora, виж таблицата. Камшичести, Flagellata) клас протозои (Protozoa). Както всички останали представители от този тип, те имат тяло, състоящо се само от една клетка, представляваща протоплазма и ядро с ядро.... ... Енциклопедичен речник F.A. Брокхаус и И.А. Ефрон
Или протозои. Съдържание на статията: Характеристики и класификация. Исторически очерк. Морфология; протоплазма с включвания (трихоцисти, ядро, контрактилни вакуоли, хроматофори и др.). Корици и скелет. Движение П.; псевдоподии, флагели и... Енциклопедичен речник F.A. Брокхаус и И.А. Ефрон
А. КОНТРАКТИВНИ ВАКУОЛИ
1. Морфология на контрактилните вакуоли
Повечето протозои (с изключение на Споровици)Те имат специален, повече или по-малко строго локализиран, понякога много сложен отделителен апарат, повечето от които имат характер на така наречените контрактилни или пулсиращи вакуоли.
Като общо правило всички сладководни животни имат вакуоли. МастигофораИ Саркодинаи при всички ресничести.
U Мастигофорав по-голямата си част има само 1 контрактилна вакуола, рядко 2 (Phytomastigina).
Размерите на вакуоларния апарат обикновено са правопропорционални на размера на тялото и обратно пропорционални на броя на вакуолите в тялото на животното. Така че, в големите реснички Heterotricha (Stentor, Spirostomum)има само 1 вакуола, но тя (заедно с каналите) достига много големи размери. U Трахелия яйцеклеткадо 30 вакуоли, но те са много малки.
Понякога се забелязва увеличаване на броя на вакуолите, докато животното расте, както при Collinia branchiarumи някои други Астомати.Това умножаване на броя на вакуолите, които Колиния (Anoplophrya)възниква по време на живота на индивида, очевидно се е случило по време на еволюцията на цялата група Астомати(фиг. 124). Всъщност сред тях могат да се идентифицират редица форми с постепенно по-сложен вакуоларен апарат. Да, да Догиелелаима само 1 субтерминално разположена вакуола в рода Анополфрия 1 надлъжен ред от малък брой вакуоли; при раждането БушлиелаИ Моподонтофрияброят на вакуолите в този ред се увеличава значително и в Радиофриявакуолите образуват 2 надлъжни реда. И накрая видът Хаптофрияредица многобройни вакуоли се сливат в общ надлъжен вакуоларен канал, чийто начин на възникване е очевиден от наличието по него на редица отделни изходни отвори.
U СаркодинаИ Мастигофорацелият отделителен апарат е ограничен до самата вакуола. В няколко флагелати (напр. еуглена)контрактилната вакуола е заобиколена от ръб от малки образователни вакуоли, свързани с тях чрез тънки тубули.
Папас и Бранд (1958) смятат, че водата във вакуолата Амебанавлиза в резултат на изливането на малки везикули, разположени наоколо в него, чрез сливане на мембраните на тези везикули с централната вакуола. При ресничките контрактилните вакуоли обикновено са снабдени с един или повече аферентни каналчета, които пулсират. U спиростом - 1 такъв канал, Стентор- 2, г Фронтония- IO дълги канали и т.н. Каналите очевидно се разминават в цитоплазмата на много по-голямо разстояние, отколкото се вижда под микроскоп. Поне по време на осмоза Парамецийстава забележимо продължението на периферните краища на каналите в дълги, тънки крайни участъци. Образователните вакуоли и аферентните канали събират течност (т.е. главно вода) от цитоплазмата и я доставят до контрактилната вакуола. Целият вакуоларен апарат на ресничките лежи дълбоко в ендоплазмата и не променя позицията си в тялото.
Наскоро Шнайдер (1960) публикува интересни данни за ултрамикроскопската структура на контрактилната вакуола и адукторните канали Paramecium caudatum.Според Д. Н. Насонов (1924) аддукторните канали са заобиколени от осмиофилно свързване, а според
наблюдения на Гелей (Gelei, 1925a, 1925b), пореста плазма (Nierenplasma или Nephridialplasma).
Според изследването на Шнайдер, тази област от плазма, заобикаляща адукторните канали, се състои от множество тънки и извити

ориз. 125. Схема на структурата на контрактилна вакуола с аддукторен канал и околната цитоплазма в Paramecium според електронна микроскопия. (След Шнайдер, 1960). А - радиален канал в систола, контрактилна вакуола в диастола; B - радиален канал в диастола, контрактилна вакуола в систола. AK - вакуолен изходен канал; Amp - ампула на периферния канал; ЕК - канал, свързващ ампулата с контрактилната вакуола; EPR - канали на ендоплазмения ретикулум; FB - контрактилни фибрили на вакуолата; KV - контрактилна вакуола; NK - контрактилен вакуолен аддукторен канал; NT - мрежа от "нефридиални канали", свързани с каналите на ендоплазмения ретикулум; RS - клъстер от "нефридиални канали" в напречно сечение.
тръби с осмиофилни стени (фиг. 125). Вътрешният лумен на тези тръби по време на систола е 100 Å, а по време на диастола 150 Å. Целият слой нефридиална плазма е с дебелина 1-1,5 μ. По периферията си тръбите преминават директно в тубулите на ендоплазмения ретикулум, които проникват в цялото тяло на парамециума. В допълнение, нефридиалната плазма е заобиколена от специални тръби с осмиофилни стени. Понякога са подредени на гроздове. Диаметърът им достига 500 Å. Дължината на тези тръби е около 1 μ.
Аддукторният канал има осмиофилна мембрана. По време на систола този канал става подобен на процеп и има лумен от около 300 Å; в диастола той се разширява до 4000 Å. Тубулите, обграждащи адукторния канал, се отварят в него само по време на диастола, когато увеличават диаметъра си, докато по време на систола тези тубули са затворени и не са свързани с канала. Аферентният канал в своя проксимален край директно преминава в ампулата, която се отваря в резервоара на вакуолата през терминалния каналикулус. Стената на ампулата и терминалния канал съдържат тубуларни контрактилни фибрили, подредени в няколко снопа. От стените на терминалния канал те преминават директно към стената на резервоара на контрактилната вакуола, като се разпределят и там под формата на отделни снопчета.
Според Rudzinska (1957), в цитоплазмата около вакуолата Tokophrya infusionumМалки мехурчета от ergastoplasma се концентрират и се натрупват митохондрии. В допълнение, около вакуолата се намират структури, които могат да бъдат сравнени с диктиозомите. Те изглежда участват в секрецията и натрупването на течност, идваща от цитоплазмата.

А. А. Стрелков (1939) открива около отделителния канал при Циклопостиумясно видимо пръстеновидно влакно, което по всяка вероятност е мионема, която регулира изхода на течност през канала навън. При всички реснички екскреторният отвор на контрактилната вакуола е предварително оформена пора. Обикновено е кръгла или овална и има напълно постоянна позиция сред редовете реснички за всеки вид. В семейството Conchophthirusдупката изглежда като тесен прорез с удебелен ръб. Може би тази форма на порите е Conchophthirusима известно функционално значение при затварянето му. А именно, много вероятно е тук затварянето да става не чрез пръстеновидно стесняване на изходния канал, а чрез свиване на ръбовете на процепа като две устни. Пората се отваря в отделителния канал, обикновено много къса, но понякога (при Paramecium trichium)образувайки няколко подобни на бримка завои.
Електронно микроскопско изследване на контрактилната вакуола Tokophrya infusionumпотвърди наличието на постоянно съществуваща пора и отделителен канал, около който са разположени фибрили (с дебелина 180 A), които го компресират (Rudzinska, 1957). Подобни фибрили са открити в Парамеций(Шнайдер, 1960).
2. Функционално значение на контрактилните вакуоли
1) Процесът на свиване на пулсиращи вакуоли
Вакуолите работят ритмично, последователно се разширяват (стадий на диастола) и бавно се пълнят с течност, след което внезапно се свиват (стадий на систола) и изтласкват съдържанието си през отделителния канал. Там, където има специални адукторни канали, цикълът на свиване на целия апарат става по-сложен; а именно, започва с разширяването на каналите, което се случва, докато вакуолата е в състояние на систола (диастола на канала); това е последвано от свиване на каналите (систола), изливане на съдържанието им в контрактилната вакуола, което предизвиква нейната систола; в същото време каналите започват да се разширяват отново. Всъщност съответните етапи присъстват и във вакуоли, които нямат канална система, но там те са малко или невидими, тъй като по време на диастола на вакуолата в нея се изливат много малки и неправилно разположени вакуоли, разположени в околната плазма (вж. . Ophryoscolecidaeориз. 126).
Скоростта на пулсация варира сред различните протозои и също се променя сред един и същи вид под влияние на промените във факторите на околната среда.
И така, за различни сладководни Протозоиинтервалите между пулсациите при различни температури са показани в табл. 6.
ТАБЛИЦА 6
Промени в скоростта на пулсация на контрактилната вакуола при ресничките(в секунда) при различни температури

Олигохета.Оказа се, че скоростта на пулсация на контрактилната вакуола не е еднаква за всички Астоматии е във връзка с осмотичните условия на средата, в която собственикът живее. U Mesnilella clavataw. Hoplitophrya secansсред сладководните олигохети пулсацията е толкова честа, колкото и при свободно живеещите сладководни реснички: скоростта на пулсация не надвишава 1 минута. (20-30 сек.).
U Радиофрияот морски олигохети Енхитреус sp., живеещи в крайбрежната зона на Баренцово море, пулсацията се появява на всеки 6-8 минути.
Накрая при Metaradiophrya lumbriciИ Anoplophrya lumbriciот червата на почвен олигохет Eisenia foetidaи при Mesnilella fastigataИ Radiophrya prolijeraот почвата Енхитреус sp. скоростта на пулсация е по-бавна (1,5-4 мин.) от тази на сладководните реснички и малко по-бърза от тази на Астоматиот морски олигохети. По този начин, в тясно свързани видове реснички Астоматиразкрива се значителна физиологична разлика поради адаптиране към осмотична среда. За ресничките от сладководни червеи изотоничната среда е 0,4% рингер, докато за ресничките от червата на почвените червеи изотоничността съответства на 0,75-0,8% разтвор на рингер, а за морските форми - 3% разтвор.
Влиянието на промените в количеството соли в околната среда върху скоростта на пулсация на вакуола е демонстрирано от Zuelzer (1910). Тя показа това Amoeba verrucosaкогато свикне с живот в морска вода, той разкрива, като се започне с концентрация на сол от 0,3%, постепенно забавяне на пулсацията на вакуолата и когато концентрацията на сол достигне 1,5%, вакуолата напълно изчезва, само за да се появи отново, когато културата постепенно се разрежда с прясна вода. По подобен начин Herfs (1922) получава за парамеций,свикнали да живеят в разтвори на NaGl и в прясна вода, следните данни:
| Съдържание на сол във вода (в%) | 0 | 0.25 | 0.5 | 0.75 | 1 |
| Интервал между пулсациите (в сек.) | 6.2 | 9.3 | 18.4 | 24.8 | 163 |
| Количеството секретирана течност за 1 час, изразено в телесни обеми | 4.8 | 2.82 | 1.38 | 1.08 | 0.16 |
Като цяло, количеството течност, отделено през вакуолите за определен период от време, при равни други условия, е пропорционално на скоростта на пулсация и за различни Протозоимогат да бъдат доста различни. Например, контрактилна вакуола Uronema nigricansпри температура 28° за 2 минути. освобождава обем вода, равен на обема на тялото на ресничките, и Euplotes пателатова изисква 14 минути, Парамециум аурелия- 46 мин.
2) Физиологично значение на контрактилните вакуоли
Функциите на вакуолите очевидно са разнообразни. На първо място, има основание да се мисли, че вакуолите са отговорни за отстраняването от тялото на крайните метаболитни продукти, които се натрупват в него.
Предположения за тази функция са направени през миналия век от Stein (1878) и други изследователи, но Howland (1924a, 1924b) е първият, който индиректно показва, че пикочната киселина се секретира от протозои. Нейните култури от парамеций и амеби съдържат пикочна киселина, чиято концентрация нараства приблизително пропорционално на продължителността на съхраняване на културата. M. Novikov (1908), Shumway (1917) и Flather (1919) показват, че при хранене на ресничките на щитовидната жлеза, под действието на епинефрин и екстракт от епифиза, пулсацията на вакуолите става по-честа, а самите вакуоли се увеличават, т.е. стимулиране се получава екскреция под въздействието на същите лекарства, които предизвикват диуреза при гръбначните животни.
Weatherby (1927) открива урея в течността на контрактилната вакуола спиростом,екстрахиран с помощта на микропипета. Въпреки това, съдейки по изчисленията на този автор, вакуолата премахва само 1% от общото количество, отделено от масовите култури. спиростомумурея.
Като цяло данните за естеството на азотните екскреционни продукти са много противоречиви. В допълнение към пикочната киселина и уреята, някои автори посочват амоняка като екскреторни продукти вместо тези вещества (напр. ГлаукомаИ спиростом).Единственото обстоятелство, което не е напълно съгласно с екскреторната функция на контрактилните вакуоли, е, че те никога не участват в екскрецията на интравитални багрила, въведени в плазмата, които много често се екскретират при висшите животни през бъбреците и други отделителни органи.
Многократно е изразено мнението, че контрактилните вакуоли участват в респираторния метаболизъм на протозоите, като улесняват отстраняването на въглеродната киселина от цитоплазмата; обаче няма действителни доказателства за специфична активност на вакуолата в тази посока. Само косвени данни от Лудвиг (Ludwig, 1928) за съответствието между действителното количество въглена киселина, отделена през вакуолата, и изчисленото количество въглена киселина, секретирана от животни, се интерпретират от този автор в полза на дихателната функция на вакуолата.
Повечето автори в момента подчертават важната роля, която контрактилните вакуоли играят като осморегулаторен апарат. Дори Деген (1905) посочва, че плазмата на сладководните протозои, поради наличието на соли в нея, има по-високо осмотично налягане от околната среда; и това обстоятелство води до постоянното поглъщане на вода от цитоплазмата. Контрактилната вакуола служи за отстраняване на излишната вода, непрекъснато навлизаща в плазмата. Правилността на тази гледна точка се потвърждава както от естеството на разпределението на контрактилните вакуоли сред протозоите, така и от експериментите за аклиматизиране на сладководни протозои към вода с нарастващи концентрации на сол (Firiley, 1930; Loefer, 1939 и др.).
Наскоро експериментално изследване на Jl. H. Seravini (1958) над пулсацията на вакуолата Paramecium caidatum,който предоставя данни, показващи, че когато ресничките се прехвърлят в физиологични разтвори с повишена концентрация, те се развиват през първите 15-30 минути. Има няколкократно намаляване на скоростта на пулсация на вакуолата, след което скоростта на пулсация постепенно се ускорява и се доближава до скоростта, характерна за контролните реснички, но обикновено не достига нивото си. Например, в 0,2% NaCl след 30 минути. Скоростта на пулсация е средно 40,5 секунди, а след 48 часа. достига 13.1 сек., докато при контролната група пулсацията протича със скорост 10.3 сек. Същото се наблюдава в разтвори от 0,1% LiCi, 0,5% CaCl2, 1% захароза и разтвор на урея. Забавянето се случи 7-15 пъти, но след 2 дни
скоростта на пулсация в тези разтвори се забави само 1,5-3 пъти. При по-ниски концентрации на сол се наблюдава пълно възстановяване на скоростта на пулсация на вакуолата. Вещества като HGl, NaOH, GaGl 2, формалин в незначителни концентрации предизвикват леко забавяне на пулсацията, която след това бързо се нормализира. Адреналинът в концентрация 0,0005% дори увеличи водния обмен, но с течение на времето пулсацията също се нормализира. По този начин осмотично активните и неактивните вещества причиняват подобни промени в активността на контрактилната вакуола. В този случай има обща тенденция за регулиране на водния поток, преминаващ през цитоплазмата на ресничките чрез промяна на скоростта на пулсация на вакуолата. В допълнение, ресничките се адаптират към променящите се условия на осмотично налягане на външната среда. Това също е отбелязано от Kitching (1952) по отношение на АрхезиумИ Подофрия.При тези реснички, при високи концентрации на етиленгликол и захароза, активността на вакуолата напълно спира, но след известно време пулсацията на вакуолата и обменът на вода се възстановяват.
Всичко това показва сложен механизъм на действие на контрактилната вакуола, която играе несъмнена роля във водния обмен на ресничките. Seravin обаче вярва, че въз основа на факта, че осмотично неактивните вещества причиняват забавяне на пулсацията на вакуолата, е възможно да се предположи, че обменът на вода в ресничките не се определя от осмотични модели и следователно ролята на контрактилната вакуола в осморегулацията не може да се счита за доказана .
За да се разбере функцията на вакуолата, разбира се е необходимо да се разбере и механизмът на циркулация на водата от цитоплазмата в резервоара на вакуолата и механизмът на свиване на самата вакуола. I. Gelei и G. Gelei (Gelei, 1928; Gelei, 1939), подобно на редица други изследователи, смятат, че само тургорното напрежение, създадено в цитоплазмата, и следователно осмотичните условия определят намаляването на вакуолния резервоар. Възможно е да има и активен механизъм за секреция на вода във вакуолата (Kitching, 1956). Според Шнайдер (Schneider, 1950) пулсацията на вакуолата се причинява от активността на контрактилните фибрили, разположени в стената на вакуолата, и пълненето на аферентните канали с вода се извършва по следния начин. Течността се събира в ергастоплазмените тубули и след това, поради по-големия диаметър на тези тубули от нефридиалните плазмени тубули, се създава воден поток от ергастоплазмените тубули в нефридиалните плазмени тубули и от тях в лумена на канала. Разбира се, необходими са допълнителни изследвания, за да се разбере по-точно механизмът на тези процеси. Екскреторната и дихателната активност на контрактилната вакуола изглежда много възможна, но не е категорично доказана.
3) Контрактилна вакуола и апарат на Голджи
Въпросът за морфологичното значение на контрактилната вакуола навлезе в нова фаза след работата на Д. Н. Насонов (1924, 1925), който изрази идеята, че контрактилната вакуола на протозоите е хомоложна на апарата на Голджи в клетките Metazoa.Насонов (1924) и почти едновременно с него И. Гелей (J. Gelei, 1925a, 1925b), прилагайки техниката на осмация към ресничките, откриха важни нови характеристики в структурата на контрактилните вакуоли (фиг. 127). Насонов установи, че около контрактилната вакуола на много реснички (Campanella, Lionotus, Parameciuin, Nassula),както и Хиломонасе силно почерняла от
редуцирана осмиева обвивка, с гладка повърхност или заобиколена от слой от малки, осмиофилни зърна. Обвивката или обгражда цялата вакуола изцяло (при горните видове), или има формата на мрежест пръстен, опасващ вакуолата по екватора (Chilodon, Dogielella).При форми със система от адукторни канали (парамеций),черупката продължава към последната; в този случай каналите могат да бъдат проследени в ендоплазмата много по-далеч, отколкото с всеки друг изследователски метод, а дисталните им части са заобиколени от сив мрежест ръкав. Насонов смята, че този участък от каналите е същинската екскреторна част на апарата, докато проксималната част на каналите и самата вакуола са само екскреторната част на вакуоларната система. U ДогиелелаНасонов би могъл да твърди, че по време на систола на вакуолата черният пръстен, който я заобикаля, не следва последната при свиването й, а остава на първоначалното си място. Gelei (1925a, 1925b) се опитва да ги сравни с нефридиалната система Metazoa,приравнявайки самата контрактилна вакуола към пикочния мехур и други части на апарата към други части на нефридиите. Насонов се фокусира върху сходството на осмотичната мембрана на вакуолите с апарата на Голджи в Metazoa.Всъщност чертежите, които той предоставя на различни видове контрактилни вакуоли и апарати на Голджи, взети от различни видове клетки в Metazoa,изключително убедителни в своята прилика.

Въз основа на това Насонов счита за възможно търсенето на адукторни канали в осмиофилната спонгиозна мембрана парамециурни мрежест пръстен Догитлелане само морфологичен, но и функционален паралелизъм между тези образувания и апарата на Голджи. Мрежовата структура на пръстенното вещество зависи от наличието на малки образователни вакуоли в него - етапът на свързана секреция. След това тези вакуоли се сливат, за да образуват централна вакуола, разположена в центъра на пръстена - етапът на свободна секреция, секретирана навън. Според концепцията на Насонов апаратът на Голджи служи за събиране на известни вещества от плазмата, които се подготвят в нея за извеждането им от тялото чрез вакуола. Цялата фактическа страна на откритията на Насонов е напълно потвърдена от по-късни изследвания на редица автори: А. А. Стрелков (1939 г.) CycloposlhiidaeФоре-Фремие, 1925 г.) Ворличела,Крал (1928) Paramecium trichiumИ Euplotesи т.н. Вярно е, че в някои протозои не беше възможно да се открие специална осмиофилна мембрана около вакуолата, което можеше да зависи от методологични проблеми.
в клетките на всеки Metazoaпод формата на мрежеста формация или състояща се от отделни диктиозоми. Следователно можем да мислим, че той присъства и във всички едноклетъчни организми. Наистина, проучванията на Hirschler (1927), Hovasse (1937), Joyet-Lavererie (1926) и други показват наличието на много протозои в плазмата (Gregarina, Coccidia, Flagellata, Sarcodina)малки пръчици, пръстени, топчета със силно осмиофилен ръб и клетки, много подобни на диктиозоми Metazoa.
Признаци на апарата на Голджи MetazoaНа първо място, този уред е в състояние специфично да редуцира определени съединения на тежки метали – сребро и осмий, което се изразява в почерняване на структурите на уреда. Тези структури не се оцветяват интравитално с неутрално червено и метиленово синьо. Често характерните структури на апарата на Голджи са диктиозоми, които се състоят от две части - хромофилна и хромофобна. Обикновено секреторните гранули са потопени в хромофобната част. По правило апаратът на Голджи е локализиран точно в секреторната зона на клетката.
Изследванията с електронен микроскоп показват единството на структурите на апарата на Голджи в различни клетки Metazoa.Във всички изследвани клетки са идентифицирани двойни мембрани (или γ-цитомембрани), разположени успоредно една на друга и образуващи система от 3-7 или повече двойки. Всяка двойка обхваща по-светло пространство с ширина 50-200 Å и образува нещо като плоска торбичка. Понякога се разширява до голяма вакуола. Мембраните са потопени в финозърнест основен материал. В някои случаи мембраните могат да обграждат вакуоли с различни размери. Всички тези структури са доста стабилни.
Показателно е, че има пълно съответствие между снимките на апарата на Голджи, описани с помощта на светлинен микроскоп от Насонов и други изследователи на клетките Metazoa,и съвременни изображения, получени с електронен микроскоп.
Dalton и Felix (1957) също отбелязват, че контрактилната вакуола Хламидомонадаограничени от мембрани, подобни на тези на апарата на Голджи на клетките Metazoa,и по този начин авторите подкрепят хипотезата на Насонов за хомологията на тези органели. Около контрактилната вакуола при ресничестите КампанелаИ Офридиумима множество тръби с плътни осмиофилни стени, които могат да бъдат до известна степен сравними с ултраструктурите на апарата на Голджи (Fauré-Fremiet et Rouiller, 1959). Горните данни от Шнайдер за структурата на нефридиалната плазма около адукторните канали на контрактилната вакуола Парамециииимзатруднява обаче хомологизирането на неговата структура с ултраструктурата на диктиозомите.
Още по-рано френски автори (Duboscq et Grassé, 1933; Grassé et Hollande, 1941) също отхвърлиха хомологията на осмиофилните мембрани на контрактилните вакуоли с апарата на Голджи. Те вярват, че осмиофилните стени на вакуолите, както и самите контрактилни вакуоли, са образувания, напълно независими от апарата на Голджи.
Трябва да се отбележи, че сърповидни или пръстеновидни диктиозоми са открити в някои инфузории, като напр. Balantidium elongatum,извън връзка, с вакуоли (Villeneuve-Brachori, 1940). Noirot-Timothee (1957) идентифицира с електронен микроскоп типичните структури на апарата на Голджи в ендоплазмата на някои Ophryoscolecidaeне е свързан с контрактилната вакуола.
Френските автори хомологизират парабазалното тяло с апарата на Голджи (виж стр. 115), тъй като тази формация, особено когато се изследва с електронен микроскоп, се оказа подобна на диктиозомите на клетките Metazoa(фиг. 80).
Несъмнено са необходими допълнителни сравнителни изследвания на протозоите, за да се разреши въпросът за хомологията на определени структури с апарата на Голджи на клетките Metazoa.