ستُعرّف هذه المقالة القارئ ببنية أبسط الكائنات الحية، أي أنها ستركز على بنية الفجوة الانقباضية، التي تؤدي وظيفة إخراجية (وليس فقط)، وتتحدث عن أهمية أبسط الكائنات الحية، وتصف طرق لوجودهم في البيئة.
فجوة مقلصة. مفهوم
فجوة (من الفجوة الفرنسية، من الكلمة اللاتينية فراغ - فارغة)، تجاويف صغيرة كروية الشكل في الخلايا النباتية والحيوانية أو الكائنات الحية وحيدة الخلية. الفجوات الانقباضية شائعة في المقام الأول بين الكائنات الحية البسيطة التي تعيش في المياه العذبة، مثل الطلائعيات مثل بروتي الأميبا والنعال الهدبية، والتي حصلت على اسمها الأصلي بسبب شكل الجسم المشابه لشكل نعل الحذاء. بالإضافة إلى الكائنات الأولية المدرجة، تم العثور على هياكل متطابقة أيضًا في خلايا إسفنجيات المياه العذبة المختلفة التي تنتمي إلى عائلة بادياغوف.

هيكل الفجوة مقلص. مميزاته
الفجوة المقلصة عبارة عن عضية غشائية تطلق السوائل الزائدة من السيتوبلازم. يختلف توطين وبنية هذا الجهاز بين الكائنات الحية الدقيقة المختلفة. من مجموعة من الفجوات الحويصلية أو الأنبوبية تسمى الإسفنجية، يدخل السائل إلى الفجوة الانقباضية. بفضل التشغيل المستمر لهذا النظام، يتم الحفاظ على حجم خلية مستقر. تحتوي الأوليات على فجوات مقلصة، وهي جهاز ينظم الضغط الأسموزي ويعمل أيضًا على إخراج الفضلات من الجسم. يتكون جسم الأوليات من خلية واحدة فقط، والتي بدورها تقوم بجميع الوظائف الحيوية اللازمة. يمتلك ممثلو هذه المملكة الفرعية، مثل هدب النعال والأميبا الشائعة والكائنات الحية الأخرى أحادية الخلية، جميع خصائص كائن حي مستقل.
دور الكائنات الحية البسيطة
تقوم الخلية بجميع الوظائف الحيوية: الإخراج، التنفس، التهيج، الحركة، التكاثر، التمثيل الغذائي. البروتوزوا موجودة في كل مكان. يعيش العدد الأكبر من الأنواع في المياه البحرية والمياه العذبة، ويعيش العديد منها في التربة الرطبة، ويمكن أن تصيب النباتات، وتعيش في أجسام الحيوانات متعددة الخلايا والبشر. في الطبيعة، تؤدي الأوليات دورًا صحيًا؛ فهي تشارك أيضًا في دورة المواد وتكون غذاءً للعديد من الحيوانات.

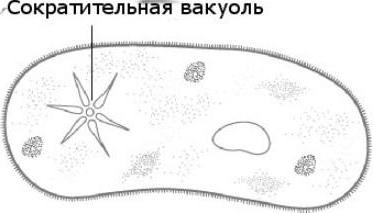
فجوة مقلصة في الأميبا الشائع
الأميبا الشائعة هي ممثلة لفئة الجذور، على عكس الممثلين الآخرين، ليس لها شكل جسم ثابت. تتم الحركة بمساعدة الأرجل الكاذبة. الآن دعونا نتعرف على الوظيفة التي تؤديها الفجوة الانقباضية في الأميبا. هذا هو تنظيم مستوى الضغط الاسموزي داخل خليته. يمكن أن تتشكل في أي جزء من الخلية. من خلال الغشاء الخارجي، يدخل الماء من البيئة بشكل تناضحي. تركيز المواد الذائبة في خلية الأميبا أعلى منه في البيئة. وهكذا ينشأ اختلاف في الضغط داخل الخلية الأولية وخارجها. وظائف الفجوة المقلصة في الأميبا هي نوع من أجهزة الضخ التي تزيل الماء الزائد من خلية أبسط كائن حي. يمكن للأميبا بروتيوس أن تطلق السائل المتراكم في البيئة في أي مكان على سطح الجسم.
بالإضافة إلى التنظيم التناضحي، فإنه يؤدي وظيفة التنفس في الحياة، لأنه نتيجة للتناضح، يسلم الماء الوارد الأكسجين المذاب فيه. ما هي الوظيفة الأخرى التي تؤديها الفجوة الانقباضية؟ كما أنه يؤدي وظيفة إفرازية، وهي إزالة المنتجات الأيضية مع الماء إلى بيئتها.

التنفس، الإفراز، التنظيم الأسموزي في النعال الهدبي
جسم الأوليات مغطى بقشرة كثيفة ذات شكل ثابت. والطحالب، بما في ذلك بعض الأوليات. يحتوي جسم الهدب على بنية أكثر تعقيدًا من بنية الأميبا. يوجد في خلية النعال فجوتان انقباضيتان تقعان في الأمام والخلف. في هذا الجهاز يمكن تمييز الخزان والعديد من الأنابيب الصغيرة. وتتوضع الفجوات الانقباضية باستمرار، بفضل هذا الهيكل (من الأنابيب الدقيقة)، في مكان دائم في الخلية.
وتتمثل المهمة الرئيسية للفجوة المقلصة في حياة هذا الممثل للكائنات الأولية في التنظيم التناضحي؛ كما أنها تزيل الماء الزائد من الخلية، والذي يخترق الخلية بسبب التناضح. أولا، تنتفخ القنوات المقربة، ثم يتم ضخ الماء منها إلى خزان خاص. ينقبض الخزان، وينفصل عن قنوات الإمداد، ويتم طرح الماء عبر المسام. يوجد في الخلية الهدبية فجوتان مقلصتان تعملان بدورها في الطور المضاد. بفضل تشغيل اثنين من هذه الأجهزة، يتم ضمان عملية مستمرة. بالإضافة إلى ذلك، يدور الماء بشكل مستمر بسبب نشاط الفجوات الانقباضية. يتم ضغطها بالتناوب، ويعتمد تواتر الانقباضات على درجة الحرارة المحيطة.
وهكذا، في درجة حرارة الغرفة (+18 - +20 درجة مئوية)، فإن وتيرة تقلصات الفجوة، وفقا لبعض البيانات، 10-15 ثانية. وباعتبار أن الموطن الطبيعي للشبشب هو أي مسطح مائي عذب به مياه راكدة ووجود مواد عضوية متحللة فيه فإن درجة حرارة هذه البيئة تتغير بعدة درجات حسب الوقت من السنة وبالتالي تكرار التقلصات يمكن أن تصل إلى 20-25 ثانية. في غضون ساعة، تكون الفجوة الانقباضية لكائن حي بسيط قادرة على إطلاق كميات كبيرة من الماء من الخلية. بما يتناسب مع حجمه. أنها تتراكم العناصر الغذائية، وبقايا الطعام غير المهضومة، والمنتجات النهائية لعملية التمثيل الغذائي، ويمكن أيضا العثور على الأكسجين والنيتروجين.
معالجة مياه الصرف الصحي مع الأوليات
إن تأثير الأوليات على دورة المواد في الطبيعة له أهمية كبيرة. في الخزانات، بسبب تصريف مياه الصرف الصحي، تتكاثر البكتيريا بأعداد كبيرة. ونتيجة لذلك، تظهر كائنات حية بسيطة مختلفة، تستخدم هذه البكتيريا كغذاء وبالتالي تساهم في الحياة الطبيعية

خاتمة
وعلى الرغم من البنية البسيطة لهذه الكائنات وحيدة الخلية، التي يتكون جسمها من خلية واحدة، إلا أنها تؤدي وظائف كائن حي كامل، وتتكيف بشكل مذهل مع البيئة. يمكن ملاحظة ذلك حتى في مثال بنية الفجوة المقلصة. اليوم، تم بالفعل إثبات الأهمية الهائلة للأبسط في الطبيعة ومشاركتها في دورة المواد.
الفراغ التعاقدي، انظر الفراغ... القاموس الموسوعي العلمي والتقني
فجوة مقلصة فجوة مقلصة. نوع من الفجوة في بعض مجموعات الطلائعيات، يشارك في إزالة الماء (المحاليل) من الخلية أثناء الانكماش وفي امتصاص الماء بواسطة الخلية أثناء التمدد، مما يعمل على تنظيم الضغط الاسموزي.... ... البيولوجيا الجزيئية وعلم الوراثة. القاموس التوضيحي.
هيكل الخلية حقيقية النواة. الفجوة مدرجة بالرقم 10. الفجوة هي عضية ذات غشاء واحد توجد في بعض الخلايا حقيقية النواة و... ويكيبيديا
أو رتبة هليوزوا من فئة ساركوديكا (q.v.) نوع من الحيوانات الأولية (q.v.). الخصائص المورفولوجية. وتتميز بجسم بروتوبلازمي كروي، منه رفيع يشبه الخيط، غير... ... يمتد في كل الاتجاهات مثل الأشعة. القاموس الموسوعي ف. بروكهاوس وآي. إيفرون
أو الهدبيات، وهي مفرزة من فئة الهدبيات، أو الهدبيات (qv)، وهي نوع من الأوليات (qv). الهدبية. أنا (أسبيروتريشا). معنى الحروف: مسحوق؛ الطبقة السنخية من الإكتوبلازم. ad.Z الصف adoral من الأهداب. ب سيتا اللمس. أهداب CL. ... القاموس الموسوعي ف. بروكهاوس وآي. إيفرون
- (Lobosea) وهي طائفة من الأوليات الأكثر بساطة تنظيما فوق طائفة الريزومات. محروم من الداخل الهيكل العظمي والخارجي قذائف. شكل الجسم متغير، تتراوح أحجامه عادةً من 20 إلى 700 ميكرون، ونادرًا ما يزيد قليلاً. شكل وحجم الأرجل الكاذبة من سمات ... ... القاموس الموسوعي البيولوجي
- (Flagellata s. Mastigophora، انظر الجدول. Flagellata، Flagellata) فئة الأوليات (البروتوزوا). مثل جميع الممثلين الآخرين لهذا النوع، لديهم جسم يتكون من خلية واحدة فقط، تمثل البروتوبلازم ونواة مع نواة.... ... القاموس الموسوعي ف. بروكهاوس وآي. إيفرون
أو البروتوزوا. محتويات المقال: الخصائص والتصنيف. رسم تاريخي. مورفولوجيا. بروتوبلازم مع شوائب (الأكياس الثلاثية، النواة، الفجوات الانقباضية، الكروماتوفورز، إلخ). الأغطية والهيكل العظمي. الحركة ص. الأرجل الكاذبة، الأسواط و... القاموس الموسوعي ف. بروكهاوس وآي. إيفرون
أ. الفجوات التعاقدية
1. مورفولوجيا الفجوات المقلصة
معظم الأوليات (باستثناء سبوروزوا)لديهم جهاز إفراز خاص، موضعي بشكل أو بآخر، ومعقد للغاية في بعض الأحيان، ومعظمه له طابع ما يسمى بالفجوات الانقباضية أو النابضة.
كقاعدة عامة، جميع حيوانات المياه العذبة لديها فجوات. الضرعو ساركوديناوفي جميع الشركات العملاقة.
ش الضرعبالنسبة للجزء الأكبر، لا يوجد سوى فجوة انقباضية واحدة، ونادرًا ما يكون هناك فجوة انقباضية واحدة (فيتوماستيجينا).
تتناسب أبعاد الجهاز الفجوي بشكل عام بشكل مباشر مع حجم الجسم وتتناسب عكسيًا مع عدد الفجوات في جسم الحيوان. لذلك، في الشركات العملاقة الكبيرة Heterotricha (ستينتور، سبيروستوموم)لا يوجد سوى فجوة واحدة، لكنها (مع قنواتها) تصل إلى أحجام كبيرة جدًا. ش البويضة القصبة الهوائيةيصل عددها إلى 30 فجوة، لكنها صغيرة جدًا.
في بعض الأحيان يلاحظ زيادة في عدد الفجوات مع نمو الحيوان، كما هو الحال في كولينيا برانشياروموبعض الآخرين فضالات.أن الضرب لعدد الفجوات التي كولينيا (أنوبلوفريا)يحدث أثناء حياة الفرد، ويبدو أنه حدث أثناء تطور المجموعة بأكملها فضالات(الشكل 124). في الواقع، من الممكن تحديد عدد من الأشكال من بينها مع جهاز فجوي أكثر تعقيدا تدريجيا. نعم ي دوجيليلالا يوجد سوى فجوة فرعية واحدة فقط في الجنس أنوبلوفريا 1 صف طولي لعدد صغير من الفجوات. عند الولادة بوتشيليلاو موبودونتوفريايزداد عدد الفجوات في هذا الصف بشكل كبير، وفي راديوفرياتشكل الفجوات صفين طوليين. وأخيرا النوع هابتوفريايندمج عدد من الفجوات العديدة في قناة فراغية طولية مشتركة، ويمكن رؤية طريقة منشأها من خلال وجود عدد من فتحات الخروج المنفصلة على طولها.
ش ساركوديناو الضرعيقتصر جهاز الإخراج بأكمله على الفجوة نفسها. في عدد قليل من السوطيات (على سبيل المثال. اليوجلينا)وتحيط بالفجوة الانقباضية حافة من الفجوات التعليمية الصغيرة المتصلة بها بواسطة أنابيب رفيعة.
يعتقد باباس وبراندت (1958) أن الماء موجود في الفجوة الأميبايدخل نتيجة تدفق الحويصلات الصغيرة الموجودة حوله، وذلك عن طريق اندماج أغشية هذه الحويصلات مع الفجوة المركزية. في الأهداب، يتم تزويد الفجوات الانقباضية عادةً بواحدة أو أكثر من القنوات الواردة التي تنبض. ش سبيروستوموم - 1 مثل هذه القناة، ستينتور- 2، ص فرونتونيا- قنوات IO الطويلة، وما إلى ذلك. يبدو أن القنوات تتباعد في السيتوبلازم إلى مسافة أكبر بكثير مما يمكن رؤيته تحت المجهر. على الأقل أثناء التناضح البراميسيوميصبح استمرار النهايات المحيطية للقنوات في مقاطع نهاية طويلة ورفيعة ملحوظًا. تقوم الفجوات التعليمية والقنوات الواردة بجمع السوائل (أي الماء بشكل رئيسي) من السيتوبلازم وتوصيلها إلى الفجوة المقلصة. يقع الجهاز الفراغي بأكمله للأهداب في أعماق الإندوبلازم ولا يغير موضعه في الجسم.
في الآونة الأخيرة، نشر شنايدر (1960) بيانات مثيرة للاهتمام حول البنية المجهرية للفجوة الانقباضية والقنوات المقربة المتناعلة المذنبة.وفقًا لـ D. N. Nasonov (1924)، فإن القنوات المقربة محاطة باقتران أوسميوفيل، ووفقًا لـ
ملاحظات Gelei (Gelei، 1925a، 1925b)، البلازما الإسفنجية (Nierenplasma أو Nephridialplasma).
ووفقا لبحث شنايدر، فإن هذه المنطقة من البلازما المحيطة بالقنوات المقربة تتكون من العديد من الخلايا الرقيقة والملتوية

أرز. 125. مخطط هيكل الفجوة المقلصة مع القناة المقربة والسيتوبلازم المحيط بها في المتناعلة حسب المجهر الإلكتروني. (بعد شنايدر، 1960). أ - القناة الشعاعية في الانقباض، والفجوة الانقباضية في الانبساط. ب - القناة الشعاعية في الانبساط، والفجوة الانقباضية في الانقباض. AK - قناة خروج الفجوة. أمبير - أمبولة القناة الطرفية. EK - قناة تربط الأمبولة بالفجوة الانقباضية. EPR - قنوات الشبكة الإندوبلازمية. FB - اللييفات المتقلصة للفجوة. KV - فجوة مقلصة. NK - قناة المقربة الفجوة المتقلصة. NT - شبكة "القنوات الكلوية" المرتبطة بقنوات الشبكة الإندوبلازمية؛ RS - مجموعة من "القنوات الكلوية" في المقطع العرضي.
أنابيب ذات جدران أوسميوفيلية (الشكل 125). يبلغ التجويف الداخلي لهذه الأنابيب أثناء الانقباض 100 أنجستروم، وأثناء الانبساط 150 أنجستروم. يبلغ سمك الطبقة الكاملة من البلازما النيفريدية 1-1.5 ميكرومتر. في محيطها، تمر الأنابيب مباشرة إلى أنابيب الشبكة الإندوبلازمية، التي تخترق جسم البراميسيوم بأكمله. بالإضافة إلى ذلك، فإن البلازما النيفريدية محاطة بأنابيب خاصة ذات جدران أوسميوفيلية. يتم ترتيبها في بعض الأحيان في عناقيد. قطرها يصل إلى 500 Å. يبلغ طول هذه الأنابيب حوالي 1 ميكرومتر.
تحتوي القناة المقربة على غشاء أسموفيلي. أثناء الانقباض، تصبح هذه القناة على شكل شق ويبلغ تجويفها حوالي 300 أنجستروم، وفي حالة الانبساط تتوسع إلى 4000 أنجستروم. الأنابيب المحيطة بالقناة المقربة لا تفتح فيها إلا أثناء الانبساط، عندما يزيد قطرها، بينما أثناء الانقباض تكون هذه الأنابيب مغلقة وغير متصلة بالقناة. تمر القناة الواردة في نهايتها القريبة مباشرة إلى الأمبولة، والتي تفتح في خزان الفجوة من خلال القناة الطرفية. يحتوي جدار الأمبولة والقناة الطرفية على ألياف ليفية انقباضية أنبوبية مرتبة في عدة حزم. من جدران القناة الطرفية تمر مباشرة إلى جدار خزان الفجوة المتقلصة، وتتوزع هناك أيضًا على شكل حزم منفصلة.
وفقا لرودزينسكا (1957)، في السيتوبلازم حول الفجوة توكوفريا إنفيونومتتركز فقاعات الإرغاستوبلازما الصغيرة وتتراكم الميتوكوندريا. بالإضافة إلى ذلك، توجد هياكل يمكن مقارنتها بالديكتيوسومات حول الفجوة. ويبدو أنها تشارك في إفراز وتراكم السوائل القادمة من السيتوبلازم.

تم العثور على A. A. Strelkov (1939) حول قناة الإخراج في سيكلوبوستيومألياف حلقية مرئية بوضوح، وهي في جميع الاحتمالات عبارة عن ورم عضلي ينظم خروج السائل عبر القناة إلى الخارج. في جميع الأهداب، تكون الفتحة الإخراجية للفجوة الانقباضية عبارة عن مسام مُشكل مسبقًا. وعادة ما تكون مستديرة أو بيضاوية ولها موضع ثابت تمامًا بين صفوف الأهداب لكل نوع. في العائلة كونكوفثيروسيشبه الثقب شقًا ضيقًا ذو حافة سميكة. ربما هذا هو شكل المسام كونكوفثيروسله أهمية وظيفية معروفة عند إغلاقه. وهي أنه من المحتمل جدًا أن يحدث الإغلاق هنا ليس من خلال الانقباض الحلقي لقناة المخرج، ولكن من خلال انهيار حواف الفجوة مثل الشفتين. ينفتح المسام في قناة الإخراج، وعادةً ما يكون قصيرًا جدًا، ولكن في بعض الأحيان (في المتناعلة التريشيوم)تشكيل عدة انحناءات تشبه الحلقة.
الفحص المجهري الإلكتروني للفجوة المقلصة توكوفريا إنفيونومأكد وجود مسام وقناة إخراجية موجودة بشكل دائم، حيث توجد حولها ألياف ليفية (بسمك 180 أمبير) تضغط عليها (Rudzinska، 1957). تم العثور على ألياف مماثلة في البراميسيوم(شنايدر، 1960).
2. الأهمية الوظيفية للفجوات مقلص
1) عملية انقباض الفجوات النابضة
تعمل الفجوات بشكل إيقاعي، وتتوسع بالتناوب (مرحلة الانبساط) وتمتلئ ببطء بالسوائل، ثم تنقبض فجأة (مرحلة الانقباض) وتدفع محتوياتها للخارج عبر قناة الإخراج. عندما تكون هناك قنوات تقريبية خاصة، تصبح دورة انكماش الجهاز بأكمله أكثر تعقيدًا؛ أي أنه يبدأ بتمدد القنوات، والذي يحدث عندما تكون الفجوة في حالة انقباض (انبساط القناة)؛ ويعقب ذلك انقباض القنوات (الانقباض)، وصب محتوياتها في الفجوة الانقباضية، مما يسبب انقباضها؛ وفي الوقت نفسه، تبدأ القنوات في التوسع مرة أخرى. في الواقع، توجد المراحل المقابلة أيضًا في الفجوات التي تفتقر إلى نظام القناة، ولكنها قليلة أو غير مرئية، لأنه أثناء انبساط الفجوة، تُسكب فيها فجوات صغيرة جدًا وغير منتظمة تقع في البلازما المحيطة (راجع . المبيضياتأرز. 126).
يختلف معدل النبض بين الكائنات الأولية المختلفة، ويتغير أيضًا بين نفس الأنواع تحت تأثير التغيرات في العوامل البيئية.
لذلك، للمياه العذبة المختلفة البروتوزوايظهر في الجدول الفترات الفاصلة بين النبضات عند درجات حرارة مختلفة. 6.
الجدول 6
التغييرات في معدل نبض الفجوة المقلصة في الشركات الهدبية(في الثانية) في درجات حرارة مختلفة

Oligochaeta.اتضح أن معدل نبض الفجوة الانقباضية ليس هو نفسه لدى الجميع فضالاتويرتبط بالظروف الاسموزية للبيئة التي يعيش فيها المالك. ش ميسنيليلا كلافاتاو. هوبليتوفريا سيكانزبين قليلات الأشواك في المياه العذبة، يكون النبض متكررًا كما هو الحال في أهداب المياه العذبة التي تعيش بحرية: لا يتجاوز معدل النبض دقيقة واحدة. (20-30 ثانية).
ش راديوفريامن قليلات الأشواك البحرية إنتشيترايوس sp.، الذين يعيشون في المنطقة الساحلية لبحر بارنتس، يحدث النبض كل 6-8 دقائق.
وأخيرا، في ميتاراديوفريا لومبريسيو أنوبلوفريا لومبريسيمن أمعاء قليلات التربة إيسينيا فويتيداوفي ميسنيليلا فاستيجاتاو راديوفريا بروليجيرامن التربة إنتشيترايوس sp. معدل النبض أبطأ (1.5-4 دقائق) من أهداب المياه العذبة، وأسرع إلى حد ما من ذلك في أهداب المياه العذبة. فضالاتمن oligochaetes البحرية. وهكذا، في الأنواع ذات الصلة الوثيقة من الشركات العملاقة فضالاتتم الكشف عن اختلاف فسيولوجي كبير بسبب التكيف مع البيئة التناضحي. بالنسبة للأهداب من ديدان المياه العذبة، فإن الوسط متساوي التوتر هو 0.4٪ من الجرس، بينما بالنسبة لأهداب أمعاء ديدان التربة، فإن متساوي التوتر يتوافق مع محلول رنين بنسبة 0.75-0.8٪، وبالنسبة للأشكال البحرية - محلول 3٪.
وقد أظهر زويلزر (1910) تأثير التغيرات في كمية الأملاح في البيئة على معدل نبض الفجوة. لقد أظهرت ذلك الأميبا الثؤلوليةعند التعود على الحياة في مياه البحر، يكشف بدءاً من تركيز الملح 0.3%، عن تباطؤ تدريجي في نبض الفجوة، وعندما يصل تركيز الملح إلى 1.5%، تختفي الفجوة تمامًا، لتعود للظهور عند الاستزراع. يتم تخفيفه تدريجياً بالماء العذب. وبالمثل، حصل هرفس (1922) على براميسيوم,اعتادوا على الحياة في محاليل NaGl وفي المياه العذبة، البيانات التالية:
| محتوى الملح في الماء (%) | 0 | 0.25 | 0.5 | 0.75 | 1 |
| الفاصل الزمني بين النبضات (بالثواني) | 6.2 | 9.3 | 18.4 | 24.8 | 163 |
| كمية السوائل التي يتم إفرازها خلال ساعة واحدة، معبرًا عنها بحجم الجسم | 4.8 | 2.82 | 1.38 | 1.08 | 0.16 |
وبشكل عام، فإن كمية السائل التي تفرز من خلال الفجوات في فترة زمنية معينة، مع تساوي العوامل الأخرى، تتناسب مع معدل النبض وباختلاف البروتوزوايمكن أن تكون مختلفة تمامًا. على سبيل المثال، فجوة مقلص أورونيما نيجريكانزعند درجة حرارة 28 درجة لمدة دقيقتين. يطلق كمية من الماء تساوي حجم جسم الهدب، و Euplotes الرضفةوهذا يتطلب 14 دقيقة، باراميسيوم أوريليا- 46 دقيقة.
2) الأهمية الفسيولوجية للفجوات مقلص
يبدو أن وظائف الفجوات متنوعة. بادئ ذي بدء، هناك سبب للاعتقاد بأن الفجوات هي المسؤولة عن إزالة المنتجات الأيضية النهائية التي تتراكم فيه من الجسم.
تم تقديم اقتراحات حول هذه الوظيفة في القرن الماضي بواسطة ستاين (1878) وباحثين آخرين، لكن هاولاند (1924 أ، 1924 ب) كان أول من أظهر بشكل غير مباشر أن حمض البوليك يفرز بواسطة الأوليات. تحتوي مزارعها من البراميسيوم والأميبات على حمض البوليك، الذي يزداد تركيزه تقريبًا بما يتناسب مع طول فترة الاحتفاظ بالثقافة. أظهر M. Novikov (1908)، Shumway (1917) وFlather (1919) أنه عند تغذية أهداب الغدة الدرقية، تحت تأثير مستخلص الإبينفرين والغدة الصنوبرية، يصبح نبض الفجوات أكثر تواتراً، وتزداد الفجوات نفسها، أي يتم الحصول على التحفيز عن طريق الإفراز تحت تأثير نفس الأدوية التي تسبب إدرار البول في الفقاريات.
اكتشف ويذربي (1927) اليوريا في سائل الفجوة المقلصة سبيروستوموم,المستخرجة باستخدام micropipette. ومع ذلك، إذا حكمنا من خلال حسابات هذا المؤلف، فإن الفجوة تزيل 1٪ فقط من إجمالي ما تفرزه الثقافات الجماعية. لولبية الفماليوريا.
بشكل عام، البيانات المتعلقة بطبيعة منتجات إفراز النيتروجين متناقضة للغاية. بالإضافة إلى حمض البوليك واليوريا، أشار بعض المؤلفين إلى الأمونيا كمنتجات إخراجية بدلاً من هاتين المادتين (على سبيل المثال، في الجلوكوماو سبيروستوموم).الظرف الوحيد الذي لا يتفق تمامًا مع وظيفة الإخراج للفجوات الانقباضية هو أنها لا تشارك أبدًا في إفراز الأصباغ أثناء الحياة التي يتم إدخالها في البلازما، والتي غالبًا ما تفرز في الحيوانات العليا من خلال الكلى وأعضاء الإخراج الأخرى.
لقد تم التعبير مرارًا وتكرارًا عن رأي مفاده أن الفجوات الانقباضية تلعب دورًا معينًا في عملية التمثيل الغذائي التنفسي للأوالي ، مما يسهل إزالة حمض الكربونيك من السيتوبلازم. ومع ذلك، لا يوجد دليل فعلي على نشاط محدد للفجوة في هذا الاتجاه. فقط البيانات غير المباشرة من لودفيغ (لودفيج، 1928) حول المراسلات بين الكمية الفعلية لحمض الكربونيك التي تفرز من خلال الفجوة والكمية المقدرة لحمض الكربونيك التي تفرزها الحيوانات يتم تفسيرها من قبل هذا المؤلف لصالح الوظيفة التنفسية للفجوة.
يسلط معظم المؤلفين حاليًا الضوء على الدور الهام الذي تلعبه الفجوات الانقباضية كجهاز تنظيم تناضحي. حتى ديجين (1905) أشار إلى أن بلازما الكائنات الأولية في المياه العذبة، بسبب وجود الأملاح فيها، لديها ضغط أسموزي أعلى من البيئة؛ وهذا الظرف يؤدي إلى امتصاص السيتوبلازم المستمر للماء. تعمل الفجوة المقلصة على إزالة الماء الزائد الذي يدخل البلازما بشكل مستمر. يتم تأكيد صحة هذا الرأي من خلال طبيعة توزيع الفجوات المقلصة بين الأوليات، ومن خلال التجارب التي أجريت على تعويد كائنات أولية المياه العذبة على الماء مع زيادة تركيزات الملح (فيريلي، 1930؛ لوفر، 1939، وما إلى ذلك).
في الآونة الأخيرة، دراسة تجريبية لJl. H. سيرافيني (1958) على نبض الفجوة براميسيوم كايداتوم,الذي يوفر بيانات توضح أنه عندما يتم نقل الأهداب إلى المحاليل الملحية ذات التركيز المتزايد، فإنها تتطور خلال أول 15-30 دقيقة. هناك انخفاض عدة مرات في معدل نبض الفجوة، ثم يتسارع معدل النبض تدريجياً ويقترب من السرعة المميزة لأهداب التحكم، لكنه عادة لا يصل إلى مستواه. على سبيل المثال، في 0.2% كلوريد الصوديوم بعد 30 دقيقة. وكان معدل النبض في المتوسط 40.5 ثانية، وبعد 48 ساعة. وصلت إلى 13.1 ثانية، بينما في المجموعة الضابطة استمر النبض بسرعة 10.3 ثانية. وقد لوحظ نفس الشيء في المحاليل 0.1% LiCi، 0.5% CaCl 2، 1% سكروز، ومحلول اليوريا. حدث التباطؤ 7-15 مرة، ولكن بعد يومين
تم إبطاء معدل النبض في هذه المحاليل بمقدار 1.5-3 مرات فقط. في تركيزات الملح المنخفضة، لوحظ الاستعادة الكاملة لمعدل نبض الفجوة. تسببت مواد مثل HGl، وNaOH، وGaGl 2، والفورمالين، بتركيزات ضئيلة، في تباطؤ طفيف في النبض، ثم عاد بسرعة إلى طبيعته. أدى الأدرينالين بتركيز 0.0005٪ إلى زيادة تبادل المياه، ولكن مع مرور الوقت، عاد النبض أيضًا إلى طبيعته. وبالتالي، فإن المواد النشطة وغير النشطة تناضحيًا تسبب تغيرات مماثلة في نشاط الفجوة المقلصة. في هذه الحالة، هناك ميل عام لتنظيم تدفق الماء الذي يمر عبر سيتوبلازم الهدب عن طريق تغيير معدل نبض الفجوة. بالإضافة إلى ذلك، تتكيف الشركات العملاقة مع الظروف المتغيرة للضغط الأسموزي للبيئة الخارجية. وقد لاحظ ذلك أيضًا كيتشينغ (1952) بخصوص الأركسيومو بودوفريا.في هذه الشركات الهدبية، في التركيزات العالية من جلايكول الإيثيلين والسكروز، يتوقف نشاط الفجوة تمامًا، ولكن بعد مرور بعض الوقت يتم استعادة نبض الفجوة وتبادل الماء.
كل هذا يشير إلى آلية عمل معقدة للفجوة المقلصة، والتي تلعب دورا لا شك فيه في تبادل المياه من الشركات الهدبية. ومع ذلك، يعتقد سيرافين، استنادًا إلى حقيقة أن المواد غير النشطة تناضحيًا تسبب تأخيرًا في نبض الفجوة، فمن الممكن افتراض أن تبادل الماء في الشركات الهدبية لا يتم تحديده من خلال الأنماط التناضحية وبالتالي لا يمكن اعتبار دور الفجوة الانقباضية في التنظيم التناضحي مثبتًا .
لفهم وظيفة الفجوة، بالطبع، من الضروري أيضًا فهم آلية دوران الماء من السيتوبلازم إلى خزان الفجوة وآلية تقلص الفجوة نفسها. يعتقد I. Gelei و G. Gelei (Gelei، 1928؛ Gelei، 1939)، مثل عدد من الباحثين الآخرين، أن توتر الامتلاء الناتج في السيتوبلازم، وبالتالي الظروف التناضحية، هو الذي يحدد تقليل خزان الفجوة. من الممكن أن تكون هناك أيضًا آلية نشطة لإفراز الماء في الفجوة (Kitching، 1956). وفقًا لشنايدر (شنايدر، 1950)، فإن نبض الفجوة يحدث بسبب نشاط الألياف الانقباضية الموجودة في جدار الفجوة، ويحدث ملء القنوات الواردة بالماء على النحو التالي. يتجمع السائل في الأنابيب الإرجستوبلازما وبعد ذلك، نظرًا لقطر هذه الأنابيب الأكبر من أنابيب البلازما النيفريدية، يتم إنشاء تدفق الماء من الأنابيب الإرجستوبلازما إلى أنابيب البلازما النيفريدية ومنها إلى تجويف القناة. وبطبيعة الحال، هناك حاجة إلى مزيد من البحث لفهم آلية هذه العمليات بشكل أكثر دقة. يبدو النشاط الإخراجي والتنفسي للفجوة الانقباضية ممكنًا جدًا، ولكن لم يتم إثباته بشكل قاطع.
3) الفجوة الانقباضية وجهاز جولجي
دخلت مسألة الأهمية المورفولوجية للفجوة الانقباضية مرحلة جديدة بعد عمل د. ن. ناسونوف (1924، 1925)، الذي عبر عن فكرة أن الفجوة الانقباضية في الأوليات متماثلة مع جهاز جولجي في الخلايا ميتازوا. Nasonov (1924) وفي نفس الوقت تقريبًا معه I. Gelei (J. Gelei، 1925a، 1925b)، الذي يطبق تقنية التناضح على الشركات العملاقة، اكتشف ميزات جديدة مهمة في بنية الفجوات الانقباضية (الشكل 127). وجد ناسونوف ذلك حول الفجوة الانقباضية للعديد من الشركات العملاقة (كامبانيلا، ليونوتوس، باراميسيوين، ناسولا)،إلى جانب شيلوموناسهو اسودت بقوة من قبل
قشرة أوزميوم مخفضة، ذات سطح أملس أو محاطة بطبقة من الحبيبات الصغيرة المحبة للأوسميوفيلية. تحيط القشرة بالفجوة بالكامل (في الأنواع المذكورة أعلاه)، أو تكون على شكل حلقة شبكية تحيط بالفجوة على طول خط الاستواء (تشيلودون، دوجيليلا).في أشكال مع نظام القنوات المقربة (البرامسيوم)،تستمر القذيفة حتى الأخيرة. في هذه الحالة، يمكن تتبع القنوات في الإندوبلازم بشكل أبعد بكثير من أي طريقة بحث أخرى، وتحاط أجزائها البعيدة بغلاف شبكي رمادي. يعتبر ناسونوف أن هذا القسم من القنوات هو الجزء الإخراجي الفعلي للجهاز، في حين أن الجزء القريب من القنوات والفجوة نفسها هما فقط الجزء الإخراجي من الجهاز الفراغي. ش دوجيليلايمكن أن يذكر ناسونوف أنه أثناء انقباض الفجوة، فإن الحلقة السوداء المحيطة بها لا تتبع الأخيرة أثناء تقلصها، ولكنها تبقى في مكانها الأصلي. حاول جيلي (1925أ، 1925ب) مقارنتها بالجهاز النيفريدي ميتازوا،مساواة الفجوة المقلصة نفسها بالمثانة، وأجزاء أخرى من الجهاز بأقسام أخرى من النيفريديا. ركز ناسونوف على تشابه الغشاء الأسموزي للفجوات مع جهاز جولجي في ميتازوا.في الواقع، فإن الرسومات التي يقدمها لأنواع مختلفة من الفجوات الانقباضية وأجهزة جولجي مأخوذة من أنواع مختلفة من الخلايا في ميتازوا،مقنعة للغاية في تشابهها.

وبناءً على ذلك، يرى ناسونوف أنه من الممكن البحث عن القنوات المقربة في الغشاء الإسفنجي الأسموفيلي باراميكورنوحلقة شبكية دوجيتليلاليس فقط المورفولوجية، ولكن أيضا التوازي الوظيفي بين هذه التشكيلات وجهاز جولجي. يعتمد التركيب الشبكي للمادة الحلقية على وجود فجوات تعليمية صغيرة فيها - مرحلة الإفراز المقيد. ثم تندمج هذه الفجوات لتشكل فجوة مركزية تقع في وسط الحلقة - وهي مرحلة الإفراز الحر المفرز إلى الخارج. وفقًا لمفهوم ناسونوف، يعمل جهاز جولجي على جمع المواد المعروفة من البلازما، والتي يتم تحضيرها فيها لإزالتها من الجسم من خلال فجوة. تم تأكيد الجانب الواقعي بأكمله من النتائج التي توصل إليها ناسونوف بالكامل من خلال الأبحاث اللاحقة التي أجراها عدد من المؤلفين: أ. أ. ستريلكوف (1939) Cycloposlhiidaeفوري فريميت، 1925) فورليسيلا,كينغ (1928) باراميسيوم تريشيومو يوبلوتسإلخ. صحيح أنه في بعض الأوليات لم يكن من الممكن اكتشاف غشاء أوسميوفيل خاص حول الفجوة، الأمر الذي قد يعتمد على القضايا المنهجية.
في خلايا الجميع ميتازواعلى شكل تشكيل شبكي أو يتكون من دكتيوسومات فردية. ولذلك، يمكننا أن نعتقد أنه موجود أيضا في جميع الكائنات وحيدة الخلية. في الواقع، أظهرت الدراسات التي أجراها هيرشلر (1927)، وهوفاس (1937)، وجويت لافيريري (1926) وآخرون وجود العديد من الأوليات في البلازما. (غريجارينا، الكوكسيديا، فلاجيلاتا، ساركودينا)قضبان وحلقات وكرات صغيرة ذات حافة أوسميوفيلية قوية وخلايا تشبه إلى حد كبير الدكتيوسومات ميتازوا.
علامات جهاز جولجي ميتازوابادئ ذي بدء، هذا الجهاز قادر على تقليل مركبات معينة من المعادن الثقيلة - الفضة والأوزميوم، والتي يتم التعبير عنها في اسوداد هياكل الجهاز. هذه الهياكل غير ملطخة بشكل داخلي باللون الأحمر المحايد وأزرق الميثيلين. غالبًا ما تكون الهياكل المميزة لجهاز جولجي عبارة عن دكتيوسومات، والتي تتكون من جزأين - محبة للون وكارهة للون. عادة، يتم غمر الحبيبات الإفرازية في الجزء الكاره للون. كقاعدة عامة، يتم تحديد موقع جهاز جولجي على وجه التحديد في المنطقة الإفرازية للخلية.
أظهرت دراسات المجهر الإلكتروني وحدة هياكل جهاز جولجي في الخلايا المختلفة ميتازوا.في جميع الخلايا المدروسة، تم تحديد الأغشية المزدوجة (أو الأغشية الخلوية)، وتقع بالتوازي مع بعضها البعض وتشكل نظامًا من 3-7 أزواج أو أكثر. يحيط كل زوج بمساحة أخف بعرض 50-200 Å ويشكل نوعًا من الكيس المسطح. في بعض الأحيان يتوسع إلى فجوة كبيرة. يتم غمر الأغشية في مادة أساسية دقيقة الحبيبات. في بعض الحالات، قد تحيط الأغشية بفجوات بأحجام مختلفة. كل هذه الهياكل مستقرة تمامًا.
ومن الجدير بالذكر أن هناك تطابقًا كاملاً بين صور جهاز جولجي الموصوف باستخدام المجهر الضوئي بواسطة ناسونوف وباحثين آخرين في الخلايا ميتازوا،والصور الحديثة التي تم الحصول عليها بالمجهر الإلكتروني.
لاحظ دالتون وفيليكس (1957) أيضًا أن الفجوة الانقباضية الكلاميدوموناسيحدها أغشية مشابهة لتلك الموجودة في جهاز جولجي للخلايا ميتازوا،ومن ثم، يدعم المؤلفون فرضية ناسونوف حول تماثل هذه العضيات. حول الفجوة المقلصة في الشركات الهدبية كامبانيلاو أوفريديومهناك العديد من الأنابيب ذات الجدران الأسموفيلية الكثيفة، والتي يمكن مقارنتها إلى حد ما بالبنى التحتية لجهاز جولجي (Fauré-Fremiet et Rouiller، 1959). البيانات المذكورة أعلاه من شنايدر حول بنية البلازما النيفريدية حول القنوات المقربة للفجوة المقلصة باراميسيمومع ذلك، من الصعب تجانس هيكلها مع البنية التحتية للديكتيوسومات.
وحتى في وقت سابق، رفض المؤلفون الفرنسيون (Duboscq et Grassé، 1933؛ Grassé et Hollande، 1941) أيضًا تماثل الأغشية الأسميوفيلية للفجوات الانقباضية مع جهاز جولجي. وهم يعتقدون أن جدران الفجوات الأسموفيلية، مثل الفجوات الانقباضية نفسها، هي تكوينات مستقلة تمامًا عن جهاز جولجي.
تجدر الإشارة إلى أنه تم العثور على ديكتيوسومات على شكل منجل أو على شكل حلقة في بعض الشركات الهدبية، كما هو الحال في بالانتيديوم إلونجاتوم،خارج الاتصال، مع فجوات (فيلنوف-براشوري، 1940). حدد نيروت-تيموثي (1957) باستخدام المجهر الإلكتروني الهياكل النموذجية لجهاز جولجي في الإندوبلازم لبعض الخلايا. المبيضياتلا يرتبط مع فجوة مقلص.
يتجانس المؤلفون الفرنسيون مع الجسم المجاور للقاعدة مع جهاز جولجي (انظر ص 115)، حيث تبين أن هذا التكوين، خاصة عند فحصه بالمجهر الإلكتروني، يشبه الديكتيوزومات في الخلايا. ميتازوا(الشكل 80).
مما لا شك فيه أن هناك حاجة إلى مزيد من الدراسات المقارنة للطفيليات لحل مشكلة تماثل بعض الهياكل مع جهاز جولجي للخلايا ميتازوا.