VACUOL CỦA TẾ BÀO THỰC VẬT
Các bào quan có tầm quan trọng chung, có cấu trúc đơn màng và xuất hiện chỉ một trong tế bào thực vật.
Kích thước và số lượng: được xác định bởi tuổi của tế bào. Ở tế bào non, không bào phát sinh từ các túi nhỏ thoát ra khỏi EPS. Khi tế bào phát triển và biệt hóa, các không bào nhỏ hợp nhất với nhau và tạo thành một hoặc nhiều không bào lớn, chiếm tới 80% thể tích của toàn bộ tế bào. Kết quả là tế bào chất với nhân và các bào quan bị đẩy ra ngoại vi.
Kết cấu: không bào trung tâm được ngăn cách với tế bào chất bằng một màng duy nhất - màng không bào, có độ dày tương tự như plasmalemma. Khoang không bào được lấp đầy Tế bào nhựa cây. Thành phần của nhựa tế bào bao gồm muối vô cơ, đường, axit hữu cơ và muối của chúng, các chất có trọng lượng phân tử thấp khác, cũng như một số hợp chất có trọng lượng phân tử cao (ví dụ: protein).
Chức năng: 1) điều hòa thẩm thấu: do tính bán thấm của tonoplast và plasmalemma, nồng độ phân tử tương ứng của nhựa tế bào được duy trì, tức là. không bào có chức năng như một máy đo thẩm thấu;
2) bài tiết: tất cả các sản phẩm trao đổi chất hòa tan trong nước (alkaloid - nicotin, caffeine; polyphenol) có thể được loại bỏ thông qua tonoplast;
3) lưu trữ: photphat K + , Na + , Ca 2+ , muối của axit hữu cơ (oxalat, citrate, v.v.), đường và protein tích tụ trong nhựa tế bào.
RIBOsome
Một bào quan không có cấu trúc màng. Đây là cơ quan duy nhất có tầm quan trọng chung hiện diện trong tế bào của cả sinh vật nhân sơ và sinh vật nhân chuẩn. Ribosome lần đầu tiên được mô tả ở 1955 G. J. Palade (hạt Palade), người đã chứng minh rằng chúng là phức hợp ribonucleoprotein (RNP).
Tính chất hóa học: RNP= rRNA+protein.
Ribosome chiếm 85% ARN, được trình bày trong một ô.
Hình thức: Ribosome có hình nấm vì nó bao gồm hai tiểu đơn vị: to lớn Và bé nhỏ, giữa chúng nằm trung tâm chức năng của ribosome (FCR) , trong đó, trong quá trình sinh tổng hợp protein (giai đoạn dịch mã), mRNA nằm ở hai bộ ba của nó và một phức hợp enzyme hoạt động, đảm bảo sự lắp ráp một phân tử protein từ các axit amin.
Kích thước: 15 – 35nm. Kích thước của ribosome hoàn chỉnh ở tế bào nhân sơ là 20x17x17 nm, ở tế bào nhân chuẩn - 25x20x20 nm.
Nơi giáo dục: sự hình thành các tiểu đơn vị ribosome xảy ra trong nhân của nhân. Sự tập hợp các tiểu đơn vị thành ribosome hoàn chỉnh xảy ra trong tế bào chất khi nồng độ ion magie (Mg 2+) đạt 0,001 M; nếu nồng độ này giảm thì xảy ra sự phân ly của các tiểu đơn vị. Khi nồng độ Mg 2+ tăng gấp 10 lần, đạt giá trị 0,01 M, hai ribosome tương tác với nhau tạo thành dimer.
Không bào và nhựa tế bào
Hầu hết các tế bào thực vật trưởng thành đều có đặc điểm là không bào trung tâm lớn, chiếm tới 70-90% thể tích tế bào. Trong trường hợp này, protoplast với tất cả các bào quan nằm ở dạng một lớp thành rất mỏng lót thành tế bào. Không bào tế bào chất nhỏ thường được tìm thấy trong protoplast của thành. Đôi khi nhân nằm ở trung tâm tế bào túi hạt nhân tế bào chất, được kết nối với lớp vách bằng các sợi tế bào chất mỏng nhất đi qua không bào trung tâm.
Nhựa tế bào là dung dịch nước của nhiều chất khác nhau, là sản phẩm của hoạt động sống của tế bào trần, chủ yếu là chất dự trữ và chất thải. Phản ứng của nhựa tế bào thường có tính axit nhẹ hoặc trung tính, ít có tính kiềm hơn. Các chất tạo nên nhựa tế bào vô cùng đa dạng. Đó là carbohydrate, protein, axit hữu cơ và muối, axit amin, ion khoáng, alkaloid, glycoside, tannin, sắc tố và các hợp chất hòa tan trong nước khác. Hầu hết chúng thuộc nhóm chất ergastic - sản phẩm của quá trình chuyển hóa protoplast, có thể xuất hiện và biến mất ở các giai đoạn sống khác nhau của tế bào. Nhiều chất nhựa tế bào chỉ được hình thành trong tế bào thực vật.
Carbohydrate trong nhựa tế bào thực vật được thể hiện bằng monosaccharides (glucose, fructose), disaccharides (sucrose) và polysaccharides (mucus, inulin).
Glucose (đường nho) và fructose (đường trái cây) tích tụ với số lượng lớn trong trái cây mọng nước. Sucrose (đường củ cải) tích tụ với số lượng lớn trong rễ củ cải đường và thân cây mía. Một số họ thực vật (cactaceae, Crassulaceae, lan) có đặc điểm là tích tụ chất nhầy trong nhựa tế bào, có tác dụng giữ nước. Inulin là một polysaccharide dự trữ, lắng đọng dưới dạng dung dịch keo trong nhựa tế bào của các cơ quan ngầm của họ Cúc thay vì tinh bột.
Protein tích tụ dưới dạng dung dịch keo trong không bào của tế bào hạt chín. Khi hạt bị mất nước trong giai đoạn phát triển sau này, nước sẽ bị loại bỏ khỏi không bào, nồng độ protein trong nhựa tế bào tăng lên và chuyển sang trạng thái gel rắn. Không bào mất nước của hạt trưởng thành được gọi là hạt aleuron.
Trong số các axit hữu cơ trong nhựa tế bào, phổ biến nhất là citric, malic, succinic và oxalic. Những axit này được tìm thấy với số lượng lớn trong nhựa tế bào của trái cây chưa chín, khiến chúng có vị chua. Khi quả chín, axit hữu cơ có thể được sử dụng làm chất nền hô hấp nên vị chua của quả thường biến mất. Muối của axit hữu cơ cùng với các ion khoáng đóng vai trò quan trọng trong quá trình thẩm thấu.
Tannin (tannin) là các hợp chất phenolic polyme có vị chát. Chúng có đặc tính sát trùng và bảo vệ các mô thực vật khỏi bị nhiễm trùng và sâu răng. Các tế bào vỏ thân và rễ (sồi, liễu), quả chưa chín (quả óc chó), lá (chè) và một số bệnh phát triển - mật đặc biệt giàu tannin. Tannid được sử dụng trong y học, thuộc da và nhuộm vải màu nâu sẫm.
Alkaloid là các chất hữu cơ chứa nitơ đa dạng về mặt hóa học, có vị đắng. Chúng có đặc tính của bazơ và được tìm thấy trong nhựa tế bào, thường ở dạng muối. Nhiều loại thực vật chứa alkaloid có độc và không được động vật ăn cỏ ăn. Trong các tế bào có chứa ancaloit, bào tử và vi trùng của vi sinh vật không phát triển và thực vật không bị ảnh hưởng bởi các bệnh do nấm và vi khuẩn. Đại diện của các họ Solanaceae, Poppy, Rubiaceae, Ranunculaceae, v.v. đặc biệt giàu alkaloid.
Glycoside là một nhóm lớn các chất tự nhiên, hợp chất của đường với rượu, aldehyd, phenol và các chất khác. Một số glycosid thực vật được sử dụng trong y học. Glycoside cũng bao gồm các sắc tố nhựa tế bào - chất flavonoid. Một trong số chúng - anthocyanin– làm cho nhựa tế bào có màu đỏ, xanh hoặc tím; khác - flavon- màu vàng. Anthocyanin có liên quan đến màu sắc hoa của nhiều loại cây. Bảng màu được xác định bởi phản ứng của nhựa tế bào: nếu có tính axit thì tông màu đỏ chiếm ưu thế, nếu trung tính, tông màu tím chiếm ưu thế và với phản ứng hơi kiềm, tông màu xanh lam chiếm ưu thế. Sự xuất hiện của các sắc thái cũng bị ảnh hưởng bởi sự hình thành phức hợp anthocyanin với các kim loại khác nhau. Flavones chịu trách nhiệm tạo ra màu vàng của cánh hoa của một số loại cây.
Tầm quan trọng của axit hữu cơ, tannin, alkaloid và glycoside của nhựa tế bào trong quá trình chuyển hóa tế bào vẫn chưa được làm rõ đầy đủ. Trước đây, chúng được coi là sản phẩm trao đổi cuối cùng. Hiện nay người ta đã chứng minh rằng nhiều chất trong số chúng có thể tham gia lại vào quá trình trao đổi chất và do đó có thể được coi là chất dự trữ.
Ngoài chức năng tích lũy chất dự trữ và chất thải, không bào trong tế bào thực vật còn thực hiện một chức năng quan trọng khác - duy trì sức trương. Nồng độ các ion và đường trong nhựa tế bào của không bào trung tâm thường cao hơn trong thành tế bào; Tonoplast làm chậm đáng kể sự khuếch tán của các chất này từ không bào, đồng thời dễ dàng thấm vào nước. Vì vậy, nước sẽ đi vào không bào. Quá trình khuếch tán nước một chiều qua màng thấm chọn lọc này được gọi là sự thẩm thấu. Nước đi vào nhựa tế bào gây áp lực lên tế bào trần và xuyên qua nó lên thành tế bào, gây ra trạng thái căng, đàn hồi của nó, hoặc cơn co giật tế bào. Turgor đảm bảo rằng các cơ quan thực vật không được phân loại vẫn giữ được hình dạng và vị trí trong không gian cũng như khả năng chống lại các yếu tố cơ học.
Nếu một tế bào được đặt trong dung dịch ưu trương chứa một số loại muối hoặc đường không độc hại (tức là trong dung dịch có nồng độ cao hơn nồng độ nhựa tế bào), thì sẽ xảy ra sự giải phóng thẩm thấu nước từ không bào. Kết quả là thể tích của nó giảm đi, protoplast của thành đàn hồi di chuyển ra khỏi thành tế bào, sức trương biến mất và quá trình plasmolysis tế bào( Hình.2.9).
Cơm. 2.9. Sơ đồ plasmolysis: 1 – tế bào ở trạng thái trương lực; 2 – bắt đầu quá trình plasmolysis; 3 – quá trình plasmolysis hoàn toàn.
Plasmolysis thường có thể đảo ngược. Khi tế bào được đặt trong nước hoặc dung dịch nhược trương, nước lại được hấp thụ mạnh mẽ bởi không bào trung tâm, protoplast lại được ép vào thành tế bào và sức trương được phục hồi. Quá trình plasmolysis có thể đóng vai trò là một chỉ số về trạng thái sống của tế bào; một tế bào chết không bị plasmolyzed, vì nó không có màng thấm chọn lọc.
Mất sức trương làm cho cây bị héo. Khi héo trong không khí trong điều kiện không đủ nước, thành tế bào mỏng co lại đồng thời với tế bào trần và bị gấp lại.
Áp suất trương lực không chỉ duy trì hình dạng của các bộ phận không bị hóa gỗ của cây mà còn là một trong những yếu tố tăng trưởng tế bào, cung cấp chiều cao tế bào kéo dài, tức là do sự hấp thụ nước và tăng kích thước của không bào. Tế bào động vật không có không bào trung tâm, sự phát triển của chúng diễn ra chủ yếu do tăng số lượng tế bào chất nên kích thước tế bào động vật thường nhỏ hơn tế bào thực vật.
Không bào trung tâm phát sinh từ sự hợp nhất của nhiều không bào nhỏ có trong tế bào mô phân sinh (phôi). Những không bào tế bào chất này được cho là được hình thành bởi màng của mạng lưới nội chất hoặc bộ máy Golgi.
Bao gồm
Sự hình thành các tạp chất là do sự tích tụ quá mức của một số sản phẩm trao đổi chất trong một số bộ phận của tế bào - trong không bào, hyaloplasm, các bào quan khác nhau và ít gặp hơn trong thành tế bào. Các chất này thường kết tủa ở dạng vô định hình hoặc tinh thể - sự bao gồm. Các tạp chất có hình dạng nhất định và có thể nhìn thấy rõ dưới kính hiển vi ánh sáng. Bằng sự hiện diện của các tạp chất nhất định, hình dạng và sự phân bố của chúng, một loài, chi và họ thực vật có thể được phân biệt với các loài khác, do đó chúng thường đóng vai trò là đặc điểm chẩn đoán quan trọng trong phân tích nguyên liệu cây thuốc.
Bao gồm là một trong hai chất phụ tùng(các hợp chất tạm thời bị loại bỏ khỏi quá trình trao đổi chất), hoặc sản phẩm cuối cùng trao đổi. Loại bao gồm đầu tiên bao gồm hạt tinh bột,giọt lipid Và tiền gửi protein; đến thứ hai - tinh thể một số chất.
Hạt tinh bột- những thể vùi phổ biến nhất của tế bào thực vật. Polysacarit tinh bột– Là loại chất dinh dưỡng dự trữ chính cho cây trồng. Nó cũng là hợp chất quan trọng nhất được động vật ăn cỏ sử dụng làm thức ăn. Tinh bột từ các loại ngũ cốc, củ khoai tây, quả chuối là nguồn dinh dưỡng quan trọng nhất của con người. Bột mì bao gồm gần 75% hạt tinh bột, trong củ khoai tây, tinh bột chiếm 20-30%. Về mặt hóa học, tinh bột là một alpha-1,4-D-glucan, các phân tử có dạng mạch nhánh, trong hạt tinh bột chúng nằm dọc theo bán kính.
Các hạt tinh bột được hình thành trong chất nền của plastid. Ở lục lạp, hạt được lắng đọng dưới ánh sáng tinh bột đồng hóa (sơ cấp), được hình thành khi có quá nhiều đường - sản phẩm của quá trình quang hợp. Sự hình thành tinh bột không hoạt động thẩm thấu ngăn cản sự gia tăng áp suất thẩm thấu trong lục lạp. Vào ban đêm, khi quá trình quang hợp không diễn ra, tinh bột đồng hóa sẽ bị thủy phân thành đường với sự trợ giúp của enzyme và vận chuyển đến các bộ phận khác của cây. Tinh bột dự phòng (thứ cấp) lắng đọng trong amyloplast của tế bào của các cơ quan thực vật khác nhau (rễ, chồi ngầm, hạt) từ đường chảy từ tế bào quang hợp. Nếu cần thiết, tinh bột dự trữ cũng được chuyển hóa thành đường.
Sự hình thành các hạt tinh bột bắt đầu tại một số điểm nhất định trong chất nền của plastid, được gọi là trung tâm giáo dục. Sự phát triển của hạt xảy ra bằng cách lắng đọng liên tiếp các lớp tinh bột xung quanh trung tâm giáo dục. Các lớp liền kề trong một hạt có thể có chiết suất ánh sáng khác nhau và sau đó chúng có thể nhìn thấy được dưới kính hiển vi - nhiều lớp hạt tinh bột. Việc sắp xếp các lớp có thể đồng tâm(lúa mì) hoặc kỳ dị(khoai tây) ( cơm. 2.10). Nếu có một trung tâm giáo dục trong amyloplast, xung quanh đó các lớp tinh bột được lắng đọng, thì đơn giản, nếu hai hoặc nhiều hơn thì nó được hình thành hạt phức tạp, bao gồm một số cái đơn giản. hạt bán phức hợpđược hình thành khi tinh bột lần đầu tiên lắng đọng xung quanh một số điểm, và sau đó, sau khi tiếp xúc với các hạt đơn giản, các lớp chung xuất hiện xung quanh chúng ( cơm. 2.10).
Hình dạng, kích thước, số lượng trong amyloplast và cấu trúc (vị trí của trung tâm giáo dục, phân lớp, có hay không có vết nứt) của hạt tinh bột thường đặc trưng cho từng loài thực vật ( cơm. 2.10). Thông thường, các hạt tinh bột có dạng hình cầu, hình trứng hoặc hình thấu kính, nhưng ở khoai tây thì chúng không đều. Các hạt lớn nhất (lên tới 100 micron) là đặc trưng của tế bào củ khoai tây, trong hạt lúa mì chúng có hai kích cỡ - nhỏ (2-9 micron) và lớn hơn (30-45 micron). Tế bào hạt ngô có đặc điểm là hạt nhỏ (5-30 micron). Gạo, yến mạch và kiều mạch có các hạt tinh bột phức tạp.
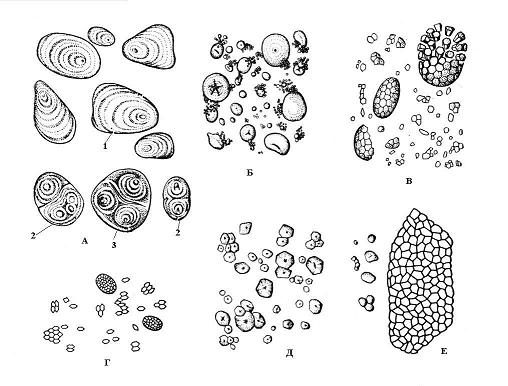
Cơm. 2.10. Hạt tinh bột của các loại thực vật khác nhau: A – khoai tây; B - lúa mì; B – yến mạch; G - gạo; D – ngô; E - kiều mạch; 1 – hạt đơn giản; 2 – hạt phức tạp; 3 – hạt bán phức tạp.
Thuốc thử tinh bột là dung dịch iốt trong dung dịch kali iodua - thuốc thử Lugol. Nó tạo màu cho hạt tinh bột có màu xanh tím.
Tinh bột tích tụ phổ biến trong tất cả các cơ quan của thực vật, nhưng hạt, chồi ngầm (củ, củ, thân rễ) và nhu mô của các mô dẫn điện của rễ và thân của cây gỗ đặc biệt giàu tinh bột.
Giọt lipidđược tìm thấy ở hầu hết các tế bào thực vật. Dầu béo tích tụ trong một số lượng lớn thực vật và là dạng dinh dưỡng dự trữ quan trọng thứ hai sau tinh bột. Hạt và trái cây đặc biệt giàu chúng. Hạt của một số loại cây (hướng dương, bông, lạc) có thể chứa tới 40% dầu tính theo trọng lượng chất khô. Vì vậy, chất béo thực vật được lấy chủ yếu từ hạt.
Các giọt lipid tích tụ trực tiếp trong hyaloplasm. Chúng trông giống như những vật thể hình cầu nhỏ, mỗi giọt được ngăn cách với hyaloplasm bằng một màng. Đôi khi giọt lipid được gọi là thể cầu.
Thuốc thử dầu béo là thuốc nhuộm SudanIII, giọt lipid có màu đỏ cam.
Bao gồm protein dưới dạng các chất lắng đọng vô định hình hoặc tinh thể khác nhau được hình thành trong các bào quan khác nhau của tế bào. Thông thường, các tinh thể protein có thể được tìm thấy trong nhân, ít gặp hơn - trong hyaloplasm, stroma plastid, trong phần mở rộng của mạng lưới nội chất, ma trận peroxisome và ty thể. Kích thước của tinh thể protein thường vượt quá độ phân giải của kính hiển vi ánh sáng.
Protein lưu trữ thuộc loại protein đơn giản - chất đạm, không giống như các protein phức tạp - protein, tạo thành cơ sở của protoplast. Chúng được lắng đọng với số lượng lớn nhất trong mô dự trữ hạt khô ở dạng hạt aleuron, hoặc thể protein.
Các hạt Aleurone thường có dạng hình cầu và kích thước khác nhau (0,2-20 µm). Chúng được bao quanh bởi một màng và chứa một ma trận protein vô định hình, trong đó các thể vùi tinh thể được nhúng vào - một (ít thường xuyên hơn, 2-3) tinh thể protein có hình thoi và tròn hình cầu(từ một đến nhiều) ( cơm. 2.11). Globoids bao gồm phytin (muối của axit inositol hexaphosphoric) và là nơi lưu trữ phốt pho dự trữ. Hạt aleurone chứa tinh thể được gọi là tổ hợp. Chúng là đặc trưng của các tế bào lưu trữ hạt có dầu (cây lanh, hướng dương, bí ngô, mù tạt, dầu thầu dầu, v.v.). Ít phổ biến đơn giản Các hạt aleurone không chứa tinh thể mà chỉ có protein vô định hình (các loại đậu, gạo, ngô) ( cơm. 2.12).
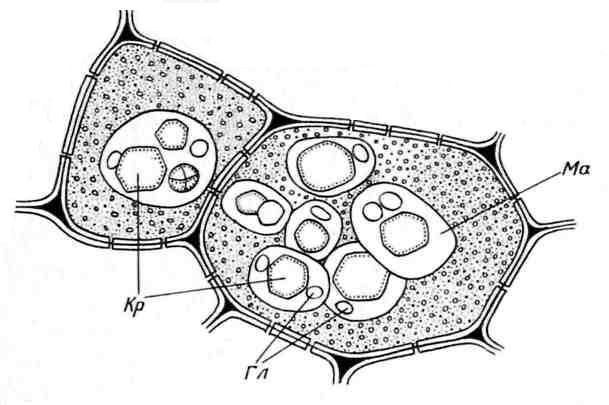
Cơm. 2.11. Hạt Aleurone trong tế bào nội nhũ của hạt thầu dầu:Kr- tinh thể protein; Gl– hình cầu; Mã– chất nền protein.
Trong quá trình phát triển của hạt, protein dự trữ được tích trữ trong không bào. Khi hạt chín, kèm theo tình trạng mất nước, không bào protein bị khô, protein và phytin kết tủa ra khỏi dung dịch và có thể kết tinh. Trong quá trình hạt nảy mầm, hạt aleurone hút nước, trương nở và chuyển hóa dần thành không bào điển hình. Protein và các chất của globoids được sử dụng cho sự sinh trưởng và phát triển của cây con.
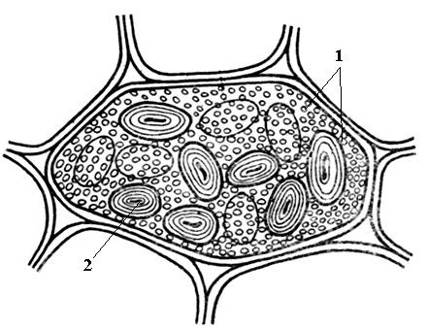
Cơm. 2.12. Các hạt aleuron đơn giản và tinh bột trong tế bào lá mầm hạt đậu: 1 – hạt aleurone đơn giản; 2 – hạt tinh bột.
Các thể vùi protein có thể được nhuộm màu vàng vàng bằng thuốc thử Lugol.
Tinh thể canxi oxalat thường thấy ở tế bào thực vật. Chúng chỉ được lắng đọng trong không bào. Hình dạng của tinh thể canxi oxalat khá đa dạng ( cơm. 2.13) và thường đặc trưng cho một số loại cây nhất định, được sử dụng để chẩn đoán nguyên liệu cây thuốc. Nó có thể đơn tinh thể hình thoi, bát diện hoặc thon dài (lá henbane), Druze– sự phát triển xen kẽ hình ngôi sao của các tinh thể hình cầu (lá cây hà thủ ô, cà độc dược, senna, rễ đại hoàng), rệp– các tinh thể hình kim nhỏ tập hợp thành chùm (lá hoa huệ, thân rễ cây điên thảo), bút chì– tinh thể lớn hơn, hình que (lá hoa huệ) và cát pha lê– cụm nhiều tinh thể đơn nhỏ (lá belladonna). Phổ biến nhất là drusen. 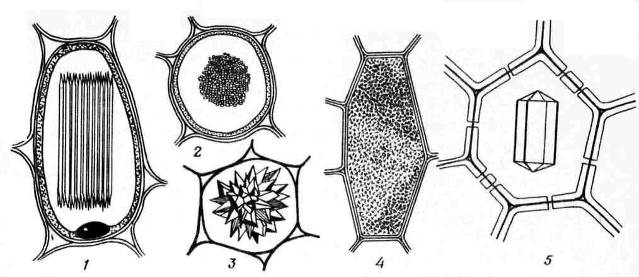
Cơm. 2.13. Các dạng tinh thể canxi oxalat: 1,2 – củ cải (1 – mặt bên, 2 – mặt cắt ngang); 3 – drusen; 4 – cát kết tinh; 5 – đơn tinh thể.
Dọc theo các sợi ở vỏ hoặc dọc theo gân lá của một số loại cây (vỏ sồi, rễ cam thảo, lá senna) lớp lót chứa tinh thể– Các tế bào xếp thành hàng song song với các tinh thể canxi oxalat đơn lẻ ( Hình.2.14).
![]()
Cơm. 2.14. Tĩnh mạch có lớp lót tinh thể trong lá senna.
Không giống như động vật giải phóng các ion dư thừa ra môi trường bên ngoài cùng với nước tiểu, thực vật chưa phát triển cơ quan bài tiết buộc phải tích tụ chúng trong mô của chúng. Người ta thường tin rằng tinh thể canxi oxalat là sản phẩm cuối cùng của đời sống tế bào, nhằm loại bỏ lượng canxi dư thừa. Thật vậy, các tinh thể được hình thành với số lượng lớn trong các cơ quan và mô mà thực vật thỉnh thoảng rụng đi (lá và vỏ cây). Tuy nhiên, có bằng chứng cho thấy tinh thể có thể biến mất khỏi không bào. Trong trường hợp này, chúng có thể được coi là nơi lắng đọng canxi dự trữ.
Bản chất hóa học của tinh thể canxi oxalat có thể được xác nhận bằng tác dụng của axit khoáng đậm đặc. Dưới tác dụng của axit clohydric, các tinh thể hòa tan. Khi tiếp xúc với axit sulfuric, canxi oxalat chuyển thành canxi sunfat (thạch cao) không hòa tan, tạo thành nhiều tinh thể hình kim.
Gần với các thể vùi tinh thể sỏi bàng quang. Chúng thường bao gồm canxi cacbonat hoặc silica và là các khối hình thành cụm phát sinh trên các hình chiếu của thành tế bào nhô vào trong tế bào ( cơm. 2,15). Cystoliths là đặc trưng của thực vật thuộc họ cây tầm ma và dâu tằm. Tầm quan trọng của sỏi bàng quang vẫn chưa được làm rõ.
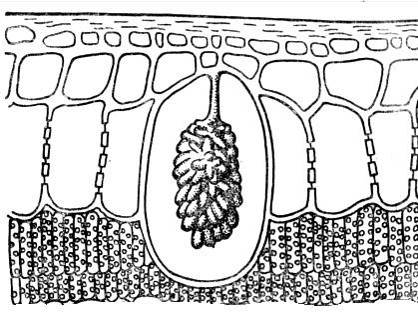
Cơm. 2.15. Cystolith trong tế bào biểu bì của lá ficus.
Vách tế bào
Vách tế bào (màng tế bào)- Đặc điểm nổi bật của tế bào thực vật để phân biệt nó với tế bào động vật. Thành tế bào tạo cho tế bào hình dạng cụ thể của nó. Tế bào thực vật được nuôi cấy trên môi trường dinh dưỡng đặc biệt, trong đó thành tế bào được loại bỏ bằng enzyme, luôn có dạng hình cầu. Thành tế bào mang lại sức mạnh cho tế bào và bảo vệ tế bào trần; nó cân bằng áp suất trương và do đó ngăn ngừa sự vỡ của plasmalemma. Tập hợp các thành tế bào tạo thành bộ xương bên trong, hỗ trợ cơ thể thực vật và mang lại cho nó độ bền cơ học.
Thành tế bào không màu và trong suốt, dễ dàng truyền ánh sáng mặt trời. Thông thường các bức tường được bão hòa với nước. Hệ thống thành tế bào vận chuyển nước và các hợp chất có trọng lượng phân tử thấp hòa tan trong đó (vận chuyển dọc theo apoplast).
Thành tế bào bao gồm chủ yếu là các polysaccharid, có thể được chia thành chất xương Và chất nền.
Chất xương vách tế bào thực vật là xenlulo (chất xơ), đó là beta-1,4-D-glucan. Đây là chất hữu cơ phổ biến nhất trong sinh quyển. Các phân tử cellulose là những chuỗi không phân nhánh rất dài, chúng nằm song song với nhau thành từng nhóm vài chục và được liên kết với nhau bằng nhiều liên kết hydro. Kết quả là, những sợi nhỏ, tạo nên khung kết cấu của bức tường và quyết định độ bền của nó. Các vi sợi cellulose chỉ có thể nhìn thấy được bằng kính hiển vi điện tử, đường kính của chúng là 10-30nm và chiều dài của chúng đạt tới vài micron.
Cellulose không hòa tan và không trương nở trong nước. Nó rất trơ về mặt hóa học và không hòa tan trong dung môi hữu cơ, kiềm đậm đặc và axit loãng. Các vi sợi cellulose có tính đàn hồi và rất bền (tương tự như thép). Những đặc tính này quyết định việc sử dụng rộng rãi cellulose và các sản phẩm của nó. Sản lượng sợi bông trên thế giới, chủ yếu bao gồm cellulose, là 1,5 10 7 tấn mỗi năm. Thuốc súng không khói, lụa axetat và viscose, giấy bóng kính và giấy được lấy từ xenlulo. Phản ứng định tính cellulose được thực hiện với thuốc thử clo-kẽm-iốt, thành tế bào xenlulo chuyển sang màu xanh tím.
Ở nấm, chất nền của thành tế bào là chitin- một polysacarit được tạo ra từ dư lượng glucosamine. Chitin thậm chí còn bền hơn xenlulo.
Các vi sợi được ngâm trong vô định hình ma trận, thường là gel nhựa bão hòa nước. Chất nền là một hỗn hợp phức tạp của các polysacarit, các phân tử bao gồm dư lượng của một số loại đường khác nhau và là chuỗi ngắn hơn và phân nhánh hơn cellulose. Polysacarit ma trận xác định các đặc tính của thành tế bào như độ trương nở mạnh, tính thấm nước cao và các hợp chất trọng lượng phân tử thấp hòa tan trong đó và đặc tính trao đổi cation. Polysacarit ma trận được chia thành hai nhóm - chất pectin Và hemicellulose.
Chất pectin trương nở hoặc tan mạnh trong nước. Chúng dễ dàng bị phá hủy bởi chất kiềm và axit. Đại diện đơn giản nhất của các chất pectin là chất tan trong nước axit pectic– Sản phẩm trùng hợp của axit alpha-D-galacturonic (lên đến 100 đơn vị), được liên kết bằng liên kết 1,4 thành chuỗi tuyến tính (alpha-1,4-D-galacturonan). Axit pectic (pectin)– đây là các hợp chất polyme có trọng lượng phân tử cao hơn (100-200 đơn vị) của axit alpha-D-galacturonic, trong đó các nhóm carboxyl được methyl hóa một phần. Pectat Và nhổ lông- muối canxi và magie của axit pectic và pectic. Axit pectic, pectat và pectinate hòa tan trong nước khi có mặt đường và axit hữu cơ để tạo thành gel đậm đặc.
Thành tế bào thực vật chứa chủ yếu protopectin– polyme có trọng lượng phân tử cao của axit polygalacturonic methoxylated với arabinans và galactan; ở thực vật hai lá mầm, chuỗi galacturonan chứa một lượng nhỏ rhamnose. Protopectin không hòa tan trong nước.
Hemicellulose là các chuỗi phân nhánh được tạo thành từ dư lượng đường trung tính, phổ biến hơn là glucose, galactose, mannose, xyloza; mức độ trùng hợp 50-300. Hemicellulose ổn định hơn về mặt hóa học so với các chất pectin; chúng khó thủy phân hơn và khó trương nở hơn trong nước. Hemicellulose có thể được lắng đọng trong thành tế bào của hạt dưới dạng chất dự trữ (chà là, quả hồng). Các chất pectic và hemicellulose được kết nối với nhau bằng sự chuyển tiếp lẫn nhau. Ngoài polysacarit, một loại protein cấu trúc đặc biệt còn có trong ma trận thành tế bào. Nó liên kết với dư lượng đường arabinose và do đó là một glycoprotein.
Polysacarit ma trận làm được nhiều việc hơn là chỉ đơn giản lấp đầy khoảng trống giữa các vi sợi cellulose. Các chuỗi của chúng được sắp xếp một cách có trật tự và tạo thành nhiều liên kết với nhau và với các vi sợi, làm tăng đáng kể độ bền của thành tế bào.
Thành tế bào thực vật thường trải qua những biến đổi hóa học. sự hóa gỗ, hoặc sự hóa gỗ xảy ra khi nó được gửi vào ma trận chất gỗ- hợp chất polyme có bản chất phenolic, không tan trong nước. Thành tế bào được hóa gỗ mất đi tính đàn hồi, độ cứng và cường độ nén tăng mạnh, khả năng thấm nước giảm. Thuốc thử lignin là: 1) phloroglucinol Và axit clohydric đậm đặc hoặc axit sulfuric(các bức tường được sơn lót có màu đỏ anh đào) và 2) sulfatanilin, dưới ảnh hưởng của nó, các bức tường được sơn hóa trở thành màu vàng chanh. Sự hóa gỗ là đặc trưng của thành tế bào của mô dẫn gỗ (gỗ) và mô cơ học của mô xơ cứng.
Lấy mẫu, hoặc suberin hóa xảy ra do sự lắng đọng của các polyme kỵ nước ở bên trong thành tế bào - suberina Và sáp. Suberin là hỗn hợp este của axit béo polyme. Monome sáp là rượu béo và este sáp. Sáp dễ dàng được chiết xuất bằng dung môi hữu cơ và nhanh chóng tan chảy, tạo thành tinh thể. Suberin là một hợp chất vô định hình không tan hoặc hòa tan trong dung môi hữu cơ. Suberin và sáp tạo thành các lớp song song xen kẽ, lót toàn bộ khoang tế bào từ bên trong dưới dạng màng. Màng suberin thực tế không thấm nước và khí nên sau khi hình thành, tế bào thường chết. Suberization là đặc điểm của thành tế bào của mô tích hợp của nút chai. Thuốc thử cho thành tế bào suberized là Sudan III, màu đỏ cam.
đồng hóa Các bức tường bên ngoài của các tế bào mô biểu bì bị lộ ra. Kutin Và sáp lắng đọng thành các lớp xen kẽ trên bề mặt ngoài của thành tế bào dưới dạng màng - biểu bì. Cutin là một hợp chất polymer giống như chất béo có tính chất và tính chất hóa học tương tự như suberin. Lớp biểu bì bảo vệ cây khỏi sự bốc hơi nước quá mức từ bề mặt của cây. Bạn có thể nhuộm nó bằng thuốc thử Sudan III có màu đỏ cam.
khoáng hóa Thành tế bào xảy ra do sự lắng đọng trong ma trận của một lượng lớn khoáng chất, thường là silica (silicon oxit), ít thường xuyên hơn là oxalate và canxi cacbonat. Khoáng chất tạo nên độ cứng và độ giòn cho tường. Sự lắng đọng silic là đặc trưng của tế bào biểu bì đuôi ngựa, cói và cỏ. Độ cứng của thân và lá thu được do quá trình silic hóa có tác dụng như một chất bảo vệ chống lại ốc sên, đồng thời cũng làm giảm đáng kể vị ngon và giá trị dinh dưỡng của thực vật.
Một số tế bào chuyên biệt có chất nhầy vách tế bào. Trong trường hợp này, thay vì có thành thứ cấp xenlulô, các polysaccharid có tính axit vô định hình, có tính hydrat hóa cao được lắng đọng ở dạng nhớt Và nướu răng, có bản chất hóa học gần với chất pectin. Chất nhầy hòa tan tốt trong nước tạo thành dung dịch chất nhầy. Nướu dính và căng thành sợi. Khi khô, chúng có độ đặc như sừng. Khi chất nhầy lắng đọng, protoplast bị đẩy dần về phía trung tâm tế bào, thể tích của nó và thể tích không bào giảm dần. Cuối cùng, khoang tế bào có thể chứa đầy chất nhầy và tế bào sẽ chết. Trong một số trường hợp, chất nhầy có thể xuyên qua thành tế bào sơ cấp để nổi lên bề mặt. Bộ máy Golgi đóng vai trò chính trong việc tổng hợp và bài tiết chất nhầy.
Chất nhầy do tế bào thực vật tiết ra thực hiện nhiều chức năng khác nhau. Vì vậy, chất nhầy của chóp rễ có tác dụng như chất bôi trơn, tạo điều kiện cho chóp rễ phát triển trong đất. Các tuyến chất nhầy của cây ăn côn trùng (sundews) tiết ra chất nhầy để côn trùng dính vào. Chất nhầy tiết ra bởi các tế bào bên ngoài của vỏ hạt (lây, mộc qua, chuối) giữ hạt giống trên bề mặt đất và bảo vệ cây con khỏi bị khô. Chất nhầy được nhuộm bằng thuốc thử xanh methylen có màu xanh lam.
Việc giải phóng nướu thường xảy ra khi cây bị thương. Ví dụ, hiện tượng chảy mủ từ những vùng bị thương trên thân và cành thường thấy ở quả anh đào và mận. Keo anh đào là một loại kẹo cao su cứng. Kẹo cao su thực hiện chức năng bảo vệ, che vết thương khỏi bề mặt. Nướu được hình thành chủ yếu ở thực vật thân gỗ thuộc họ đậu (keo, tragacanth astragalus) và Rosaceae thuộc phân họ mận (anh đào, mận, mơ). Nướu và chất nhầy được sử dụng trong y học.
Thành tế bào là sản phẩm của hoạt động sống còn của protoplast. Polysaccharides ma trận, glycoprotein vách, lignin và chất nhầy được hình thành trong bộ máy Golgi. Sự tổng hợp, hình thành và định hướng cellulose của các vi sợi được thực hiện bởi plasmalemma. Vai trò chính trong việc định hướng của các vi sợi thuộc về các vi ống, nằm song song với các vi sợi lắng đọng gần plasmalemma. Nếu các vi ống bị phá hủy thì chỉ có các tế bào đẳng đường được hình thành.
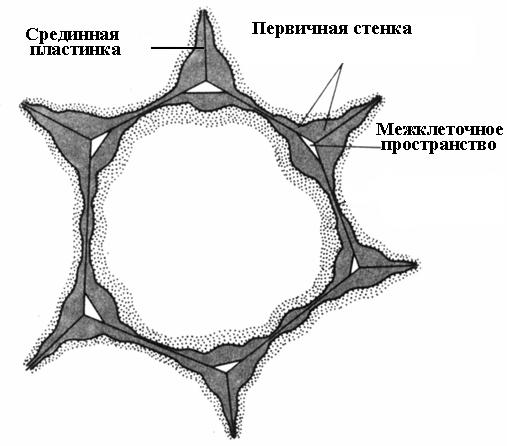
Sự hình thành thành tế bào bắt đầu trong quá trình phân chia tế bào. Trong mặt phẳng phân chia, một tấm tế bào được hình thành, một lớp chung cho hai tế bào con. Nó bao gồm các chất pectin có độ đặc bán lỏng; không có xenlulozơ. Ở tế bào trưởng thành, tấm tế bào được bảo tồn nhưng trải qua những thay đổi, đó là lý do tại sao nó được gọi là Trung bình, hoặc tấm gian bào (chất gian bào)(cơm. 2.16). Tấm giữa thường rất mỏng và gần như không thể phân biệt được.
Ngay sau khi hình thành tấm tế bào, các nguyên mẫu của tế bào con bắt đầu hình thành thành tế bào của chính chúng. Nó được lắng đọng từ bên trong cả trên bề mặt của tấm tế bào và trên bề mặt của các thành tế bào khác trước đây thuộc về tế bào mẹ. Sau khi phân chia, tế bào bước vào giai đoạn tăng trưởng kéo dài, nguyên nhân là do tế bào hấp thụ nước với cường độ thẩm thấu cao liên quan đến sự hình thành và phát triển của không bào trung tâm. Áp suất Turgor bắt đầu làm căng bức tường, nhưng nó không bị rách do các phần mới của vi sợi và chất nền liên tục được lắng đọng vào đó. Sự lắng đọng các phần vật chất mới diễn ra đồng đều trên toàn bộ bề mặt của protoplast nên độ dày của thành tế bào không giảm.
Vách ngăn tế bào đang phân chia và phát triển được gọi là sơ đẳng. Chúng chứa rất nhiều (60-90%) nước. Chất khô chủ yếu là polysaccharide nền (60-70%), hàm lượng cellulose không vượt quá 30% và không có lignin. Độ dày của thành sơ cấp rất nhỏ (0,1-0,5 micron).
Đối với nhiều tế bào, sự lắng đọng thành tế bào chấm dứt đồng thời với sự ngừng phát triển của tế bào. Những tế bào như vậy được bao quanh bởi một bức tường sơ cấp mỏng cho đến cuối đời ( cơm. 2.16).
Cơm. 2.16. Tế bào nhu mô có vách sơ cấp.
Ở các tế bào khác, quá trình lắng đọng thành vẫn tiếp tục ngay cả sau khi tế bào đạt đến kích thước cuối cùng. Trong trường hợp này, độ dày của thành tăng lên và thể tích chiếm giữ của khoang tế bào giảm đi. Quá trình này được gọi là dày lên thứ cấp bức tường, và bản thân bức tường được gọi là sơ trung(cơm. 2.17).
Bức tường thứ cấp có thể được coi là bổ sung, chủ yếu thực hiện chức năng hỗ trợ cơ học. Bức tường thứ cấp chịu trách nhiệm về các tính chất của gỗ, sợi dệt và giấy. Bức tường thứ cấp chứa ít nước hơn đáng kể so với bức tường sơ cấp; nó bị chi phối bởi các vi sợi cellulose (40-50% trọng lượng chất khô), nằm song song với nhau. Trong số các polysaccharide nền, hemicellulose (20-30%) là điển hình và có rất ít chất pectin. Thành tế bào thứ cấp thường trải qua quá trình hóa gỗ. Trong các bức tường thứ cấp không được lót gỗ (sợi lanh, lông bông), hàm lượng xenlulo có thể đạt tới 95%. Hàm lượng cao và sự định hướng có trật tự nghiêm ngặt của các vi sợi quyết định tính chất cơ học cao của các vách thứ cấp. Thông thường, các tế bào có thành tế bào thứ cấp sẽ chết sau khi quá trình dày lên thứ cấp hoàn tất.
Lớp màng ở giữa dán các tế bào lân cận lại với nhau. Nếu nó bị hòa tan, các thành tế bào sẽ mất liên lạc với nhau và tách ra. Quá trình này được gọi là sự ngâm. Quá trình ngâm tự nhiên khá phổ biến, trong đó các chất pectin ở tấm giữa được chuyển hóa thành trạng thái hòa tan nhờ enzyme pectinase rồi rửa sạch bằng nước (các loại quả quá chín như lê, dưa, đào, chuối). Người ta thường quan sát thấy hiện tượng ngâm một phần, trong đó tấm ở giữa không hòa tan trên toàn bộ bề mặt mà chỉ ở các góc của tế bào. Do áp suất trương lực, các tế bào lân cận ở những nơi này được làm tròn, dẫn đến sự hình thành khoảng gian bào(cơm. 2.16). Các khoảng gian bào tạo thành một mạng lưới phân nhánh duy nhất chứa đầy hơi nước và khí. Vì vậy, không gian giữa các tế bào cải thiện sự trao đổi khí của tế bào.
Một đặc điểm đặc trưng của tường thứ cấp là sự lắng đọng không đồng đều của nó trên đỉnh tường chính, do đó các phần không dày vẫn còn trên tường thứ cấp - lỗ chân lông. Nếu thành thứ cấp không đạt độ dày lớn thì các lỗ rỗng trông giống như những vết lõm nhỏ. Trong các tế bào có thành thứ cấp chắc chắn, các lỗ ở mặt cắt ngang trông giống như các kênh xuyên tâm kéo dài từ khoang tế bào đến thành sơ cấp. Dựa vào hình dạng của lỗ chân lông, có hai loại lỗ chân lông: đơn giản và về có lưỡi(Hình 2.17).
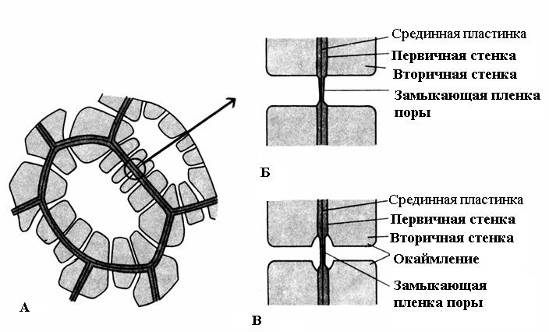
Cơm. 2.17. Các loại lỗ chân lông: A – tế bào có thành thứ cấp và nhiều lỗ đơn giản; B – một cặp lỗ chân lông đơn giản; B – cặp lỗ chân lông có viền.
bạn lỗ chân lông đơn giảnĐường kính của kênh lỗ rỗng bằng nhau dọc theo toàn bộ chiều dài của nó và có dạng hình trụ hẹp. Các lỗ đơn giản là đặc trưng của tế bào nhu mô, sợi libe và sợi gỗ.
Các lỗ ở hai ô liền kề có xu hướng xuất hiện đối diện nhau. Các lỗ thông thường này trông giống như một kênh, được ngăn cách bởi một vách ngăn mỏng của tấm giữa và vách sơ cấp. Sự kết hợp của hai lỗ trên thành liền kề của các tế bào lân cận được gọi là cặp lỗ chân lông và hoạt động như một tổng thể. Phần tường ngăn cách chúng được gọi là màng đóng lỗ chân lông, hoặc màng lỗ chân lông. Trong tế bào sống, màng đóng của lỗ chân lông được thấm nhiều chất plasmodesmata(cơm. 2.18).
Plasmodesmata chỉ có ở tế bào thực vật. Chúng là những sợi tế bào chất xuyên qua thành tế bào lân cận. Số lượng plasmodesmata trong một tế bào rất lớn - từ vài trăm đến hàng chục nghìn; thông thường plasmodesmata được tập hợp thành nhóm. Đường kính của kênh plasmodesmal là 30-60 nm. Các bức tường của nó được lót bằng plasmalemma, liên tục với plasmalemma của các tế bào lân cận. Ở trung tâm của plasmodesmata có một hình trụ màng - thanh trung tâm của plasmadesmata, liên tục với màng của các thành phần của lưới nội chất của cả hai tế bào. Giữa thanh trung tâm và màng sinh chất trong ống có hyaloplasm, liên tục với hyaloplasm của các tế bào lân cận.
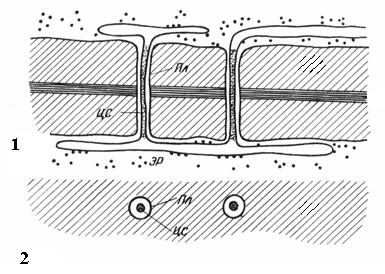
Cơm. 2.18. Plasmodesmata dưới kính hiển vi điện tử (sơ đồ): 1 – trên mặt cắt dọc; 2 – trên mặt cắt ngang; xin vui lòng- màng sinh chất; CA– thanh trung tâm của plasmodesmata; phòng cấp cứu- Thành phần của lưới nội chất.
Do đó, các nguyên sinh chất của tế bào không hoàn toàn tách biệt với nhau mà giao tiếp thông qua các kênh plasmodesmata. Chúng thực hiện sự vận chuyển giữa các ion và các phân tử nhỏ, đồng thời truyền các kích thích nội tiết tố. Thông qua plasmodesmata, các nguyên sinh chất của tế bào trong cơ thể thực vật tạo thành một tổng thể duy nhất gọi là đơn giản nhất, và việc vận chuyển các chất qua plasmodesmata được gọi là tượng trưng không giống khải huyền vận chuyển dọc theo thành tế bào và khoảng gian bào.
bạn lỗ chân lông có viền(cơm. 2.17) kênh thu hẹp mạnh trong quá trình lắng đọng của thành tế bào, do đó lỗ bên trong của lỗ mở vào khoang tế bào hẹp hơn nhiều so với lỗ bên ngoài tiếp giáp với thành sơ cấp. Các lỗ có viền là đặc điểm của các tế bào chết sớm của các bộ phận dẫn nước của gỗ. Trong đó, kênh lỗ rỗng mở rộng theo hình phễu về phía màng đóng và bức tường thứ cấp treo dưới dạng con lăn trên phần mở rộng của kênh, tạo thành khoang lỗ rỗng. Cái tên lỗ chân lông viền xuất phát từ thực tế là khi nhìn từ bề mặt, lỗ bên trong trông giống như một vòng tròn nhỏ hoặc khe hẹp, trong khi lỗ bên ngoài dường như viền lỗ bên trong dưới dạng một vòng tròn có đường kính lớn hơn hoặc rộng hơn. rạch.
Các lỗ chân lông tạo điều kiện thuận lợi cho việc vận chuyển vào
BÀI HỌC: Đặc điểm của tế bào thực vật
Tổng thành phần hóa học gần như giống nhau.
Điểm tương đồng với tế bào động vật: sự hiện diện của màng, bào tương, nhân và các bào quan cụ thể.
Tuy nhiên, giữa các loại sinh vật nhân chuẩn này có 2 điểm khác biệt về cấu trúc: sự hiện diện của các bào quan cụ thể và sự trao đổi chất.
Trong tế bào thực vật có Trung tâm không bào. Không bào của tế bào thực vật hấp thụ lượng nước dư thừa, phồng lên dẫn đến tế bào bị giãn ra, không bào chiếm phần lớn thể tích của tế bào thực vật, do đó trong tế bào thực vật chỉ còn lại những sợi tế bào chất mỏng, đi qua toàn bộ tế bào. Đây là một cách tiết kiệm để tế bào thực vật tăng kích thước của nó. Trong trường hợp này, năng lượng tiêu thụ ít hơn so với trường hợp lấp đầy cùng một thể tích bằng tế bào chất giàu protein.
Tế bào thực vật non có thể có một số không bào, khi tế bào phát triển sẽ hợp nhất với nhau và tạo thành một hoặc một số không bào lớn, chiếm tới 90% thể tích của toàn bộ tế bào. Trong trường hợp này, các bào quan được đẩy ra ngoại vi tế bào và nhân cũng vậy. Màng của không bào là tonoplast. Khoang không bào chứa đầy nhựa tế bào, là dung dịch nước của muối vô cơ, đường, axit hữu cơ và muối, protein và các hợp chất trọng lượng phân tử thấp của chúng.
Chức năng chính của không bào trung tâm- duy trì áp suất trương lực. Nó được xác định bởi các phân tử hòa tan trong nhựa tế bào của không bào để duy trì nồng độ thẩm thấu. Nồng độ này đảm bảo rằng không bào hoạt động như một cấu trúc duy trì áp suất bên trong tế bào, mang lại cho tế bào sức mạnh và sức căng. Tonoplast có bơm proton phụ thuộc ATP để đảm bảo vận chuyển đường và cũng có các kênh ion cung cấp sự bài tiết (loại bỏ) khỏi không bào của các chất chuyển hóa khác nhau. Nồng độ proton trong nhựa tế bào tương ứng với môi trường axit 2-5pH. Không bào trung tâm đảm bảo tích lũy các chất dinh dưỡng dự trữ - đường và protein. Việc cung cấp các chất này có liên quan đến khả năng tương tác của màng với tonoplast. Quá trình tiêu hóa cũng có thể diễn ra bên trong không bào nhựa tế bào, bởi vì chứa các enzyme thủy phân, trong khi tonoplast hình thành các túi tiết, chúng tách ra khỏi không bào, tương tác với các túi ngoại bào, đảm bảo quá trình tiêu hóa các thành phần có trong chúng. Họ cũng thực hiện các phản ứng của chu trình tự động, tham gia vào quá trình thủy phân chất làm đầy bị lỗi.
Vách tế bào thực vật.
Được hình thành với sự tham gia của plasmalemma. Nó là một cấu trúc đa lớp ngoại bào có chức năng bảo vệ bề mặt tế bào, về cơ bản là bộ xương bên ngoài của nó. Nó bao gồm hai cấu trúc: một khung (được làm từ các sợi fibril cellulose) và một ma trận giống như gel. Chất nền chứa các polysaccharides: hemicelluloses và các chất pectin. Hemiaellulose là chuỗi polymer phân nhánh bao gồm nhiều hexose khác nhau (glucose, manose, galactose), cũng có thể có pentose (xyloza, arabinose) và axit uronic (glucuronic và galacturonic). Các thành phần này của hemicellulose được kết hợp với nhau theo các tỷ lệ định lượng khác nhau và tạo thành các tổ hợp khác nhau. Nhưng các chuỗi phân tử hemiaellulose không bao giờ kết tinh và các sợi nhỏ của chúng không cố định (không được phát hiện). Chứa lượng nước cao.
Pectin. Một nhóm không đồng nhất bao gồm các polyme phân nhánh, được hydro hóa cao, mang điện tích âm do có nhiều gốc axit galacturonic.
Nhờ các thành phần này, ma trận là một khối nhựa mềm đóng vai trò là đế (khung).
Các polyme cellulose riêng lẻ được đóng gói thành các vi sợi bằng liên kết hydro. Cung cấp độ cứng và sức mạnh. Giữa chúng có các dây chằng biến thành tế bào thành một khối nguyên khối. Các vi sợi cellulose liên kết như vậy được bao quanh bởi pectin. Chúng có thể tương tác với canxi và silicon. Điều gì mang lại cho nó sự cứng nhắc. Do cấu trúc giống như gel, ma trận cung cấp khả năng truyền nước và phân tử nhỏ khuếch tán.
Thành tế bào được chia thành 3 loại: sơ cấp, thứ cấp, thứ ba.
Tế bào sơ cấp bao gồm 90% carbohydrate. Trong quá trình phân chia tế bào, một bó vi ống xuất hiện trong mặt phẳng xích đạo của tế bào, nằm giữa các nhiễm sắc thể phân kỳ (phragmoplast). Trong số các vi ống có nhiều túi nhỏ của hệ thống không bào, CG, ở phần trung tâm các thể lạp bắt đầu hợp nhất với nhau. Tạo thành một đĩa phẳng hoặc tấm trung gian. Bao gồm một polysaccharide gọi là callose. Đàn hồi hơn so với cellulose (không chứa bên trong bong bóng CG, hình thành trên plasmalemma). Các túi CG chứa các thành phần cần thiết để tạo nên màng của hai tế bào con. Quá trình hợp nhất của các không bào nhỏ xảy ra từ trung tâm tế bào đến ngoại vi và tiếp tục cho đến khi các túi màng hợp nhất với bề mặt của bề mặt bên của tế bào.
Thành tế bào sơ cấp đang phát triển đã bao gồm 3 lớp (tấm giữa, bao gồm chất nền vô định hình và 2 lớp ngoại vi, chứa hemicellulose và các sợi nhỏ). Thành sơ cấp được hình thành do sự tiết ra các sợi hemicellulose và cellulose bởi hai cấu trúc tế bào mới. Độ dày của thành tế bào sẽ tăng lên do hoạt động của các tế bào con, mỗi tế bào con sẽ tiết ra các chất của màng tế bào, các chất này sẽ dày lên bằng cách xếp ngày càng nhiều lớp mới. Tại màng sinh chất, các sợi cellulose được tổng hợp và trùng hợp, dẫn đến sự hình thành thành tế bào thứ cấp (tạo cho tế bào hình dạng cuối cùng). Ngay sau khi quá trình làm dày thành hoàn tất, nó được biến tính bằng lignin, một loại polyme kỵ nước, được tổng hợp thông qua phản ứng trùng hợp oxy hóa của tối đa ba dư lượng rượu thơm và đóng vai trò là thành phần chính của gỗ (20-35% gỗ là lignin). ). Do tính chất kỵ nước của nó, lignin tạo ra khả năng chống thấm cho tường và đóng vai trò như một vật liệu tăng cường bổ sung. Thành tế bào trở nên bị hóa gỗ hoặc bị suber hóa (do cutin và suberin). Ở tế bào biểu bì, sáp được tiết ra trên bề mặt màng tế bào.
Nấm có chitin (N-acetylglucosamine).
Plastid.
Tính chất chung của tất cả các plastid:
Chúng được bao quanh bởi hai màng đóng chặt dọc theo toàn bộ bề mặt của bào quan.
Nội dung bên trong là stroma. Nó chứa các đĩa màng được hình thành do sự xâm lấn và nảy chồi của các phần của màng bên trong. Những đĩa này được gọi là thylakoid và có khả năng gấp lại thành chồng (granas). Chúng tương tác với loại màng thứ tư – lamella.
Plastid có bộ gen riêng, mỗi plastid có nhiều bản sao DNA vòng chứa khoảng 100 gen. Bộ gen mã hóa các protein cần thiết để thực hiện các chức năng chuyên biệt. Các protein RNA liên quan đến phiên mã và dịch mã cũng nằm ở đây. Chúng có nguồn gốc từ plastid, nhưng hầu hết các protein khác đều phụ thuộc vào bộ gen hạt nhân. Những protein này được tổng hợp trong tế bào chất và phải được nhập vào lạp thể.
Plastid không được kết nối với các bào quan khác bằng liên kết mụn nước.
Plastid đại diện cho một loạt các biến đổi lẫn nhau.
Proplastid -> leucoplast -> lục lạp -> sắc lạp.
Bạch cầu có thể biến đổi thành: amyloplast, elaioplast, hạt aleurone và sắc lạp.
lục lạp.
Chúng có hình dạng thon dài, kích thước 2-4 micron và chiều dài có thể là 10 micron. Tế bào sắc tố (lục lạp khổng lồ).
Không gian bên trong của lục lạp là lumen. 2 hiện tượng quan trọng: quang hợp, tổng hợp năng lượng. Màng thylakoid được đặc trưng bởi sự hiện diện của chất diệp lục; nó có khả năng hấp thụ lượng tử ánh sáng mặt trời và chuyển đổi năng lượng mặt trời thành năng lượng hóa học. Sự hấp thụ ánh sáng có bước sóng cụ thể gây ra sự thay đổi chất diệp lục. Anh ta rơi vào trạng thái phấn khích. Và năng lượng giải phóng của diệp lục hoạt hóa được truyền qua một loạt các giai đoạn trung gian đến chuỗi vận chuyển điện tử, dẫn đến sự tổng hợp ATP và phục hồi chất mang là 2 mononucleotide (NAD và FAD). Năng lượng được sử dụng cho quá trình tổng hợp carbon dioxide và tổng hợp đường.
Quang hợp: pha sáng và pha tối.
Pha sáng chỉ xảy ra trong ánh sáng và gắn liền với sự hấp thụ ánh sáng của sắc tố và tiến hành phản ứng hóa học - phản ứng Hill.
Trong pha tối, carbon dioxide được cố định và khử (từ khí quyển), dẫn đến sự tổng hợp carbon, AMK và các chất khác. Kết quả của pha sáng xảy ra quá trình phosphoryl hóa, tổng hợp ADP và ATP. Cũng như sự phục hồi chất mang xảy ra trong quá trình thủy phân và quang phân nước.
Trong pha sáng của quá trình quang hợp, năng lượng của ánh sáng mặt trời kích thích các electron của diệp lục. Chúng được vận chuyển dọc theo các thành phần của chuỗi oxy hóa trong màng thylakoid, tương tự như cách các electron được vận chuyển dọc theo chuỗi hô hấp ở ty thể. Ở lục lạp, proton được bơm từ chất nền (pH = 8) vào lòng (pH = 5) tạo ra một gradient 3 đơn vị pH. Độ dốc proton này tạo ra một động lực proton có công suất 200 mW ở màng proton, nhưng điều này gần như hoàn toàn là do độ dốc pH. So với chuỗi hô hấp của ty thể, các electron di chuyển theo một hướng khác. Các electron được lấy từ nước trong quá trình quang phân và chuyển sang chất mang với mức tiêu hao năng lượng. Cái đó. ở pha sáng chúng ta có: các phân tử ATP được tổng hợp và chất vận chuyển khử. Cả hai sản phẩm đều được sử dụng trong giai đoạn tối.
Trong giai đoạn tối, carbohydrate được hình thành do NAD và ATP bị khử. Quá trình này là nhiều giai đoạn. Một số lượng lớn các enzyme có liên quan đến nó. Chu trình calvin. Sự gia tăng số lượng lục lạp và sự hình thành các dạng lạp thể khác xảy ra từ các cấu trúc tiền thân (proplastid).
Proplastids là những túi màng đôi nhỏ không có đặc điểm gì đặc biệt. Chúng khác với không bào tế bào chất ở thành phần đậm đặc hơn và sự hiện diện của hai màng. Proplastids nằm trong việc phân chia các mô thực vật. Số lượng tiền lạp thể tăng lên thông qua quá trình phân chia và nảy chồi. Trong điều kiện chiếu sáng bình thường, các proplastid biến thành lục lạp (tăng kích thước, hình thành các nếp gấp màng dọc. Một số tạo thành lamellae, một số khác - thylakoid). Nếu chúng phát triển trong bóng tối, thì lúc đầu thể tích plastid tăng lên, nhưng hệ thống màng bên trong không được hình thành, một khối túi nhỏ được hình thành, chúng tích tụ thành các vùng riêng biệt, tạo thành các cấu trúc mạng tinh thể phức tạp - các thể prolamellar (etioplast). Màng của chúng chứa protochlorophyll (màu vàng), dưới tác dụng của ánh sáng, lục lạp được hình thành từ chúng và protochlorophyll có thể biến thành diệp lục thực sự (màng, thành phần của chuỗi chuyển được tổng hợp).
Bạch cầu. Khác với lục lạp. Chúng được tìm thấy trong các tế bào mô dự trữ. Hình thái không chắc chắn của chúng khiến chúng khó phân biệt với các tiền lạp thể. Chúng có thể nhìn thấy dưới dạng phiến mỏng nhưng có khả năng hình thành các cấu trúc thylakoid bình thường. Trong bóng tối, chúng tích tụ nhiều chất dự trữ khác nhau. Nếu là carbohydrate thì bạch cầu được hình thành thành amyloplast (cũng có thể có protein ở dạng hạt aleurone). Và nếu chất béo là elaioplast.
Chúng tôi quan sát quá trình đổi màu và thay đổi lục lạp trong quá trình phát triển của cánh hoa và quá trình chín của quả. Các màng bổ sung tích tụ trong plastid, chất diệp lục và tinh bột bị phá hủy. Khi các phiến mỏng bị phá hủy, các giọt lipid được giải phóng, trong đó các sắc tố (carotenoid) hòa tan tốt và xuất hiện sự thay đổi màu sắc. Sắc lạp là một dạng plastid thoái hóa, có liên quan đến sự phân hủy các phức hợp lipoprotein.
Bộ gen plastid.
Hệ thống di truyền riêng của nó đảm bảo quá trình tổng hợp các protein được định vị bên trong plastid (protein màng thylakoid). Chúng có chiều dài lên tới 60 micron. 1,3*10 8 Có. Thời lượng chu kỳ và tốc độ sao chép không khớp. DNA không có cấu trúc cố định. Tất cả các đặc điểm của phân tử vòng của lục lạp đều gần giống với ở sinh vật nhân sơ. Sự giống nhau của DNA lục lạp được thể hiện ở chỗ các quá trình điều hòa cơ bản của phiên mã đều giống nhau. Tổng hợp tất cả các loại RNA. Mã DNA lục lạp cho RNA ribosome. Hằng số phân đoạn ribosome 70S. Chúng chứa 17S và 23S. Nhạy cảm với kháng sinh (chloramphenicol).
BÀI HỌC: Tiếp xúc giữa các tế bào
Lý thuyết cộng sinh.
Komintsin và Mereshkovsky.
Lý thuyết này được hỗ trợ bởi sự giống nhau đáng kinh ngạc trong cấu trúc của lục lạp và tảo xanh lam. Sự tương đồng về chức năng chính của chúng, khả năng gần như giống hệt nhau trong quá trình quang hợp. Là kết quả của sự cộng sinh của các tế bào nhân sơ khác nhau, các tế bào nhân (nhân chuẩn) đã xuất hiện. Theo giả thuyết này, một tế bào nhân chuẩn trong quá trình phát triển của nó đã trải qua nhiều giai đoạn cộng sinh với các tế bào khác.
Trong giai đoạn đầu tiên, các tế bào như sinh vật nhân sơ dị dưỡng kỵ khí trở thành vật chủ cho vi khuẩn hiếu khí, dẫn đến sự hình thành các sinh vật nhân sơ hiếu khí (ty thể, trong các sinh vật nhân sơ này). Song song với quá trình này, trong tế bào chủ, bộ máy nhân nhân sơ (genophore) được hình thành thành nhân cách ly với tế bào chất. Đây là cách các tế bào nhân chuẩn dị dưỡng có thể phát sinh. Một tế bào như vậy sẽ tiến hành cộng sinh lặp đi lặp lại. Ví dụ, với xoắn khuẩn hoặc vi khuẩn giống roi. Sự bao gồm các sinh vật cộng sinh này dẫn đến sự xuất hiện của roi và lông mao trong các tế bào nhân chuẩn dị dưỡng. Và là kết quả của sự hấp thụ bổ sung. Điều này dẫn đến sự xuất hiện của các tế bào động vật nhân chuẩn có tiên mao. Một sinh vật như vậy cũng có thể hấp thụ tảo xanh lam và điều này dẫn đến sự hình thành các plastid và hình thành tế bào thực vật. Giả thuyết về nguồn gốc nội cộng sinh của các bào quan trong màng là không thể nghi ngờ. Có rất nhiều sự thật đã biết về quá trình nội cộng sinh thực sự của tảo xanh lục và tế bào của thực vật bậc thấp và động vật nguyên sinh, nơi các sinh vật cộng sinh hoạt động hoàn hảo và cung cấp cho tế bào chủ các sản phẩm quang hợp. Ví dụ. Một loại ciliate dép, nó có chứa tảo chlorella. Lục lạp có thể được một số tế bào chọn lọc và sử dụng làm chất nội cộng sinh. Luân trùng ăn tảo. Lục lạp tìm thấy chính mình bên trong các tế bào của tuyến tiêu hóa và tiếp tục thực hiện các chức năng của chúng.
Đặc điểm chức năng của tế bào thực vật. Phương pháp tổng hợp các chất hữu cơ là dinh dưỡng tự dưỡng.
Tế bào thực vật do có thành tế bào nên không cho tế bào thực vật thay đổi hình dạng và di chuyển. Và tế bào thực vật bù đắp cho sự thiếu vận động bằng cách tổng hợp thức ăn của chính chúng, tức là. là sinh vật tự dưỡng. Thực vật phát triển trong suốt cuộc đời, mô phân sinh không ngừng phân chia và hình thành tế bào mới, tất cả thực vật đều thuộc về sinh vật mô đun. Hình thức sống cuối cùng của họ phụ thuộc vào môi trường của họ. Tế bào thực vật cũng có những đặc điểm trong quá trình nguyên phân:
Vị trí của thành tế bào mới. Nó được xác định từ rất lâu trước khi hình thành trục chính, trong khi bản thân trục chính không tham gia vào vị trí của ô này. Ngay cả trước khi nguyên phân, trong pha G 2 của chu kỳ tế bào, một cấu trúc cụ thể đã được hình thành trong tế bào thực vật, được gọi là vòng tiền kỳ. Nó được hình thành từ các vi ống của vỏ não (tế bào) ở dạng dải rộng bao quanh tế bào. Hạt nhân vẫn còn nguyên (rắn). Trong khu vực của vòng này, các sợi Actin tập hợp lại, đảm bảo nó thu hẹp lại cho đến khi biến thành một bó vi ống dày đặc hình thành xung quanh nhân. Cấu trúc thu được giống như một bánh xe, trong đó vành và các nan hoa được cấu tạo từ các vi ống và Actin, còn lõi tạo thành trục. Cấu trúc của vòng được làm phong phú thêm với các phần tử EPR và KG. Ở giai đoạn đầu của quá trình nguyên phân, các vi ống của vòng tiền kỳ bị khử polyme và bị phá hủy. Và trong kỳ đầu tiên, một thành tế bào mới được hình thành chính xác trên mặt phẳng được chỉ định bởi vòng tiền kỳ đầu.
Nhân trong tế bào thực vật di chuyển đến trung tâm của tế bào, nơi sẽ xảy ra sự phân chia. Các lớp (sợi) tế bào chất hẹp kéo dài từ nhân theo mọi hướng. Khi quá trình nguyên phân xảy ra, các lớp hợp nhất với nhau, tạo thành một mặt phẳng nơi từng có vòng tiền tiên tri và mặt phẳng này được gọi là phragmosome.
Ở thực vật, các cực của trục phân bào không chứa các trung thể và có bản chất khuếch tán hơn. Nói cách khác, chúng dường như bị mờ đi. Ban đầu, trục chính tiên tri được hình thành từ các vi ống. Từ đó, sau khi màng nhân bị phá hủy, một trục chính thực sự sẽ được hình thành và với sự hình thành của trục phân bào thực sự này, không còn vi ống tế bào chất nào còn lại trong tế bào. Cái gì điều khiển trục quay này, cái gì được hình thành? Nhiễm sắc thể. Đầu tiên, quá trình trùng hợp các vi ống xảy ra gần các nhiễm sắc thể, được định hướng ngẫu nhiên. Sau đó, các protein vận động (dynein) sắp xếp các vi ống, đầu dương của chúng được gửi đến nhiễm sắc thể, cho phép chúng gắn vào kinetochores của nhiễm sắc thể và các đầu khác tương tác với đầu âm của vi ống. Là kết quả của quá trình trùng hợp, phân loại và gắn kết, một trục chính lưỡng cực được hình thành. Sau khi phân chia hạt nhân, sự phân chia tế bào chất xảy ra. Điều này trùng hợp với thời điểm hình thành vách ngăn tế bào mới, vị trí của vách ngăn này được xác định bởi vòng tiền kì đầu. Mối quan hệ giữa hướng trục chính và vách ngăn mới không chặt chẽ như ở tế bào động vật.
Phức hợp Golgi của tế bào thực vật. Chức năng: glycosyl hóa (sơ cấp), phân loại (tách), tổng hợp các polysaccharide thành tế bào (trừ cellulose và callose), vận chuyển các phức hợp enzyme (hoa hồng lục giác, tổng hợp cellulose) đến màng, vận chuyển trực tiếp các túi vào không bào.
KG được định vị trong các tế bào thực vật gần với huyết tương hơn, xa nhân hơn và được phân tán dưới dạng nhiều chồng bể. Tất cả các yếu tố của phức hợp được kết nối bằng các sợi Actin, tạo thành một mạng lưới đảm bảo vận chuyển nội bào. Cấu trúc Actin cung cấp khả năng di chuyển (cùng với myosin). Tế bào thực vật có một loại myosin đặc biệt - myosin 8. Myosin 8 này không hình thành sợi và tích tụ trong thành tế bào sơ cấp mới hình thành, túi CG chứa polysaccharides và protein cấu trúc tạo thành thành tế bào thứ cấp.
Liên hệ giữa các tế bào.
Chức năng tiếp xúc của màng đảm bảo sự tiếp xúc của các tế bào với nhau và với các tế bào khác. Tất cả các tế bào trong mô đều tiếp xúc với chất nền ngoại bào. Ma trận ngoại bào tham gia vào việc duy trì tính toàn vẹn của mô và tạo thành một khuôn khổ có trật tự trong đó các tế bào di chuyển và tương tác với nhau. Sự tương tác của các tế bào được thực hiện do các vùng tiếp xúc, được gọi là tiếp xúc giữa các tế bào.
Có một số cách phân loại trong số chúng, chúng được cấu trúc khác nhau. Phân loại của Alberts và Gray (1986). Theo đó: địa chỉ liên lạc có thể được chia thành 3 nhóm.
Cơ khí (= Chất kết dính). Liên quan đến khả năng của các tế bào dính vào nhau, do sự hiện diện của glycoprotein ma trận ngoại bào trong màng tế bào. Khi sự bám dính xảy ra giữa các màng sinh chất, luôn tồn tại một khoảng trống (20nm) chứa đầy glycocalyx. Các protein CAM đặc biệt (cam) chịu trách nhiệm trực tiếp kết nối các tế bào với nhau: fibronectin, cadherins, selectins, integrins và các protein khác. Một số trong số chúng kết nối các tế bào với nhau thông qua tương tác giữa các phân tử. Những người khác hình thành các kết nối giữa các tế bào đặc biệt. Sự tương tác giữa các protein này có thể là đồng tính, tức là trong trường hợp này các tế bào lân cận giao tiếp với nhau bằng các phân tử đồng nhất. Hoặc dị thể, khi nhiều loại protein trên các tế bào lân cận tham gia vào quá trình bám dính. Ngoài ra còn có những hợp chất phức tạp hơn khi liên kết giữa các tế bào được đảm bảo bởi các phân tử trung gian liên kết. Ngoài các kết nối đơn giản như vậy, còn có một số cấu trúc đặc biệt thực hiện các chức năng cụ thể (desmosome). Chúng tương tác với các yếu tố của tế bào học. Có 3 loại desmosome: có dấu lấm chấm, bao quanh và hemidesmosome.
Các dấu chấm kết nối các ô với nhau dưới dạng nút. Khoảng cách giữa các màng tế bào tiếp xúc thay đổi từ 22 đến 35 nm. Giữa các tế bào, do sự biến đổi của phức hợp siêu màng, một ma trận sợi được hình thành và ở phần trung tâm của nó có một “tấm” bao gồm các hạt protein. Chúng được thể hiện bằng cách tương tác với các cadderin và desmoglein của protein màng tích hợp. Tấm này tương tác với màng tế bào thông qua hệ thống các sợi ngang. Về phía tế bào chất, một tấm thứ hai được hình thành từ protein desmoplakin, từ đó các cấu trúc sợi liên quan đến các sợi 10 nm (sợi keratin) kéo dài theo hướng ngang vào sâu. Do đó, desmosome có dấu lấm chấm được tìm thấy trong biểu mô, cơ tim và mạch nội mô.
Desmosome dạng thắt lưng. Khoảng cách giữa các ô lân cận là 15-20 nm. Tấm trung tâm không quá sáng. Nhưng có những glycoprotein xuyên màng bám dính vào nhau và tạo ra sự kết nối cơ học của màng. Các sợi Actin mỏng (6-7 nm) tích tụ ở phía tế bào chất. Chúng nằm dọc theo plasmalemma và chạy dọc theo toàn bộ chu vi của tế bào. Chức năng: kết dính cơ học của các tế bào với nhau; khi các sợi Actin co lại, hình dạng của tế bào có thể thay đổi; có thể gây ra những thay đổi về hình dạng tế bào; truyền áp lực cơ học đến tế bào. Do sự thay đổi thể tích tế bào.
Hemidesmosome. Sự kết nối của tế bào với ma trận ngoại bào (Ví dụ với màng đáy). Vai trò chức năng là cơ học, cho phép các lớp biểu mô chịu được các tác động cơ học lớn. Desmosome là một phần của tế bào, nhưng chúng không phải là cấu trúc tĩnh. Họ rất năng động. Có khả năng biến mất và xuất hiện trở lại. Chúng bị phá hủy bởi phản ứng thực bào.
Đóng (tiếp xúc chặt chẽ). Chúng là đặc trưng của biểu mô một lớp. Vùng mà các lớp bên ngoài của hai màng sinh chất càng gần nhau càng tốt. Do đó, khi nhìn qua kính hiển vi điện tử, có thể nhìn thấy cấu trúc ba lớp. Hai lớp ngoài của cả hai màng hợp nhất thành một lớp dày 2-3 nm. Không phải trên toàn bộ khu vực tiếp xúc chặt chẽ mà thể hiện một loạt điểm tiếp xúc của các protein màng nguyên khối. Hơn 24 loại protein đã được tìm thấy ở vùng tiếp giáp chặt chẽ. Nhô ra khỏi màng của lớp bilipid, các protein như vậy giao nhau và tạo thành một mạng lưới (mạng lưới). Về phía liên kết chặt chẽ, trong tế bào chất có rất nhiều sợi nhỏ (7nm), nằm song song với bề mặt của plasmalemma. Các mối nối chặt chẽ là đặc điểm của biểu mô tuyến và ruột. Chúng kết nối các tế bào với nhau. Ngoài ra, đây là những cấu trúc phân tử điều chỉnh sự vận chuyển của tế bào, tức là vận chuyển các chất trong không gian giữa các tế bào. Rào cản vật lý đối với việc vận chuyển ion có bản chất khác nhau. Các ion truyền qua ngay lập tức, nhưng các thành phần khác mất nhiều thời gian hơn (thậm chí hàng giờ) để đi qua phần tiếp xúc chặt chẽ.
Rào cản thấm trong một tiếp xúc chặt chẽ được tạo ra bởi các lớp mang điện tích tạo thành cấu trúc mạng gồm các phần tử giống như sợi fibrillar. Các ion có thể đi qua các lỗ như vậy, kích thước của chúng nhỏ. Tuy nhiên, để các thành phần hòa tan đi qua, tính toàn vẹn của các sợi này phải bị phá vỡ. Các nguyên tố hòa tan phá vỡ các sợi này, sau đó chúng được tập hợp lại sau khi các phân tử đi qua. Và do đó phân tử di chuyển qua hàng rào tiếp xúc. Về mặt cấu trúc và chức năng, các mối nối chặt chẽ chia plasmalemma thành 2 phần (đỉnh và đáy) và mối nối chặt chẽ hoạt động như một vách ngăn, do đó thành phần phân tử được duy trì trong giới hạn.
Địa chỉ liên lạc dẫn điện. Nó khác nhau ở động vật và thực vật.
Có 2 loại tế bào động vật: khoảng cách và tổng hợp hóa học. Thông qua các mối nối khoảng cách, các phân tử có thể di chuyển từ tế bào này sang tế bào khác. Nhưng trong quá trình tổng hợp hóa học, tế bào không có sự kết nối trực tiếp. Các mối nối khoảng cách được coi là các mối nối giao tiếp của các tế bào. Đây là một cấu trúc có liên quan đến việc chuyển hóa chất trực tiếp. Sự tiếp xúc đặc trưng của loại này là sự gắn kết các màng sinh chất của hai tế bào lân cận lại với nhau ở khoảng cách 2-3 nm. Bản thân khe hở tiếp xúc có kích thước 0,5-5 µm. Nó bao gồm 2 nửa, kích thước mỗi nửa là 7-8 nm, chúng nằm xung quanh một kênh rộng 1-2 nm. Và 2 nửa này được gọi là connexon (nửa kênh). Chúng bao gồm 6 tiểu đơn vị protein connectin. Trọng lượng phân tử của protein là khoảng 30 nghìn. Bằng cách kết hợp với nhau, các protein tạo thành một cấu trúc hình trụ và trong hai tế bào, kích thước, số lượng, kích thước và cách sắp xếp các kết nối hoàn toàn đối xứng. Các tiếp điểm khe hở cho phép khuếch tán tự do các phân tử nặng 1200 Dalton, ngoại trừ các phân tử có khối lượng đạt tới 2000 Dalton. Tính thấm của các mối nối khe hở được kiểm soát bằng cách mở và đóng các cổng kênh. Quá trình này được gọi là cổng (cơ chế cổng). Nó được kiểm soát bởi những thay đổi về độ pH nội bào, dòng ion canxi hoặc sự phosphoryl hóa trực tiếp của các tiểu đơn vị protein connexin. Chức năng của các mối nối khe hở: dẫn truyền các chất hữu cơ và vô cơ. (Ví dụ, trong tế bào cơ tim).
Plasmodesmata là một loại tiếp xúc tiến hành ở thực vật. Đây là những kênh tế bào chất hình ống mỏng nối 2 tế bào lân cận. Đường kính của các kênh này thay đổi từ 20 đến 40 nm. Điểm đặc biệt của các kênh này là màng sinh chất của một tế bào đi vào màng sinh chất của tế bào khác. Dọc theo trục của kênh này, một ống hình trụ của mô giải thể trải dài từ tế bào này sang tế bào khác. Lumen của nó giao tiếp với các tế bào ER. Khoảng trống giữa desmotubule và màng plasmodesmata được lấp đầy bằng cytosol. Plasmalemma bao quanh các tế bào này dễ dàng đi vào plasmodesmata và màng của các tế bào khác. Kết quả là hyaloplasm của các tế bào lân cận được kết nối thành một hệ thống duy nhất (symplast). Và về mặt hình thức không có sự tách biệt (ô này với ô khác). Vận chuyển tự do qua plasmodesmata bị giới hạn ở trọng lượng phân tử 800 Dalton. Plasmodesmata được hình thành ở giai đoạn hình thành chính của tấm trung vị từ các phần tử của ER. Trong một tế bào mới được phân chia, số lượng plasmodesmata lên tới 1000 mỗi tế bào. Với sự lão hóa, số lượng của chúng giảm đi, nhưng độ dày của chúng tăng lên. Vai trò chức năng: đảm bảo sự lưu thông giữa các tế bào của dung dịch (ion, đường, chất dinh dưỡng). Mặt tiêu cực là dễ mắc bệnh hơn.