Вид, его критерии. Популяция - структурная единица вида и элементарная единица эволюции. Микроэволюция. Образование новых видов. Способы видообразования. Сохранение многообразия видов как основа устойчивости биосферы
Вид, его критерии
Основатель современной систематики К. Линней рассматривал вид как группу сходных по морфологическим признакам организмов, которые свободно скрещиваются между собой. По мере развития биологии были получены свидетельства того, что различия между видами намного глубже, и затрагивают химический состав и концентрацию веществ в тканях, направление и скорость химических реакций, характер и интенсивность процессов жизнедеятельности, число и форму хромосом, т. е. вид является наименьшей группой организмов, отражающей их близкое родство. К тому же виды не существуют вечно - они возникают, развиваются, дают начало новым видам и исчезают.
Вид - это совокупность особей, сходных по строению и особенностям процессов жизнедеятельности, имеющих общее происхождение, свободно скрещивающихся между собой в природе и дающих плодовитое потомство.
Все особи одного вида имеют одинаковый кариотип и занимают в природе определенную географическую область - ареал.
Признаки сходства особей одного вида называют критериями вида . Так как ни один из критериев не является абсолютным, для правильного определения вида необходимо использовать совокупность критериев.
Основными критериями вида являются морфологический, физиологический, биохимический, экологический, географический, этологический (поведенческий) и генетический.
- Морфологический - совокупность внешних и внутренних признаков организмов одного вида. Несмотря на то, что у некоторых видов существуют уникальные признаки, с помощью одних только морфологических черт часто очень трудно различить близкородственные виды. Так, в последнее время открыт ряд видов-двойников, обитающих на одной территории, например домовая и курганчиковая мыши, поэтому использовать исключительно морфологический критерий для определения вида недопустимо.
- Физиологический - сходство процессов жизнедеятельности у организмов, в первую очередь, размножения. Он также не является универсальным, поскольку некоторые виды в природе скрещиваются и дают плодовитое потомство.
- Биохимический - сходство химического состава и протекания процессов обмена веществ. Несмотря на то, что данные показатели могут значительно варьировать у разных особей одного вида, в настоящее время им уделяется большое внимание, поскольку особенности строения и состава биополимеров помогают идентифицировать виды даже на молекулярном уровне и устанавливать степень их родства.
- Экологический - различие видов по их принадлежности к определенным экосистемам и экологическим нишам, которые они занимают. Однако многие неродственные виды занимают сходные экологические ниши, поэтому данный критерий может быть использован для выделения вида только в совокупности с другими признаками.
- Географический - существование популяции каждого вида в определенной части биосферы - ареале, который отличается от ареалов всех остальных видов. В связи с тем, что у множества видов границы ареалов совпадают, а также имеется ряд видов-космополитов, ареал которых охватывает огромные пространства, географический критерий также не может служить маркерным «видовым» признаком.
- Генетический - постоянство признаков хромосомного набора - кариотипа - и нуклеотидного состава ДНК у особей одного вида. В связи с тем, что негомологичные хромосомы не могут конъюгировать во время мейоза, потомство от скрещивания особей разных видов с неодинаковым набором хромосом либо не появляется вовсе, либо не плодовито. Это создает репродуктивную изоляцию вида, поддерживает его целостность и обеспечивает реальность существования в природе. Данное правило может нарушаться в случае скрещивания близких по происхождению видов с одинаковым кариотипом или возникновения различных мутаций, однако исключение лишь подтверждает общее правило, и виды следует рассматривать как устойчивые генетические системы. Генетический критерий является основным в системе критериев вида, но также не исчерпывающим.
При всей сложности системы критериев вид нельзя представить как группу абсолютно идентичных по всем параметрам организмов, то есть клонов. Наоборот, для многих видов характерно значительное разнообразие даже внешних признаков, как, например, для одних популяций божьих коровок характерно преобладание в окраске красного цвета, а для других - черного.
Популяция - структурная единица вида и элементарная единица эволюции
Трудно представить, чтобы в реальности особи одного вида были равномерно распределены по земной поверхности в пределах ареала, поскольку, например, лягушка озерная обитает в основном в достаточно редких стоячих пресных водоемах, и вряд ли ее можно встретить на полях и в лесах. Виды в природе чаще всего распадаются на отдельные группы, в зависимости от подходящих по совокупности условий местам обитания - популяции.
Популяция - группа особей одного вида, занимающих часть его ареала, свободно скрещивающихся между собой и относительно обособленных от других совокупностей особей того же вида в течение более или менее длительного времени.
Популяции могут быть разделены не только пространственно, они могут обитать даже на одной территории, но иметь различия в пищевых предпочтениях, сроках размножения и т. д..
Таким образом, вид - это совокупность популяций особей, обладающих рядом общих морфологических, физиологических, биохимических признаков и типов взаимоотношений с окружающей средой, населяющих определенный ареал, а также способных скрещиваться между собой с образованием плодовитого потомства, но почти или совсем не скрещивающихся с другими группами особей того же вида.
Внутри видов с большими ареалами, охватывающими территории с разными условиями жизни, иногда различают и подвиды - большие популяции или группы соседних популяций, имеющих стойкие морфологические отличия от других популяций.
Популяции разбросаны по земной поверхности не случайным образом, они привязаны к конкретным ее участкам. Совокупность всех факторов неживой природы, необходимых для проживания особей данного вида, называется местообитанием . Однако только этих факторов может быть недостаточно для занятия этого участка популяцией, поскольку она должна быть еще вовлечена в тесное взаимодействие с популяциями других видов, то есть занять определенное место в сообществе живых организмов - экологическую нишу . Так, австралийский сумчатый медведь коала при всех прочих равных условиях не может существовать без своего основного источника питания - эвкалипта.
Образующие неразрывное единство в одних и тех же местообитаниях популяции различных видов обыкновенно обеспечивают более или менее замкнутый круговорот веществ и являются элементарными экологическими системами (экосистемами) - биогеоценозами.
При всей своей требовательности к условиям окружающей среды популяции одного вида неоднородны по занимаемой площади, численности, плотности и пространственному размещению особей, часто образующих более мелкие группы (семьи, стаи, стада и др.), полу, возрасту, генофонду и т. д., поэтому различают их размерную, возрастную, половую, пространственную, генетическую, этологическую и другие структуры, а также динамику.
Важными характирестиками популяции являются генофонд - совокупность генов, характерных для особей данной популяции или вида, а также частоты определенных аллелей и генотипов. Разные популяции одного и того же вида изначально имеют неодинаковый генофонд, так как новые территории осваивают особи со случайными, а не специально подобранными генами. Под действием внутренних и внешних факторов генофонд подвергается еще более существенным изменениям: обогащается за счет возникновения мутаций и нового сочетания признаков и обедняется в результате выпадения отдельных аллелей при гибели или миграции некоторого числа особей.
Новые признаки и их сочетания могут быть полезными, нейтральными или вредными, поэтому в популяции выживают и успешно размножаются только приспособленные к данным условиям окружающей среды особи. Однако в двух различных точках земной поверхности условия окружающей среды никогда не бывают полностью идентичными, поэтому и направление изменений даже в двух соседних популяциях может быть совершенно противоположным или они будут протекать с различной скоростью. Результатом изменений генофонда является расхождение популяций по морфологическим, физиологическим, биохимическим и другим признакам. Если популяции при этом еще и изолированы друг от друга, то они могут давать начало новым видам.
Так, возникновение каких-либо препятствий в скрещивании особей различных популяций одного вида, например, вследствие образования горных массивов, изменения русел рек, различий в сроках размножения и т. д., приводит к тому, что популяции постепенно приобретают все больше различий и, в конце концов, становятся различными видами. Некоторое время на границах этих популяций происходит скрещивание особей и возникают гибриды, однако со временем и эти контакты исчезают, т. е. популяции из открытых генетических систем становятся закрытыми.
Несмотря на то, что действию факторов окружающей среды подвергаются в первую очередь отдельные особи, изменение генетического состава у единичного организма является незначительным и проявится в лучшем случае только у его потомков. Подвиды, виды и более крупные таксоны также не подходят на роль элементарных единиц эволюции, поскольку они не отличаются морфологическим, физиологическим, биохимическим, экологическим, географическим и генетическим единством, тогда как популяции как наименьшие структурные единицы вида, накапливающие разнообразие случайных изменений, худшие из которых будут отсеяны, соответствуют этому условию и являются элементарными единицами эволюции.
Микроэволюция
Изменение генетической структуры популяций не всегда приводит к образованию нового вида, а может лишь улучшать приспособление популяции к конкретным условиям среды, однако виды не являются вечными и неизменными - они способны развиваться. Этот процесс необратимого исторического изменения живого называется эволюцией. Первично эволюционные преобразования происходят внутри вида на уровне популяций. В их основе лежат, прежде всего, мутационный процесс и естественный отбор, приводящие к изменению генофонда популяций и вида в целом, или даже к образованию новых видов. Совокупность этих элементарных эволюционных событий называется микроэволюцией.
Популяции характеризуются огромным генетическим разнообразием, которое часто не проявляется фенотипически. Генетическое разнообразие возникает вследствие спонтанного мутагенеза, который происходит непрерывно. Большинство мутаций неблагоприятны для организма и снижают жизнеспособность популяции в целом, но, если они рецессивны, то могут сохраняться в гетерозиготе в течение продолжительного времени. Некоторые мутации, не имеющие приспособительной ценности в данных условиях существования, способны приобрести такую ценность в будущем или при освоении новых экологических ниш, создавая, таким образом, резерв наследственной изменчивости.
Значительное влияние на микроэволюционные процессы оказывают колебания численности особей в популяциях, миграции и катастрофы, а также изоляция популяций и видов.
Новый вид является промежуточным результатом эволюции, но никак не ее итогом, поскольку на этом микроэволюция не прерывается - она продолжается далее. Возникающие новые виды в случае удачного сочетания признаков заселяют новые местообитания, и, в свою очередь, дают начало новым видам. Такие группы близкородственных видов объединяются в роды, семейства и т. д. Эволюционные процессы, происходящие в надвидовых группах, называются уже макроэволюцией. В отличие от макроэволюции, микроэволюция протекает в гораздо более сжатые сроки, тогда как первой требуются десятки и сотни тысяч и миллионов лет, как, например, эволюция человека.
В результате микроэволюции формируется все многообразие видов живых организмов, когдалибо существовавших и ныне живущих на Земле.
Вместе с тем эволюция необратима, и уже исчезнувшие виды никогда не возникают вновь. Появляющиеся виды закрепляют все достигнутое в процессе эволюции, однако это не гарантирует того, что в будущем не появятся новые виды, которые будут иметь более совершенные приспособления к условиям окружающей среды.
Образование новых видов
В широком смысле под образованием новых видов понимается не только отщепление от основного ствола вида нового или распад материнского вида на несколько дочерних, а и общее развитие вида как целостной системы, приводящее к существенным изменениям его морфоструктурной организации. Однако чаще все же видообразование рассматривают как процесс формирования новых видов посредством разветвления «родословного древа» вида.
Принципиальное решение проблемы видообразования было предложено Ч. Дарвином. Согласно его теории расселение особей одного вида приводит к образованию популяций, которые вследствие различий условий окружающей среды вынуждены адаптироваться к ним. Это, в свою очередь, влечет за собой обострение внутривидовой борьбы за существование, направляемой естественным отбором. В настоящее время считается, что борьба за существование вовсе не является обязательным фактором видообразования, наоборот, давление отбора в ряде популяций может снижаться. Различие условий существования способствует возникновению неодинаковых приспособительных изменений в популяциях вида, следствием которого является расхождение признаков и свойств популяций - дивергенция.
Однако накопления различий, даже на генетическом уровне, отнюдь не достаточно для появления нового вида. До тех пор, пока различающиеся по каким-либо признакам популяции не только контактируют, но и способны к скрещиванию с образованием плодовитого потомства, они относятся к одному виду. Лишь невозможность перетока генов из одной группы особей в другую, даже в случае разрушения разделяющих их преград, т. е. скрещивания, означает завершение сложнейшего эволюционного процесса формирования нового вида.
Видообразование является продолжением микроэволюционных процессов. Существует точка зрения, что видообразование не может быть сведено к микроэволюции, оно представляет качественный этап эволюции и осуществляется благодаря другим механизмам.
Способы видообразования
Выделяют два основных способа видообразования: аллопатрическое и симпатрическое.
Аллопатрическое , или географическое видообразование является следствием пространственного разделения популяций физическими преградами (горные хребты, моря и реки) вследствие их возникновения или расселения в новые места обитания (географическая изоляция). Поскольку в данном случае генофонд отделившейся популяции существенно отличается от материнской, да и условия в месте ее обитания не будут совпадать с исходными, со временем это приведет к дивергенции и формированию нового вида. Ярким примером географического видообразования является обнаруженное Ч. Дарвином во время путешествия на корабле «Бигль» разнообразие видов вьюрков на Галапагосских островах вблизи побережья Эквадора. По-видимому, отдельные особи единственного обитающего на южноамериканском континенте вьюрка каким-то образом попали на острова, и, вследствие различия условий (в первую очередь доступности пищи) и географической изоляции, они постепенно эволюционировали, образовав группу родственных видов.
В основе симпатрического , или биологического видообразования лежит какая-либо из форм репродуктивной изоляции, при этом новые виды возникают внутри ареала исходного вида. Обязательным условием симпатрического видообразования является быстрая изоляция образовавшихся форм. Это более быстрый процесс, чем аллопатрическое видообразование, а новые формы похожи на исходные предковые.
Симпатрическое видообразование может быть вызвано быстрыми изменениями хромосомного набора (полиплоидизация) или хромосомными перестройками. Иногда новые виды возникают вследствие гибридизации двух исходных видов, как, например, у домашней сливы, являющейся гибридом терна и алычи. В некоторых случаях симпатрическое видообразование связано с разделением экологических ниш у популяций одного вида в пределах единого ареала или сезонной изоляции - расхождения сроков репродукции у растений (разные виды сосны в Калифорнии пылят в феврале и апреле) и сроков размножения у животных.
Из всего многообразия вновь возникающих видов только немногие, наиболее приспособленные, могут существовать длительное время и дать начало новым видам. Причины гибели большинства видов до сих пор неизвестны, скорее всего это обусловлено резкими изменениями климата, геологическими процессами и вытеснением их более приспособленными организмами. В настоящее время одной из причин гибели значительного числа видов является человек, который истребляет наиболее крупных животных и самые красивые растения, и если в XVII веке этот процесс только начался истреблением последнего тура, то в XXI веке ежечасно исчезает более 10 видов.
Сохранение многообразия видов как основа устойчивости биосферы
Несмотря на то, что на планете, по разным оценкам, обитает 5–10 млн еще не описанных видов организмов, о существовании большинства из них мы никогда не узнаем, поскольку ежечасно с лица Земли исчезает около 50 видов. Исчезновение живых организмов в настоящее время совсем не обязательно связано с их физическим истреблением, чаще это обусловлено разрушением в результате человеческой деятельности их природных мест обитания. Гибель отдельного вида вряд ли способна привести к фатальным последствиям для биосферы, однако уже давно установлено, что вымирание одного вида растений влечет за собой гибель 10–12 видов животных, а это уже представляет угрозу как для существования отдельных биогеоценозов, так и для глобальной экосистемы в целом.
Накопленные за предыдущие десятилетия печальные факты вынудили Международный союз охраны природы и природных ресурсов (МСОП) начать в 1949 году сбор информации о редких и находящихся под угрозой исчезновения видах растений и животных. В 1966 году МСОП издал первую «Красную книгу фактов».
Красная книга - это официальный документ, содержащий регулярно обновляемые данные о состоянии и распространении редких и находящихся под угрозой исчезновения видов растений, животных и грибов.
В этом документе принята пятиступенчатая шкала статуса охраняемого вида, причем к первой ступени охраны относят виды, спасение которых невозможно без осуществления специальных мероприятий, а к пятой - восстановленные виды, состояние которых благодаря принятым мерам не вызывает опасений, но они еще не подлежат промышленному использованию. Разработка такой шкалы позволяет направить первоочередные усилия в области охраны именно на самые редкие виды, такие как амурские тигры.
Помимо международного варианта Красной книги, существуют также национальные и региональные ее варианты. В СССР Красная книга была учреждена в 1974 году, а в Российской Федерации порядок ее ведения регламентируется Федеральными законами «Об охране окружающей среды», «О животном мире» и Постановлением Правительства РФ «О Красной книге Российской Федерации». Сегодня в Красную книгу РФ занесено 610 видов растений, 247 видов животных, 42 вида лишайников и 24 вида грибов. Популяции некоторых из них, в свое время оказавшиеся под угрозой исчезновения (бобр европейский, зубр), уже довольно успешно восстановлены.
Под охрану в России взяты следующие виды животных: русская выхухоль, тарбаган (монгольский сурок), белый медведь, кавказская европейская норка, калан, манул, амурский тигр, леопард, снежный барс, сивуч, морж, тюлени, дельфины, киты, лошадь Пржевальского, кулан, розовый пеликан, фламинго обыкновенный, аист черный, лебедь малый, орел степной, беркут, журавль черный, стерх, дрофа, филин, чайка белая, черепаха средиземноморская, полоз японский, гюрза, жаба камышовая, минога каспийская, все виды осетровых рыб, лосось озерный, жук-олень, шмель необыкновенный, аполлон обыкновенный, рак-богомол, жемчужница обыкновенная и др.
К растениям Красной книги РФ относятся 7 видов подснежников, некоторые виды полыни, женьшень настоящий, 7 видов колокольчиков, дуб зубчатый, пролеска пролесковидная, 11 видов касатиков, рябчик русский, тюльпан Шренка, лотос орехоносный, башмачок венерин настоящий, пион тонколистный, ковыль перистый, первоцвет Юлии, прострел (сон-трава) луговой, красавка белладонна, сосна пицундская, тис ягодный, щитовник китайский, полушник озерный, сфагнум мягкий, филлофора курчавая, хара нитевидная и др.
Редкие грибы представлены трюфелем летним, или русским черным трюфелем, трутовиком лакированным и др.
Охрана редких видов в большинстве случаев связана с запретом их уничтожения, сохранением их самих в искусственно созданной среде обитания (зоопарках), охране их местообитаний и созданием низкотемпературных генетических банков.
Наиболее эффективной мерой охраны редких видов является сохранение их местообитаний, которое достигается организацией сети особо охраняемых заповедных территорий, имеющих, в соответствии с Федеральным законом «Об особо охраняемых природных территориях» (1995), международное, федеральное, региональное или местное значение. К ним относятся государственные природные заповедники, национальные парки, природные парки, государственные природные заказники, памятники природы, дендрологические парки, ботанические сады и др.
Государственный природный заповедник - это полностью изъятый из хозяйственного использования особо охраняемый природный комплекс (земля, водные объекты, недра, растительный и животный мир), имеющий природоохранное, научное, эколого-просветительское значение как образец естественной природной среды, типичные или редкие ландшафты, места сохранения генетического фонда растительного и животного мира.
Заповедники, которые входят в международную систему биосферных резерватов, осуществляющих глобальный экологический мониторинг, имеют статус государственных природных биосферных заповедников . Заповедник является природоохранным, научно-исследовательским и эколого- просветительским учреждением, имеющим целью сохранение и изучение естественного хода природных процессов и явлений, генетического фонда растительного и животного мира, отдельных видов и сообществ растений и животных, типичных и уникальных экологических систем.
В настоящее время в России насчитывается около 100 государственных природных заповедников, 19 из которых носят статус биосферного, в т. ч. Байкальский, Баргузинский, Кавказский, «Кедровая падь», Кроноцкий, Приокско-Террасный и др.
В отличие от заповедников, территории (акватории) национальных парков включают в себя природные комплексы и объекты, имеющие особую экологическую, историческую и эстетическую ценности, и предназначены для использования в природоохранных, просветительских, научных и культурных целях и для регулируемого туризма. Такой статус имеют 39 особо охраняемых природных территорий, в т. ч. Забайкальский и Сочинский национальные парки, а также национальные парки «Куршская коса», «Русский Север», «Шушенский бор» и др.
Природные парки являются природоохранными рекреационными учреждениями, находящимися в ведении субъектов Российской Федерации, территории (акватории) которых включают в себя природные комплексы и объекты, имеющие значительную экологическую и эстетическую ценности, и предназначены для использования в природоохранных, просветительских и рекреационных целях.
Государственными природными заказниками являются территории (акватории), имеющие особое значение для сохранения или восстановления природных комплексов или их компонентов и поддержания экологического баланса.
Развитие эволюционных идей. Значение эволюционной теории Ч. Дарвина. Взаимосвязь движущих сил эволюции. Формы естественного отбора, виды борьбы за существование. Синтетическая теория эволюции. Элементарные факторы эволюции. Исследования С. С. Четверикова. Роль эволюционной теории в формировании современной естественнонаучной картины мира
Развитие эволюционных идей
Все теории происхождения и развития органического мира могут быть сведены к трем основным направлениям: креационизму, трансформизму и эволюционизму. Креационизм - это концепция постоянства видов, рассматривающая многообразие органического мира как результат его творения Богом. Данное направление сформировалось в результате установления господства христианской церкви в Европе, опиравшейся на библейские тексты. Видными представителями креационизма были К. Линней и Ж. Кювье.
«Князь ботаников» К. Линней, открывший и описавший сотни новых видов растений, и создавший первую стройную их систему, тем не менее, доказывал, что общее количество видов организмов неизменно со времени сотворения Земли, то есть они не только не появляются вновь, но и не исчезают. Только к концу жизни он пришел к заключению, что делом рук Божьих являются роды, тогда как виды могут развиваться вследствие приспособления к местным условиям.
Вкладом выдающегося французского зоолога Ж. Кювье (1769–1832) в биологию было опиравшееся на многочисленные данные палеонтологии, сравнительной анатомии и физиологии учение о корреляциях - взаимосвязях частей организма. Благодаря этому стало возможным реконструировать по отдельным частям внешний облик животного. Однако в процессе палеонтологических исследований Ж. Кювье не мог не обратить внимания как на явное изобилие ископаемых форм, так и на резкие изменения групп животных в течение геологической истории. Эти данные послужили отправной точкой для формулировки теории катастроф , согласно которой все или почти все организмы на Земле неоднократно погибали в результате периодических стихийных бедствий, а потом планета вновь заселялась видами, пережившими катастрофу. Последователи Ж. Кювье насчитывали до 27 таких катастроф в истории Земли. Соображения же об эволюции представлялись Ж. Кювье оторванными от реальности.
Противоречия в исходных положениях креационизма, которые становились все более очевидными по мере накопления научных фактов, послужили отправной точкой для формирования другой системы взглядов - трансформизма , признающего реальное существование видов и их историческое развитие. Представители данного течения - Ж. Бюффон, И. Гете, Э. Дарвин и Э. Жоффруа Сент-Илер, будучи не в силах вскрыть истинные причины эволюции, сводили их к приспособлению к условиям окружающей среды и наследованию приобретенных признаков. Корни трансформизма можно найти в работах еще древнегреческих и средневековых философов, которые признавали исторические изменения органического мира. Так, Аристотель высказал идею о единстве природы и постепенности перехода от тел неживой природы к растениям, а от них - к животным - «лестнице природы». Основной причиной изменений живых организмов он считал их внутреннее стремление к совершенству.
Французский естествоиспытатель Ж. Бюффон (1707–1788), главным трудом жизни которого была 36-томная «Естественная история», вопреки представлениям креационистов, раздвинул рамки истории Земли до 80–90 тыс. лет. При этом он констатировал единство растительного и животного мира, а также возможность изменения родственных организмов под воздействием факторов окружающей среды в результате одомашнивания и гибридизации.
Английский врач, философ и поэт Э. Дарвин (1731–1802), дед Ч. Дарвина, полагал, что история органического мира насчитывает миллионы лет, а разнообразие животного мира является результатом смешения нескольких «естественных» групп, влияния внешней среды, упражнения и неупражнения органов, и других факторов.
Единство плана строения групп животных считал одним из главных доказательств развития живого мира Э. Жоффруа Сент-Илер (1772–1844). Однако, в отличие от своих предшественников, он склонялся к тому, что изменение видов обусловлено воздействием факторов среды не на взрослые особи, а на зародыши.
Несмотря на то, что в разгоревшейся в 1831 году дискуссии между Ж. Кювье и Э. Жоффруа Сент-Илером в виде серии докладов в Академии наук, явный перевес остался на стороне первого, именно трансформизм стал предтечей эволюционизма. Эволюционизм (теория эволюции, эволюционное учение) - это система взглядов, признающая развитие природы по определенным законам. Он является теоретической вершиной биологии, которая позволяет объяснять наблюдаемые нами многообразие и сложность живых систем. Однако в связи с тем, что эволюционное учение описывает труднодоступные для наблюдения явления, оно сталкивается со значительными затруднениями. Иногда теорию эволюции называют «дарвинизмом» и отождествляют с учением Ч. Дарвина, что является в корне неверным, поскольку, хотя теория Ч. Дарвина и внесла неоценимый вклад в развитие не только эволюционного учения, но и биологии в целом (а также многих других наук), основы эволюционной теории были заложены другими учеными, она продолжает развиваться и по сей день, а «дарвинизм» во многих аспектах имеет лишь историческое значение.
Создателем первой эволюционной теории - ламаркизма - был французский естествоиспытатель Ж. Б. Ламарк (1744–1829). Движущей силой эволюции он считал внутреннее стремление организмов к совершенству (закон градации ), однако приспособление к условиям внешней среды вынуждает их к отклонению от этой магистральной линии. При этом органы, которые интенсивно используются животным в процессе жизнедеятельности, развиваются, а ненужные ему, наоборот, ослабляются и даже могут исчезать (закон упражнения и неупражнения органов ). Приобретенные в процессе жизни признаки закрепляются и передаются потомкам. Так, наличие перепонок между пальцами ног у водоплавающих птиц он объяснял попытками их предков передвигаться в водной среде, а длинная шея жирафов, по Ламарку, является следствием того, что их предки старались доставать листья с верхушек деревьев.
Недостатками ламаркизма были теоретичность многих построений, а также допущение вмешательства Творца в эволюцию. В процессе развития биологии выяснилось, что индивидуальные изменения, приобретаемые организмами в процессе жизни, большей частью укладываются в пределы фенотипической изменчивости, а их передача практически невозможна. Например, немецкий зоолог и теоретик эволюционного учения А. Вейсман (1834–1914) на протяжении многих поколений отрезал мышам хвосты и всегда получал в потомстве только хвостатых грызунов. Теория Ж. Б. Ламарка не была принята современниками, но на рубеже веков легла в основу так называемого неоламаркизма.
Значение эволюционной теории Ч. Дарвина
Предпосылками для создания наиболее известной эволюционной теории Ч. Дарвина, или дарвинизма, стали публикация в 1778 году труда английского экономиста Т. Мальтуса «Трактат о народонаселении», работы геолога Ч. Лайеля, формулировка клеточной теории, успехи селекции в Англии и собственные наблюдения Ч. Дарвина (1809–1882), сделанные в годы учебы в Кембридже, в экспедиции в качестве натуралиста на корабле «Бигль» и по ее завершении.
Так, Т. Мальтус утверждал, что население Земли увеличивается в геометрической прогрессии, что существенно превышает возможности планеты по обеспечению его продовольствием и ведет к гибели части потомства. Параллели, проведенные Ч. Дарвиным и его соавтором А. Уоллесом (1823–1913), указывали на то, что и в природе особи размножаются с очень большой скоростью, однако численность популяций остается относительно постоянной. Исследования английского геолога Ч. Лайеля позволили установить, что поверхность Земли далеко не всегда была такой, как в настоящее время, а ее изменения были вызваны воздействием воды, ветра, вулканическими извержениями и деятельностью живых организмов. Самого же Ч. Дарвина еще в студенческие годы поразила крайняя степень изменчивости жуков, а во время путешествия - сходство флоры и фауны континентальной Южной Америки и лежащих вблизи нее Галапагосских островов, и в то же время значительное разнообразие видов, например вьюрков и черепах. Кроме того, в экспедиции он мог наблюдать скелеты гигантских вымерших млекопитающих, сходных с современными броненосцами и ленивцами, что существенно поколебало его веру в сотворение видов.
Основные положения теории эволюции были высказаны Ч. Дарвином в 1859 году на заседании Лондонского Королевского общества, а впоследствии развиты в книгах «Происхождение видов путем естественного отбора, или сохранение благоприятствуемых пород в борьбе за жизнь» (1859), «Изменение домашних животных и культурных растений» (1868), «Происхождение человека и половой отбор» (1871), «Выражение эмоций у человека и животных» (1872) и др.
Сущность разработанной Ч. Дарвиным концепции эволюции можно свести к ряду вытекающих друг из друга положений, имеющих соответствующие доказательства:
- Особи, составляющие любую популяцию, производят гораздо больше потомков, нежели это необходимо для поддержания численности популяции.
- В связи с тем, что жизненные ресурсы для любого вида живых организмов ограничены, между ними неизбежно возникает борьба за существование . Ч. Дарвин различал внутривидовую и межвидовую борьбу, а также борьбу с факторами окружающей среды. При этом он указывал и на то, что речь идет не только о борьбе конкретной особи за существование, но и за оставление потомства.
- Следствием борьбы за существование является естественный отбор - преобладающее выживание и размножение организмов, случайно оказавшихся наиболее приспособленными к данным условиям существования. Естественный отбор во многом аналогичен искусственному, который человек использует с глубокой древности для выведения новых сортов растений и пород домашних животных. Отбирая особей, которые имеют какой-нибудь желательный признак, человек сохраняет эти признаки путем искусственного разведения с помощью избирательного размножения или опыления. Особой формой естественного отбора является половой отбор по признакам, которые обычно не имеют прямого приспособительного значения (длинные перья, огромные рога и т. п.), однако способствуют достижению успеха в размножении, поскольку делают особь более привлекательной для противоположного пола или более грозной для соперников того же пола.
- Материалом для эволюции служат различия организмов, возникающие вследствие их изменчивости. Ч. Дарвин различал неопределенную и определенную изменчивости. Определенная (групповая) изменчивость проявляется у всех особей вида одинаково под действием определенного фактора и исчезает у потомков, когда действие этого фактора прекращается. Неопределенная (индивидуальная) изменчивость - это изменения, которые возникают у каждой отдельной особи, независимо от колебаний значений факторов окружающей среды, и передаются потомкам. Такая изменчивость не имеет приспособительного (адаптивного) характера. Впоследствии выяснилось, что определенная изменчивость является ненаследственной, а неопределенная - наследственной.
- Естественный отбор в конечном итоге приводит к расхождению признаков отдельных изолированных разновидностей - дивергенции, и, в конце концов - к образованию новых видов.
Теория эволюции Ч. Дарвина не только постулировала процесс возникновения и развития видов, но и вскрыла сам механизм эволюции, который зиждется на принципе естественного отбора. Дарвинизм также отрицал запрограммированность эволюции и постулировал ее непрерывный характер.
Вместе с тем эволюционная теория Ч. Дарвина не могла ответить на целый ряд вопросов, например, о природе генетического материала и его свойствах, сущности наследственной и ненаследственной изменчивости, их эволюционной роли. Это привело к кризису дарвинизма и возникновению новых теорий: неоламаркизма, сальтационизма, концепции номогенеза и др. Неоламаркизм основывается на положении теории Ж. Б. Ламарка о наследовании приобретенных признаков. Сальтационизм - это система взглядов на процесс эволюции как скачкообразные изменения, приводящие к быстрому появлению новых видов, родов и более крупных систематических групп. Концепция номогенеза постулирует запрограммированность направления эволюции и развитие различных признаков на основе внутренних законов. Лишь синтез дарвинизма и генетики в 20–30-е годы ХХ века смог преодолеть противоречия, неизбежно возникавшие при объяснении ряда фактов.
Взаимосвязь движущих сил эволюции
Эволюция не может быть связана с действием какого-либо одного фактора, поскольку мутации сами по себе являются случайными и ненаправленными изменениями, и не могут обеспечивать приспособление особей к факторам среды, тогда как естественный отбор уже сортирует эти изменения. Равным образом и сам отбор не может быть единственным фактором эволюции, поскольку для отбора нужен соответствующий материал, поставляемый мутациями.
Тем не менее можно отметить, что мутационный процесс и поток генов создают изменчивость, тогда как естественный отбор и дрейф генов сортируют эту изменчивость. Это значит, что факторы, создающие изменчивость, запускают процесс микроэволюции, а сортирующие изменчивость продолжают ее, приводя к установлению новых частот вариантов. Таким образом, эволюционное изменение в пределах популяции можно рассматривать как результат действия противоположных сил, создающих и сортирующих генотипическую изменчивость.
Примером взаимодействия между мутационным процессом и отбором служит гемофилия у человека. Гемофилия - это заболевание, обусловленное пониженной свертываемостью крови. Ранее оно приводило к смерти в дорепродуктивном периоде, поскольку любое повреждение мягких тканей потенциально могло привести к большой кровопотере. Это заболевание обусловлено рецессивной мутацией сцепленного с полом гена Н (Xh). Женщины болеют гемофилией крайне редко, чаще бывают гетерозиготными носительницами, однако их сыновья могут унаследовать данное заболевание. Теоретически в течение нескольких поколений такие мужчины умирают до половой зрелости и постепенно данный аллель должен исчезнуть из популяции, однако частота встречаемости этого заболевания не снижается ввиду повторных мутаций в данном локусе, как это произошло у королевы Виктории, передавшей заболевание трем поколениям королевских домов Европы. Постоянная частота данного заболевания свидетельствует о равновесии между мутационным процессом и давлением отбора.
Формы естественного отбора, виды борьбы за существование
Естественным отбором называют избирательное переживание и оставление потомства наиболее приспособленными особями и гибель наименее приспособленных.
Сущность естественного отбора в теории эволюции заключается в дифференцированном (неслучайном) сохранении в популяции определенных генотипов и избирательном их участии в передаче генов следующему поколению. При этом он влияет не на отдельный признак (или ген), а на весь фенотип, который формируется вследствие взаимодействия генотипа с факторами окружающей среды. Естественный отбор в разных условиях среды будет носить неодинаковый характер. В настоящее время различают несколько форм естественного отбора: стабилизирующий, движущий и разрывающий.

Стабилизирующий отбор направлен на закрепление узкой нормы реакции, которая оказалась наиболее благоприятной при данных условиях существования. Он характерен для тех случаев, когда фенотипические признаки являются оптимальными для неизменяющихся условий среды. Ярким примером действия стабилизирующего отбора является сохранение относительно постоянной температуры тела теплокровных животных. Эта форма отбора была детально исследована выдающимся отечественным зоологом И. И. Шмальгаузеном.
Движущий отбор возникает в ответ на изменения условий среды, в результате чего сохраняются мутации, отклоняющиеся от среднего значения признака, тогда как господствовавшая ранее форма подвергается уничтожению, так как недостаточно отвечает новым условиям существования. Например, в Англии в результате загрязнения воздуха промышленными выбросами широко распространились доселе невиданные во многих местах бабочки березовые пяденицы с темной окраской крыльев, которые были хуже заметны для птиц на фоне закопченных стволов берез. Движущий отбор не способствует полному уничтожению формы, против которой он действует, поскольку вследствие мер, предпринятых правительством и природоохранными организациями, ситуация с загрязнением атмосферы резко улучшилась, и наблюдается возвращение окраски крыльев бабочек к исходному варианту.
Разрывающий , или дизруптивный отбор благоприятствует сохранению крайних вариантов признака и удаляет промежуточные, как, например, в результате применения ядохимикатов появляются устойчивые к нему группы особей насекомых. По своему механизму дизруптивный отбор является противоположностью стабилизирующему отбору. Благодаря этой форме отбора в популяции возникают несколько резко разграниченных фенотипов. Это явление называется полиморфизмом . Возникновение репродуктивной изоляции между резко выраженными формами может приводить к видообразованию.
Иногда также отдельно рассматривают дестабилизирующий отбор , который сохраняет мутации, приводящие к широкому разнообразию какого-либо признака, например, расцветки и строения раковин некоторых моллюсков, живущих в разнородных микроусловиях скалистой полосы прибоя моря. Эта форма отбора была открыта Д. К. Беляевым при изучении одомашнивания животных.
В природе ни одна из форм естественного отбора не существует в чистом виде, а наоборот, имеются разнообразные их комбинации, и по мере изменения условий среды на первый план выходит то одна, то другая из них. Так, по завершении изменений в среде движущий отбор сменяется стабилизирующим, который оптимизирует группу особей в новых условиях существования.
Естественный отбор происходит на различных уровнях, в связи с чем различают также индивидуальный, групповой и половой отбор. Индивидуальный отбор устраняет менее приспособленные особи от участия в размножении, тогда как групповой направлен на сохранение признака, полезного не отдельной особи, а группе в целом. Под давлением группового отбора могут полностью вымирать целые популяции, виды и более крупные группы организмов без оставления потомства. В отличие от индивидуального отбора, групповой сокращает разнообразие форм в природе.
Половой отбор осуществляется внутри одного пола. Он способствует выработке признаков, которые обеспечивают успех в оставлении наибольшего потомства. Благодаря этой форме естественного отбора развился половой диморфизм, выражающийся в величине и окраске хвоста у павлина, рогов у оленей и т. д.
Естественный отбор является результатом борьбы за существование на основе наследственной изменчивости. Под борьбой за существование понимают всю совокупность взаимосвязей между особями своего и других видов, а также с абиотическими факторами окружающей среды. Эти отношения определяют успех или неудачу определенной особи в выживании и оставлении потомства. Причиной борьбы за существование является появление избыточного числа особей по отношению к доступным ресурсам. Помимо конкуренции, в эти взаимоотношения следует включать и взаимопомощь, которая повышает шансы особей на выживание.
Взаимодействие с факторами окружающей среды также может приводить к гибели подавляющего большинства особей, например, у насекомых, лишь незначительная часть которых переживает зиму.
Синтетическая теория эволюции
Успехи генетики в начале ХХ века, например открытие мутаций, наводили на мысль о том, что наследственные изменения фенотипа организмов происходят внезапно, а не формируются длительное время, как это постулировала эволюционная теория Ч. Дарвина. Однако дальнейшие исследования в области популяционной генетики привели к формулировке в 20–50-е годы ХХ века новой системы эволюционных воззрений - синтетической теории эволюции . Значительный вклад в ее создание внесли ученые разных стран: советские ученые С. С. Четвериков, И. И. Шмальгаузен и А. Н. Северцов, английский биохимик и генетик Д. Холдейн, американские генетики С. Райт и Ф. Добржанский, эволюционист Д. Хаксли, палеонтолог Д. Симпсон и зоолог Э. Майр.
Основные положения синтетической теории эволюции:
- Элементарным материалом эволюции является наследственная изменчивость (мутационная и комбинативная) у особей популяции.
- Элементарной единицей эволюции является популяция, в которой происходят все эволюционные изменения.
- Элементарное эволюционное явление - изменение генетической структуры популяции.
- Элементарные факторы эволюции - дрейф генов, волны жизни, поток генов - имеют ненаправленный, случайный характер.
- Единственным направленным фактором эволюции является естественный отбор, который носит творческий характер. Естественный отбор бывает стабилизирующим, движущим и разрывающим (дизруптивным).
- Эволюция имеет дивергентный характер, то есть один таксон может дать начало нескольким новым таксонам, тогда как каждый вид имеет только одного предка (вид, популяцию).
- Эволюция имеет постепенный и продолжительный характер. Видообразование как этап эволюционного процесса является последовательной сменой одной популяции чередой других временных популяций.
- Различают два вида эволюционного процесса: микроэволюция и макроэволюция. Макроэволюция не имеет своих особых механизмов и осуществляется только благодаря микроэволюционным механизмам.
- Любая систематическая группа может или процветать (биологический прогресс), или вымирать (биологический регресс). Биологический прогресс достигается благодаря изменениям в строении организмов: ароморфозам, идиоадаптациям или общей дегенерации.
- Основными закономерностями эволюции являются ее необратимый характер, прогрессивное усложнение форм жизни и развитие приспособленности видов к среде обитания. Вместе с тем эволюция не имеет конечной цели, т. е. процесс это ненаправленный.
Несмотря на то, что эволюционная теория за прошедшие десятилетия обогатилась данными смежных наук - генетики, селекции и др., все же она не учитывает целого ряда аспектов, например направленного изменения наследственного материала, поэтому в будущем возможно создание новейшей концепции эволюции, которая заменит синтетическую теорию.
Элементарные факторы эволюции
Согласно синтетической теории эволюции элементарное эволюционное явление заключается в изменении генетического состава популяции, а события и процессы, которые приводят к изменению генофондов, называются элементарными факторами эволюции . К ним относятся мутационный процесс, популяционные волны, дрейф генов, изоляция и естественный отбор. Ввиду исключительности значения естественного отбора в эволюции он будет рассмотрен отдельно.
Мутационный процесс, который так же непрерывен, как и сама эволюция, поддерживает генетическую неоднородность популяции за счет возникновения все новых и новых вариантов генов. Мутации, возникающие под воздействием внешних и внутренних факторов, относят к генным, хромосомным и геномным.
Генные мутации возникают с частотой 10 –4 –10 –7 на одну гамету, однако, ввиду того, что у человека и большинства высших организмов общее число генов может достигать нескольких десятков тысяч, невозможно себе представить, чтобы два организма были абсолютно идентичными. Большинство возникающих мутаций является рецессивными, тем более что доминантные мутации тотчас же подвергаются естественному отбору. Рецессивные мутации и создают тот самый резерв наследственной изменчивости, однако прежде чем проявиться в фенотипе, они должны закрепиться у многих особей в гетерозиготном состоянии благодаря свободному скрещиванию в популяции.
Хромосомные мутации, связанные с потерей или переносом части хромосомы (целой хромосомы) на другую, также довольно часты у различных организмов, например, разница между некоторыми видами крыс заключается в единственной паре хромосом, что затрудняет их скрещивание.
Геномные мутации , связанные с полиплоидизацией, также ведут к репродуктивной изоляции вновь возникшей популяции ввиду нарушений в митозе первого деления зиготы. Тем не менее у растений они довольно широко распространены и такие растения могут произрастать в Арктике и на альпийских лугах ввиду их большей устойчивости к факторам окружающей среды.
Комбинативная изменчивость, которая обеспечивает возникновение новых вариантов объединения генов в генотипе, и, соответственно, повышает вероятность появления новых фенотипов, также вносит свой вклад в эволюционные процессы, поскольку только у человека количество вариантов комбинаций хромосом составляет 2 23 , то есть появление организма, аналогичного уже существующему, практически невозможно.
Популяционные волны. К противоположному результату (обеднению генного состава) приводят зачастую колебания численности организмов в природных популяциях, которая у некоторых видов (насекомые, рыбы и др.) может изменяться в десятки и сотни раз - популяционные волны , или «волны жизни» . Увеличение или уменьшение численности особей в популяциях может быть как периодическим , так и непериодическим . Первые являются сезонными или многолетними, как, например, миграции у перелетных птиц, или размножение у дафний, у которых весной и летом имеются только женские особи, а к осени появляются мужские, необходимые для полового размножения. Непериодические колебания численности обусловлены зачастую резким увеличением количества пищи в благоприятный год, нарушением условий обитания, размножением вредителей или хищников.
Поскольку восстановление численности популяции происходит за счет небольшого количества особей, имеющих не весь набор аллелей, новая и исходная популяции будут иметь неодинаковую генетическую структуру. Изменение частоты генов в популяции под действием случайных факторов называется дрейфом генов , или генетико-автоматическими процессами . Он имеет место также при освоении новых территорий, ведь на них попадает чрезвычайно ограниченное число особей данного вида, которые могут дать начало новой популяции. Поэтому здесь особое значение приобретают генотипы данных особей (эффект основателя ). В результате дрейфа генов зачастую выщепляются новые гомозиготные формы (по мутантным аллелям), которые могут оказаться приспособительно ценными, и будут в дальнейшем подхвачены естественным отбором.
Так, у индейского населения американского континента и лапландцев очень высока доля лиц с I (0) группой крови, тогда как III и IV группы крайне редки. Вероятно, в первом случае основателями популяции были лица, не имевшие аллеля I B , либо он был утрачен в процессе отбора.
До определенного момента между соседними популяциями происходит обмен аллелями в результате скрещивания между особями различных популяций - поток генов , который уменьшает расхождение между отдельными популяциями, однако с возникновением изоляции он прекращается. По сути, поток генов представляет собой запаздывающий мутационный процесс.
Изоляция. Любые изменения генетической структуры популяции должны быть закреплены, что и происходит благодаря изоляции - возникновению любых барьеров (географических, экологических, поведенческих, репродуктивных и др.), затрудняющих и делающих невозможным скрещивание особей различных популяций. Хотя сама по себе изоляция не создает новых форм, она, тем не менее, сохраняет генетические различия между популяциями, подвергающихся действию естественного отбора. Различают две формы изоляции: географическую и биологическую.
Географическая изоляция возникает в результате разделения ареала физическими барьерами (водными препятствиями для наземных организмов, участками суши для видов-гидробионтов, чередованием приподнятых участков и равнин); этому оказывает содействие малоподвижный или прикрепленный (у растений) образ жизни. Иногда географическая изоляция может быть вызвана расширением ареала какого-нибудь вида с последующим вымиранием его популяций на промежуточных территориях.
Биологическая изоляция является следствием определенных расхождений организмов в пределах одного вида, которые каким-то образом предотвращают свободное скрещивание. Существует несколько разновидностей биологической изоляции: экологическая, сезонная, этологическая, морфологическая и генетическая. Экологическая изоляция достигается благодаря разделению экологических ниш (например, предпочтение определенных местообитаний или характера пищи, как у клеста-еловика и клеста-сосновика). Сезонная (временная) изоляция наблюдается в случае размножения особей одного вида в разные сроки (разные стада сельди). Этологическая изоляция зависит от особенностей поведения (особенности ритуала ухаживания, окраска, «пение» самок и самцов из разных популяций). При морфологической изоляции препятствием к скрещиванию служат расхождение в структуре органов размножения или даже размерах тела (пекинес и дог). Генетическая изоляция оказывает наибольшее влияние и проявляется в несовместимости половых клеток (гибель зиготы после оплодотворения), стерильности или сниженной жизнеспособности гибридов. Причинами этого являются особенности числа и формы хромосом, вследствие чего становится невозможным полноценное деление клеток (митоз и мейоз).
Нарушая свободное скрещивание между популяциями, изоляция тем самым закрепляет в них те различия, которые возникли на генотипическом уровне вследствие мутаций и колебаний численности. При этом каждая из популяций подвергается действию естественного отбора отдельно от другой, и это в конечном итоге приводит к дивергенции.
Творческая роль естественного отбора в эволюции
Естественный отбор выполняет функцию своеобразного «сита», сортирующего генотипы по степени приспособленности. Однако еще Ч. Дарвин подчеркивал, что отбор не только и не столько направлен на сохранение исключительно наилучших, но на удаление наихудших, то есть он позволяет сохранить многовариантность. Этим функция естественного отбора не ограничивается, поскольку он обеспечивает размножение приспособленных генотипов, и, таким образом, определяет направление эволюции, последовательно слагая случайные и многочисленные уклонения. Естественный отбор не имеет определенной цели: на основе одного и того же материала (наследственной изменчивости) в различных условиях могут быть получены неодинаковые результаты.
В связи с этим рассматриваемый фактор эволюции нельзя сравнивать с работой скульптора, обтесывающего мраморную глыбу, скорее он действует как далекий предок человека, изготавливающий орудие труда из каменного обломка, не представляя себе конечного результата, который зависит не только от природы камня и его формы, но и от силы, направления удара и т. д. Однако в случае неудачи отбор, как и человекообразное существо, отбраковывает «неправильную» форму.
Платой за отбор является возникновение генетического груза , то есть накопление в популяции мутаций, которые со временем могут стать преобладающими вследствие внезапной гибели большей части особей или миграции их небольшого количества.
Под давлением естественного отбора формируется не только разнообразие видов, но и повышается их уровень организации, в том числе происходит их усложнение или специализация. Однако, в отличие от искусственного отбора, осуществляемого человеком только на хозяйственно ценные признаки, зачастую в ущерб приспособительным свойствам, естественный отбор не может способствовать этому, поскольку ни одно приспособление в природе не может компенсировать вред от снижения жизнеспособности популяции.
Исследования С. С. Четверикова
Один из важных шагов к примирению дарвинизма и генетики был сделан московским зоологом С. С. Четвериковым (1880–1959). Основываясь на результатах исследования генетического состава природных популяций плодовой мушки дрозофилы, доказал, что они несут множество рецессивных мутаций в гетерозиготной форме, не нарушающих фенотипического однообразия. Большинство этих мутаций являются неблагоприятными для организма и создают так называемый генетический груз , снижающий приспособленность популяции в целом к среде обитания. Некоторые мутации, не имеющие приспособительного значения в данный момент развития вида, могут приобрести определенную ценность впоследствии, и тем самым являются резервом наследственной изменчивости. Распространение таких мутаций среди особей популяции вследствие последовательных свободных скрещиваний может привести в конечном итоге к их переходу в гомозиготное состояние и проявлению в фенотипе. Если данное состояние признака - фен - является приспособительным, то через несколько поколений оно полностью вытеснит из популяции менее соответствующий данным условиям доминантный фен вместе с его носителями. Таким образом, вследствие подобных эволюционных изменений сохраняется только рецессивная мутантная аллель, а ее доминантная аллель исчезает.
Попробуем доказать это на конкретном примере. При исследовании какой-либо конкретной популяции можно обнаружить, что не только ее фенотипическая, но и генотипическая структуры могут оставаться неизменными в течение длительного времени, что обусловлено свободным скрещиванием, или панмиксией диплоидных организмов.
Данное явление описывается законом Харди – Вайнберга , согласно которому в идеальной популяции неограниченно больших размеров в отсутствие мутаций, миграций, популяционных волн, дрейфа генов, естественного отбора и при условии свободного скрещивания частоты аллелей и генотипов диплоидных организмов не будут изменяться на протяжении ряда поколений.
Например, в популяции некий признак кодируется двумя аллелями одного и того же гена - доминантной (А ) и рецессивной (а ). Частота доминантной аллели обозначается при этом как р , а рецессивной - q . Сумма частот данных аллелей составляет 1: p + q = 1. Следовательно, если нам известна частота доминантной аллели, то можно определить и частоту рецессивной аллели: q = 1 – p . Фактически частоты аллелей равны вероятностям образования соответствующих гамет. Тогда после формирования зигот частоты генотипов уже в первом поколении составят:
(pA + qa ) 2 = p 2 AA + 2pqAa + q 2 aa = 1,
где p 2 AA - частота доминантных гомозигот;
2pqAa - частота гетерозигот;
q 2 aa - частота рецессивных гомозигот.
Несложно рассчитать, что и в последующих поколениях частоты генотипов останутся прежними, поддерживая генетическое разнообразие популяции. Но в природе идеальные популяции отсутствуют, и поэтому в них мутантные аллели не только могут сохраняться, но и распространяться, и даже замещать ранее более распространенные аллели.
С. С. Четвериков четко осознавал, что естественный отбор не просто устраняет отдельные менее удачные признаки, и, соответственно, кодирующие их аллели, но и действует на весь комплекс генов, влияющих на проявление определенного гена в фенотипе, или генотипическую среду . В качестве генотипической среды в настоящее время рассматривается весь генотип как совокупность генов, которая способна усиливать или ослаблять проявление конкретных аллелей.
Не менее важны в развитии эволюционного учения и исследования С. С. Четверикова в области динамики популяций, в частности «волн жизни», или популяционных волн. Еще будучи студентом, в 1905 году он опубликовал статью о возможности вспышек массового размножения насекомых и столь же быстром снижении их численности.
Роль эволюционной теории в формировании современной естественнонаучной картины мира
Значение эволюционной теории в развитии биологии и других естественных наук трудно переоценить, так как она впервые объяснила условия, причины, механизмы и результаты исторического развития жизни на нашей планете, т. е. дала материалистическое объяснение развития органического мира. Кроме того, теория естественного отбора явилась первой подлинно научной теорией биологической эволюции, так как при ее создании Ч. Дарвин опирался не на умозрительные построения, а исходил из собственных наблюдений и опирался на реальные свойства живых организмов. Вместе с тем она обогатила биологический инструментарий историческим методом.
Формулировка эволюционной теории не только вызвала бурную научную дискуссию, но и дала толчок развитию таких наук, как общая биология, генетика, селекция, антропология и целый ряд других. В связи с этим нельзя не согласиться с утверждением, что теория эволюции увенчала собой очередной этап развития биологии и стала отправной точкой для ее прогресса в ХХ веке.
Доказательства эволюции живой природы. Результаты эволюции: приспособленность организмов к среде обитания, многообразие видов
Доказательства эволюции живой природы
В разных областях биологии еще до Ч. Дарвина и после публикации его теории эволюции был получен целый ряд свидетельств, подкрепляющих ее. Эти свидетельства называют доказательствами эволюции . Наиболее часто приводят палеонтологические, биогеографические, сравнительно- эмбриологические, сравнительно-анатомические и сравнительно-биохимические доказательства эволюции, хотя нельзя сбрасывать со счетов и данные систематики, а также селекции растений и животных.
Палеонтологические доказательства основываются на изучении ископаемых остатков организмов. К ним относятся не только хорошо сохранившиеся организмы, вмерзшие в лед или заключенные в янтарь, но и «мумии», обнаруженные в кислых торфяных болотах, а также сохранившиеся в осадочных породах остатки организмов и окаменелости. Наличие в древних породах более простых организмов, чем в позднейших слоях, и то, что виды, встречающиеся на одном уровне, исчезают на другом, считают одним из наиболее значимых доказательств эволюции и объясняют возникновением и вымиранием видов в соответствующие эпохи вследствие изменения условий окружающей среды.
Несмотря на то, что ископаемых остатков обнаружено пока немного и в палеонтологической летописи отсутствуют многие фрагменты вследствие низкой вероятности сохранения органических остатков, все же найдены формы организмов, у которых имеются признаки как эволюционно более древних, так и более молодых групп организмов. Такие формы организмов называют переходными формами . Яркими представителями переходных форм, иллюстрирующими переход от рыб к наземным позвоночным, являются кистеперые рыбы и стегоцефалы, а между рептилиями и птицами определенное место занимает археоптерикс.
Ряды ископаемых форм, последовательно связанных между собой в процессе эволюции не только общими, но и частными чертами строения, называются филогенетическими рядами . Они могут быть представлены ископаемыми остатками с разных континентов, и претендовать на большую или меньшую полноту, однако их изучение невозможно без сравнения с живущими ныне формами, чтобы продемонстрировать поступательность эволюционного процесса. Классическим примером филогенетического ряда является эволюция предков лошади, исследованная основателем эволюционной палеонтологии В. О. Ковалевским.
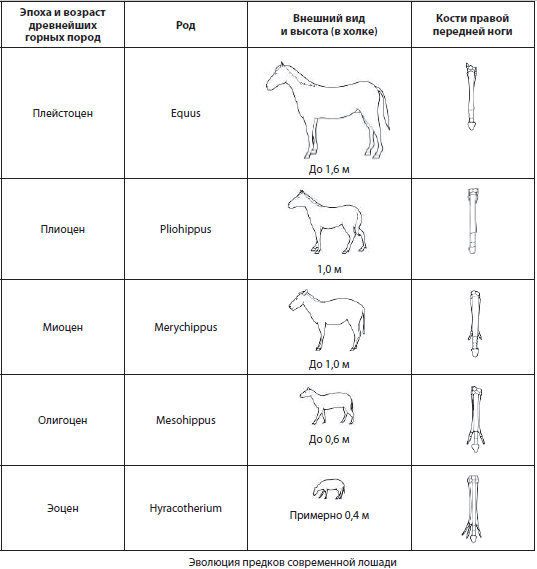
Биогеографические доказательства. Биогеография как наука изучает закономерности распространения и распределения по поверхности нашей планеты видов, родов и других групп живых организмов, а также их сообществ.
Отсутствие в какой-либо части земной поверхности видов организмов, которые приспособлены к такой среде обитания и хорошо приживаются при искусственном завозе, как кролики в Австралии, а также наличие близких форм организмов в отстоящих на значительных расстояниях друг от друга частях суши свидетельствуют, прежде всего, о том, что облик Земли не всегда был таким, и геологические преобразования, в частности, дрейф континентов, образование гор, подъем и опускание уровня Мирового океана влияют на эволюцию организмов. Например, в тропических областях Южной Америки, в Южной Африке и Австралии обитают четыре сходных вида двоякодышащих рыб, ареалы же относящихся к одному отряду верблюдов и лам располагаются в Северной Африке, большей части Азии и в Южной Америке. Палеонтологические исследования показали, что верблюды и ламы происходят от общего предка, обитавшего некогда в Северной Америке, а затем распространившегося в Азию через существовавший ранее перешеек на месте Берингова пролива, а также через Панамский перешеек в Южную Америку. Впоследствии все представители данного семейства в промежуточных областях вымерли, а в краевых в процессе эволюции сформировались новые виды. Более раннее отделение Австралии от остальных массивов суши позволило сформироваться там совершенно особой флоре и фауне, в которой сохранились такие формы млекопитающих, как однопроходные - утконос и ехидна.
С точки зрения биогеографии можно объяснить и разнообразие дарвиновых вьюрков на Галапагосских островах, отстоящих от побережья Южной Америки на 1200 км и имеющих вулканическое происхождение. По-видимому, некогда на них залетели или были занесены представители единственного в Эквадоре вида вьюрков, а затем, по мере размножения, часть особей расселялась по остальным островам. На центральных крупных островах борьба за существование (пищу, места гнездования и т. д.) была наиболее острой, поэтому и сформировались незначительно отличающиеся друг от друга по внешним признакам виды, потребляющие различную пищу (семена, плоды, нектар, насекомых и т. д.).
Влияли на распространение различных групп организмов и изменения климатических условий на Земле, способствовавшие процветанию одних групп и вымиранию других. Отдельные виды или группы организмов, сохранившиеся от широко распространенных ранее флор и фаун, называют реликтами . К ним относятся гинкго, секвойя, тюльпанное дерево, кистеперая рыба латимерия и др. В более широком смысле виды растений и животных, обитающие на ограниченных участках территории или акватории, называются эндемичными , или эндемиками . Например, эндемиками являются все представители аборигенной флоры и фауны Австралии, а во флоре и фауне озера Байкал таковых до 75 %.
Сравнительно-анатомические доказательства. Изучение анатомии родственных групп животных и растений дает убедительные свидетельства сходства строения их органов. Несмотря на то, что экологические факторы, безусловно, накладывают свой отпечаток на строение органов, у покрытосеменных растений при всем их поразительном разнообразии цветки имеют чашелистики, лепестки, тычинки и пестики, а у наземных позвоночных животных конечность построена по плану пятипалой. Органы, имеющие сходное строение, занимающие одно и то же положение в организме и развивающиеся из одних и тех же зачатков у родственных организмов, но выполняющие разные функции, называются гомологичными . Так, слуховые косточки (молоточек, наковальня и стремечко) гомологичны жаберным дугам рыб, ядовитые железы змей - слюнным железам других позвоночных, молочные железы млекопитающих - потовым, ласты тюленей и китообразных - крыльям птиц, конечностям лошадей и кротов.

Нефункционирующие длительное время органы, вероятнее всего, в процессе эволюции превращаются в рудиментарные (рудименты) - недоразвитые по сравнению с предковыми формами структуры, которые потеряли основное значение. К ним относят малую берцовую кость у птиц, глаза у кротов и слепышей, волосяной покров, копчик и червеобразный отросток (аппендикс) у человека и др.
У отдельных особей, тем не менее, могут проявляться признаки, отсутствующие у данного вида, но имевшиеся у отдаленных предков - атавизмы , например, трехпалость у современных лошадей, развитие дополнительных пар молочных желез, хвоста и волосяного покрова на всем теле человека.
Если гомологичные органы являются свидетельством в пользу родства организмов и дивергенции в процессе эволюции, то аналогичные органы - сходные у организмов различных групп структуры, выполняющие одинаковые функции, наоборот, относятся к примерам конвергенции (конвергенцией называется в целом независимое развитие сходных признаков у разных групп организмов, существующих в одинаковых условиях) и подтверждают тот факт, что окружающая среда накладывает значительный отпечаток на организм. Аналогами являются крылья насекомых и птиц, глаза позвоночных и головоногих (кальмаров, осьминогов), членистые конечности членистоногих и наземных позвоночных.

Сравнительно-эмбриологические доказательства. Изучая эмбриональное развитие у представителей разных групп позвоночных, К. Бэр обнаружил их поразительное структурное единство, особенно на ранних стадиях развития (закон зародышевого сходства ). Позднее Э. Геккель сформулировал биогенетический закон , согласно которому онтогенез является кратким повторением филогенеза, т. е. стадии, которые организм проходит в процессе своего индивидуального развития, повторяют историческое развитие той группы, к которой он принадлежит.
Так, зародыш позвоночного на первых стадиях развития приобретает черты строения, характерные для рыб, а затем земноводных и, в конце концов, той группы, к которой он относится. Это превращение объясняется тем, что каждый из вышеперечисленных классов имеет общих предков с современными рептилиями, птицами и млекопитающими.

Однако биогенетический закон обладает целым рядом ограничений, и поэтому русский ученый А. Н. Северцов существенно ограничил область его применения повторением в онтогенезе исключительно особенностей зародышевых стадий развития предковых форм.
Сравнительно-биохимические доказательства. Разработка более точных методов биохимического анализа предоставила ученым-эволюционистам новую группу данных в пользу исторического развития органического мира, поскольку наличие одинаковых веществ у всех организмов указывает на возможную биохимическую гомологию, подобной таковой на уровне органов и тканей. Сравнительно-биохимические исследования первичной структуры таких широко распространенных белков, как цитохром с и гемоглобин, а также нуклеиновых кислот, особенно рРНК, показали, что многие из них имеют практически одинаковое строение и выполняют те же функции у представителей различных видов, при этом, чем ближе родство, тем большее сходство обнаруживается в строении исследуемых веществ.
Таким образом, теория эволюции подтверждается значительным количеством данных из различных источников, что лишний раз свидетельствует о ее достоверности, но она еще будет изменяться и уточняться, поскольку многие аспекты жизни организмов остаются вне поля зрения исследователей.
Результаты эволюции: приспособленность организмов к среде обитания, многообразие видов
Помимо общих признаков, свойственных представителям того или иного царства, виды живых организмов характеризуются поразительным разнообразием особенностей внешнего и внутреннего строения, жизнедеятельности и даже поведения, появившихся и отобранных в процессе эволюции и обеспечивающих приспособление к условиям обитания. Однако не следует считать, что поскольку у птиц и насекомых есть крылья, то это связано с непосредственным действием воздушной среды, ведь и бескрылых насекомых и птиц предостаточно. Вышеупомянутые приспособления были отобраны в процессе естественного отбора из всего спектра имеющихся мутаций.
Эпифитные растения, обитающие не на почве, а на деревьях, приспособились к поглощению атмосферной влаги при помощи корней без корневых волосков, но со специальной гигроскопичной тканью - веламеном . Некоторые бромелии могут впитывать водяные пары во влажной атмосфере тропиков с помощью волосков на листьях.
У насекомоядных растений (росянки, венериной мухоловки), обитающих на почвах, где азот недоступен по тем или иным причинам, выработался механизм привлечения и поглощения мелких животных, чаще всего насекомых, являющихся для них источником искомого элемента.
Для защиты от поедания травоядными животными у многих растений, ведущих прикрепленный способ жизни, сформировались пассивные средства защиты, такие как колючки (боярышник), шипы (роза), жгучие волоски (крапива), накопление кристаллов щавелевокислого кальция (щавель), биологически активных веществ в тканях (кофе, боярышник) и др. У некоторых из них даже семена в незрелых плодах окружены каменистыми клетками, не дающими вредителям добраться до них, и лишь к осени происходит процесс раздревеснения, что позволяет семенам попасть в почву и прорасти (груша).
Среда оказывает формирующее влияние и на животных. Так, многие рыбы и водные млекопитающие имеют обтекаемую форму тела, которая облегчает им передвижение в ее толще. Однако не стоит считать, что вода непосредственно влияет на форму тела, просто в процессе эволюции наиболее приспособленными к ней оказались именно те животные, которые обладали данным признаком.
Тело китов и дельфинов не покрыто при этом волосяным покровом, тогда как у родственной им группы ластоногих имеется в той или иной мере редуцированный шерстный покров, поскольку, в отличие от первых, они часть времени проводят на суше, где без шерсти их кожа тотчас бы обледенела.
Тело большинства рыб покрыто чешуей, которая на нижней стороне более светло окрашена, нежели на верхней, вследствие чего сверху эти животные малозаметны для естественных врагов на фоне дна, а снизу - на фоне неба. Окраска, обеспечивающая незаметность животных для их врагов или жертв, называется покровительственной . Она широко распространена в природе. Ярким примером такой окраски является окраска нижней стороны крыльев бабочки каллимы, которая, сев на веточку и сложив крылья вместе, оказывается похожей на сухой листочек. Другие насекомые, например палочники, маскируются под веточки растений.
Пятнистая или полосатая окраска также имеет приспособительное значение, поскольку на фоне почвы таких птиц, как перепела или гаги, не видно даже на близком расстоянии. Незаметны и пятнистые яйца птиц, гнездящихся на земле.
Окраска животных не всегда настолько же постоянна, как у зебры, например, камбала и хамелеон способны менять ее в зависимости от характера того места, где они находятся. Кукушки же, подкладывая свои яйца в гнезда различных птиц, могут варьировать окраску их скорлупы таким образом, чтобы «хозяева» гнезда не заметили различий между ним и собственными яйцами.
Окраска животных далеко не всегда может делать их незаметными - многие из них просто бросаются в глаза, что должно предупреждать об опасности. Большинство таких насекомых и пресмыкающихся в той или иной степени ядовиты, как, например, божья коровка или оса, поэтому хищник, несколько раз испытав неприятные ощущения после употребления в пищу подобного объекта, избегает его. Тем не менее, предупреждающая окраска не является универсальной, поскольку некоторые птицы приспособились питаться ими (осоед).
Увеличение шансов на выживание у особей с предупреждающей окраской способствовало ее появлению у представителей других видов без должных для того оснований. Это явление носит название мимикрии . Так, неядовитые гусеницы некоторых видов бабочек подражают ядовитым, а божьим коровкам - один из видов тараканов. Однако птицы довольно быстро могут научиться отличать ядовитые организмы от неядовитых и потреблять последних, избегая особей, послуживших образцом для подражания.
В некоторых случаях может наблюдаться и обратное явление - хищные животные подражают по окраске безобидным, что позволяет им приближаться к жертве на близкое расстояние, а затем нападать (саблезубая морская собачка).
Защиту многим видам обеспечивает и приспособительное поведение, которое связано с запасанием пищи на зиму, заботой о потомстве, замирание на месте или наоборот, принятие угрожающей позы. Так, речные бобры заготавливают на зиму несколько кубометров веток, частей стволов и другой растительной пищи, затапливая ее в воде возле «хаток».
Забота о потомстве присуща в основном млекопитающим и птицам, однако и у представителей других классов хордовых она также встречается. Например, известно агрессивное поведение самцов колюшки, отгоняющих всех врагов от гнезда, в котором находится икра. Самцы шпорцевых лягушек наматывают на лапки икру и носят ее до вылупливания из нее головастиков.
Даже некоторые насекомые способны обеспечивать своему потомству более благоприятную среду обитания. Например, пчелы выкармливают своих личинок, а молодые пчелы первое время «работают» только в улье. Муравьи переносят своих куколок вверх и вниз в муравейнике, в зависимости от температуры и влажности, а при угрозе наводнения вообще уносят их с собой. Жукискарабеи заготавливают для своих личинок специальные шарики из отходов жизнедеятельности животных.
Многие насекомые при угрозе нападения застывают на месте и принимают вид сухих палочек, веточек и листочков. А гадюки, наоборот, поднимаются и раздувают свой капюшон, тогда как гремучая змея издает специальный звук погремушкой, расположенной на конце хвоста.
Поведенческие адаптации дополняются и физиологическими, связанными с особенностями среды обитания. Так, человек способен находиться под водой без акваланга всего несколько минут, после чего он может потерять сознание и погибнуть из-за недостатка кислорода, а киты не всплывают на протяжении достаточно длительного времени. Объем легких у них не слишком велик, однако существуют другие физиологические приспособления, например, в мышцах высока концентрация дыхательного пигмента - миоглобина, который как бы запасает кислород и отдает его во время погружения. Кроме того, у китов есть особое образование - «чудесная сеть», которая позволяет использовать кислород даже венозной крови.
Животные жарких мест обитания, например пустыни, постоянно подвергаются риску перегрева и потери избыточного количества влаги. Поэтому лисичка-фенек имеет чрезвычайно большие ушные раковины, позволяющие излучать тепло. Земноводные пустынных регионов во избежание потери влаги через кожу вынуждены перейти к ночному образу жизни, когда влажность повышается и появляется роса.
Птицы, освоившие воздушную среду обитания, помимо анатомо-морфологических приспособлений к полету, имеют и важные физиологические особенности. Например, из-за того, что передвижение в воздухе требует чрезвычайно больших затрат энергии, для этой группы позвоночных характерна высокая интенсивность обмена веществ, а выделяемые продукты метаболизма выводятся тотчас же, что способствует снижению удельной плотности тела.
Приспособления к среде обитания, несмотря на все их совершенство, относительны. Так, некоторые виды молочая вырабатывают ядовитые для большинства животных алкалоиды, однако гусеницы одного из видов бабочек - данаиды - не только питаются тканями молочая, но и накапливают эти алкалоиды, становясь несъедобными для птиц.
Кроме того, адаптации являются целесообразными только в конкретной среде обитания и бесполезны в другой среде. Например, редкий и крупный хищник уссурийский тигр, как и все кошки, имеет мягкие подушечки на лапах и втягивающиеся острые когти, острые зубы, отличное зрение даже в темноте, острый слух и сильные мышцы, что позволяет ему обнаружить жертву, незаметно подкрасться к ней и напасть из засады. Однако его полосатая окраска маскирует его только весной, летом и осенью, тогда как на снегу он становится хорошо заметным и тигр может рассчитывать только на молниеносное нападение.
Соцветия инжира, дающие ценные соплодия, имеют настолько специфическое строение, что опыляются только осами бластофагами, и поэтому, введенные в культуру, они длительное время не плодоносили. Только выведение партенокарпических сортов инжира (образующих плоды без оплодотворения) смогло спасти ситуацию.
Несмотря на то, что описаны примеры видообразования в течение совсем коротких промежутков времени, как в случае с погремком на кавказских лугах, который из-за регулярного скашивания сначала разделился на две популяции - раноцветущие и плодоносящие и поздноцветущие, на самом деле микроэволюция, скорее всего, требует гораздо больших сроков - многих столетий, ведь человечество, разные группы которого тысячелетиями были оторваны друг от друга, тем не менее, так и не разделилось на разные виды. Однако, поскольку эволюция располагает практически неограниченным временем, за сотни миллионов и миллиарды лет на Земле обитало уже несколько миллиардов видов, большая часть которых вымерла, а дошедшие до нас являются качественными этапами этого незатухающего процесса.
Согласно современным данным на Земле насчитывается свыше 2 млн видов живых организмов, большая часть из которых (приблизительно 1,5 млн видов) относится к царству животных, около 400 тыс. - к царству растений, свыше 100 тыс. - к царству грибов, а остальные - к бактериям. Такое поразительное разнообразие является результатом дивергенции (расхождения) видов по различным морфологическим, физиолого-биохимическим, экологическим, генетическим и репродуктивным признакам. Например, один из наибольших родов растений, относящийся к семейству Орхидные, - дендробиум - включает свыше 1 400 видов, а род жуков-калоедов - свыше 1 600 видов.
Классификация организмов является задачей систематики, которая в течение уже 2 тыс. лет пытается построить не просто стройную иерархию, а «естественную» систему, отражающую степень родства организмов. Однако все попытки сделать это пока еще не увенчались успехом, так как в ряде случаев в процессе эволюции наблюдалась не только дивергенция признаков, но и конвергенция (схождение), в результате чего у весьма отдаленных групп органы приобрели черты сходства, как, например, глаз головоногих и глаз млекопитающих.
Макроэволюция. Направления и пути эволюции (А. Н. Северцов, И. И. Шмальгаузен). Биологический прогресс и регресс, ароморфоз, идиоадаптация, дегенерация. Причины биологического прогресса и регресса. Гипотезы возникновения жизни на Земле. Основные ароморфозы в эволюции растений и животных. Усложнение живых организмов в процессе эволюции
Макроэволюция
Образование вида ознаменовывает собой новый виток эволюционного процесса, поскольку особи этого вида, будучи более приспособленными к условиям среды, нежели особи материнского вида, постепенно расселяются на новые территории, и уже в его популяциях играют свою созидательную роль мутагенез, популяционные волны, изоляция и естественный отбор. Со временем эти популяции дают начало новым видам, которые вследствие генетической изоляции имеют гораздо больше признаков сходства между собой, чем с видами того рода, от которого отпочковался видродоначальник, и, таким образом, возникает новый род, затем - новое семейство, отряд (порядок), класс и т. д. Совокупность эволюционных процессов, которые приводят к возникновению надвидовых таксонов (родов, семейств, отрядов, классов и т. п.), называется макроэволюцией. Макроэволюционные процессы как бы обобщают микроэволюционные изменения, происходящие в течение длительного времени, выявляя при этом основные тенденции, направления и закономерности эволюции органического мира, которые не поддаются наблюдению на более низком уровне. До сих пор никаких специфических механизмов макроэволюции не выявлено, поэтому считается, что она осуществляется только посредством микроэволюционных процессов, однако эта позиция постоянно подвергается вполне обоснованной критике.
Возникновение сложной иерархической системы органического мира в значительной степени является результатом неодинаковой скорости эволюционирования различных групп организмов. Так, уже упоминавшийся гинкго двулопастный как бы «законсервировался» на тысячи лет, тогда как достаточно близкие к нему сосны существенно изменились за это время.
Направления и пути эволюции (А. Н. Северцов, И. И. Шмальгаузен). Биологический прогресс и регресс, ароморфоз, идиоадаптация, дегенерация
Анализируя историю органического мира, можно заметить, что в определенные промежутки времени господствовали отдельные группы организмов, которые затем клонились к упадку либо исчезали вовсе. Таким образом, можно различить три магистральных направления эволюции : биологический прогресс, биологический регресс и биологическая стабилизация. Значительный вклад в разработку учения о направлениях и путях эволюции внесли русские эволюционисты А. Н. Северцов и И. И. Шмальгаузен.
Биологический прогресс связан с биологическим процветанием группы в целом и характеризует ее эволюционный успех. Он отражает закономерное развитие живой природы от простого к сложному, от более низкой степени организации к более высокой. По А. Н. Северцову, критериями биологического прогресса являются увеличение численности особей данной группы, расширение ее ареала, а также появление и развитие в ее составе групп низшего ранга (преобразование вида в род, рода в семейство и т. д.). В настоящее время биологический прогресс наблюдается у покрытосеменных растений, насекомых, костистых рыб и млекопитающих.
По А. Н. Северцову, биологический прогресс может достигаться вследствие определенных морфофизиологических преобразований организмов, при этом он выделил три основные пути достижения: арогенез, аллогенез и катагенез.

Арогенез , или морфофизиологический прогресс, связан со значительным расширением ареала данной группы организмов вследствие приобретения больших изменений строения - ароморфозов.
Ароморфозом называют эволюционное преобразование строения и функций организма, которое повышает его уровень организации и открывает новые возможности для приспособления к разнообразным условиям существования.
Примерами ароморфозов являются возникновение эукариотической клетки, многоклеточности, появление сердца у рыб и разделение его полной перегородкой у птиц и млекопитающих, формирование цветка у покрытосеменных и т. п.
Аллогенез , в отличие от арогенеза, не сопровождается расширением ареала, однако внутри старого возникает значительное разнообразие форм, имеющих частные приспособления к среде обитания - идиоадаптации.
Идиоадаптация - это мелкое морфофизиологическое приспособление к специальным условиям среды, полезное в борьбе за существование, но не изменяющее уровня организации. Эти изменения иллюстрируют покровительственная окраска у животных, разнообразие ротовых аппаратов у насекомых, колючки растений и др. Не менее удачным примером являются дарвиновы вьюрки, специализирующиеся на различных видах пищи, у которых преобразования вначале затронули клюв, а затем и другие части тела - оперение, хвост и т. п.
Как это ни парадоксально, но и упрощение организации может вести к биологическому прогрессу. Этот путь называется катагенезом.
Дегенерация - это упрощение организмов в процессе эволюции, которое сопровождается потерей определенных функций или органов.
Фаза биологического прогресса сменяется фазой биологической стабилизации , сущность которой заключается в сохранении признаков данного вида как наиболее благоприятных в данном микроокружении. По И. И. Шмальгаузену, она вовсе «не означает прекращения эволюции, наоборот, означает максимальную согласованность организма с изменениями среды». В фазе биологической стабилизации находятся «живые ископаемые» латимерия, гингко и др.
Антиподом биологического прогресса является биологический регресс - эволюционный упадок данной группы вследствие невозможности приспособиться к изменениям окружающей среды. Он проявляется в снижении численности популяций, сужении ареалов, уменьшении количества групп низшего ранга в составе высшего таксона. Группе организмов, которая пребывает в состоянии биологического регресса, угрожает вымирание. В истории органического мира можно увидеть много примеров такого явления, и в настоящее время регресс характерен для некоторых папоротников, амфибий и рептилий. С появлением человека биологический регресс зачастую обусловлен его хозяйственной деятельностью.
Направления и пути эволюции органического мира не являются взаимоисключающими, то есть появление ароморфоза не означает, что идиоадаптация или дегенерация произойти уже не может. Напротив, согласно разработанному А. Н. Северцовым и И. И. Шмальгаузеном правилу смены фаз , различные направления эволюционного процесса и пути достижения биологического прогресса закономерно сменяют друг друга. В ходе эволюции эти пути сочетаются: достаточно редкие ароморфозы переводят группу организмов на качественно новый уровень организации, а в дальнейшем историческое развитие идет по пути идиоадаптации либо дегенерации, обеспечивающих приспособление к конкретным условиям среды обитания.
Причины биологического прогресса и регресса
В процессе эволюции планку естественного отбора преодолевают и, соответственно, прогрессируют только те группы организмов, у которых наследственная изменчивость создает достаточное количество комбинаций, которые могут обеспечить выживание группы в целом.
Те же группы, у которых по каким-то причинам не имеется такого резерва, в большинстве случаев обречены на вымирание. Зачастую это связано с низким давлением отбора на предыдущих этапах эволюционного процесса, приведшим к узкой специализации группы или даже дегенеративным явлениям. Следствием этого является невозможность приспособиться к новым условиям среды при ее резких изменениях. Ярким примером тому является внезапная гибель динозавров вследствие падения гигантского небесного тела на Землю 65 млн лет назад, которое повлекло за собой землетрясение, поднятие в воздух миллионов тонн пыли, резкое похолодание, гибель большей части растений и растительноядных животных. В то же время предки современных млекопитающих, не имея узких предпочтений источников питания и будучи теплокровными, смогли пережить эти условия и занять на планете господствующее положение.
Гипотезы возникновения жизни на Земле
Из всего спектра гипотез образования Земли наибольшее количество фактов свидетельствует в пользу теории «Большого взрыва». Ввиду того, что данное научное допущение зиждется в основном на теоретических расчетах, подтвердить его экспериментально призван большой адронный коллайдер, сооруженный в Европейском центре ядерных исследований вблизи г. Женева (Швейцария). Согласно теории «Большого взрыва», Земля образовалась свыше 4,5 млрд лет назад вместе с Солнцем и другими планетами Солнечной системы в результате конденсации газопылевого облака. Снижение температуры планеты и миграция химических элементов на ней способствовали ее расслоению на ядро, мантию и кору, а происходившие затем геологические процессы (движение тектонических плит, вулканическая деятельность и т. д.) стали причиной формирования атмосферы и гидросферы.
Жизнь существует на Земле также очень давно, о чем свидетельствуют ископаемые остатки разнообразных организмов в горных породах, однако физические теории не могут дать ответа на вопрос о времени и причинах ее возникновения. Существуют две противоположные точки зрения на возникновение жизни на Земле: теории абиогенеза и биогенеза. Теории абиогенеза утверждают возможность происхождения живого из неживого. К ним относят креационизм, гипотезу самозарождения и теорию биохимической эволюции А. И. Опарина.
Фундаментальным положением креационизма являлось сотворение мира неким сверхъестественным существом (Творцом), что нашло свое отражение в мифах народов мира и религиозных культах, однако возраст планеты и жизни на ней намного превышает указанные в этих источниках сроки, да и несоответствий в них предостаточно.
Основателем теории самозарождения жизни считается древнегреческий ученый Аристотель, который утверждал, что возможно многократное появление новых существ, например, дождевых червей из луж, а червей и мух - из гнилого мяса. Однако эти воззрения были опровергнуты в XVII–XIX веках смелыми опытами Ф. Реди и Л. Пастера.
Итальянский врач Франческо Реди в 1688 году поместил кусочки мяса в горшки и плотно запечатал их, однако никаких червей в них не завелось, тогда как в открытых горшках они появились. Дабы опровергнуть бытовавшее тогда убеждение, что жизненное начало содержится в воздухе, он повторил свои опыты, однако горшки не запечатал, а закрыл несколькими слоями кисеи, и вновь жизнь не появилась. Несмотря на убедительные данные, полученные Ф. Реди, исследования А. ван Левенгука дали новую пищу для дискуссий о «жизненном начале», которые продолжились в течение всего следующего века.
Другой итальянский исследователь - Ладзаро Спалланцани - в 1765 году видоизменил опыты Ф. Реди, прокипятив в течение нескольких часов мясные и овощные отвары и запечатав их. По прошествии нескольких дней он также не обнаружил там никаких признаков жизни и сделал вывод, что живое может возникнуть только от живого.
Последний удар теории спонтанного самозарождения нанес великий французский микробиолог Луи Пастер в 1860 году, поместивший прокипяченный бульон в колбу с S-образным горлышком и не получивший никаких зародышей. Казалось бы, это свидетельствовало в пользу теорий биогенеза, однако оставался открытым вопрос о том, каким же путем возник самый-самый первый организм.
Ответить на него попытался советский биохимик А. И. Опарин, пришедший к выводу о том, что состав атмосферы Земли на первых этапах ее существования был совсем не таким, как в наше время. Скорее всего, она состояла из аммиака, метана, углекислого газа и водяных паров, но не содержала свободного кислорода. Под действием электрических разрядов высокой мощности и при высокой температуре в ней могли синтезироваться простейшие органические соединения, что и было подтверждено экспериментами С. Миллера и Г. Юри в 1953 году, получивших из вышеупомянутых соединений несколько аминокислот, простые углеводы, аденин, мочевину, а также простейшие жирные, муравьиную и уксусную кислоты.
Тем не менее синтез органических веществ еще не означает возникновения жизни, поэтому А. И. Опарин выдвинул гипотезу биохимической эволюции , согласно которой разнообразные органические вещества возникали и объединялись в более крупные молекулы на мелководьях морей и океанов, где условия для химического синтеза и полимеризации являются наиболее благоприятными. Первыми носителями жизни в настоящее время считаются молекулы РНК.
Некоторые из этих веществ постепенно образовывали в воде устойчивые комплексы - коацерваты , или коацерватные капли , напоминающие капли жира в бульоне. В эти коацерваты поступали разнообразные вещества из окружающего раствора, которые подвергались химическим превращениям, происходящим в каплях. Как и органические вещества, коацерваты сами по себе не являлись живыми существами, а были очередной ступенью в их возникновении.
Те из коацерватов, которые имели удачное соотношение веществ в своем составе, в особенности белки и нуклеиновые кислоты, благодаря каталитическим свойствам белков-ферментов со временем приобрели способность воспроизводить себе подобных и осуществлять реакции обмена веществ, при этом структуру белков кодировали нуклеиновые кислоты.
Однако, помимо размножения, для живых систем характерна зависимость от поступления энергии извне. Эта проблема первоначально решалась за счет бескислородного расщепления органических веществ из окружающей среды (кислорода в атмосфере на тот момент не было), т. е.
гетеротрофного питания. Некоторые из поглощаемых органических веществ оказались способными аккумулировать энергию солнечного света, как, например, хлорофилл, что дало возможность ряду организмов перейти к автотрофному питанию. Выделение кислорода в атмосферу в процессе фотосинтеза привело к появлению более эффективного кислородного дыхания, возникновению озонового слоя и, в конечном итоге, выходу организмов на сушу.
Таким образом, результатом химической эволюции явилось появление протобионтов - первичных живых организмов, от которых в результате биологической эволюции произошли все существующие в настоящее время виды.
Теория биохимической эволюции в наше время является наиболее подтвержденной, однако представление о конкретных механизмах возникновения жизни изменились. Например, выяснилось, что образование органических веществ начинается еще в космосе, а органические вещества играют важную роль даже в самом образовании планет, обеспечивая слипание мелких частей. Также формирование органических веществ происходит и в недрах планеты: при одном извержении вулкан выбрасывает до 15 т органики. Существуют и другие гипотезы относительно механизмов концентрирования органических веществ: замораживания раствора, абсорбции (связывания) на поверхности определенных минеральных соединений, действия природных катализаторов и т. п. Возникновение жизни на Земле в настоящее время невозможно, поскольку любые органические вещества, спонтанно образовавшиеся в любой точке планеты, тотчас же были бы окислены свободным кислородом атмосферы или использованы гетротрофными организмами. Это понимал еще в 1871 году Ч. Дарвин.
Теории биогенеза отрицают самопроизвольное зарождение жизни. Основными из них являются гипотеза стационарного состояния и гипотеза панспермии. Первая из них базируется на том, что жизнь существует вечно, тем не менее, на нашей планете есть очень древние породы, в которых следы деятельности органического мира отсутствуют.
Гипотеза панспермии утверждает, что зародыши жизни были занесены на Землю из космоса некими пришельцами либо божественным провидением. В пользу этой гипотезы свидетельствуют два факта: необходимость для всего живого достаточно редкого на планете, но часто встречающегося в метеоритах молибдена, а также находка организмов, похожих на бактерии, на метеоритах с Марса. Однако каким образом жизнь возникла на других планетах, остается невыясненным.
Основные ароморфозы в эволюции растений и животных
Растительные и животные организмы, представляющие различные ветви эволюции органического мира, в процессе исторического развития независимо приобретали определенные черты строения, которые будут охарактеризованы далее.
У растений важнейшими из них являются переход от гаплоидности к диплоидности, независимость от воды в процессе оплодотворения, переход от наружного оплодотворения к внутреннему и возникновение двойного оплодотворения, расчленение тела на органы, развитие проводящей системы, усложнение и усовершенствование тканей, а также специализация опыления с помощью насекомых и распространения семян и плодов.
Переход от гаплоидности к диплоидности сделал растения более устойчивыми к действию факторов окружающей среды вследствие снижения риска проявления рецессивных мутаций. Повидимому, это преобразование коснулось предков сосудистых растений, в число которых не входят моховидные, характеризующиеся преобладанием в жизненном цикле гаметофита.
Главные ароморфозы в эволюции животных связаны с возникновением многоклеточности и все большим расчленением всех систем органов, возникновением прочного скелета, развитием центральной нервной системы, а также общественного поведения в различных группах высокоорганизованных животных, что дало толчок и к прогрессу человека.
Усложнение живых организмов в процессе эволюции
Историю органического мира на Земле изучают по сохранившимся остаткам, отпечаткам и другим следам жизнедеятельности живых организмов. Она является предметом науки палеонтологии . Исходя из того, что остатки разных организмов расположены в различных пластах горных пород, была создана геохронологическая шкала, согласно которой историю Земли разделили на определенные промежутки времени: эоны, эры, периоды и века.
Эоном называют большой промежуток времени в геологической истории, объединяющий несколько эр. В настоящее время выделяют только два эона: криптозой (скрытая жизнь) и фанерозой (явная жизнь). Эра - это промежуток времени в геологической истории, являющийся подразделением эона, объединяющий, в свою очередь, периоды. В криптозое выделяют две эры (архей и протерозой), тогда как в фанерозое - три (палеозой, мезозой и кайнозой).
Важную роль в создании геохронологической шкалы сыграли руководящие ископаемые - остатки организмов, которые были многочисленны в определенные промежутки времени и хорошо сохранились.
Развитие жизни в криптозое. Архей и протерозой составляют большую часть истории жизни (период 4,6 млрд лет - 0,6 млрд лет назад), однако сведений о жизни в тот период недостаточно. Первые остатки органических веществ биогенного происхождения имеют возраст около 3,8 млрд лет, а прокариотические организмы существовали уже 3,5 млрд лет назад. Первые прокариоты входили в состав специфических экосистем - цианобактериальных матов, благодаря деятельности которых образовались специфические осадочные породы строматолиты («каменные ковры»).
Понять жизнь давних прокариотических экосистем помогло открытие их современных аналогов - строматолитов в заливе Шарк-Бей в Австралии и специфических пленок на поверхности почвы в заливе Сиваш в Украине. На поверхности цианобактериальных матов расположены фотосинтезирующие цианобактерии, а под их слоем - чрезвычайно разнообразные бактерии других групп и археи. Минеральные вещества, которые оседают на поверхность мата и образовываются за счет его жизнедеятельности, откладываются пластами (приблизительно 0,3 мм в год). Такие примитивные экосистемы могут существовать лишь в непригодных для жизни других организмов местах, и действительно, оба вышеупомянутые местообитания характеризуются чрезвычайно высокой соленостью.
Многочисленные данные свидетельствуют о том, что поначалу Земля имела атмосферу возобновляемого характера, в состав которой входили: углекислый газ, водяной пар, оксид серы, а также угарный газ, водород, сероводород, аммиак, метан и т. п. Первые организмы Земли были анаэробами, однако благодаря фотосинтезу цианобактерий в среду выделялся свободный кислород, который сначала быстро связывался с восстановителями, находящимися в среде, и лишь после связывания всех восстановителей среда начала приобретать окислительные свойства. О таком переходе свидетельствуют отложение окисленных форм железа - гематита и магнетита.
Около 2 млрд лет назад в результате геофизических процессов практически все несвязанное в осадочных породах железо переместилось к ядру планеты, а кислород начал накапливаться в атмосфере из-за отсутствия этого элемента - произошла «кислородная революция». Она явилась переломным этапом в истории Земли, который повлек за собой не только смену состава атмосферы и образование озонового экрана в атмосфере - главной предпосылки для заселения суши, но и состава пород, формирующихся на поверхности Земли.
В протерозое произошло и другое важное событие - возникновение эукариот. В последние годы удалось собрать убедительные доказательства теории эндосимбиогенетического происхождения эукариотической клетки - путем симбиоза нескольких прокариотических клеток. Вероятно, «главным» предком эукариот стали археи, которые перешли к поглощению пищевых частиц путем фагоцитоза. Наследственный аппарат переместился вглубь клетки, сохранив, тем не менее, связь с мембраной благодаря переходу внешней мембраны возникшей ядерной оболочки в мембраны эндоплазматической сети.
Геохронологическая история Земли Эон Эра Период Начало, млн лет назад Продолжительность, млн лет Развитие жизни Фанерозой Кайнозой Антропоген 1,5 1,5 Четыре ледниковых периода, сменявшихся потопами, привели к формированию холодоустойчивой флоры и фауны (мамонты, овцебыки, северные олени, лемминги). Обмен животными и растениями между материками вследствие возникновения сухопутных мостов. Доминирование плацентарных млекопитающих. Вымирание многих крупных млекопитающих. Становление человека как биологического вида и его расселение. Одомашнивание животных и окультуривание растений. Исчезновение многих видов живых организмов вследствие хозяйственной деятельности человека Неоген 25 23,5 Распространение злаковых. Формирование всех современных отрядов млекопитающих. Возникновение человекообразных обезьян Палеоген 65 40 Доминирование цветковых растений, млекопитающих и птиц. Возникновение копытных, хищных, ластоногих, приматов и др. Мезозой Мел 135 70 Возникновение покрытосеменных растений, млекопитающие и птицы становятся многочисленными Юра 195 60 Эпоха рептилий и головоногих моллюсков. Возникновение сумчатых и плацентарных млекопитающих. Господство голосеменных Триас 225 30 Первые млекопитающие и птицы. Рептилии многочисленны. Распространение травянистых споровых Палеозой Пермь 280 55 Возникновение современных насекомых. Развитие рептилий. Вымирание ряда групп беспозвоночных. Распространение хвойных Карбон 345 65 Первые рептилии. Возникновение крылатых насекомых. Преобладают папоротники и хвощи Девон 395 50 Многочисленны рыбы. Первые амфибии. Возникновение основных групп споровых, первых голосеменных и грибов Силур 430 35 Обильны водоросли. Первые наземные растения и животные (пауки). Обычны челюстноротые рыбы и ракоскорпионы Ордовик 500 70 Обильны кораллы и трилобиты. Расцвет зеленых, бурых и красных водорослей. Возникновение первых хордовых Кембрий 570 70 Многочисленные окаменелости рыб. Обычны морские ежи и трилобиты. Возникновение многоклеточных водорослей Криптозой Протерозой 2600 2000 Возникновение эукариот. Распространены в основном одноклеточные зеленые водоросли. Возникновение многоклеточности. Вспышка разнообразия многоклеточных животных (возникновение всех типов беспозвоночных) Архей 3500 1500 Первые следы жизни на Земле - бактерии и цианобактерии. Возникновение фотосинтеза
Поглощенные клеткой бактерии могли не перевариваться, а оставаться живыми и продолжать свое функционирование. Считают, что митохондрии ведут свое происхождение от пурпурных бактерий, утративших способность к фотосинтезу и перешедших к окислению органических веществ. Симбиоз с другими фотосинтезирующими клетками привел к возникновению пластид у растительных клеток. Вероятно, жгутики эукариотических клеток возникли вследствие симбиоза с бактериями, которые, подобно современным спирохетам, были способны к извивающимся движениям. Поначалу наследственный аппарат эукариотических клеток был устроен приблизительно так же, как у прокариот, и лишь позднее, вследствие необходимости управления большой и сложной клеткой, образовались хромосомы. Геномы внутриклеточных симбионтов (митохондрий, пластид и жгутиков) в целом сохранили прокариотическую организацию, но большая часть их функций перешла к ядерному геному.
Эукариотические клетки возникали неоднократно и независимо друг от друга. Например, красные водоросли возникли в результате симбиогенеза с цианобактериями, а зеленые водоросли - с бактериями-прохлорофитами.
Остальные одномембранные органеллы и ядро эукариотической клетки, согласно эндомембранной теории, возникли из впячиваний мембраны прокариотической клетки.
Точное время появления эукариот неизвестно, поскольку уже в отложениях возрастом около 3 млрд лет присутствуют отпечатки клеток, имеющих похожие размеры. Точно эукариоты зафиксированы в породах возрастом около 1,5–2 млрд лет, но только после кислородной революции (около 1 млрд лет назад) сложились условия, благоприятные для них.
В конце протерозойской эры (не менее 1,5 млрд лет назад) уже существовали и многоклеточные эукариотические организмы. Многоклеточность, как и эукариотическая клетка, неоднократно возникала у разных групп организмов.
Существуют различные взгляды на происхождение многоклеточных животных. По одним данным их родоначальниками были многоядерные, подобные инфузориям, клетки, которые затем распались на отдельные одноядерные клетки.
Другие гипотезы связывают происхождение многоклеточных животных с дифференцировкой клеток колониальных одноклеточных. Расхождения между ними касаются возникновения слоев клеток у первоначального многоклеточного животного. Согласно гипотезы гастреи Э. Геккеля, это происходит путем впячивания одной из стенок однослойного многоклеточного организма, как у кишечнополостных. В противовес ей И. И. Мечников сформулировал гипотезу фагоцителлы, считая предками многоклеточных однослойные шарообразные колонии наподобие вольвокса, которые поглощали пищевые частицы путем фагоцитоза. Клетка, захватившая частицу, теряла жгутик и переходила вглубь организма, где и осуществляла пищеварение, а по окончании процесса возвращалась на поверхность. Со временем произошло разделение клеток на два слоя с определенными функциями - внешний обеспечивал движение, а внутренний - фагоцитоз. Такой организм И. И. Мечников назвал фагоцителлой.
В течение продолжительного времени многоклеточные эукариоты проигрывали в конкурентной борьбе прокариотическим организмам, однако в конце протерозоя (800–600 млн лет тому назад) вследствие резкого изменения условий на Земле - снижения уровня морей, роста концентрации кислорода, уменьшения концентрации карбонатов в морской воде, регулярных циклов похолодания - многоклеточные эукариоты получили преимущества над прокариотами. Если до этого времени встречались только отдельные многоклеточные растения и, возможно, грибы, то с этого момента в истории Земли известны и животные. Из возникших в конце протерозоя фаун лучше других изучены эдиакарская и вендская. Животных вендского периода принято включать в состав особой группы организмов или относить к таким типам, как кишечнополостные, плоские черви, членистоногие и др. Однако ни у одной из этих групп нет скелетов, что может свидетельствовать об отсутствии хищников.
Развитие жизни в палеозойской эре. Палеозойская эра, длившаяся более 300 млн лет, делится на шесть периодов: кембрийский, ордовикский, силурийский, девонский, каменноугольный (карбон) и пермский.
В кембрийском периоде суша состояла из нескольких материков, расположенных преимущественно в Южном полушарии. Самыми многочисленными фотосинтезирующими организмами в этот период были цианобактерии и красные водоросли. В толще воды жили фораминиферы и радиолярии. В кембрии появляется огромное количество скелетных животных организмов, о чем свидетельствуют многочисленные ископаемые остатки. Эти организмы относились примерно к 100 типам многоклеточных животных, как современным (губки, кишечнополостные, черви, членистоногие, моллюски), так и исчезнувшим, например: огромный хищник аномалокарис и колониальные граптолиты, которые плавали в толще воды или были прикреплены ко дну. Суша на протяжении кембрия оставалась почти незаселенной, однако процесс почвообразования уже начали бактерии, грибы и, возможно, лишайники, а в конце периода на сушу вышли малощетинковые черви и многоножки.
В ордовикском периоде уровень вод Мирового океана поднялся, что привело к затоплению материковых низменностей. Основными продуцентами в этот период были зеленые, бурые и красные водоросли. В отличие от кембрия, в котором рифы строили губки, в ордовике их сменяют коралловые полипы. Расцвет переживали брюхоногие и головоногие моллюски, а также трилобиты (ныне вымершие родственники паукообразных). В этом периоде впервые зафиксированы и хордовые, в частности бесчелюстные. В конце ордовика произошло грандиозное вымирание, которое уничтожило около 35 % семейств и более 50 % родов морских животных.
Силурийский период характеризуется усилением горообразования, которое привело к осушению материковых платформ. Ведущую роль в фауне беспозвоночных силура играли головоногие моллюски, иглокожие и гигантские ракоскорпионы, тогда как среди позвоночных сохраняется большое разнообразие бесчелюстных и появляются рыбы. В конце периода на сушу вышли первые сосудистые растения - риниофиты и плауновидные, которые начали колонизацию мелководья и приливно-отливной зоны побережий. На сушу вышли и первые представители класса паукообразных.
В девонском периоде вследствие поднятия суши образовались большие мелководья, которые пересыхали и даже промерзали, поскольку климат становился еще более континентальным, чем в силуре. В морях преобладают кораллы и иглокожие, тогда как головоногие моллюски представлены спирально закрученными аммонитами. Среди позвоночных девона расцвета достигли рыбы, причем на смену панцирным пришли и хрящевые, и костные, а также двоякодышащие и кистеперые. В конце периода появляются первые амфибии, которые сначала жили в воде.
В среднем девоне на суше появились первые леса из папоротников, плаунов и хвощей, которые были заселены червями и многочисленными членистоногими (многоножками, пауками, скорпионами, бескрылыми насекомыми). В конце девона появились первые голосеменные. Освоение суши растениями привело к уменьшению выветривания и усилению почвообразования. Закрепление почв привело к возникновению русел рек.
В каменноугольном периоде суша была представлена двумя материками, разделенными океаном, а климат стал заметно более теплым и влажным. К концу периода произошло небольшое поднятие суши, а климат сменился более континентальным. В морях господствовали фораминиферы, кораллы, иглокожие, хрящевые и костные рыбы, а пресные водоемы населяли двухстворчатые моллюски, ракообразные и разнообразные земноводные. В середине карбона возникли мелкие насекомоядные рептилии, а среди насекомых появились крылатые (тараканы, стрекозы).
Для тропиков были характерны заболоченные леса, в которых доминировали гигантские хвощи, плауны и папоротники, отмершие остатки которых образовали впоследствии залежи каменного угля. В середине периода в умеренной зоне, благодаря их независимости от воды в процессе оплодотворения и наличию семени, началось распространение голосеменных.
Пермский период отличался слиянием всех материков в единый суперконтинент Пангею, отступлением морей и усилением континентальности климата до такой степени, что во внутренних районах Пангеи образовались пустыни. К концу периода на суше почти исчезли древовидные папоротники, хвощи и плауны, а господствующее положение заняли засухоустойчивые голосеменные. Несмотря на то, что крупные амфибии еще продолжали существовать, возникли разные группы рептилий, в том числе крупных растительноядных и хищных. В конце перми произошло самое большое вымирание в истории жизни, так как исчезли многие группы кораллов, трилобиты, большинство головоногих, рыб (в первую очередь хрящевых и кистеперых), а также амфибий. Морская фауна потеряла при этом 40–50 % семейств и около 70 % родов.
Развитие жизни в мезозое. Мезозойская эра продолжалась около 165 млн лет и характеризовалась поднятием суши, интенсивным горообразованием и снижением влажности климата. Она делится на три периода: триасовый, юрский и меловой.
В начале триасового периода климат был засушливым, однако позднее вследствие поднятия уровня морей он стал более влажным. Среди растений преобладали голосеменные, папоротники и хвощи, однако древесные формы споровых практически полностью вымерли. Высокого развития достигли некоторые кораллы, аммониты, новые группы фораминифер, двухстворчатых моллюсков и иглокожих, тогда как разнообразие хрящевых рыб уменьшилось, изменились и группы костных рыб. Господствовавшие на суше рептилии начали осваивать и водную среду, как ихтиозавры и плезиозавры. Из пресмыкающихся триаса до нашего времени дожили крокодилы, гаттерии и черепахи. В конце триаса появились динозавры, млекопитающие и птицы.
В юрском периоде суперконтинент Пангея раскололся на несколько меньших. Большая часть юры была очень влажной, а к его концу климат стал более засушливым. Доминирующей группой растений были голосеменные, из которых от того времени сохранились секвойи. В морях процветали моллюски (аммониты и белемниты, двухстворчатые и брюхоногие), губки, морские ежи, хрящевые и костные рыбы. Крупные амфибии практически полностью вымерли в юрском периоде, однако появились современные группы земноводных (хвостатые и бесхвостые) и чешуйчатых (ящериц и змей), возросло разнообразие млекопитающих. К концу периода возникли и возможные предки первых птиц - археоптериксы. Однако во всех экосистемах доминировали пресмыкающиеся - ихтиозавры и плезиозавры, динозавры и летающие ящеры - птерозавры.
Меловой период получил название в связи с образованием мела в осадочных породах того времени. На всей Земле, кроме приполярных областей, был стойкий теплый и влажный климат. В этом периоде возникли и приобрели широкое распространение покрытосеменные, вытеснявшие голосеменных, что повлекло за собой резкое увеличение разнообразия насекомых. В морях, помимо моллюсков, костистых рыб, плезиозавров, вновь появилось огромное количество фораминифер, раковинки которых и образовали залежи мела, а на суше преобладали динозавры. Лучше приспособленные к воздушной среде птицы начали постепенно вытеснять летающих ящеров.
В конце периода произошло глобальное вымирание, в результате которого исчезли аммониты, белемниты, динозавры, птерозавры и морские ящеры, древние группы птиц, а также некоторые голосеменные. С лица Земли в целом исчезло около 16 % семейств и 50 % родов животных. Кризис в конце мела связывают с падением большого метеорита в Мексиканский залив, однако он, скорее всего, не был единственной причиной глобальных изменений. В ходе последующего похолодания выжили только небольшие рептилии и теплокровные млекопитающие.
Развитие жизни в кайнозое. Кайнозойская эра началась около 66 млн лет назад и продолжается до настоящего времени. Она характеризуется господством насекомых, птиц, млекопитающих и покрытосеменных растений. Кайнозой делят на три периода - палеоген , неоген и антропоген - последний из которых является самым коротким в истории Земли.
В раннем и среднем палеогене климат оставался теплым и влажным, к концу периода стало прохладнее и суше. Доминирующей группой растений стали покрытосеменные, однако, если в начале периода преобладали вечнозеленые леса, то в конце появилось много листопадных, а в засушливых зонах образовались степи.
Среди рыб господствующее положение заняли костистые рыбы, а количество видов хрящевых, несмотря на их заметную роль в соленых водоемах, незначительно. На суше из рептилий сохранились только чешуйчатые, крокодилы и черепахи, тогда как млекопитающие заняли большую часть их экологических ниш. В середине периода появились основные отряды млекопитающих, в том числе насекомоядные, хищные, ластоногие, китообразные, копытные и приматы. Изоляция материков сделала фауну и флору географически более разнообразными: Южная Америка и Австралия стали центрами развития сумчатых, а другие материки - плацентарных млекопитающих.
Неогеновый период. Земная поверхность в неогене приобрела современный вид. Климат стал более прохладным и сухим. В неогене уже сформировались все отряды современных млекопитающих, а в африканских саванах возникло семейство Гоминид и род Человек. К концу периода в приполярных областях континентов распространились хвойные леса, появились тундры, а степи умеренного пояса заняли злаки.
Четвертичный период (антропоген) характеризуется периодическими сменами оледенений и потеплений. Во время оледенений высокие широты покрывались ледниками, резко снижался уровень океана, суживались тропический и субтропический пояса. На близлежащих к ледникам территориях устанавливался холодный и сухой климат, который способствовал формированию холодоустойчивых групп животных - мамонтов, гигантских оленей, пещерных львов и др. Сопутствовавшее процессу оледенения снижение уровня Мирового океана привело к образованию сухопутных мостов между Азией и Северной Америкой, Европой и Британскими островами и т. д. Миграции животных, с одной стороны, привели к взаимообогащению флор и фаун, а с другой, к вытеснению реликтов пришельцами, например, сумчатых и копытных в Южной Америке. Эти процессы, однако, не затронули Австралию, оставшуюся изолированной.
В целом, периодические изменения климата привели к формированию чрезвычайно обильного видового разнообразия, характерного для нынешнего этапа эволюции биосферы, а также оказали влияние на эволюцию человека. На протяжении антропогена несколько видов рода Человек расселились из Африки в Евразию. Около 200 тысяч лет назад в Африке возник вид Человек разумный, который после продолжительного периода существования в Африке около 70 тысяч лет назад вышел в Евразию и около 35–40 тыс. лет назад - в Америку. После периода сосуществования с близкородственными видами он вытеснил их и расселился по всей территории земного шара. Около 10 тыс. лет назад хозяйственная деятельность человека в умеренно теплых областях земного шара начала оказывать влияние как на облик планеты (распашка земель, выжигание лесов, перевыпас пастбищ, опустынивание и т. д.), так и на животный и растительный мир вследствие сокращения ареалов их обитания и истребления, и вступил в действие антропогенный фактор.
Происхождение человека. Человек как вид, его место в системе органического мира. Гипотезы происхождения человека. Движущие силы и этапы эволюции человека. Человеческие расы, их генетическое родство. Биосоциальная природа человека. Социальная и природная среда, адаптации к ней человека
Происхождение человека
Еще 100 лет назад подавляющее большинство людей на планете даже и мысли не допускало о том, что человек может происходить от столь «малопочтенных» животных, как обезьяны. В дискуссии с одним из защитников дарвиновской теории эволюции, профессором Томасом Гексли его ярый противник епископ оксфордский Сэмюэль Уилберфорс, опиравшийся на религиозные догмы, даже обратился к нему с вопросом, считает ли он себя связанным с обезьяньими предками через дедушку или бабушку.
Тем не менее, мысли об эволюционном происхождении высказывали еще античные философы, а великий шведский систематик К. Линней в XVIII веке по совокупности признаков дал человеку видовое название Homo sapiens L. (человек разумный) и отнес его вместе с обезьянами к одному отряду - Приматы. Ж. Б. Ламарк поддержал К. Линнея и считал, что человек даже имел общих предков с современными обезьянами, однако в какой-то момент своей истории он спустился с дерева, что и послужило одной из причин становления человека как вида.
Ч. Дарвин также не обошел вниманием данный вопрос и в 70-е годы XIX века опубликовал труды «Происхождение человека и половой отбор» и «О выражении эмоций у животных и человека », в которых привел не менее убедительные доказательства общности происхождения человека и обезьян, чем немецкий исследователь Э. Геккель («Естественная история творения», 1868; «Антропогенез, или История происхождения человека», 1874), составивший даже родословную царства животных. Однако эти исследования касались лишь биологической стороны становления человека как вида, тогда как социальные аспекты вскрыл классик исторического материализма - немецкий философ Ф. Энгельс.
В настоящее время происхождение и развитие человека как биологического вида, а также разнообразие популяций современного человека и закономерности их взаимодействия изучает наука антропология.
Человек как вид, его место в системе органического мира
Человек разумный (Homo sapiens ) как биологический вид относится к царству животных, подцарству многоклеточных. Наличие в процессе эмбрионального развития хорды, жаберных щелей в глотке, нервной трубки и двусторонней симметрии позволяет отнести его к типу хордовых, тогда как развитие позвоночника, наличие двух пар конечностей и расположение сердца на брюшной стороне тела свидетельствует о его родстве с другими представителями подтипа позвоночных.
Выкармливание детенышей молоком, которое выделяют молочные железы, теплокровность, четырехкамерное сердце, наличие волос на поверхности тела, семи позвонков в шейном отделе позвоночника, преддверия рта, альвеолярных зубов и смена молочных зубов на постоянные являются признаками класса млекопитающих, а внутриутробное развитие зародыша и его связь с организмом матери через плаценту - подкласса плацентарных.
Более частные признаки, такие как хватательные конечности с противопоставленным большим пальцем и ногтями на пальцах, развитие ключиц, направленные вперед глаза, увеличение размеров черепа и головного мозга, а также наличие всех групп зубов (резцов, клыков и коренных) не оставляют сомнения в том, что его место - в отряде приматов.
Значительное развитие головного мозга и мимической мускулатуры, а также особенности строения зубов позволяют отнести человека к подотряду высших приматов, или обезьян.
Отсутствие хвоста, наличие изгибов позвоночника, развитие больших полушарий переднего мозга, покрытых корой с многочисленными бороздами и извилинами, присутствие верхней губы и изреженность волосяного покрова дают основания размещать его среди представителей семейства высших узконосых, или человекообразных обезьян.
Однако даже от наиболее высокоорганизованных обезьян человека отличают резкое увеличение объема головного мозга, прямохождение, широкий таз, выступающий подбородок, членораздельная речь и наличие в кариотипе 46 хромосом и определяют его принадлежность к роду Человек.
Использование верхних конечностей для трудовой деятельности, изготовление орудий труда, абстрактное мышление, коллективная деятельность и развитие на основе больше социальных, чем биологических законов, являются видовыми признаками Человека разумного.
Все современные люди принадлежат к одному виду - Человек разумный (Homo sapiens ), и подвиду H. sapiens sapiens . Этот вид является совокупностью популяций, которые дают при скрещивании плодовитое потомство. Несмотря на достаточно значительное разнообразие морфофизиологических признаков, они не являются свидетельствами более высокой или более низкой степени организации определенных групп людей - все они находятся на одном уровне развития.
В наше время собрано уже достаточное количество научных фактов в интересах становления человека как вида в процессе эволюции - антропогенеза . Конкретный ход антропогенеза еще до конца не выяснен, но благодаря новым палеонтологическим находкам и современным методам исследования можно надеяться, что четкая картина появится уже достаточно скоро.
Гипотезы происхождения человека
Если не принимать во внимание не относящиеся к области биологии гипотезы божественного творения человека и его проникновения с других планет, то все более или менее состоятельные гипотезы происхождения человека возводят его к общим предкам с современными приматами.
Так, гипотеза происхождения человека от древнетропического примата долгопята , или тарзиальная гипотеза , сформулированная английским биологом Ф. Вудом Джонсом в 1929 году, опирается на сходство пропорций тел человека и долгопята, особенности волосяного покрова, укороченность лицевого отдела черепа последнего и т. д. Однако различия в строении и жизнедеятельности этих организмов настолько велики, что она не снискала всеобщего признания.
С человекообразными же обезьянами у человека даже слишком много черт сходства. Так, помимо уже упоминавшихся выше анатомо-морфологических особенностей, следует акцентировать внимание на их постэмбриональном развитии. Например, волосяной покров у маленьких шимпанзе гораздо более редкий, отношение объема головного мозга к объему тела намного больше, а возможности передвижения на задних конечностях несколько шире, чем у взрослых особей. Даже половое созревание у высших приматов наступает намного позже, чем у представителей других отрядов млекопитающих, имеющих сходные размеры тела.
В ходе цитогенетических исследований было выявлено, что одна из хромосом человека образовалась в результате слияния хромосом двух разных пар, присутствующих в кариотипе человекообразных обезьян, и это объясняет различие в числе их хромосом (у человека 2n = 46, а у крупных человекообразных обезьян 2n = 48), а также является еще одним свидетельством родства этих организмов.
Очень велико сходство человека и человекообразных обезьян и по молекулярно-биохимическим данным, поскольку у человека и шимпанзе одинаковы белки групп крови АВ0 и резус, многие ферменты, а аминокислотные последовательности цепей гемоглобина имеют всего 1,6 % различий, тогда как с другими обезьянами это расхождение несколько больше. Да и на генетическом уровне различия в последовательностях нуклеотидов в ДНК между этими двумя организмами составляют менее 1 %. Если учитывать среднюю скорость эволюции таких белков у родственных групп организмов, можно определить, что предки человека отделились от других групп приматов около 6–8 млн лет назад.
Поведение обезьян во многом напоминает поведение человека, так как они живут группами, в которых четко распределены социальные роли. Совместная защита, взаимопомощь и охота - не единственные цели создания группы, поскольку внутри нее обезьяны испытывают привязанность друг к другу, всячески выражают ее, эмоционально реагируют на разнообразные раздражители. Кроме того, в группах происходит обмен опытом между особями.
Таким образом, сходство человека с другими приматами, особенно высшими узконосыми обезьянами, обнаруживается на разных уровнях биологической организации, а отличия человека как вида в значительной степени определяются особенностями этой группы млекопитающих.
К группе гипотез, не подвергающих сомнению происхождение человека от общих предков с современными человекообразными обезьянами, относятся гипотезы полицентризма и моноцентризма.
Исходным положением гипотезы полицентризма является возникновение и параллельная эволюция современного вида человека сразу в нескольких районах земного шара от разных форм древнего или даже древнейшего человека, но это противоречит основным положениям синтетической теории эволюции.
Гипотезы единого происхождения современного человека, напротив, постулируют возникновение человека в одном месте, однако расходятся в том, где это произошло. Так, гипотеза внетропического происхождения человека базируется на том, что только суровые климатические условия высоких широт Евразии могли способствовать «очеловечиванию» обезьян. В ее пользу свидетельствовало открытие на территории Якутии стоянок времен древнейшего палеолита - дирингской культуры, но впоследствии было установлено, что возраст этих находок составляет не 1,8–3,2 млн лет, а 260–370 тыс. лет. Таким образом, эта гипотеза также недостаточно подтверждена.
Наибольшее число доказательств в настоящее время собрано именно в пользу гипотезы африканского происхождения человека , но и она не лишена недостатков, которые призвана учесть комплексная гипотеза широкого моноцентризма , объединяющая аргументы гипотез полицентризма и моноцентризма.
Движущие силы и этапы в эволюции человека
В отличие от других представителей животного мира, человек в процессе своей эволюции подвергался действию не только биологических факторов эволюции, но и социальных, что и способствовало возникновению вида качественно новых существ с биосоциальными свойствами. Социальные факторы обусловили прорыв в принципиально новую адаптивную среду, которая давала огромные преимущества для выживания популяций человека и резко убыстряла темпы его эволюции.
Биологическими факторами эволюции, играющими определенную роль в антропогенезе и по сей день, являются наследственная изменчивость, а также поток генов, поставляющих первичный материал для естественного отбора. Вместе с тем изоляция, популяционные волны и дрейф генов почти полностью утратили свое значение в результате научно-технического прогресса. Это дает основания некоторым ученым считать, что в будущем даже минимальные различия между представителями разных рас исчезнут вследствие их смешения.
Поскольку изменение условий окружающей среды вынудило предков человека спуститься с деревьев на открытое пространство, и перейти к передвижению на двух конечностях, высвобожденные верхние конечности были использованы ими для переноса пищи и детей, а также изготовления и использования орудий труда. Однако изготовить такое орудие можно только при условии наличия четкого представления о конечном результате - образа предмета, поэтому развивалось и абстрактное мышление. Хорошо известно, что сложные движения и процесс мышления являются необходимыми для развития определенных зон коры головного мозга, что и произошло в процессе эволюции. Однако унаследовать такие знания и умения невозможно, их можно лишь передать от одной особи к другой на протяжении жизни последней, следствием чего стало создание особенной формы общения - членораздельной речи.
Таким образом, к социальным факторам эволюции следует отнести трудовую деятельность человека, абстрактное мышление и членораздельную речь. Не следует отбрасывать и проявления альтруизма первобытного человека, который заботился о детях, женщинах и лицах преклонного возраста.
Трудовая деятельность человека не только повлияла на внешний вид его самого, но и позволила вначале частично облегчить условия существования благодаря использованию огня, изготовлению одежды, сооружению жилья, а в дальнейшем и активно изменять их посредством сведения лесов, распахивания земель и т. д. В наше время бесконтрольная хозяйственная деятельность поставила человечество перед угрозой глобальной катастрофы в результате эрозии почв, высыхания пресноводных водоемов, разрушение озонового экрана, что, в свою очередь, может усилить давление биологических факторов эволюции.
Дриопитек , живший около 24 млн лет назад, вероятнее всего, был общим предком человека и человекообразных обезьян. Несмотря на то, что по деревьям он лазил и бегал на всех четырех конечностях, он мог передвигаться и на двух ногах, а в руках переносить пищу. Полное разделение высших обезьян и линии, ведущей к человеку, произошло около 5–8 млн лет назад.

Австралопитек . От дриопитеков, по-видимому, произошел род ардипитек , который сформировался свыше 4 млн лет назад в саваннах Африки в результате похолодания и отступления лесов, что заставило этих обезьян перейти к передвижению на задних конечностях. Это небольшое животное, по всей видимости, дало начало достаточно многочисленному роду австралопитеков («южная обезьяна»).
Австралопитеки появились около 4 млн лет назад и жили в африканских саваннах и сухих лесах, где в полной мере сказались преимущества двуногого перемещения. От австралопитеков пошли две ветви - большие растительноядные с мощными челюстями парантропы и более мелкие и менее специализированные люди . На протяжении определенного времени эти два рода развивались параллельно, что, в частности, проявлялось в увеличении объема мозга, и усложнении используемых орудий. Особенностями нашего рода являются изготовление каменных орудий (парантропы использовали только кость) и относительно крупный мозг.
Первые представители рода Человек появились около 2,4 млн лет назад. Они принадлежали к виду человек умелый (Homo habilis) и были невысокими существами (около 1,5 м) с объемом мозга приблизительно 670 см 3 . Они использовали грубые галечные орудия. По-видимому, у представителей данного вида была хорошо развита мимика и имелась зачаточная речь. Человек умелый сошел с исторической сцены около 1,5 млн лет назад, дав начало следующему виду - человеку выпрямленному.
Человек выпрямленный (H. erectus ) как биологический вид сформировался в Африке около 1,6 млн лет назад и просуществовал на протяжении 1,5 млн лет, быстро расселившись на огромных территориях в Азии и Европе. Представитель этого вида с острова Ява был в свое время описан как питекантроп («обезьяночеловек»), обнаруженный в Китае получил название синантропа , тогда как их европейским «коллегой» является гейдельбергский человек.
Все эти формы также называют архантропами (древнейшими людьми). Человек выпрямленный отличался низким лбом, большими надбровными дугами и скошенным назад подбородком, объем его мозга составлял 900–1200 см 3 . Туловище и конечности человека выпрямленного напоминали таковые у современного человека. Вне всякого сомнения, представители этого рода пользовались огнем и изготавливали обоюдоострые рубила. Как показали недавние находки, этот вид даже освоил мореплавание, ибо его потомки были найдены на отдаленных островах.
Палеоантроп. Около 200 тыс. лет назад от гейдельбергского человека произошел неандертальский человек (H. neandertalensis) , которого относят к палеоантропам (древним людям), жившим в Европе и Западной Азии в пределах 200–28 тыс. лет назад, в том числе в эпохи оледенений. Это были крепкие, физически достаточно сильные и выносливые люди с большим объемом мозга (даже бульшим, чем у современного человека). Они имели членораздельную речь, изготовляли сложные орудия и одежду, хоронили своих умерших, и, возможно, имели даже некоторые зачатки искусства. Неандертальцы не были предками человека разумного, эта группа развивалась параллельно. Их вымирание связано с исчезновением мамонтовой фауны после последнего оледенения, а возможно, является также результатом конкурентного вытеснения со стороны нашего вида.
Наиболее древняя находка представителя человека разумного (Homo sapiens) имеет возраст 195 тысяч лет и происходит из Африки. Скорее всего, предками современного человека являются не неандертальцы, а какая-то форма архантропов, например гейдельбергский человек.
Неоантроп. Около 60 тыс. лет назад в результате неизвестных событий наш вид едва не вымер, поэтому все следующие люди являются потомками небольшой группы, которая насчитывала всего несколько десятков особей. Переборов этот кризис, наш вид начал расселяться по Африке и Евразии. От других видов он отличается более стройным телосложением, более высокой скоростью размножения, агрессивностью и, конечно, самым сложным и самым гибким поведением. Людей современного типа, населявших Европу 40 тыс. лет назад, называют кроманьонцами и относят к неоантропам (современным людям). Они биологически не отличались от современных людей: рост 170–180 см, объем мозга около 1600 см 3 . У кроманьонцев появились искусство и религия, они одомашнили многие виды диких животных и окультурили многие виды растений. От кроманьон цев произошли современные люди.
Человеческие расы, их генетическое родство
При расселении человечества по планете между разными группами людей возникали определенные расхождения, касающиеся цвета кожи, черт лица, характера волос, а также частот встречаемости некоторых биохимических особенностей. Совокупность таких наследственных признаков характеризует группу особей одного вида, различия между которыми являются менее существенными, чем подвиды - расу.
Изучение и классификация рас усложнены отсутствием четких границ между ними. Все современное человечество принадлежит к одному виду, внутри которого выделяют три большие расы: австрало-негроидную (черную), европеоидную (белую) и монголоидную (желтую). Каждая из них делится на малые расы. Различия между расами сводятся к особенностям цвета кожи, волос, формы носа, губ и т. д.
Австрало-негроидная , или экваториальная раса характеризуется темным цветом кожи, волнистыми или курчавыми волосами, широким и мало выступающим носом, поперечным расположением ноздрей, толстыми губами и рядом черепных признаков. Европеоидная , или евразийская раса характеризуется светлой или смуглой кожей, прямыми или волнистыми мягкими волосами, хорошим развитием волосяного покрова на лице мужчин (борода и усы), узким выступающим носом, тонкими губами и рядом черепных признаков. Монголоидная (азиатско-американская) раса характеризуется смуглой или светлой кожей, часто жесткими волосами, средней шириной носа и губ, уплощенностью лица, сильным выступанием скул, сравнительно крупными размерами лица, заметным развитием «третьего века».
Эти три расы различаются и расселением. До эпохи европейской колонизации австрало-негроидная раса была распространена в Старом Свете к югу от тропика Рака; европеоидная раса - в Европе, Северной Африке, Передней Азии и Северной Индии; монголоидная раса - в Юго-Восточной, Северной, Центральной и Восточной Азии, Индонезии, Северной и Южной Америке.
Тем не менее, различия между расами касаются лишь второстепенных признаков, имеющих приспособительное значение. Так, кожа негроидов получает ожог при десятикратно большей дозе ультрафиолетового облучения, чем кожа европеоидов, однако европеоиды меньше страдают от рахита в высоких широтах, где может ощущаться нехватка необходимого для образования витамина D ультрафиолета.
Ранее некоторые люди стремились доказать совершенство одной из рас, чтобы получить моральный перевес над другими. В настоящее время понятно, что расовые особенности отражают лишь разные исторические пути групп людей, но никак не связаны с преимуществом или биологической отсталостью той или другой группы. Человеческие расы менее четко очерчены, чем подвиды и расы других животных, и никак не могут быть сравнены, например, с породами домашних животных (которые являются результатом целеустремленного отбора). Как показывают медико-биологические исследования, последствия межрасового брака зависят от индивидуальных особенностей мужчины и женщины, а не от их расовой принадлежности. Поэтому любые запрещения на межрасовые браки или определенные суеверия являются ненаучными и антигуманными.
Более конкретными, чем расы, группами людей являются народности - исторически сформированные языковые, территориальные, экономические и культурные общности людей. Население определенной страны образует ее народ. При взаимодействии многих народностей в составе какого-либо народа может возникнуть нация. Сейчас на Земле не существует «чистых» рас, а каждый достаточно большой народ представлен людьми, которые принадлежат к разным расам.
Биосоциальная природа человека
Несомненно, человек как биологический вид должен испытывать давление факторов эволюции, таких как мутагенез, популяционные волны и изоляция. Однако по мере становления человеческого общества одни из них ослабевают, а другие, наоборот, усиливаются, поскольку на планете, захваченной процессами глобализации, почти не осталось изолированных популяций человека, в которых осуществляются близкородственные скрещивания, а численность самих популяций не подвержена резким колебаниям. Соответственно, и движущий фактор эволюции - естественный отбор - благодаря успехам медицины не играет уже той роли в популяциях человека, какая свойственна ему в популяциях других организмов.
К сожалению, ослабление давления отбора ведет к возрастанию частоты наследственных заболеваний в популяциях. Например, в индустриально развитых странах до 5 % населения страдают цветовой слепотой (дальтонизмом), тогда как в менее развитых государствах этот показатель составляет до 2 %. Негативные последствия данного явления могут быть преодолены благодаря профилактическим мероприятиям и прогрессу в таких областях науки, как генная терапия.
Однако это не означает, что эволюция человека завершилась, поскольку естественный отбор продолжает действовать, устраняя, например, гаметы и особей с неблагоприятными комбинациями генов еще на проэмбриональном и эмбриональном периодах онтогенеза, а также по устойчивости к возбудителям различных заболеваний. Кроме того, материал для естественного отбора поставляет не только мутационный процесс, но и накопление знаний, способность к обучению, восприятие культуры и других признаков, которые могут передаваться от человека к человеку. В отличие от генетической информации, накопленный в процессе индивидуального развития опыт передается как от родителей потомкам, так и в обратном направлении. А конкуренция возникает уже между сообществами, различающимися в культурном отношении. Эта форма эволюции, свойственная исключительно человеку, получила название культурной , или социальной эволюции.
Тем не менее, культурная эволюция не исключает биологической, поскольку она стала возможной только вследствие формирования человеческого мозга, а сама биология человека в настоящее время определяется культурной эволюцией, так как в отсутствие общества и разнообразия движений в мозге не формируются определенные зоны.
Таким образом, человек имеет биосоциальную природу, которая накладывает отпечаток на проявление биологических, в том числе генетических закономерностей, которым подчиняется его индивидуальное и эволюционное развитие.
Социальная и природная среда, адаптации к ней человека
Под социальной средой понимают, прежде всего, окружающие человека общественные материальные и духовные условия его существования и деятельности. Помимо экономической системы, общественных отношений, общественного сознания и культуры, в нее включают и непосредственное окружение человека - семью, рабочий и учащийся коллективы, а также другие группы. Среда, с одной стороны, оказывает решающее воздействие на формирование и развитие личности, а с другой, и сама изменяется под влиянием человека, что влечет за собой новые изменения людей и т. д.
Приспособление индивидов или их групп к социальной среде для реализации собственных потребностей, интересов, жизненных целей и включает в себя адаптацию к условиям и характеру учебы, труда, межличностным отношениям, экологической и культурной среде, условиям проведения досуга и быта, а также их активное изменение для удовлетворения своих потребностей. Большую роль в этом играет и изменение самого себя, своих мотивов, ценностей, потребностей, поведения и т. д.
Информационные нагрузки и эмоциональные переживания в современном обществе зачастую являются основной причиной стресса, преодолеть который возможно с помощью четкой самоорганизации, физической подготовки и аутотренинга. В некоторых, особо тяжелых случаях, требуется обращение к врачу-психотерапевту. Попытка найти забвение этих проблем в переедании, курении, употреблении алкоголя и других вредных привычках не приводит к желаемому результату, а лишь усугубляет состояние организма.
Не меньшее влияние оказывает на человека и окружающая природная среда, несмотря на то, что человек уже около 10 тыс. лет пытается сформировать для себя комфортную искусственную среду. Так, подъем на значительную высоту из-за снижения концентрации кислорода в воздухе приводит к возрастанию количества эритроцитов в крови, усилению дыхания и сердцебиения, а длительное нахождение на открытом солнце способствует усилению пигментации кожи - загару. Однако перечисленные изменения укладываются в норму реакции и не наследуются. Тем не менее, у народов, длительное время проживающих в подобных условиях, могут иметься некоторые приспособления. Так, у северных народов носовые пазухи имеют гораздо больший объем для согревания воздуха, а размер выступающих частей тела уменьшается для снижения потери тепла. Африканцы отличаются более темным цветом кожи и курчавыми волосами, поскольку пигмент меланин защищает органы тела от проникновения губительных ультрафиолетовых лучей, а шапка волос обладает термоизоляционными свойствами. Светлые глаза европейцев являются приспособлением к более острому восприятию зрительной информации в сумерках и в тумане, а монголоидный разрез глаз - это результат естественного отбора на действие ветров и пыльных бурь.
Для этих изменений требуются века и тысячелетия, однако и жизнь в цивилизованном обществе влечет за собой некоторые изменения. Так, уменьшение физических нагрузок приводит к облегчению скелета и снижению его прочности, уменьшению мышечной массы. Низкая подвижность, избыток высококалорийной еды, стрессы влекут за собой рост числа людей с избыточным весом, а полноценное белковое питание и продолжение светового дня с помощью искусственного освещения способствуют акселерации - ускорению роста и полового созревания, увеличению размеров тела.
Сади Карно, сын военного министра при Наполеоне Бонапарте и дядя будущего президента Французской республики, анализируя последствия завоевательных войн Франции начала XIX века, пришел к выводу, что одной из причин поражения Франции явилось ее отставание в использовании энергии.
В последние двести лет люди потратили много сил, чтобы разобраться в этом понятии. Сегодня стало ясно, что благосостояние людей связано с потреблением энергии. Например, объем валового национального продукта страны почти пропорционален объему потребляемой энергии.
Изучением способов превращения энергии в физических телах занимается термодинамика. Она отвечает на вопросы: как получить тепло, как сделать процесс получения тепла управляемым, как его сохранить и как лучше его использовать. Чтобы ответить на них, ученые ввели определения и сформулировали основные законы, или начала, термодинамики.
Энергия (от греческого energeia - действие, деятельность) определяется как общая количественная мера различных форм движения материи.
В природе любым физическим процессам соответствует тот или иной вид энергии: механической, химической, тепловой, гравитационной, электромагнитной, ядерной и так далее. Другими словами, она выражает способность тела или системы совершать работу.
Фундаментальные исследования в области теории информации привели к понятию информационной энергии , или энергии информационного воздействия , как к количественной мере изменения количества информации.
Первое начало термодинамики кратко формулируется так: “Энергия сохраняется”. Она не возникает из ничего и не исчезает бесследно; она может только переходить из одной формы в другую. Этим основополагающим открытием 50-х годов прошлого столетия мы обязаны лорду Кельвину (Уильямсу Томсону) и Рудольфу Клаузису. Кельвин, как религиозный человек, считал, что Творец в момент создания мира наделил его запасом энергии, и что этот божественный дар будет существовать вечно.
Второе начало термодинамики устанавливает наличие в природе фундаментальной асимметрии, то есть однонаправленности всех происходящих в ней самопроизвольных процессов рассеивания энергии: горячие тела с течением времени охлаждаются, однако холодные сами по себе не становятся горячими, вращающийся волчок, в конце концов, останавливается, однако покоящийся волчок самопроизвольно не начнет вращаться.
То есть распределение имеющейся энергии изменяется необратимым образом. Это свойство материи отражается принципом энтропии . Если есть термин энтропия , характеризующий степень беспорядка в природе, значит должен существовать термин, определяющий степень порядка. Энтальпия и является функцией, оценивающей степень упорядоченности системы (локальной неоднородности).
Естественно протекающие в природе процессы сопровождаются увеличением энтропии (беспорядка).
Удивительным образом эти представления отражаются в модели Вселенной. Она расширяется, при этом происходит рассеяние энергии в пространстве. Непрерывное и глобальное естественное разрушение является основным свойством окружающего нас материального мира. В этом контексте то, что стоит в сознании человека за образом Мефистофеля, играет важную роль в природе, обеспечивая процессыдиссипации (от латинского dissipatio - рассеяние). Вместе с тем из-за локальных неоднородностей распределения вещества в природе происходит его накопление в локальных зонах Вселенной. Так образуются галактики, созвездия, солнечные системы, планеты.
Если мы не поддерживаем сознательно в доме порядок, то независимо от нашего желания в нем будет увеличиваться беспорядок. И наоборот, целенаправленная созидательная деятельность сопровождается уменьшением энтропии (беспорядка) в некоторой локальной зоне пространства. Правда, это возможно только за счет создания еще большего беспорядка вне этой зоны: так рождаются свалки и другие отходы - побочные продукты созидательной деятельности.
Из практики мы знаем, что созидательная деятельность невозможна без использования разума или некоего рожденного природой алгоритма. В этом случае рождение кристалла в аморфной породе является примером целенаправленной созидательной деятельности в природе на основе рожденного ею же самой алгоритма.
Любой живой организм также является примером ограниченного пространства, в котором в течение жизни энтропия не возрастает. В отличие от кристалла организм обладает свойством саморегулирования, то есть способностью поддерживать постоянство внутренней среды при изменении внешних и внутренних возмущений. Это свойство не увеличивать энтропию в течение жизни является одним из основных, отличающих элемент живой природы от неживой.
Таким образом, поддержание порядка связано с алгоритмом саморегулирования, сознанием. Рассуждая понятиями гетевских аллегорий, можно сказать, что Творец отвечает за процессы, связанные с жизнью, не увеличивающие энтропию. Он как бы использует материю как экспериментальный материал для создания и отработки алгоритмов, поддерживающих различные жизни в поисках их совершенства.
Поскольку на практике часто трудно непосредственно измерить величину энергии тела, люди научились оценивать ее через количество работы, которую можно с ее помощью выполнить. Из школьного курса физики мы знаем, что работа оценивается как произведение силы , приложенной к телу, умноженной на расстояние, на которое тело переместилось. Так или иначе, понятие силы в природе связано с видом взаимодействия.
Как мы уже отмечали, в основе всех известных сил лежат четыре основных типа взаимодействия физических тел (таблица 1.1.1). При рассмотрении динамических процессов важно помнить, что любое движение материального тела может быть разложено на три основных: поступательное, колебательное и вращательное.
Движение и сила имеют направления, которые не всегда совпадают. Это несовпадение учитывается коэффициентом, пропорциональным величине угла между направлением силы и направлением движения. Теперь мы можем объяснить, что показывающие время часы, неподвижно висящие на стене, потребляют энергию за счет колебания маятника и других частей. Ось юлы, практически неподвижно стоит относительно пола. Однако юла имеет энергию вращения и тратит ее на преодоление сопротивления воздуха и трения оси о пол.
Для оценки скорости использования энергии или ее приобретения применяется понятие мощности, как скорости изменения общей энергии некоторой системы.
Таким образом, произведение мощности на время дает энергию. Мощность измеряется в ваттах. За одну секунду накопитель энергии с мощностью один ватт способен дать системе один джоуль (Дж) энергии. Одна лошадиная сила определяется как ежесекундный расход энергии, равной 746 Дж.
Итак, перечислим необходимые для понимания материала книги основные законы, которые лежат в основе процессов, происходящих во Вселенной (макромире):
1. Закон иерархического подобия.
2. Закон ритма.
3. Закон сохранения энергии.
4. Закон энтропии.
Законы экологии — общие закономерности и принципы взаимодействия человеческого общества с природной средой.
Значение этих законов состоит в регламентации характера и направленности человеческой деятельности в пределах экосистем различного уровня. Среди законов экологии, сформулированных разными авторами, наибольшую известность получили четыре закона-афоризма американского ученого-эколога Барри Коммонера (1974):
- «все связано со всем» (закон всеобщей связи вещей и явлений в природе);
- «все должно куда-то деваться» (закон сохранения массы вещества);
- «ничто не дается даром» (о цене развития);
- «природа знает лучше» (о главном критерии эволюционного отбора).
Из закона всеобщей связи вещей и явлений в природе («все связано со всем») вытекает несколько следствий:
- закон больших чисел - совокупное действие большого числа случайных факторов приводит к результату, почти не зависящему от случая, т.е. имеющему системный характер. Так, мириады бактерий в почве, воде, телах живых организмов создают особую, относительно стабильную микробиологическую среду, необходимую для нормального существования всего живого. Или другой пример: случайное поведение большого числа молекул в некотором объеме газа обусловливает вполне определенные значения температуры и давления;
- принцип Ле Шателье (Брауна) - при внешнем воздействии, выводящем систему из состояния устойчивого равновесия, это равновесие смещается в направлении, при котором эффект внешнего воздействия уменьшается. На биологическом уровне он реализуется в виде способности экосистем к саморегуляции;
- закон оптимальности — любая система функционирует с наибольшей эффективностью в некоторых характерных для нее пространственно-временных пределах;
- любые системные изменения в природе оказывают прямое или опосредованное воздействие на человека — от состояния индивидуума до сложных общественных отношений.
Из закона сохранения массы вещества («все должно куда-то деваться») вытекают по меньшей мере два постулата, имеющих практическое значение:
Барри Коммонер писан «...глобальная экосистема представляет собой единое целое, в рамках которого ничего не может быть выиграно или потеряно и которое не может являться объектом всеобщего улучшения; все, что было извлечено из нее человеческим трудом, должно быть возмещено. Платежа по этому векселю нельзя избежать; он может быть только отсрочен. Нынешний кризис окружающей среды говорит о том, что отсрочка очень затянулась».
Принцип «природа знает лучше» определяет прежде всего то, что может и что не должно иметь места в биосфере. Все в природе — от простых молекул до человека — прошло жесточайший конкурс на право существования. В настоящее время планету населяет лишь 1/1000 испытанных эволюцией видов растений и животных. Главный критерий этого эволюционного отбора — вписанность в глобальный биотический круговорот , заполненность всех экологических ниш. У любого вещества, выработанного организмами, должен существовать разлагающий его фермент, и все продукты распада должны вновь вовлекаться в круговорот. С каждым биологическим видом, который нарушал этот закон, эволюция рано или поздно расставалась. Человеческая индустриальная цивилизация грубо нарушает замкнутость биотического круговорота в глобальном масштабе, что не может остаться безнаказанным. В этой критической ситуации должен быть найден компромисс, что под силу только человеку, обладающему разумом и стремлением к этому.
Помимо формулировок Барри Коммонера современные экологи вывели еще один закон экологии - «на всех не хватит» (закон ограниченности ресурсов). Очевидно, что масса питательных веществ для всех форм жизни на Земле конечна и ограничена. Ее не хватает на всех появляющихся в биосфере представителей органического мира, поэтому значительное увеличение численности и массы каких-либо организмов в глобальном масштабе может происходить только за счет уменьшения численности и массы других. На противоречие между скоростью размножения и ограниченностью ресурсов питания применительно к народонаселению планеты впервые обратил внимание английский экономист Т.Р. Мальтус (1798), который именно этим пытался обосновать неизбежность социальной конкуренции. В свою очередь, Ч. Дарвин заимствовал у Мальтуса понятие «борьба за существование» для объяснения механизма естественного отбора в живой природе.
Закон ограниченности ресурсов — источник всех форм конкуренции, соперничества и антагонизма в природе и, к сожалению, в обществе. И сколько бы ни считали классовую борьбу, расизм, межнациональные конфликты чисто социальными явлениями — все они своими корнями уходят во внутривидовую конкуренцию, принимающую иногда гораздо более жестокие формы, чем у животных.
Существенное различие в том, что в природе в результате конкурентной борьбы выживают лучшие, а в человеческом обществе — это отнюдь не так.
Обобщенную классификацию экологических законов представил известный советский ученый Н.Ф. Реймерс. Им даны следующие формулировки:
- закон социально-экологического равновесия (необходимости сохранения равновесия между давлением на среду и восстановлением этой среды, как природным, так и искусственным);
- принцип культурного управления развитием (наложение ограничений на экстенсивное развитие, учет экологических ограничений);
- правило социально-экологического замещения (необходимость выявления путей замещения человеческих потребностей);
- закон социально-экологической необратимости (невозможность поворота эволюционного движения вспять, от сложных форм к более простым);
- закон ноосферы Вернадского (неизбежность трансформации биосферы под влиянием мысли и человеческого труда в ноосферу — геосферу, в которой разум становится доминирующим в развитии системы «человек-природа»).
Соблюдение этих законов возможно при условии осознания человечеством своей роли в механизме поддержания стабильности биосферы. Известно, что в процессе эволюции сохраняются только те виды, которые способны обеспечивать устойчивость жизни и окружающей среды. Только человек, используя силу своего разума, может направить дальнейшее развитие биосферы по пути сохранения дикой природы, сохранения цивилизации и человечества, создания более справедливой социальной системы, перехода от философии войны к философии мира и партнерства, любви и уважения к будущим поколениям. Все это составляющие нового биосферного мировоззрения, которое должно стать общечеловеческим.
Законы и принципы экологии
Закон минимума
В 1840 г. Ю. Либих установил, что урожай часто ограничивается не теми питательными веществами, которые требуются в больших количествах, а теми, которых нужно немного, но которых мало и в почве. Сформулированный им закон гласил: «Веществом, находящимся в минимуме, управляется урожай, определяется величина и устойчивость последнего во времени». Впоследствии к питательным веществам добавили ряд других факторов, например температуру. Действие данного закона ограничивают два принципа. Первый закон Либиха строго действует только в условиях стационарного состояния. Более точная формулировка: «при стационарном состоянии лимитирующим будет то вещество, доступные количества которого наиболее близки к необходимому минимуму». Второй принцип касается взаимодействия факторов. Высокая концентрация или доступность некоторого вещества может изменять потребление минимального питательного вещества. Следующий закон сформулирован в самой экологии и обобщает закон минимума.
Закон толерантности
Этот закон формулируется следующим образом: отсутствие или невозможность развития экосистемы определяется не только недостатком, но и избытком любого из факторов (тепло, свет, вода). Следовательно, организмы характеризуются как экологическим минимумом, так и максимумом. Слишком много хорошего тоже плохо. Диапазон между двумя величинами составляет пределы толерантности, в которых организм нормально реагирует на влияние среды. Закон толерантности предложил В. Шелфорд в 1913 г. Можно сформулировать ряд дополняющих его предложений.
- Организмы могут иметь широкий диапазон толерантности в отношении одного фактора и узкий в отношении другого.
- Организмы с широким диапазоном толерантности ко всем факторам обычно наиболее широко распространены.
- Если условия по одному экологическому фактору не оптимальны для вида, то может сузиться диапазон толерантности к другим экологическим факторам.
- В природе организмы очень часто оказываются в условиях, не соответствующих оптимальному значению того или иного фактора, определенному в лаборатории.
- Период размножения обычно является критическим; в этот период многие факторы среды часто оказываются лимитирующими.
Живые организмы изменяют условия среды, чтобы ослабить лимитирующее влияние физических факторов. Виды с широким географическим распространением образуют адаптированные к местным условиям популяции, которые называются экотипами. Их оптимумы и пределы толерантности соответствуют местным условиям.
Обобщающая концепция лимитирующих факторов
Наиболее важными факторами на суше являются свет, температура и вода (осадки), а в море — свет, температура и соленость. Эти физические условия существования могут быть лимитирующими и влияющими благоприятно. Все факторы среды зависят друг от друга и действуют согласованно. Из других лимитирующих факторов можно отметить атмосферные газы (углекислый газ, кислород) и биогенные соли. Формулируя «закон минимума», Либих и имел в виду лимитирующее воздействие жизненно важных химических элементов, присутствующих в среде в небольших и непостоянных количествах. Они называются микроэлементами и к ним относятся железо, медь, цинк, бор, кремний, молибден, хлор, ванадий, кобальт, йод, натрий. Многие микроэлементы подобно витаминам действуют как катализаторы. Фосфор, калий, кальций, сера, магний, требующиеся организмам в больших количествах, называются макроэлементами. Важным лимитирующим фактором в современных условиях является загрязнение природной среды. Главный лимитирующий фактор, по Ю. Одуму, - размеры и качество «ойкоса », или нашей «природной обители», а не просто число калорий, которые можно выжать из земли. Ландшафт не только склад запасов, но и дом, в котором мы живем. «Следует стремиться к тому, чтобы сохранить, по меньшей мере, треть всей суши в качестве охраняемого открытого пространства. Это означает, что треть всей нашей среды обитания должны составлять национальные или местные парки, заповедники, зеленые зоны, участки дикой природы и т.п.». Территория, необходимая одному человеку, по разным оценкам колеблется от 1 до 5 га. Вторая из этих цифр превосходит площадь, которая приходится ныне на одного жителя Земли.
Плотность населения приближается к одному человеку на 2 га суши. Пригодны же для сельского хозяйства только 24% суши. Хотя с площади всего лишь 0,12 га можно получить достаточно калорий, чтобы поддержать существование одного человека, для полноценного питания с большим количеством мяса, фруктов и зелени необходимо около 0,6 га на человека. Кроме того, требуется еще около 0,4 га для производства разного рода волокна (бумаги, древесины, хлопка) и еще 0,2 га для дорог, аэропортов, зданий и т.п. Отсюда концепция «золотого миллиарда», в соответствии с которой оптимальным количеством населения является 1 млрд человек, и стало быть, уже сейчас около 5 млрд «лишних людей». Человек впервые за свою историю столкнулся с предельными, а не локальными ограничениями. Преодоление лимитирующих факторов требует огромных затрат вещества и энергии. Для удвоения урожая необходимо десятикратное увеличение количества удобрений, ядохимикатов и мощности (животных или машин). К лимитирующим факторам относится и численность популяции.
Закон конкурентного исключения
Данный закон формулируется следующим образом: два вида, занимающие одну экологическую нишу, не могут сосуществовать в одном месте неограниченно долго.
То, какой вид побеждает, зависит от внешних условий. В сходных условиях победить может каждый. Важным для победы обстоятельством является скорость роста популяции. Неспособность вида к биотической конкуренции ведет к его оттеснению и необходимости приспособления к более трудным условиям и факторам.
Закон конкурентного исключения может работать и в человеческом обществе. Особенность его действия в настоящее время заключается в том, что цивилизации не могут разойтись. Им некуда уйти со своей территории, потому что в биосфере нет свободного места для расселения и нет избытка ресурсов, что приводит к обострению борьбы со всеми вытекающими отсюда последствиями. Можно говорить об экологическом соперничестве между странами и даже экологических войнах или войнах, обусловленных экологическими причинами. В свое время Гитлер оправдывал агрессивную политику нацистской Германии борьбой за жизненное пространство. Ресурсы нефти, угля и т.п. и тогда были важны. Еще больший вес они имеют в XXI в. К тому же добавилась необходимость территорий для захоронения радиоактивных и прочих отходов. Войны — горячие и холодные — приобретают экологическую окраску. Многие события в современной истории, например распад СССР, воспринимаются по-новому, если на них посмотреть с экологических позиций. Одна цивилизация может не только завоевать другую, но использовать ее для корыстных с экологической точки зрения целей. Это и будет экологический колониализм. Так переплетаются политические, социальные и экологические проблемы.
Основной закон экологии
Одним из главных достижений экологии стало открытие, что развиваются не только организмы и виды, но и . Последовательность сообществ, сменяющих друг друга в данном районе, называется сукцессией. Сукцессия происходит в результате изменения физической среды под действием сообщества, т.е. контролируется им.
Высокая продуктивность дает низкую надежность — еще одна формулировка основного закона экологии, из которой вытекает следующее правило: «Оптимальная эффективность всегда меньше максимальной». Разнообразие в соответствии с основным законом экологии непосредственно связано с устойчивостью. Однако пока неизвестно, до какой степени эта связь является причинно-следственной.
Некоторые другие важные для экологии законы и принципы.
Закон эмерджентности : целое всегда имеет особые свойства, отсутствующие у его части.
Закон необходимого разнообразия : система не может состоять из абсолютно идентичных элементов, но может иметь иерархическую организацию и интегративные уровни.
Закон необратимости эволюции : организм (популяция, вид) не может вернуться к прежнему состоянию, осуществленному в ряду его предков.
Закон усложнения организации : историческое развитие живых организмов приводит к усложнению их организации путем дифференциации органов и функций.
Биогенетический закон (Э. Геккель): онтогенез организма есть краткое повторение филогенеза данного вида, т.е. индивид в своем развитии повторяет сокращенно историческое развитие своего вида.
Закон неравномерности развития частей системы : системы одного уровня иерархии развиваются не строго синхронно, в то время как одни достигают более высокой стадии развития, другие остаются в менее развитом состоянии. Этот закон непосредственно связан с законом необходимого разнообразия.
Закон сохранения жизни : жизнь может существовать только в процессе движения через живое тело потока веществ, энергии, информации.
Принцип сохранения упорядоченности (Я. Пригожий): в открытых системах энтропия не возрастает, а уменьшается до тех пор, пока не достигается минимальная постоянная величина, всегда больше нуля.
Принцип Ле Шателье-Брауна : при внешнем воздействии, выводящем систему из состояния устойчивого равновесия, это равновесие смещается в том направлении, при котором эффект внешнего воздействия ослабляется.
Принцип экономии энергии (Л. Онсагер): при вероятности развития процесса в некотором множестве направлений, допускаемых началами термодинамики, реализуется то, которое обеспечивает минимум рассеивания энергии.
Закон максимизации энергии и информации : наилучшими шансами на самосохранение обладает система, в наибольшей степени способствующая поступлению, выработке и эффективному использованию энергии и информации; максимальное поступление вещества не гарантирует системе успеха в конкурентной борьбе.
Закон развития системы за счет окружающей среды : любая система может развиваться только за счет использования материально-энергетических и информационных возможностей окружающей ее среды; абсолютно изолированное саморазвитие невозможно.
Правило Шредингера «о питании» организма отрицательной энтропией: упорядоченность организма выше окружающей среды, и организм отдает в эту среду больше неупорядоченности, чем получает. Это правило соотносится с принципом сохранения упорядоченности Пригожина.
Правило ускорения эволюции : с ростом сложности организации биосистем продолжительность существования вида в среднем сокращается, а темпы эволюции возрастают. Средняя продолжительность существования вида птиц — 2 млн лет, вида млекопитающих — 800 тыс. лет. Число вымерших видов птиц и млекопитающих в сравнении со всем их числом велико.
Закон относительной независимости адаптации : высокая адаптивность к одному из экологических факторов не дает такой же степени приспособления к другим условиям жизни (наоборот, она может ограничивать эти возможности в силу физиолого-морфологических особенностей организмов).
Принцип минимального размера популяций : существует минимальный размер популяции, ниже которого ее численность не может опускаться.
Правило представительства рода одним видом : в однородных условиях и на ограниченной территории таксономический род, как правило, представлен только одним видом. По-видимому, это связано с близостью экологических ниш видов одного рода.
Закон обеднения живого вещества в островных его сгущениях (Г.Ф. Хильми): «Индивидуальная система, работающая в среде с уровнем организации более низким, чем уровень самой системы, обречена: постепенно теряя структуру, система через некоторое время растворится в окружающей среде». Из этого следует важный вывод для человеческой природоохранной деятельности: искусственное сохранение экосистем малого размера (на ограниченной территории, например, заповедника) ведет к их постепенной деструкции и не обеспечивает сохранения видов и сообществ.
Закон пирамиды энергий (Р. Линдеман): с одного трофического уровня экологической пирамиды переходит на другой, более высокий уровень в среднем около 10% поступившей на предыдущий уровень энергии. Обратный поток с более высоких на более низкие уровни намного слабее — не более 0,5-0,25%, и потому говорить о круговороте энергии в биоценозе не приходится.
Правило обязательности заполнения экологических ниш : пустующая экологическая ниша всегда и обязательно бывает естественно заполнена («природа не терпит пустоты»).
Принцип формирования экосистемы : длительное существование организмов возможно лишь в рамках экологических систем, где их компоненты и элементы дополняют друг друга и взаимно приспособлены. Из этих экологических законов и принципов следуют некоторые выводы, справедливые для системы «человек — природная среда». Они относятся к типу закона ограничения разнообразия, т.е. накладывают ограничения на деятельность человека по преобразованию природы.
Закон бумеранга : все, что извлечено из биосферы человеческим трудом, должно быть возвращено ей.
Закон незаменимости биосферы : биосферу нельзя заменить искусственной средой, как, скажем, нельзя создать новые виды жизни. Человек не может построить вечный двигатель, в то время как биосфера и есть практически «вечный» двигатель.
Закон шагреневой кожи : глобальный исходный природно-ресурсный потенциал в ходе исторического развития непрерывно истощается. Это следует из того, что никаких принципиально новых ресурсов, которые могли бы появиться, в настоящее время нет. Для жизни каждого человека в год необходимо 200 т твердых веществ, которые он с помощью 800 т воды и в среднем 1000 Вт энергии превращает в полезный для себя продукт. Все это человек берет из уже имеющегося в природе.
Принцип удаленности события : потомки что-нибудь придумают для предотвращения возможных отрицательных последствий. Вопрос о том, насколько законы экологии можно переносить на взаимоотношения человека с окружающей средой, остается открытым, так как человек отличается от всех других видов. Например, у большинства видов скорость роста популяции уменьшается с увеличением ее плотности; у человека, наоборот, рост населения в этом случае ускоряется. Некоторые регулирующие механизмы природы отсутствуют у человека, и это может служить дополнительным поводом для технологического оптимизма у одних, а для экологических пессимистов свидетельствовать об опасности такой катастрофы, которая невозможна ни для одного иного вида.
Применительно к деятельности по природопользованию и охране природной среды часто приходится слышать о необходимости учета законов развития природы. Человек, осознав свою роль в биосфере лишь как одного из многочисленных видов, образующих ее многообразие, как часть ее, должен, как и все остальные, подчиняться законам природы. При этом сила homo sapiens заключается не в том, чтобы, проявляя свою мощь, перестраивать природу, а в том, чтобы, правильно поняв законы ее развития, следовать им. Законы развития природы - законы более высокого порядка для человека в сравнении с законами развития общества. Это - объективные законы. В силу их действия и благодаря им человек появился и может существовать. Законы общества написаны человеком для собственного социального, политического и экономического удобства, организации и обеспечения общежития.
Знание и соблюдение законов развития природы в деятельности человека и общества имеет решающее значение и оценивается как императив. Проявляющиеся во взаимодействии общества и природы законы развития природы создают естественно-научные и философские основы разнообразной деятельности по природопользованию и охране окружающей среды, в том числе в сфере права. Учет законов природы при планировании и осуществлении экологически вредной деятельности и их соблюдение должно служить основным критерием экологической обоснованности и допустимости такой деятельности. Их знание и учет особенно важны при осуществлении таких правовых мер охраны природы, как нормирование предельно допустимых воздействий на природу, оценка воздействия планируемой деятельности на окружающую среду, экологическая экспертиза, планирование мер по охране природы и др. Законы развития природы должны учитываться также при подготовке законопроектов об охране окружающей среды. Обеспечение учета и соблюдение законов природы при принятии хозяйственных, управленческих и иных экологически значимых решений - одно из условий, методологическая основа выхода из экологического кризиса.
Рассмотрим некоторые основные законы развития природы в интерпретации одного из ведущих ученых-экологов России профессора Н.Ф. Реймерса.
Закон биогенной миграции атомов (В.И. Вернадского). Миграция химических элементов на земной поверхности и в биосфере в целом осуществляется или при непосредственном участии живого вещества (биогенная миграция) или протекает в среде, геохимические особенности которой (О 2 , С0 2 , Н 2 и т.д.) обусловлены живым веществом - как тем, которое в настоящее время населяет биосферу, так и тем, которое было на Земле в течение всей геологической истории.
Согласно этому Закону, имеющему важное теоретическое и практическое значение, понимание общих химических процессов, протекавших и протекающих на поверхности суши, в атмосфере и в заселенных организмами глубинах литосферы и вод, а также геологических слоях, сложенных прошлой деятельностью организмов, невозможно без учета биотических и биогенных факторов, в том числе эволюционных. Поскольку люди воздействуют прежде всего на биосферу и ее живое население, они тем самым изменяют условия биогенной миграции атомов, создавая предпосылки для еще более глубоких химических перемен в исторической перспективе. Таким образом, процесс может стать саморазвивающимся, не зависящим от желания человека и практически, при глобальном размахе, неуправляемым. Отсюда одна из самых насущных потребностей - сохранение живого покрова Земли в относительно неизменном состоянии. Тот же Закон определяет и необходимость учета воздействий на биоту при любых проектах преобразования природы. В этих случаях происходят региональные и локальные изменения в химических процессах, ведущие при любых крупных ошибках к деградации среды - опустыниванию.
Закон внутреннего динамического равновесия. Вещество, энергия, информация и динамические качества отдельных природных систем и их иерархия взаимосвязаны настолько, что любое изменение одного из этих показателей вызывает сопутствующие функциональные структурные качественные и количественные перемены, сохраняющие общую сумму вещественно-энергетических, информационных и динамических качеств систем, где эти изменения происходят, или в их иерархии.
Эмпирическим путем установлен ряд следствий действия данного Закона:
- а) любое изменение среды (веществ, энергии, информации, динамических качеств экосистем) неизбежно приводит к развитию природных цепных реакций, направленных в сторону нейтрализации произведенного изменения или формирования новых природных систем, образование которых при значительных изменениях среды может принять необратимый характер;
- б) взаимодействие вещественно-энергетических экологических компонентов (энергии, газов, жидкостей и др.), информации и динамических качеств природных систем количественно не является линейным, т.е. слабое воздействие или изменение одного из показателей может вызвать сильные отклонения в других (и во всей системе в целом);
- в) производимые в крупных экосистемах перемены относительно необратимы. Проходя по иерархии снизу вверх - от места воздействия до биосферы в целом, они меняют глобальные процессы и тем самым переводят их на новый эволюционный уровень;
- г) любое местное преобразование природы вызывает в глобальной совокупности биосферы и в ее крупнейших подразделениях ответные реакции, приводящие к относительной неизменности эколого-экономического потенциала («правило Тришкина кафтана»), увеличение которого возможно лишь путем значительного возрастания энергетических вложений.
Закон внутреннего динамического равновесия - одно из узловых положений в природопользовании. Пока изменения среды слабы и произведены на относительно небольшой площади, они или ограничиваются конкретным местом, или «гаснут» в цепи иерархии экосистем. Но как только перемены достигают существенных значений для крупных экосистем, например происходят в масштабах больших речных бассейнов, они приводят к существенным сдвигам в этих обширных природных образованиях, а через них, в соответствии со следствием б), и во всей биосфере Земли.
Закон «все или ничего» (X. Боулича). Слабые воздействия могут не вызывать у природной системы ответных реакций до тех пор, пока, накопившись, не приведут к развитию бурного динамического процесса. Закон полезен при экологическом прогнозировании.
Закон константности (В.И. Вернадского). Количество живого вещества природы (для данного геологического периода) есть константа. Любое изменение количества живого вещества в одном из регионов биосферы неминуемо влечет за собой такую же по размеру его перемену в каком-либо регионе, но с обратным знаком. Полярные изменения могут быть использованы в процессах управления природой, но следует учитывать, что не всегда происходит адекватная замена. Обычно высокоразвитые виды и экосистемы вытесняются другими, стоящими на относительно эволюционно более низком уровне (крупные организмы - более мелкими), а полезные для человека формы- менее полезными, нейтральным или даже вредными.
Закон минимума (Ю. Либиха). Выносливость организма определяется самым слабым звеном в цепи его экологических потребностей, т.е. жизненные возможности лимитируют экологические факторы, количество и качество которых близки к необходимому организму или экосистеме минимуму; дальнейшее их снижение ведет к гибели организма или деструкции экосистемы.
Закон ограниченности природных ресурсов. Все природные ресурсы (и условия) Земли конечны. Закон основан на том, что, поскольку планета представляет собой естественно ограниченное целое, на ней не могут существовать бесконечные части. Следовательно, категория «неисчерпаемых» природных ресурсов возникла по недоразумению.
Закон развития природной системы за счет окружающей ее среды. Любая природная система может развиваться только за счет использования материально-энергетических и информационных возможностей окружающей ее среды. Абсолютно изолированное саморазвитие невозможно. Этот же Закон имеет чрезвычайно важное теоретическое и практическое значение благодаря основным своим следствиям:
- а) абсолютно безотходное производство невозможно;
- б) любая более высокоорганизованная биотическая система (например, вид живого), используя и видоизменяя среду жизни, представляет потенциальную угрозу для менее организованных систем;
- в) биосфера Земли как система развивается не только за счет ресурсов планеты, но опосредованно за счет и под управляющим воздействием космических систем (прежде всего Солнечной).
Согласно первому следствию мы можем рассчитывать лишь на малоотходное производство. Поэтому первым этапом развития технологий должна быть их малая ресурсоемкость (как на входе, так и на выходе - экономность и незначительные выбросы), вторым этапом будет создание цикличности производств (отходы одних могут быть сырьем для других) и третьим - организация разумного захоронения неминуемых остатков и нейтрализация неустранимых энергетических отходов. Представление, будто биосфера работает по принципу безотходности, ошибочно, так как в ней всегда накапливаются выбывающие из биологического круговорота вещества, формирующие осадочные породы.
Согласно второму следствию рассматриваемого Закона воздействие человека на природу требует мероприятий по нейтрализации этих воздействий, поскольку они могут оказаться разрушающими для остальной природы и, согласно правилу соответствия условий среды генетической предопределенности организма, угрожают и самому человеку. В связи с этим охрана природы - одно из обязательных составляющих социально-экономического развития высокоразвитого общества.
Третье следствие Закона имеет особое значение для долгосрочного прогнозирования. Оно должно учитываться при рассмотрении всех процессов, происходящих на Земле.
Закон (закономерность) снижения природоемкости готовой продукции. Удельное содержание природного вещества в усредненной единице общественного продукта исторически неуклонно снижается. Это не означает, что в процесс производства вовлекается меньше природного вещества. Наоборот, его количество увеличивается - выбрасывается до 95-98% потребляемого в производстве природного вещества. Однако в конечных аналогичных продуктах общественного производства в наши дни, вероятно, усредненно меньше природного вещества, чем в отдаленном прошлом. Объясняется это миниатюризацией изделий, заменой естественных материалов и продуктов синтетическими, а также другими явлениями. Поскольку точных расчетов еще не сделано (методически они затруднительны), этот Закон имеет характер экспертного умозаключения.
Закон снижения энергетической эффективности природопользования. С ходом исторического времени при получении из природных систем полезной продукции на ее единицу затрачивается в среднем все больше энергии.
Расход энергии на одного человека (в ккал/сут.) в каменном веке был порядка 4 тыс., в аграрном обществе - 12 тыс., в индустриальную эпоху - 7 тыс., а в передовых развитых странах настоящего времени - 230-250 тыс., т.е. в 58-62 раза больше, чем у далеких предков. С начала нашего века количество энергии, затрачиваемой на одну единицу сельскохозяйственной продукции, в развитых странах мира возросло в 8-10 раз. Общая энергетическая эффективность сельскохозяйственного производства (соотношение вкладываемой и получаемой с готовой продукцией энергии) в промышленно развитых странах примерно в 30 раз ниже, чем в условиях примитивного земледелия. В ряде случаев увеличение в десятки раз затрат энергии на удобрения и обработку полей приводит лишь к весьма незначительному (на 10-15%) повышению урожайности. Это связано с необходимостью параллельно с улучшением агротехники учитывать общую экологическую обстановку, налагаемые ею ограничения. В начале 80-х гг. удельные затраты энергии на производство единицы валового национального продукта (ВНП) в ходе решительных мер по экономии энергии в промышленно развитых странах сократились на 15%. В течение последнего десятилетия ВНП возрос тут на 20%, а потребление энергии - лишь на 2% (это стало возможно в результате устранения неоправданных потерь энергии).
Закон убывающего (естественного) плодородия. В связи с постоянным изъятием урожая и нарушением естественных процессов почвообразования, а также при длительной монокультуре, в результате накопления токсичных веществ, выделяемых растениями, на культивируемых землях постепенно происходит снижение естественного плодородия почв. Этот процесс частично нейтрализуется накоплением биомассы подземных частей культурных растений, но главным образом внесением удобрений (созданием искусственного плодородия). К настоящему времени плодородие в той или иной степени потеряно приблизительно у 50% всех пахотных угодий мира (от 1,5-1,6 до 2 млрд. га) при средней скорости потерь в 70-х гг. 6,8, в 80-х - около 7 млн. га в год. Интенсификация сельского хозяйства позволяет получать все большие урожаи при меньших затратах человеческого труда и частично нейтрализовать действие Закона убывающего плодородия, но в то же время падает энергетическая эффективность производства.
Закон физико-химического единства живого вещества (В.И. Вернадского). Все живое вещество Земли физико-химически едино. Из Закона естественно вытекает следствие: вредное для одной части живого вещества не может быть безразлично для другой его части, или: вредное для одних видов существ вредно и для других. Отсюда - любые физико-химические агенты, смертельные для одних организмов (например, средства борьбы с вредителями), не могут не оказывать вредного влияния на другие организмы. Вся разница состоит лишь в степени устойчивости видов к агенту. Поскольку в любой многочисленной популяции всегда находятся разнокачественные особи, в том числе менее или более устойчивые к физико-химическим влияниям, скорость отбора по выносливости популяций к вредному агенту прямо пропорциональна скорости размножения организмов, быстроте чередования поколений. Исходя из этого, при растущем воздействии физико-химического фактора, к которому организм с относительно медленной сменой поколений устойчив, на менее устойчивый, но быстрее размножающийся вид их способность противостоять рассматриваемому фактору уравнивается. Именно поэтому длительное применение химических методов борьбы с вредителями растений и возбудителями болезней человека и теплокровных животных экологически неприемлемо. С отбором устойчивых особей быстро размножающихся членистоногих нормы обработки приходится увеличивать. Однако и эти увеличенные концентрации оказываются малоэффективными, но тяжело отражаются на здоровье людей и позвоночных животных.
Закон экологической корреляции. В экосистеме, как и в любом другом целостном природно-системном образовании, особенно в биотическом сообществе, все входящие в нее виды живого и абиотические экологические компоненты функционально соответствуют друг другу. Выпадение одной части системы (например, уничтожение вида) неминуемо ведет к исключению всех тесно связанных с этой частью системы других ее частей и функциональному изменению целого в рамках Закона внутреннего динамического равновесия. Закон экологической корреляции особенно важен для сохранения видов живого, никогда не исчезающих изолированно, но всегда взаимосвязанной группой. Действие Закона приводит к скачкообразности в изменении экологической устойчивости: при достижении порога изменения функциональной целостности происходит срыв (часто неожиданный) - экосистема теряет свойство надежности. Например, многократное увеличение концентрации вещества-загрязнителя может не приводить к катастрофическим последствиям, но затем ничтожная его прибавка приведет к катастрофе.
Известный американский ученый-эколог Б. Коммонер сводит основные законы экологии к следующему: 1) все связано со всем; 2) все должно куда-то деваться; 3) природа «знает» лучше; 4) ничто не дается даром.
Обитание жизни развивается в результате постоянного обмена веществ информацией на базе потока энергии в совокупном единстве среды и населяющих её организмов.
40. Закон минимума (Либиха): Веществом, присутствующим в минимуме, управляет урожай, определяется его величина, и стабильность во времени.
41. Законы Коммонера:
«Все связано со всем»;
«Все должно куда-то деваться»;
«Ничто не дается даром»;
«Природа знает лучше».
42. Закон максимума (Шелфорда): Процветание организма ограничено зонами максимума и минимума определенных экологических факторов; между ними располагается зона экологического оптимума, в пределах которого организм нормально реагирует на условия среды.
43. Деградация биосферы - это разрушение или существенное нарушение экологических связей в природе, сопровождающееся ухудшением условий жизни человека, вызванное стихийными бедствиями или хозяйственной деятельностью самого человека, производимой без учета знания законов развития природы.
44.Этапы деградации биосферы:
использование огня (ранний палеолит);
развитие сельского хозяйства;
промышленная революция.
экологический кризис.
45. Источники деградации биосферы могут быть естественными (природными) и искусственными (антропогенными). Естественные загрязнение ОС вызвано природными процессами (пыльные бури, вулканизм, лесные пожары и т.д.). Искусственные загрязнение связи с выбросами в ОС различных загрязняющих веществ в процессе деятельности человека (сельское хозяйство, транспорт, промышленность и т.д.)
46. Последствия деградации биосферы:
Заметное уменьшение биоразнообразия экосистемы, разрушение и уничтожение ещё сохранившихся участков дикорастущей растительности, варварское уничтожение лесов и болот, сокращение численности диких животных, исчезновение многих представителей флоры и фауны. В результате всех этих действий к середине ХХ столетия антропогенное воздействие на биосферу по своему значению вошло на один уровень с естественным, приняв планетарные масштабы. Таким образом, человечество превратилось в один из основных геоэкологических судьбоносных факторов эволюции планеты.
47. Загрязнение – любое внесения в ту или иную экологическую систему (биоценоз) не свойственных ей живых или неживых компонентов, любых изменений, прерывающих или нарушающих процессы круговорота и обмена веществ, потоки энергии, следствием которых является снижения продуктивности или разрушения данной системы.
48.Основные загрязняющие вещества:
диоксид углерода (СО 2);
оксид углерода (СО);
диоксид серы (SO 2);
оксиды азота (NO, NO 2 , N 2 O);
тяжелые металлы и в первую очередь ртуть, свинец и кадмий;
радионуклиды и другие радиоактивные вещества;
диоксиды (хлоруглеводороды);
твердые примеси (аэрозоли): пыль, сажа, дым;
нефть и нефтепродукты.
канцерогенные вещества, в частности, бензапирен;
пестициды;
49. По агрегатному состоянию различают 3 вида загрязнителей: твердые, жидкие и газообразные.
50. По происхождению природы, агрегатного состояния, масштабу распространения, вызванным последствиям, степени токсичности
51. По природе загрязняющие вещества классифицируют на такие группы: химические, физические, биологические,эстетические.