Hücre duvarı. Farklı elektron yoğunluklarına sahip 9...10 katmandan oluşan çok katmanlı bir kabuktur. Amorf bir matris içine gömülü bir mikrofibril sistemi hücre iskeletini oluşturur. Fibriller türlerine göre selüloz, glikon ve kitinden oluşabilir. Diğer polisakkaritler, proteinler, pigmentler, lipitler, hücre duvarının mikrofibriler kısmı ile kimyasal bağlar oluşturan bağlayıcı maddeler olarak görev yapar. Bu tür komplekslerin varlığı, bazı maddeler için seçici geçirgenlik sağlarken diğerlerinin blokajını sağlar.
Hücre duvarının destekleyici mikrofibrilleri ve matrisi, oluşum ve biyosentez mekanizması bakımından farklılık gösterir. Liflerin ve matrisin oluşumu eş zamanlı olarak gerçekleşmez; öncelikle duvarın fibriller iskeleti yenilenir. Hücre duvarının bu iki bölümünün biyosentezi enzimlerin katılımıyla gerçekleştirilir.
Hücre duvarı oluşumu süreci iki şekilde gerçekleşir: yeni malzeme ya duvara polarize bir şekilde sokulabilir ya da tüm yüzeyine eşit şekilde uygulanabilir. İlk durumda, ikinci küresel olarak silindirik hücreler oluşur.
Hücre duvarı koruyucu bir cihaz görevi görür ve mantar hücresini çeşitli çevresel faktörlerin, örneğin çeşitli maddelere karşı seçici geçirgenliği belirleyen bir ozmotik bariyerin etkilerinden korur. Hiflerin ve üreme organlarının bitkisel hücrelerine şekil verir. Hücre duvarı ve sitoplazmik membranın yüzeyinde, hücre tarafından asimile edilmeyen (suda çözünmeyen) polimerlerin dönüşümünü gerçekleştiren enzimler lokalizedir.
Lizis sonucunda mantarların hücre duvarı, diğer hücreler tarafından salgılanan ve mantarın kendi hücresinde oluşan enzimlerin etkisi altında tahrip edilebilir.
Mantar hücre duvarının ana bileşenleri kitin, glukanlar, protein ve yağlardır. Azotlu ve azotsuz polisakkaritler, yağlı maddelerle çözünür ve çözünmez kompleksler oluşturur. Hücre duvarı, oranı farklı mantarlar arasında biraz değişen 4...6 monosakaritten oluşur. Polisakkarit fraksiyonlarının bileşimi glukozamin, mannoz, glikoz, ksiloz vb. içerir. Aynı mantarın farklı hücrelerinin hücre zarı bileşiminin aynı olmadığı vurgulanmalıdır.
Protoplast- hücre duvarı içine alınmış hücre içeriği: Sitoplazmik bir zar, endoplazmik retikulum, nükleoluslu bir veya daha fazla çekirdek, ayrıca mitokondri, RNA'lı ribozomlar, lizozomlar, Golgi aparatı, vakuoller, lameller kompleks, salgı granülleri ve ayrıca diğer yapılar ve çeşitli kapanımlar.
Sitoplazmik membran.İnce üç katmanlı bir zar doğrudan hücre duvarının altında bulunur ve onu sitoplazmadan ayırır. Sitoplazmik membran, hücreye giren ve çıkan maddelere karşı seçici olarak geçirgendir. Sitoplazmik membran %40'a kadar lipit ve %38'e kadar protein içerir. Sitoplazmik membranın çeşitli istila ve ihlal biçimlerine mezozom adı verilir.
Sitoplazmik zarın temel işlevsel amacı şu şekildedir: çeşitli maddelerin hücreye girişi, enzimatik işlem ve metabolik ürünlerin salınması. Sitoplazmik membranda işlenen maddeler hücrenin protoplastına girer ve metabolizmaya katılır.
Endoplazmik retikulum. Bir çeşit besin deposu görevi gören kesecikler, tübüller ve vakuollerden oluşur.
Mitokondri. Bir veya iki katmanlı bir kabukla kaplanmış, bölmeli, elipsoidal şekilli çok sayıda hareketli kapalı oluşum. Mitokondrilerin kendilerine ait DNA halka yapıları nedeniyle üreme yeteneğine sahip oldukları varsayılmaktadır. Mitokondri, üzerinde enzimlerin lokalize olduğu bir zar ile çevrilidir: piruvat oksidaz, süksindehidrojenaz, alkalin ve asit fosfatazlar, peroksidaz, vb. Mitokondri, hücrede enerji jeneratörü görevi görür. Yetiştirme koşullarına ve hücrenin fizyolojik durumuna bağlı olarak mitokondrinin şekli ve hücredeki sayısı değişir.
Ribozomlar. Hücresel proteinlerin sentezinde, boyutu 200'e kadar olan yuvarlak ribonükleoprotein taneleri yer alır. Ribozom sayısı, farklı mantar türleri arasında önemli ölçüde değişiklik gösterir ve dış faktörlere, kültürün yaşına vb. bağlıdır.
Golgi aygıtı.Çok küçük çaplı (0,000 002...O,OOO 01 mikron) bir grup kabarcık veya paralel disk şeklindeki plakalarla temsil edilir. Bu organel hücrede ribozomların bulunmadığı bir bölgede bulunur.
Lizozomlar. Golgi aygıtının türevleri hücre zarı ile sitoplazmik zar arasında bulunur. Tek katmanlı bir lipoprotein membranla çevrelenmiş granüler oluşumlardır. Proteini hidrolize eden ve hücreleri ekzo ve endojen kökenli toksik maddelerin olumsuz etkilerinden koruma işlevini yerine getiren bir enzim içerirler.
Lipozomlar. Tek katmanlı zarlarla çevrelenmiş yağlı madde damlacıkları.
Çekirdek. Hücrenin merkezinde veya kutuplarında bulunur. Mantar hücreleri tek veya birden fazla çekirdeğe sahip olabilir. Kalıtsal işlevlerden sorumludurlar. Çekirdeklerin şekli yuvarlak veya uzundur. Her çekirdek, yoğun tanecikler ve ince fibrillerden oluşan bir nükleolus içeren iki katmanlı gözenekli bir nükleomembranla çevrilidir. Nükleoller, kromozomların bir parçası olarak DNA içerir. Anastomozlar yoluyla çekirdekler bir hücreden diğerine geçebilir.
Kapsamalar. Mantar hücreleri çok sayıda kapanım içerir: volutin, glikojen, lipitler, pigmentler, miyeloid oluşumlar, organik asit tuzları, amino asitler vb. Glikojenin endojen solunumdan sorumlu olduğuna ve volutinin enerji süreçlerinde yer alan bir yedek besin maddesi olarak görev yaptığına inanılmaktadır.
Yaşam süreci boyunca mantar hücrelerinde çeşitli metabolik ürünlerin - antibiyotikler, enzimler, toksinler, vitaminler vb. - biriktiğine dikkat edilmelidir.
Mikroskobik mantarların sayısız morfolojik elementlerinin tümü iki gruba ayrılır: miselyum ve sporlar. Çeşitli şekil ve boyutlarda gelirler. Sporlar ve miselyum arasındaki morfolojik fark, mantarın tipini belirlerken önemli bir ayırıcı özellik olarak hizmet eder.
Miselyum.Çapı mikromisetler arasında bir ila birkaç mikron arasında değişen dar, yuvarlak bir tüptür.
Bol dallanma ile birbirleriyle temas halinde olan misel hifleri, hücreler - anastomozlar arasında füzyonlar oluşturabilir. Çok sayıda varsa miselyum karakteristik bir ağ görünümü kazanır. Çok hücreli miselyumlu çeşitli mantarlarda anastomoz gelişimi gözlenir. Onlar sayesinde hücre çekirdeğinin bir hücreden diğerine taşınması ve haploidden diploid miselyuma geçiş mümkün olmaktadır. Ancak çoğu durumda vejetatif fonksiyonlarını yerine getirirler ve beslenme eksikliğinden dolayı birçok formda gelişirler. Misel hücrelerinin uzunluğu birkaç mikrondan onlarca ve daha az sıklıkla yüzlerce mikrona kadar değişir.
Miselyum, genç kültürlerde daha hassas olan çift devreli bir kabukla çevrilidir. Miselyumu tek tek hücrelere bölen bölmelerde, büyüme sürecinde sitoplazmanın ve onunla birlikte besinlerin aktığı gözenekler vardır. Hücrelerde birçok farklı kapanım vardır: eskilerde, birçok vakuol nedeniyle sitoplazma granüler hale gelir. Genç miselyum uzun dikdörtgen hücrelerden, eskilerden - kısa yuvarlak veya çok yönlü olanlardan oluşur. Bölümleri olan miselyuma septat denir. Bununla birlikte, bazı alt mantarlarda miselyum, enine bölmelerden yoksun hiflerden oluşur ve çok sayıda çekirdeğe sahip, oldukça dallanmış dev bir hücreye benzer ve bölmeli olmayan miselyum olarak adlandırılır.
Miselyum nasıl gelişir? Spordan bir mikrop tüpü çıkar; bu tüp uzar ve daha sonra sporu içeren orta kısımdan bir septum ile ayrılır. Büyüme tüpleri daha sonra daha da uzar ve distal veya apikal hücre ve proksimal veya iç hücreye bölünen yeni bir septum alır. Daha sonra apikal hücre uzar ve tekrar bölünerek birinciden daha genç ikinci bir iç hücreyi ayırır. Bu olmaya devam ediyor. Bu süreçte iç hücreler yalnızca uzar, enine bölünmeleri nadiren meydana gelir, ancak onlardan yan dallar gelişir. İç hücrenin uzak ucunda, silindirik bir şekil alan ve daha sonra onu üreten hücreden bir septum ile ayrılan yanal bir çıkıntı oluşur. Yeni hücre daha sonra ana hücreyle aynı şekilde büyüyüp dallanan bir yan dal haline gelir. Ana hipha boyunca dalların gelişmesi nedeniyle, daha yaşlı ve daha gelişmişlerdir, dalları tabana ne kadar yakınsa - akropetal dallanma.
Bölmeli olmayan miselyumun gelişimi genel olarak aynı şekilde gerçekleşir, ancak enine bölümlerin oluşumu olmadan. Büyüme, bol miktarda protoplazmanın biriktiği, tüm lümeni doldurduğu hifanın uçlarında meydana gelir ve daha arka kısımlarda merkezi vakuollerin önemli bir gelişimi vardır. Homojen bir ortamda, örneğin besleyici jelatinin yüzeyinde, miselyumun hifleri (hem hücresiz hem de çok hücreli) eşit ve radyal olarak büyür, böylece miselyum kenarlardan büyüyen bir daire şekline sahiptir. Orta kısmı en yaşlı, hatta bazen ölü, çevre kısmı ise en genç olanıdır.
Tipik olarak adlandırılabilecek misel gelişiminin genel tekdüzeliğine rağmen, bazı durumlarda hem makroskobik görünümde hem genel büyüme düzeninde hem de mikroskobik yapıda bir takım spesifik özellikler gözlenir. Miselyumun makroskobik görünümü öncelikle hava hifleri tarafından belirlenir. Bazı durumlarda, alt tabakanın tam yüzeyinde ve kısmen içinde oluşurlar ve daha sonra miselyum, alt tabakaya bastırılmış düz bir daire görünümüne sahiptir; diğer durumlarda, ek olarak az çok miktarda hif gelişir, havaya yükselir ve miselyuma, örneğin alt tabakanın üzerinde yükselen bir pamuk yünü parçasına bir miktar benzerlik verir. Aynı mantarın büyüme şekli, neme, beslenmeye vb. bağlı olarak farklı olabilir. Bununla birlikte, bazı mantar türlerinin belirli özellikleri vardır; örneğin, bir odun yok edici olan yemyeşil hava miselyumunun oluşumu.
Miselyumun rengi çoğunlukla kar beyazıdır, ancak yaşlandıkça farklı tonlarda kahverengiye döner. Bunun nedeni hücre duvarlarında ve daha az sıklıkla hücrenin içinde pigment birikmesidir.
Gerçek miselyum ve psödomiselyum vardır. İkincisi, bireysel hücrelerin birbirine bağlı olmaması ve ortak bir zara sahip olmaması ile karakterize edilir. Burada gerçek dallanma yerine ağaç benzeri bir hücre dizilişi gözlenir.
Bazı mantarlar, evrim sırasında alt tabakaya bağlanmak ve ondan besinleri çıkarmak için bunun için özel olarak tasarlanmış organlar oluşturmuştur: mantarları tanımlarken dikkate alınan rizoidler ve apressoria. Rizoidler kök şeklindedir ve apressorialar kısa, genişlemiş, bazen miselyumun lob şeklinde çıkıntılarıdır.
Sklerotia, şeritler, rizomorflar ve klamidyasporlar da miselyal büyümenin modifikasyonlarıdır.
Sclerotia, özel cisimler oluşturan bölmeli mantar hiphalarıdır. Sklerotia oluştuğunda hifal membranlar kalınlaşır ve rengi koyulaşır. Sklerotyumun dış katmanındaki hifanın duvarı büyük ölçüde kalınlaşmıştır, ancak içerideki hifler daha ince duvarlıdır ve genellikle renkli değildir. Sclerotia, mantarın çevrede uzun süre hayatta kalmasına izin veren ve çeşitli dış etkenlere karşı direncini sağlayan koruyucu adaptif gövdelerdir: sıcaklık, güneş ışığı vb. Olgun sklerotia, miselyum ve birçok rezerv maddesi - lipitler, glikojen ile karşılaştırıldığında daha az nem içerir.
Sklerotia'nın boyutları birkaç milimetreden birkaç on santimetreye kadar değişir ve şekli çok çeşitli olabilir: küresel, düzensiz, düz veya kavisli boynuz şeklinde, vb.
Sklerotia hücrelerinin yapısı ve oluşma mekanizması farklıdır ancak oluşumları miselyumun dallanmasının ve hiflerin bölünmesinin artmasıyla gerçekleşir. Sklerotia oluşturmanın bilinen iki yolu vardır: terminal - hiflerin uçlarında; interkalar - ana hiphanın ayrı parçalarında.
Birçok mantarda, meyve veren cisimlerin ve bazı bitkisel yapıların gelişimi sırasında sahte doku oluşur - plektenkima (psödoparenkima). Üç yönde hücre bölünmesinden kaynaklanan gerçek parankim dokusunun aksine, plektenkima pleksus ve füzyonla oluşur. Az ya da çok izodiametrik hücrelerden oluşuyorsa buna paraplektenkima denir; net bir hifa benzeri yapıya (uzun hücreler) sahipse buna prosoplectenchyma denir.
Misel şeritleri- doğrusal olarak toplanmış hiflerin bitkisel yapısı. Misel şeritlerinin çapı, merkezi taban çevresinde yoğunlaşan hiflerin sayısına bağlıdır.
En basit durumda az sayıda paralel hif, mukoza dış zarları tarafından birbirine yapıştırılır veya oluşturularak daha dayanıklı bir bağlantıya girer. çok sayıda kısa anastomoz oluşumu. Diğer durumlarda Çaylarda kordonlar masif olduğunda hifleri belli bir farklılaşmaya uğrar. Dış elementler daha incedir ve merkezi kalın gövdenin etrafında bir tür kabuk oluşturur.
Rizomorflar- Merkezi hipanın büyüme yoğunluğu, yan dalların kapsamı ve hif hücrelerinin farklılaşma derecesi açısından farklı mantarlar arasında farklılık gösteren, toplanma açısından daha karmaşık olan hifler.
Rhizomorpha'nın dış kısımları genellikle koyu renklidir ve yüksek bitkilerin köklerine belli bir benzerlik gösterir. Büyük meyve veren gövdelere sahip mantarlarda yaygındırlar: basidiomycetes, keseli hayvanlar vb.
Misel iplikçiklerinin ve rizomorfların temel amacı, mantarların substratta yayılmasını ve besin maddelerinin hifler boyunca hareketini sağlamaktır.
Klamidosporlar- bunlar olgun ve eski kültürlerde miselyumun uçlarında veya seyri boyunca meydana gelen değişikliklerdir. Klamidosporların temel işlevi üreme değil, türün korunmasıdır. Şekilleri genellikle yuvarlak, oval veya hafif uzundur, çapı miselyumun çapını aşar. Bazı mantarların çift devreli bir duvarı vardır, yüzeyi pürüzsüz veya pürüzlüdür. Klamidosporlar miselyumun uçlarında görünebilir, daha sonra miselyum - interpolar (orta) boyunca terminal olarak adlandırılırlar.
Eski kültürlerde, büyük klamidospor kümeleri genellikle bir tespih veya kolyeyi anımsatan tuhaf bir biçimde görülür. Genç ve olgun klamidosporlar çimlenme yeteneğine sahiptir. Eski hücreler dejenere olur.
Tartışma. Sporların yardımıyla mantarlar sadece üremekle kalmaz, aynı zamanda çevreye de yayılır. Bu, spor kabuklarının agresif faktörlere karşı yüksek direnci ile kolaylaştırılmıştır. Sporlar, miselyum üzerinde bulunan özel kaplar - sporangia (torbalar) ve ekzosporlar içinde oluşturulan endosporlara ayrılır.
Mükemmel mantarlarda sporlar oosporlara, zigosporlara, askosporlara, basidiosporlara, endosporlara, fialosporlara ve klamidyasporlara ayrılır. Kusurlu mantarların sporları da büyüklük ve kökene göre çeşitli gruplara ayrılır. Miselyumun içinde miselyumun segmentasyonuyla oluşturulan endosporlar arasında artrosporları, klamidosporları ve blastosporları içeren tallosporlar bulunur. Ek olarak kusurlu mantarlar, kusurlu konidia olarak kabul edilen konidia, makrokonidia, aleuria (mikrokonidia) ve hemisporların oluşumuyla karakterize edilir.
Hemisporlar miselyum ile daha sıkı bir şekilde ilişkilidir ve misel ipliğinin enine bölünmesinden sonra ayrılan bir veya iki segmenti temsil eder. Şekilleri silindirik, bazen yuvarlak veya çok yönlüdür, kabuk çift devrelidir.
Mantarların hücre duvarının kimyasal bileşimi ve yapısı
Mantarların hücre duvarı çok katmanlıdır; kimyasal bileşimleri farklı olan yapısal karbonhidratların oluşturduğu farklı katmanlar, kimyasal bileşimlerine göre 3 gruba ayrılabilir:
glikoz polimerleri(glukan, kitin, selüloz). Glukanlar çoğu mantarın hücre duvarının dış katmanını oluşturur. Mantar hücre duvarının iç tabakası, ona sertlik kazandıran kitin zincirlerinden oluşur. Kitin, çoğu mantarda bulunmayan ancak şu anda tipik mantarlara ait olmayan oomisetlerin hücre duvarının bir parçası olan selülozun yerini alır. Deasetillenmiş kitin, kitin ile kombinasyon halinde zigomisetlerin hücre duvarını oluşturan kitosan olarak adlandırılır.
diğer monosakaritlerin polimerleri (mannoz, galaktoz vb.) Hemiselüloz genel adı altında matrisin temelini oluşturdukları yüksek bitkilerden farklı olarak, mantarlara daha az karakteristiktirler. Bunun istisnası, hücre duvarları mannan adı verilen mannoz polimerleri açısından özellikle zengin olan mayadır. Bu duvar bileşiminin tomurcuklanmayı daha iyi sağladığı varsayılmaktadır.
Peptitlere (glikoproteinler) kovalent olarak bağlanan karbonhidrat polimerleriÇok katmanlı bir hücre duvarının orta katmanını oluştururlar ve hem hücrelerin yapısal bütünlüğünün korunmasında hem de çevre ile olan metabolik süreçlerinde önemli rol oynarlar.
Mantar hücresinin diğer spesifik özellikleri şunları içerir: plastid yokluğu bu da onu bir hayvan hücresine yaklaştırıyor;
Nişasta yok eumycetes'te hayvan nişastası glikojenine yakın bir polisakarit ile değiştirilir, oomisetlerde kahverengi alglerin laminarinine yakın bir polisakkarit ile değiştirilir. Mantarlara özgü bir takım yedek karbonhidratlar da üretilir.
Spesifik gelişimi ikincil metabolitler Bunlar arasında antibiyotikler, fito- ve mikotoksinler ve fitohormonlar önemli bir rol oynamaktadır.
Mantarların kendine özgü özellikleri arasında ayrıca Heterokaryoz ve paraseksüel süreç.
Mantarlarda, bazı genler için heteroalelik olan çekirdeklerin uzun süre bir hücrede kaldığı heterokaryoz veya heteronükleerlik olgusu çok yaygındır. Heterokaryoz, haploid mantarlarda heterozigotluğun yerini alır ve mantarların değişen koşullara hızlı adaptasyonunu destekler. Farklı çekirdeklerin varlığı, mantarların bir takım spesifik özelliklerinden kaynaklanmaktadır:
1. Bir hücrede birden fazla çekirdeğin bulunması
2. çekirdeklerin bir hücreden diğerine geçebildiği, gözenek adı verilen bir veya daha fazla açık deliğin bulunduğu hücre duvarının spesifik yapısı
3. Bir koloni içindeki hifler ve hatta aynı mantar türünün farklı sporlarından büyüyen farklı yakın aralıklı koloniler sıklıkla birlikte büyür, bunun sonucunda farklı suşların çekirdek değişimi mümkündür.
Paraseksüel (psödoseksüel) işlem. Heterokaryotik hücrelerin çekirdekleri herhangi bir lokus sigortasında heteroalelik ise, heterozigot diploid bir çekirdek ortaya çıkar. Bir spora girebilir ve diploid heterozigot bir klona yol açabilir. Mitoz sırasında diploid çekirdekler, bir kromozom setinin kaybı nedeniyle haploid duruma dönebilir veya içlerinde kromozom bölümleri değişimi meydana gelebilir (mitotik geçiş). Her iki sürece de ebeveyn genlerinin ve dolayısıyla fenotiplerin rekombinasyonu eşlik eder. Paraseksüel (aseksüel) rekombinasyon, milyonda bir çekirdeği aşmayan çok nadir bir olgudur, ancak miselyumdaki çok sayıda çekirdek nedeniyle mantar popülasyonlarında sürekli gözlenir.
Üreme– bitkisel, aseksüel, cinsel.
Bitkisel- Thallus'un parçalanması, bir süre dinlenme döneminden sonra miselyumda filizlenen ve maya içinde tomurcuklanan klamidosporların oluşumu.
aseksüel Farklı mantarlarda üreme hareketli ve hareketsiz sporlar tarafından gerçekleştirilebilir. Zoosporlar, suda yaşayan mantarlar ve alglerle genetik bağlantıların açıkça görülebildiği, suda yaşayan ve bazıları karada yaşayan nispeten küçük bir mantar ve mantar benzeri organizma grubu oluşturur. Kamçının yapısı, belirli bir krallığa atandığında önemli bir teşhis özelliğidir. Mantarların büyük çoğunluğunda eşeysiz üreme sırasında hareketsiz sporlar oluşur, bu da onların karada çok eski bir şekilde ortaya çıktıklarını gösterir. Oluşum ve lokalizasyon yerine göre, sporangia'da oluşan endojen sporangiosporlar ve özel hipha - conidiophores üzerinde gelişen ekzojen (conidia) ayırt edilir. Konidialar çoğu mantarda (ascomycetes, basidiomycetes, bazı zygomycetes) oluşur ve son derece çeşitli olan ve mantarların teşhisinde yaygın olarak kullanılan konidial sporülasyonu oluşturur.
Mantarların cinsel üremesi hem cinsel sürecin morfolojisinde hem de cinsiyetin genetik ve fizyolojik düzenleme mekanizmalarında ve kalıtsal bilgilerin aktarımında kendine özgü bir özelliği vardır.
Somatogami- En yaygın ve en basit cinsel süreç türü, gametlere farklılaşmamış iki somatik hücrenin füzyonundan oluşur. Bazen hücre füzyonu olmadan da meydana gelir; hücre içindeki çekirdekler birleşir. Çoğu basidiomiset, keseli maya ve diğer bazı taksonlarda bulunur.
Gametangiogami– gametangia haploid miselyumda izole edilir ve içerikleri cinsel süreç sırasında boşaltılır. Bu cinsel süreç çoğu keseli mantarın karakteristiğidir. Gametangiogaminin bir çeşidi, zygomycetes'teki zigogamidir.
Oyuntogami izo-hetero ve oogami biçiminde mantarlarda diğer ökaryotlara göre çok daha az sıklıkta meydana gelir. İzo ve heterogami yalnızca chytridiomycetes'te meydana gelir. Sperm ve yumurta oluşumuyla oluşan klasik oogami mantarlarda ifade edilmez, ancak oldukça değiştirilmiş varyantlar bulunur.
Mantarlarda cinsiyet düzenlemesinin özelliklerine göre, çeşitli cinsel süreç türleri ayırt edilir.
Jinandromiksis Oogonia ve anteridianın, örneğin geç yanıklık veya patates mantarı gibi farklı tahaller üzerinde geliştiği diocious oomycetes örneği kullanılarak düşünülebilir. Genetik olarak homojen bir tür monokültürde yetiştirilirse, yalnızca eşeysiz olarak çoğalır. İki suşun miselleri yakındaysa, steroid niteliğindeki kimyasal salgıların (seks feromonları) etkisi altında morfogenetik değişiklikler izlenebilir. Anteridiol, partnerde anteridia oluşumunu indükler ve oogoniol, oogonium oluşumunu indükler. Bu durumda cinsiyetin düzenlenmesi görecelidir: Bir suşun anteridia mı yoksa oogonia mı oluşturacağı, içindeki ve partnerindeki karşılık gelen feromonların niceliksel oranına bağlıdır. Dolayısıyla cinsel sürecin adı - gynandromixis.
Dimixis veya heterotalizm. Mantarların homo veya heterotalik olabileceği uzun zamandır bilinmektedir. Homotalik türlerde cinsel süreç sırasında miselyum içindeki genetik olarak özdeş çekirdekler birleşir. Heterotalik türlerde, cinsel döngüyü geçmek için, bir aşamada (farklı mantar türleri için farklı), sporların soyundan gelenlerin (daha doğrusu çekirdeklerinin) füzyonu gereklidir. İki suşun cinsel olarak uyumlu olabilmesi için, çiftleşme lokusları adı verilen belirli lokuslarda genetik farklılığın (heteroalelisite) olması gerekir. Çoğu mantar (zygomycetes, ascomycetes ve bazı basidiomycetes) iki alel ile bir çiftleşme lokusuna sahiptir. Çiftleşme lokusu, cinsiyet feromonlarının sentezini kontrol eden birkaç genden oluşur. Bu tür heterothallizme tek faktörlü veya iki kutuplu denir. Mayozdan sonra bu tür mantarların yavruları, 1:1 oranında kendi kendine uyumsuz, ancak karşılıklı olarak uyumlu iki gruba ayrılır; Akraba (akrabalı çiftleşme) ve akraba olmayan (yabancı çiftleşme) melezleme olasılığı, biseksüel yüksek ökaryotlarda olduğu gibi %50'dir.
Daha yüksek basidiomisetlerin genomunda iki çiftleşme lokusu vardır - A ve B ve yalnızca her iki lokusta heteroalelik olan suşlar birbiriyle uyumludur (Ax Bx, Ay By ile uyumludur, ancak Ax By ve Ay Bx ile uyumlu değildir). Bu tür heterotalizme iki faktörlü veya tetrapolar denir. Akrabalı çiftleşme olasılığını %25'e kadar azaltır.
Diyaforomiks– daha yüksek basidiomisetler, popülasyonu oluşturan farklı türlerde rastgele bulunan çiftleşme lokusunun iki değil birçok aleline sahiptir. Bu çiftleşme kontrolü, farklı alellere sahip suşların karşılıklı olarak uyumlu olması ve çok sayıda alel bulunması nedeniyle, %100 yavrulama şansı sağlar. Sonuç olarak panmix hibrit popülasyonları oluşur.
Mantarların yaşam döngüleri mantarlar kadar çeşitlidir. Ana döngüler, mantarların bölümlerine ait olmaları
1. Eşeysiz döngü, cinsel üremeyi kaybetmiş büyük bir kusurlu veya mitofungus grubunun karakteristiğidir. Çekirdeklerinin bölünmesi yalnızca mitotiktir. Mitofungusların büyük çoğunluğu keseli mantarlara aittir, ancak cinsel sürecin kaybı nedeniyle kusurlu mantarların veya döteromisetlerin resmi bir grubunu oluştururlar.
2. Haploid döngüsü. Bitkisel thallus haploid çekirdekler taşır. Cinsel süreçten (eş anlamlılık) sonra, diploid zigot (genellikle bir süre dinlendikten sonra) mayotik olarak - zigotik mayoz olarak bölünür. Zygomycetes ve birçok chytridiomycetes'in karakteristiği.
3. Haploid-dikaryotik döngü, gametangia (gametangiogamy) veya haploid miselyumun somatik hücrelerinin (somatogamy) içeriklerinin füzyonundan sonra çekirdeklerin dikaryonlar (genetik olarak farklı çekirdek çiftleri) oluşturmasıyla karakterize edilir. Eşzamanlı olarak bölünerek dikaryotik miselyum oluştururlar. Cinsel süreç, dikaryon çekirdeklerinin birleşmesi ile sona erer, ortaya çıkan zigot, dinlenme süresi olmaksızın mayoz bölünmeyle bölünür. Meiosporlar, keseli ve basidiomisetlerin askosporlar ve basidiosporlar formunda cinsel sporulasyonunu oluşturur. Çimlendiklerinde haploid miselyum oluşur. Keseli mantarların büyük çoğunluğunda (maya ve taphrin mantarları hariç), yaşam döngüsüne bitkisel miselyum (anamorf) formundaki haploid faz hakimdir, dikaryotik faz kısa ömürlüdür ve üzerinde torbaların bulunduğu askojen hiflerle temsil edilir. oluşur (teleomorf). Basidiomycetes'te yaşam döngüsünde dikaryotik faz baskındır; haploid faz kısa ömürlüdür.
4. Nesillerin izomorfik değişimi şeklindeki haploid-diploid döngüsü mantarlarda (bazı mayalar ve suda yaşayan chytridiomycetes) nadirdir.
5. Diploid döngü, oomisetlerin ve bazı keseli mayaların karakteristiğidir. Bitkisel thallus diploiddir, gametangia veya gamet oluşumu sırasında gametik mayoz gözlenir.
3. Ekolojik mantar grupları.
Mantarlar ve mantar benzeri organizmalar, ayrıştırıcıların trofik düzeyini işgal eden bakterilerle birlikte, tüm kara ve su ekosistemlerinde heterotrofik bloğun en önemli parçası olarak yer alır. Mantarların biyosferdeki geniş dağılımı bir dizi önemli özellik tarafından belirlenir:
1. Çoğunluğun thallus misel yapısına sahiptir. (alt tabakaya hızlı bir şekilde hakim olmanızı ve çevre ile geniş bir temas yüzeyine sahip olmanızı sağlar).
2. Kısa sürede büyük substrat kütlelerinin kolonileşmesine, çok sayıda spor oluşturmasına ve bunları uzun mesafelere dağıtmasına olanak tanıyan yüksek büyüme ve üreme hızı.
3. Çok çeşitli çevresel faktörlerle kendini gösteren yüksek metabolik aktivite.
4. Yüksek oranda genetik rekombinasyon, önemli biyokimyasal değişkenlik, çevresel esneklik.
5. Hızlı bir şekilde askıya alınmış animasyon durumuna geçme ve olumsuz koşullarda uzun süre hayatta kalma yeteneği.
Mantarlar için ana çevresel faktör gıda substratıdır. Bu faktöre bağlı olarak, trofik gruplar adı verilen ana mantar grupları ayırt edilir.
4. Mantar bölümlerinin kısa açıklaması.
MİKROBİYOLOJİ, 2010, cilt 79, sayı 6, s. 723-733
UDC 582.281(047)
MANTAR HÜCRE DUVARI: BİLEŞİM VE BİYOLOJİK FONKSİYON HAKKINDA MODERN KAVRAMLAR
© 2010 E.P. Feofilova1
Rusya Bilimler Akademisi Mikrobiyoloji Enstitüsü'nün adını taşıyan kurum. S.N. Vinogradsky RAS, Moskova
Editörün eline geçtiği tarih 05.11.2009
İnceleme, filamentli mantar hücresinin az çalışılmış yüzey yapısına - hücre duvarına (CW) ayrılmıştır. CS fraksiyonunda sitoplazmik içeriğin bulunmamasına ve kimyasal bileşimine ilişkin izolasyon yöntemleri ve saflık testlerine ilişkin veriler sağlanmaktadır. CS'nin yapısal (çerçeve) ve yapısal bileşenleri - aminopolisakkaritler, a- ve b-glukanlar, proteinler, lipitler, üronik asitler, hidrofobinler, sporopollenin ve melaninler - ayrıntılı olarak tartışılmaktadır. Anti-stres hücre korumasındaki yeni işlevi olan kitinin yanı sıra bu mantar aminopolisakkariti ile alg ve Arthropoda kitinin arasındaki farklara özellikle dikkat edilir. Apikal hipal büyüme olgusu ve özel mikropartiküllerin mantar hücresinin morfogenezine katılımı tartışılmaktadır. CS'nin sentezinde ve lizizinde yer alan enzimler hakkında veriler sağlanmaktadır. Son olarak, yüksek ökaryotların yüzey yapılarıyla karşılaştırıldığında SC'nin mantarlardaki işlevsel önemi tartışılmaktadır.
Anahtar kelimeler: filamentli mantarlar, hücre duvarı, izolasyon yöntemleri, kimyasal bileşim, apikal büyüme, fizyolojik fonksiyonlar, morfogenez.
Modern biyolojinin temel sorunlarından biri, gelişim sürecinde morfolojik olarak özdeş organizmaların oluşumunun nasıl gerçekleştiği, milyarlarca yıldır gezegenimizde meydana gelen bu sürece hangi biyokimyasal mekanizmaların ve hangi hücresel yapıların dahil olduğu sorusudur. yıllar. Stresin morfogenezi nasıl etkilediğine ve hangi biyopolimerlerin hücrelerin dış şeklini kontrol ettiğine dair veriler de daha az ilgi çekici değil. Son araştırmaların sonuçlarının gösterdiği gibi, filamentli mantarların büyüyen hifleri, hücre oluşum sürecini ve hücre morfolojisinin sabitliğininontogenezde nasıl korunduğunu anlamamızı sağlayan eşsiz bir modeldir. Bu fenomenin çalışmasına çok büyük bir katkı, filamentli mantarların CS'sinin bileşimi ve biyolojik fonksiyonu hakkındaki verilerle yapılmıştır.
Bitkilerin CS'si 1665'te, mantarların CS'si ise ancak 18. yüzyılın başında keşfedildi. Bununla birlikte, CS'nin yalnızca bir "çerçeve" işlevi yerine getirdiğine inanıldığından ve destekleyici bir yük taşıyan bir evin duvarlarıyla karşılaştırıldığından, bu yüzey hücresel yapısı pratikte uzun süredir incelenmemiştir. Ancak 20. yüzyılın başında CS'ye yönelik tutum dramatik bir şekilde değişti; yoğun bir şekilde incelenmeye başlandı, ancak esas olarak bitkiler ve bakteriler üzerinde. Bununla birlikte, filamentöz mantarların biyolojik olarak aktif madde üreticileri olduğu mantar yetiştiriciliği ve biyoteknolojik üretimin yoğun gelişimi, gelişmeyi yoğunlaştırdı.
1 Yazışma adresi (e-posta: [e-posta korumalı]).
Mantar CS çalışmalarına ilişkin bilimsel araştırmaların yapılması. 1980'lere kadar olan döneme ait bu yüzey yapısına ilişkin veriler, dünyanın ilk mantar CS kitabında özetlenmiştir. Daha sonraki yıllarda çalışmaların ana kısmı 20. yüzyılın sonu ve 21. yüzyılın başında ortaya çıktı. KS'nin sistematiği, kimyasal bileşimi, apikal hif büyümesi, kitin metabolizması, antifungal ilaçlar, hidrofobinler, kovalent bağlı proteinler, KS oluşumunda rol oynayan enzimler, hifal dallanma ve KS lizizi üzerine temel çalışmalar yapılmıştır. Bu veriler genelleştirilmemiştir, ancak hücrenin bu yüzey yapısının çok işlevli bir yük taşıdığını ve örneğin hücreyi olumsuz faktörlerin etkisinden korumak gibi önemli işlevleri yerine getirdiğini göstermemizi sağlayan, bunların birlikte tartışılmasıdır. morfogenezi kontrol etmek, üreme süreçlerine katılmak, antijenik ve yapışkan özellikleri belirlemek, dimorfizm süreçlerinin kontrolü ve hareketsiz mantar hücrelerinin oluşumu, harici bir sinyalin algılanması ve bunun zara ve hücre içi habercilere iletilmesi. Yukarıdakileri göz önünde bulundurarak, şu verileri tartışmanın uygun olduğunu düşündük: CS'nin kimyasal bileşimi, ana biyopolimerlerinin biyolojik fonksiyonu, CS'nin yapısı ve mantar hiflerindeki hücreler arası etkileşimler, apikal hif büyümesi olgusu ve CS'nin oluşumunda ve parçalanmasında rol oynayan ana enzimler.
HÜCRESELİN KİMYASAL BİLEŞİMİ
MANTAR DUVARLARI VE ANA BİLEŞENLERİNİN BİYOLOJİK FONKSİYONU
Mantar CS'nin kimyasal bileşiminin incelenmesi, bu yapının saf bir kısmının elde edilmesiyle başlar; CS'nin sitoplazmik kontaminasyondan saflaştırılması. İlk prosedür - mantar hücrelerinin yok edilmesi ve hücre içeriklerinin yıkanması - parçalayıcı enzimlerin yıkıcı aktivitesini önlemek amacıyla düşük bir sıcaklıkta (yaklaşık 4-5°C) gerçekleştirilir. Hücreler yok edilmeden önce sıvı nitrojen sıcaklığında dondurulur. Hücre imhası ya özel homojenizatörlerde ya da “katı basınç” yöntemi kullanılarak preslerde gerçekleştirilir. Ultrasonik parçalayıcının daha fazla kullanılması iyi sonuçlar verir. Hücresel içeriklerin yıkanması soğuk suyla gerçekleştirilir ve 4-5 kez tekrarlanır. Sitoplazmik içerikleri daha iyice çıkarmak için NaCl, 8 M üre, 1 M amonyum veya 0,5 M asetik asit ile yıkayın. Şu anda, mantar CS'leriniontogenezleri sırasında izole etmek için yöntemler geliştirilmiştir ve bu yöntemler miselyum ve dinlenme hücreleri için önemli ölçüde farklılık gösterir. Araştırmanın yönüne bağlı olarak, özellikle polisakkarit bileşimini belirlerken, lipitler CS'den organik çözücüler, örneğin 2: 1 oranında kloroform ve metanol kullanılarak ekstrakte edilir, bazen CS'nin sülfürik eter ile ilave işlenmesi iyi sonuç verir; sonuçlar. Bu solvent CS'yi iyi kurutur ve bazen dondurarak kurutmanın yerini alabilir. Bir sonraki adım, elde edilen CS fraksiyonunun saflığını belirlemektir. Bu amaçla ışık ve elektron mikroskobu ve özel boyama yöntemleri kullanılır. En yaygın kullanılan reaksiyon I3-'nin kitosan ile etkileşimine dayanmaktadır. Lugol çözeltisi ile izole edilmiş saf CS pembe veya mor renkliyken, bozulmamış CS kırmızıdır. DAPI (4,6-ekt1to-2-rkepyTsn-yo1) ile çekirdeklerin varlığına yönelik boyama da kullanılır. Saf bir CS fraksiyonunun izolasyonunun, kimyasal bileşiminin analizinin sonraki sonuçlarının bağlı olduğu çok sorumlu bir prosedür olduğu özellikle vurgulanmalıdır. Bu bağlamda, önceki tüm çalışmalar Mucorales miselyumunda glukan bulunmadığını göstermesine rağmen, mucorales mantarlarının bir temsilcisinin miselyumunda glukanın veya daha doğrusu bir kitosan-glukan kompleksinin bulunduğu ilginç bir çalışmadır.
Şu anda, mantar CS'nin bileşenleri yapısal bileşenlere (kitin, p-(1-3)-p(1-6)-glukanlar, p-(1-4)-glukan (selüloz) ve intrayapısal bileşenlere (bunlar mannoproteinleri, galakto-mannoproteinleri, ksilo-mannoproteinleri, glukurono-
mannoproteinler ve a-(1-3)-glukan. En az çalışılanlar, doğrusal bir glikoz polimeri olan a-glukanlardır (Schizosaccharomyces pombe'de bu glukan yaklaşık 260 glikoz kalıntısı içerir). Bu polimer, polimer molekülünün uçlarında yaklaşık 120 (1-3)-a-D-glikoz kalıntısı ve (1-4)-a-D-glikoz kalıntısı içeren iki bağlantılı doğrusal zincirden oluşur. Bu glukanın morfogenez süreci için gerekli olduğuna inanılmaktadır. Suda çözünmeyen bir glukan, kuruduktan sonra alkali içinde çözünme yeteneğini kaybeden Penicillium roqueforti miselyumunun CS'sinden izole edildi, ancak dimetil sülfoksit içinde% 10'luk bir lityum klorür çözeltisi içinde çözülebildi. Metilasyon yoluyla glukan incelendiğinde, yaklaşık 5: 2 oranında monosakkaritler arasındaki 1 - 3 ve 1 - "- 4 bağlarına ve tetra-O- izlerine karşılık gelen iki tri-O-metil glikoz türevi elde edildi. terminal indirgeyici olmayan mono-nosakarit kalıntılarına karşılık gelen metil türevi. Bu sonuç, glikoz kalıntıları arasında yalnızca 1 -»- 3 ve 1 -»- 4 bağı içeren moleküllerin doğrusal bir yapısını gösterir. Bu veriler polisakaritin 13C-NMR spektrumunun analizi ile doğrulandı; buradan tüm glukoz kalıntılarının a-konfigürasyonu da takip edilir. Bu tür polisakkaritler, özellikle onları suda çözünür hale getirebilen kimyasal modifikasyondan (sülfasyon veya karbosimetilasyon) sonra, biyolojik olarak aktif polimerler olarak kullanılabilir ve mantarların ve bitkilerin hücre duvarındaki biyolojik işlevlerinin incelenmesi açısından ilgi çekicidir.
Suda çözünmeyen (1 -»- 3)-a-glukanlar daha önce Penicillium chrysogenum dahil olmak üzere çeşitli yüksek ve düşük mantar türlerinden ve iki tür bağa (1-3) ve (1 -») sahip ilgili a-glukanlardan izole edilmişti. - 4), oranları farklı olan, daha yüksek bitkilerde (Aconitum kusnezoffii Reichb) bulundu.
Yapısal bileşen - selüloz - modern taksonomiye göre psödomantarlar olarak sınıflandırılan oomiset mantarlarının karakteristiğidir. CS'nin monosakarit bileşimi glikoz, mannoz, ksiloz içerir ve baskın şeker %68'e kadar olan glikozdur. Mantar CS'nin bileşimi ayrıca amino asitleri, lipitleri (%3'ten fazla değil) ve N-asetil-glukozamini içerir. 1,3-glukan gibi glukanlar, kitin ile mantar hücresinin iskeletini oluşturan kitin-glukan kompleksi (CGC) adı verilen güçlü bir kompleks oluşturur. Bu kompleks, güvenilir bir sistematik özellik olarak hizmet eden zygomycetes hariç hemen hemen tüm mantarların CS'sinde mevcuttur. Bazı mantarlarda, CS'nin yüzeyinde her üç glikoz birimine ß-1,6 bağları ile bağlanan yalnızca glikoz birimleri içeren ß-1,3-glukanlarla temsil edilen polisakkaritlerden oluşan bir mukoza malzemesi bulunmuştur.
Mantarların kuru miselyum kütlesinin oldukça büyük bir kısmı, yani% 5 ila 15'i hücre duvarıdır. Bileşimi büyük ölçüde değişir ve genellikle mantarların belirli taksonomik grupları için çok spesifiktir. Bu, Tabloda verilen maya, chytrid ve mucor mantarları - zygomycetes temsilcilerindeki kompozisyon örneklerinden görülebilir. 1.3 (Aronson, 1965).
* (Fosfatsız kül miktarı.)
Yapısal olarak mantar kabukları, matrisin amorf kütlesine dahil edilen mikrofibrillerin bulunduğu iki fazlı bir sistem temelinde inşa edilmiştir. Elektron mikroskobuna göre farklı yönelimli fibrillere sahip en az iki katmandan oluşur. İç katman genellikle hücrenin ana ekseni boyunca, dış katman ise ona açılı olarak yönlendirilir (Şekil 1.1). Maya kabuğu genellikle çok katmanlıdır; dıştaki kalın katmanda mannan ve içteki ince katmanda glukan yer alır. Su mantarlarında, örneğin Allomyces'te, kabuk, bir tekerleğin jant tellerine benzeyen sahte bölmeler - psödosepta oluşturur. Keseli ve basidiomisetlerde gerçek bölümler - septa - gözlenir. Keseli hayvanlarda, hücreler arasındaki bölmeler genellikle basit bir gözenek içerir ve oluşumu sırasında her iki yanında bir çift ozmofilik Voronin cisimciği görülebilir. Basidiomisetlerde bu gözenekler genellikle çok karmaşıktır ve kapaklar - parentozomlarla donatılmıştır (Şekil 1.2). Bu tür gözenekler, bir dizi yüksek basidiomisetin meyve veren gövdelerinde ve rizomorflarında bulunmuştur (Moore, 1965; Burnett, 1968). Bununla birlikte, bugüne kadar, asco- ve basidiomycetes'in septal gözeneklerinin yapısındaki bu farklılığın, bunların taksonomik bağlantısına mı yoksa keseli genomunun haploid ve homokaryotik yapısına ve di- ve heterokaryotik doğasına mı atfedilebileceği tam olarak açık değildir. basidiomisetler. Henüz böyle bir çalışma yok, ancak bunların mantar krallığındaki evrimsel ve taksonomik yapılar açısından önemi çok temeldir.
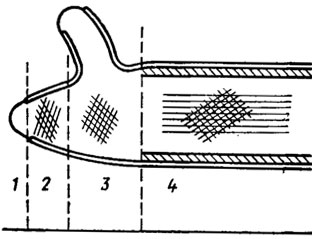

Elektron mikroskobik düzeydeki çalışmalarla mantar septal gözeneklerinin incelenmesine pek çok yeni bilgi eklenmiştir (Flegler ve diğerleri, 1976; Kamaletdinova ve Vasiliev, 1982). İlk olarak, basidiomisetlerin doliporlarının, misel hücrelerini, meyve veren cisimcikler oluşturmaya başlayana kadar birbirlerinden görünüşte hermetik olarak izole eden yapılar olduğu anlaşıldı (Flegler ve diğerleri, 1976). Bu izolasyon, çözünebilir proteolitik enzimler ve aynı zamanda meyve veren cisimcikler oluştuğunda kaybolan osmofilik (proteinler ve lipitler içeren) iki taraflı tıkaçlar tarafından gerçekleştirilir. Kaybolmalarına parentozomların delinmesi eşlik eder ve hifal hücreler arasındaki iletişim yeniden sağlanır.
Diskomisetlerin (örneğin, Peziza badia - Kamaletdinova, Vasiliev, 1982) meyve veren gövdelerindeki septalara ilişkin son gözlemler, hif hücrelerini ayıran benzer yapıların keseli mantarlar sınıfında da mevcut olduğunu gösterdi. Voronin cisimcikleri, hücre zarının şişe şeklindeki girintilerinde oluşan, osmofilik içerikleri (Voronin cisimcikleri) septumun yakınında salınan ve septumun açıklığında bulunan, yavaş yavaş içine nüfuz ederek bir oluşum oluşturan oluşumlarında yer alır. onu kapatan cihaz. Voronin gövdesinin üstteki hymenial katmana daha fazla nüfuz etmesi, gelecekteki bursa'nın ana hücresindeki septal gözeneğin üzerinde yer alan özel bir delikli yapı tarafından açıkça önlenmektedir; bu, subhymenial katmandan submikroskobik organellerin nüfuzundan tamamen izole olduğu ortaya çıkmaktadır. . Meyve veren gövdelerde oluşan parafizlerde de benzer yalıtım yapıları gözlenir.
Benzer yapılar, örneğin Arthrobotrys conoides gibi Voronin gövdeleri içeren kusurlu mantarların sporlu miselyumunda da mevcuttur. Penisilin üreten döteromiset Penicillium chrysogenum'da, septumda Kamaletdinova ve Vasiliev tarafından diskomiset Peziza badia'da bulunan yapıya tam olarak karşılık gelen bir yapı bulunmuştur (Kurilovich ve diğerleri, 1980).
Mantar kabuklarının iskelet temelleri kristal polisakkaritlerden oluşur: selüloz, kitin, kitosan, mannan, glukanlar ve diğerleri. Hepsi, başlangıç bileşenlerinin - heksoz monomerleri, amino ve asetaminoheksozların - β-1,4 bağları ile doğrusal bir yapıya sahiptir. Mikrokimyasal testlerin sonuçlarına (rutenyum kırmızısı boyama) dayanarak, daha önce mantarların hücre duvarlarının pektin içerdiğine inanılıyordu. Ancak kimyasal analiz sonuçları pektinin yapısında yer alan galakturonik asit monomerini doğrulamamıştır (Aronson, 1965).
Çoğu mantar için kitin ve kitosan, nitrojen içeren polimerler olarak kabuklarının çok karakteristik özelliğidir. Aynı zamanda, mantarların kitinini, böceklerin ve kabukluların kitinine çok benzemektedir ve bu, X-ışını kırınım desenleriyle de doğrulanmaktadır. Ancak mantarların kitininde hayvanlara göre daha az nitrojen bulunur ve bileşenleri arasında misetoz adı verilen metilpentoz bulunur. Mantarlardaki kitin, Van Wisseling yöntemi kullanılarak, kısmi alkalin deasetilasyonu ve ardından kitosanla reaksiyon ve X-ışını kırınımı kullanılarak mikrokimyasal olarak tespit edilebilir. Sadece saprolegnia ve peronosporlu mantarlar gibi oomisetlerde bulunmadı. Daha önce, mayada kitinin bulunmadığına inanılıyordu, ancak hücre bölümlerinde - Saccharomycetes septalarında - bulunduğuna inanılıyordu (Kulaev, 1975).
Kitinin Chitrydiales, Monoblepharidales, Protomycetales, Hyphochyiridiales, tüm Endomycetales, Blastocladiales, Mucorales, Entomophthorales, tüm keseli ve basidiomycetes ve bunlardan türetilen Fungi imperfekti (deuteromycetes) türlerinde bulunabileceği tespit edilmiştir. Bunun istisnası, kabuğundaki polisakkaritler arasında selüloz bulunan ve maya mantarlarının temsilcilerinde tamamen bulunmayan Oomycetes'tir.
Son zamanlarda, polimerlerin sentezi için mantar kitininin pratik kullanım olasılığının belirlenmesiyle bağlantılı olarak, kabuklu kitin ile karşılaştırıldığında bunun daha ayrıntılı bir çalışmasına ilişkin verilerle oldukça fazla sayıda çalışma ortaya çıkmıştır (Tablo 1.4). Asetat ve D-glukozamin, Aspergillus niger'de en az bulunanlardır; bu durum, mantar kabuğunun β-glukanı ve iki α-glukanındaki pentoz ve glikozun daha yüksek içeriğiyle açıklanmaktadır. Kırınım desenlerinin analizi, mantarlarda ve yengeçlerde kitinin aynı kristal yapısını gösterdi; ikincisinde biraz daha büyük bir düzen vardı. Ayrıca mantarların kitininin, yengeçlerin kitininin lamelli yapısının aksine, lifli bir yapıya sahip olduğu görülmüştür (Feofilova ve ark., 1980).

Bazı mukor mantarlarında kitinin yerini alan ve kitosan sülfatla reaksiyona girerek zayıf asit hidrolizi sırasında bulunan kitosan, Mucor rouxii'nin yanı sıra Phycomyces blakesleeanus'ta da bulunur. M. rouxii'de asetil gruplarının sayısının farklı olduğu ve sıfıra kadar değiştiği ortaya çıktı. Diğer aminopolisakkaritlerin yanı sıra, fosfatları bağlayabilen, kitosan tipi yapıya sahip, serbest bir amino grubuna sahip bir galaktozamin polimeri, Neurospora crassa, N. sitophitla, A. niger ve Botrytls drierea'nın hücre duvarlarından izole edildi (Aronson, 1965). Bazı mantarlar mannan, glukan ve proteinlerle ilişkili amino şeker polimerleri içerir.
Mantar hücre duvarlarındaki selüloz genellikle kitin ile aynı anda oluşmaz. Bunun istisnası, aynı anda bulundukları Hyphochytrid mantarları takımından bir Rhizidiomyces'tir. Selüloz, Acrasiales, Lagenidiales, Saprolegniales, Leptomitales ve Peronosporales takımına ait oomisetlerden elde edilen mantarlarda keşfedilmiştir (Aronson, 1965).
Su ortamlarında yaşayan mantarlar arasında selüloz genellikle yalnızca çift kamçılı zoosporlara sahip gruplarda bulunur. Monoflagellate zoosporlara sahip olan Blastocladiales ve Monoblepharidales'te bu özellik yoktur. Hem kitin hem de selüloz içeren ve Nable tarafından kitin içeren Chytridiales ve Blastocladiales ile selüloz içeren oomisetler arasında bir geçiş formu olarak kabul edilen Hyphochytridiales takımından Rhizidiomyces'in istisnası anlaşılabilir. Bu tek kamçılı form, Blastocladiales ve Monoblepharidales'te olduğu gibi kırbaç benzeri bir kamçı yerine tüylü bir zoospor kamçısına sahiptir. Oomisetlerin tüylü flagellalarının villus yapısının bakteri flagellalarının yapısına benzemesi, kırbaç benzeri olanların ise flagellatların flagellalarına tamamen benzemesi ilginçtir.
Mantarlardaki selüloz, Speitzer reaktifi veya %70'lik sülfürik asit çözeltisinin eklenmesiyle potasyum iyodür içindeki bir iyot çözeltisinden oluşan bir reaktif kullanılarak mikrokimyasal olarak kolayca tespit edilir.
Mantarlarda glukanlar çok bol miktarda bulunur ve yapı olarak selülozdan farklıdır. Monomerleri de glikozdur. Ancak en çok çalışılan maya glukanları selülozdan daha az kristal yapıya sahiptir. Glukanların kristalliği, sülfürik asit ile muamele edildiklerinde hidroglukan oluşumuna bağlı olarak artar. Proteinlerle ilişkili benzer glukanlar mayalarda ve Penicillunt notatum'da bulunmuştur (Aronson, 1965). Mantarların kabuklarının bir parçası olan başka bir glukan, yüksek bitkilerin elek tüplerinde bulunana benzer ve bazik boyalarla güçlü bir şekilde lekelenmiş olan kallozdur, yani doğası gereği asidiktir ve selülozun aksine β-1'e sahiptir, 3-glukosidik bağlar. Bazik boyalarla lekelenen benzer bir glukan Sclerofinia'da bulunmuştur. Glukanlar ayrıca Aspergillus fischeri, Allomyces macrogynus ve Neurospora crassa'da da bulunur. Mantarlar ayrıca, özellikle Penicillium cinsinde yaygın olan, çeşitli şekerlerin monomerlerinden oluşan heteropolisakkaritleri de içerir. Coccidioides ve Cryptococcus gibi hayvanlar için patojen olan maya benzeri formlarda benzer ancak asidik polisakkaritler, hücrelerini çevreleyen kapsüllerin bir parçasıdır. Mantarların hücre duvarlarını ve depo maddelerini oluşturan monosakkaritlerin ve şeker asitlerinin glukan ve diğer polimerizasyon ürünlerinin örnekleri Tablo'da verilmiştir. 1.5.

Mannanlar, mannoz monomerlerinden oluşan polisakkaritlerdir. Özellikle mayalarda bol miktarda bulunur ve ağaçların kabuğu altındaki kambiyumun yüzeyinde yaşayan maya türlerinde sık görülür. Bu formlar arasında iğne yapraklı ağaçların kabuğu altında yaşayan Hansenula; Bu mayaların hücrelerini çevreleyen kapsülde polisakkarit, fosfomannan formunda bulunur. Bu hidrofilik ve sümüksü polisakkarit, kabuk böceklerinin gövdesini kaplayan kıllara maya ile birlikte yapışır ve bu şekilde maya, onların yardımıyla bir ağaçtan diğerine aktarılır (Wickerham ve Barton, 1961). Mannanlar henüz hif oluşturan mantarlarda bulunmamıştır, ancak mannoz bunların hücre duvarlarının hidrolizatlarında bulunur.
Mantarlar ayrıca galaktoz, 6-deoksipentoz, metilpentozlar ve özellikle mucoraceae'de çoğunlukla fruktoz içeren polisakkaritler içerir. Penicillium chrysogenum'un hücre duvarlarında bulunan pentozlar arasında 6-deoksipentoz, ramnoz ve ksiloz vardı ve bunlar aynı zamanda kav mantarı Polysiictus sanguineus'ta da bulundu.
Mayalarda, örneğin bebeklerde pamukçuk etkeni olan Candida albicans'ta polisakkarit-protein kompleksleri keşfedilmiştir. Mannan-protein kompleksi Saccharomyces'te bulundu.
Mantarlardaki lipitlerin miktarı çevre koşullarına ve kültürün yaşına bağlı olarak büyük ölçüde değişir. Bazen sayıları hücrelerin kuru madde kütlesinin% 35-36'sına ulaşır. Mayalarda lipitlerin %3'ünden fazlası membranlarında bulunur. Ayrıca mukor mantarlarının kabuklarında da bulunurlar, örneğin Mycor rouxii ve Phycomyces'te, sporangioforlarda kuru kütlelerinden yaklaşık %25 oranında lipit bulunur. Muhtemelen orada kütikül tipi oluşumlarda bulunurlar (Aronson, 1965).
Pigmentler ayrıca mantarların hücre duvarlarında gözle görülür miktarlarda bulunur. Pigmentler, özellikle de genellikle özel bir katmanda lokalize olan siyah pigment melanin, miselyumun hücre duvarlarında veya birçok mantarın spor kabuklarında sıklıkla bulunur. Böyle bir melanin tabakası Neurospora tetrasperma'nın askospor membranlarında mevcuttur (Aronson, 1965).
Biyosentezinde yer alan düşük polifenol oksidaz aktivitesine sahip ve metabolizmada aktif dehidrojenazların baskın olduğu mantarlarda melanin yoktur. Fusarium, Trichothecium, Arthrobotris, Cephalosporium ve diğer pek çok türün temsilcilerini içeren bu tür mantarlar, hafif koruyucu bir rol üstlenen karotenoidlerin bolluğuna bağlı olarak çoğunlukla pembe veya turuncu sporlu yapı rengiyle karakterize edilir. koyu renkli mantarlarda melanine ait olan koruyucu ve antioksidandır. Karotenoidlerin varlığı ile yüksek düzeyde dehidrojenaz aktivitesi arasındaki bağlantı, dehidrojenazların etkisi için redoks rejiminin optimal koşullarının, karotenoidlerin ve diğer terpenoid şant ürünlerinin biyosentezi için optimal koşullarla örtüşmesiyle açıklanmaktadır. Karotenoid pigmentler artık genellikle mantarların hücre duvarlarının bir parçası değildir, ancak hücre zarında veya sitoplazmada dağılmış lipit damlacıklarında lokalizedir. Bazı durumlarda, mantarlarda, böceklerin göz pigmentleri olan ommokromlar, Pycnoporus (Polyporus) cinnabarinus (Shivrina, 1965) poliporunda veya Russula emetica ve R. paludosa (Efimenko, russula türlerinde) fikobilinler gibi tamamen özel pigmentler bulunur. 1972), mavi-yeşil ve kırmızı alglerde fotosentez sisteminin bir parçasıdır.
Actinomycetes (Actinomycetales, Yunanca aktis - ray, mykes - mantar kelimelerinden gelir), Actinobacteria filumuna ait dallanan bakterilerdir. Karasal omurgalıların ve omurgasızların sindirim sisteminin normal mikroflorasının bir parçasıdırlar ve ayrıca toprakta bol miktarda bulunurlar ve topraktaki maddelerin ekolojisi ve döngüsünde kritik bir rol oynarlar.
Bu mikroorganizmalar, vücudun bağışıklık sisteminin işlevinin azalması sonucu ortaya çıkan birçok fırsatçı patolojinin etken maddeleridir. Aktinomisetler, bir dizi antibakteriyel ve antitümör maddesinin kaynağı olduklarından biyoteknolojide yaygın olarak kullanılmaktadır.
Pirinç. 1. Streptomycetes, büyük miktarda antibakteriyel ve antitümör ilacı sentezler.
Aktinomisetlerin yapısı: neden mantar değil de bakteridirler?
1. Genetik materyalin organizasyonu
Actinomycetes'in kalıtsal materyali, halka şeklinde olan ve sitoplazmada serbestçe yer alan bir deoksiribonükleik asit molekülünde bulunur - nükleoid adı verilen genetik materyalin aynı organizasyon şekli, diğer bakterilerin karakteristiğidir. Mantarlarda genetik materyal organize edilmiştir ve hücre çekirdeğinin bir parçasıdır.
Aktinomisetlerin DNA'sı çok sayıda GC çifti içerir (toplam nükleotid sayısının %65-75'i). Bu özellik sabittir, mutasyonlara bağlı değildir ve bu nedenle mikroorganizmaların taksonomisinde kullanılır. GC çiftlerinin bu içeriği aktinomisetlerin DNA'sını oldukça dirençli hale getirir; dolayısıyla aktinomisetlerin DNA analizi diğer bakterilere göre daha fazla zaman alır.

Pirinç. 2. Gr+ bakterisinin hücre duvarının şematik yapısı.

Pirinç. 3. Aktinomisetler, Gram boyalı.
Aktinomisetler, sitoplazmik membranın dışında yer alan ve pozitif Gram boyamalarına neden olan yoğun bir bakteri hücre duvarına sahiptir. Diğer Gram-pozitif bakteriler gibi, teikoik ve lipoteikoik asitlerin nüfuz ettiği birkaç düzine murein polimeri (peptidoglikan) katmanından oluşur. Lipoteikoik asitler bakterinin sitoplazmik zarına sabitlenir ve onu hücre duvarına bağlar. Teikoik asitler hücre duvarına negatif yük verir. Mantarların hücre duvarı diğer polimerlerden (kitin ve glukan) oluşur.
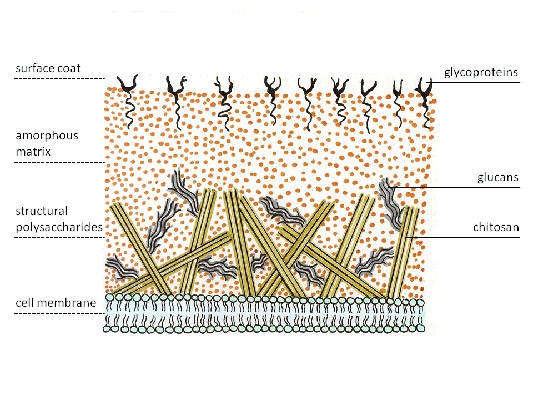
Pirinç. 4. Mantar hücre duvarı.
3. Hücresel organeller
Aktinomisetlerin diğer bakteriler gibi membran organelleri yoktur. Aktinomisetlerde 70S ribozom bulunurken mantarlarda diğer ökaryotik organizmalarda olduğu gibi 80S ribozom bulunur.
4. Kolonilerin büyümesi
Büyüme sırasında miselyum oluşumu, aktinomisetlerin mantarlara en çok benzediği şeydir. Aktinomisetlerde miselyum, dallanan bir hif topluluğudur. Hifler septalarla birkaç nükleoid içeren uzun bakteri hücrelerine bölünür. Bazı türlerdeki bölmeler karşılıklı olarak dik yönde ilerleyebilir. Tomurcuklanarak hif dalı.
Bir substrata (toprak, silt veya besin ortamı) dönüşen miselyuma substrat miselyum adı verilir. Koloniye besin sağlar. Bir hava miselyumu substratın üzerinde yükselir ve koloniye "kabarıklık" verir - sporların yanı sıra "ikincil metabolitler" olarak adlandırılan (substrat miselyumunun "birincil metabolitlerinin" aksine) aralarında çok sayıda bulunan sporlar oluşturur. antibakteriyel maddeler.
Aktinomisetlerin yaşam döngüsü ve fizyolojisi
Yaşam döngüsü boyunca çoğu aktinomiset spor oluşturur. Bazı aktinomisetler miselyumun parçalanmasıyla çoğalır.
1. Sporülasyon
Aktinomiset sporları havadaki miselyumdan kaynaklanır. Bunlar ekzosporlardır; ana hücrenin dışında gelişirler. Sporların geliştiği hava miselyumunun hiphalarına spor taşıyıcıları denir. Sporlar, sporangiumun - bir sporangiumun (örneğin, streptomisetler, aktinoplanlar ve plymelia'da) ucundaki bir kalınlaşma içinde bulunabilir veya sporangium boyunca bir zincir halinde yerleştirilebilir (örneğin, nokardia ve aktinomadurada).
Oluşan spor sayısına göre aktinomisetler ikiye ayrılır:
- Monosporlu (örneğin, Saccaromonospora, Micromonospora, Thermomonospora) - genellikle tomurcuklanarak ve daha sonra ana hiphadan bir septumla ayrılarak tek sporlar oluşturur;
- Oligosporlu (örneğin Actinomadura) - spor taşıyıcısı boyunca kısa spor zincirleri oluşturur;
- Polisoroz (diğer aktinomisetlerin çoğu, örneğin Streptomyces, Frankia, Geodermatophilus) - sporangia içine alınmış birçok spor oluşturur.

Pirinç. 5. Frankia cinsinin aktinomisetlerin sporangium'u.
Aktinomiset sporları hareketli olabilir - bu durumda sporun bir kamçısı vardır ve hareket edebilir (aktinoplan sporları, jeodermatofiller ve dermatofiller). Çoğu durumda sporlar hareketsizdir ve rüzgar, su veya hayvanlar tarafından yayılır.

Pirinç. 6. Dermatofiller, ışık mikroskobu.
Aktinomisetlerde sporlanma özellikle olumsuz koşullar altında aktiftir. Sporların ısıya karşı direnci diğer bakterilerin sporlarıyla karşılaştırıldığında düşüktür, ancak kurumaya diğerlerinden daha kötü dayanamazlar ve bu nedenle çok büyük adaptif öneme sahiptirler. Aktinomisetler kuru çöl topraklarındaki diğer mikroorganizmalara baskındır.
Oğulların çimlenmesi, dış ortamda belirli bir nemin olmasını gerektirir. Suyun varlığında spor şişer, enzimler aktive edilir ve büyüme tüplerinin (gelecekteki bakteri gövdeleri) salınması ve nükleik asitlerin sentezi ile birlikte metabolik süreçler başlatılır.
2. Solunum tipi
Çoğu aktinomiset aerobdur (yaşamı sürdürmek için oksijene ihtiyaç duyarlar). Fakültatif anaeroblar (oksijenin hem varlığında hem de yokluğunda yaşayabilen bakteriler), miselyumun parçalanmasıyla üreyen, kısa misel evresine sahip türler arasında bulunur.
3.Asit direnci
Aktinomisetlerin asit toleransı vardır; asitli ortamda direnç, asitle doymuş orman topraklarında yaşamalarına olanak tanır. Laboratuvarda asit direnci, aktinomisetler içeren bir preparatın Ziehl-Neelsen'e göre boyanması (fuksin, ardından sülfürik asitle aşındırma ve metilen mavisi ile boyama) yoluyla belirlenebilir. Bu renge sahip aktinomisetlerin çoğu, asitle aşındırdıktan sonra rengi solmaz ve kırmızı macenta rengini korur. Alkali ortam bu bakteriler için elverişsizdir; yüksek pH değerlerinde sporlanmaya eğilimlidirler.
4. Metabolizmanın özellikleri

Pirinç. 7. Aerobik aktinomisetler agarın eğimleri üzerinde pigment oluşturur. Soldan sağa: Actinomadura madurae, Nocardia asteroides, Micromonospora.
Hava miselyumunun “ikincil metabolitlerin” oluşumundan yukarıda bahsedilmişti. Aralarında:
- ortamda büyürken hava miselyumunun farklı renklerine neden olan pigmentler;
- yağmurdan sonra toprağa, durgun suya ve bazı hayvanların derisine karakteristik bir koku veren uçucu kokulu maddeler;
- antibiyotikler:
A. antifungal – polienler;
B. antibakteriyel - örneğin streptomisin, eritromisin, tetrasiklin, vankomisin;
C. antitümör - antrasiklinler, bleomisin.
Aktinomisetler nerede yaşar?
Aktinomisetler toprakta çok sayıda bulunur ve misel formları sporlara göre çok daha azdır. Diğer bakterilerin kullanması zor olan organik maddeleri parçalayarak humus oluşumunda önemli rol oynarlar. Bu bağlamda, aktinomisetler sıhhi ve epidemiyolojik konularda sıhhi indikatör mikroorganizmalar olarak kullanılır: toprakta veya suda büyük miktarlarda tespit edilmeleri, ilgili substratta kompostun varlığını gösterir.

Pirinç. 8. Komposttaki aktinomisetler.
Aktinomisetler birçok bitkinin simbiyontudur ve azotu sabitlemelerine yardımcı olur. Aynı zamanda bu sınıfa ait birçok mikroorganizma bitki hastalıklarının patojenleridir.

Pirinç. 9. Patates streptomikozu.
Ayrıca toprak annelidlerinden (solucanlar gibi) büyükbaş hayvanlara kadar birçok hayvanın sindirim sisteminin normal mikroflorasında da bulunurlar.
Bu mikroorganizmalar bitkisel besinlerde bol miktarda bulunan selülozun parçalanmasına yardımcı olur. İnsanlarda aktinomisetler ağız boşluğunda (diş etleri ve plak), bağırsaklarda (kalın bağırsağın uzak kısımları), deride (yüz, burun kanatları, kulak arkası, parmakların arasında) ve solunum sisteminde bulunur. (esas olarak üst solunum yollarında).
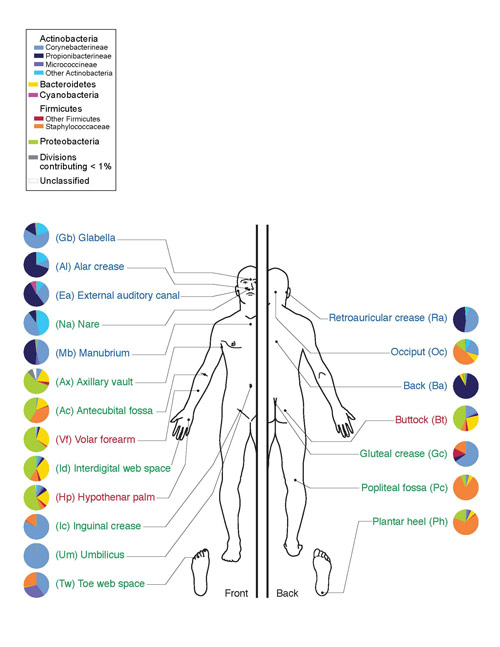
Pirinç. 10. İnsan derisinin mikroflorası. Actinobacteria filumu mavinin tonlarında gösterilir, Actinomycetes sınıfı ise parlak mavi ile gösterilir.
Aktinomisetler, vücudun bağışıklık reaktivitesi azalırsa, aktinomikoz - aktinomikoz granülomlarının oluşumundan oluşan fırsatçı hastalıklar - bağışıklık sistemi yetkin hücrelerle çevrelenmiş sarı kükürt tanelerine ("drusen") benzeyen bakteriyel cisimlerin birikmesine neden olabilir.

Enflamatuar reaksiyon, granülomların erimesine, fistül oluşumuna, organların delinmesine ve bakterilerin kan yoluyla yayılmasına yol açar.

Pirinç. 11. Aktinomikoz drusen, Gram boyama.

Pirinç. 12. Bir inekte üst çenenin aktinomikozu.
Pirinç. 13. İnsan maksiller aktinomikozu.