Actinomycetes (Actinomycetales จากภาษากรีก aktis - ray, mykes - เห็ด) เป็นแบคทีเรียที่แตกแขนงออกจากไฟลัม Actinobacteria พวกมันเป็นส่วนหนึ่งของจุลินทรีย์ปกติ ระบบย่อยอาหารสัตว์มีกระดูกสันหลังบนบกและสัตว์ไม่มีกระดูกสันหลัง และยังมีอยู่มากมายในดิน และมีบทบาทสำคัญในระบบนิเวศและวัฏจักรของสารในดิน
จุลินทรีย์เหล่านี้เป็นสาเหตุของโรคฉวยโอกาสหลายอย่างซึ่งเกิดขึ้นจากการทำงานที่ลดลง ระบบภูมิคุ้มกันร่างกาย. Actinomycetes ถูกนำมาใช้กันอย่างแพร่หลายในเทคโนโลยีชีวภาพ เนื่องจากเป็นแหล่งของสารต้านเชื้อแบคทีเรียและสารต้านมะเร็งหลายชนิด
ข้าว. 1. Streptomycetes สังเคราะห์ยาต้านแบคทีเรียและยาต้านมะเร็งจำนวนมาก
โครงสร้างของ actinomycetes: ทำไมพวกมันถึงเป็นแบคทีเรียไม่ใช่เชื้อรา?
1. การจัดระเบียบของสารพันธุกรรม
วัสดุทางพันธุกรรมของ Actinomycetes มีอยู่ในโมเลกุลหนึ่งของกรดดีออกซีไรโบนิวคลีอิก ซึ่งมีรูปร่างเป็นวงแหวนและตั้งอยู่อย่างอิสระในไซโตพลาสซึม ซึ่งเป็นลักษณะเฉพาะของแบคทีเรียชนิดอื่นที่จัดรูปแบบเดียวกันของการจัดระเบียบของสารพันธุกรรมที่เรียกว่านิวเคลียส ในเชื้อรามีการจัดระเบียบสารพันธุกรรมและเป็นส่วนหนึ่งของ นิวเคลียสของเซลล์.
Actinomycete DNA ประกอบด้วย จำนวนมากคู่ GC (65-75% ของจำนวนนิวคลีโอไทด์ทั้งหมด) ลักษณะนี้เป็นลักษณะคงที่ ไม่ขึ้นอยู่กับการกลายพันธุ์ จึงถูกนำมาใช้ในการจัดอนุกรมวิธานของจุลินทรีย์ เนื้อหาของคู่ GC นี้ทำให้ DNA ของ actinomycetes ทนไฟได้มาก ดังนั้นการวิเคราะห์ DNA ของ actinomycetes จึงใช้เวลานานกว่าเมื่อเปรียบเทียบกับแบคทีเรียชนิดอื่น

ข้าว. 2. โครงสร้างแผนผังผนังเซลล์ของแบคทีเรีย Gr+

ข้าว. 3. Actinomycetes ย้อมด้วยแกรม
Actinomycetes มีผนังเซลล์แบคทีเรียหนาแน่นซึ่งตั้งอยู่ด้านนอก เมมเบรนไซโตพลาสซึมและทำให้เกิดการย้อมสีแกรมที่เป็นบวก เช่นเดียวกับแบคทีเรียแกรมบวกอื่น ๆ ประกอบด้วยโพลีเมอร์มูรินหลายสิบชั้น (เปปทิโดไกลแคน) ซึ่งถูกแทรกซึมด้วยกรดเตอิโคอิกและไลโปเตโชอิก กรดไลโปเตโชอิกถูกยึดไว้ในเยื่อหุ้มเซลล์ของแบคทีเรียและเชื่อมต่อกับผนังเซลล์ กรดไทโชอิกส่งไปที่ผนังเซลล์ ประจุลบ- ผนังเซลล์ของเชื้อราประกอบด้วยโพลีเมอร์อื่น - ไคตินและกลูแคน
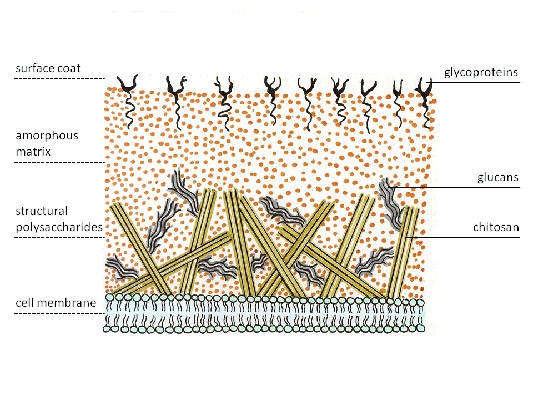
ข้าว. 4. ผนังเซลล์เชื้อรา
3. ออร์แกเนลล์ของเซลล์
Actinomycetes ก็เหมือนกับแบคทีเรียชนิดอื่นไม่มี ออร์แกเนลล์เมมเบรน- Actinomycetes มีไรโบโซม 70S ในขณะที่เชื้อรามีไรโบโซม 80S เช่นเดียวกับสิ่งมีชีวิตยูคาริโอตอื่น ๆ
4. การเติบโตของอาณานิคม
การก่อตัวของไมซีเลียมในระหว่างการเจริญเติบโตคือสิ่งที่แอคติโนไมซีตมีความคล้ายคลึงกับเชื้อรามากที่สุด ไมซีเลียมในกรณีของแอคติโนไมซีตคือกลุ่มของเส้นใยที่แตกแขนง เส้นใยจะถูกแบ่งโดยผนังกั้นเป็นเซลล์แบคทีเรียขนาดยาวที่มีนิวเคลียสหลายตัว ฉากกั้นในหลายสายพันธุ์สามารถวิ่งไปในทิศทางตั้งฉากกัน กิ่ง Hyphae โดยการแตกหน่อ
ไมซีเลียมเจริญเติบโตในสารตั้งต้น (ดิน ตะกอน หรือ สารอาหารปานกลาง) เรียกว่าสารตั้งต้น ให้สารอาหารแก่อาณานิคม ไมซีเลียมทางอากาศลอยอยู่เหนือสารตั้งต้นทำให้อาณานิคมมี "ความฟู" - มันก่อตัวเป็นสปอร์เช่นเดียวกับที่เรียกว่า "สารรอง" (ตรงกันข้ามกับ "สารหลัก" ของไมซีเลียมสารตั้งต้น) ซึ่งมีอยู่มากมาย สารต้านเชื้อแบคทีเรีย
วงจรชีวิตและสรีรวิทยาของแอคติโนไมซีต
ในช่วงวงจรชีวิต ที่สุด actinomycetes สร้างสปอร์ แอกติโนไมซีตบางชนิดแพร่พันธุ์โดยการแตกตัวของไมซีเลียม
1. การสร้างสปอร์
สปอร์ของ Actinomycete มีต้นกำเนิดมาจากไมซีเลียมในอากาศ สิ่งเหล่านี้คือเอ็กโซสปอร์ - พวกมันพัฒนานอกเซลล์แม่ เส้นใยของไมซีเลียมทางอากาศซึ่งสปอร์พัฒนาขึ้นเรียกว่าพาหะของสปอร์ สปอร์อาจมีความหนาขึ้นที่ส่วนปลายของ sporangium - sporangium (ตัวอย่างเช่น ใน streptomycetes, actinoplanes และ plymelia) หรือสามารถอยู่ในสายโซ่ตามแนว sporangium (เช่น ใน nocardia และ actinomadura)
ตามจำนวนสปอร์ที่เกิดขึ้น Actinomycetes แบ่งออกเป็น:
- Monosporous (เช่น Saccaromonospora, Micromonospora, Thermomonospora) - สร้างสปอร์เดี่ยว บ่อยครั้งโดยการแตกหน่อและแยกออกจากเส้นใยแม่ด้วยผนังกั้น;
- Oligosporous (เช่น Actinomadura) - สร้างสปอร์สายสั้น ๆ ตามแนวพาหะของสปอร์
- Polysporous (actinomycetes อื่นๆ ส่วนใหญ่ เช่น Streptomyces, Frankia, Geodermatophilus) - สร้างสปอร์จำนวนมากที่อยู่ใน sporangia

ข้าว. 5. Sporangium ของ actinomycetes ในสกุล Frankia
สปอร์ของ Actinomycete สามารถเคลื่อนที่ได้ - ในกรณีนี้ สปอร์มีแฟลเจลลัมและสามารถเคลื่อนที่ได้ (สปอร์ของแอคติโนเพลน, geodermatophiles และ dermatophiles) ในกรณีส่วนใหญ่ สปอร์จะไม่เคลื่อนไหวและแพร่กระจายโดยลม น้ำ หรือสัตว์

ข้าว. 6. Dermatophils กล้องจุลทรรศน์แบบใช้แสง
การสร้างสปอร์ใน actinomycetes จะออกฤทธิ์โดยเฉพาะอย่างยิ่งภายใต้สภาวะที่ไม่เอื้ออำนวย ความต้านทานของสปอร์ต่อความร้อนต่ำเมื่อเทียบกับสปอร์ของแบคทีเรียอื่น ๆ แต่พวกมันสามารถทนต่อการอบแห้งได้ไม่เลวร้ายไปกว่าตัวอื่น ๆ ดังนั้นจึงมีความสำคัญในการปรับตัวอย่างมาก Actinomycetes ครองจุลินทรีย์อื่น ๆ ในดินทะเลทรายแห้ง
การงอกของฝูงต้องมีความชื้นพอสมควร สภาพแวดล้อมภายนอก- ในที่ที่มีน้ำ สปอร์จะพองตัว เอนไซม์จะถูกกระตุ้นและกระบวนการเผาผลาญจะเกิดขึ้นพร้อมกับการปล่อยท่อการเจริญเติบโต (ร่างกายของแบคทีเรียในอนาคต) และการสังเคราะห์กรดนิวคลีอิก
2. ประเภทของการหายใจ
แอคติโนไมซีตส่วนใหญ่เป็นแอโรบี (พวกมันต้องการออกซิเจนเพื่อรักษาชีวิต) แบคทีเรียแบบไม่ใช้ออกซิเจน (แบคทีเรียที่สามารถมีชีวิตอยู่ได้ทั้งที่มีและไม่มีออกซิเจน) พบได้ในสปีชีส์ที่มีระยะเส้นใยสั้น โดยสืบพันธุ์โดยการแยกส่วนของไมซีเลียม
3. ความต้านทานต่อกรด
Actinomycetes มีความทนทานต่อกรด - ต้านทานในสภาพแวดล้อมที่เป็นกรด ซึ่งช่วยให้พวกมันอาศัยอยู่ในดินป่าที่มีกรดอิ่มตัว ความต้านทานต่อกรดในห้องปฏิบัติการสามารถกำหนดได้โดยการย้อมสารเตรียมที่มีแอคติโนไมซีตตาม Ziehl-Neelsen (ฟูชซินตามด้วยการกัดกรดซัลฟิวริกและย้อมด้วยเมทิลีนบลู) แอกติโนไมซีตส่วนใหญ่ที่มีสีนี้จะไม่เปลี่ยนสีหลังจากการกัดด้วยกรดและคงสีม่วงแดงไว้ สภาพแวดล้อมที่เป็นด่างไม่เอื้ออำนวยต่อแบคทีเรียเหล่านี้: ที่ pH สูงขึ้นพวกมันมีแนวโน้มที่จะสร้างสปอร์
4. คุณสมบัติของการเผาผลาญ

ข้าว. 7. แอกติโนไมซีตแบบแอโรบิกสร้างเม็ดสีบนวุ้นเอียง จากซ้ายไปขวา: Actinomadura madurae, ดาวเคราะห์น้อย Nocardia, Micromonospora
การก่อตัวของ "สารทุติยภูมิ" โดยไมซีเลียมทางอากาศได้ถูกกล่าวถึงข้างต้น ในหมู่พวกเขา:
- เม็ดสีที่ทำให้เกิดสีต่าง ๆ ของไมซีเลียมทางอากาศเมื่อเติบโตบนสื่อ
- สารระเหยที่ให้กลิ่นเฉพาะตัวแก่ดินหลังฝนตก น้ำนิ่ง และผิวหนังของสัตว์บางชนิด
- ยาปฏิชีวนะ:
ก. ต้านเชื้อรา – โพลีอีน;
ข. ต้านเชื้อแบคทีเรีย - ตัวอย่างเช่น streptomycin, erythromycin, tetracycline, vancomycin;
ค. ต่อต้าน - แอนทราไซคลีน, เบโลมัยซิน
Actinomycetes อาศัยอยู่ที่ไหน?
แอกติโนไมซีเตสใน จำนวนที่ยิ่งใหญ่ที่สุดพบได้ในดินและมีรูปแบบเส้นใยน้อยกว่าสปอร์อย่างมาก มีบทบาทสำคัญในการสร้างฮิวมัส โดยสลายสารอินทรีย์ที่แบคทีเรียชนิดอื่นนำไปใช้ได้ยาก ในเรื่องนี้ actinomycetes ถูกใช้เป็นจุลินทรีย์บ่งชี้ด้านสุขอนามัยในด้านสุขอนามัยและระบาดวิทยา: การตรวจหาใน ปริมาณมากในดินหรือน้ำบ่งชี้ว่ามีปุ๋ยหมักอยู่ในสารตั้งต้นที่เกี่ยวข้อง

ข้าว. 8. Actinomycetes ในปุ๋ยหมัก
Actinomycetes เป็นส่วนของพืชหลายชนิดที่ช่วยตรึงไนโตรเจน ในเวลาเดียวกันจุลินทรีย์หลายชนิดในชั้นนี้ก็เป็นเชื้อโรคของพืช

ข้าว. 9. Streptomycosis มันฝรั่ง
นอกจากนี้ยังพบได้ในจุลินทรีย์ปกติของระบบย่อยอาหารของสัตว์หลายชนิดตั้งแต่ดิน annelids(เช่นน้ำฝน) และปิดท้ายด้วยปศุสัตว์ขนาดใหญ่
จุลินทรีย์เหล่านี้ช่วยสลายเซลลูโลสซึ่งมีอยู่มากในอาหารจากพืช ในมนุษย์ Actinomycetes พบได้ในช่องปาก (เหงือกและคราบจุลินทรีย์) ลำไส้ (ลำไส้ใหญ่ส่วนปลาย) ผิวหนัง (ใบหน้า จมูก หลังหู ระหว่างนิ้ว) และในอวัยวะต่างๆ ระบบทางเดินหายใจ(ส่วนใหญ่อยู่ในทางเดินหายใจส่วนบน)
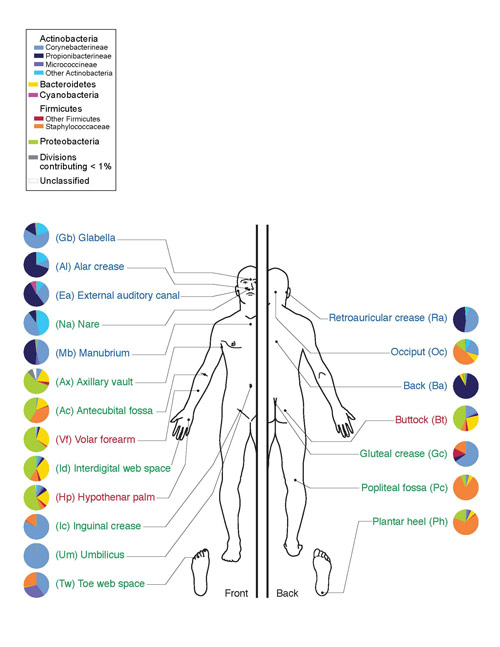
ข้าว. 10. จุลินทรีย์ของผิวหนังมนุษย์ ไฟลัมแอคติโนแบคทีเรียจะแสดงด้วยสีน้ำเงิน ส่วนคลาสแอคติโนไมซีตจะแสดงด้วยสีน้ำเงินสดใส
Actinomycetes หากปฏิกิริยาทางภูมิคุ้มกันของร่างกายลดลงสามารถทำให้เกิด actinomycosis - โรคฉวยโอกาสซึ่งประกอบด้วยการก่อตัวของ actinomycosis granulomas - การสะสมของแบคทีเรียที่มีลักษณะคล้ายเมล็ดกำมะถันสีเหลือง ("drusen") ล้อมรอบด้วยเซลล์ภูมิคุ้มกันบกพร่อง

ปฏิกิริยาการอักเสบนำไปสู่การละลายของ granulomas การก่อตัวของรูทวารนำไปสู่การเจาะอวัยวะและการแพร่กระจายของแบคทีเรียทางเลือด

ข้าว. 11. Actinomycosis drusen, คราบแกรม

ข้าว. 12. Actinomycosis ของกรามบนในวัว
ข้าว. 13. แอคติโนมัยโคซิสบนขากรรไกรของมนุษย์ แอกติโนมัยซีเตส –สิ่งมีชีวิตที่น่าทึ่ง
หา
ผนังเซลล์ของเชื้อรามีหลายชั้น โดยชั้นต่างๆ เกิดขึ้นจากคาร์โบไฮเดรตที่มีโครงสร้างซึ่งมีองค์ประกอบทางเคมีที่แตกต่างกัน ซึ่งสามารถแบ่งออกเป็น 3 กลุ่มตามองค์ประกอบทางเคมี:
กลูโคสโพลีเมอร์(กลูแคน, ไคติน, เซลลูโลส) กลูแคนประกอบขึ้นเป็นชั้นนอกของผนังเซลล์ของเชื้อราส่วนใหญ่ ชั้นในของผนังเซลล์เชื้อราถูกสร้างขึ้นด้วยสายไคตินทำให้มีความแข็งแกร่ง ไคตินเข้ามาแทนที่เซลลูโลสซึ่งไม่มีอยู่ในเชื้อราส่วนใหญ่ แต่เป็นส่วนหนึ่งของผนังเซลล์ของโอไมซีต ซึ่งปัจจุบันไม่ได้อยู่ในเชื้อราทั่วไป ไคตินแบบดีอะซิติเลตเรียกว่าไคโตซาน ซึ่งเมื่อรวมกับไคตินจะทำให้เกิดผนังเซลล์ของไซโกไมซีต
โพลีเมอร์ของโมโนแซ็กคาไรด์อื่นๆ (มานโนส กาแลคโตส ฯลฯ)ตรงกันข้ามกับพืชชั้นสูงซึ่งเป็นพื้นฐานของเมทริกซ์ที่อยู่ด้านล่าง ชื่อสามัญเฮมิเซลลูโลส ซึ่งพบได้น้อยสำหรับเชื้อรา ข้อยกเว้นคือยีสต์ซึ่งมีผนังเซลล์อุดมไปด้วยโพลีเมอร์แมนโนสที่เรียกว่าแมนแนนเป็นพิเศษ สันนิษฐานว่าองค์ประกอบของผนังนี้ช่วยให้มั่นใจได้ดีกว่า
โพลีเมอร์ของคาร์โบไฮเดรตที่เชื่อมโยงกับเปปไทด์โควาเลนต์ (ไกลโคโปรตีน)สร้างชั้นกลางของผนังเซลล์หลายชั้นแล้วเล่น บทบาทที่สำคัญทั้งในการรักษาความสมบูรณ์ของโครงสร้างเซลล์และในกระบวนการเผาผลาญกับสิ่งแวดล้อม
คุณสมบัติเฉพาะอื่น ๆ ของเซลล์เชื้อรา ได้แก่ : ไม่มีพลาสติกซึ่งนำมันเข้าใกล้เซลล์สัตว์มากขึ้น
ไม่มีแป้งซึ่งในยูไมซีตจะถูกแทนที่ด้วยโพลีแซ็กคาไรด์ใกล้กับแป้งไกลโคเจนของสัตว์ ในโอไมซีตจะมีโพลีแซ็กคาไรด์ใกล้กับลามินาริน สาหร่ายสีน้ำตาล- นอกจากนี้ยังมีการผลิตคาร์โบไฮเดรตสำรองจำนวนหนึ่งสำหรับเห็ดโดยเฉพาะ
การพัฒนาเฉพาะ สารทุติยภูมิซึ่ง บทบาทใหญ่ยาปฏิชีวนะ ไฟโตและสารพิษจากเชื้อรา และไฟโตฮอร์โมนก็มีบทบาทเช่นกัน
ลักษณะเฉพาะของเห็ดยังได้แก่ โรคเฮเทอโรคาริโอซิสและกระบวนการพาราเซ็กชวล
ในเชื้อราปรากฏการณ์ของเฮเทอโรคาริโอซิสหรือเฮเทอโรนิวเคลียร์เป็นที่แพร่หลายมากซึ่งในเซลล์เดียว เวลานานนิวเคลียสที่เป็นเฮเทอโรอัลลีลิกสำหรับยีนบางตัวจะถูกเก็บรักษาไว้ Heterokaryosis แทนที่เฮเทอโรไซโกซิตี้ในเชื้อราเดี่ยวและส่งเสริมการปรับตัวของเชื้อราอย่างรวดเร็วต่อสภาวะที่เปลี่ยนแปลง การมีอยู่ของเฮเทอโรนิวเคลียร์เกิดจากสาเหตุหลายประการ คุณสมบัติเฉพาะเห็ด:
1. การมีอยู่ของนิวเคลียสมากกว่าหนึ่งนิวเคลียสในเซลล์
2. โครงสร้างเฉพาะของผนังเซลล์ซึ่งมีรูทะลุหนึ่งรูขึ้นไปเรียกว่ารูพรุนซึ่งนิวเคลียสสามารถเคลื่อนย้ายจากเซลล์หนึ่งไปยังอีกเซลล์หนึ่งได้
3. เส้นใยภายในอาณานิคมเดียวและแม้แต่โคโลนีที่มีระยะห่างใกล้เคียงกันซึ่งเติบโตจากสปอร์ต่าง ๆ ของเชื้อราชนิดเดียวกันมักจะเติบโตร่วมกันอันเป็นผลมาจากการแลกเปลี่ยนนิวเคลียสของสายพันธุ์ต่าง ๆ เป็นไปได้
พาราเซ็กชวล (นามแฝง)กระบวนการ. หากในนิวเคลียสของเซลล์เฮเทอโรคาริโอตที่เป็นเฮเทอโรอัลลีลิกที่ฟิวส์ตำแหน่งใด ๆ นิวเคลียสซ้ำเฮเทอโรไซกัสจะปรากฏขึ้น มันสามารถเข้าไปในสปอร์และก่อให้เกิดโคลนเฮเทอโรไซกัสแบบดิพลอยด์ ในระหว่างกระบวนการไมโทซิส นิวเคลียสซ้ำสามารถกลับสู่สถานะเดี่ยวได้เนื่องจากการสูญเสียโครโมโซมหนึ่งชุด หรือการแลกเปลี่ยนส่วนของโครโมโซมอาจเกิดขึ้นได้ (การข้ามไมโทติค) กระบวนการทั้งสองจะมาพร้อมกับการรวมตัวกันใหม่ของยีนของผู้ปกครองและฟีโนไทป์ตามมาด้วย การรวมตัวกันอีกครั้งของพาราเซ็กชวล (ไม่อาศัยเพศ) เป็นปรากฏการณ์ที่หายากมาก โดยไม่เกินหนึ่งนิวเคลียสต่อล้าน แต่เนื่องจากมีนิวเคลียสจำนวนมากในไมซีเลียม จึงพบเห็นได้อย่างต่อเนื่องในประชากรเชื้อรา
การสืบพันธุ์– เป็นพืช, ไม่อาศัยเพศ, ทางเพศ.
พืชผัก– การกระจายตัวของแทลลัส, การก่อตัวของคลาไมโดสปอร์ซึ่งหลังจากช่วงพักตัวจะงอกเป็นไมซีเลียมและแตกหน่อในยีสต์
กะเทยการสืบพันธุ์ในเชื้อราต่าง ๆ สามารถทำได้โดยสปอร์ที่เคลื่อนที่ได้และที่ไม่สามารถเคลื่อนที่ได้ ซูสปอร์ก่อตัวเป็นกลุ่มเล็กๆ ของเชื้อราและสิ่งมีชีวิตคล้ายเชื้อรา ทั้งในน้ำและบนบก โดยที่ การเชื่อมต่อทางพันธุกรรมด้วยเห็ดน้ำและสาหร่าย โครงสร้างของแฟลเจลลามีความสำคัญ สัญญาณการวินิจฉัยเมื่อได้รับมอบหมายให้อยู่ในอาณาจักรใดอาณาจักรหนึ่งโดยเฉพาะ เชื้อราส่วนใหญ่ การสืบพันธุ์แบบไม่อาศัยเพศสปอร์ที่ไม่สามารถเคลื่อนที่ได้ถูกสร้างขึ้นซึ่งบ่งบอกถึงการเกิดขึ้นของพวกมันโบราณบนบก ตามสถานที่ของการก่อตัวและการแปลเป็นภาษาท้องถิ่น sporangiospores ภายนอกมีความโดดเด่นก่อตัวใน sporangia และภายนอก (conidia) ซึ่งพัฒนาบน hyphae พิเศษ - conidiophores Conidia เกิดขึ้นในเชื้อราส่วนใหญ่ (ascomycetes, basidiomycetes, zygomycetes บางชนิด) ซึ่งประกอบเป็นการสร้างสปอร์ของ conidial ซึ่งมีความหลากหลายอย่างมากและใช้กันอย่างแพร่หลายในการวินิจฉัยเชื้อรา
การสืบพันธุ์แบบอาศัยเพศของเชื้อรามีลักษณะเฉพาะของตัวเองทั้งในด้านสัณฐานวิทยาของกระบวนการทางเพศและในกลไกทางพันธุกรรมและ การควบคุมทางสรีรวิทยาเพศและการถ่ายทอดข้อมูลทางพันธุกรรม
โซมาโตกามี- กระบวนการทางเพศประเภทที่พบบ่อยที่สุดและง่ายที่สุดประกอบด้วยการรวมกันของเซลล์สืบพันธุ์สองตัวที่ไม่แตกต่างกัน เซลล์ร่างกาย- บางครั้งมันเกิดขึ้นแม้ไม่มีการหลอมรวมของเซลล์ - นิวเคลียสภายในเซลล์จะรวมกัน พบได้ใน basidiomycetes ส่วนใหญ่ ยีสต์ที่มีกระเป๋าหน้าท้อง และแท็กซ่าอื่นๆ
เกมแทงจิโอกามี– gametangia ถูกแยกออกจากเส้นใยเดี่ยวและเนื้อหาจะถูกระบายออกไปในระหว่างกระบวนการทางเพศ กระบวนการทางเพศนี้เป็นลักษณะของเชื้อราที่มีกระเป๋าหน้าท้องส่วนใหญ่ เกมทังจิโอกามีที่แตกต่างจากเดิมคือไซโกกามีในไซโกไมซีต
เกมโทกามีในรูปของ iso-hetero และ oogamy มักพบในเชื้อราน้อยกว่ายูคาริโอตอื่นมาก Iso- และ heterogamy เกิดขึ้นเฉพาะใน chytridiomycetes oogamy แบบคลาสสิกที่มีการก่อตัวของสเปิร์มและไข่ไม่แสดงออกในเชื้อรา แต่พบสายพันธุ์ที่มีการดัดแปลงอย่างมาก
ตามลักษณะของการควบคุมเพศในเชื้อรากระบวนการทางเพศหลายประเภทมีความโดดเด่น
ไจแอนโดรมิกซิสสามารถพิจารณาได้โดยใช้ตัวอย่างของ oomycetes ที่แตกต่างกัน ซึ่ง oogonia และ antheridia พัฒนาบน thalli ที่แตกต่างกัน เช่น โรคใบไหม้ปลายหรือเชื้อรามันฝรั่ง หากสายพันธุ์ที่เป็นเนื้อเดียวกันทางพันธุกรรมถูกปลูกในการปลูกพืชเชิงเดี่ยว มันจะแพร่พันธุ์เท่านั้น แบบไม่อาศัยเพศ- หากมีไมซีเลียของสองสายพันธุ์อยู่ใกล้ ๆ การเปลี่ยนแปลงทางสัณฐานวิทยาสามารถตรวจสอบได้ภายใต้อิทธิพลของการหลั่งสารเคมีที่มีลักษณะเป็นสเตียรอยด์ - ฟีโรโมนเพศ Antheridiol ทำให้เกิดการก่อตัวของ antheridia ในคู่หู และ oogoniol ทำให้เกิดการก่อตัวของ oogonia ในกรณีนี้ การควบคุมเรื่องเพศสัมพันธ์กัน: ไม่ว่าสายพันธุ์จะก่อให้เกิดแอนเธอริเดียหรือโอโอโกเนียหรือไม่นั้นขึ้นอยู่กับอัตราส่วนเชิงปริมาณของฟีโรโมนที่เกี่ยวข้องในฟีโรโมนและคู่ของมัน ดังนั้นชื่อของกระบวนการทางเพศ - gynandromixis
Dimixis หรือ Heterallismเป็นที่สังเกตมานานแล้วว่าเชื้อราสามารถเป็นโฮโมหรือเฮเทอโรทัลลิกได้ ในสายพันธุ์โฮโมทัลลิก นิวเคลียสที่เหมือนกันทางพันธุกรรมภายในฟิวส์ไมซีเลียมในระหว่างกระบวนการทางเพศ ในสายพันธุ์เฮเทอโรทัลลิก จำเป็นต้องมีวงจรทางเพศในบางช่วง (ต่างกันใน ประเภทต่างๆเชื้อรา) การรวมกันของลูกหลานของสปอร์ (แม่นยำยิ่งขึ้นคือนิวเคลียสของพวกมัน) เพื่อให้สายพันธุ์สองสายพันธุ์สามารถเข้ากันได้ทางเพศได้ จะต้องมีความแตกต่างทางพันธุกรรม (เฮเทอโรอัลลิลิซิตี) ในบางตำแหน่งที่เรียกว่าตำแหน่งผสมพันธุ์ เชื้อราส่วนใหญ่ (zygomycetes, ascomycetes และ basidiomycetes บางชนิด) มีตำแหน่งผสมพันธุ์หนึ่งตำแหน่งและมีอัลลีลสองตัว ตำแหน่งการผสมพันธุ์ประกอบด้วยยีนหลายตัวที่ควบคุมการสังเคราะห์ฟีโรโมนเพศ ภาวะเฮเทอโรทัลลิซึมดังกล่าวเรียกว่าปัจจัยเดียวหรือไบโพลาร์ ลูกหลานของเชื้อราดังกล่าวหลังไมโอซิสจะถูกแบ่งออกเป็นสองกลุ่มที่เข้ากันไม่ได้ แต่เข้ากันได้ในอัตราส่วน 1:1 เช่น ความน่าจะเป็นของการผสมข้ามพันธุ์ที่เกี่ยวข้อง (ผสมพันธุ์) และไม่เกี่ยวข้อง (ผสมพันธุ์ผสมพันธุ์) คือ 50% เช่นเดียวกับในยูคาริโอตที่สูงกว่าแบบกะเทย
ในจีโนมของ basidiomycetes ที่สูงกว่า มีตำแหน่งผสมพันธุ์สองตำแหน่ง - A และ B และมีเพียงสายพันธุ์ที่เป็นเฮเทอโรอัลลีลิกที่ตำแหน่งทั้งสองเท่านั้นที่เข้ากันได้ (Ax Bx เข้ากันได้กับ Ay By แต่ไม่ใช่กับ Ax By และ Ay Bx) ภาวะเฮเทอโรทัลลิซึมดังกล่าวเรียกว่า ทู-แฟคเตอร์ หรือ เตตราโพลาร์ จะช่วยลดโอกาสในการผสมพันธุ์ได้ถึง 25%
ไดอะโฟโรมิกซิส- basidiomycetes ที่สูงกว่านั้นไม่ได้มีสองอัลลีล แต่มีอัลลีลจำนวนมากของโลคัสผสมพันธุ์ซึ่งพบแบบสุ่มในสายพันธุ์ต่าง ๆ ที่ประกอบกันเป็นประชากร การควบคุมการผสมพันธุ์นี้รับประกันโอกาส 100% ที่จะผสมพันธุ์ เนื่องจากสายพันธุ์ที่มีอัลลีลต่างกันเข้ากันได้และมีอัลลีลจำนวนมาก เป็นผลให้เกิดประชากรลูกผสมแพนมิกซ์
วงจรชีวิตของเชื้อรามีความหลากหลายพอๆ กับเห็ดนั่นเอง วงจรหลักเป็นของแผนกเชื้อรา
1. วงจรไม่อาศัยเพศเป็นลักษณะของกลุ่มเชื้อราที่ไม่สมบูรณ์หรือไมโตเชื้อรากลุ่มใหญ่ที่สูญเสียการสืบพันธุ์แบบอาศัยเพศ การแบ่งนิวเคลียสของพวกมันเป็นแบบไมโทติคโดยเฉพาะ mitofungi ส่วนใหญ่เป็นของ marsupial fungi แต่เนื่องจากการสูญเสียกระบวนการทางเพศ พวกมันจึงประกอบขึ้นเป็น กลุ่มที่เป็นทางการเชื้อราหรือดิวเทอโรไมซีตที่ไม่สมบูรณ์
2. วัฏจักรเดี่ยว แทลลัสที่เป็นพืชมีนิวเคลียสเดี่ยว หลังจากกระบวนการทางเพศ (syngamy) ไซโกเทตซ้ำ (โดยปกติหลังจากช่วงเวลาที่เหลือ) จะแบ่งไมโอซิสแบบไมโอซิส - ไซโกติกไมโอซิส ลักษณะของ zygomycetes และ chytridiomycetes หลายชนิด
3. วงจรฮาพลอยด์-ไดคาริโอตมีลักษณะเฉพาะคือหลังจากการหลอมรวมของเนื้อหาของเกมทังเกีย (เกมทังจิโอกามี) หรือเซลล์ร่างกายของไมซีเลียมเดี่ยว (โซมาโตกามี) นิวเคลียสจะก่อตัวเป็นไดคาริโอต (คู่ของนิวเคลียสที่แตกต่างกันทางพันธุกรรม) พวกมันแบ่งตัวพร้อมกันทำให้เกิดไดคาริโอตไมซีเลียม กระบวนการทางเพศสิ้นสุดลงด้วยการหลอมรวมของนิวเคลียสไดคาริออน ส่งผลให้ไซโกตแบ่งตัวด้วยไมโอซิสโดยไม่มีระยะเวลาพัก Meiospores ประกอบด้วยการสร้างสปอร์ทางเพศของ marsupial และ basidiomycetes ในรูปแบบของ ascospores และ basidiospores เมื่องอกจะเกิดไมซีเลียมเดี่ยว ในเชื้อราที่มีกระเป๋าหน้าท้องส่วนใหญ่ (ยกเว้นยีสต์และเห็ดทัฟฟริน) วงจรชีวิตระยะเดี่ยวมีชัยเหนือในรูปของเส้นใยพืช (anamorph) ระยะไดคาริโอตมีอายุสั้นและแสดงด้วยเส้นใยแอสโคเจนิกซึ่งเกิดถุง (เทเลโอมอร์ฟ) ใน basidiomycetes ระยะไดคาริโอตมีอิทธิพลเหนือวงจรชีวิต ระยะเดี่ยวนั้นมีอายุสั้น
4. วงจรเดี่ยว-ไดพลอยด์ในรูปแบบของการเปลี่ยนแปลงไอโซมอร์ฟิกของรุ่นนั้นหาได้ยากในเชื้อรา (ยีสต์บางชนิดและไคไตรดิโอไมซีตในน้ำ)
5. วงจรดิพลอยด์เป็นลักษณะของโอไมซีตและยีสต์ที่มีกระเป๋าหน้าท้องบางชนิด แทลลัสของพืชเป็นแบบซ้ำซ้อนไมโอซิสที่เกิดจากเซลล์สืบพันธุ์จะสังเกตได้ในระหว่างการก่อตัวของเซลล์สืบพันธุ์หรือเซลล์สืบพันธุ์
3. กลุ่มสิ่งแวดล้อมเห็ด
เชื้อราและสิ่งมีชีวิตคล้ายเชื้อราพบได้ในทุกภาคพื้นดินและ ระบบนิเวศทางน้ำ, ยังไง ส่วนที่สำคัญที่สุดบล็อกเฮเทอโรโทรฟิกซึ่งอยู่ร่วมกับแบคทีเรีย ระดับโภชนาการตัวย่อยสลาย แพร่หลายเชื้อราในชีวมณฑลถูกกำหนดโดยจำนวน คุณสมบัติที่สำคัญที่สุด:
1. ส่วนใหญ่มีโครงสร้างไมซีเลียมของแทลลัส (ช่วยให้คุณควบคุมพื้นผิวได้อย่างรวดเร็วและมีพื้นผิวสัมผัสกับสิ่งแวดล้อมขนาดใหญ่)
2. การเจริญเติบโตและการสืบพันธุ์ที่รวดเร็วช่วยให้ เงื่อนไขระยะสั้นเติม ฝูงใหญ่สารตั้งต้นสร้างสปอร์จำนวนมากและกระจายไปในระยะทางไกล
3. กิจกรรมการเผาผลาญสูง แสดงออกในปัจจัยด้านสิ่งแวดล้อมที่หลากหลาย
4. อัตราการรวมตัวกันทางพันธุกรรมสูง, ความแปรปรวนทางชีวเคมีที่สำคัญ, ความเป็นพลาสติกในสิ่งแวดล้อม
5. ความสามารถในการเปลี่ยนไปสู่สถานะของแอนิเมชั่นที่ถูกระงับอย่างรวดเร็วเพื่อสัมผัส เงื่อนไขที่ไม่เอื้ออำนวยเป็นเวลานาน
หลัก ปัจจัยด้านสิ่งแวดล้อมสำหรับเห็ดนั้นเป็นสารตั้งต้นของอาหาร เกี่ยวกับปัจจัยนี้ กลุ่มเชื้อราหลัก มีความโดดเด่นซึ่งเรียกว่ากลุ่มโภชนาการ
4. คำอธิบายสั้น ๆแผนกเห็ด
เซลล์โปรคาริโอต
โปรคาริโอต- สิ่งมีชีวิตที่ไม่มีนิวเคลียสของเซลล์ที่ก่อตัวขึ้นและภายในอื่น ๆ ต่างจากยูคาริโอต ออร์แกเนลล์เมมเบรน(ยกเว้นถังเก็บน้ำแบบแบนในสายพันธุ์สังเคราะห์แสง เช่น ไซยาโนแบคทีเรีย) โมเลกุล DNA แบบเกลียวคู่ขนาดใหญ่เพียงชนิดเดียว (ในบางสปีชีส์ - เชิงเส้น) ซึ่งมีสารพันธุกรรมจำนวนมากของเซลล์ (ที่เรียกว่านิวครอยด์) ไม่ได้ก่อตัวที่ซับซ้อนด้วยโปรตีนฮิสโตน (ที่เรียกว่าโครมาติน ). โปรคาริโอตประกอบด้วยแบคทีเรีย รวมถึงไซยาโนแบคทีเรีย (สาหร่ายสีน้ำเงินแกมเขียว) และอาร์เคีย ทายาทของเซลล์โปรคาริโอตคือออร์แกเนลล์ของเซลล์ยูคาริโอต - ไมโตคอนเดรียและพลาสติด เนื้อหาหลักของเซลล์ที่เติมปริมาตรทั้งหมดคือไซโตพลาสซึมแบบเม็ดหนืด
เซลล์ยูคาริโอต
ยูคาริโอต- สิ่งมีชีวิตซึ่งแตกต่างจากโปรคาริโอตที่มีนิวเคลียสของเซลล์ที่ก่อตัวขึ้นซึ่งคั่นด้วยไซโตพลาสซึมด้วยเปลือกนิวเคลียร์ สารพันธุกรรมนั้นมีอยู่ในโมเลกุล DNA ที่มีเกลียวคู่เชิงเส้นหลายเส้น (ขึ้นอยู่กับประเภทของสิ่งมีชีวิตจำนวนต่อนิวเคลียสอาจมีตั้งแต่สองถึงหลายร้อย) ติดจากด้านในถึงเยื่อหุ้มนิวเคลียสของเซลล์และก่อตัวในอันกว้างใหญ่ ส่วนใหญ่ (ยกเว้นไดโนแฟลเจลเลต) สารเชิงซ้อนที่มีโปรตีนฮิสโตนเรียกว่าโครมาติน เซลล์ยูคาริโอตมีระบบของเยื่อหุ้มภายในซึ่งนอกเหนือจากนิวเคลียสแล้ว ยังก่อให้เกิดออร์แกเนลล์อื่นๆ อีกจำนวนหนึ่ง (เอนโดพลาสมิกเรติคูลัม อุปกรณ์กอลไจ ฯลฯ) นอกจากนี้ส่วนใหญ่มีเซลล์โปรคาริโอต symbionts แบบถาวรในเซลล์ - ไมโตคอนเดรียและสาหร่ายและพืชก็มีพลาสติดเช่นกัน
2) ไซโตพลาสซึม
ไซโตพลาสซึม - สภาพแวดล้อมภายในเซลล์ที่มีชีวิตหรือเซลล์ที่ตายแล้ว ยกเว้นนิวเคลียสและแวคิวโอล มีจำกัด พลาสมาเมมเบรน- ประกอบด้วยไฮยาพลาสซึมซึ่งเป็นสารโปร่งใสหลักของไซโตพลาสซึมส่วนประกอบของเซลล์ที่จำเป็นที่พบในนั้น - ออร์แกเนลล์รวมถึงโครงสร้างที่ไม่ถาวรต่างๆ - การรวม
องค์ประกอบของไซโตพลาสซึมประกอบด้วยสารอินทรีย์ทุกประเภทและ สารอนินทรีย์- นอกจากนี้ยังมีของเสียที่ไม่ละลายน้ำจากกระบวนการเผาผลาญและอะไหล่ สารอาหาร- สารหลักของไซโตพลาสซึมคือน้ำ
ไซโตพลาสซึมมีการเคลื่อนไหวอย่างต่อเนื่อง ไหลภายในเซลล์ของสิ่งมีชีวิต และเคลื่อนที่ไปพร้อมกับมัน สารต่างๆการรวมตัวและออร์แกเนลล์ การเคลื่อนไหวนี้เรียกว่าไซโคลซิส กระบวนการเผาผลาญทั้งหมดเกิดขึ้นในนั้น
ไซโตพลาสซึมมีความสามารถในการเจริญเติบโตและการสืบพันธุ์ และหากเอาออกไปบางส่วนก็สามารถกลับคืนสภาพเดิมได้ อย่างไรก็ตามไซโตพลาสซึมทำงานได้ตามปกติเมื่อมีนิวเคลียสเท่านั้น หากไม่มีมัน ไซโตพลาสซึมจะไม่สามารถดำรงอยู่ได้เป็นเวลานาน เช่นเดียวกับนิวเคลียสที่ไม่มีไซโตพลาสซึม
บทบาทที่สำคัญที่สุดของไซโตพลาสซึมคือการรวมโครงสร้างเซลล์ (ส่วนประกอบ) ทั้งหมดเข้าด้วยกันและจัดเตรียมไว้ ปฏิกิริยาทางเคมี- ไซโตพลาสซึมยังรักษา turgor (ปริมาตร) ของเซลล์และรักษาอุณหภูมิ
3) ผนังเซลล์
ผนังเซลล์- เยื่อหุ้มเซลล์แข็งที่อยู่นอกเยื่อหุ้มเซลล์ไซโตพลาสซึมและทำหน้าที่ด้านโครงสร้าง การป้องกัน และการขนส่ง พบได้ในแบคทีเรีย อาร์เคีย เชื้อรา และพืชส่วนใหญ่ สัตว์และโปรโตซัวหลายชนิดไม่มีผนังเซลล์
ผนังเซลล์โปรคาริโอต
ผนังเซลล์ของแบคทีเรียประกอบด้วย peptidoglycan (murein) และมีสองประเภท: แกรมบวกและแกรมลบ ผนังเซลล์ประเภทแกรมบวกประกอบด้วยชั้น peptidoglycan หนาซึ่งอยู่ติดกับเยื่อหุ้มเซลล์อย่างแน่นหนาและซึมซับด้วยกรด teichoic หรือ poteichoic ในประเภทแกรมลบชั้น peptidoglycan จะบางกว่ามากระหว่างมันกับพลาสมาเมมเบรนจะมีช่องว่าง periplasmic และด้านนอกเซลล์ถูกล้อมรอบด้วยเมมเบรนอีกอันหนึ่งซึ่งแสดงโดยสิ่งที่เรียกว่า lipopolysaccharide และเป็นสารก่อมะเร็งในแบคทีเรียแกรมลบ
ผนังเซลล์ของเชื้อรา
ผนังเซลล์ของเชื้อราประกอบด้วยไคตินและกลูแคน
ลักษณะของเห็ด
ไปจนถึงเห็ด ในความหมายกว้างๆรวมถึงยูคาริโอตเฮเทอโรโทรฟิกที่มีสารอาหารประเภทออสโมโทรฟิค เกี่ยวข้องกับนิสัยการบริโภคอาหาร คุณสมบัติลักษณะโครงสร้างและวิถีชีวิตของสิ่งมีชีวิตเหล่านี้ ที่สุด คุณสมบัติลักษณะเห็ด:
– รูปแบบของพืชที่พบมากที่สุดคือไมซีเลียมซึ่งประกอบด้วยเส้นใยที่แตกแขนงซ้ำ ๆ ซึ่งเจาะทะลุสารตั้งต้นและดูดซับสารอาหารที่ละลายในน้ำจากนั้น
– สารอินทรีย์ในสารตั้งต้นซึ่งเป็นแหล่งโภชนาการของเชื้อราโดยส่วนใหญ่จะอยู่ในรูปของไบโอโพลีเมอร์ที่ไม่สามารถทะลุผ่านได้ เยื่อหุ้มเซลล์- เซลล์เชื้อราผลิตเอนไซม์ดีโพลีเมอเรส ซึ่งถูกปล่อยออกสู่สารตั้งต้นและสลายโพลีเมอร์ให้กลายเป็นสารที่ง่ายกว่า สารประกอบอินทรีย์ซึ่งสามารถขนส่งเข้าสู่เซลล์เชื้อราได้ เอนไซม์ดังกล่าวมักเรียกว่าเอ็กโซไซม์
– การดูดซึมสารจากสารตั้งต้นนั้นเกิดขึ้นเนื่องจากแรงดัน turgor มหาศาลซึ่งในเซลล์ของเชื้อรานั้นสูงกว่ามากเมื่อเปรียบเทียบกับสิ่งมีชีวิตยูคาริโอตอื่น ๆ
– เชื้อรามีลักษณะเฉพาะมากคือการก่อตัวของสปอร์ซึ่งปรากฏขึ้นทั้งในระหว่างการสืบพันธุ์แบบไม่อาศัยเพศและการสืบพันธุ์แบบอาศัยเพศ การรับรู้ของเห็ดนั้นเป็นไปไม่ได้โดยร่างกายของพืช แต่โดยการสร้างสปอร์ เนื่องจากที่ตั้งของร่างกายพืชภายในสารตั้งต้นและวิถีชีวิตที่ไม่สามารถเคลื่อนที่ได้ในช่วงเวลาของการสืบพันธุ์โครงสร้างสปอร์ที่มีสปอร์ต่างๆจึงถูกสร้างขึ้นบนไมซีเลียมเห็ดซึ่งสูงขึ้นเหนือสารตั้งต้นและสปอร์ที่เกิดขึ้นบนพวกมันจะถูกกระจายโดยกระแสอากาศ ในระยะทางที่ไกลมาก
ระบบที่ทันสมัยเห็ด
โดย ความคิดที่ทันสมัยจากการศึกษาจีโนมของระบบนิเวศ “เห็ด” เป็นกลุ่มรวมที่จำแนกเชื้อราที่แท้จริงได้ คือ อาณาจักร Mycota หรือ Fungi ซึ่งส่วนใหญ่เป็นสายพันธุ์แท็กซ่าและน้อยกว่า กลุ่มที่หลากหลายสิ่งมีชีวิตคล้ายเชื้อรา (pseudomycetes) เป็นส่วนหนึ่งของอาณาจักร Straminopila ซึ่งรวมถึงสาหร่ายสี (Ochrophyta) ด้วย อาณาจักรแห่งเห็ดที่แท้จริงรวม 4 ส่วนที่แตกต่างกันในโครงสร้างของร่างกายพืชและลักษณะของการสืบพันธุ์: Chytridiomycota, Zygomycota, Ascomycota, Basidiomycota
อนุกรมวิธาน สัญญาณสำคัญใช้ในการสร้างระบบเห็ด
– โครงสร้างของร่างกายพืช
– องค์ประกอบทางเคมีและโครงสร้างของผนังเซลล์
– ประเภทของกระบวนการทางเพศ
– ประเภทของการสร้างสปอร์
ประเภทของเชื้อราและสิ่งมีชีวิตคล้ายเชื้อรา
อะมีบอยหรือพลาสโมเดียลแทลลัสเป็นเซลล์โมโนนิวเคลียร์หรือเซลล์หลายนิวเคลียสที่ไม่มีผนังเซลล์ ล้อมรอบด้วยพลาสมาเมมเบรนและชั้นโปรตีนส่วนปลายที่ถูกบดอัด - เพริพลาสต์ ในการยึดเหนี่ยวกับสารตั้งต้นและเพิ่มพื้นผิวการดูดซึม สามารถสร้างผลพลอยได้คล้ายรากซึ่งรวมกันเป็นไรโซไมซีเลียม ในระหว่างการสืบพันธุ์ แทลลัสทั้งหมดจะกลายเป็นสปอรังเจียมเดี่ยว (โฮโลคาร์ปิก แทลลัส) หรือสปอรังเกียหลายอันเกิดขึ้น (ยูคาร์ปิก แทลลัส)
ไมซีเลียลแทลลัสและการปรับเปลี่ยน ขั้นพื้นฐาน องค์ประกอบโครงสร้าง– เส้นใย ไมซีเลียมแบบแบ่งส่วนและไม่แบ่งส่วน Septa และรูปแบบของพวกเขา
การดัดแปลงไมซีเลียม
โครงสร้างที่พัก: chlamydospores และ sclerotia
โครงสร้างการนำไฟฟ้าและรองรับ: เส้นและไรโซมอร์ฟ
โครงสร้างการติดเชื้อ: appressoria และ haustoria
แทลลัสที่มีลักษณะคล้ายยีสต์หมายถึงแต่ละเซลล์ที่มีเยื่อหุ้มเซลล์หรือเซลล์ที่ไม่เสถียรและสลายตัวได้ง่ายซึ่งเกิดขึ้นระหว่างการแตกหน่อเนื่องจากการไม่แยกเซลล์แม่และลูกสาว สถานะคล้ายยีสต์ถาวรและชั่วคราวของ thalli
องค์ประกอบทางเคมีและโครงสร้างผนังเซลล์ของเชื้อรา
ผนังเซลล์ของเชื้อรามีหลายชั้น โดยชั้นต่างๆ เกิดขึ้นจากคาร์โบไฮเดรตที่มีโครงสร้างซึ่งมีองค์ประกอบทางเคมีที่แตกต่างกัน ซึ่งสามารถแบ่งออกเป็น 3 กลุ่มตามองค์ประกอบทางเคมี:
กลูโคสโพลีเมอร์(กลูแคน, ไคติน, เซลลูโลส) กลูแคนประกอบขึ้นเป็นชั้นนอกของผนังเซลล์ของเชื้อราส่วนใหญ่ ชั้นในของผนังเซลล์เชื้อราถูกสร้างขึ้นด้วยสายไคตินทำให้มีความแข็งแกร่ง ไคตินเข้ามาแทนที่เซลลูโลสซึ่งไม่มีอยู่ในเชื้อราส่วนใหญ่ แต่เป็นส่วนหนึ่งของผนังเซลล์ของโอไมซีต ซึ่งปัจจุบันไม่ได้อยู่ในเชื้อราทั่วไป ไคตินแบบดีอะซิติเลตเรียกว่าไคโตซาน ซึ่งเมื่อรวมกับไคตินจะทำให้เกิดผนังเซลล์ของไซโกไมซีต
โพลีเมอร์ของโมโนแซ็กคาไรด์อื่นๆ (มานโนส กาแลคโตส ฯลฯ)ต่างจากพืชชั้นสูงที่พวกมันสร้างพื้นฐานของเมทริกซ์ภายใต้ชื่อทั่วไปว่าเฮมิเซลลูโลส พวกมันมีลักษณะน้อยกว่าของเชื้อรา ข้อยกเว้นคือยีสต์ซึ่งมีผนังเซลล์อุดมไปด้วยโพลีเมอร์แมนโนสที่เรียกว่าแมนแนนเป็นพิเศษ สันนิษฐานว่าองค์ประกอบของผนังนี้ช่วยให้มั่นใจได้ดีกว่า
โพลีเมอร์ของคาร์โบไฮเดรตที่เชื่อมโยงกับเปปไทด์โควาเลนต์ (ไกลโคโปรตีน)สร้างชั้นกลางของผนังเซลล์หลายชั้นและมีบทบาทสำคัญในการรักษาความสมบูรณ์ของโครงสร้างเซลล์และในกระบวนการเผาผลาญกับสิ่งแวดล้อม
คุณสมบัติเฉพาะอื่น ๆ ของเซลล์เชื้อรา ได้แก่ : ไม่มีพลาสติกซึ่งนำมันเข้าใกล้เซลล์สัตว์มากขึ้น
ไม่มีแป้งซึ่งในยูไมซีตจะถูกแทนที่ด้วยโพลีแซ็กคาไรด์ใกล้กับไกลโคเจนในแป้งของสัตว์ และในโอไมซีตจะมีโพลีแซ็กคาไรด์ใกล้กับลามินารินของสาหร่ายสีน้ำตาล นอกจากนี้ยังมีการผลิตคาร์โบไฮเดรตสำรองจำนวนหนึ่งสำหรับเห็ดโดยเฉพาะ
การพัฒนาเฉพาะ สารทุติยภูมิซึ่งยาปฏิชีวนะ ไฟโตและสารพิษจากเชื้อรา และไฟโตฮอร์โมนมีบทบาทสำคัญ
ลักษณะเฉพาะของเห็ดยังได้แก่ โรคเฮเทอโรคาริโอซิสและกระบวนการพาราเซ็กชวล
ในเชื้อราปรากฏการณ์ของเฮเทอโรคาริโอซิสหรือเฮเทอโรนิวเคลียร์เป็นที่แพร่หลายมากซึ่งนิวเคลียสที่เป็นเฮเทอโรอัลลีลิกสำหรับยีนบางตัวยังคงอยู่ในเซลล์เดียวเป็นเวลานาน Heterokaryosis แทนที่เฮเทอโรไซโกซิตี้ในเชื้อราเดี่ยวและส่งเสริมการปรับตัวของเชื้อราอย่างรวดเร็วต่อสภาวะที่เปลี่ยนแปลง การมีอยู่ของนิวเคลียสที่แตกต่างกันนั้นเกิดจากคุณสมบัติเฉพาะหลายประการของเห็ด:
1. การมีอยู่ของนิวเคลียสมากกว่าหนึ่งนิวเคลียสในเซลล์
2. โครงสร้างเฉพาะของผนังเซลล์ซึ่งมีรูทะลุหนึ่งรูขึ้นไปเรียกว่ารูพรุนซึ่งนิวเคลียสสามารถเคลื่อนย้ายจากเซลล์หนึ่งไปยังอีกเซลล์หนึ่งได้
3. เส้นใยภายในอาณานิคมเดียวและแม้แต่โคโลนีที่มีระยะห่างใกล้เคียงกันซึ่งเติบโตจากสปอร์ต่าง ๆ ของเชื้อราชนิดเดียวกันมักจะเติบโตร่วมกันอันเป็นผลมาจากการแลกเปลี่ยนนิวเคลียสของสายพันธุ์ต่าง ๆ เป็นไปได้
พาราเซ็กชวล (นามแฝง)กระบวนการ. หากในนิวเคลียสของเซลล์เฮเทอโรคาริโอตที่เป็นเฮเทอโรอัลลีลิกที่ฟิวส์ตำแหน่งใด ๆ นิวเคลียสซ้ำเฮเทอโรไซกัสจะปรากฏขึ้น มันสามารถเข้าไปในสปอร์และก่อให้เกิดโคลนเฮเทอโรไซกัสแบบดิพลอยด์ ในระหว่างกระบวนการไมโทซิส นิวเคลียสซ้ำสามารถกลับสู่สถานะเดี่ยวได้เนื่องจากการสูญเสียโครโมโซมหนึ่งชุด หรือการแลกเปลี่ยนส่วนของโครโมโซมอาจเกิดขึ้นได้ (การข้ามไมโทติค) กระบวนการทั้งสองจะมาพร้อมกับการรวมตัวกันใหม่ของยีนของผู้ปกครองและฟีโนไทป์ตามมาด้วย การรวมตัวกันอีกครั้งของพาราเซ็กชวล (ไม่อาศัยเพศ) เป็นปรากฏการณ์ที่หายากมาก โดยไม่เกินหนึ่งนิวเคลียสต่อล้าน แต่เนื่องจากมีนิวเคลียสจำนวนมากในไมซีเลียม จึงพบเห็นได้อย่างต่อเนื่องในประชากรเชื้อรา
การสืบพันธุ์– เป็นพืช, ไม่อาศัยเพศ, ทางเพศ.
พืชผัก– การกระจายตัวของแทลลัส, การก่อตัวของคลาไมโดสปอร์ซึ่งหลังจากช่วงพักตัวจะงอกเป็นไมซีเลียมและแตกหน่อในยีสต์
กะเทยการสืบพันธุ์ในเชื้อราต่าง ๆ สามารถทำได้โดยสปอร์ที่เคลื่อนที่ได้และที่ไม่สามารถเคลื่อนที่ได้ Zoospores ก่อตัวเป็นกลุ่มเล็กๆ ของเชื้อราและสิ่งมีชีวิตที่มีลักษณะคล้ายเชื้อรา - ในน้ำและบนบก ซึ่งการเชื่อมต่อทางพันธุกรรมกับเชื้อราในน้ำและสาหร่ายจะมองเห็นได้ชัดเจน โครงสร้างของแฟลเจลลาเป็นคุณลักษณะการวินิจฉัยที่สำคัญเมื่อกำหนดให้กับอาณาจักรเฉพาะ ในเชื้อราส่วนใหญ่ในระหว่างการสืบพันธุ์แบบไม่อาศัยเพศสปอร์ที่ไม่สามารถเคลื่อนที่ได้จะเกิดขึ้นซึ่งบ่งบอกถึงการเกิดขึ้นของพวกมันโบราณบนบก ตามสถานที่ของการก่อตัวและการแปลเป็นภาษาท้องถิ่น sporangiospores ภายนอกมีความโดดเด่นก่อตัวใน sporangia และภายนอก (conidia) ซึ่งพัฒนาบน hyphae พิเศษ - conidiophores Conidia เกิดขึ้นในเชื้อราส่วนใหญ่ (ascomycetes, basidiomycetes, zygomycetes บางชนิด) ซึ่งประกอบเป็นการสร้างสปอร์ของ conidial ซึ่งมีความหลากหลายอย่างมากและใช้กันอย่างแพร่หลายในการวินิจฉัยเชื้อรา
การสืบพันธุ์แบบอาศัยเพศของเชื้อรามีลักษณะเฉพาะของตัวเองทั้งในด้านสัณฐานวิทยาของกระบวนการทางเพศและในกลไกของการควบคุมทางพันธุกรรมและสรีรวิทยาของเพศและการถ่ายทอดข้อมูลทางพันธุกรรม
โซมาโตกามี- กระบวนการทางเพศประเภทที่พบบ่อยที่สุดและง่ายที่สุดประกอบด้วยการหลอมรวมของเซลล์ร่างกายสองเซลล์ซึ่งไม่ได้แยกความแตกต่างออกเป็นเซลล์สืบพันธุ์ บางครั้งมันเกิดขึ้นแม้ไม่มีการหลอมรวมของเซลล์ - นิวเคลียสภายในเซลล์จะรวมกัน พบได้ใน basidiomycetes ส่วนใหญ่ ยีสต์ที่มีกระเป๋าหน้าท้อง และแท็กซ่าอื่นๆ
เกมแทงจิโอกามี– gametangia ถูกแยกออกจากเส้นใยเดี่ยวและเนื้อหาจะถูกระบายออกไปในระหว่างกระบวนการทางเพศ กระบวนการทางเพศนี้เป็นลักษณะของเชื้อราที่มีกระเป๋าหน้าท้องส่วนใหญ่ เกมทังจิโอกามีที่แตกต่างจากเดิมคือไซโกกามีในไซโกไมซีต
เกมโทกามีในรูปของ iso-hetero และ oogamy มักพบในเชื้อราน้อยกว่ายูคาริโอตอื่นมาก Iso- และ heterogamy เกิดขึ้นเฉพาะใน chytridiomycetes oogamy แบบคลาสสิกที่มีการก่อตัวของสเปิร์มและไข่ไม่แสดงออกในเชื้อรา แต่พบสายพันธุ์ที่มีการดัดแปลงอย่างมาก
ตามลักษณะของการควบคุมเพศในเชื้อรากระบวนการทางเพศหลายประเภทมีความโดดเด่น
ไจแอนโดรมิกซิสสามารถพิจารณาได้โดยใช้ตัวอย่างของ oomycetes ที่แตกต่างกัน ซึ่ง oogonia และ antheridia พัฒนาบน thalli ที่แตกต่างกัน เช่น โรคใบไหม้ปลายหรือเชื้อรามันฝรั่ง หากสายพันธุ์ที่เป็นเนื้อเดียวกันทางพันธุกรรมถูกปลูกในพืชเชิงเดี่ยว มันจะสืบพันธุ์แบบไม่อาศัยเพศเท่านั้น หากมีไมซีเลียของสองสายพันธุ์อยู่ใกล้ ๆ การเปลี่ยนแปลงทางสัณฐานวิทยาสามารถตรวจสอบได้ภายใต้อิทธิพลของการหลั่งสารเคมีที่มีลักษณะเป็นสเตียรอยด์ - ฟีโรโมนเพศ Antheridiol ทำให้เกิดการก่อตัวของ antheridia ในคู่หู และ oogoniol ทำให้เกิดการก่อตัวของ oogonia ในกรณีนี้ การควบคุมเรื่องเพศสัมพันธ์กัน: ไม่ว่าสายพันธุ์จะก่อให้เกิดแอนเธอริเดียหรือโอโอโกเนียหรือไม่นั้นขึ้นอยู่กับอัตราส่วนเชิงปริมาณของฟีโรโมนที่เกี่ยวข้องในฟีโรโมนและคู่ของมัน ดังนั้นชื่อของกระบวนการทางเพศ - gynandromixis
Dimixis หรือ Heterallismเป็นที่สังเกตมานานแล้วว่าเชื้อราสามารถเป็นโฮโมหรือเฮเทอโรทัลลิกได้ ในสายพันธุ์โฮโมทัลลิก นิวเคลียสที่เหมือนกันทางพันธุกรรมภายในฟิวส์ไมซีเลียมในระหว่างกระบวนการทางเพศ ในสายพันธุ์เฮเทอโรทอลลิกเพื่อที่จะผ่านวงจรทางเพศในบางระยะ (แตกต่างกันไปตามเชื้อราประเภทต่าง ๆ ) จำเป็นต้องมีการหลอมรวมของลูกหลานของสปอร์ (แม่นยำยิ่งขึ้นคือนิวเคลียสของพวกมัน) เพื่อให้สายพันธุ์สองสายพันธุ์สามารถเข้ากันได้ทางเพศได้ จะต้องมีความแตกต่างทางพันธุกรรม (เฮเทอโรอัลลิลิซิตี) ในบางตำแหน่งที่เรียกว่าตำแหน่งผสมพันธุ์ เชื้อราส่วนใหญ่ (zygomycetes, ascomycetes และ basidiomycetes บางชนิด) มีตำแหน่งผสมพันธุ์หนึ่งตำแหน่งและมีอัลลีลสองตัว ตำแหน่งการผสมพันธุ์ประกอบด้วยยีนหลายตัวที่ควบคุมการสังเคราะห์ฟีโรโมนเพศ ภาวะเฮเทอโรทัลลิซึมดังกล่าวเรียกว่าปัจจัยเดียวหรือไบโพลาร์ ลูกหลานของเชื้อราดังกล่าวหลังไมโอซิสจะถูกแบ่งออกเป็นสองกลุ่มที่เข้ากันไม่ได้ แต่เข้ากันได้ในอัตราส่วน 1:1 เช่น ความน่าจะเป็นของการผสมข้ามพันธุ์ที่เกี่ยวข้อง (ผสมพันธุ์) และไม่เกี่ยวข้อง (ผสมพันธุ์ผสมพันธุ์) คือ 50% เช่นเดียวกับในยูคาริโอตที่สูงกว่าแบบกะเทย
ในจีโนมของ basidiomycetes ที่สูงกว่า มีตำแหน่งผสมพันธุ์สองตำแหน่ง - A และ B และมีเพียงสายพันธุ์ที่เป็นเฮเทอโรอัลลีลิกที่ตำแหน่งทั้งสองเท่านั้นที่เข้ากันได้ (Ax Bx เข้ากันได้กับ Ay By แต่ไม่ใช่กับ Ax By และ Ay Bx) ภาวะเฮเทอโรทัลลิซึมดังกล่าวเรียกว่า ทู-แฟคเตอร์ หรือ เตตราโพลาร์ จะช่วยลดโอกาสในการผสมพันธุ์ได้ถึง 25%
ไดอะโฟโรมิกซิส- basidiomycetes ที่สูงกว่านั้นไม่ได้มีสองอัลลีล แต่มีอัลลีลจำนวนมากของโลคัสผสมพันธุ์ซึ่งพบแบบสุ่มในสายพันธุ์ต่าง ๆ ที่ประกอบกันเป็นประชากร การควบคุมการผสมพันธุ์นี้รับประกันโอกาส 100% ที่จะผสมพันธุ์ เนื่องจากสายพันธุ์ที่มีอัลลีลต่างกันเข้ากันได้และมีอัลลีลจำนวนมาก เป็นผลให้เกิดประชากรลูกผสมแพนมิกซ์
วงจรชีวิตของเชื้อรามีความหลากหลายพอๆ กับเห็ดนั่นเอง วงจรหลักเป็นของแผนกเชื้อรา
1. วงจรไม่อาศัยเพศเป็นลักษณะของกลุ่มเชื้อราที่ไม่สมบูรณ์หรือไมโตเชื้อรากลุ่มใหญ่ที่สูญเสียการสืบพันธุ์แบบอาศัยเพศ การแบ่งนิวเคลียสของพวกมันเป็นแบบไมโทติคโดยเฉพาะ mitofungi ส่วนใหญ่เป็นของเชื้อรา marsupial แต่เนื่องจากการสูญเสียกระบวนการทางเพศ พวกมันจึงรวมตัวกันเป็นกลุ่มของเชื้อราหรือดิวเทอโรไมซีตที่ไม่สมบูรณ์อย่างเป็นทางการ
2. วัฏจักรเดี่ยว แทลลัสที่เป็นพืชมีนิวเคลียสเดี่ยว หลังจากกระบวนการทางเพศ (syngamy) ไซโกเทตซ้ำ (โดยปกติหลังจากช่วงเวลาที่เหลือ) จะแบ่งไมโอซิสแบบไมโอซิส - ไซโกติกไมโอซิส ลักษณะของ zygomycetes และ chytridiomycetes หลายชนิด
3. วงจรฮาพลอยด์-ไดคาริโอตมีลักษณะเฉพาะคือหลังจากการหลอมรวมของเนื้อหาของเกมทังเกีย (เกมทังจิโอกามี) หรือเซลล์ร่างกายของไมซีเลียมเดี่ยว (โซมาโตกามี) นิวเคลียสจะก่อตัวเป็นไดคาริโอต (คู่ของนิวเคลียสที่แตกต่างกันทางพันธุกรรม) พวกมันแบ่งตัวพร้อมกันทำให้เกิดไดคาริโอตไมซีเลียม กระบวนการทางเพศสิ้นสุดลงด้วยการหลอมรวมของนิวเคลียสไดคาริออน ส่งผลให้ไซโกตแบ่งตัวด้วยไมโอซิสโดยไม่มีระยะเวลาพัก Meiospores ประกอบด้วยการสร้างสปอร์ทางเพศของ marsupial และ basidiomycetes ในรูปแบบของ ascospores และ basidiospores เมื่องอกจะเกิดไมซีเลียมเดี่ยว ในเชื้อราที่มีกระเป๋าหน้าท้องส่วนใหญ่ (ยกเว้นเชื้อรายีสต์และทาพริน) วงจรชีวิตถูกครอบงำโดยระยะเดี่ยวในรูปแบบของไมซีเลียมพืช (anamorph) ระยะไดคาริโอตมีอายุสั้นและแสดงด้วยเส้นใย ascogenous ซึ่งถุง ก่อตัวขึ้น (เทเลโอมอร์ฟ) ใน basidiomycetes ระยะไดคาริโอตมีอิทธิพลเหนือวงจรชีวิต ระยะเดี่ยวนั้นมีอายุสั้น
4. วงจรเดี่ยว-ไดพลอยด์ในรูปแบบของการเปลี่ยนแปลงไอโซมอร์ฟิกของรุ่นนั้นหาได้ยากในเชื้อรา (ยีสต์บางชนิดและไคไตรดิโอไมซีตในน้ำ)
5. วงจรดิพลอยด์เป็นลักษณะของโอไมซีตและยีสต์ที่มีกระเป๋าหน้าท้องบางชนิด แทลลัสของพืชเป็นแบบซ้ำซ้อนไมโอซิสที่เกิดจากเซลล์สืบพันธุ์จะสังเกตได้ในระหว่างการก่อตัวของเซลล์สืบพันธุ์หรือเซลล์สืบพันธุ์
3. กลุ่มนิเวศวิทยาของเชื้อรา
เชื้อราและสิ่งมีชีวิตที่มีลักษณะคล้ายเชื้อรารวมอยู่ในระบบนิเวศทั้งทางบกและทางน้ำโดยเป็นส่วนที่สำคัญที่สุดของบล็อกเฮเทอโรโทรฟิค พร้อมด้วยแบคทีเรียที่ครอบครองระดับสารอาหารของผู้ย่อยสลาย การแพร่กระจายของเชื้อราในวงกว้างในชีวมณฑลนั้นพิจารณาจากคุณสมบัติที่สำคัญหลายประการ:
1. ส่วนใหญ่มีโครงสร้างไมซีเลียมของแทลลัส (ช่วยให้คุณควบคุมพื้นผิวได้อย่างรวดเร็วและมีพื้นผิวสัมผัสกับสิ่งแวดล้อมขนาดใหญ่)
2. การเจริญเติบโตและการสืบพันธุ์ที่มีความเร็วสูง ทำให้สามารถตั้งอาณานิคมของสารตั้งต้นจำนวนมากได้ในเวลาอันสั้น ก่อตัวเป็นสปอร์จำนวนมากและกระจายไปในระยะทางไกล
3. กิจกรรมการเผาผลาญสูง แสดงออกในปัจจัยด้านสิ่งแวดล้อมที่หลากหลาย
4. อัตราการรวมตัวกันทางพันธุกรรมสูง, ความแปรปรวนทางชีวเคมีที่สำคัญ, ความเป็นพลาสติกในสิ่งแวดล้อม
5. ความสามารถในการเปลี่ยนไปสู่สถานะของแอนิเมชั่นที่ถูกระงับอย่างรวดเร็วและอยู่รอดในสภาวะที่ไม่เอื้ออำนวยเป็นเวลานาน
ปัจจัยด้านสิ่งแวดล้อมหลักสำหรับเห็ดคือสารตั้งต้นของอาหาร เกี่ยวกับปัจจัยนี้ กลุ่มเชื้อราหลัก มีความโดดเด่นซึ่งเรียกว่ากลุ่มโภชนาการ
4. คำอธิบายโดยย่อของส่วนเชื้อรา
จุลชีววิทยา, 2553, เล่มที่ 79, ครั้งที่ 6, น. 723-733
ยูดีซี 582.281(047)
ผนังเซลล์เชื้อรา: แนวคิดสมัยใหม่เกี่ยวกับองค์ประกอบและการทำงานทางชีววิทยา
© 2010 E. P. Feofilova1
สถานประกอบการ สถาบันการศึกษารัสเซียสถาบันวิทยาศาสตร์จุลชีววิทยาตั้งชื่อตาม เอส.เอ็น. Vinogradsky RAS, มอสโก
รับโดยบรรณาธิการ 05.11.2009
การตรวจสอบนี้เน้นไปที่โครงสร้างพื้นผิวเซลล์ของเชื้อราที่เป็นเส้นใยซึ่งก็คือผนังเซลล์ (CW) ที่ได้รับการศึกษาเพียงเล็กน้อย ข้อมูลมีให้เกี่ยวกับวิธีการแยกเดี่ยวและการทดสอบความบริสุทธิ์สำหรับการไม่มีปริมาณไซโตพลาสซึมในส่วน CS และองค์ประกอบทางเคมี ส่วนประกอบโครงสร้าง (กรอบ) และโครงสร้างพื้นฐานของ CS - อะมิโนโพลีแซ็กคาไรด์, a- และ b-glucans, โปรตีน, ไขมัน, กรดยูโรนิก, ไฮโดรโฟบิน, สปอโรโพลเลนินและเมลานิน - มีการกล่าวถึงในรายละเอียด เอาใจใส่เป็นพิเศษมอบให้กับไคติน มัน คุณลักษณะใหม่ในการต่อต้านความเครียดของเซลล์ เช่นเดียวกับความแตกต่างระหว่างอะมิโนโพลีแซ็กคาไรด์ของเชื้อราและไคตินของสาหร่ายและ Arthropoda มีการกล่าวถึงปรากฏการณ์ของการเจริญเติบโตของปลายยอดและการมีส่วนร่วมของ microvesicles พิเศษในการสร้างสัณฐานวิทยาของเซลล์เชื้อรา ข้อมูลมีให้เกี่ยวกับเอนไซม์ที่เกี่ยวข้องกับการสังเคราะห์และการสลาย CS ท้ายที่สุด จะมีการหารือถึงความสำคัญเชิงหน้าที่ของ SC ในเชื้อราเมื่อเปรียบเทียบกับโครงสร้างพื้นผิวของยูคาริโอตที่สูงกว่า
คำสำคัญ: ราใย ผนังเซลล์ วิธีการแยก องค์ประกอบทางเคมี การเจริญเติบโตยอด ฟังก์ชั่นทางสรีรวิทยา, การสร้างสัณฐานวิทยา
หนึ่งในปัญหาสำคัญ ชีววิทยาสมัยใหม่คำถามคือในระหว่างกระบวนการพัฒนาการก่อตัวของสิ่งมีชีวิตที่มีลักษณะทางสัณฐานวิทยาเหมือนกันเกิดขึ้นได้อย่างไรกลไกทางชีวเคมีและโครงสร้างเซลล์ใดที่เกี่ยวข้องกับกระบวนการนี้ซึ่งเกิดขึ้นบนโลกของเราเป็นเวลาหลายพันล้านปี สิ่งที่น่าสนใจไม่น้อยคือข้อมูลที่บอกว่าความเครียดส่งผลต่อการเกิดสัณฐานวิทยาอย่างไร และโพลีเมอร์ชีวภาพชนิดใดที่ควบคุม แบบฟอร์มภายนอกเซลล์ ตามที่ผลการวิจัยแสดง ปีที่ผ่านมาการเจริญเติบโตของเส้นใยของเชื้อราที่เป็นเส้นใยเป็นรูปแบบพิเศษที่ช่วยให้เข้าใจกระบวนการสร้างเซลล์และวิธีรักษาความคงตัวของสัณฐานวิทยาของเซลล์ในระหว่างการสร้างเซลล์ การมีส่วนร่วมอย่างมากในการศึกษาปรากฏการณ์นี้เกิดขึ้นจากข้อมูลจากการศึกษาองค์ประกอบและ ฟังก์ชั่นทางชีวภาพ CS ของเชื้อราที่มีเส้นใย
CS ของพืชถูกค้นพบในปี 1665 และ CS ของเชื้อราเมื่อต้นศตวรรษที่ 18 เท่านั้น อย่างไรก็ตาม นี่เป็นเรื่องผิวเผิน โครงสร้างเซลล์เป็นเวลานานแล้วที่ไม่ได้ศึกษาในทางปฏิบัติเนื่องจากเชื่อกันว่า CS ทำหน้าที่เพียง "เฟรม" เท่านั้นและเปรียบเทียบกับผนังของบ้านที่รองรับน้ำหนัก แต่ในตอนต้นของศตวรรษที่ 20 ทัศนคติต่อ CS เปลี่ยนไปอย่างมาก เริ่มมีการศึกษาอย่างเข้มข้น แต่ส่วนใหญ่อยู่ในพืชและแบคทีเรีย อย่างไรก็ตาม การพัฒนาอย่างเข้มข้นของการเพาะเห็ดและการผลิตเทคโนโลยีชีวภาพ ซึ่งเชื้อราที่เป็นเส้นใยเป็นผู้ผลิตสารออกฤทธิ์ทางชีวภาพ ทำให้การพัฒนามีความเข้มข้นมากขึ้น
1 ที่อยู่สำหรับติดต่อสื่อสาร (อีเมล: [ป้องกันอีเมล]).
ผูก การวิจัยทางวิทยาศาสตร์ในการศึกษาเชื้อรา CS ข้อมูลเกี่ยวกับโครงสร้างพื้นผิวนี้ในช่วงทศวรรษ 1980 ได้รับการสรุปไว้ในหนังสือเล่มแรกของโลกเกี่ยวกับ CS เชื้อรา ในปีต่อๆ มา งานหลักเกิดขึ้นในช่วงปลายวันที่ 20 และ จุดเริ่มต้นของ XXIศตวรรษ มีการศึกษาพื้นฐานเกี่ยวกับระบบ องค์ประกอบทางเคมีของ CS การเจริญเติบโตของเส้นใยปลายยอด เมแทบอลิซึมของไคติน ยาต้านเชื้อรา ไฮโดรโฟบิน โควาเลนต์ โปรตีนที่ถูกผูกไว้เอนไซม์ที่เกี่ยวข้องกับการก่อตัวของ KS การแตกแขนงของเส้นใยและการสลายของ KS ข้อมูลเหล่านี้ไม่ได้รับการสรุป แต่เป็นการอภิปรายร่วมกันซึ่งทำให้สามารถแสดงให้เห็นว่าโครงสร้างพื้นผิวของเซลล์นี้รับภาระแบบมัลติฟังก์ชั่นและดำเนินการดังกล่าว ฟังก์ชั่นที่จำเป็นเช่นการปกป้องเซลล์จากการออกฤทธิ์ ปัจจัยที่ไม่เอื้ออำนวย, การควบคุมการเกิดสัณฐานวิทยา, การมีส่วนร่วมในกระบวนการสืบพันธุ์, การกำหนดคุณสมบัติของแอนติเจนและกาว, การควบคุมกระบวนการของพฟิสมอร์ฟิซึมและการก่อตัวของเซลล์เชื้อราที่พักผ่อน, การรับรู้สัญญาณภายนอกและการส่งสัญญาณไปยังเยื่อหุ้มเซลล์และผู้ส่งสารภายในเซลล์ เมื่อพิจารณาจากข้างต้น เราถือว่าเหมาะสมที่จะหารือเกี่ยวกับข้อมูลต่อไปนี้: องค์ประกอบทางเคมีของ CS, หน้าที่ทางชีวภาพของพอลิเมอร์ชีวภาพหลัก, โครงสร้างของ CS และปฏิสัมพันธ์ระหว่างเซลล์ในเส้นใยของเชื้อรา, ปรากฏการณ์ของการเจริญเติบโตของเส้นใยปลายยอด และ เอนไซม์หลักที่เกี่ยวข้องกับการก่อตัวและการสลายของ CS
องค์ประกอบทางเคมีของเซลล์
ผนังของเห็ดและการทำงานทางชีวภาพของส่วนประกอบหลัก
กำลังเรียน องค์ประกอบทางเคมี CS ของเชื้อราเริ่มต้นด้วยการได้รับเศษส่วนที่บริสุทธิ์ของโครงสร้างนี้ เช่น การทำให้ CS บริสุทธิ์จากการปนเปื้อนของไซโตพลาสซึม ขั้นตอนเริ่มแรก - การทำลายเซลล์เชื้อราและการล้างสิ่งที่อยู่ในเซลล์ - ดำเนินการที่อุณหภูมิต่ำ (ประมาณ 4-5°C) เพื่อป้องกันการทำลายของเอนไซม์ที่ย่อยสลาย ก่อนที่เซลล์จะถูกทำลาย เซลล์เหล่านั้นจะถูกแช่แข็งที่อุณหภูมิ ไนโตรเจนเหลว- การทำลายเซลล์จะดำเนินการในโฮโมจีไนเซอร์แบบพิเศษหรือแบบกดโดยใช้ " แรงกดดันที่มั่นคง"การใช้เครื่องสลายตัวแบบอัลตราโซนิกต่อไปจะให้ผลลัพธ์ที่ดี เนื้อหาในเซลล์จะถูกล้าง น้ำเย็นและทำซ้ำ 4-5 ครั้ง เพื่อการกำจัดเนื้อหาในไซโตพลาสซึมอย่างละเอียดยิ่งขึ้น ให้ล้างด้วย NaCl, ยูเรีย 8 โมลาร์, แอมโมเนียม 1 โมลาร์ หรือ 0.5 โมลาร์ กรดอะซิติก- ปัจจุบันวิธีการได้รับการพัฒนาเพื่อแยก CS ของเชื้อราในระหว่างการสร้างเซลล์ และวิธีการเหล่านี้แตกต่างกันอย่างมีนัยสำคัญสำหรับไมซีเลียมและเซลล์ที่พัก โดยเฉพาะอย่างยิ่งขึ้นอยู่กับทิศทางของการวิจัยเมื่อพิจารณาองค์ประกอบโพลีแซ็กคาไรด์ไขมันจะถูกสกัดจาก CS โดยใช้ตัวทำละลายอินทรีย์เช่นคลอโรฟอร์มและเมทานอลในอัตราส่วน 2: 1 บางครั้ง ผลลัพธ์ที่ดีให้การบำบัด CS เพิ่มเติมด้วยซัลฟิวริกอีเทอร์ ตัวทำละลายนี้ทำให้ CS แห้งได้ดี และบางครั้งอาจใช้แทนการทำแห้งแบบเยือกแข็งได้ ขั้นตอนต่อไป- การกำหนดความบริสุทธิ์ของเศษส่วน CS ที่เป็นผลลัพธ์ เพื่อจุดประสงค์นี้แสงและ กล้องจุลทรรศน์อิเล็กตรอนและเทคนิคพิเศษในการลงสี ปฏิกิริยาที่ใช้บ่อยที่สุดขึ้นอยู่กับอันตรกิริยาของ I3- กับไคโตซาน CS บริสุทธิ์ที่แยกได้พร้อมสารละลายของ Lugol จะมีสีชมพูหรือ สีม่วงในขณะที่ CS ที่สมบูรณ์จะมีสีแดง นอกจากนี้ยังใช้การย้อมสีสำหรับการมีอยู่ของนิวเคลียสด้วย DAPI (4,6-ekt1to-2-rkepytsn-yo1) ควรเน้นเป็นพิเศษว่าการแยกเศษส่วนบริสุทธิ์ของ CS เป็นขั้นตอนที่มีความรับผิดชอบมากซึ่งขึ้นอยู่กับผลการวิเคราะห์องค์ประกอบทางเคมีในภายหลัง ในเรื่องนี้งานที่น่าสนใจคือพบกลูแคนหรือคอมเพล็กซ์ไคโตซาน - กลูแคนอย่างแม่นยำมากขึ้นในไมซีเลียมของตัวแทนของเชื้อราเมือกแม้ว่าทั้งหมด การศึกษาก่อนหน้าบ่งชี้ว่าไม่มีกลูแคนในไมซีเลียมของมูโคราเลส
ปัจจุบันส่วนประกอบของเชื้อรา CS แบ่งออกเป็นส่วนประกอบโครงสร้าง (ไคติน, p-(1-3)-p(1-6)-กลูแคน, p-(1-4)-กลูแคน (เซลลูโลส) และโครงสร้างภายใน (ได้แก่ เรียกว่าเมทริกซ์) ซึ่งเริ่มรวมถึงแมนโนโปรตีน, กาแลคโต-แมนโนโปรตีน, ไซโล-แมนโนโปรตีน, กลูคูโรโน-
แมนโนโปรตีนและเอ-(1-3)-กลูแคน การศึกษาน้อยที่สุดคือกลูแคนซึ่งเป็นโพลีเมอร์เชิงเส้นของกลูโคส (ใน Schizosaccharomyces pombe กลูแคนนี้มีกลูโคสตกค้างประมาณ 260 ตัว) โพลีเมอร์นี้ประกอบด้วยสายโซ่เชิงเส้นสองเส้นที่เชื่อมต่อกันซึ่งประกอบด้วยเรซิดิวของกลูโคสประมาณ 120 (1-3)-a-D-และ (1-4)-a-D-กลูโคสที่ส่วนปลายของโมเลกุลโพลีเมอร์ เชื่อกันว่ากลูแคนนี้จำเป็นต่อกระบวนการสร้างสัณฐานวิทยา กลูแคนที่ไม่ละลายน้ำถูกแยกได้จาก CS ของ Penicillium roqueforti mycelium ซึ่งหลังจากการอบแห้งจะสูญเสียความสามารถในการละลายในด่าง แต่สามารถละลายได้ในสารละลายลิเธียมคลอไรด์ 10% ในไดเมทิลซัลฟอกไซด์ เมื่อศึกษากลูแคนโดยเมทิลเลชั่นจะได้รับอนุพันธ์ของกลูโคสไตรโอเมทิลสองตัวซึ่งสอดคล้องกับพันธะ 1 - 3 และ 1 - "- 4 ระหว่างโมโนแซ็กคาไรด์ในอัตราส่วนประมาณ 5: 2 และร่องรอยของเตตร้า -O -อนุพันธ์ของเมทิลที่สอดคล้องกับสารตกค้างโมโนโนแซ็กคาไรด์ที่ไม่รีดิวซ์ที่ปลาย ผลลัพธ์นี้บ่งชี้ว่า โครงสร้างเชิงเส้นโมเลกุลที่มีเพียง 1 -»- 3 และ 1 -»- 4 พันธะระหว่างกลูโคสที่ตกค้าง ข้อมูลเหล่านี้ได้รับการยืนยันโดยการวิเคราะห์สเปกตรัม 13C-NMR ของโพลีแซ็กคาไรด์ ซึ่งจะมีการกำหนดค่า a ของกลูโคสที่ตกค้างทั้งหมดตามมาด้วย โพลีแซ็กคาไรด์ดังกล่าวโดยเฉพาะอย่างยิ่งหลังจากการดัดแปลงทางเคมี (ซัลเฟตหรือคาร์โบซิเมทิลเลชั่น) ซึ่งสามารถละลายในน้ำได้ สามารถใช้เป็นโพลีเมอร์ที่มีฤทธิ์ทางชีวภาพได้ และเป็นที่สนใจสำหรับการศึกษาการทำงานทางชีวภาพของพวกมันในผนังเซลล์ของเชื้อราและพืช
ก่อนหน้านี้สารที่ไม่ละลายน้ำ (1 -»- 3)-α-กลูแคนถูกแยกได้จากเชื้อราสูงและต่ำหลายชนิด รวมถึงเชื้อราเพนิซิลเลียม ไครโซจีนัม และα-กลูแคนที่เกี่ยวข้องซึ่งมีพันธะสองประเภท (1-3) และ (1 -» - 4) พบว่ามีอัตราส่วนต่างกัน พืชที่สูงขึ้น(Aconitum kusnezoffii Reichb).
ส่วนประกอบโครงสร้าง- เซลลูโลส - ลักษณะของเชื้อรา oomycete ซึ่งตามอนุกรมวิธานสมัยใหม่จัดอยู่ในประเภท pseudofungi องค์ประกอบโมโนแซ็กคาไรด์ของ CS ประกอบด้วยกลูโคส มานโนส ไซโลส และน้ำตาลหลักคือกลูโคส ซึ่งคิดเป็นสัดส่วนมากถึง 68% องค์ประกอบของเห็ด CS ยังรวมถึงกรดอะมิโน, ไขมัน (ไม่เกิน 3%) และ N-acetyl-glucosamine กลูแคน เช่น 1,3-กลูแคน ก่อให้เกิดสารเชิงซ้อนที่แข็งแกร่งด้วยไคติน เรียกว่าไคติน-กลูแคนคอมเพล็กซ์ (CGC) ซึ่งประกอบขึ้นเป็นโครงกระดูกของเซลล์เชื้อรา สารเชิงซ้อนนี้มีอยู่ใน CS ของเชื้อราเกือบทั้งหมด ยกเว้น zygomycetes ซึ่งทำหน้าที่เป็นคุณสมบัติที่เป็นระบบที่เชื่อถือได้ ในเชื้อราบางชนิด วัสดุเมือกที่ประกอบด้วยโพลีแซ็กคาไรด์ที่แสดงโดย ß-1,3-กลูแคนที่มีเพียงหน่วยกลูโคสที่เชื่อมต่อกันด้วยพันธะ ß-1,6 กับหน่วยกลูโคสทุกๆ สามหน่วยถูกพบบนพื้นผิวของ CS